Submitted:
04 December 2023
Posted:
05 December 2023
You are already at the latest version
Abstract
In recent years, magnetic fields have emerged as a non-thermophysical treatment with a signifi-cant impact on microbial fermentation processes. Brassica trispora is a microorganism known for its indus-trial-scale production of lycopene and high yield of single cells. This study aimed to in-vestigate the impact of low frequency magnetic fields on lycopene synthesis by Brassica trispora and elucidate the underlying mechanism for enhancing lycopene yield. The results indicated that both the intensity and duration of the magnetic field treatment influenced the cell. Exposing the cell to a 0.5 mT magnetic field for 48 hours on the second day of fermentation resulted in a lyco-pene yield of 1473.16 mg/L, representing a re-markable increase of 216.1% compared to the con-trol group. Transcriptome analysis revealed that al-ternating magnetic field significantly up-regulated genes related to ROS and cell membrane structure, leading to a substantial increase in lycopene production. Scanning electron microscopy revealed that the magnetic field treatment resulted in rough, loose, wrinkled surface morphology of the mycelium, along with a few mi-cropores, thereby altering the cell membrane permeability to some extent. Moreover, there was a significant increase in intracellular ROS content, cell membrane permeability, key enzyme activi-ty involved in lycopene metabolism, and ROS-related enzyme activity. In conclusion, the alter-nating fre-quency magnetic field can activate a self-protective mechanism that enhances lyco-pene synthesis by modulating intracellular ROS content and cell membrane structure. These find-ings not only deepen our understanding of the impact of magnetic fields on microbial growth and metabolism but also provide valuable insights for developing innovative approaches to en-hance carotenoid fermentation.
Keywords:
magnetic field
; lycopene
; Brassica trispora
; transcriptome
; reactive oxygen species
1. Introduction
The Earth itself harbors a substantial magnetic field. As an environmental factor, the magnetic field (MF) pervades nature and profoundly influences the behavior and physiol-ogy of organisms. Recent years have seen extensive research by domestic and international scientists on the biological magnetic effects of artificial magnetic fields on microorganisms, plants, and animals [1]. The manipulability of microorganisms has led to widespread re-search on the effects of magnetic fields on microbial growth and metabolic product syn-thesis. Magnetic fields exert both specific and non-specific effects on microorganisms, with the specific effects garnering increased research attention in recent years. According to Zhang's [2]study, the yield of yellow and red pigments showed a significant increase at an intensity of 0.4 mT in the frequency-modulated magnetic field, while the increase was smaller and not statistically significant at 0.8 mT and 1.0 mT. Additionally, the pigment yield varied with the duration of exposure. Magnetic fields have the potential to affect both metabolic products and enzyme activity, either enhancing or inhibiting the latter. Magnetic fields have been shown to enhance the activity of black fungus oxidoreductase and peroxidase[3]. Aboneima [4]and colleagues discovered that magnetic fields inhibit the activity of carboxymethylcellulosease (CMCase) in black fungi. Enzyme activity starts to decline after 2 hours of exposure, and nearly half of the activity is lost after 10 hours. Lycopene, a red natural pigment, serves as an intermediate in the synthesis of various ca-rotenoids. It is found in plants, microorganisms, and humans, with the highest concentra-tion occurring in vegetables and fruits like carrots, tomatoes, watermelons, and pome-granates [5]. Additionally, lycopene can be synthesized through microbial fermentation. Ly-copene exhibits antioxidant [6], anti-cancer [7], cardiovascular disease prevention [8], and immune enhancement properties [9], leading to its widespread application in the food, pharmaceutical, health product, and other industries. Research on lycopene has increas-ingly become a focal point in scientific investigations [10]. However, no studies have been reported on the influence of magnetic fields on the metabolism of Brassica trispora, a cru-cial microorganism in lycopene production. This study aims to examine the impact of var-ious magnetic fields on lycopene production by Brassica trispora and preliminarily explore the underlying reasons and mechanisms. This study aims to examine the impact of various magnetic fields on lycopene production by Brassica trispora and preliminarily explore the underlying reasons and mechanisms. This study establishes a theoretical foundation for utilizing magnetic fields to enhance microbial fermentation.
2. Materials and Methods
2.1. Strain and Culture Medium
B. trispora Fly-916 (-) and B. trispora Fly-915 (+).
Seed medium: corn starch 40 g / L, soybean cake powder 23 g / L, KH2PO40.5 g/L,VB1 0.02 g, pH values were adjusted to 6.3 and sterilized at 121℃ for 20 min.
Fermentation medium: corn starch 25 g / L, soybean cake powder 50 g / L, soybean oil 40 g / L, KH2 PO4 1 g / L, MgSO4 0.1 g / L, pH value was adjusted to 6.7 and sterilized at 121℃ for 20 min.
2.2.Main Reagents and Instruments
Ethyl acetate, magnesium sulfate, corn starch, soybean cake powder and potassium dihydrogen phosphate were purchased from Tianjin Yongda Chemical Reagent Co., Ltd. Soybean oil, vitamin B1, potato glucose Agar and potato glucose broth were purchased from Beijing Soleibao Technology Co., Ltd. ROS detection kit, Sod detection kit and cat de-tection kit were purchased from Beijing Suolaibao Technology Co.,Ltd.; PDS ELISA kit was purchased from Shanghai Fuyu Biological Co., Ltd. NEBNext Ultra Directional RNA Li-brary Prep Kit for Illumina kit were purchased from NewEnglandBiolabsInc., Ltd.The PrimeScript TM 1st stand cDNA Synthesis Kit kit was purchased from Bao Nippon Bio-technology Co., Ltd., the spectrophotometer was purchased from Shanghai Yuanxian in-strument Co., Ltd., the electrothermal blast drying box was purchased from Shanghai Yi-heng Scientific instrument Co., Ltd., the high-speed centrifuge was purchased from Shang-hai Anting Science instrument Factory, and the magnetic field and light incubator was purchased from Inductor (Wuxi) Induction Technology Co., Ltd.
2.3.Training Method
Solid culture: strain line inoculation and PDA solid plate, inverted culture at 28 ℃ for 4 days.
Seed culture: under aseptic condition, the spores were scraped off for 4 days with normal saline, and the concentrations of B. trispora (-) and B. trispora (+) spores were con-trolled to 104/mL and 105/mL. 1 mL of positive and negative spore liquid were inoculated in seed medium and cultured at 28 ℃ and 180 r/min for two days.
Fermentation medium: under aseptic condition, the positive and negative seed media growing for two days were inoculated in the fermentation medium at the proportion of 1:8, the inoculum amount was 10% r/min at 28 ℃ for 5 days, and 0.2 g gamma L imidazole (cyclase inhibitor) was added at 48 h of fermentation.
2.4.Magnetic Field Treatment
The fermentation broth was inoculated and subjected to cultivation and fermentation under varying magnetic field intensities (0, 0.2, 0.5, 0.7, 1 mT), exposure periods (1, 2, 3 days), and treatment durations (4, 12, 24, 48, 72 hours) until the 6th day, leading to the isolation of mycelia. The biomass and lycopene production were quantified. The fermented products treated with magnetic fields served as controls, and each experiment was repli-cated three times.
2.5.Analytical Method
2.5.1. Detection of Biomass and Lycopene Yield
The strain underwent a fermentation process for 144 hours, eventually transitioning into the decline phase. Thus, in this experiment, the biomass and lycopene content were assessed after 144 hours of fermentation. Mycelial dry weight was determined using the weighing method. Following fermentation, the fermentation broth underwent centrifuga-tion at 8000 rpm for 5 minutes, followed by three washes with distilled water. Subsequent-ly, it was dried at 80°C until a constant weight was attained. Lycopene content was deter-mined using the colorimetric method. The centrifuged mycelia were vacuum-dried at 40°C, ground to disrupt the cell wall, and subsequently extracted with ethyl acetate until color-less [11]. The extracted solution was filtered, diluted, and its absorbance was measured at a wavelength of 502 nm. Lycopene content was quantified using a standard curve of lyco-pene [12].
2.5.2. Determination of Process Curve
The lycopene content and biomass of Trichoderma sp. were measured every 24 hours throughout the entire fermentation cycle under the optimal magnetic field treatment con-ditions. Process curves were generated for both the untreated magnetic field conditions and the treated magnetic field conditions.
