Submitted:
30 November 2023
Posted:
01 December 2023
You are already at the latest version
Abstract
The receptor tyrosine kinase (EphB4), which produces erythropoietin, was identified as a molecular target for cancer. We identified six peptides as potential inhibitors of EphB4 using an integrated approach combining online bioinformatics tools. According to PeptideRanker, CSM-peptides, SCMTHP, and AcPEP analysis, all peptides are effective against cancer, with the exception of LTAPGQATLPT and VEQNLVPGLK. Furthermore, Toxin-Pred predicts that none of the peptides are toxic, and that only VQLPGWRVFP and TMKGPPDTLQ are potentially allergenic using AllerTop server. PepSite2, HPEPDOCK, and MDockPeP servers were utilized to perform molecular docking, and PacDock was used to examine molecular interactions. The docking score (kcal/mol) range of six promising anticancer peptides against EphB4 was recorded as follows: VQLPGWRVFP (-240.72), FTKDDEWSCFPF (-201.74), SYLPPLSAEVTAK (-177.52), LTAPGQATLPT (-162.23), VEQNLVPGLK (-157.30), TMKGPPDTLQ (-149.67). The main bindings on the allosteric site to VQLPGWRVFP were with R706, S815, R819 and P843 on EphB4. Collectively, our results demonstrated that M. oleifera peptides ameliorate cancer by inhibiting EphB4, and they are promising candidates for the prevention and treatment of cancer.
Keywords:
Moringa oleifera
; anticancer peptides
; EphB4
; Bioinformatics.
1. Introduction
The Eph (erythropoietin-producing hepatocellular carcinoma) receptors represent the largest known family of receptor tyrosine kinases in mammals. Eph receptors are highly conserved proteins that are divided into two subfamilies of nine EphA and five EphB receptors based on sequence similarity [1,2]. The Eph (erythropoietin-producing hepatocellular carcinoma) B receptors are important in a variety of cellular processes through their roles in cell-to-cell contact and signaling; their up-regulation and down-regulation has been shown to have implications in a variety of cancers, and are implicated in a range of processes including angiogenesis, stem cell maintenance and metastasis. They are thus of great interest as targets for cancer therapy [3]. Eph receptors are transmembrane proteins with an extracellular domain capable of recognizing signals from the cells' environment. Ephrins are the ligands to Eph receptors and stimulate bi-directional signaling of the Eph/ephrin axis. Data suggest that Eph/ephrin signaling could play an important role in the development of novel inhibition strategies and cancer treatments to potentially target this receptor tyrosine kinase and/or its ligand [4].
Biopeptides (BPs) are used to kill cancer cells in anticancer peptide therapy, which is an emerging field. BPs can be easily designed using bioinformatics tools to target some protein of interest, including the amino acid sequence, structure, and interaction partners of various oncogenic proteins. Anticancer therapy is effective depending on the stage and type of cancer, but it is expensive and extremely toxic to patients. As a result, there is an urgent need for new research on inexpensive and non-toxic anti-cancer treatments. In this context, anticancer peptides are an alternative to drugs (small molecules) traditionally used in cancer treatments. These fragments of proteins induce cell death through various mechanisms, such as apoptosis, disruption of the tubulin-microtubule balance, or inhibition of angiogenesis [5].
Moringa oleifera Lam. (Moringaceae) has been researched for its chemopreventive capabilities and has been demonstrated to suppress the development of numerous human cancer cells [6]. Many studies have found that M. oleifera leaves can protect organisms and cells against oxidative DNA damage, which is linked to cancer and degenerative illnesses [7]. The extract of M. oleifera leaves decreased the viability of acute myeloid leukemia, acute lymphoblastic leukemia, and hepatocellular carcinoma cells[8]. M. oleifera contains several bioactive chemicals that may be responsible for its anti-cancer activities, including 4-(-l-rhamnosyloxy) benzyl isothiocyanate, niazimicin, and β-sitosterol-3-O—d-glucopyranoside [9]. Moreover, Al-Asmari AK et al., (2015) also proposed that the M. oleifera extracts act as an anti-cancer agent by decreasing cell motility and colony formation in colorectal and breast cancer cell lines. Current research also indicated that the M. oleifera solvent fractions possess in vitro anticancer activity against HeLa cancer cell line [10,11].
Moringa oleifera leaves and seeds have a high percentage of protein, as well as all the necessary amino acids, making them a potential source of functional protein isolate for use in the food and biomedical sectors [12]. Total hydrolysates (TH) and peptide fractions from Moringa seed protein isolates demonstrated that the fraction > 10 kDa of pepsin-trypsin digested for 5 h had higher Angiotensin Converting Enzyme (ACE) inhibitory activity, with an IC50 of 0.224 g/l. Also, antidiabetic efficacy was increased in pepsin-trypsin treatment with 5 hours of hydrolysis, with an IC50 of 0.123 g/L. Ultimately, the hydrolysates of Moringa seed proteins shown great in vitro nutraceutical potential [13]. In addition, eight unique antioxidant peptides of GY, PFE, YTR, FG, QY, IN, SF, SP, and three recognized antioxidant peptides of YFE, IY, and LY were extracted and characterized from M. oleifera seed protein hydrolysate. SF and QY demonstrated considerable protection from H2O2-induced Chang liver cells by boosting the activities of endogenous antioxidant enzymes (superoxide dismutase and catalase) and scavenging intracellular ROS. The findings suggested that SF and QY might be used as natural antioxidants in pharmaceutical or functional foods [14]. Also, it was reported that M. oleifera leaves peptides released by in vitro gastrointestinal digestion might be a potential resource for natural antioxidant and anti-inflammatory components [15]. However, there are no reports on anticancer peptides from M. oleifera. Therefore, the aim of this study is to determine if the previously reported peptides possess this therapeutic property. We have chosen six antioxidant peptides (Figure 1) derived from M. oleifera and compared their anticancer potency against EphB4 by molecular docking.
2. Results
QSAR/QSPR models have been utilized by hundreds of studies in recent years to aid in screening bioactive peptides and predicting their activity. Several of these models have been included in simple online programs that researchers may use as preliminary screening tools. Before conducting wet chemistry studies, researchers can obtain approximated BP profiles with expected bioactivities using servers and protein hydrolysis modeling tools. Researchers can benefit from it by choosing better enzyme-substrate combinations for BP synthesis and identification.
