
Submitted:
03 December 2023
Posted:
05 December 2023
You are already at the latest version
Abstract
The recent rise in ocean temperatures, accompanied by other environmental changes, has notably increased the occurrence and spread of diseases in Octocorallia, of which many species are integral to shallow tropical and subtropical coral reef ecosystems. This study focuses on the understanding of these diseases, which has been largely limited to symptomatic descriptions, with clear etiological factors identified in only a fraction of cases. A key example is the multifocal purple spots syndrome (MPSS) affecting the common Caribbean octocoral sea fan Gorgonia ventalina, linked to the gall-forming copepods of the genus Sphaerippe, a member of the widespread family Lamippidae. The specialized nature of these copepods as endoparasites in octocorals suggests the potential for the discovery of similar diseases across this host spectrum. Our investigation employed four molecular markers to study disease hotspots in Saint Eustatius, Curaçao, northwest and southwest Cuba, and Bonaire. This led to the discovery of a group of copepod species in these varied Caribbean locations. Importantly, these species are morphologically indistinguishable through traditional methods, challenging established taxonomic approaches. The observed diversity of symbionts, despite the host species' genetic uniformity, is likely due to variations in larval dispersal mechanisms. Our phylogenetic analyses confirmed that the Lamippidae copepods belong to the order Poecilostomatoida (Copepoda) and revealed their sister group relationship with the Anchimolgidae, Rhynchomolgidae, and Xarifiidae clades, known for their symbiotic relationships with scleractinian corals. These results add to our understanding of the evolutionary and ecological interactions of copepods and their hosts, and the diseases they cause, important data in a changing climate.
Keywords:
parasites
; gorgonian octocorals
; integrative taxonomic approach
; phylogenetic analysis
; Caribbean region
; copepod crustaceans
; Lamippidae
1. Introduction
In an era characterized by the relentless advance of global climate change and the ever-escalating impact of anthropogenic pollution, the resilience of marine ecosystems is facing unprecedented challenges. Among the denizens of the oceanic realm, octocorals (Cnidaria, Octocorallia) are becoming increasingly susceptible to an onslaught of novel and potent infectious agents. This heightened vulnerability has manifested in a concerning surge of epidemic outbreaks, accompanied by substantial mass mortalities, with profound repercussions for the diversity and extent of octocoral populations (Weil et al., 2017; Calderón-Hernández et al., 2021; Becker et al., 2023).
Octocorals, with their intricate ecological roles, stand as keystones in the intricate web of life within shallow-water tropical and subtropical coral reef ecosystems. They function as essential contributors to ecosystem productivity and as indispensable havens and sustenance sources for a multitude of invertebrate species interconnected within their holobiont networks. The diminishment in octocoral numbers, however, has left an indelible mark on the overall composition, structural integrity, and functional dynamics of these vital marine ecosystems (Knowlton and Rohwer, 2003; Ritchie, 2006; Kimes et al., 2010; Burge et al., 2013; Weil et al., 2017).
Nowhere is this ecological crisis more evident than in the Caribbean region, a global epicenter of octocoral diversity. Home to approximately 70% of the world's infection-prone octocoral species, this region has borne the brunt of the mounting environmental pressures (Smith, Weil, 2004; Harvell et al., 2007; Ruiz-Moreno et al., 2012; Weil et al., 2017). Within the confines of this ecological hotspot yet our understanding of the infectious agents, transmission mechanisms, and the holistic impacts of these diseases remains confined to only eight (Weil et al., 2017). Of particular note, Gorgonia ventalina (Linnaeus, 1758), an endemic Caribbean sea fan (Bayer, 1961), exhibits susceptibility to nine of these diseases, distinguishing it as the most disease-prone species among octocorals. The multifocal purple spots syndrome (MPSS) that afflicts G. ventalina is particularly enigmatic, as it is incited by gall-forming copepods related to the genus Sphaerippe Grygier, 1980 within the family Lamippidae (Ivanenko et al., 2017; Korzhavina et al., 2019, 2021, 2023).
The Lamippidae family, notable for its extensive yet homogenous distribution, contains highly specialized obligate endoparasites, characterized by their notably modified body shapes and remarkable reduction of appendages (Humes 1957; Grygier, 1980, 1983; Williams et al., 2018; Korzhavina et al., 2019). These Lamippids find residence within the mesoglea, coenosarcal channels, or galls of octocorals on a global scale, to the exclusion of the Indian Ocean, and thrive across an astonishing depth range, spanning from the shallows to bathyal depths of 2258 m (Korzhavina et al., 2019). Presently, the scientific community has documented 54 lamippid species, with 115 recorded observations worldwide, while the Caribbean region has yielded only 14 of eight distinct species (Korzhavina et al., 2019, 2021, 2023). It is imperative to acknowledge, however, that a significant portion of lamippid species remains concealed, a consequence of the inherent challenges associated with their detection (Williams et al., 2018). This gap in our exploration of lamippid biology and virulence imposes substantial obstacles in our endeavors to model potential epizootic events and formulate effective control measures (Weil, 2004; Weil and Rogers, 2011; Kim and Rypien, 2015).
In light of these pressing ecological concerns, the principal objective of this study was to unravel the elusive causative agents responsible for the MPSS in Gorgonia ventalina. Through comprehensive investigation, this research aims to enrich our understanding of the intricate interactions between octocorals and their parasitic copepods. Ultimately, our efforts are aimed at contributing to the preservation and management of octocoral populations, striving to mitigate the dire consequences of mounting environmental challenges on these vital marine organisms.
2. Material and methods
Specimen collection
Our research involved the collection of 30 octocoral colonies from depths of 1–20 m across 18 different Caribbean reefs and three ecoregions (Hoeksema et al., 2017). These collections occurred at St. Eustatius (11 samples in 2015), Curaçao (four samples in 2017), southwest Cuba (four samples in 2019), northwest Cuba (nine samples in 2019), and Bonaire (four samples in 2019) (Table 1, Figure 1). The targeted octocoral colonies were Gorgonia ventalina (Alcyonacea: Gorgoniidae), selected through SCUBA diving by V.N. Ivanenko (V.N.I.) and O.A. Korzhavina (OAK). Underwater photographs of the hosts were taken by V.N.I. (Figures S1–S7), and the colonies were then carefully placed in plastic bags and transported to the surface.
Subsequent examinations of these colonies focused on identifying the presence of purple spots. The spotted tissues were preserved in 96% ethanol for later analysis. These spots were dissected to isolate copepods under a binocular microscope, Olympus SZX7 (Tokyo, Japan). The copepods were then prepared for morphological examination on glass slides in glycerine or placed in individual tubes for molecular analyses. Additionally, small sections of healthy coral tissue were separately preserved in tubes for DNA identification purposes. To complete the preservation process, bulk coral samples were stored in formalin.
Morphological examinations
In the study of copepods and their exuviae (exoskeletons) for light microscopy, post-DNA extraction, we employed the "hanging drop method" using Olympus CX41RF and Olympus BX 51 microscopes (Tokyo, Japan), following the methodologies outlined by Ivanenko and Defaye (2004) and Ivanenko et al., (2018). For scanning electron microscopy (SEM), we prepared copepods initially fixed in formalin by washing them in distilled water containing detergent. Subsequently, these specimens underwent a dehydration process involving two or three ethanol washes with increasing concentrations, followed by a transfer to acetone. The specimens were then dried using a critical point dryer (Hitachi HCP-2) (Tokyo, Japan). They were subsequently mounted on aluminium stubs using double-sided sticky tape and gold-coated in an IB-3. Imaging was carried out using a JEOL JSM-6380LA (Tokyo, Japan) and CamScan-S2 (Cambridge, United Kingdom), resulting in a total of 112 photographs. The electron microscopy was conducted as part of the research at the General Faculty Laboratory of Electron Microscopy, located within the Faculty of Biology at Lomonosov Moscow State University (MSU). These specimens have been added to the collection at the Biological Faculty of MSU for further study and reference.