2.5.3. Determination of Cell Permeability of Bacteria
The relative electrical conductivity of mycelial cells served as an indicator of mem-brane permeability. The filtered and washed mycelia were dried using filter paper to elim-inate moisture. Each sample was standardized for mass and placed in a tightly sealed tri-angular flask containing 5 mL of distilled water. The extraction was conducted at a con-stant temperature of 20°C in an incubator for 24 hours with intermittent shaking. The electrical conductivity of the extraction solution was determined using a digital conduc-tivity meter. Subsequently, the samples were immersed in a boiling water bath for 30 minutes, cooled to room temperature, and the total electrical conductivity was measured. The relative electrical conductivity (%) =(extract conductivity/total conductivity)×100.
2.5.5. Scanning Electron Microscope (SEM) Observation
The sample processing method described by Cheng et al. [13]was adopted. A volume of 1 mL of fermentation broth was extracted and transferred to a 2 mL centrifuge tube. The mycelia were then sedimented by centrifugation at 8000 rpm/min, fixed with a 2.5% glu-taraldehyde solution, and stored at 4°C for 24 hours. The mycelial precipitate was then washed three times with PBS buffer and subjected to gradient dehydration using ethanol. Two consecutive replacements were performed using ethyl acetate, with each replacement lasting 20 minutes. Lastly, the samples were dried at room temperature for 2 days, coated with a layer of gold using sputter-coating, and observed using scanning electron micros-copy.
2.5.6. Transcriptome Analysis
The total RNA of mycelium samples was extracted and the library was established by NEBNext Ultra II RNA Library Prep Kit for Illumina (New England Biolabs Inc;Ipswich,Massachusetts,USA) kit. The transcriptional library of the samples was se-quenced by Illumina sequencing platform. The differentially expressed genes were ana-lyzed by DESeq (v1.38.3) software, and the conditions for screening differentially ex-pressed genes were as follows:multiple of differential expression |log2FoldChange|>1,significant Pmurvaluevaluation<0.05. Through the functional analy-sis of GO and KEGG database, the differentially expressed genes were annotated and en-riched in metabolic pathway.
2.5.7. RT-qPCR
Six related genes were screened for real-time fluorescence quantitative PCR to verify the transcriptome data. The primers used are shown in Table 1. cDNA was synthesized by kit (PrimeScript TM 1st stand cDNA Synthesis Kit). RT-qPCR reaction system (20 μL): 2 × SYBR real-time PCR premixture 10 μL primers specific upstream and downstream (10 μmol/L), each 0.4 μL primer free ddH2O 8 μL. The reaction conditions were 95 ℃, 5 min, 95 ℃, 15 min, 60 ℃, 30 min, 40 cycles. With tef1 as the internal reference gene [14], the rela-tive mRNA expression of the target gene was calculated by 2-Ct analysis method [15].
2.5.8. Data Analysis
The experimental data were counted by SPSS 20.0. The drawing was made by Origin 2018 software. Microbiological analyses were performed in, at minimum, triplicate.All data are expressed as mean ± standard deviation.
3. Results
3.1. Magnetic Field Intensity
Previous studies have demonstrated that magnetic fields exert diverse effects on mi-croorganisms, encompassing magnetic field intensity, treatment timing, and exposure du-ration [16]. Consequently, the influence of magnetic fields on lycopene production in Spo-robolomyces sp. was examined under the specified magnetic field conditions. Liquid fer-mentation experiments were performed on Sporobolomyces sp. within a magnetic field range of 0 to 1 mT. Lycopene production was quantified after 144 hours of fermentation, as depicted in Figure 1. Figure 1 reveals a notable enhancement in lycopene production at magnetic field intensities of 0.2, 0.5, and 0.7 mT when compared to the control group. Conversely, lycopene production decreased at a magnetic field intensity of 1 mT, corrobo-rating the findings of Jialan et al. [17]. Lycopene production reached 870.2 mg/L at 0.4 mT, signifying an 86.5% increase compared to the control group's lycopene yield of 466.4 mg/L. Consequently, the optimal magnetic field intensity for treatment was identified as 0.5 mT.
3.2. Magnetic Field Processing Time
The impact of magnetic field treatment duration on lycopene production necessitates the investigation of the optimal duration. The fermentation broth was exposed to a 0.5 mT magnetic field for varying durations (4, 12, 24, 48, and 72 hours) on days 1, 2, and 3. Sub-sequently, the fermentation was continued for 144 hours, during which lycopene produc-tion and biomass were measured (Table 1). Table 1 reveals a minor, albeit insignificant, in-hibition in biomass when compared to the control group. Lycopene production remained nearly unchanged upon magnetic field treatment on the first day of fermentation, while a substantial increase was observed on the second and third days. This can be attributed to the primary production of lycopene during the microorganisms' stable growth phase [12]. The highest lycopene production was achieved after 48 hours of exposure, followed by a gradual decline in production with longer treatment durations when compared to the con-trol group. This decline in production may be attributed to the prolonged exposure to the magnetic field, which inhibits normal microbial metabolism [18]. On the second day of fermentation, lycopene production reached 1473.16 mg/L after 48 hours of magnetic field exposure, resulting in a 216.1% increase compared to the control group's production of 466.46 mg/L. On the third day of fermentation, lycopene production reached 911.76 mg/L after 48 hours of magnetic field exposure, resulting in a 95.6% increase. Thus, the optimal magnetic field treatment duration was identified as 48 hours of exposure on the second day of fermentation.

Table 2.
effects of magnetic field intervention period and exposure time on biomass and lycopene yield.
Table 2.
effects of magnetic field intervention period and exposure time on biomass and lycopene yield.
| Exposure period(d) | Exposure time(h) | Dry cell weight(g/L) | Lycopene yield(mg/L) |
|---|---|---|---|
| Control | 0 | 63.3±3.40 | 466.46±23.2 |
| 1 | 4 12 24 48 72 |
60.4±4.30 58.3±3.50 56.1±3.80 57.2±4.20 58.3±5.30 |
411.36±22.28e 467.45±16.52b 437.63±18.37c 492.17±14.23a 421.32±21.41d |
| 2 | 4 12 24 48 72 |
59.3±2.70 58.1±3.40 56.3±4.50 58.1±4.60 60.2±3.20 |
575.25±19.48e 723.26±25.25d 759.53±21.37c 1473.16±29.42a 892.13±19.64b |
| 3 | 4 12 24 48 72 |
59.3±2.80 61.2±2.40 58.5±5.20 60.2±3.20 57.1±4.60 |
598.26±18.33e 672.44±19.22d 692.48±15.53c 911.76±18.43a 742.21±17.53b |
*The lowercase alphabet with different right shoulder in the same column showed significant differ-ence (p<0.05).
3.3. Process Metabolic Curve
Figure 3 presents the metabolic profiles of Sporobolomyces sp. during fermentation under the optimized magnetic field treatment conditions, which involved exposing the culture to a 0.5 mT magnetic field for 48 hours on the second day of fermentation. Both the control group and the treatment group displayed comparable patterns in biomass and ly-copene production, demonstrating a logarithmic growth phase from 72 to 120 hours, fol-lowed by a subsequent stabilization. The magnetic field exerted a significant positive effect on lycopene production while displaying a minor inhibitory effect on microbial growth, albeit with negligible consequences.