In vitro gastrointestinal digestion of M. oleifera leaves resulted in the selection of peptides for this study. In Avilés-Gaxiola, S. et al., (2021) reported fourteen peptides of different sizes, but only the largest ones were taken into account [15]. Using the CSM-peptides server, Table 1 displays the potential therapeutic effects of six peptides from M. oleifera. All peptides can have multiple activities, with anticancer and anti-inflammatory activities standing out. There are only two peptides (TMKGPPDTLQ and VEQNLVPGLK) that have identical activity, and only the VQLPGWRVFP peptide had anti-viral activity. However, none of the peptides has anti-bacterial activity. Finally, it is interesting to mention that the TMKGPPDTLQ and VEQNLVPGLK peptides have very high quorum sensing activity (Table 1).
Tumor homing peptides (THPs), which consist of 3 to 15 amino acids, have specialized recognition and binding to tumor cells or tumor vasculature. To confirm that anticancer activity is present in all peptides and which of them are THPS, we used other online servers (SCMTHP, ACPred, TumorHPD, and Neptune). The ACPred server only detected the VQLPGWRVFP peptide with a high score for its anti-tancer activity, however, the other peptides were not considered due to their lack of this activity (Table 2). SCMTHP and tumorHPD validated that the VQLPGWRVFP peptide belongs to THP. In addition, FTKDDEWSCFPF and SYLPPLSAEVTAK were the other THP peptides that corresponded to two employed servers. It is important to mention that the LTAPGQATLPT peptide was the only peptide that did not exhibit anticancer activity (Table 2). On the other hand, the peptides VQLPGWRVFP and FTKDDEWSCFPF were the only active peptides according to PeptideRanker. While the ToxinPred server proposes that no peptide is toxic to cells. Finally, the AllerTop2 server suggests that the peptides VQLPGWRVFP and TMKGPPDTLQ could probably be allergens (Table 2).
To identify which type of cancer and the therapeutic concentrations for each peptide, we use the AcPEP server. Table 3 shows that the VQLPGWRVFP peptide presented a dose of 74.75 μM for skin cancer, but a high dose of 478 μM for prostate cancer. The LTAPGQATLPT peptide presented the highest tentative doses compared to the other peptides and these data confirm the results of the previous tables. The TMKGPPDTLQ peptide presented a dose of 163.37 μM for cervical cancer, but a high dose of 551.05 μM for breast cancer. The FTKDDEWSCFPF peptide presented the best dose of 64.49 μM for skin cancer and the lowest dose for lung cancer (Table 3). The SYLPPLSAEVTAK peptide presented the best tentative dose of 83.60 μM for prostate cancer and presented a low dose (267.4 μM) for breast cancer. It is important to mention that three peptides presented doses close to 110 μM for cervical cancer (Table 3).
After discovering anticancer peptides that are favorable, it is crucial to comprehend the mechanism by which they exhibit bioactivities. Molecular docking is an in silico approach to explain the interaction between ligands (BPs) and receptors (target proteins) at the atom level [16]. Based on anticancer prediction, toxicity prediction and allergenic prediction, we performed a molecular docking analysis of peptides using servers (PePSite2, MDockPeP and HPEPDOCK 2.0). According to the Pepsite2 server, all peptides bind to the receptor, but TMKGPPDTLQ has the best probability (Table 4). In contrast, MDockPeP and HPEPDOCK 2.0 use bioactivity analysis to predict bioactivity and take into account the binding energy between the peptides and the receptor. MdockpEP suggests that the FTKDDEWSCFPF peptide presents the best binding energy, while HPEPDOCK 2.0 indicates that the VQLPGWRVFP peptide is the most affinity to the receptor and the second option is FTKDDEWSCFPF (Table 4).
In the course of cancer, the development of tumors, invasiveness, resistance, and angiogenesis are facilitated by EphB4/ephrinB2 signaling [17,18]. In colorectal cancer, EphB4 has been shown to not only promote tumor growth, but also tumor-associated angiogenesis [19]. In ovarian cancer research, it has been found that EphB4 expression is associated with lower survival, and the use of EphB4 targeting has shown promising preclinical activity [20]. Epidermal growth factor receptor (EGFR) signaling partially regulates EphB4 overexpression in bladder cancer and allows for cancer survival through anti-apoptosis signaling [21]. The EphB4 gene appears to be amplified in esophageal tumors and plays a role in tumor cell survival and migration [22]. The EphB4 gene appears to be amplified in esophageal tumors and plays a role in tumor cell survival and migration [23]. In an EphB4 knockdown screen of prostate cancer, EphB4 was shown to regulate integrin β8, a key determinant of prostate cancer invasiveness [24]. Overexpression of the EphB4 ligand ephrinB2 has also been correlated with poor outcome in several tumors. Targeting EphB4/ephrinB2 with specific antibodies was found to be effective in animal models of solid tumors [25]. Epithelial cancers, such as lung cancer, have been found to have EphB4 mutations, while almost one-third of head and neck cancers have EphB4 gene amplification [26].
Based on the above information, we decided to use the crystallographic structure of EphB4 (PDB ID: 3ZEW) to determine if the peptides with anticancer property could interact with this receptor. To gain more insight into how EphB4 interferes with VQLPGWRVFP and FTKDDEWSCFPF, molecular docking was applied to elucidate interactions between the peptide and receptor. The EphB4 showed the greatest peptide binding affinity of -240.72 kcal/mol (Table 4). In the Figure 2, the molecular docking results showed that two peptides could bind EphB4 tightly by hydrogen bonding and hydrophobic interactions. Figure 2B shows that the VQLPGWRVFP peptide binds at the same binding site as staurosporine. While the FTKDDEWSCFPF peptide binds at a more distant site to the pocket occupied by stauroporine (Figure 2D). As shown in Figure 2C, our molecular docking analysis revealed that the binding model suggested that binding between EphB4 and VQLPGWRVFP occurred via hydrogen bonds involving D702 and R744 in the binding pocket, five hydrophobic contacts were observed between the residues F695, V629, D702 and R819. The VQLPGWRVFP also interacted with D702 residue via ionic interaction. The molecular interactions of the peptide FTKDDEWSCFPF with EphB4 showed bonding interactions with the R706, S815, R819, and P843 amino acid residues (Figure 2F).