DNA extraction
In our study, we implemented a refined non-destructive DNA extraction methodology, based on the protocol established by Porco et al., (2010) and further elaborated in Ivanenko et al., (2018). This process involved individual copepods, secured in 1.5 mL eppendorf tubes, from which 96% ethanol was carefully removed using a pipette with 200 μl tips. Each specimen was then treated with 50 μL of a specially formulated lysis solution (30 mM Tris-HCl, 20 mM EDTA, 1% SDS, 0.1 mg/ml proteinase K), with varying incubation times tailored to sample origin: two hours for Curaçao and St. Eustatius, 30 min for Cuba, and a variable 30 to 90 min for Bonaire samples, contingent upon specimen size. The incubation for samples was attentively monitored, concluding upon the sample's transition to a translucent state. Subsequently, the lysis solution was transferred to new tubes using a pipette equipped with slender 10 μl tips.
For the extraction of DNA from the lysis buffer, a silica-based DNA extraction kit (Diatom DNAprep 100, Isogene, Moscow, Russia) was utilized, following the manufacturer’s guidelines for fresh blood samples. The extracted DNA, in volumes of 20–30 μl, was then stored in appropriately labeled sterile tubes at -20°C for subsequent molecular analyses. Additionally, to preserve the morphological integrity of the copepod exuviae, a mixture of 100 μL of 1:1 ethanol-glycerol was applied.
The coral tissue DNA extraction commenced with the introduction of 300 μl of guanidine buffer, as per the standard instructional guidelines (Ivanenko et al., 2018). This was followed by a 2-hr incubation at 65°C, interspersed with vortex shaking at 30-min intervals, paralleling the protocol used for copepod DNA extraction (Ivanenko et al., 2018). A set of 91 samples was prepared for molecular study, encompassing 66 samples from lamippid copepods and 25 from their corresponding hosts.
DNA amplification and sequencing
Amplification of genetic material was conducted utilizing an Encyclo Plus PCR kit (Evrogen, Moscow, Russia) on BIO-RAD Dyad and BIO-RAD MJ Mini thermal cyclers. For analyses, three molecular markers were selected: mitochondrial cytochrome c-oxidase subunit I (COI), nuclear transcribed spacer 2 (ITS2), and nuclear ribosomal DNA (18S). These markers were chosen due to the availability of their sequences for most copepod families in existing databases. The COI marker was amplified using the forward copepod-specific primer LCO1490cop3 (Ivanenko et al., 2018) and the universal reverse primer jgH2198 (Geller et al., 2013). ITS2 amplification utilized a pair of copepod-specific primers, 58d-cop and 28-1-cop (Ivanenko et al., 2018), while universal primers 18d1 (Aleshin, unpublished) and Q39 (Medlin et al., 1988) were employed for the 18S marker. Octocoral DNA markers included ITS2 (Aguilar andSanchez, 2007) and msh1 (Sanchez et al., 2003), and were selected based on their previous application in octocoral phylogenetic studies and the availability of sequences for the genus Gorgonia Linnaeus, 1758 in databases. Details of the primers, amplified region lengths, and annealing temperatures are provided in Tables S1 and S2.
Post-amplification, PCR products were visualized through electrophoresis in 1% or 1.2% agarose gel. For processing, Shrimp Alkaline Phosphatase (SAP) and Exonuclease I E. coli enzymes were added to the PCR products, followed by incubation for one hour at 37°C and subsequent deactivation for 15 minutes at 85°C. Sequencing from both ends was performed using a BigDye Terminator reagent kit on ABI 3730 capillary sequencers at Evrogen (Moscow) following the manufacturer's instructions.
The resulting sequences were assembled and edited using Geneious 8.1. (Kearse et al., 2012) and subsequently stored in the GenBank sequence database (Tables S3 and S4). All sequences underwent verification using the NCBI BLAST tool, and protein-coding sequences (COI) were examined for an open reading frame (Altschul et al., 1990). We obtained sequences of 18S rDNA (1537-1658 bp) for three Sphaerippe samples, COI (618-695 bp) for 56 samples, and ITS2 (441-575 bp) for 59 samples. Additionally, sequences of ITS2 (215-240 bp) were retrieved for 20 Gorgonia ventalina samples, and msh1 (781-857 bp) for 21 samples. Sequence alignments were conducted using the MUSCLE algorithm in Geneious for monogenic alignments (Edgar, 2004) or MAFFT version 7 (Kuraku et al., 2013, Katoh et al., 2017) for concatenated alignments. The phylogenetic trees derived from these alignments are available for review at the TreeBASE online data exchange center.
DNA taxonomy
In this detailed phylogenetic study, we conducted Bayesian Analysis (BA) on both individual markers and a combined dataset for copepods, while Maximum Likelihood (ML) analysis was specifically applied to the concatenated alignment of these organisms. Throughout this process, uniform tree-building parameters were employed for all alignments, with the sole variation being the models of nucleotide evolution. These models were selected using MegaX for single-gene alignments and PartitionFinder for the concatenated datasets, in accordance with established protocols by Guindon et al. (2010) and Lanfear et al. (2012, 2016).
For the construction of BA phylogenetic trees, we utilized the CIPRES web interface (Ronquist et al., 2012). The procedure included runs over 25 million generations, employing four synchronous Markov Chain Monte Carlo (MCMC) chains and saving every 5000th tree. We excluded the initial 25% of trees from subsequent analyses as ‘burn-in’. The convergence of these analyses was monitored using Tracer v1.7.1 (Rambaut et al., 2018), with Effective Sample Sizes (ESS) for all parameters exceeding the threshold of 200 to ensure data reliability. Nodal support in the BA trees was assessed based on posterior probabilities, and the ML trees were constructed using the IQ-TREE web application (Trifinopoulos et al., 2016), with nodal supports determined via 1000 bootstrap replications (Felsenstein, 1985).
The COI alignment for copepods comprised 56 sequences, predominantly from Sphaerippe spp., augmented with lamippid sequences from Australia, while the ITS2 alignment included 59 sequences. Both alignments were modelled using the GTR + G + I model, identified as the most appropriate based on respective selection criteria. The concatenated alignment, encompassing COI and ITS2 sequences of lamippids, featured 66 sequences for tree construction and 51 sequences for Poisson Tree Processes (PTP) analysis. PartitionFinder was employed to recommend evolutionary models for this alignment's partitions, highlighting the complexity of the genetic data. Additionally, ITS2 and msh1 alignments of Gorgonia spp. octocorals included sequences from a range of Caribbean locations and GenBank, representing diverse species within the genus.
Species differentiation was performed using Automated Barcode Gap Detection (ABGD) and Poisson Tree Process (PTP), recognized for their efficacy in DNA taxonomy (Fontaneto et al., 2015). The ABGD analysis, executed separately for COI, ITS2, and msh1 markers, identified genetic distance gaps indicative of interspecific variability. The PTP analysis was applied to the BA trees for both COI, ITS2, and their combined datasets of Sphaerippe spp., as well as the msh1 dataset of Gorgonia spp., utilizing the bPTP online platform's standard parameters.
Host relationships and geographical isolation
In our research, we applied the DNAsp program to discern and segregate all haplotypes within our alignments, effectively removing any repetitive sequences. Additionally, DNAsp was utilized to compute Fu’s F parameter to assess genetic diversity and population dynamics.