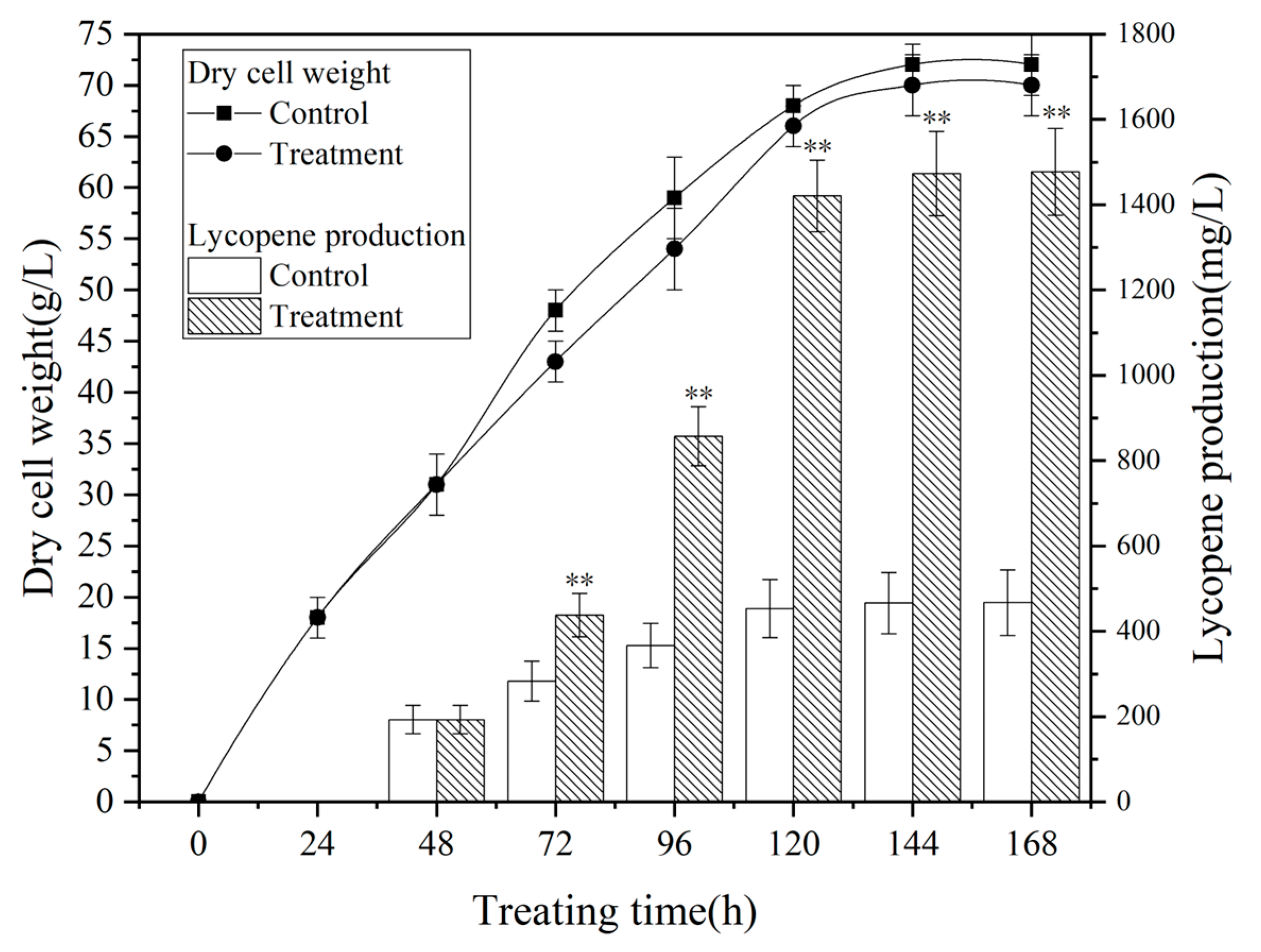
Figure 2.
fermentation curve of Brassica trispora under the optimal magnetic field conditions (exposure to 0.5mT magnetic field for 48 hours on the second day of fermentation,p<0.05).
Figure 2.
fermentation curve of Brassica trispora under the optimal magnetic field conditions (exposure to 0.5mT magnetic field for 48 hours on the second day of fermentation,p<0.05).
3.4. Transcriptome Analysis of the Effect of Magnetic Field on Bacteriae
3.4.1. Differentially Expressed Genes
To examine the impact of magnetic fields on the global transcriptome of Streptomyces brasiliensis during fermentation, we selected the previously mentioned optimal magnetic field conditions and compared them to the control conditions. The bacterial cultures un-derwent a 120-hour fermentation period, and their transcriptomes were subsequently se-quenced and analyzed. The results confirmed the reliability of the sequencing by demon-strating an accuracy of base identification exceeding 94%, as indicated by Q30 values. Dif-ferential gene expression analysis was performed using DESeq, with the criteria for select-ing differentially expressed genes set as |log2FoldChange| > 1 and a significant P-value < 0.05. Figure 5 displays the volcano plot, which visualizes the differentially expressed genes between the two conditions. This transcriptome sequencing identified a total of 581 dif-ferentially expressed genes, including 405 significantly upregulated genes and 176 signifi-cantly downregulated genes.
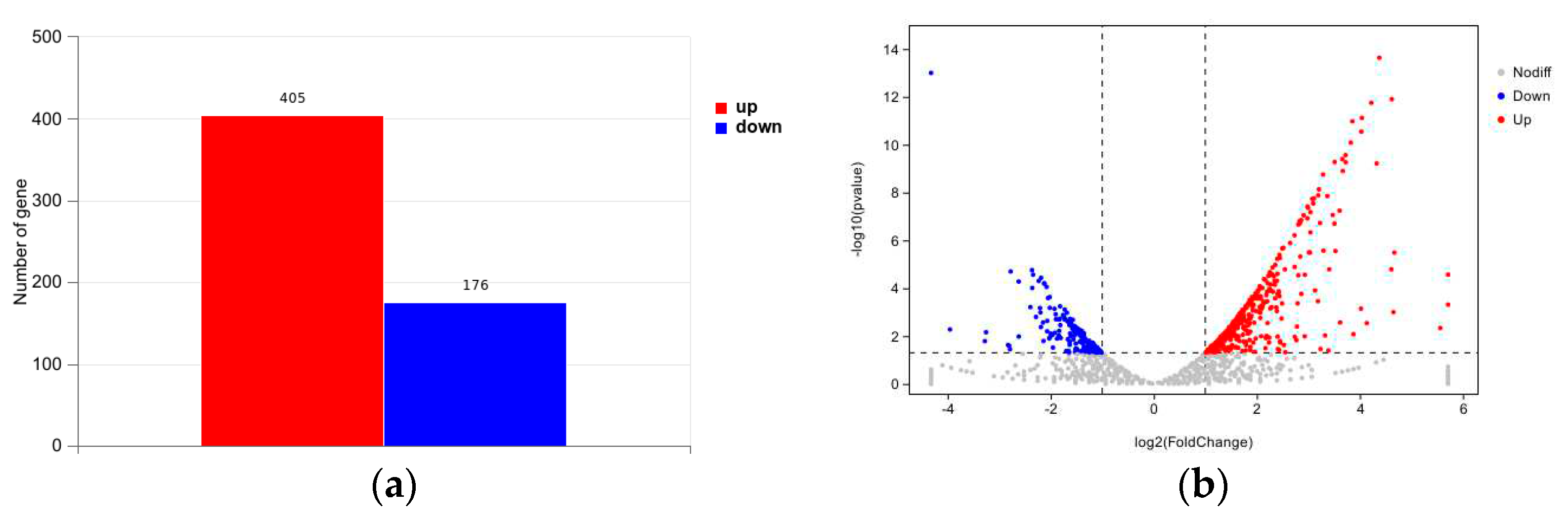
Figure 3.
Comparative transcriptomic analysis of magnetic field and control group. (a) Differential expression anal-ysis of magnetic field group in comparison to control group, when growth on 120h. (b) Log2 ratio of magnetic field group/control group vs. -log10(pvalue) in either strain. Red dots represent upregulated genes (log2 FoldChange≥ 1, and DESeq P-adj value < 0.05), and blue dots down-regulated genes (log2 FoldChange ≤ -1, and DESeq P-adj value < 0.05). Gray dots represent genes whose changes in expression are not statistically.
Figure 3.
Comparative transcriptomic analysis of magnetic field and control group. (a) Differential expression anal-ysis of magnetic field group in comparison to control group, when growth on 120h. (b) Log2 ratio of magnetic field group/control group vs. -log10(pvalue) in either strain. Red dots represent upregulated genes (log2 FoldChange≥ 1, and DESeq P-adj value < 0.05), and blue dots down-regulated genes (log2 FoldChange ≤ -1, and DESeq P-adj value < 0.05). Gray dots represent genes whose changes in expression are not statistically.
3.4.2. Differentially Expressed Genes
The differentially expressed genes underwent Gene Ontology (GO) enrichment analy-sis, dividing them into three primary categories: Molecular Function (MF), Biological Pro-cess (BP), and Cellular Component (CC). Following annotation of the differentially ex-pressed genes, the top 20 significantly enriched GO terms are depicted in Figure 7. In the Molecular Function category, the most prominent and representative GO terms were cata-lytic activity (GO:0003824) and oxidoreductase activity (GO:0016491). The representative GO terms in the Biological Process category were carbohydrate metabolic process (GO:0005975) and G protein-coupled receptor signaling pathway (GO:0007186). Signifi-cant enrichment of GO terms in the Cellular Component category was detected in mem-brane-related locations, such as integral component of membrane (GO:0016021), intrinsic component of membrane (GO:0031224), and membrane (GO:0016020).