3. Discussion
M. oleifera is one such tree having tremendous nutritional and medicinal benefits. It is rich in macro- and micronutrients and other bioactive compounds which are important for normal functioning of the body and prevention of certain diseases [27]. Leaves, flowers, seeds, and almost all parts of this tree are edible and have immense therapeutic properties including antidiabetic, anticancer, antiulcer, antimicrobial, and antioxidant [28]. For this reason, there is a great interest in the therapeutic potential of Moringa protein hydrolysates and their subsequent incorporation to functional foods.
The bioactive peptides are encrypted in proteins but can be released by modifications or cleavage from original protein by means of enzymes during gastrointestinal transit or processes as fermentation, germination, heating, and pressure. BPs derived from food proteins have enormous promise as functional foods and nutraceuticals. Bioactive peptides have numerous important activities in the living body, including antioxidative, anti-inflammatory, anticancer, antibacterial, immunomodulatory, and antihypertensive properties [29]. The analysis of the main peptides of the M. oleifera leaves protein hydrolysate using the BIOPEP-UWM database (Minkiewicz, P. et al., 2022) showed the presence of angiotensin I-converting enzyme and dipeptidyl peptidase IV inhibitory peptide sequences [15,30]. These peptides could be further evaluated for their preventive potential against chronic diseases.
Eph/Ephrin signalling plays an important role in tumorigenesis, neovascularization, and vasculogenesis. The EphB receptors play important roles in cell shape, adhesion, migration, and invasion, and their abnormal activity has been associated to the genesis and progression of a variety of tumor forms. Its opposing expression patterns in cancer tissues, together with their high sequence and structural similarity, provide intriguing hurdles to those attempting to design selective therapeutic agents targeting this vast receptor family. All Eph receptors have a highly conserved overall structure with EphA and EphB receptors sharing the same structural features and domains. The primary sequence differences between EphA and EphB receptors reside in a region of the ligand binding domain determined to be a low affinity ephrin binding site which is likely involved in determining ephrin subclass binding specificity [31]. EphB4 protein was more highly expressed in colorectal cancer tissues compared with adjacent normal mucosae (P<0.05), while EphrinB2 levels were unchanged. EphB4 acts as a tumour promoter associated with proliferation, invasion, and angiogenesis, and may be used as a potential CRC therapeutic target (Lv J et al., 2016). It is important to mention that the peptides from M. oleifera are specific for EphB4 (PDB ID: 3ZEW) and present very low interaction with EphB3 (PDB ID: 3ZFY), the VQLPGWRVFP peptide was the only one that presented a positive value (0.1746) for PepSite2. While there is no interaction with the EphA (PDB ID: 3CKH).
Moreover, these peptides bind in the cavity of the stauroporine binding site. Staurosporine is an anti-cancer cell permeable alkaloid isolated from Streptomyces staurosporeus. Staurosporine inhibits protein kinases, including protein kinase C, in a non-selective manner. This chemical causes apoptosis through an unknown mechanism. As a result, these findings are exceedingly promising as anticancer peptides, which can exert anti-tumor activity via several key mechanisms: (a) Apoptosis induction, which involves an energy-dependent cascade mediated via specific proteases or caspases; strategies to overcome tumor resistance to apoptotic pathways include activation of pro-apoptotic receptors, restoration of p53 activity, caspase modulation, and proteasome inhibition [32]. (b) Blockade of intermediate tumor generation by regulating cellular mechanisms associated with cell proliferation and survival, or biosynthetic pathways that control cell growth [33]. (c) Regulation of immune system function by increasing the expression of tumor-associated antigens (antigenicity) in cancer cells, by triggering tumor cells to release danger signals that stimulate immune responses (immunogenicity), or by increasing the predisposition of tumor cells to be recognized and killed by the immune system (susceptibility) [34].
It is important to note that the peptides used in this study can present multiple activities that must be studied in depth. For example, TMKGPPDTLQ and VEQNLVPGLK peptides present very high anti-quorum sensing activity (Table 1). The quorum sensing peptides are mainly secreted by Gram-positive bacteria and can "communicate" with human cells. Some sensing peptides have shown to promote angiogenesis, tumor cell invasion and metastasis of colon cancer as well as of breast cancer cells. On the other hand, quorum sensing cyclodipeptides produced by bacteria and fungi have shown significant antitumor activities [35].
On the other hand, five peptides also possess anti-inflammatory activity. Inflammation is often associated with the development and progression of cancer (Table 1). The cells responsible for cancer-associated inflammation are genetically stable and thus are not subjected to rapid emergence of drug resistance; therefore, the targeting of inflammation represents an attractive strategy both for cancer prevention and for cancer therapy. Tumor-extrinsic inflammation is caused by many factors, including bacterial and viral infections, autoimmune diseases, obesity, tobacco smoking, asbestos exposure, and excessive alcohol consumption, all of which increase cancer risk and stimulate malignant progression. In contrast, cancer-intrinsic or cancer-elicited inflammation can be triggered by cancer-initiating mutations and can contribute to malignant progression through the recruitment and activation of inflammatory cells. Both extrinsic and intrinsic inflammations can result in immunosuppression, thereby providing a preferred background for tumor development [36].
Finally, the results in Table 3 suggest that the peptides could be more effective for skin or cervical cancer. However, studies are required with different cancer cell lines such as A-431, HT-3, HeLa, MCF-7 and A-549 for validating cancer targets and for defining drug efficacy. These results correlate with the findings reported by Al-Asmari AK et al., (2015) who reported that both the leaf and bark extracts of Moringa collected from the Saudi Arabian region possess anti-cancer activity against MDA-MB-231 and HCT-8 cancer cell lines and that can be used to develop new drugs for treatment of breast and colorectal cancers [11]. Moreover, The Moringa oleifera bioactive compounds have been effective in the suppression of cancers, making them the therapeutic agents of choice for the current investigation to treat Melanoma-associated gene expression antigens presented in non-small cell lung cancer [37].
Further research is needed to ensure appropriate preclinical and clinical evidence to meet the criterion of safe and effective treatment. However, M. oleifera is a valuable species that provides a significant but still poorly studied source of anticancer peptides.
