Submitted:
02 December 2023
Posted:
05 December 2023
You are already at the latest version
Abstract
Oral squamous cell carcinoma (OSCC) remains an under-studied and significant global cancer killer and dismal survival rates have not changed in decades. A better understanding of the molecular basis of OSCC progression and metastasis is needed to develop new approaches for treating this disease. The supportive network surrounding cancer tumor cells known as the tumor microenvironment (TME) has gained increasing interest lately since it performs essential pro-tumorigenic functions. Cancer-associated fibroblasts (CAFs) are one of the main cell types in the TME and are known to play a key role in influencing the biological behavior of tumors. CAFs present a heterogeneity both in phenotype as well as functions, leading to the suggestion of different CAF subtypes in several cancer forms. The task to subtype CAFs in OSCC has however just begun and there is today no united way of subtyping CAFs in this disease. This review aims to define the features of CAFs and to summarize CAF subtype research in OSCC including aspects as disease prognosis and therapeutic opportunities.
Keywords:
Cancer‐associated fibroblast
; CAF subtype
; oral squamous cell carcinoma
; tumor microenvironment
1. Introduction
Head and neck cancer consists of several subtypes, with oral squamous cell carcinoma (OSCC) being the most common. The 5-year survival rate for all OSCC patients is approximately 50%, and this measure decreases to approximately 30% when looking only at patients diagnosed with more advanced stages of the disease [1,2]. Frequent late-stage diagnoses and high rates of recurrence are the key drivers of this poor prognosis. Interventions such as immunotherapy and targeted therapies have enhanced cancer treatment efficacy and improved cancer patient survival in general, building on successes from standard care involving surgery, radiation, and chemotherapy. However, gains in survival have been modest. There remains a clear need to develop new approaches for treating this disease. A better understanding of the molecular basis of OSCC progression and metastasis is needed to provide new treatment/disease management options.
Recent years have seen an expanded research focus on the tumor microenvironment (TME) based on the emerging evidence that it plays a crucial role in tumor progression and metastasis. Cancer associated fibroblasts (CAFs) represent one of the main cell constituents in the TME and these are known to contribute to stromal desmoplasia, which is linked to worse outcome for many tumor types including head and neck cancer [3,4]. Expanded research in to CAFs have indicated a key role for these fibroblasts in tumor progression and metastasis, making them attractive factors for targeted therapy and immunotherapy research [5]. However, effective targeting of CAFs require a robust understanding of the heterogeneity of CAF cell populations, as well as the biological and clinical significance of various CAF subpopulations.
CAF subtypes have been widely studied in pancreatic, hepatic, and breast cancer [6,7,8,9,10,11,12,13] yet no tools currently exist for CAF subtyping analysis in the context of OSCC. Moreover, only a few groups have investigated CAF cell population heterogeneity in OSCC. The prognostic role of CAF infiltration in tumors has recently been established: in oral cancer, the presence of CAFs have been linked to negative clinical outcome by several authors [14,15,16,17]. One multicenter, retrospective analysis of 282 OSCC patients reported that the strongest independent risk factor of early OSCC death was a feature of stroma rather than tumor cells [18]. Herein, we will review and define the features of CAF subtypes, summarize CAF subtype research in OSCC, present new potential therapeutic strategies linked to CAF subtypes, and describe future research challenges for CAF subtype research in the context of OSCC.
2. Defining fibroblasts
Fibroblasts exist in almost all tissue types and are the main cell type in stroma which produces the structural framework of human tissue through creation and maintenance of the extracellular matrix (ECM). Fibroblasts are derived mainly from primitive mesenchymal cells, although alternative origins have been reported (this is described in a subsequent section). There is no single marker used to identify fibroblast cells. In practice, fibroblasts taken from tumors/ tissues are usually first defined through negative selection: they do not express epithelial (EpCAM), endothelial (CD31), leukocyte (CD45), or erythrocyte (CD235a) markers. Next, positive expression of mesenchymal markers such as Vimentin (VIM), alpha-smooth muscle actin (αSMA), and others are used to indicate that a given cell is a fibroblast. In culture, fibroblasts can also be characterized by their spindle-like appearance.
Fibroblast functions include tissue homeostasis, wound healing response through secretion of extracellular matrix proteins to help heal the area, and/or through secretion of cytokines to recruit immune cells and initiate an inflammatory response. Normal fibroblasts can also serve as progenitors for specialized mesenchymal cell types, such as bone-forming osteoblasts and lipid-filled adipocytes [19]. Further, fibroblasts activated in a TME are called CAFs and these are known to display a vast array of functional abilities affecting tumor progression.
3. Fibroblast activation and CAF markers
3.1. Origin
CAFs may originate from normal tissue fibroblasts, as reported by others [20], with lineage tracing studies in genetically engineered mouse models supporting this concept [21]. These lineage tracing mouse models utilize a specific fibroblast gene promoter attached to a reporter molecule (e.g. mCherry or GFP). This reporter is expressed once the cell begins to express the fibroblast gene used to promote the reporter in the mouse (e.g. a Col5a1 promoter). The cell continues to express the reporter once activated, and expression of the marker is passed down to all daughter cells. This approach has allowed tracking of a chosen cell type as it morphs into a CAF in a tumor setting. As an example of lineage tracing model, Garcia et al. [21] showed that mesenchymal cells driven by Hoxb6 and Gli1 expression become fibroblasts then CAFs in healthy pancreas then pancreatic ductal adenocarcinoma (PDAC), respectively.
However, lineage tracing studies have also challenged the belief that most CAFs originate from normal fibroblasts in the tissue - Helms et al. have shown that pancreatic stellate cells (PSCs, the resident fibroblast-like cell of the pancreas) which were lineage traced with Fabp4, only contributed to a fraction of the CAFs in PDAC, contrary to popular belief that most CAFs in PDAC come from PSCs [22]. This work demonstrated that there is heterogeneity in the origin of cells that become CAFs in PDAC and potentially in other tumor types as well. Other than lineage tracing, single-cell RNA-sequencing (scRNA-seq) in tandem with either flow-cytometry sorting or mass spectrometry have allowed researchers to identify specific CAF subsets by surface markers that have distinct gene signatures denoting different functions. For instance, Hutton et al. [23] showed that CD105+ and CD105- CAFs in PDAC were mutually exclusive cells that could not be converted into each other and that CD105+ CAFs induced increased tumor progression, while less abundant CD105- CAFs supported favorable anti-tumor immunity. In addition to in vivo models, many in vitro studies clearly display how fibroblasts can become CAFs [24,25]. These studies have shown many origins of CAFs, including endothelial cells through endothelial-mesenchymal transition (EndMT) [26], epithelial cells under specific circumstances through epithelial-mesenchymal transition (EMT) [27], bone marrow-derived mesenchymal stem cells (MSCs) [28], hematopoietic stem cells (HSCs) [29], cancer stem cells (CSCs) [30], adipocytes [31], pericytes [32], and stellate cells [33]. The impact of these different suspected progenitors on fibroblast activation into certain CAF subtypes during tumorigenesis is currently a busy avenue of inquiry, and the importance of cell origin heterogeneity in CAF effects remains to be explored.
3.2. Fibroblast activation
Fibroblasts can be activated into CAFs through several mechanisms. Among growth factors, the TGFβ family of ligands [34] are well-established as activating signals. Lysophosphatidic acid, a lipid mediator [35], can also drive expression of the myofibroblast marker αSMA while also increasing the activity of the contractile cytoskeleton. Fibroblast activation through Notch signaling [36,37] has been linked to poor prognosis of ductal breast carcinoma. Several inflammatory cytokines drive CAF subtype differentiation, with IL-6 activating STAT signaling [38] and IL-1 acting through NF-κB [39]. Stromatogenesis as a result of connective tissue reaction to cancer has been shown to promote desmoplastic fibroblast differentiation in three-dimensional cultures [40], and YAP-mediated enhancement of matrix stiffening helps to maintain CAF phenotypes in breast cancer [41].
Physiological and genomic stress can also drive fibroblast activation info CAFs, with factors like reactive oxygen species [42] and double-stranded DNA breaks [43] serving as examples of this phenomenon. There is increasing evidence that cancer cell-derived extracellular vesicles (EVs) can also drive CAF activation [25,44,45,46], though EV content differs and can include miRNAs, TGFβ, and cells surface proteins, as well as other proteins, mRNAs, and lncRNAs [44,47,48]. It should be noted, however, that many CAF subtypes and functions exist on a spectrum, rather than distinctive subtypes – and that these subtypes can have overlapping functions, a reality that amplifies the complexity inherent in defining CAFs subtypes.