Significant enrichment of GO terms in the Cellular Component category was detected in membrane-related locations, such as integral component of membrane (GO:0016021), intrinsic component of membrane (GO:0031224), and membrane (GO:0016020). Figure 7 shows that membrane protein-related GO terms, such as G protein-coupled receptor sig-naling pathway (GO:0007186) and ABC-type xenobiotic transporter activity (GO:0008559), were significantly enriched. This suggests that low-frequency magnetic fields might impact transmembrane proteins, resulting in elevated transcription levels and modifications in intracellular calcium and sodium ion concentrations, thereby fostering cellular metabolism [19]. Additional studies have shown that low-frequency magnetic fields can modify mem-brane permeability [20], leading to changes in ion concentrations inside and outside the membrane, and thereby enhancing cellular metabolism [21]. The upregulation of cell membrane permeability and membrane protein functionality could serve as a compensa-tory response to magnetic fields, safeguarding the cell [22]. From the GO enrichment re-sults, it is evident that the effects of magnetic fields on organisms mainly include oxida-tive-reductive enzymes, membrane-associated functions, and membrane transporters, ul-timately impacting cellular growth and metabolism.
We performed KEGG pathway enrichment analysis on differentially expressed genes to investigate the metabolic pathways of Trichoderma bisporum in response to magnetic fields. Figure 7 illustrates the top 20 enriched pathways, encompassing cellular processes, metabolism, environmental information processing, and organismal systems. Most of the differentially expressed genes are enriched in cellular engineering and metabolism. The peroxisome pathway is the main enrichment in cellular processes, while metabolism is en-riched in the biosynthesis pathway of unsaturated fatty acids, citric acid cycle (TCA cycle), and sulfur metabolism pathway. The MAPK signaling pathway is primarily enriched in environmental information processing, and the lifespan regulation pathway is dominant in organismal systems. These enrichment pathways guide further research on the response mechanism of Trichoderma bisporum to magnetic fields and validate the results of GO en-richment analysis.
The redox enzyme metabolism pathway exhibits significant enrichment among dif-ferentially expressed genes. The upregulation of enzymes involved in ROS metabolism, in-cluding superoxide dismutase (gene_8655), catalase (gene_5118), and peroxidase (gene_6907), suggests that magnetic fields can modulate intracellular and extracellular ROS levels. Consistent with the findings of S. Roy et al. [23], low-frequency magnetic fields can impact biological systems by modulating ROS or free radicals. Furthermore, peroxi-somes are crucial cellular organelles involved in intracellular ROS production [24], as xanthine oxidase in the peroxisome matrix or membrane can generate O2- [25]. Moreover, lycopene acts as an antioxidant, and cells synthesize lycopene as a protective response when the cellular ROS levels rise [26], potentially explaining the high lycopene production. Wang et al. [27] found that ROS can markedly upregulate the transcription of key lycopene metabolic enzymes, namely carotenoid synthase (carRA) and lycopene dehydrogenase (carB) in Trichoderma bisporum, leading to enhanced lycopene production. Importantly, the differentially expressed genes exhibit significant enrichment in the sulfur metabolism pathway, potentially due to the high concentration of magnetic sensor proteins in this pathway [28].
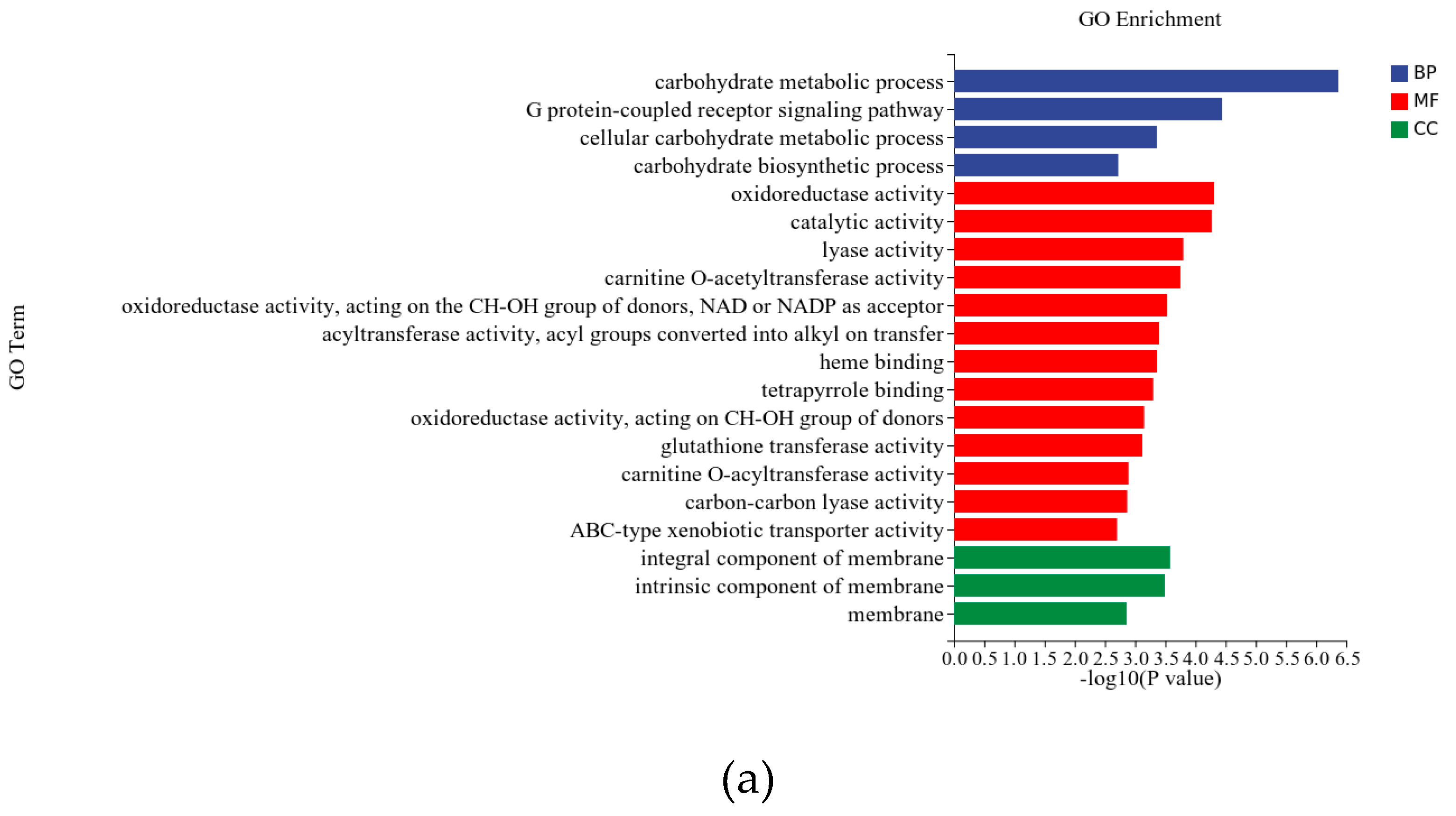
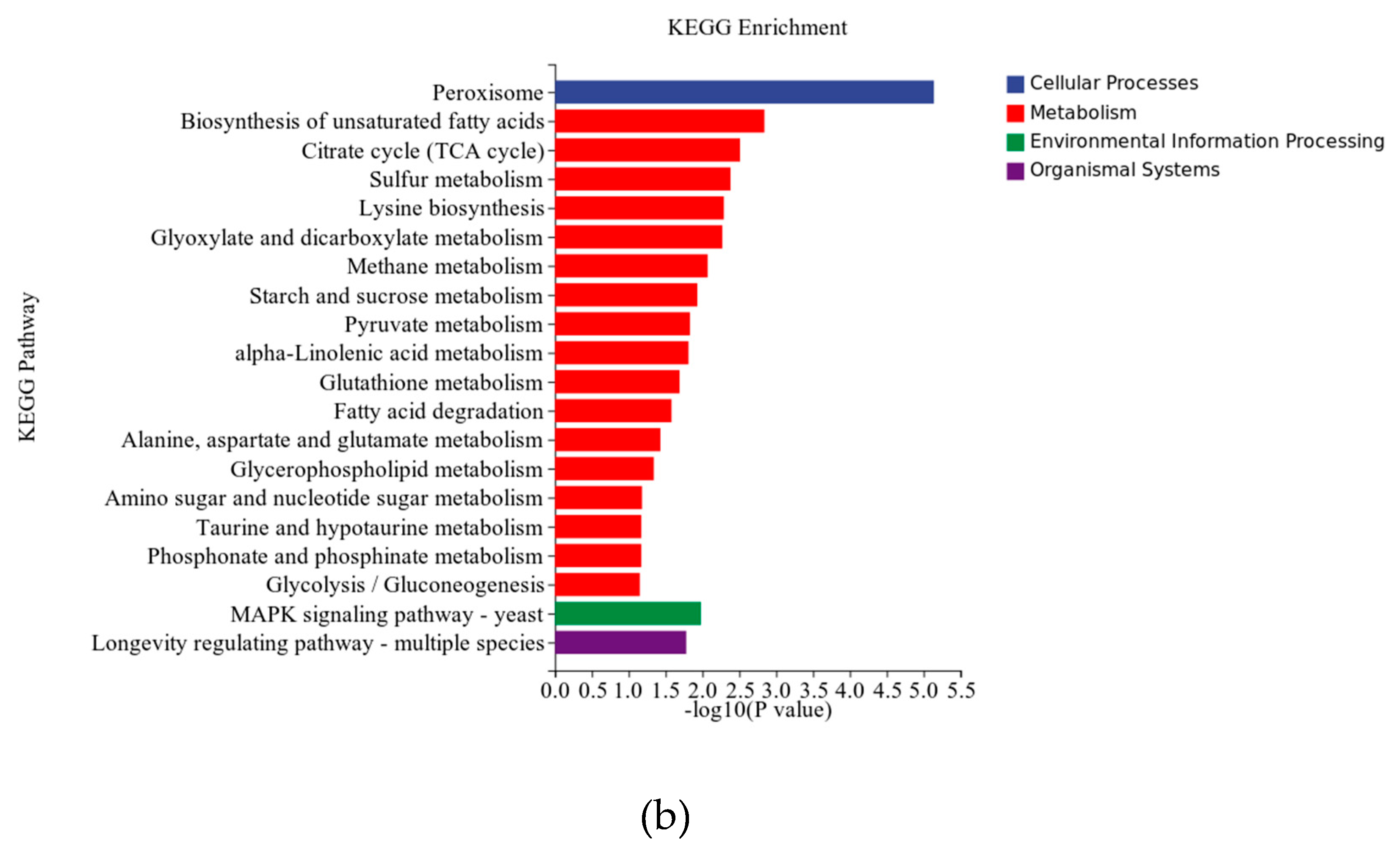
Figure 4.