4. Materials and Methods
In silico analysis can aid in the prediction of a wide variety of factors used to derive BPs and assess biological activity. Moreover, utilizing the web tools, the basic characteristics of the peptides may be examined. To predict the activity of target molecules, quantitative structure-activity relationship (QSAR) and molecular docking are utilized. AllerTOP2, for example, is the first alignment-free server for in silico allergy prediction based on the basic physicochemical features of proteins [38]. TumorHPD is a website that predicts and designs tumor homing peptides [39]. The overall bioactivity of the in silico released peptides was estimated using tools available in PeptideRanker (provider: University College Dublin, Ireland; http://distilldeep.ucd.ie/PeptideRanker/) [40]. The PeptideRanker tool can sort a set of peptides and award scores ranging from 0 to 1 based on their function-structure models. The greatest score indicates the most active peptides, while the lowest indicates the least active. Peptides for which the PeptideRanker Score was higher than 0.5 were analyzed using Pepsite2 (provider: University of Heidelberg, Germany; http://pepsite2.russelllab.org.) [41], which allows computing the potential interaction (p-value) between the peptide and target enzymes (PDB—Protein Data Bank; provider: Research Collaboratory for Structural Bioinformatics; http://www.rcsb.org/pdb/). CSM-peptides, a novel machine learning method for rapid identification of eight different types of therapeutic peptides: anti-angiogenic, anti-bacterial, anti-cancer, anti-inflammatory, anti-viral, cell-penetrating, quorum sensing, and surface binding; https://biosig.lab.uq.edu.au/csm_peptides/ [42]. AcPEP is a server of sequence-based machine learning methods for anticancer peptide (ACP) prediction. This server accepts peptide amino acid sequences in FASTA format. Firstly, it predicts if the sequences are ACPs, and then it predicts their biological activities against 6 different cancer types: breast, cervix, colon, lung, prostate, and skin; https://app.cbbio.online/acpep/home/ [43]. The PepSite 2 web server was used to predict peptide-binding sites in EphB4; http://pepsite2.russelllab.org/ [41]. Receptor grids were generated for the prepared protein structures so that peptides bind within the predicted active site. Receptor grids were generated with default values for van der Waals scaling factor (1.00) and charge cutoff (0.25). A cubic search space, centered on the centroid of the active site residues predicted by PepSite 2, was generated for each receptor. The EphB4 model was obtained from the deposited X-ray structure PDB ID 3ZEW. The SMILES format were obtained from the amino acid sequence using the server the Novoprolabs tools (https://www.novoprolabs.com/tools/convert-peptide-to-smiles-string) accessed on 05 January 2023. The conformational search was performed using the Cheminformatic tools and databases for Pharmacology (https://chemoinfo.ipmc.cnrs.fr/) accessed on 06 January 2023, and the most stable conformers were chosen and optimized [44]. PacDock is a novel web service for fully automated detection and visualization of relevant non-covalent protein-ligand contacts in 3D structures, freely available at https://pegasus.lbic.unibo.it/pacdock/PacVIEW_Receptor_Ligand_Interactions.html [45].
5. Conclusions
It can be concluded that both VQLPGWRVFP and FTKDDEWSCFPF are the best peptides with EphB4 inhibitory activity in silico. To our knowledge, this is the first time that anti-cancer biopeptides of M. oleifera are reported and thereby contributing new insights into the nutraceutical properties of this medicinal plant. We report the molecular docking analysis of anticancer peptides VQLPGWRVFP and FTKDDEWSCFPF with the target EphB4 receptor. Both peptides can be used for treatment of cancer, mainly cervix and skin cancer, although it would be necessary to perform bioavailability and clinical studies to demonstrate their efficiency in vitro and in vivo models.
Author Contributions
Conceptualization, E.S.-C., M.V.-S., C.A.-D., and A.T.-V.; Formal analysis, E.S.-C., M.V.-S., C.A.-D., and A.T.-V.; Investigation, E.S.-C., and M.V.-S.; Supervision, E.S.-C., M.V.-S., C.A.-D., and A.T.-V.; Writing—original draft, E.S.-C., and M.V.-S. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Acknowledgments
The authors gratefully acknowledge to Jorge L. Arreola-Berumen and Iván A. Corral-Guerrero for technical support.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Pasquale, E.B. The Eph Family of Receptors. Curr. Opin. Cell Biol. 1997, 9, 608–615. [Google Scholar] [CrossRef] [PubMed]
- Liang, L.-Y.; Patel, O.; Janes, P.W.; Murphy, J.M.; Lucet, I.S. Eph Receptor Signalling: From Catalytic to Non-Catalytic Functions. Oncogene 2019, 38, 6567–6584. [Google Scholar] [CrossRef] [PubMed]
- Overman, R.C.; Debreczeni, J.E.; Truman, C.M.; McAlister, M.S.; Attwood, T.K. Biochemical and Biophysical Characterization of Four EphB Kinase Domains Reveals Contrasting Thermodynamic, Kinetic and Inhibition Profiles. Biosci. Rep. 2013, 33. [Google Scholar] [CrossRef] [PubMed]