3.3. Fibroblast markers
CAFs lack a single, specific biomarker in various tumor types. This can be a significant challenge in the context of CAF research. Besides widely mentioned mesenchymal biomarkers such as VIM, αSMA, fibroblast activation protein (FAP), podoplanin (PDPN), integrin beta-1 (ITGB1, also known as CD29), FSP-1 (S100A4), and platelet-derived growth factor alfa (PDGFR-α), PDGFR-β [7,25], the role of matrix metalloproteinases (MMPs), through which CAFs can facilitate tumor growth, invasion, and metastasis has also been evaluated [49]. It is now commonplace for researchers to try to identify useful CAF markers through either flow sorting followed by RNA-seq and/or scRNA-seq to discover new potential CAF markers that are then validated in vivo. The LRRC15 protein represents one example of this - and may also serve as a putative target for precision medicine. High expression of LRRC15 has been reported in CAFs, but not in normal fibroblasts, for multiple tumor types [50]. Moreover, LRRC15 expression has been validated as an important CAF marker that can suggest poor response to immunotherapy treatment [51].
To date, exclusive CAF markers have not been reported. Heterogenous expression of some or all of the above-referenced markers in the context of CAFs is suggestive that tumors arise through distinct CAF subgroups that have different cells of origin, different stages of activation, and potentially unique cell function.
4. CAF subtypes in various cancer types
Flow cytometry and scRNA-seq studies have begun to define the phenotypic and functional heterogeneity of CAFs for individual cancer types, revealing a variety of CAF subtypes (which may also be referred to as “subsets” or “subpopulations”) (Table 1). A cell subtype is defined by specific markers, unique functional properties, and has a secretome pattern that differs from other subtypes. Turley et al. have created a resource for searching CAF and fibroblast markers called “FibroXplorer” [52]. It includes data for both human and mouse CAFs/fibroblasts. This is a scRNA-seq object that was created from pooling data from 17 tissues, 50 datasets, and 11 tumor types. Although this may not help with subtyping CAFs in a specific model or a patient sample, it is a great resource for looking at specific CAF markers in the context of various disease states or tissues of origin. Work by the group behind the FibroXplorer resource has also been used to define two universal fibroblast transcriptional sites of origin using mouse tissues, characterize Pi16+ and Col15a1+ expressing fibroblasts, and describe use of dermatopontin (Dpt) to trace fibroblasts [53]. While almost all healthy tissue fibroblasts expressed Dpt, especially in Pi16+ and Col15a1+ expressing fibroblasts, Dpt expression has been reported to be lost as a fibroblast becomes specialized either into a CAF or a specific fibroblast niche. While this work is a powerful resource for fibroblast research, emerging findings around CAF subtypes for specific cancer tissues show that there is additional complexity to unpack. Subsequent sections will describe CAF subtype-specific findings for breast cancer, pancreatic cancer, hepatocellular carcinoma, and cholangiocarcinoma.
4.1. Breast cancer
Costa et al., Givel et al., and Bonneau et al. have demonstrated the presence of four CAF subtypes in breast cancer when samples were characterized using multicolor flow cytometry [6,7,54]. The CAF-S1 subset, typically positive for FAP and CD29 and lacking CAV1, contributes to immunosuppression through attracting CD4+ T cells, increasing CD4+CD25+ T lymphocyte survival, and promoting their differentiation into FOXP3+ cells [7]. This CAF-S1 subset was preferentially detected in aggressive breast cancer [7], in cases of relapsing luminal breast cancer [6], and in mesenchymal high-grade serous ovarian cancer associated with poor prognosis [54]. On the other hand, the presence of the FAP negative CAF-S4 subset in metastatic axillary lymph nodes of breast cancer correlated with the later development of distant metastasis, a finding that suggests that evaluating CAF subtypes may be useful as a prognostic marker for recurrence or metastasis.
Using scRNA-seq, Sebastian et al. detected myofibroblast-like CAFs (myCAFs), inflammatory CAFs (iCAFs), and antigen-presenting CAFs (apCAFs) in breast cancer [55]. Interestingly, these subsets with similar transcriptomes could also be detected in pancreatic cancer, based on analyses of publicity available RNA-seq data [9].
4.2. Pancreatic and gastric cancer
Pancreatic ductal adenocarcinoma (PDAC) has been the primary subtype of focus of CAF research in pancreatic malignancy, with several authors having delineated CAFs into myCAFs and iCAFs [8,9,10,56]. These two subpopulations of CAFs have been identified using immunofluorescence (IF), immunohistochemistry (IHC), and microscopy [10]. Using scRNA-seq, 962 fibroblast cells could be analyzed in vivo and two distinct subclusters could be formed with unique gene signatures. The subcluster 1 had enriched expression of Il-6 and Il-8 and chemokines such as Cxcl1, Cxcl2, Ccl2, and Cxcl12, and was identified as iCAFs. The αSMA positive subcluster 2 was identified as myCAFs [9]. Using Gene Set Enrichment Analysis (GSEA), enriched pathways could be identified enclosing an upregulation of inflammatory pathways such as IFNγ response, TNF/NF-κB, IL2/STAT5, IL6/JAK/STAT3, and the complement pathway in iCAFs, while myCAFs displayed an upregulation in pathways such as smooth muscle contraction, focal adhesion, ECM organization and collagen formation. Biffi et al. further revealed IL1 to be critical for generation of iCAFs [8]. When analyzing PDAC tumors in Kras+/LSL-G12D; Trp53+/LSL-R172H; Pdx1-Cre (KPC) mice, a new subpopulation – namely, apCAF – were described [9]. These MHC II-expressing and Cd74-expressing apCAFs could activate CD4+ T cells in an antigen-specific fashion based on use of an ovalbumin-specific TCR transgenic OTII mouse model. An apCAF subtype has also been suggested by Zhang et al. in HNSCC [57].
Another group also divided PDAC CAFs into myCAFs and iCAFs, with IL1 inducing an iCAF phenotype similar to the one reported by Biffi et al. [58]. However, this other group also revealed that hypoxia promoted the iCAF-like state. The TME is characterized by varied extent of hypoxia and associated with poor prognosis of patients with PDAC. This finding links the direct functional impact of hypoxia on CAFs in the TME. A third group also delineated CAFs into myCAFs and iCAFs [59]. They showed in an orthotopic model of diffuse-type gastric cancer (DGC) that EVs from highly metastatic DGC cells transferred various miRNAs and induced chemokine expression (CXCL1 and CXCL8) in fibroblasts.
While most CAF subtypes are studied in their cancer-promoting functions, the discovery of cancer-restraining CAFs (rCAFs) in PDAC, which are defined by the expression of Meflin, complicates our understanding of the role of fibroblasts in cancer. These rCAFs have been reported to have inhibitory effects on tumor progression in pancreas cancer [60]. The rCAFs and the previously mentioned CD105- CAFs in PDAC both show that there are CAFs that help in the fight against cancer and must not be forgotten when identifying and studying heterogeneous CAF subtypes.
4.3. Hepatocellular carcinoma and cholangiocarcinoma
Ying et al. reviewed studies summarizing CAF subpopulations and their markers in hepatocellular carcinoma (HCC) and cholangiocarcinoma (CCA), highlighting the problematic lack of specific CAF markers [11,12]. Six distinct fibroblast subsets were detected using scRNA-seq on human intrahepatic cholangiocarcinoma (ICC) by Zhang et al.: vascular CAFs (vCAF), matrix CAFs (mCAF), inflammatory CAFs (iCAF), antigen-presenting CAFs (apCAF), EMT-like CAFs (eCAF), and lipofibroblasts. The authors determined that the vCAF subgroup was the most prevalent in the tumors. These vCAFs were shown to secrete IL-6, which induced significant epigenetic alterations in tumor cells and furthered malignancy [13].
5. CAF subtypes in oral cancer
Given the emerging complexity and varied reports about CAF subtypes, it is clear that resolving the role of these cells in cancer processes will remain challenging for the near future. It is tempting to suggest that CAF subtypes described by one group are the same as subtypes described by another group with the only difference being the name. For example, Li et al. detected two subtypes in colorectal cancer (CRC) naming them CAF-A and CAF-B [61]. CAF-A expressed markers of myofibroblasts such as Acta2, Tagln, and Pdgfa according to differential expression analysis; however, the authors did not label these as myCAFs. Bonneau et al. stated that their suggested CAF-S1 and S4 myofibroblasts have been validated in other studies, yet no other referred groups use the same nomenclature [6,62]. In fact, they have also shown that CAF-S1 fibroblasts themselves have eight distinct subtypes within that subset, some of which fit the myCAF/iCAF nomenclature [7]. Additionally, a subset expressing ECM-related genes displaying ECM-remodeling properties pathways have been suggested by several authors however without a united name [13,57,62,63]. One CAF subtype that seems to have an agreed-upon name and function are the inflammatory CAFs, iCAFs, which several authors have described and implicated in tumorigenesis [25,57,64,65,66] and a recent study demonstrates that oral cancer cell-derived EVs can be responsible for activating this iCAF subset [25]. Further, it must be noted that CAF subtypes in distribution and functional roles seem to differ between cancer types. Accordingly, it is of high interest to find similarities among different cancer forms, as summarized in Table 2.