For the analysis of haplotypes, we employed the Median Joining method via the PopArt program (Leigh and Bryant, 2015). Our dataset for this portion of the study consisted of 54 sequences in the COI alignment of Sphaerippe spp., and 57 sequences in the ITS2 alignment. The ITS2 alignment for sea fans of the genus Gorgonia encompassed 20 of our sequences and an additional three from GenBank. Similarly, the msh1 alignment included 21 sequences of Gorgonia ventalina alongside three sourced from GenBank (Table S5).
The selection of sampling points was derived from expedition data to St. Eustatius (2015), Curaçao (2017), Cuba (2019), and Bonaire (2019), as detailed in a database from a comprehensive review (Korzhavina et al., 2019). This approach not only enriched the geographical scope of our study but also facilitated the calculation of statistical parameters, including nucleotide diversity and Tajima’s D statistic (Tajima, 1989).
Molecular phylogenetic analyses
The phylogenetic positioning of Sphaerippe spp. within the copepod clade was ascertained through an analysis of the 18S rDNA alignment. This alignment incorporated 100 copepod sequences from GenBank, which included 53 species from Cyclopoida, 44 from Poecilostomatoida, and three from Misophrioida. Notably, the dataset also contained a sequence from the octocoral Junceella fragilis (AY962533.1), which was actually a lamippid sequence mistakenly categorized under the host name in GenBank. Additionally, four sequences from our samples were included, three of which were from different Caribbean regions representing Sphaerippe spp., and as well one lamippid from Lizard Island (Australia) (Table S6). The sequences utilized ranged from 564 to 1866 base pairs in length.
For model selection, the General Time Reversible model with Gamma distribution and Invariant sites (GTR+G+I) was determined as the most suitable using the Akaike Information Criterion with correction (AICc) in Mega X. Bayesian Analysis (BA) was executed with settings as previously mentioned, and convergence of the results was validated using Tracer v1.7.1 (Rambaut, Drummond, 2007). A Maximum Likelihood (ML) phylogenetic tree was constructed using the standard parameters.
Figures depicting these phylogenetic trees and their associated captions were edited only for clarity using Adobe Photoshop 21.2.9 and CorelDRAW.
3. Results
Observation of the purple galls on Gorgonia ventalina
We conducted a detailed examination of the easily detectable underwater purple galls on the sea fan. These galls predominantly appeared as isolated or, more frequently, aggregated gall-like growths. These formations, slightly thickened and diverse in structure, were primarily located on the lateral aspects of the stolons or, more typically, at the nodes of the sea fan's reticulate structure, as shown in Figure 2 and Figure S1–S7.
The dissection of these purple galls devoid of any apparent openings typically detected presence of one or rarely some more chambers containing spheroidal females, typically one and occasionally two per chamber (Figure 2). These females were often accompanied by a male, and in less frequent cases, two males. Some galls were contained elongated copepod stages, which were noticeably smaller than female and male.
The gall walls formed by the sea fans are including copious microscopic spherical capsules with a diameter of about 0.1 mm. Each capsule contained embryo covered by a membranous shell. This ranged from early-stage, undifferentiated round embryos to nearly fully formed nauplii, likely on the verge of hatching. The nauplii exhibited typical distinctive features, such as three pairs of anterior appendages, including a uniramous antennule and biramous antennae and mandibles, and have a slit-like oral opening devoid of an overlying labrum (Figure 2).
We uncovered the presence of yellowish, sclerotized structures within galls that housed living copepods and, intriguingly, in some galls where live copepods were not observed. These structures are identified as the exoskeletons of mummified copepods along with their spermatophores, seemingly isolated by Gorgonia ventalina. All these finding indicate to the complex biological and ecological interactions between copepods and their gorgonian hosts.
Morphological features of Sphaerippe spp. from the purple galls
The females are discernibly different from their male counterparts, primarily in their rounded body morphology accentuated by various projections (Figure 2). These females feature pronounced bulges and folds, dividing the body into distinct sections. Morphological features include a forward-directed conical rostrum, uniramous antennules and antennae, an oral cone, and two pairs of biramous, modified swimming legs located in the anterior portion of the body, complemented by caudal rami. A notable characteristic of the female copepods is the presence of elaborately developed modified setae on the first and second pairs of swimming legs and the caudal rami. These setae split at the base into clusters of long, slender projections. In contrast, the males are characterized by an elongated body shape, with a more extended rostrum. Their modified setae, similar to those of the females, are less developed in comparison.
The analysis of samples collected from different locations, employing both light and scanning electron microscopy, revealed a notable degree of variability among the specimens, even those inhabiting the same locale. The study did not yield any distinct diagnostic morphological features of copepods discernible through molecular methods.
Interspecies molecular diversity
In our comprehensive phylogenetic analyses using Bayesian Analysis (BA) on the COI alignment of Lamippidae copepods, we discerned a bifurcation of Caribbean lamippids into three monophyletic groups. This division was represented by a first clade consisting of samples from Bonaire and Curaçao (both Southern Caribbean), and St. Eustatius (Eastern Caribbean), supported robustly with a probability of 1. The second and third clades, encompassing samples from southwest and northwest Cuba (Greater Antilles), respectively, had supports of 0.76 and 1, respectively (Figure 3). Intriguingly, the northwestern Cuban clade was phylogenetically allied as a sister group to the Eastern + Southern Caribbean clade in the BA framework. Employing the Automatic Barcode Gap Discovery (ABGD) method, we identified three distinct species groups within the COI alignment of Sphaerippe spp. corresponding to these three clades, with intraspecific distances ranging from 0.16 to 0.31. The Poisson Tree Processes (PTP) model further corroborated this finding, delineating three potential species in the dataset: Sphaerippe sp. 1 from St. Eustatius and Curaçao (support 0.964), Sphaerippe sp. 2 from northwest Cuba (support 0.977), and Sphaerippe sp. 3 from southwest Cuba (support 0.966).
Similarly, the BA phylogenetic tree based on the ITS2 alignment of lamippids recovered a split into Cuban (Greater Antilles) and Eastern + Southern Caribbean monophyletic clades (Figure 4). The ABGD analysis, considering prior intraspecific distances from 0.06 to 0.19, and the PTP model, with supports of 1 and 0.99, respectively, confirmed the existence of two distinct species groups within the ITS2 alignment of Sphaerippe spp.
Moreover, the BA and Maximum Likelihood (ML) phylogenetic trees, derived from the copepods’ concatenated alignment (COI + ITS2), revealed three distinct monophyletic Caribbean clades (Figure S8). The PTP model applied to this dataset also identified three species: Sphaerippe sp. 1 from the islands of St. Eustatius and Curaçao (support 0.847), Sphaerippe sp. 2 from southwest Cuba (support 0.84), and Sphaerippe sp. 3 from northwest Cuba (support 0.83).
The ITS2 alignment of octocoral samples was characterized by minimal polymorphism, indicating the probable conspecific nature of all samples. The GenBank sequences of Gorgonia ventalina and Gorgonia flabellum Linnaeus, 1758 revealed only two polymorphic substitutions. The msh1 octocoral alignment presented a similar scenario, with the exception of samples 19–32, which showed nine nucleotide substitutions. Both the ABGD and PTP analyses suggested four species in this dataset: Pseudopterogorgia bipinnata (Verrill, 1864), Gorgonia mariae Bayer, 1961, the distinct samples 19–32, and a collective group comprising all other samples along with G. ventalina and G. flabellum. In the PTP analysis, samples 19-32 had a support of 0.79, while the aggregate group, including G. ventalina and G. flabellum, had a support of 0.64.