- Surawska, H.; Ma, P.C.; Salgia, R. The Role of Ephrins and Eph Receptors in Cancer. Cytokine Growth Factor Rev. 2004, 15, 419–433. [Google Scholar] [CrossRef] [PubMed]
- Kim, E.-K.; Kim, Y.-S.; Hwang, J.-W.; Lee, J.S.; Moon, S.-H.; Jeon, B.-T.; Park, P.-J. Purification and Characterization of a Novel Anticancer Peptide Derived from Ruditapes Philippinarum. Process Biochem. 2013, 48, 1086–1090. [Google Scholar] [CrossRef]
- Karim, N.A.A.; Ibrahim, M.D.; Kntayya, S.B.; Rukayadi, Y.; Hamid, H.A.; Razis, A.F.A. Moringa Oleifera Lam Targeting Chemoprevention. Asian Pacific J. Cancer Prev. 2016, 17, 3675–3686. [Google Scholar]
- Sikder, K.; Sinha, M.; Das, N.; Das, D.K.R.; Datta, S.; Dey, S. Moringa Oleifera Leaf Extract Prevents in Vitro Oxidative DNA Damage. Asian J. Pharm. Clin. Res. 2013, 6, 157–161. [Google Scholar]
- Khalafalla, M.M.; Abdellatef, E.; Dafalla, H.M.; Nassrallah, A.A.; Aboul-Enein, K.M.; Lightfoot, D.A.; El-Deeb, F.E.; El-Shemy, H.A. Active Principle from Moringa Oleifera Lam Leaves Effective against Two Leukemias and a Hepatocarcinoma. African J. Biotechnol. 2010, 9, 8467–8471. [Google Scholar]
- Abdull Razis, A.F.; Ibrahim, M.D.; Kntayya, S.B. Health Benefits of Moringa Oleifera. Asian Pacific J. Cancer Prev. 2014, 15, 8571–8576. [Google Scholar] [CrossRef]
- Mumtaz, M.Z.; Kausar, F.; Hassan, M.; Javaid, S.; Malik, A. Anticancer Activities of Phenolic Compounds from Moringa Oleifera Leaves: In Vitro and in Silico Mechanistic Study. Beni-Suef Univ. J. Basic Appl. Sci. 2021, 10, 12. [Google Scholar] [CrossRef]
- Al-Asmari, A.K.; Albalawi, S.M.; Athar, M.T.; Khan, A.Q.; Al-Shahrani, H.; Islam, M. Moringa Oleifera as an Anti-Cancer Agent against Breast and Colorectal Cancer Cell Lines. PLoS One 2015, 10, e0135814. [Google Scholar] [CrossRef]
- Kumar, M.; Selvasekaran, P.; Kapoor, S.; Barbhai, M.D.; Lorenzo, J.M.; Saurabh, V.; Potkule, J.; Changan, S.; ElKelish, A.; Selim, S.; et al. Moringa Oleifera Lam. Seed Proteins: Extraction, Preparation of Protein Hydrolysates, Bioactivities, Functional Food Properties, and Industrial Application. Food Hydrocoll. 2022, 131, 107791. [Google Scholar] [CrossRef]
- González Garza, N.G.; Chuc Koyoc, J.A.; Torres Castillo, J.A.; García Zambrano, E.A.; Betancur Ancona, D.; Chel Guerrero, L.; Sinagawa García, S.R. Biofunctional Properties of Bioactive Peptide Fractions from Protein Isolates of Moringa Seed (Moringa Oleifera). J. Food Sci. Technol. 2017, 54, 4268–4276. [Google Scholar] [CrossRef] [PubMed]
- Liang, L.; Cai, S.; Gao, M.; Chu, X.; Pan, X.; Gong, K.-K.; Xiao, C.; Chen, Y.; Zhao, Y.; Wang, B.; et al. Purification of Antioxidant Peptides of Moringa Oleifera Seeds and Their Protective Effects on H2O2 Oxidative Damaged Chang Liver Cells. J. Funct. Foods 2020, 64, 103698. [Google Scholar] [CrossRef]
- Avilés-Gaxiola, S.; León-Félix, J.; Jiménez-Nevárez, Y.B.; Angulo-Escalante, M.A.; Ramos-Payán, R.; Colado-Velázquez, J.; Heredia, J.B. Antioxidant and Anti-Inflammatory Properties of Novel Peptides from Moringa Oleifera Lam. Leaves. South African J. Bot. 2021, 141, 466–473. [Google Scholar] [CrossRef]
- Chen, Z.; Li, W.; Santhanam, R.K.; Wang, C.; Gao, X.; Chen, Y.; Wang, C.; Xu, L.; Chen, H. Bioactive Peptide with Antioxidant and Anticancer Activities from Black Soybean [Glycine Max (L.) Merr.] Byproduct: Isolation, Identification and Molecular Docking Study. Eur. Food Res. Technol. 2019, 245, 677–689. [Google Scholar] [CrossRef]
- Boyd, A.W.; Bartlett, P.F.; Lackmann, M. Therapeutic Targeting of EPH Receptors and Their Ligands. Nat. Rev. Drug Discov. 2014, 13, 39–62. [Google Scholar] [CrossRef] [PubMed]
- Xi, H.-Q.; Wu, X.-S.; Wei, B.; Chen, L. Eph Receptors and Ephrins as Targets for Cancer Therapy. J. Cell. Mol. Med. 2012, 16, 2894–2909. [Google Scholar] [CrossRef] [PubMed]
- McCall, J.L.; Gehring, D.; Clymer, B.K.; Fisher, K.W.; Das, B.; Kelly, D.L.; Kim, H.; White, M.A.; Lewis, R.E. KSR1 and EPHB4 Regulate Myc and PGC1β To Promote Survival of Human Colon Tumors. Mol. Cell. Biol. 2016, 36, 2246–2261. [Google Scholar] [CrossRef] [PubMed]
- Spannuth, W.A.; Mangala, L.S.; Stone, R.L.; Carroll, A.R.; Nishimura, M.; Shahzad, M.M.K.; Lee, S.-J.; Moreno-Smith, M.; Nick, A.M.; Liu, R.; et al. Converging Evidence for Efficacy from Parallel EphB4-Targeted Approaches in Ovarian Carcinoma. Mol. Cancer Ther. 2010, 9, 2377–2388. [Google Scholar] [CrossRef]
- Xia, G.; Kumar, S.R.; Stein, J.P.; Singh, J.; Krasnoperov, V.; Zhu, S.; Hassanieh, L.; Smith, D.L.; Buscarini, M.; Broek, D.; et al. EphB4 Receptor Tyrosine Kinase Is Expressed in Bladder Cancer and Provides Signals for Cell Survival. Oncogene 2006, 25, 769–780. [Google Scholar] [CrossRef] [PubMed]
- Hasina, R.; Mollberg, N.; Kawada, I.; Mutreja, K.; Kanade, G.; Yala, S.; Surati, M.; Liu, R.; Li, X.; Zhou, Y.; et al. Critical Role for the Receptor Tyrosine Kinase EPHB4 in Esophageal Cancers. Cancer Res. 2013, 73, 184–194. [Google Scholar] [CrossRef] [PubMed]
- Liersch-Löhn, B.; Slavova, N.; Buhr, H.J.; Bennani-Baiti, I.M. Differential Protein Expression and Oncogenic Gene Network Link Tyrosine Kinase Ephrin B4 Receptor to Aggressive Gastric and Gastroesophageal Junction Cancers. Int. J. Cancer 2016, 138, 1220–1231. [Google Scholar] [CrossRef] [PubMed]