While several research groups have detected and categorized CAF subtypes in breast, ovarian, pancreatic, and hepatocellular cancer, the work to describe CAF subtypes in oral cancer has just begun. Galbo et al. attempted to find common ground between CAF subgroupings by analyzing results from head and neck squamous cell carcinoma (HNSCC), melanoma, and lung cancer (LC) [63]. The authors identified six CAF subtypes named pan-CAFs that were shared across the different tumor types. Pan-myCAFs showed elevated expression of activated fibroblast markers and smooth muscle cell markers. Pan-dCAFs revealed elevated expression of genes coding for collagen and extracellular matrix remodeling. Pan-iCAFs and pan-iCAF-2 both showed high expression of genes related to inflammation. The authors suggests that patients with high pan-iCAFs might be linked to a poor clinical outcome due to an enrichment of gene sets and pathways related to interleukin signaling, which is linked to the activation of innate immune cells and the establishment of an immunosuppressive microenvironment [53]. Pan-nCAFs was referred as normal fibroblasts whereas pan-pCAFs showed elevated expression of genes related to cell cycle. The work by Galbo et al. and the FibroXplorer tool represent robust resources for initiating CAF subtype analysis in OSCC models and patient samples [52].
Costea et al. detected two CAF subtypes in oral cancer. CAF-N having transcriptome and secretome close to normal fibroblasts and CAF-D with more divergent expression pattern [67]. Both subtypes showed tumor-promoting capability but were mechanistically different. CAF-N, secreting significantly higher levels of keratinocyte growth factor (KGF) and acidic fibroblast growth factor (aFGF), defined more of migratory fibroblasts dependent on hyaluronan and essential for supporting cancer cell invasion. CAF-D, synthesizing higher levels of TGF-β1, was shown to increase invasion and expression of EMT markers. This was impaired by TGF-β1 inhibition which suggests CAF-Ds to have similarities with myCAFs. The protumorigenic potential of CAF-derived TGF-β1 was confirmed by Yang et al., subtyping CAFs into myofibroblasts (mCAFs) and inflammatory CAFs (iCAFs) [68]. The authors suggest the two subtypes to both promote OSCC progression, however through different regulatory roles in the TME. Patel et al. on the other hand, divided CAFs into C1 or C2 CAFs [69]. C1 CAFs, showing lower expression of αSMA, showed increased proliferation of oral cancer cells linked to BMP4. This highlights that aSMA expression is not crucial for protumorigenic CAFs. The first steps in detecting an apCAF subtype expressing Cd74 and other MHC-II related genes has also started in oral cancer [65]. Zhang et al. suggest their cluster 4 to be an apCAF, associated with poorer overall survival in HNSCC [57]. CAF subtypes linked to angiogenesis has been detected in other cancer forms [13,55,63] and future studies are encouraged to see whether CAF subtypes can be linked to pathological angiogenesis in OSCC.

Table 2.
Similarities in CAF subtypes across malignancies.
| Subtype | Marker | Transcriptome | Pathways | Additional notes | Reference |
|---|---|---|---|---|---|
| myCAF | MHCII-/Ly6C- [9,58] | Acta2, contractile proteins (Tagln, Myl9, Tpm1, Tpm2, Mmp11, Postn, Hopx), Igfbp3, Thy1, Col12a1, Thbs2 [9] | Smooth muscle contraction, focal adhesion, ECM organization, collagen formation [9] | Activates through: - TGFβ [8] |
[8,9,10,55,56,58,59,63,65,66,68] |
| αSMA+ IL-6- [10] | Acta2, TGFβ response genes (Ctgf and Col1a1) [10] | Smooth muscle contraction, vascular wound healing [63] | |||
| αSMA [59] | Contractile proteins (Acta2, Tagln, Myl9, Tnc, Tpm1, Tpm2), growth factors (Tgfb1, Tgfb2, Ccn2/Ctgf, Pgf, Vegfa, Wnt5a, Pdgfrb), Igfbp3, Cnn2, Cnn3, Tmem119 [55] | ECM-receptor interaction, vascular smooth muscle contraction, focal adhesion [66] | |||
| Increased Acta2 and reduced Cxcl12, Des [56] | Contractile fiber, blood vessel development, tissue migration, ECM organization [68] | ||||
| Acta2, contractile proteins (Myh11, Mcam, Tagln, Mylk) [63] | |||||
| Pdpn, Col1a1 [65] | |||||
| Acta2, Mylk, Myl9 [66] | |||||
| Acta2, Tagln, Mmp11, Myl9, Postn, Tpm1, Tpm2 [68] | |||||
| iCAF | Ly6C+ [9,58] | Il6, Il8, chemokines (Cxcl1, Cxcl2, Ccl2, Cxcl12), Pdgfra, Cfd, Lmna, Dpt, Has1, Has2, Agtr1 [9] | Cytokine signaling, JAK/STAT signaling [8,10] | Activates through: - NF-κB signaling through IL-1 [8] - Cancer-derived EVs [25,59] - Hypoxia induced HIF1α [58] |
[8,9,10,13,25,55,56,58,59,63,65,66,68] |
| αSMA- IL-6+ [10] | Cytokines (Il6, Il11, Lif) and chemokines (Cxcl1, Cxcl2) [10] | IFNγ response, TNF/NF-κB, IL2/STAT5, IL6/JAK/STAT3, the complement pathway [9] | |||
| CXCL8 [59] | Fbln1, Igfi, Cxcl1, Igfbp6, Slpi, Saa1, and complement genes (C3, C7) and reduced Acta2 [13] | ECM, inflammatory response regulation, complement activation [13] | |||
| Ly6c1 and inflammatory cytokines (Il6, Cxcl1, Cxcl12, Mcp-1/Ccl2, Mcp-3/Ccl7, Il33), compliment pathway members (C3, C4b, C1s1, C1s2), Clec3b, Has1, Dpt, Col14a1, Ccl7, Ly6a/Sca-1, Saa3, Ogn, Prg4, Prelp, Efemp1, Htra3, Pdgfra [55] | Inflammation (iCAF) NFκB signaling pathway (iCAF-2) [63] | Function: - Recruit M2 macrophages by secreting CXCL12 [66] |
|||
| Increased Vim, Fap, Col3a1, Des, Il6, Cxcl12 and reduced Acta2 [56] | TNF signaling pathway, IL-17 signaling pathway, ECM-receptor interaction [66] | ||||
| iCAF: Cfd, C3, Cxcl14, Cxcl12, Il33; iCAF-2: inflammatory genes (Cxcl2, Tnfaip3), Icam1, Clu, Bdkrb1, Relb [63] | Blood vessel development, tissue migration, ECM organization [68] | ||||
| Cxcl12, Cxcl14, Igf1 [65] | |||||
| Cxcl12, Pdgfra, Thy1 [66] | |||||
| Chemokines (Pdgfra, Cxcl12, Cfd, Dpt, Lmna, Cxcl2, Ccl2) [68] | |||||
| apCAF | MHCII+ [9,58] | MHC-II genes (Cd74, H2-Aa, H2-Ab1), Saa3, Slpi [9] | Antigen presentation and processing, fatty-acid metabolism, MYC targets, MTORC1 signaling [9] | Function: - Activate CD4+ T cells [9] |
[9,13,55,57,58,65] |
| MHC-II (Cd74, Hla-Dra, Hla-Drb1) [13] | Leukocyte cell-cell adhesion, response to IFN-γ, antigen processing, and antigen presentation via MHCII [13] | ||||
| MHC-II (Cd74, H2-Aa, H2-Ab1, H2-Eb), Krt8, Krt18, Fsp1 [55] | Ribosome, antigen processing and presentation, phagosome [57] | ||||
| MHC-II (Cd74, Hla-Dra, Hla-Drb1, Hla-Dqb1, Hla-Drb5, Hla-Dpa1, Hla-Dpb1) [65] | |||||
| Matrix CAF (mCAF) [13] Cluster 3 [57] mCAF [62] dCAF [63] |
Fibulin-1 and PDGFRα [62] | High levels of ECM signatures (Col5a1, Col5a2, Col6a3), Postn, Fn1, Lum, Dcn, Vcan) and low levels of Acta2 [13] | ECM and collagen fibril organization [13] | [13,57,62,63] | |
| Acta2- [57] | ECM remodeling (protein processing in the endoplasmic reticulum, oxidative phosphorylation, proteoglycans in cancer, ECM-receptor interaction) [57] | ||||
| ECM-related genes such as glycoproteins (Dcn, Lum, and Vcan), structural proteins (Col14a1), matricellular proteins (Fbln1, Fbln2, and Smoc), and matrix-modifying enzymes (Lox and Loxl1) , Cxcl14 [62] | ECM signature [62] | ||||
| Col1a1, Col3a1, Stc1, Mmp1, Mmp11, Col10a1 [63] | ECM remodeling [63] | ||||
| S1 | CD29Med FAPHi αSMAHi FSP1Low-Hi PDGFRβMed-Hi CAV1Low [6,7,54] |
Function: - Promotes immunosuppression [7,54] - Increases migration properties of luminal BC cells [6] |
[6,7,54] | ||
| S4 | CD29Hi FAPLow αSMAHi FSP1Hi PDGFRβMed-Hi CAV1Neg-Low [6,7,54] |
[6,7,54] |
6. The prognostic role of CAF subtypes in oral cancer
Looking upon CAFs as a heterogeneous population of cells with different marker proteins, transcriptomes, and secretomes, it follows that different CAF subtypes may have different functional properties – and that they may be associated with different clinical features, including prognosis. Yang et al. defined CAFs by t-SNE plots into mCAFs (highly expressing Acta2, Tagln, Mmp11, Myl9, Postn, Tpm1, and Tpm2), and iCAFs (highly expressing chemokines) [68]. The authors could see a correlation between mCAF/iCAF ratio and tumor stage in OSCC, with a higher proportion of mCAFs detected in more advanced stages of disease. Looking into survival, however, high mCAFs and iCAFs both predicted poor overall survival in OSCC, while only high iCAFs values alone predicted poor overall survival after five years. Galbo et al. could link specific pan-CAFs to clinical outcome in some cancer types - high presence of pan-iCAFs correlated to poor clinical outcome in low grade glioma - however, pan-CAFs could not be correlated to prognosis in HNSCC. This shows the importance of subtyping CAFs to better understand patient outcomes. Additionally, HNSCC is known to be a very heterogenous group of tumors, both in morphology, etiology, treatment, and prognosis. Thus it is imperative to explore CAF subtypes in OSCC specifically in order to glean meaningful insights into the role of CAFs in this disease.