Intraspecific molecular diversity
In the phylogenetic investigation, we employed a haploweb constructed from 54 COI sequences of Sphaerippe spp., revealing a clear division into a group of two or three species (Figure 4). The analysis of 36 individuals of Sphaerippe sp. 1, utilizing the DNAsp program, identified seven distinct haplotypes. These haplotypes segregated into two geographic clusters, one encompassing the islands of Bonaire with Curaçao in the Southern Caribbean ecoregion and the other St. Eustatius in the Eastern Caribbean ecoregion. Each neighbouring haplotype was differentiated by a single nucleotide substitution, with the most predominant haplotype observed in St. Eustatius, exhibiting a nucleotide distance of n = 1.61. In the dataset of Sphaerippe sp. 2, comprising ten specimens, DNAsp analysis delineated six haplotypes, with a nucleotide distance of n = 1.533. Furthermore, the analysis of eight individuals of Sphaerippe sp. 3 identified three haplotypes, showing a nucleotide distance of n = 1.107.
The ITS2 haploweb, based on the alignment of 57 sequences of Sphaerippe spp., demonstrated divergence into three species (Figure 4). The group of 36 specimens from Bonaire, Curaçao, and St. Eustatius collectively formed a single haplotype, exhibiting identical sequences except for variations in microsatellite repeats. Consequently, nucleotide distances were not computed for this group. In the Cuban Sphaerippe dataset, encompassing 21 sequences, the DNAsp program identified six haplotypes with a nucleotide distance of n = 1.867.
Tajima’s D and Fu’s F statistics (Fu, 1997) for all species of Sphaerippe spp. and both DNA markers showed no significant deviations from zero (p<0.05) (Table 2).
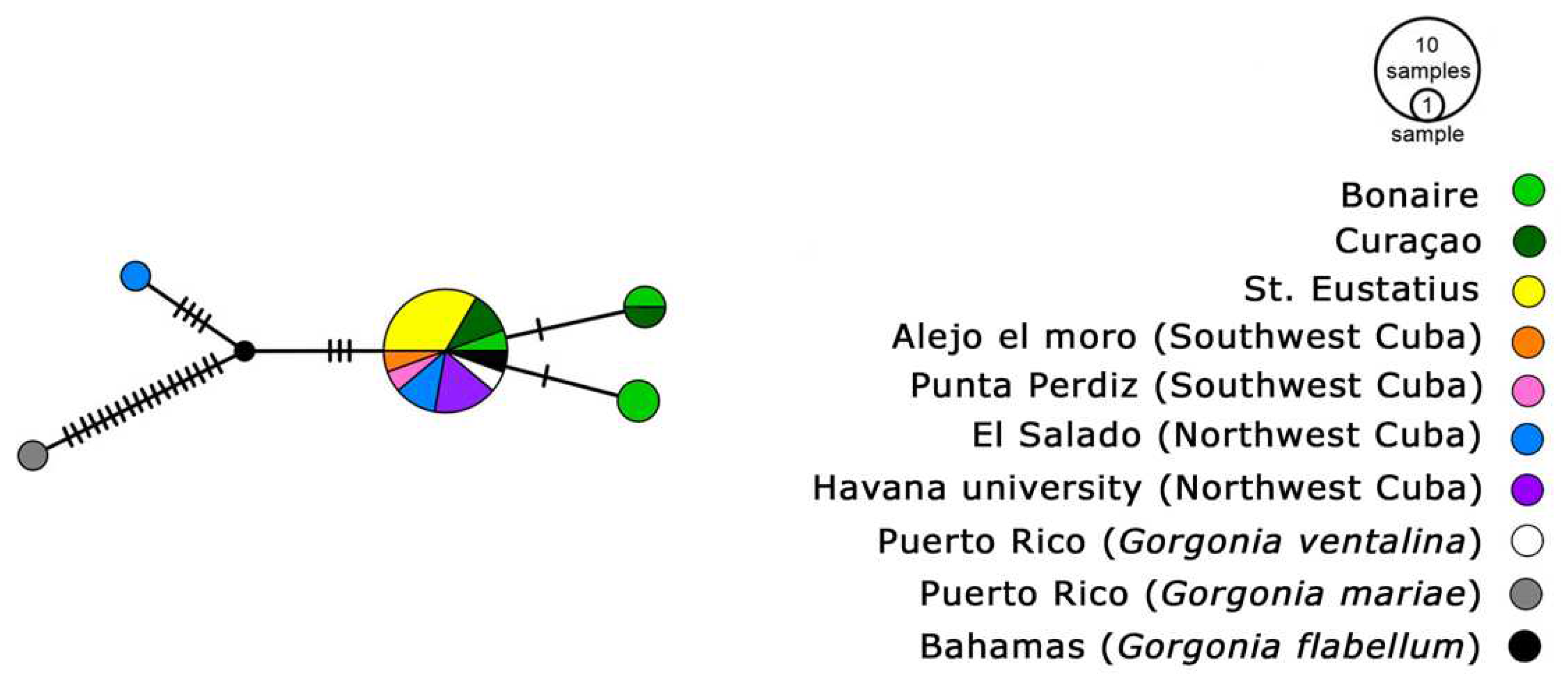
The haploweb analysis for ITS2 corals of Gorgonia Linnaeus, 1758, revealed two haplotypes: one exclusive to Gorgonia mariae Bayer, 1961, and the other inclusive of all our samples, G. flabellum, and G. ventalina. The msh1 haploweb for G. ventalina indicated a division into two species, one of which formed two distinct haplotypes (Figure 5), with a nucleotide distance of n = 4. For this species, the values of Tajima’s D and Fu’s F statistics exhibited significant differences from zero (-1.92207 and 2.499).
Figure 5.
msh1 haplotype diversity in G. ventalina across the Caribbean. Circle sizes represent haplotype frequency, notches indicate genetic substitutions. Colors denote geographic origin: yellow for St. Eustatius, light and dark green for Bonaire and Curaçao, orange and pink for Southwest Cuba, blue and purple for Northwest Cuba, white and grey for Puerto Rico samples (GenBank), and black for Bahamas samples (GenBank).
Figure 5.
msh1 haplotype diversity in G. ventalina across the Caribbean. Circle sizes represent haplotype frequency, notches indicate genetic substitutions. Colors denote geographic origin: yellow for St. Eustatius, light and dark green for Bonaire and Curaçao, orange and pink for Southwest Cuba, blue and purple for Northwest Cuba, white and grey for Puerto Rico samples (GenBank), and black for Bahamas samples (GenBank).
Phylogeny reconstruction
Phylogenetic analyses utilizing Maximum Likelihood (ML) and Bayesian Analysis (BA) based on the 18S alignment robustly positioned the genus Sphaerippe within the suborder Poecilostomatoida (Cyclopoida). These results had 100% support probability (Figures 6 and S9 and S10) Within this phylogenetic framework, the Lamippidae family was observed to cluster with the family groups Anchimolgidae, Rhynchomolgidae, Sabelliphilidae Xarifiidae, and, with strong support scores of 100 and 1 in ML and BA trees, respectively. Moreover, the clade comprising Anchimolgidae, Rhynchomolgidae, and Xarifiidae emerged as a sister group to Sphaerippe spp., with this relationship receiving high support values of 98 and 0.98 in the ML and BA analyses, respectively.
Figure 6.