- Mertens-Walker, I.; Fernandini, B.C.; Maharaj, M.S.; Rockstroh, A.; Nelson, C.C.; Herington, A.C.; Stephenson, S.-A. The Tumour-Promoting Receptor Tyrosine Kinase, EphB4, Regulates Expression of Integrin-Β8 in Prostate Cancer Cells. BMC Cancer 2015, 15, 164. [Google Scholar] [CrossRef] [PubMed]
- Abéngozar, M.A.; de Frutos, S.; Ferreiro, S.; Soriano, J.; Perez-Martinez, M.; Olmeda, D.; Marenchino, M.; Cañamero, M.; Ortega, S.; Megias, D.; et al. Blocking EphrinB2 with Highly Specific Antibodies Inhibits Angiogenesis, Lymphangiogenesis, and Tumor Growth. Blood 2012, 119, 4565–4576. [Google Scholar] [CrossRef] [PubMed]
- Ferguson, B.D.; Carol Tan, Y.-H.; Kanteti, R.S.; Liu, R.; Gayed, M.J.; Vokes, E.E.; Ferguson, M.K.; John Iafrate, A.; Gill, P.S.; Salgia, R. Novel EPHB4 Receptor Tyrosine Kinase Mutations and Kinomic Pathway Analysis in Lung Cancer. Sci. Rep. 2015, 5, 10641. [Google Scholar] [CrossRef] [PubMed]
- Valdez-Solana, M.A.; Mejía-García, V.Y.; Téllez-Valencia, A.; García-Arenas, G.; Salas-Pacheco, J.; Alba-Romero, J.J.; Sierra-Campos, E. Nutritional Content and Elemental and Phytochemical Analyses of Moringa Oleifera Grown in Mexico. J. Chem. 2015, 2015, 1–9. [Google Scholar] [CrossRef]
- Islam, Z.; Islam, S.M.R.; Hossen, F.; Mahtab-ul-Islam, K.; Hasan, M.R.; Karim, R. Moringa Oleifera Is a Prominent Source of Nutrients with Potential Health Benefits. Int. J. Food Sci. 2021, 2021, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Zaky, A.A.; Simal-Gandara, J.; Eun, J.-B.; Shim, J.-H.; Abd El-Aty, A.M. Bioactivities, Applications, Safety, and Health Benefits of Bioactive Peptides From Food and By-Products: A Review. Front. Nutr. 2022, 8. [Google Scholar] [CrossRef]
- Minkiewicz, P.; Iwaniak, A.; Darewicz, M. BIOPEP-UWM Virtual—A Novel Database of Food-Derived Peptides with In Silico-Predicted Biological Activity. Appl. Sci. 2022, 12, 7204. [Google Scholar] [CrossRef]
- Himanen, J.-P.; Henkemeyer, M.; Nikolov, D.B. Crystal Structure of the Ligand-Binding Domain of the Receptor Tyrosine Kinase EphB2. Nature 1998, 396, 486–491. [Google Scholar] [CrossRef] [PubMed]
- Burz, C.; Berindan-Neagoe, I.; Balacescu, O.; Irimie, A. Apoptosis in Cancer: Key Molecular Signaling Pathways and Therapy Targets. Acta Oncol. (Madr). 2009, 48, 811–821. [Google Scholar] [CrossRef] [PubMed]
- Kornienko, A.; Mathieu, V.; Rastogi, S.K.; Lefranc, F.; Kiss, R. Therapeutic Agents Triggering Nonapoptotic Cancer Cell Death. J. Med. Chem. 2013, 56, 4823–4839. [Google Scholar] [CrossRef] [PubMed]
- Zitvogel, L.; Galluzzi, L.; Smyth, M.J.; Kroemer, G. Mechanism of Action of Conventional and Targeted Anticancer Therapies: Reinstating Immunosurveillance. Immunity 2013, 39, 74–88. [Google Scholar] [CrossRef] [PubMed]
- Tornesello, A.L.; Buonaguro, L.; Tornesello, M.L.; Buonaguro, F.M. The Role of Sensing Peptides in the Cross-Talk between Microbiota and Human Cancer Cells. Mini-Reviews Med. Chem. 2018, 18, 1567–1571. [Google Scholar] [CrossRef] [PubMed]
- Singh, N.; Baby, D.; Rajguru, J.; Patil, P.; Thakkannavar, S.; Pujari, V. Inflammation and Cancer. Ann. Afr. Med. 2019, 18, 121. [Google Scholar] [CrossRef] [PubMed]
- Bhat, S.S.; Mahapatra, S. Das; R, S.; Sommano, S.R.; Prasad, S.K. Virtual Screening and Quantitative Structure–Activity Relationship of Moringa Oleifera with Melanoma Antigen A (MAGE-A) Genes against the Therapeutics of Non-Small Cell Lung Cancers (NSCLCs). Cancers (Basel). 2022, 14, 5052. [Google Scholar] [CrossRef] [PubMed]
- Dimitrov, I.; Bangov, I.; Flower, D.R.; Doytchinova, I. AllerTOP v.2—a Server for in Silico Prediction of Allergens. J. Mol. Model. 2014, 20, 2278. [Google Scholar] [CrossRef] [PubMed]
- Sharma, A.; Kapoor, P.; Gautam, A.; Chaudhary, K.; Kumar, R.; Chauhan, J.S.; Tyagi, A.; Raghava, G.P.S. Computational Approach for Designing Tumor Homing Peptides. Sci. Rep. 2013, 3, 1607. [Google Scholar] [CrossRef]
- Mooney, C.; Haslam, N.J.; Pollastri, G.; Shields, D.C. Towards the Improved Discovery and Design of Functional Peptides: Common Features of Diverse Classes Permit Generalized Prediction of Bioactivity. PLoS One 2012, 7, e45012. [Google Scholar] [CrossRef]
- Trabuco, L.G.; Lise, S.; Petsalaki, E.; Russell, R.B. PepSite: Prediction of Peptide-Binding Sites from Protein Surfaces. Nucleic Acids Res. 2012, 40, W423–W427. [Google Scholar] [CrossRef]
- Rodrigues, C.H.M.; Garg, A.; Keizer, D.; Pires, D.E. V.; Ascher, D.B. CSM-peptides: A Computational Approach to Rapid Identification of Therapeutic Peptides. Protein Sci. 2022, 31. [Google Scholar] [CrossRef]
- Chen, J.; Cheong, H.H.; Siu, S.W.I. XDeep-AcPEP: Deep Learning Method for Anticancer Peptide Activity Prediction Based on Convolutional Neural Network and Multitask Learning. J. Chem. Inf. Model. 2021, 61, 3789–3803. [Google Scholar] [CrossRef]
- Riniker, S.; Landrum, G.A. Better Informed Distance Geometry: Using What We Know to Improve Conformation Generation. J. Chem. Inf. Model. 2015, 55, 2562–2574. [Google Scholar] [CrossRef]
- Carbone, J.; Ghidini, A.; Romano, A.; Gentilucci, L.; Musiani, F. PacDOCK: A Web Server for Positional Distance-Based and Interaction-Based Analysis of Docking Results. Molecules 2022, 27, 6884. [Google Scholar] [CrossRef]
Figure 1.