GO and KEGG pathway enrichment taxonomic map of differentially expressed genes. (a) GO enrichment analysis (b) KEGG enrichment analysis.
Figure 4.
GO and KEGG pathway enrichment taxonomic map of differentially expressed genes. (a) GO enrichment analysis (b) KEGG enrichment analysis.
3.5. RT-qPCR Analysis
To further analyze the transcriptional activity of the key differentially expressed genes (carRA, carB, sod, cat, iscA, elo3), we utilized real-time quantitative PCR (RT-qPCR) to measure their expression levels. Lycopene synthesis in Trichoderma bisporum predomi-nantly takes place via the mevalonate (MVA) pathway, with lycopene synthase and lyco-pene cyclase (carRA, carB) serving as key enzymes [29]. Hence, carRA and carB were chosen for analysis. Furthermore, Figure 6 illustrates the influence of the magnetic field on the key enzyme genes involved in lycopene metabolism. To validate the elevated intracellular oxi-dative levels, we selected superoxide dismutase and catalase (sod, cat), which are enzymes involved in reactive oxygen species (ROS) metabolism. We chose the elo3 gene, which is associated with cell membrane functions, due to its significant upregulation, which we confirmed through RT-qPCR to validate the transcriptomic results. Notably, Qin et al. [28]dis-covered that the iron-sulfur cluster assembly protein (iscA) functions as a biomagnetic sensor in response to magnetic fields. Thus, we selected the iscA gene for analysis to exam-ine the effects of the magnetic field on the fungal organism. The RT-qPCR results showed consistency with the RNA-Seq data, displaying similar trends and confirming the reliabil-ity of the transcriptomic findings in this study.
The RT-qPCR results, presented in Table 3, indicated substantial upregulation of key enzymes involved in lycopene metabolism (carRA, carB), ROS-related genes (sod, cat), and the cell membrane-related gene elo3. This further supports the notion that the magnetic field treatment enhances lycopene production by affecting ROS levels and cell membrane integrity. During the magnetic field treatment, the expression of the iscA gene, which en-codes the iron-sulfur cluster assembly protein, was upregulated, potentially because it acts as a magnetic receptor protein (MagR) responsive to magnetic fields, as reported by [28]. IscA (MagR) can induce membrane depolarization and action potentials through external magnetic field exposure, resulting in intracellular calcium influx and subsequent activa-tion of the organism's magnetic field response [30].
3.6. Mycelial Morphologye
We used scanning electron microscopy (SEM) to observe the effects of magnetic fields on the microstructure of Trichoderma bisporum mycelia. Figure 4a illustrates the myceli-al surface, which appeared smooth, flat, and exhibited minimal bending. The mycelial sur-face in the magnetic field group (Figure 4b) displayed roughness, looseness, increased folding, and a few micropores. This can be attributed to the elevated levels of intracellular reactive oxygen species (ROS), which induce changes in the structure and permeability of the cell membrane. This finding aligns with the research conducted by Guo Li et al. [31], which documented increased folding on the surface of Grifola frondosa mycelia and an overall more porous mycelial structure. Likewise, Voychuk et al. [32] observed an increase in cell wall invaginations in Saccharomyces cerevisiae yeast cells exposed to magnetic fields, resulting in modifications to internal metabolic synthesis processes. This could be attributed to the ability of magnetic fields to induce alterations in the local structure of the cell membrane, thereby facilitating the transport of ions and molecules [33]. Microscopic examination of the mycelial color, as depicted in Figure 4c,d, demonstrated that the mycelia in the magnetic field-treated group displayed a more intense red color in compar-ison to the control group. This further supports the notion that magnetic fields enhance lycopene synthesis.

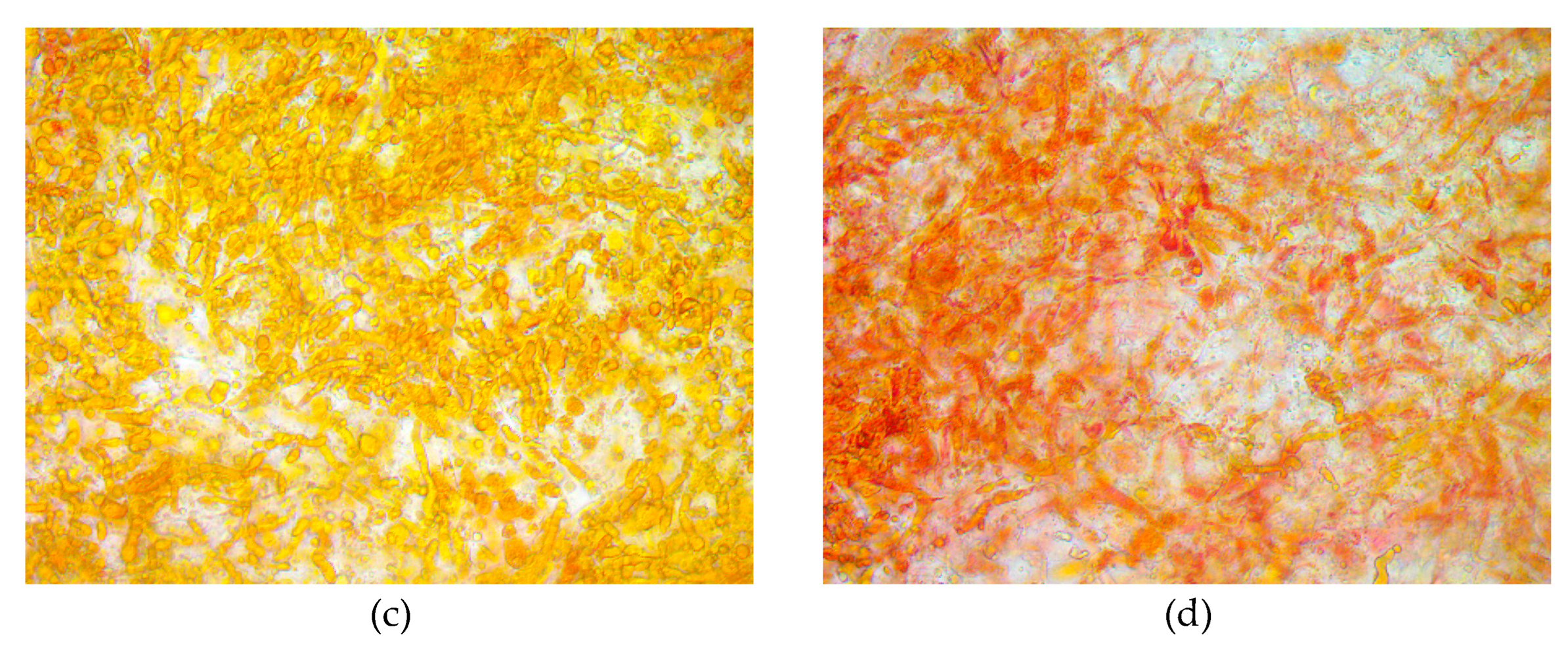
Figure 5.