7. Therapeutic opportunities linked to CAF subtypes
In PDAC, up to 90% of the tumor volume consists of stroma [58]. In HNSCC, CAFs can account for up to 80% of the tumor mass [70]. Tumor-stroma ratio has been shown to have prognostic importance [71] and evaluating this ratio can also garner insights into clinical disease [72]. Tumor stroma may block delivery of therapy to cancer cells, support cancer growth, and promote therapy resistance [48]. On the other hand, the tumor stroma may also contain potential targets for potential therapy. Recently, the number of preclinical experiments targeting CAFs to restore the anticancer immune response has increased dramatically. Further, strategies for CAF-based immunotherapy have emerged: direct CAF depletion including a novel therapeutic that uses FAP-targeting chimeric antigen receptor (CAR)-T cells [5] resulting in reduced CAF activation [73] and functional suppression of CAF-induced ECM remodeling [74,75,76] (Figure 1).
Multiple rationales exist for targeting CAFs in anti-cancer treatment although the heterogeneity of CAF populations – and the reality that CAFs are always solely tumor promoting – complicate this approach. Supporting this, recent studies aiming to target stromal cells in general have not always been successful [77,78,79]. A new therapeutic approach would be to embrace the detection of CAF heterogeneity and to understand the importance of subtypes in terms of treatment opportunities. Costa et al. found that silencing B7H3 and CD73, which are highly expressed in S1 CAFs, significantly reduced their impact on the total numbers of tumor suppressive CD25+FOXP3+ T cells [7]. The authors accordingly suggests that therapy against B7H3 or CD73 could target CAF-S1 cells and enhance anti-tumor immunity by inhibiting CAF-S1-mediated immunosuppression. Alternatively, Biffi et al. suggest a combinatorial therapy with both a TNFα-neutralizing antibody and a IL1 receptor antagonist for targeting iCAFs in vivo [8]. Converting specific CAF subtypes into a less activated state or shifting tumor-promoting CAF subtypes towards a quiescent or tumor-restraining phenotype - or even neutralizing ligands that activate fibroblasts into specific CAF subtypes such (e.g. IL-1, IL-6, LIF, and/or TGF-β) - may open up completely new druggable pathways [80].
In terms of targeting specific markers on CAFs in general, FAP is one of the most widely studied. FAP has been used as a target in preclinical therapeutic strategies with vaccines, antibodies, and CAR-T cells [81,82,83,84]. In fact, a promising new FAP targeting CAR-T cell has been shown to loosen the desmoplasia that restricts T cell invasion into tumors which enables other tumor-targeting CAR-T cells to be used to finish the job and shrink the tumor [5]. However, FAP does not appear to distinguish the iCAF subtype in PDAC or OSCC [9,25].
Numerous previous studies have linked the interaction between CAFs and tumor cells in tumorigenesis. Infiltrating immune cells like macrophages, dendritic cells, lymphocytes, and neutrophils are other key components in the TME. Cancer immunotherapy has been one of the biggest medical breakthroughs in recent years. Several reports reveal improved patient survival with those who have increased CD3+/CD8+ T cell tumor infiltration [7,85,86]. The importance of lymphocytes as a positive prognostic factor in the TME has also been studied in oral cancer [87]. The interplay between tumor cells and immune cells throughout tumor progression involves multiple steps with both stimulating and inhibitory factors. The interaction between CAFs, immune cells, and tumor cells in the TME and their role in tumor progression is an ongoing topic of study. It is, however, known that CAFs regulate and enable the pro-tumor activity of TME immune cells both directly and indirectly via the ECM and secretion of cytokines such as IL-6 [39,76,88,89,90].
Most research into iCAFs describes them as pro-tumorigenic and prognostically unfavorable, which aligns with the role of iCAFs and IL-6 secretion. Since cancer immunology has largely been focused on T cells and CD3+/CD8+ T cell response it is crucial to determine CAF subtype functions in oral cancer, especially linked to CD8+ and CD4+ T cells as done by Costa et al. in S1 CAFs in breast and ovarian cancer [7,54]. For instance, CXCL12 secreted by FAP+ CAFs decreases tumor infiltration of CD8+ T cells [81]. This makes CXCL12 or its associated signaling pathway a potential therapeutic target. Similar analyses of CAF subtypes in the context of disease will reveal additional therapeutic targets with the potential to impact patient outcomes.
8. Conclusions
Herein we have reviewed how heterogenous CAF subpopulations may be defined and the present understanding of how they are generated, particularly in the context of oral cancer. The immediate research challenges in this area include: i) development of a unified classification scheme for CAF subtypes in OSCC, ii) assessment of the relative frequency of each of these subtypes in the context of clinical oral cancer tissues and evaluate the prognostic significance of CAF subtype distribution in OSCC, iii) functional evaluation of the iCAF subtype in the context of OSCC models and iv) an evaluation of therapeutic interventions against specific CAF subtypes. Ongoing work on these areas has the potential to better illuminate our understanding of malignancy and provide new tools for to improve lives of patients.
Author Contributions
Conceptualization, J.A., C.-M.L., K.L.B. and C.G.; writing—original draft preparation, J.A.; writing—review and editing, J.A., C.-M.L., K.L.B. and C.G. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by grants from the Swedish Society of Medicine, the Royal Swedish Academy of Sciences, the Sweden-America Foundation, Lindhés Lawyes Agency, Karolinska Institutet, the Swedish Association for Otorhinolaryngology Head and Neck Surgery, Acta Oto-Laryngologica, the Svensson Sibling Foundation, Michael Smith Foundation for Health Research (MSFHR) RT-2021-1484, Canadian Institutes of Health Research (CIHR): operating grants MOP-133640 and PJT-162380 and funds from the Pacific Otolaryngology Foundation.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Suresh, G. M., R. Koppad, B. V. Prakash, K. S. Sabitha and P. S. Dhara. Prognostic indicators of oral squamous cell carcinoma. Ann Maxillofac Surg 2019, 9, 364–370. [CrossRef] [PubMed]
- American cancer society. Cancer facts & figures 2023; Atlanta, Ga: American Cancer Society, 2023.