Phylogenetic trees based on 18S alignment using the GTR + G + I nucleotide evolution model. Node numbers indicate posterior probabilities (Bayesian) and bootstrap supports (Maximum Likelihood), with asterisks (*) marking nodes with differing topologies in ML and BA trees. Circle colors represent copepod host taxa, and red frame highlights the position of Lamippidae within the Copepoda.
Figure 6.
Phylogenetic trees based on 18S alignment using the GTR + G + I nucleotide evolution model. Node numbers indicate posterior probabilities (Bayesian) and bootstrap supports (Maximum Likelihood), with asterisks (*) marking nodes with differing topologies in ML and BA trees. Circle colors represent copepod host taxa, and red frame highlights the position of Lamippidae within the Copepoda.

4. Discussion
Morphological examination of copepod specimens
The findings of this research underscore the imperative for targeted investigations focusing specifically on male specimens or females that have recently molted. Such a focused approach is essential for the delineation of definitive morphological characteristics, which, when clearly established, can be effectively integrated into molecular diagnostic protocols. This multifaceted methodology is expected to substantially enhance the accuracy and precision of species identification in future studies. Our investigation did not yield significant diagnostic markers that could facilitate a refined diagnosis at the genus level, a limitation stemming from the currently inadequate detail in the existing genus descriptions (Grygier, 1980; Williams et al., 2018; Korzhavina et al., 2021). Furthermore, the study revealed an absence of significant morphological variation among specimens from different Caribbean regions. This observation could largely be attributed to the extensive morphological variability inherent in the female specimens of the genus, particularly noted in the reduction of appendages and the challenges in preserving structural details during gall dissection and analysis.
The significant taxonomic ambiguity of the Lamippidae family, primarily due to the absence of distinct morphological features for reliable species differentiation, is a well-documented challenge in the scientific community (Korzhavina et al., 2021). Our research underscores the necessity of detailed specimen analysis, emphasizing the inclusion of both male and female specimens for accurate species identification. This is crucial given the notable morphological diversity observed between different genders and developmental stages within species, adding complexity to establishing definitive diagnostic characters for species delineation. To address these taxonomic challenges effectively and enhance genus-level diagnoses within the Lamippidae family, we advocate a dual-methodological approach, combining molecular techniques with detailed morphological analyses. This strategy aims to achieve a more refined and comprehensive taxonomic classification for the family, resolving existing taxonomic complexities and deepening our understanding of the phylogenetic and evolutionary relationships in this diverse and underexplored group of copepods.
Our observation of the dissolution of all copepod exuviae during the DNA extraction process suggests a potential weakening of the chitinous layer in these copepods. This finding deviates from expected results based on previous studies that successfully conserved copepod exoskeletons (Conradi et al., 2018; Ivanenko et al., 2018; Shelyakin et al., 2018). Possible explanations for this phenomenon include a thinned chitin exoskeleton, characteristic of endoparasitic adaptations, or an altered chemical composition of the exoskeleton in Lamippidae copepods. The substitution of chitin with a more elastic protein, such as resilin, is another speculative explanation (Michels et al., 2016). This unexpected result prompts the need for further in-depth examination of the exoskeletal structure of these unique copepods.
Molecular phylogenetic divergence
This study delineated copepod populations associated with the octocoral genus Gorgonia into three distinct phylogenetic clades, each endemic to specific geographic areas within the Caribbean. These clades are well supported and genetically distant enough to warrant the recognition of three novel, hitherto undescribed Sphaerippe species. These findings, particularly the values of Tajima’s D and Fu’s F statistics, imply a dynamic state of evolutionary flux within these populations, marked by an imbalance between genetic drift and mutations. The results are indicative of an extensive coevolutionary process between Sphaerippe copepods and their hosts. One clade, originating from the Eastern and Southern Caribbean ecoregions, predominantly inhabits the vicinity of the islands of St. Eustatius, Curaçao, and Bonaire, spanning approximately 900 km (Figure 1a). Notably, this clade exhibited minimal genetic variation over these considerable distances. The phylogeographic similarity between these different locations is not unique, since it can also be found in reef fishes (Taylor and Hellberg, 2006; Rocha et al., 2008). This can be explained by a connectivity caused by the westward Caribbean Current from the Atlantic, entering the eastern Caribbean through the Lesser Antilles Arc and flowing towards the southern Caribbean (Rocha et al., 2008).
Conversely, the Western Caribbean clades showed a distinct separation based on mitochondrial DNA sequences, with one subgroup associated with Cuba’s southern coastline and the other with its northern counterpart. Intriguingly, analyses of nuclear internal transcribed spacer (ITS2) regions in these copepods revealed genetic intermingling between some specimens from the southern clade with those from the northern clade, indicative of gene flow between these two distinct species. The occurrence of hybridization, particularly between Sphaerippe spp. from the disparate northern and southern Cuban coasts, suggests a lack of prezygotic morphological barriers to reproduction. This observation aligns with the hypothesis of larval dispersal facilitated by the currents of the Yucatan Strait, underscoring the significant influence of oceanographic factors on the evolutionary trajectory and geographic distribution of these Caribbean Sphaerippe species.
The taxonomic classification and determination of the phylogenetic order of copepods within the Lamippidae family, particularly considering their modified morphology and appendage reduction, has been long uncertain. These studies were complicated by the distinctive morphological traits of the Lamippidae, which historically led to their varied classification into orders such as Siphonostomatoida, Cyclopoida, and Poecilostomatoida (Korzhavina et al., 2021). Our phylogenetic analyses robustly place Lamippidae copepods, specialized endoparasites of octocorals (Octocorallia), within the order Poecilostomatoida (Korzhavina et al., 2021). This research additionally revealed a sister relationship between Lamippidae and families known as symbionts of scleractinian corals (Anchimoligidae, Rhynchomolgidae, and Xarifidae), known symbionts of scleractinian corals (Huys et al., 2006; Ferrari et al., 2010; Mikhailov, Ivanenko,2019, 2021). This phylogenetic arrangement not only underscores the evolutionary relationships within these taxa but also enhances the understanding of their systematic positions within the broader copepod lineage. Importantly, despite ongoing debates regarding the boundaries and validity of the orders Cyclopoida and Poecilostomatoida, which have yet to be conclusively resolved through molecular methods, a significant group of predominantly symbiotic copepod families within these orders appeared to represent a cohesive and well-diagnosable group within our analyses. This insight underlines the importance of continued molecular and morphological research to better understand the complexities of copepod taxonomy and their evolutionary relationships with various host taxa within marine ecosystems.
The Gorgonia sea fans analysed in our study were characterized by a range of morphological variations in colony branching. This diversity was subject to much discussion on its taxonomic meaning until the advent of molecular methods for identifying interspecies boundaries among closely related groups (Figure S1–S7) (Bayer, 1961; Sanchez and Wirshing 2005; McFadden et al., 2006). Our genetic analyses revealed that the sequences of most Gorgonia taxa were congruent in both the ITS2 and msh1 markers. Furthermore, Gorgonia collectively formed a monophyletic clade at the species level, which also included sequences of Gorgonia ventalina and G. flabellum. This finding underscores the limitations of current DNA markers in effectively distinguishing species within octocorals (McFadden et al., 2010). Given the impact of environmental factors on the morphological variability of corals (Sanchez et al., 2007, McFadden et al., 2010), and considering the genetic homogeneity of our Gorgonia samples, we classified all specimens within the species G. ventalina. An outlier in our analysis was sample 19-32, which, based on the msh1 marker, was distinct in both Maximum Likelihood (ML) and MrBayes phylogenetic trees. Sequences from this specimen did not cluster with either those of our specimens or those in GenBank, suggesting it may represent a significantly divergent msh1 haplotype. However, its concordance in ITS2 markers and general external morphology with other Gorgonia specimens indicates its probable affiliation with the same species as the rest of our specimens.