Microstructure of Brassica trispora under scanning electron microscope (SEM) and microscope. (a) (c)blank group;(b) (d)magnetic field treatment for 48 hours.
Figure 5.
Microstructure of Brassica trispora under scanning electron microscope (SEM) and microscope. (a) (c)blank group;(b) (d)magnetic field treatment for 48 hours.
3.7. Effects of Magnetic Field Treatment on Intracellular Reactive Oxygen Species, Cell Membrane Permeability and Activities of Key Enzymes
To further validate the transcriptome analysis results, we measured the activities of key enzymes, cell membrane permeability, and intracellular levels of reactive oxygen spe-cies (ROS), as depicted in Figure 9. Magnetic field treatment significantly increased intra-cellular ROS content compared to the control group. Furthermore, the activities of ROS metabolism-related enzymes, including superoxide dismutase (SOD), catalase (CAT), and phytoene desaturase (PDS), the key enzyme in lycopene metabolism, were significantly enhanced. These findings suggest that the elevated intracellular ROS levels stimulate lyco-pene production, contributing to the maintenance of intracellular redox balance. Quiles-Rosillo et al. [34] made similar observations, reporting that ROS markedly upregu-lated the transcription levels of carB and carRA, resulting in enhanced carotenoid accu-mulation. Wang et al. [27] also demonstrated that the supplementation of H2O2 increased lycopene production in Trichoderma bisporum. These findings align with the study con-ducted by Qian et al. [35], which revealed that low-frequency alternating magnetic fields can enhance Monascus pigment synthesis by modulating intracellular ROS levels.
Figure 9(2) demonstrates a significant increase in cell membrane permeability of the mycelia under magnetic field treatment when compared to the control group. This implies that magnetic field treatment has the ability to modify cell membrane permeability, ulti-mately enhancing cellular metabolism [36]. In a similar vein, Liu et al. [16] discovered that magnetic field stimulation induced changes in membrane fluidity and permeability of Ganoderma lucidum mycelia, influencing the exchange of substances between the intra-cellular and extracellular environments and promoting cellular metabolism. Collectively, these findings indicate that magnetic field treatment primarily amplifies intracellular ROS levels and cell membrane permeability, providing further confirmation of the results ob-tained from transcriptome analysis and scanning electron microscopy.
Third bullet.
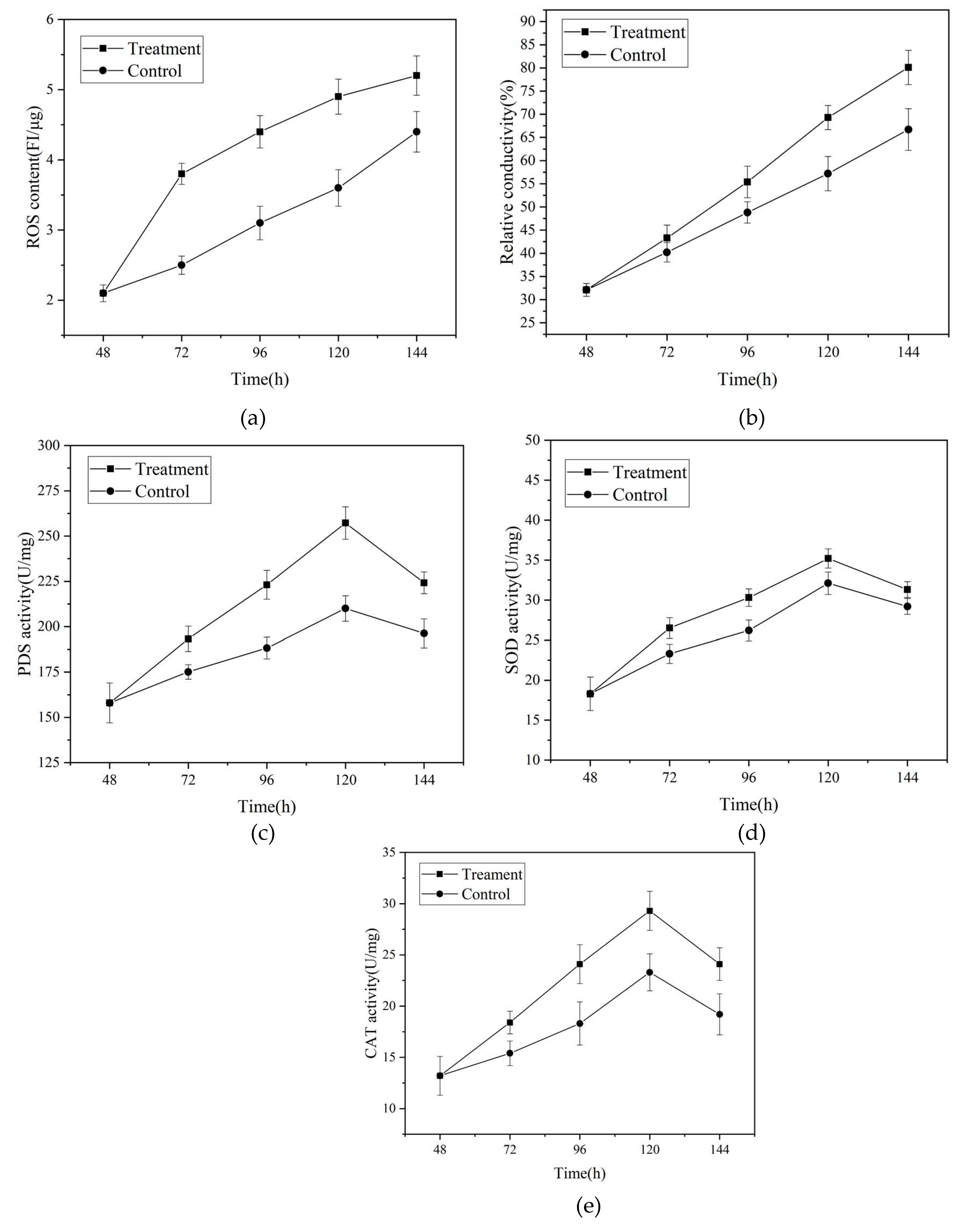
Figure 6.
The effect of magnetic fields on crucial enzymes, ROS-related enzyme activity, cellular membrane permeability, and intracellular ROS in the lycopene synthesis pathway of Brassica trispora. (a) Intracellular ROS content;(b) cell membrane permeability;(c) PDS activity;(d) SOD activity;(e) CAT activity.
Figure 6.
The effect of magnetic fields on crucial enzymes, ROS-related enzyme activity, cellular membrane permeability, and intracellular ROS in the lycopene synthesis pathway of Brassica trispora. (a) Intracellular ROS content;(b) cell membrane permeability;(c) PDS activity;(d) SOD activity;(e) CAT activity.
4. Conclusions
The results suggest that low-frequency magnetic fields can boost lycopene production by regulating intracellular ROS and modulating gene expression associated with cell membrane transport proteins. This phenomenon was investigated using scanning electron microscopy, transcriptomics, and real-time quantitative PCR analyses. A significant in-crease in lycopene production was observed after 48 hours of treatment with a 0.5mT low-frequency magnetic field during a 2-day fermentation. Transcriptomic analysis showed upregulation of genes primarily associated with oxidation-reduction enzymes and membrane-related processes. The reliability of the transcriptomic data was confirmed through scanning electron microscopy and real-time quantitative PCR results. Magnetic field exposure significantly enhanced the activities of ROS-related enzymes, key enzymes in lycopene synthesis, intracellular ROS levels, and cell membrane permeability, providing further validation of the transcriptomic findings. These findings offer theoretical insights into carotenoid fermentation under magnetic field conditions. Furthermore, the response and underlying mechanism of the iron-sulfur cluster assembly protein (IscA/MagR) in Trichoderma bisporum to magnetic fields remain unclear. The response of magnetic sens-ing proteins to magnetic fields has a specific effect on microorganisms, emphasizing the need to investigate their mechanisms to understand the impact of magnetic fields on mi-croorganisms [37]. Therefore, further research is needed to elucidate the role and mecha-nism of action of magnetic sensing proteins.