- Zainab, H., A. Sultana and Shaimaa. Stromal desmoplasia as a possible prognostic indicator in different grades of oral squamous cell carcinoma. J Oral Maxillofac Pathol 2019, 23, 338–343. [CrossRef] [PubMed]
- Graizel, D., A. Zlotogorski-Hurvitz, I. Tsesis, E. Rosen, R. Kedem and M. Vered. Oral cancer-associated fibroblasts predict poor survival: Systematic review and meta-analysis. Oral Dis 2020, 26, 733–744. [CrossRef] [PubMed]
- Xiao, Z., L. Todd, L. Huang, E. Noguera-Ortega, Z. Lu, L. Huang, M. Kopp, Y. Li, N. Pattada, W. Zhong, et al. Desmoplastic stroma restricts t cell extravasation and mediates immune exclusion and immunosuppression in solid tumors. Nat Commun 2023, 14, 5110. [CrossRef] [PubMed]
- Bonneau, C., A. Eliès, Y. Kieffer, B. Bourachot, S. Ladoire, F. Pelon, D. Hequet, J. M. Guinebretière, C. Blanchet, A. Vincent-Salomon, et al. A subset of activated fibroblasts is associated with distant relapse in early luminal breast cancer. Breast Cancer Res 2020, 22, 76. [CrossRef] [PubMed]
- Costa, A., Y. Kieffer, A. Scholer-Dahirel, F. Pelon, B. Bourachot, M. Cardon, P. Sirven, I. Magagna, L. Fuhrmann, C. Bernard, et al. Fibroblast heterogeneity and immunosuppressive environment in human breast cancer. Cancer Cell 2018, 33, 463–479.e10. [CrossRef] [PubMed]
- Biffi, G., T. E. Oni, B. Spielman, Y. Hao, E. Elyada, Y. Park, J. Preall and D. A. Tuveson. Il1-induced jak/stat signaling is antagonized by tgfβ to shape caf heterogeneity in pancreatic ductal adenocarcinoma. Cancer Discov 2019, 9, 282–301. [CrossRef]
- Elyada, E., M. Bolisetty, P. Laise, W. F. Flynn, E. T. Courtois, R. A. Burkhart, J. A. Teinor, P. Belleau, G. Biffi, M. S. Lucito, et al. Cross-species single-cell analysis of pancreatic ductal adenocarcinoma reveals antigen-presenting cancer-associated fibroblasts. Cancer Discov 2019, 9, 1102–1123. [CrossRef]
- Öhlund, D., A. Handly-Santana, G. Biffi, E. Elyada, A. S. Almeida, M. Ponz-Sarvise, V. Corbo, T. E. Oni, S. A. Hearn, E. J. Lee, et al. Distinct populations of inflammatory fibroblasts and myofibroblasts in pancreatic cancer. J Exp Med 2017, 214, 579–596. [CrossRef]
- Chiavarina, B., R. Ronca, Y. Otaka, R. B. Sutton, S. Rezzola, T. Yokobori, P. Chiodelli, R. Souche, D. Pourquier, A. Maraver, et al. Fibroblast-derived prolargin is a tumor suppressor in hepatocellular carcinoma. Oncogene 2022, 41, 1410–1420. [CrossRef]
- Ying, F., M. S. M. Chan and T. K. W. Lee. Cancer-associated fibroblasts in hepatocellular carcinoma and cholangiocarcinoma. Cell Mol Gastroenterol Hepatol 2023, 15, 985–999. [CrossRef] [PubMed]
- Zhang, M., H. Yang, L. Wan, Z. Wang, H. Wang, C. Ge, Y. Liu, Y. Hao, D. Zhang, G. Shi, et al. Single-cell transcriptomic architecture and intercellular crosstalk of human intrahepatic cholangiocarcinoma. J Hepatol 2020, 73, 1118–1130. [CrossRef] [PubMed]
- Dhanda, J., A. Triantafyllou, T. Liloglou, H. Kalirai, B. Lloyd, R. Hanlon, R. J. Shaw, D. R. Sibson and J. M. Risk. Serpine1 and sma expression at the invasive front predict extracapsular spread and survival in oral squamous cell carcinoma. Br J Cancer 2014, 111, 2114–2121. [CrossRef] [PubMed]
- Kellermann, M. G., L. M. Sobral, S. D. da Silva, K. G. Zecchin, E. Graner, M. A. Lopes, I. Nishimoto, L. P. Kowalski and R. D. Coletta. Myofibroblasts in the stroma of oral squamous cell carcinoma are associated with poor prognosis. Histopathology 2007, 51, 849–853. [CrossRef] [PubMed]
- Vered, M., A. Dobriyan, D. Dayan, R. Yahalom, Y. P. Talmi, L. Bedrin, I. Barshack and S. Taicher. Tumor-host histopathologic variables, stromal myofibroblasts and risk score, are significantly associated with recurrent disease in tongue cancer. Cancer Sci 2010, 101, 274–280. [CrossRef] [PubMed]
- Vered, M., A. Shnaiderman-Shapiro, A. Zlotogorski-Hurvitz, T. Salo and R. Yahalom. Cancer-associated fibroblasts in the tumor microenvironment of tongue carcinoma is a heterogeneous cell population. Acta Histochem 2019, 121, 151446. [CrossRef]
- Marsh, D., K. Suchak, K. A. Moutasim, S. Vallath, C. Hopper, W. Jerjes, T. Upile, N. Kalavrezos, S. M. Violette, P. H. Weinreb, et al. Stromal features are predictive of disease mortality in oral cancer patients. J Pathol 2011, 223, 470–481. [CrossRef]
- Plikus, M. V., X. Wang, S. Sinha, E. Forte, S. M. Thompson, E. L. Herzog, R. R. Driskell, N. Rosenthal, J. Biernaskie and V. Horsley. Fibroblasts: Origins, definitions, and functions in health and disease. Cell 2021, 184, 3852–3872. [CrossRef]
- Kalluri, R. The biology and function of fibroblasts in cancer. Nat Rev Cancer 2016, 16, 582–598. [Google Scholar] [CrossRef]
- Garcia, P. E., M. Adoumie, E. C. Kim, Y. Zhang, M. K. Scales, Y. S. El-Tawil, A. Z. Shaikh, H. J. Wen, F. Bednar, B. L. Allen, et al. Differential contribution of pancreatic fibroblast subsets to the pancreatic cancer stroma. Cell Mol Gastroenterol Hepatol 2020, 10, 581–599. [CrossRef]
- Helms, E. J., M. W. Berry, R. C. Chaw, C. C. DuFort, D. Sun, M. K. Onate, C. Oon, S. Bhattacharyya, H. Sanford-Crane, W. Horton, et al. Mesenchymal lineage heterogeneity underlies nonredundant functions of pancreatic cancer-associated fibroblasts. Cancer Discov 2022, 12, 484–501. [CrossRef] [PubMed]
- Hutton, C., F. Heider, A. Blanco-Gomez, A. Banyard, A. Kononov, X. Zhang, S. Karim, V. Paulus-Hock, D. Watt, N. Steele, et al. Single-cell analysis defines a pancreatic fibroblast lineage that supports anti-tumor immunity. Cancer Cell 2021, 39, 1227–1244.e20. [CrossRef] [PubMed]
- Lawson, J., C. Dickman, R. Towle, J. Jabalee, A. Javer and C. Garnis. Extracellular vesicle secretion of mir-142-3p from lung adenocarcinoma cells induces tumor promoting changes in the stroma through cell-cell communication. Mol Carcinog 2019, 58, 376–387. [CrossRef] [PubMed]
- Arebro, J., R. Towle, C. M. Lee, K. L. Bennewith and C. Garnis. Extracellular vesicles promote activation of pro-inflammatory cancer-associated fibroblasts in oral cancer. Front Cell Dev Biol 2023, 11, 1240159. [CrossRef] [PubMed]
- Yeon, J. H., H. E. Jeong, H. Seo, S. Cho, K. Kim, D. Na, S. Chung, J. Park, N. Choi and J. Y. Kang. Cancer-derived exosomes trigger endothelial to mesenchymal transition followed by the induction of cancer-associated fibroblasts. Acta Biomater 2018, 76, 146–153. [CrossRef] [PubMed]
- Krzysiek-Maczka, G., A. Targosz, U. Szczyrk, M. Strzalka, T. Brzozowski and A. Ptak-Belowska. Involvement of epithelial-mesenchymal transition-inducing transcription factors in the mechanism of helicobacter pylori-induced fibroblasts activation. J Physiol Pharmacol 2019, 70. [CrossRef] [PubMed]
- Tan, H. X., Z. B. Cao, T. T. He, T. Huang, C. L. Xiang and Y. Liu. Tgfβ1 is essential for mscs-cafs differentiation and promotes hct116 cells migration and invasion via jak/stat3 signaling. Onco Targets Ther 2019, 12, 5323–5334. [CrossRef] [PubMed]