Geographical heterogeneity of parasite and host populations
In our study, we have discerned a conspicuous disparity in species differentiation of Sphaerippe among copepods across distinct Caribbean regions, accompanied by a comparatively restricted intraspecific variability in the composition of their host Gorgonia populations and other symbionts associated with the same host. (Figure 3, Figure 4 and Figure 5) (Andras et al., 2011, 2013). This pattern appears to be influenced by the relatively limited dispersal capability of both Sphaerippe and Gorgonia. Throughout our field research, it was recurrently noted that colonies afflicted with Multifocal Purple Spot Syndrome (MFPS) were often located in proximity to healthy sea fan colonies. This proximity may be indicative of the copepods' ability for self-infection within sea fan colonies and their active role in attracting dispersal stages to parts of the population already parasitized by these copepods.
Our hypothesis posits that the nauplii of Sphaerippe spp., which develop inside the gall, or their first copepodid stage, acting as a dispersal phase in many parasitic copepods, are responsible for rupturing the gall coverings. These nauplii then disseminate within the Gorgonia colony of the maternal gall and the infected host colony and may also spread to and infect adjacent sea fan colonies (Ivanenko et al., 2001; Ivanenko and Ferrari 2003; Cheng et al., 2013; Williams, Bunkley-Williams 2019). Contrasting with the copepods, the planktonic larvae of Gorgonia spp. probably exhibit a prolonged pelagic phase, suggesting a more effective dispersal capability (Andras et al., 2013, fig 2). The data obtained from our research corroborate findings from another Caribbean symbiont-host relationship involving the pea crab Dissodactylus primitivus Bouvier, 1917 and the sea urchin Meoma ventricosa (Lamarck, 1816) (Jossart, 2017). In this relationship, geographically separate populations of the symbiotic crab and a uniformity in the host population were observed (Jossart, 2017), underscoring the complexity of symbiotic interactions in marine ecosystems.
Coral diseases and the multifocal purple spot syndrome (MFPS)
Coral diseases, initially detected in the 1970s, are characterized by alterations in coral structures and functions, resulting from the intricate interplay among the corals, their environmental context, and various pathogenic agents (Martin et al., 1987; Work, Abey, 2006; Rogers, 2010; Weil et al., 2017; van de Water et al., 2018). With the advent of climate change, corals are increasingly subjected to physiological stressors, leading to compromised immune responses. This heightened vulnerability transforms previously innocuous agents into potential pathogens (Rosenberg and Ben-Haim, 2002; Lesser et al., 2007; Weil and Rogers, 2011; Shelyakin et al., 2018; van de Water et al., 2018). Research into coral pathologies is further complicated by the inaccessible nature of their habitats and the lack of universally accepted methodologies for diagnosing disease etiologies (Burge et al., 2012). As a result, the majority of current literature on coral diseases primarily focuses on symptomatology, often omitting detailed etiological information (Weil, 2004; Work and Abey, 2006; Weil 2011; Montano et al., 2020).
The multifocal purple spot syndrome (MFPS), identified in the widely distributed and shallow-water coral species Gorgonia ventalina in the Caribbean in 2005 (Harvell et al., 2007), is characterized by the presence of multiple purple swellings or galls on the octocoral colony. These galls are distinctively devoid of any openings (Ivanenko et al., 2017). Research into the pathology of these conspicuous galls has implicated organisms from the Labyrinthulomycetes group, particularly genera Aplanochytrium and Thraustochytrium (Burge et al., 2012, 2013). However, a more detailed anatomical investigation of G. ventalina specimens affected by MFPS revealed the presence of copepods from the genus Sphaerippe. Notably, galls that lacked external openings contained female copepods, occasionally with males, as well as numerous embryos, developing nauplii, and sizeable spermatophores (Ivanenko et al., 2015; Shelyakin et al., 2018; present observations). This new insight into the condition has introduced a nuanced perspective on the etiological factors of MFPS, complicating the accurate diagnosis and characterization of the syndrome in this widespread, shallow-water coral species in the Caribbean (Weill et al., 2017; Tracy et al., 2018; van de Water et al., 2018; present observations).
The etiological investigation of Multifocal Purple Spot Syndrome (MFPS) in Gorgonia ventalina necessitates a comprehensive experimental framework to elucidate the pathogenicity of coral-associated microorganisms. This approach is essential due to the current reliance on indirect evidence. A salient diagnostic characteristic of MFPS is the specific size and morphology of the lesions, signifying an initial immunological response of Gorgonia species aimed at mitigating pathogen proliferation. This response is evidenced by a change in the coloration of Gorgonia surface tissues, characterized by an abundance of purple sclerites, as reported in multiple studies (Petes et al., 2003; Burge et al., 2012; Ivanenko et al., 2017; van Weil et al., 2017; Shelyakin et al., 2018; de Water et al., 2018). Notably, the lesions associated with MFPS, typically small with smooth edges, are markedly distinct from other types of lesions that are larger, irregular in shape, and exhibit purple colouring at the edges, as commonly observed in sea fans (Weil and Hooten, 2008).
Furthermore, the spatial distribution of MFPS, governed by the transmission dynamics of the pathogen, requires further detailed examination. Extensive observational data from dives across different regions of the Caribbean Sea indicate a higher prevalence of MFPS in shallower waters, correlating with the presence of Sphaerippe copepods. This finding is contrasted by the deeper distribution of the Labyrinthulomycetes genera Aplanochytrium and Thraustochytrium, which are associated with similar disease manifestations in La Parguera Natural Reserve of the southwest coast of Puerto Rico (Burge et al., 2012; Tracy et al., 2018). The contrasting features between MFPS and diseases induced by other organisms suggest that copepods of the genus Sphaerippe are likely the principal pathogens of MFPS.
With regards to the life cycle of Sphaerippe copepods, following coral infestation, both male and female copepods consume coral tissue and undergo significant morphological transformations. Females develop into a spherical form, while males assume a seed-like shape, contained within the coral gall. This gall environment facilitates their growth, molting, and reproduction, as well as the development of numerous nauplii. The prevailing hypothesis posits that the emergence of copepods into the external environment occurs during the late naupliar or early copepodid stages, often leading to the rupture of coral tissues. Dissections of various galls have revealed instances where, despite the absence of living copepods, the galls contained only their exuviae and spermatophores, encapsulated in a dense yellowish substance, presumably secreted by the coral cells. This observation suggests that the lifespan of the female copepod may limit the duration of gall formation. Additionally, dissections have shown that in cases where galls, devoid of living copepods, contained only their exuviae, indicating that the manifestation period of galls is potentially constrained by the lifespan of the female copepod. The penetration of copepods into the coral and gall formation by the female likely occurs during a dispersive, immature stage of the copepod, either through the polyp or directly through the coral's covering. However, the precise mechanisms of this penetration and subsequent gall formation remain unexplored.
The scarcity of prior documentation of the distinct purple lesions characteristic of Multifocal purple spot syndrome (MFPS) in the shallow-water sea fans of the extensively studied Caribbean basin could be attributed to an oversight in scientific focus on this specific symptom. Alternatively, this absence might be indicative of a relatively recent emergence of MFPS in the Caribbean region, possibly driven by climatic changes over the last 25 years (Chollet et al., 2012). Observational studies have noted a significant 34% increase in the proportion of infected Gorgonia colonies relative to healthy ones within a seven-year period following the disease's identification (Weil and Rogers, 2010; Weil et al., 2014, 2016). Given the observed peak in disease prevalence during summer months (Weil and Rogers, 2010), it is reasonable to speculate that climatic shifts may play a role in the increased manifestation of MFPS, likely influenced by the presence of gall-inducing copepods (Rosenberg and Ben-Haim, 2002; Weil et al., 2011).