Supplementary Materials
Figure S1: Lycopene standard curve; Table S1: effect of magnetic field intensity on lycopene yield; Table S2: fermentation curve of Brassica trispora under the optimal magnetic field conditions; Table S3: The effect of magnetic fields on ROS-related enzyme activity in the lycopene synthesis pathway of Brassica trispora; Table S4: The effect of magnetic fields on crucial enzymes activity in the lycopene synthesis pathway of Brassica trispora; Table S5: The effect of magnetic fields on intracellular ROS in the lycopene synthesis pathway of Brassica trispora; Table S6: The effect of magnetic fields on cellular membrane permeability in the lycopene synthesis pathway of Brassica trispora.
Author Contributions
J.H. and H.W. contributed equally to this paper; J.H. and H.W. performed the main ex-periments and prepared figures and tables; J.L. contributed the experiment materials and performed some experiments; J.H., H.W. and D.W. wrote the manuscript text; J.L. and M.Y. designed the experiments. All authors reviewed the manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by Natural Science Foundation of Hebei Province.(Grant No.C2021208019)
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
The anthors gratefully acknowledge Jinlong Liu from Hebei University of Science and Technology (Shijiazhang, China) for providing suggestions for this experimental study, and gratefully acknowledge Maohua Yang from Institute of process engineering, Chinese Academy of Sciences(Beijing, China) for providing suggestions for this experimental study.
Conflicts of Interest
Not applicable.
References
- Zhang, X.; Yarema, K.; Xu, A. Impact of Static Magnetic Field (SMF) on Microorganisms, Plants and Animals. Springer Singapore 2017, 10.1007/978-981-10-3579–1, 133–172.
- Zhang, J.; Zeng, D.; Xu, C.; Gao, M. Effect of low-frequency magnetic field on formation of pigments of Monascus purpureus. European Food Research and Technology 2014, 240, 577–582. [Google Scholar] [CrossRef]
- Makarov, I.O.K. Effect of Low-Frequency Pulsed Magnetic Field and Low-Level Laser Radiation on Oxidoreductase Activity and Growth of FungiActive Destructors of Polymer Materials. Microbiology 2019, 88. [Google Scholar] [CrossRef]
- Aboneima, S.; El-Metwally, M.M. Effect of extremely low frequency magnetic field in growth, CMCase, electric conductivity and DNA of Aspergillus niger. National Information and Documentation Center (NIDOC), Academy of Scientific Research and Technology (ASRT) 2020.
- Tagging, A.N.I.M. Re: Tomato and Lycopene and Multiple Health Outcomes: Umbrella Review. The Journal of Urology 2022, 207. [Google Scholar]
- Khan, U.M.; Sevindik, M.; Zarrabi, A.; Nami, M.; Ozdemir, B.; Kaplan, D.N.; Selamoglu, Z.; Hasan, M.; Kumar, M.; Alshehri, M.M. Lycopene: Food Sources, Biological Activities, and Human Health Benefits. Hindawi Limited, 2021. [Google Scholar]
- Nigussie, A.; Tura, A.K.; Sisay, M.; Hagos, B.; Motbaynor, B. Anti-Cancer Effects of Lycopene in Animal Models of Hepatocellular Carcinoma: A Systematic Review and Meta-Analysis. Frontiers in pharmacology 2020, 11. [Google Scholar] [CrossRef]
- Cheng, H.M.; Koutsidis, G.; Lodge, J.K.; Ashor, A.; Siervo, M.; Lara, J. Tomato and lycopene supplementation and cardiovascular risk factors: A systematic review and meta-analysis. Atherosclerosis 2017, 257, 100–108. [Google Scholar] [CrossRef] [PubMed]
- Crowe-White, K.M.; Phillips, T.A.; Ellis, A.C. Lycopene and cognitive function. 2019.
- Srivastava, S.; Srivastava, A.K. Lycopene; chemistry, biosynthesis, metabolism and degradation under various abiotic parameters. Journal of Food Science & Technology 2015, 52, 41–53. [Google Scholar] [CrossRef]
- Qiang, W.; Ling-Ran, F.; Luo, W.; Han-Guang, L.; Lin, W.; Ya, Z.; Xiao-Bin, Y. Mutation Breeding of Lycopene-Producing Strain Blakeslea Trispora by a Novel Atmospheric and Room Temperature Plasma (ARTP). Applied Biochemistry and Biotechnology 2014. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Feng, L.R.; Luo, W.; Li, H.G.; Zhou, Y.; Yu, X.B. Effect of Inoculation Process on Lycopene Production by Blakeslea trispora in a Stirred-Tank Reactor. Applied Biochemistry & Biotechnology 2015, 175, 770–779. [Google Scholar] [CrossRef]
- 13. Sun; Cheng; Mueller; Erich; Meffert; Matthias; Gerthsen; Dagmar On the Progress of Scanning Transmission Electron Microscopy (STEM) Imaging in a Scanning Electron Microscope. Microscopy & Microanalysis the Official Journal of Microscopy Society of America Microbeam Analysis Society Microscopical Society of Canada 2018.
- André D., Schmidt; Heinekamp, T.; Matuschek, M.; Liebmann, B.; Bollschweiler, C.; Brakhage, A.A. Analysis of mating-dependent transcription of Blakeslea trispora carotenoid biosynthesis genes carB and carRA by quantitative real-time PCR. Applied Microbiology & Biotechnology 2005, 67, 549–555. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. 2013.
- Liu, D.; Zhu, L.; Guo, Y.; Zhao, Y.; Betchem, G.; Yolandani, Y.; Ma, H. Enhancing submerged fermentation of Antrodia camphorata by low-frequency alternating magnetic field. Innovative Food Science & Emerging Technologies 2023, 86. [Google Scholar] [CrossRef]
- Xu; Cui; Zhang; Jialan; Gao; Mengxiang; Zeng; Dongjie Effect of low-frequency magnetic field on formation of pigments of Monascus purpureus. Zeitschrift Fur Lebensmittel Untersuchung Und Forschung A 2015.
- Masood, S.; Saleem, I.; Smith, D.; Chu, W.K. Growth Pattern of Magnetic Field Treated Bacteria 2018.
- Falone, S.; Grossi, M.R.; Cinque, B.; Barbara D’Angelo; Tettamanti, E. ; Cimini, A.; Ilio, C.D.; Amicarelli, F. Fifty hertz extremely low-frequency electromagnetic field causes changes in redox and differentiative status in neuroblastoma cells. International Journal of Biochemistry & Cell Biology 2007, 39, 2093–2106. [Google Scholar] [CrossRef]
- Mermall; V Unconventional Myosins in Cell Movement, Membrane Traffic, and Signal Transduction. Science 1998, 279, 527–533. [CrossRef] [PubMed]
- Klotz; J; L CELL BIOLOGY SYMPOSIUM: Membrane trafficking and signal transduction. J. Anim. Sci. 2017, 95, 2183–2184. [CrossRef]
- Zablotskii, V.; Polyakova, T.; Dejneka, A. Modulation of the Cell Membrane Potential and Intracellular Protein Transport by High Magnetic Fields. Bioelectromagnetics 2020. [Google Scholar] [CrossRef] [PubMed]
- Roy, S.; Noda, Y.; Eckert, V.; Traber, M.G.; Packer, L. The phorbol 12-myristate 13- acetate (PMA)-induced oxidative burst in rat peritoneal neutrophils is increased by a 0.1 mT (60 Hz) magnetic field. FEBS Letters 1995, 376, 164–166. [Google Scholar] [CrossRef] [PubMed]
- Dansen, T.B.; Wirtz, K.W.A. The Peroxisome in Oxidative Stress. IUBMB Life 2001. [Google Scholar] [CrossRef] [PubMed]
- Bonekamp, N.A.; Vlkl, A.; Fahimi, H.D.; Schrader, M. Reactive oxygen species and peroxisomes: Struggling for balance. Biofactors 2009, 35, 346–355. [Google Scholar] [CrossRef] [PubMed]
- Nanou, K.; Roukas, T.; Papadakis, E. Oxidative stress and morphological changes in Blakeslea trispora induced by enhanced aeration during carotene production in a bubble column reactor. Biochem. Eng. J. 2011, 54, 172–177. [Google Scholar] [CrossRef]
- Wang, H.B.; Luo, J.; Huang, X.Y.; Lu, M.B.; Yu, L.J. Oxidative stress response of Blakeslea trispora induced by HO during β-carotene biosynthesis. Journal of Industrial Microbiology & Biotechnology 2014, 41, 555–561. [Google Scholar] [CrossRef]
- Qin, S.; Yin, H.; Yang, C.; Xie, C. A magnetic protein biocompass. 2015.