- Abangan, R. S., Jr., C. R. Williams, M. Mehrotra, J. D. Duncan and A. C. Larue. Mcp1 directs trafficking of hematopoietic stem cell-derived fibroblast precursors in solid tumor. Am J Pathol 2010, 176, 1914–1926. [CrossRef] [PubMed]
- Nair, N., A. S. Calle, M. H. Zahra, M. Prieto-Vila, A. K. K. Oo, L. Hurley, A. Vaidyanath, A. Seno, J. Masuda, Y. Iwasaki, et al. A cancer stem cell model as the point of origin of cancer-associated fibroblasts in tumor microenvironment. Sci Rep 2017, 7, 6838. [CrossRef]
- Bochet, L., C. Lehuédé, S. Dauvillier, Y. Y. Wang, B. Dirat, V. Laurent, C. Dray, R. Guiet, I. Maridonneau-Parini, S. Le Gonidec, et al. Adipocyte-derived fibroblasts promote tumor progression and contribute to the desmoplastic reaction in breast cancer. Cancer Res 2013, 73, 5657–5668. [CrossRef]
- Ning, X., H. Zhang, C. Wang and X. Song. Exosomes released by gastric cancer cells induce transition of pericytes into cancer-associated fibroblasts. Med Sci Monit 2018, 24, 2350–2359. [CrossRef]
- Tan, H. X., W. Z. Gong, K. Zhou, Z. G. Xiao, F. T. Hou, T. Huang, L. Zhang, H. Y. Dong, W. L. Zhang, Y. Liu, et al. Cxcr4/tgf-β1 mediated hepatic stellate cells differentiation into carcinoma-associated fibroblasts and promoted liver metastasis of colon cancer. Cancer Biol Ther 2020, 21, 258–268. [CrossRef] [PubMed]
- De Wever, O., Q. D. Nguyen, L. Van Hoorde, M. Bracke, E. Bruyneel, C. Gespach and M. Mareel. Tenascin-c and sf/hgf produced by myofibroblasts in vitro provide convergent pro-invasive signals to human colon cancer cells through rhoa and rac. Faseb j 2004, 18, 1016–1018. [CrossRef] [PubMed]
- Radhakrishnan, R., J. H. Ha, M. Jayaraman, J. Liu, K. M. Moxley, C. Isidoro, A. K. Sood, Y. S. Song and D. N. Dhanasekaran. Ovarian cancer cell-derived lysophosphatidic acid induces glycolytic shift and cancer-associated fibroblast-phenotype in normal and peritumoral fibroblasts. Cancer Lett 2019, 442, 464–474. [CrossRef]
- Su, Q., B. Zhang, L. Zhang, T. Dang, D. Rowley, M. Ittmann and L. Xin. Jagged1 upregulation in prostate epithelial cells promotes formation of reactive stroma in the pten null mouse model for prostate cancer. Oncogene 2017, 36, 618–627. [CrossRef] [PubMed]
- Strell, C., J. Paulsson, S. B. Jin, N. P. Tobin, A. Mezheyeuski, P. Roswall, C. Mutgan, N. Mitsios, H. Johansson, S. M. Wickberg, et al. Impact of epithelial-stromal interactions on peritumoral fibroblasts in ductal carcinoma in situ. J Natl Cancer Inst 2019, 111, 983–995. [CrossRef] [PubMed]
- Sanz-Moreno, V., C. Gaggioli, M. Yeo, J. Albrengues, F. Wallberg, A. Viros, S. Hooper, R. Mitter, C. C. Féral, M. Cook, et al. Rock and jak1 signaling cooperate to control actomyosin contractility in tumor cells and stroma. Cancer Cell 2011, 20, 229–245. [CrossRef]
- Erez, N., M. Truitt, P. Olson, S. T. Arron and D. Hanahan. Cancer-associated fibroblasts are activated in incipient neoplasia to orchestrate tumor-promoting inflammation in an nf-kappab-dependent manner. Cancer Cell 2010, 17, 135–147. [CrossRef]
- Amatangelo, M. D., D. E. Bassi, A. J. Klein-Szanto and E. Cukierman. Stroma-derived three-dimensional matrices are necessary and sufficient to promote desmoplastic differentiation of normal fibroblasts. Am J Pathol 2005, 167, 475–488. [CrossRef]
- Calvo, F., N. Ege, A. Grande-Garcia, S. Hooper, R. P. Jenkins, S. I. Chaudhry, K. Harrington, P. Williamson, E. Moeendarbary, G. Charras, et al. Mechanotransduction and yap-dependent matrix remodelling is required for the generation and maintenance of cancer-associated fibroblasts. Nat Cell Biol 2013, 15, 637–646. [CrossRef]
- Weinberg, F., N. Ramnath and D. Nagrath. Reactive oxygen species in the tumor microenvironment: An overview. Cancers (Basel) 2019, 11. [CrossRef] [PubMed]
- Fordyce, C., T. Fessenden, C. Pickering, J. Jung, V. Singla, H. Berman and T. Tlsty. DNA damage drives an activin a-dependent induction of cyclooxygenase-2 in premalignant cells and lesions. Cancer Prev Res (Phila) 2010, 3, 190–201. [CrossRef] [PubMed]
- Dai, G., X. Yao, Y. Zhang, J. Gu, Y. Geng, F. Xue and J. Zhang. Colorectal cancer cell-derived exosomes containing mir-10b regulate fibroblast cells via the pi3k/akt pathway. Bull Cancer 2018, 105, 336–349. [CrossRef] [PubMed]
- Ringuette Goulet, C., G. Bernard, S. Tremblay, S. Chabaud, S. Bolduc and F. Pouliot. Exosomes induce fibroblast differentiation into cancer-associated fibroblasts through tgfβ signaling. Mol Cancer Res 2018, 16, 1196–1204. [CrossRef] [PubMed]
- Ping, Q., R. Yan, X. Cheng, W. Wang, Y. Zhong, Z. Hou, Y. Shi, C. Wang and R. Li. Cancer-associated fibroblasts: Overview, progress, challenges, and directions. Cancer Gene Ther 2021, 28, 984–999. [CrossRef] [PubMed]
- Zhang, X., Y. Zhang, X. Qiu, J. Cai, Z. Yang and F. Song. Extracellular vesicles derived from lung cancer cells induce transformation of normal fibroblasts into lung cancer-associated fibroblasts and promote metastasis of lung cancer by delivering lncrna hotair. Stem Cells Int 2022, 2022, 3805013. [CrossRef] [PubMed]
- Shoucair, I., F. Weber Mello, J. Jabalee, S. Maleki and C. Garnis. The role of cancer-associated fibroblasts and extracellular vesicles in tumorigenesis. Int J Mol Sci 2020, 21, 103390/ijms21186837.
- Öhlund, D., E. Elyada and D. Tuveson. Fibroblast heterogeneity in the cancer wound. J Exp Med 2014, 211, 1503–1523. [CrossRef] [PubMed]
- Purcell, J. W., S. G. Tanlimco, J. Hickson, M. Fox, M. Sho, L. Durkin, T. Uziel, R. Powers, K. Foster, T. McGonigal, et al. Lrrc15 is a novel mesenchymal protein and stromal target for antibody-drug conjugates. Cancer Res 2018, 78, 4059–4072. [CrossRef]
- Dominguez, C. X., S. Müller, S. Keerthivasan, H. Koeppen, J. Hung, S. Gierke, B. Breart, O. Foreman, T. W. Bainbridge, A. Castiglioni, et al. Single-cell rna sequencing reveals stromal evolution into lrrc15(+) myofibroblasts as a determinant of patient response to cancer immunotherapy. Cancer Discov 2020, 10, 232–253. [CrossRef]
- Available online: https://fibroxplorer.com/.
- Buechler, M. B., R. N. Pradhan, A. T. Krishnamurty, C. Cox, A. K. Calviello, A. W. Wang, Y. A. Yang, L. Tam, R. Caothien, M. Roose-Girma, et al. Cross-tissue organization of the fibroblast lineage. Nature 2021, 593, 575–579. [CrossRef]
- Givel, A. M., Y. Kieffer, A. Scholer-Dahirel, P. Sirven, M. Cardon, F. Pelon, I. Magagna, G. Gentric, A. Costa, C. Bonneau, et al. Mir200-regulated cxcl12β promotes fibroblast heterogeneity and immunosuppression in ovarian cancers. Nat Commun 2018, 9, 1056. [CrossRef]
- Sebastian, A., N. R. Hum, K. A. Martin, S. F. Gilmore, I. Peran, S. W. Byers, E. K. Wheeler, M. A. Coleman and G. G. Loots. Single-cell transcriptomic analysis of tumor-derived fibroblasts and normal tissue-resident fibroblasts reveals fibroblast heterogeneity in breast cancer. Cancers (Basel) 2020, 12, 103390/cancers12051307.