Chemical structures of the selected peptides previously reported [15].
Figure 1.
Chemical structures of the selected peptides previously reported [15].
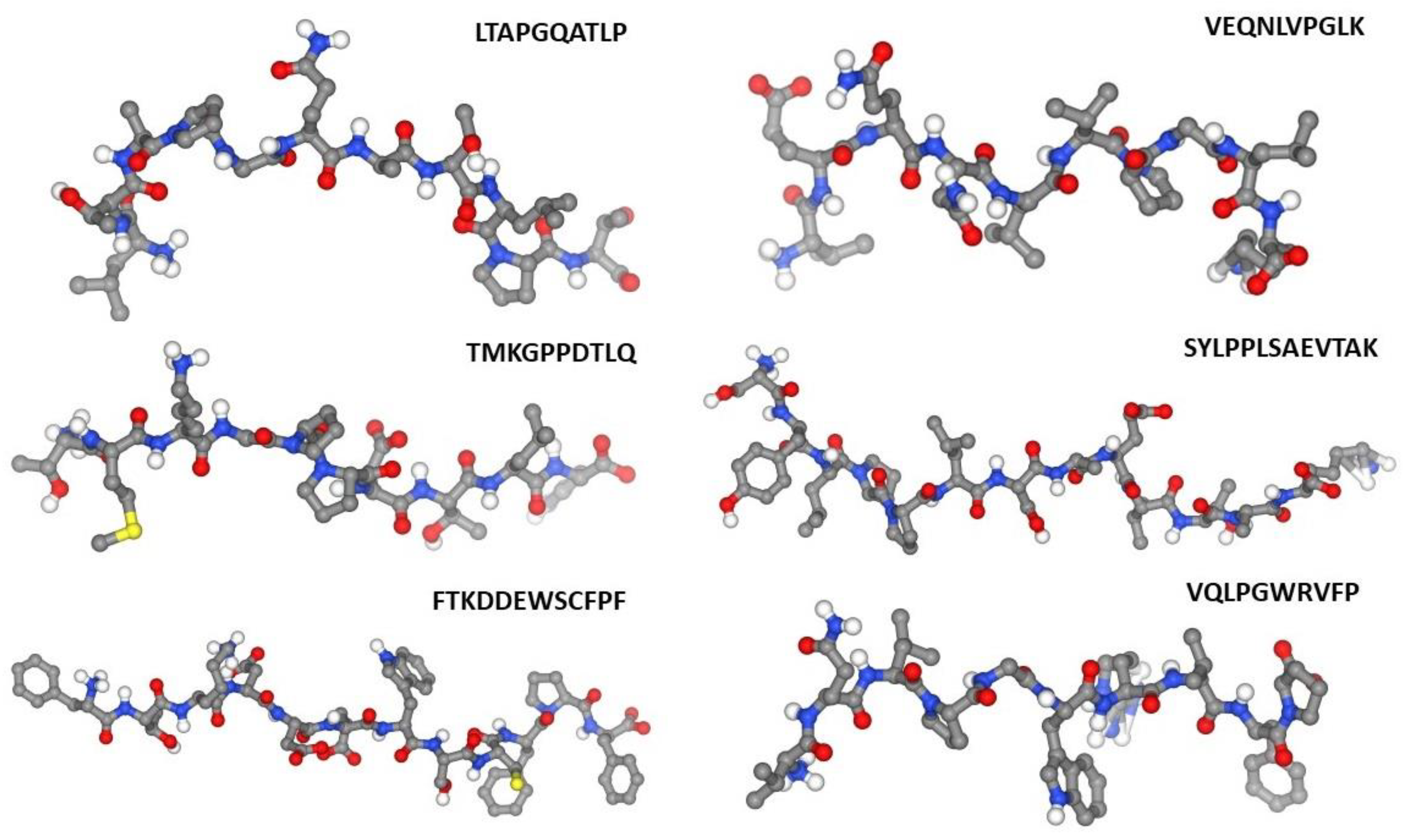
Figure 2.
The conformations with the lowest docking energy were selected for demonstration. The protein is shown in ribbon representation (A and D), whereas peptides are shown as mesh representation: B) VQLPGWRVFP (blue), E) FTKDDEWSCFPF (pink). Figure 2C and 2F were obtained with PacDOCK web server.
Figure 2.
The conformations with the lowest docking energy were selected for demonstration. The protein is shown in ribbon representation (A and D), whereas peptides are shown as mesh representation: B) VQLPGWRVFP (blue), E) FTKDDEWSCFPF (pink). Figure 2C and 2F were obtained with PacDOCK web server.