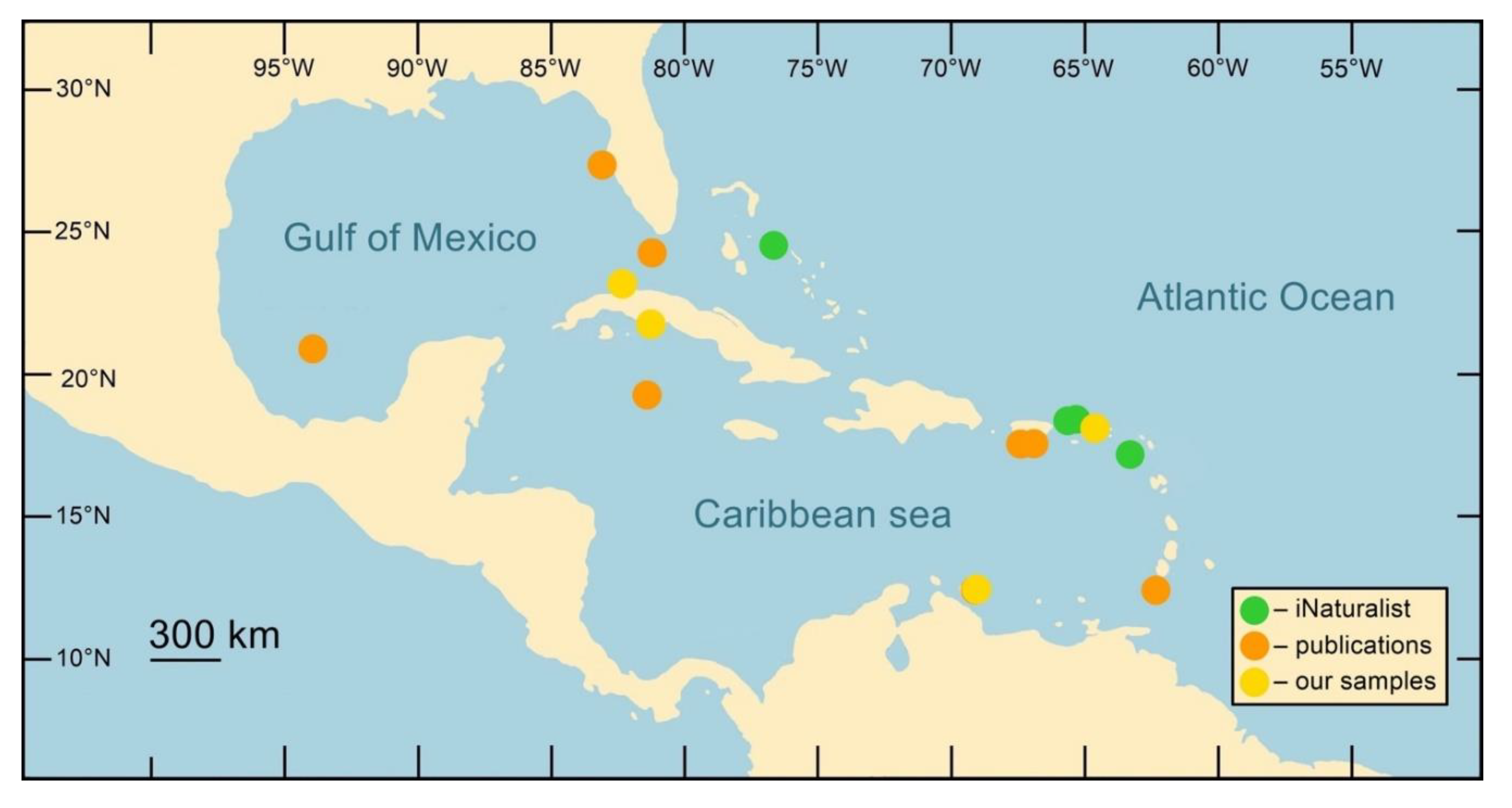
The current literature delineates the distribution of MFPS, spanning depths of 3–20 m along the coasts of Florida, Mexico, and the islands of Puerto Rico, Grand Cayman, Curaçao, St. Eustatius, and Grenada (Harvell et al., 2007; Weil and Hooten, 2008; Weil and Rogers, 2011; Burge et al., 2012; Ivanenko et al., 2017; Weil et al., 2017; Tracy et al., 2018). However, our analysis of underwater photographs from the iNaturalist web site (https://www.inaturalist.org) indicates a potentially broader spread of both Gorgonia and MFPS. Additionally, our data revealed the syndrome's presence in various regions of Cuba, and on the islands of Curaçao, Bonaire, and St. Eustatius (Figure 7, Table S7). There is a pressing need for more comprehensive data on the presence or absence of MFPS in other Caribbean regions, particularly given the current limited understanding of the syndrome’s impact on the health of the host Gorgonia octocorals.
Figure 7.
Localities of the multifocal purple spot syndrome (MFPS) records in the Caribbean Sea (see Table S7).
Figure 7.
Localities of the multifocal purple spot syndrome (MFPS) records in the Caribbean Sea (see Table S7).