- Bernard, V., A. Semaan, J. Huang, F. A. San Lucas, F. C. Mulu, B. M. Stephens, P. A. Guerrero, Y. Huang, J. Zhao, N. Kamyabi, et al. Single-cell transcriptomics of pancreatic cancer precursors demonstrates epithelial and microenvironmental heterogeneity as an early event in neoplastic progression. Clin Cancer Res 2019, 25, 2194–2205. [CrossRef]
- Zhang, Q., Y. Wang, C. Xia, L. Ding, Y. Pu, X. Hu, H. Cai and Q. Hu. Integrated analysis of single-cell rna-seq and bulk rna-seq reveals distinct cancer-associated fibroblasts in head and neck squamous cell carcinoma. Ann Transl Med 2021, 9, 1017. [CrossRef]
- Schwörer, S., F. V. Cimino, M. Ros, K. M. Tsanov, C. Ng, S. W. Lowe, C. Carmona-Fontaine and C. B. Thompson. Hypoxia potentiates the inflammatory fibroblast phenotype promoted by pancreatic cancer cell-derived cytokines. Cancer Res 2023, 83, 1596–1610. [CrossRef]
- Naito, Y., Y. Yamamoto, N. Sakamoto, I. Shimomura, A. Kogure, M. Kumazaki, A. Yokoi, M. Yashiro, T. Kiyono, K. Yanagihara, et al. Cancer extracellular vesicles contribute to stromal heterogeneity by inducing chemokines in cancer-associated fibroblasts. Oncogene 2019, 38, 5566–5579. [CrossRef]
- Mizutani, Y., H. Kobayashi, T. Iida, N. Asai, A. Masamune, A. Hara, N. Esaki, K. Ushida, S. Mii, Y. Shiraki, et al. Meflin-positive cancer-associated fibroblasts inhibit pancreatic carcinogenesis. Cancer Res 2019, 79, 5367–5381. [CrossRef]
- Li, H., E. T. Courtois, D. Sengupta, Y. Tan, K. H. Chen, J. J. L. Goh, S. L. Kong, C. Chua, L. K. Hon, W. S. Tan, et al. Reference component analysis of single-cell transcriptomes elucidates cellular heterogeneity in human colorectal tumors. Nat Genet 2017, 49, 708–718. [CrossRef] [PubMed]
- Bartoschek, M., N. Oskolkov, M. Bocci, J. Lövrot, C. Larsson, M. Sommarin, C. D. Madsen, D. Lindgren, G. Pekar, G. Karlsson, et al. Spatially and functionally distinct subclasses of breast cancer-associated fibroblasts revealed by single cell rna sequencing. Nat Commun 2018, 9, 5150. [CrossRef] [PubMed]
- albo, P. M., Jr., X. Zang and D. Zheng. Molecular features of cancer-associated fibroblast subtypes and their implication on cancer pathogenesis, prognosis, and immunotherapy resistance. Clin Cancer Res 2021, 27, 2636–2647. [CrossRef] [PubMed]
- Hu, C., Y. Zhang, C. Wu and Q. Huang. Heterogeneity of cancer-associated fibroblasts in head and neck squamous cell carcinoma: Opportunities and challenges. Cell Death Discov 2023, 9, 124. [CrossRef] [PubMed]
- Song, H., C. Lou, J. Ma, Q. Gong, Z. Tian, Y. You, G. Ren, W. Guo, Y. Wang, K. He, et al. Single-cell transcriptome analysis reveals changes of tumor immune microenvironment in oral squamous cell carcinoma after chemotherapy. Front Cell Dev Biol 2022, 10, 914120. [CrossRef] [PubMed]
- Wang, Z., H. Zhang, Y. Zhai, F. Li, X. Shi and M. Ying. Single-cell profiling reveals heterogeneity of primary and lymph node metastatic tumors and immune cell populations and discovers important prognostic significance of ccdc43 in oral squamous cell carcinoma. Front Immunol 2022, 13, 843322. [CrossRef] [PubMed]
- Costea, D. E., A. Hills, A. H. Osman, J. Thurlow, G. Kalna, X. Huang, C. Pena Murillo, H. Parajuli, S. Suliman, K. K. Kulasekara, et al. Identification of two distinct carcinoma-associated fibroblast subtypes with differential tumor-promoting abilities in oral squamous cell carcinoma. Cancer Res 2013, 73, 3888–3901. [CrossRef] [PubMed]
- Yang, W., S. Zhang, T. Li, Z. Zhou and J. Pan. Single-cell analysis reveals that cancer-associated fibroblasts stimulate oral squamous cell carcinoma invasion via the tgf-β/smad pathway. Acta Biochim Biophys Sin (Shanghai) 2022, 55, 262–273. [CrossRef] [PubMed]
- Patel, A. K., K. Vipparthi, V. Thatikonda, I. Arun, S. Bhattacharjee, R. Sharan, P. Arun and S. Singh. A subtype of cancer-associated fibroblasts with lower expression of alpha-smooth muscle actin suppresses stemness through bmp4 in oral carcinoma. Oncogenesis 2018, 7, 78. [CrossRef]
- Bienkowska, K. J., C. J. Hanley and G. J. Thomas. Cancer-associated fibroblasts in oral cancer: A current perspective on function and potential for therapeutic targeting. Front Oral Health 2021, 2, 686337. [CrossRef]
- Almangush, A., I. Heikkinen, N. Bakhti, L. K. Mäkinen, J. H. Kauppila, M. Pukkila, J. Hagström, J. Laranne, Y. Soini, L. P. Kowalski, et al. Prognostic impact of tumour-stroma ratio in early-stage oral tongue cancers. Histopathology 2018, 72, 1128–1135. [CrossRef] [PubMed]
- Hendry, S., R. Salgado, T. Gevaert, P. A. Russell, T. John, B. Thapa, M. Christie, K. van de Vijver, M. V. Estrada, P. I. Gonzalez-Ericsson, et al. Assessing tumor-infiltrating lymphocytes in solid tumors: A practical review for pathologists and proposal for a standardized method from the international immuno-oncology biomarkers working group: Part 2: Tils in melanoma, gastrointestinal tract carcinomas, non-small cell lung carcinoma and mesothelioma, endometrial and ovarian carcinomas, squamous cell carcinoma of the head and neck, genitourinary carcinomas, and primary brain tumors. Adv Anat Pathol 2017, 24, 311–335. [CrossRef]
- Froeling, F. E., C. Feig, C. Chelala, R. Dobson, C. E. Mein, D. A. Tuveson, H. Clevers, I. R. Hart and H. M. Kocher. Retinoic acid-induced pancreatic stellate cell quiescence reduces paracrine wnt-β-catenin signaling to slow tumor progression. Gastroenterology 2011, 141, 1486–97, 97.e1-14. [CrossRef] [PubMed]
- Li, Z. L., H. L. Zhang, Y. Huang, J. H. Huang, P. Sun, N. N. Zhou, Y. H. Chen, J. Mai, Y. Wang, Y. Yu, et al. Autophagy deficiency promotes triple-negative breast cancer resistance to t cell-mediated cytotoxicity by blocking tenascin-c degradation. Nat Commun 2020, 11, 3806. [CrossRef] [PubMed]
- Cazet, A. S., M. N. Hui, B. L. Elsworth, S. Z. Wu, D. Roden, C. L. Chan, J. N. Skhinas, R. Collot, J. Yang, K. Harvey, et al. Targeting stromal remodeling and cancer stem cell plasticity overcomes chemoresistance in triple negative breast cancer. Nat Commun 2018, 9, 2897. [CrossRef] [PubMed]
- Mao, X., J. Xu, W. Wang, C. Liang, J. Hua, J. Liu, B. Zhang, Q. Meng, X. Yu and S. Shi. Crosstalk between cancer-associated fibroblasts and immune cells in the tumor microenvironment: New findings and future perspectives. Mol Cancer 2021, 20, 131. [CrossRef] [PubMed]
- Özdemir, B. C., T. Pentcheva-Hoang, J. L. Carstens, X. Zheng, C. C. Wu, T. R. Simpson, H. Laklai, H. Sugimoto, C. Kahlert, S. V. Novitskiy, et al. Depletion of carcinoma-associated fibroblasts and fibrosis induces immunosuppression and accelerates pancreas cancer with reduced survival. Cancer Cell 2015, 28, 831–833. [CrossRef] [PubMed]
- Rhim, A. D., P. E. Oberstein, D. H. Thomas, E. T. Mirek, C. F. Palermo, S. A. Sastra, E. N. Dekleva, T. Saunders, C. P. Becerra, I. W. Tattersall, et al. Stromal elements act to restrain, rather than support, pancreatic ductal adenocarcinoma. Cancer Cell 2014, 25, 735–747. [CrossRef]
- Lee, J. J., R. M. Perera, H. Wang, D. C. Wu, X. S. Liu, S. Han, J. Fitamant, P. D. Jones, K. S. Ghanta, S. Kawano, et al. Stromal response to hedgehog signaling restrains pancreatic cancer progression. Proc Natl Acad Sci U S A 2014, 111, E3091–100. [CrossRef] [PubMed]
- Biffi, G. and D. A. Tuveson. Diversity and biology of cancer-associated fibroblasts. Physiol Rev 2021, 101, 147–176. [CrossRef] [PubMed]
- Feig, C., J. O. Jones, M. Kraman, R. J. Wells, A. Deonarine, D. S. Chan, C. M. Connell, E. W. Roberts, Q. Zhao, O. L. Caballero, et al. Targeting cxcl12 from fap-expressing carcinoma-associated fibroblasts synergizes with anti-pd-l1 immunotherapy in pancreatic cancer. Proc Natl Acad Sci U S A 2013, 110, 20212–20217. [CrossRef]
- Kraman, M., P. J. Bambrough, J. N. Arnold, E. W. Roberts, L. Magiera, J. O. Jones, A. Gopinathan, D. A. Tuveson and D. T. Fearon. Suppression of antitumor immunity by stromal cells expressing fibroblast activation protein-alpha. Science 2010, 330, 827–830. [CrossRef]
- Lo, A., C. P. Li, E. L. Buza, R. Blomberg, P. Govindaraju, D. Avery, J. Monslow, M. Hsiao and E. Puré. Fibroblast activation protein augments progression and metastasis of pancreatic ductal adenocarcinoma. JCI Insight 2017, 2. [CrossRef] [PubMed]
- Peltier, A., R. D. Seban, I. Buvat, F. C. Bidard and F. Mechta-Grigoriou. Fibroblast heterogeneity in solid tumors: From single cell analysis to whole-body imaging. Semin Cancer Biol 2022, 86, 262–272. [CrossRef] [PubMed]
- Fridman, W. H., F. Pagès, C. Sautès-Fridman and J. Galon. The immune contexture in human tumours: Impact on clinical outcome. Nat Rev Cancer 2012, 12, 298–306. [CrossRef] [PubMed]
- Jimenez, D. G., A. Sobti, D. Askmyr, C. Sakellariou, S. C. Santos, S. Swoboda, O. Forslund, L. Greiff and M. Lindstedt. Tonsillar cancer with high cd8(+) t-cell infiltration features increased levels of dendritic cells and transcriptional regulation associated with an inflamed tumor microenvironment. Cancers 2021, 13. [CrossRef] [PubMed]
- Almangush, A., I. O. Bello, I. Heikkinen, J. Hagström, C. Haglund, L. P. Kowalski, P. Nieminen, R. D. Coletta, A. A. Mäkitie, T. Salo, et al. Stromal categorization in early oral tongue cancer. Virchows Arch 2021, 478, 925–932. [CrossRef] [PubMed]
- Harper, J. and R. C. Sainson. Regulation of the anti-tumour immune response by cancer-associated fibroblasts. Semin Cancer Biol 2014, 25, 69–77. [CrossRef] [PubMed]
- Liu, T., C. Han, S. Wang, P. Fang, Z. Ma, L. Xu and R. Yin. Cancer-associated fibroblasts: An emerging target of anti-cancer immunotherapy. J Hematol Oncol 2019, 12, 86. [CrossRef] [PubMed]
- Karakasheva, T. A., E. W. Lin, Q. Tang, E. Qiao, T. J. Waldron, M. Soni, A. J. Klein-Szanto, V. Sahu, D. Basu, S. Ohashi, et al. Il-6 mediates cross-talk between tumor cells and activated fibroblasts in the tumor microenvironment. Cancer Res 2018, 78, 4957–4970. [CrossRef]
Figure 1.