- Zhang, Y.; Navarro, E.; Cánovas-Márquez, J.T.; Almagro, L.; Chen, H.; Chen, Y.Q.; Zhang, H.; Torres-Martínez, S.; Chen, W.; Garre, V. A new regulatory mechanism controlling carotenogenesis in the fungus Mucor circinelloides as a target to generate β-carotene over-producing strains by genetic engineering. Microbial Cell Factories 2016, 15, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Long, X.; Ye, J.; Zhao, D.; Zhang, S.J.; Sciences, S.O.L.; University, T.; Medicine, S.O.; Brain, I.G.I.F.; Sciences, T.P.C.F.L. Magnetogenetics: remote non-invasive magnetic activation of neuronal activity with a magnetoreceptor. Sci. Bull. 2015. [Google Scholar] [CrossRef] [PubMed]
- Guo, L.N.; Li, X.Y.; Zhang, X.Y.; Ma, H.L. Effect of low-intensity magnetic field on the growth and metabolite of Grifola frondosa in submerged fermentation and its possible mechanisms. Food Res. Int. 2022, 159. [Google Scholar] [CrossRef] [PubMed]
- Voychuk, S.I.; Gromozova, E.N.; Lytvyn, P.M.; Podgorsky, V.S. Changes of Surface Properties of Yeast Cell Wall Under Exposure of Electromagnetic Field (40.68 MHz) and Action of Nystatin. Environmentalist 2005, 25, 139–144. [Google Scholar] [CrossRef]
- Weaver, J.C. Electroporation: a general phenomenon for manipulating cells and tissues. Journal of Cellular Biochemistry 1993, 51, 426–435. [Google Scholar] [CrossRef]
- María, D. Quiles-Rosillo; Rosa M. Ruiz-Vázquez; Santiago Torres-Martínez; Garre, V. Light induction of the carotenoid biosynthesis pathway in Blakeslea trispora. Fungal Genetics & Biology 2005, 42, 141–153. [Google Scholar] [CrossRef]
- Liao, Q.; Liu, Y.; Zhang, J.; Li, L.; Gao, M. A low-frequency magnetic Field regulates Monascus pigments synthesis via reactive oxygen species in M. purpureus. Process Biochem. 2019, 86, 16–24. [Google Scholar] [CrossRef]
- Ikehara, T.; Yamaguchi, H.; Hosokawa, K.; Miyamoto, H.; Aizawa, K. Effects of ELF magnetic field on membrane protein structure of living HeLa cells studied by Fourier transform infrared spectroscopy. Bioelectromagnetics 2010, 24. [Google Scholar] [CrossRef] [PubMed]
- Binhi, V.N.; Rubin, A.B. Theoretical Concepts in Magnetobiology after 40 Years of Research. Cells 2022, 11, 274. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
effect of magnetic field intensity on lycopene yield. Different lowercase letters indicate signif-icant differences (p<0.05).
Figure 1.
effect of magnetic field intensity on lycopene yield. Different lowercase letters indicate signif-icant differences (p<0.05).
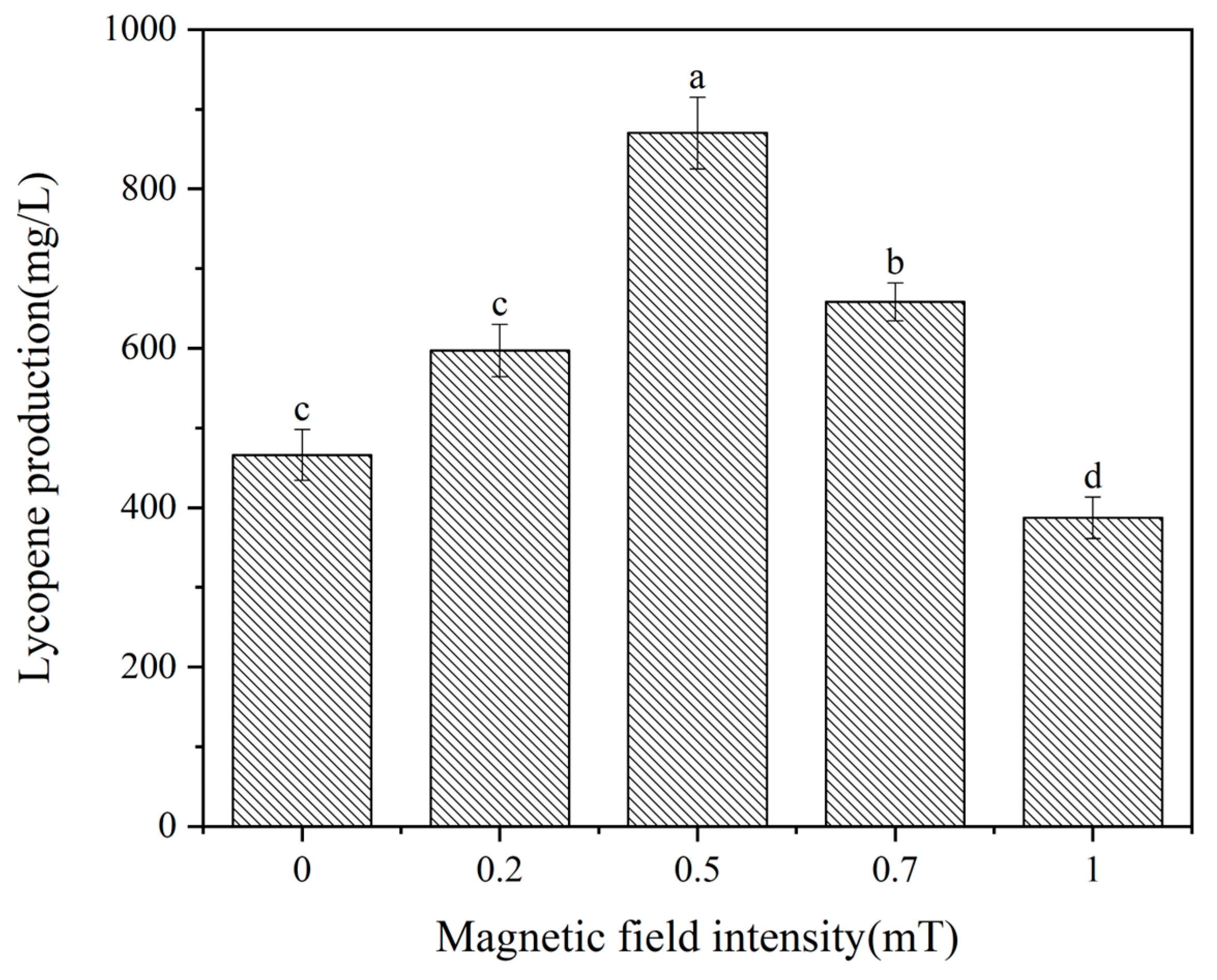
Figure 6.
effect of magnetic field on transcriptional level of genes encoded by key enzymes in lycopene metabolic pathway.
Figure 6.
effect of magnetic field on transcriptional level of genes encoded by key enzymes in lycopene metabolic pathway.

Table 1.
Primers used for RT-qPCR assays.
| Gene name | Gene code | Forward primer sequence(5’-3’) | Reverse primer sequence(5’-3’) |
| carRA | gene_8655 | CATCTCGTCGTTGGTTCA | AAGCATAGGCAATAACACAAG |
| carB | gene_5071 | GGCACAGATATAACTTGA | TTATTCTTATTGGCTTCCT |
| sod | gene_5118 | ATCACTACAATCCTACTG | ACCATACTTCTTCCAATA |
| cat | gene_6144 | CTATGCTACCAGAGATATG | CCAGACCTTAGTTACATC |
| iscA | gene_10317 | CTGCTGCCAACTCGTTAA | CTGCTGGTGTCAGTGTAAG |
| elo3 | gene_7519 | TGGTCATCAAGAAGAAGA | GTCAAGTTCAGGATAATAGG |
| tef1 | GGTAAGTCTACCACCACTGGTCACT | CAAGAGGAGGGTAGTCAGTGTAAGC |
Table 3.
results of RT-qPCR verification of some differentially expressed genes.
| Name | Gene ID | Description | Fold change of RNA-Seq | 2⊿⊿Ctof RT-qPCR |
|---|---|---|---|---|
| carRA | gene_8655 | phytoene synthase | 1.12 | 2.15 |
| carB | gene_5071 | Phytoene desaturase | 2.23 | 1.47 |
| sod | gene_5118 | Superoxide dismutase | 2.42 | 2.15 |
| cat | gene_6144 | catalase | 8.18 | 7.80 |
| iscA | gene_10317 | iron-sulfur cluster assembly protein | 1.27 | 1.08 |
| elo3 | gene_7519 | ELO family | 4.26 | 3.19 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