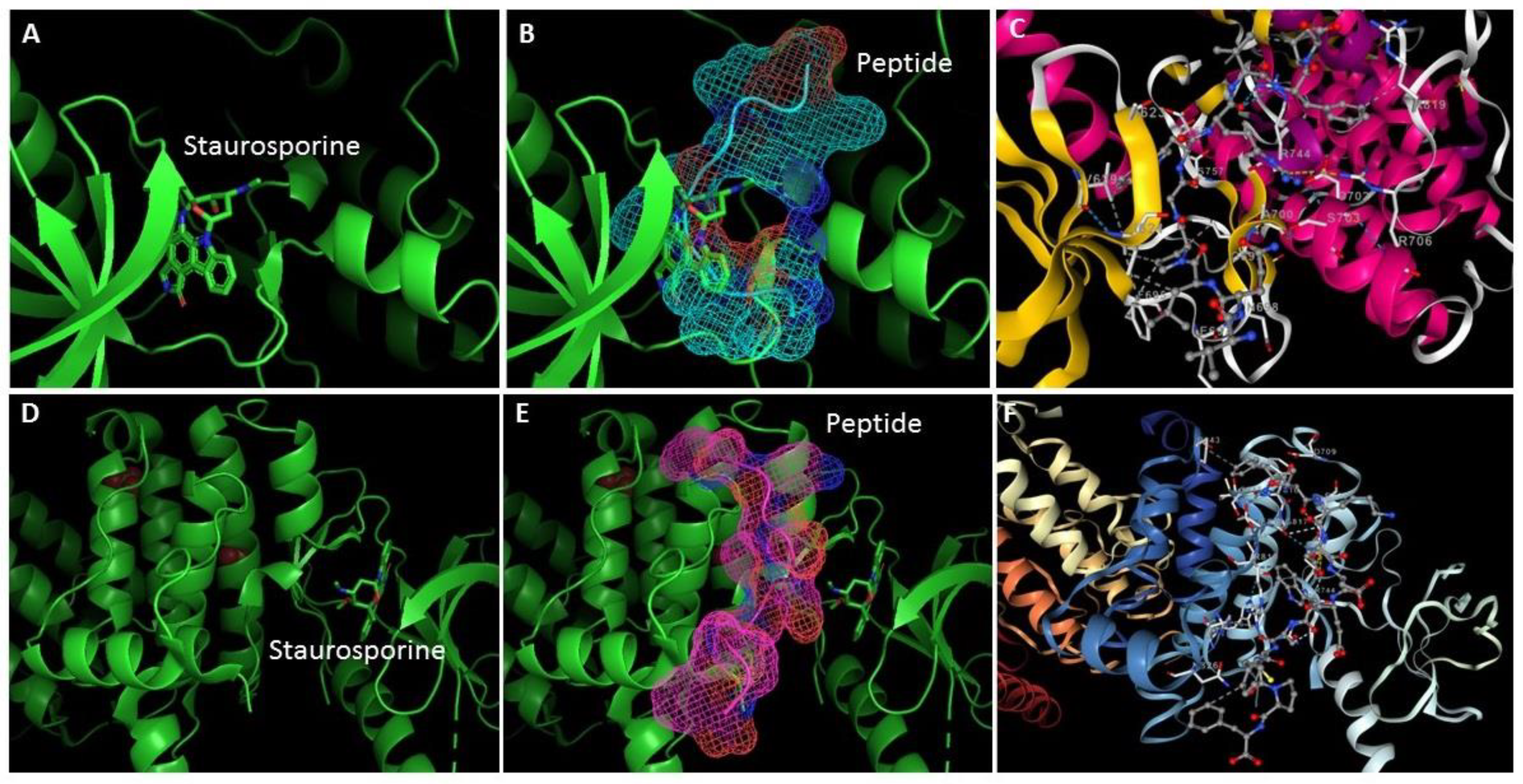

Table 1.
Therapeutic activity Prediction for M. oleifera peptides by CSM-peptides.
|
Peptide sequence |
Activity Prediction | |||||
| Anti-angiogenic | Anti-Bacterial | Anti-cancer | Anti-inflammatory | Anti-viral | Quorum Sensing | |
| VQLPGWRVFP | ---------- | --------- | 0.72 | 0.64 | 0.52 | -------- |
| LTAPGQATLPT | ---------- | --------- | 0.52 | -------- | --------- | -------- |
| TMKGPPDTLQ | 0.51 | --------- | 0.57 | 0.71 | --------- | 0.92 |
| FTKDDEWSCFPF | ---------- | --------- | 0.54 | 0.54 | --------- | -------- |
| SYLPPLSAEVTAK | ---------- | --------- | 0.53 | 0.53 | --------- | -------- |
| VEQNLVPGLK | 0.57 | --------- | 0.79 | 0.77 | --------- | 0.97 |
Table 2.
Tumor homing peptides, toxicity and allergenic characteristics for M. oleifera peptides.
|
Peptide sequence |
Multiple peptide servers | ||||||
| SCMTHP | ACPred | TumorHPD | Neptune | PR* | ToxinPred | AllerTop | |
| VQLPGWRVFP | THP (392.9) | ACP (0.91) | THP (0.33) | NON-THP (0.283) | 0.72 | Non-toxin | Allergen |
| LTAPGQATLPT | NON-THP (277.0) | NON-ACP (0.932) | NON-THP (-0.86) | NON-THP (0.097) | 0.18 | Non-toxin | Non- Allergen |
| TMKGPPDTLQ | THP (392.9) | NON-ACP (0.977) | NON-THP (-0.86) | NON-THP (0.368) | 0.14 | Non-toxin | Allergen |
| FTKDDEWSCFPF | THP (392.9) | NON-ACP (0.901) | THP (0.89) | NON-THP (0.063) | 0.80 | Non-toxin | Non- Allergen |
| SYLPPLSAEVTAK | THP (392.9) | NON-ACP (0.973) | NON-THP (-0.69) | THP (0.935) | 0.34 | Non-toxin | Non- Allergen |
| VEQNLVPGLK | NON-THP (209.3) | NON-ACP (0.986) | NON-THP (-0.1) | NON-THP (0.053) | 0.27 | Non-toxin | Non- Allergen |
*PR, PeptideRanker.
Table 3.
Anticancer prediction for M. oleifera peptides by AcPEP server.
|
Peptide sequence |
Activity Prediction | |||||
| Breast (μM) | Cervix (μM) | Colon (μM) | Lung (μM) | Prostate (μM) | Skin (μM) | |
| VQLPGWRVFP | 309.61 | 110.02 | 412.54 | 277.14 | 478.23 | 74.75 |
| LTAPGQATLPT | 551.05 | 408.95 | 490.98 | 822.35 | 301.11 | 503.92 |
| TMKGPPDTLQ | 520.78 | 163.37 | 617.55 | 684.13 | 386.68 | 372.89 |
| FTKDDEWSCFPF | 180.87 | 291.62 | 368.83 | 126.08 | 479.94 | 64.497 |
| SYLPPLSAEVTAK | 267.4 | 114.22 | 244.69 | 588.18 | 83.60 | 299.26 |
| VEQNLVPGLK | 474.14 | 109.63 | 557.37 | 457.76 | 315.45 | 297.65 |
Table 4.
Molecular docking between EPHRIN TYPE B receptor 4 and Moringa oleifera leaves peptides.
| Peptide sequence | PepSite2 p-value | ITScorePeP | HPEPDOCK 2.0 |
| VQLPGWRVFP | 0.0196 | -165.2 | -240.72 |
| LTAPGQATLPT | 0.0673 | -109.8 | -162.23 |
| TMKGPPDTLQ | 0.0078 | -120.2 | -149.67 |
| FTKDDEWSCFPF | 0.0720 | -217.1 | -201.74 |
| SYLPPLSAEVTAK | 0.0862 | -151.9 | -177.52 |
| VEQNLVPGLK | 0.0760 | -121.6 | -157.30 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