CAF-related therapeutics.
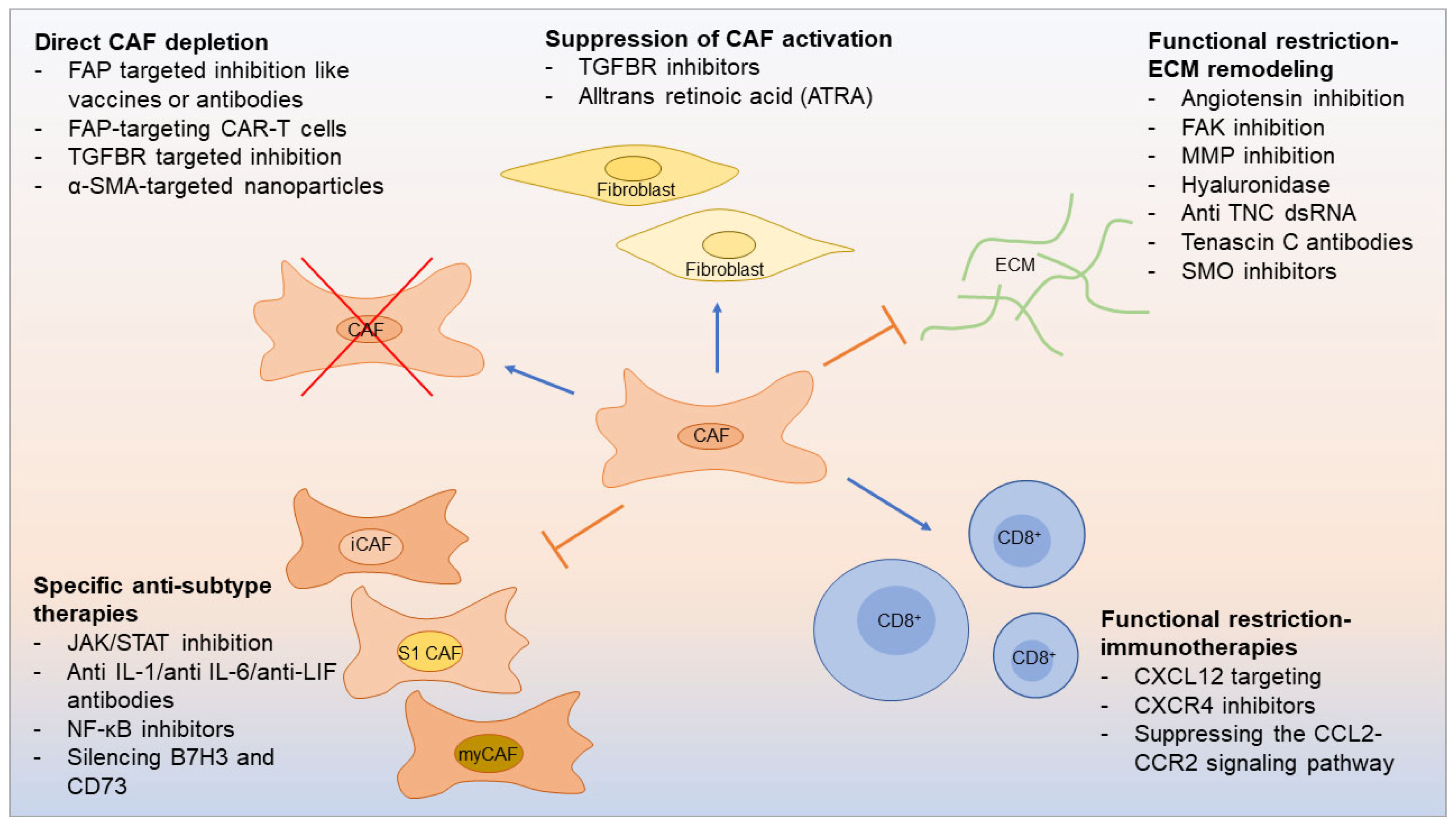
Table 1.
Differences in CAF subtypes across malignancies.
| Cancer form | CAF subtype | Subtyping method | Reference | Year |
|---|---|---|---|---|
| Breast | CAF-S1, -S2, -S3, -S4 | Flow cytometry and IHC | [6,7] | 2018, 2020 |
| myCAF, iCAF, apCAF. Six clusters | ScRNA-seq | [55] | 2020 | |
| vCAF, mCAF, dCAF | ScRNA-seq, IHC | [62] | 2018 | |
| Ovarian | CAF-S1, -S2, -S3, -S4 | Flow cytometry and IHC | [54] | 2018 |
| PDAC * | myCAF, iCAF | IF, IHC, flow cytometry, RNA in situ hybridization, RNA-seq | [10] | 2017 |
| myCAF, iCAF, apCAF | RNA-seq, RNA in situ hybridization, flow cytometry, IHC | [9] | 2019 | |
| myCAF, iCAF | ScRNA-seq | [8] | 2019 | |
| myCAF, iCAF | Single-cell digital microdissection | [56] | 2019 | |
| myCAF, iCAF | Flow cytometry | [58] | 2023 | |
| DGC * | myCAF, iCAF | IF | [59] | 2019 |
| ICC * | vCAF, mCAF, iCAF, apCAF, eCAF, lipofibroblast | ScRNA-seq | [13] | 2020 |
| HCC * | CAF_Port, CAF_HSC, CAF_VSMC | Proteomics, scRNA-seq | [11] | 2022 |
| CRC * | CAF-A, CAF-B (myofibroblast markers) | ScRNA-seq | [61] | 2017 |
| HNSCC * | CAF cluster 0-8 (cluster 4: apCAF) | ScRNA-seq | [57] | 2021 |
| OSCC * | iCAF, apCAF, myCAF | ScRNA-seq | [65] | 2022 |
| iCAF, myo-CAF (mCAF) | ScRNA-seq | [66] | 2022 | |
| CAF-N, CAF-D | RT PCR | [67] | 2013 | |
| Myofibroblast (mCAF), iCAF | ScRNA-seq | [68] | 2022 | |
| Melanoma, HNSCC, LC * | myCAF, dCAF, iCAF, iCAF-2, nCAF, pCAF | ScRNA-seq | [63] | 2021 |
| Gingivobuccal OC * | C1-type CAF, C2-type CAF | Gene expression microarray | [69] | 2018 |
* Pancreatic ductal adenocarcinoma (PDAC), diffuse-type gastric cancer (DGC), intrahepatic cholangiocarcinoma (ICC), hepatocellular carcinoma (HCC), colorectal cancer (CRC), head and neck squamous cell carcinoma (HNSCC), oral squamous cell carcinoma (OSCC), lung cancer (LC), oral cancer (OC).
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