5. Conclusions
Gorgonia affected by MFPS and their associated gall-forming copepods, with their relatively straightforward diagnostic features, have the potential to become model organisms for research on shallow-water communities in the Caribbean. Their distinctive characteristics, conducive to identification and observation, provide valuable insights into the ecological dynamics and health of these ecosystems. By designating these corals and copepods as model systems, researchers will be able to gain profound insights into the interplay between corals and pathogens, the impact of environmental changes on marine biodiversity, and the mechanisms of disease spread and response in coral ecosystems. This knowledge is crucial for developing effective conservation and management strategies for these vital marine habitats. Additionally, the ease of identifying these organisms and the symptoms of MFPS renders them suitable for broader involvement in scientific studies, including by citizen scientists and SCUBA diving enthusiasts (Korein et al., 2023), thus popularizing scientific research and promoting a more inclusive approach to marine conservation.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org, Tables S1–S7; Figure S1–S10. Table S1. List of primers used in molecular genetic analysis. Table S2. List of amplification temperature regimes for different primers. Table S3. GenBank accession numbers for ITS2 and msh1 sequences of coral specimens. For additional details, including geographic coordinates, please refer to Table 1. Table S4. GenBank accession numbers for 18S, COI and ITS2 sequences of copepod specimens. For additional details, including geographic coordinates, please refer to Table 1. Table S5. GenBank accession numbers ITS2 and msh1 sequences of species used for phylogenetic analyses. Table S6. GenBank accession numbers 18S sequences of species used for phylogenetic analyses. Table S7. List of MFPS geographic distribution with coordinates, depth data and references. Figure S1. Specimens of Gorgonia ventalina (Linnaeus 1758) labeled as follows: Statia15-99 – 1, 2; Statia15-134 – 3, 4; Statia15-135 – 5, 6; Statia15-141 – 7, 8. Figure S2. Specimens of Gorgonia ventalina (Linnaeus 1758) labeled as follows: Statia15-142 – 9, 10; Statia15-146 – 11, 12; Statia15-163 – 13, 14; Statia15-170 – 15, 16. Figure S3. Specimens of Gorgonia ventalina (Linnaeus 1758) labeled as follows: Statia15-174 – 17, 18; CUR17-39 – 19, 20; CUR17-81 – 21, 22; CUR17-88 – 23, 24. Figure S4. Specimens of Gorgonia ventalina (Linnaeus 1758) labeled as follows: CUR17-96 – 25, 26; Cuba19-1 – 27, 28; Cuba19-2 – 29, 30; Cuba19-3 – 31, 32. Figure 5. Specimens of Gorgonia ventalina (Linnaeus 1758) labeled as follows: Cuba19-5 – 33, 34; Cuba19-21 – 35, 36; Cuba19-22 – 37; Cuba19-25 – 38, no; Cuba19-32 – 39, no; Cuba19-33 – 40. Figure S6. Specimens of Gorgonia ventalina (Linnaeus 1758) labeled as follows: Cuba19-23 – 41, 42; Cuba19-27 – 43, 44; Cuba19-28 – 45, 46; Cuba19-30 – 47, 48. Figure S7. Specimens of Gorgonia ventalina (Linnaeus 1758) labeled as follows: Bonaire19-28 – 50, 51; Bonaire19-31 – 52, 53; Bonaire19-47 – 54, 55; Bonaire19-91 – 56, 57. Figure S8. The phylogenetic tree was constructed based on the concatenated alignment of COI (Cytochrome c oxidase subunit I) and ITS2 (Internal Transcribed Spacer 2) sequences of Lamippidae copepods, including Sphaerippe spp. Model of nucleotide evolution is HKY+G for COI_pos1, K81UF+I for ITS and COI_pos2, HKY+I for COI_pos3. Numbers in the nodes is posterior probabilities / bootstrap supports. Color bars on the right denote the species delimitation results. For additional details, including geographic coordinates, please refer to Table 1 and Table S4. Figure S9. In the Bayesian (BA) phylogenetic tree based on the alignment of 18S sequences of crustacean copepods, as indicated in Table S6, the nucleotide evolution model applied was GTR + G + I. The numbers associated with the nodes represent Bayesian (BA) posterior probabilities. For additional information, refer to Figure 6 and Supplementary Figure 10 (Figure 6 and Figure S10). Figure S10. Maximum Likelihood phylogenetic tree based on the 18S alignment of copepod crustaceans indicated in Table S6. The nucleotide evolution model applied is GTR + G + I. The numbers associated with the nodes represent bootstrap support values. For additional information, refer to Figure 6 and Supplementary Figure 9 (Figure 6 and Figure S9).
Author Contributions
Conceptualization and methodology, V.N.I., M.A.N., B.W.H.; software, O.A.K., M.A.N.; validation, O.A.K., M.A.N.; formal analysis, O.A.K., V.N.I., M.A.N.; investigation, O.A.K., V.N.I., M.A.N., M.A., B.W.H., J.D.R.; data curation, M.A.N., V.N.I., O.A.K.; writing—original draft preparation, O.A.K., V.N.I., M.A.N.; writing—review and editing, O.A.K., V.N.I., M.A.N., M.A., B.W.H., J.D.R.; visualization, O.A.K., V.N.I., M.A.N.; supervision, V.N.I.; project administration, V.N.I.; funding acquisition, V.N.I., M.A., B.W.H. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the Russian Science Foundation Grant No. 22-24-00365.
Institutional Review Board Statement
Not applicable.
Data Availability Statement
The data presented in this study are openly available in doi.org/хххх (accessed ххххх).
Conflicts of Interest
The authors declare no conflict of interest. The funder had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
References
- Aguilar, C.; Sánchez, J. A. Phylogenetic hypotheses of gorgoniid octocorals according to ITS2 and their predicted RNA secondary structures. Molecular Phylogenetics and Evolution 2007, 43, 774–786. [Google Scholar] [CrossRef] [PubMed]
- Altschul, S. F.; Gish, W.; Miller, W.; Myers, E. W.; Lipman, D. J. Basic local alignment search tool. Journal of Molecular Biology 1990, 215, 403–410. [Google Scholar] [CrossRef] [PubMed]
- Andras, J. P.; Kirk, N. L.; Harvell, C. D. Range–wide population genetic structure of Symbiodinium associated with the Caribbean Sea fan coral, Gorgonia ventalina. Molecular Ecology 2011, 20, 2525–42. [Google Scholar] [CrossRef]
- Andras, J. P.; Rypien, K. L.; Harvell, C. D. Range–wide population genetic structure of the Caribbean sea fan coral, Gorgonia ventalina. Molecular Ecology 2013, 22, 56–73. [Google Scholar] [CrossRef] [PubMed]
- Bayer, F. M. The shallow–water Octocorallia of the West Indian region. Studies on the Fauna of Curaçao and other Caribbean Islands.
- Becker, A. A. M. J.; Freeman, M. A.; Dennis, M. M. A combined diagnostic approach for the investigation of lesions resembling aspergillosis in Caribbean sea fans (Gorgonia spp.). Veterinary Pathology. [CrossRef]
- Burge, C. A.; Douglas, N.; Conti–Jerpe, I.; Weil, E.; Roberts, S.; Friedman, C. S.; Harvell, C. D. Friend or foe: the association of Labyrinthulomycetes with the Caribbean sea fan Gorgonia ventalina. Diseases of Aquatic Organisms 2012, 101, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Burge, C. A.; Mouchka, M. E.; Harvell, C. D.; Roberts, S. Immune response of the Caribbean seafan, Gorgonia ventalina exposed to an Aplanochytrium parasite as revealed by transcriptome sequencing. Frontiers in Physiology 2013, 4, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Calderón–Hernández, A.; Urbina–Villalobos, A.; Mora–Barboza, C. , Lesions in octocorals of the Costa Rican Caribbean During The 2015–2016 El Niño. Coral Reefs 2021, 40, 1167–1179. [Google Scholar] [CrossRef]
- Cheng, Y. R.; Lin, C. Y.; Yu, J. K. Embryonic and post–embryonic development in the parasitic copepod Ive ptychoderae (Copepoda: Iviidae): Insights into its phylogenetic position. PLoS One 2023, 18, e0281013. [Google Scholar] [CrossRef] [PubMed]
- Chollett, I.; Müller–Karger, F. E.; Heron, S. F.; Skirving, W.; Mumby, P. J. Seasonal and spatial heterogeneity of recent sea surface temperature trends in the Caribbean Sea and southeast Gulf of Mexico. Marine Pollution Bulletin 2012, 64, 956–965. [Google Scholar] [CrossRef]
- Conradi, M.; Bandera, E.; Mudrova, S. V.; Ivanenko, V. N. Five new coexisting species of copepod crustaceans of the genus Spaniomolgus (Poecilostomatoida: Rhynchomolgidae), symbionts of the stony coral Stylophora pistillata (Scleractinia). ZooKeys 2018, 791, 71–95. [Google Scholar] [CrossRef]
- Edgar, R. C. MUSCLE: multiple sequence alignment with high accuracy and high throughput. Nucleic Acids Research 2004, 32, 1792–1797. [Google Scholar] [CrossRef] [PubMed]
- Felsenstein, J. Phylogenies and the comparative method. The American Naturalist 1985, 125, 1–15. [Google Scholar] [CrossRef]
- Ferrari, F. D.; Ivanenko, V. N.; Dahms, H. –U. Body architecture and relationships among basal copepods. Journal of Crustacean Biology 2010, 30, 465–477. [Google Scholar] [CrossRef]
- Fontaneto, D.; Flot, J. F.; Tang, C. Q. Guidelines for DNA taxonomy, with a focus on the meiofauna. Marine Biodiversity 2015, 45, 1–19. [Google Scholar] [CrossRef]
- Fu, Y. –X. Statistical tests of neutrality against population growth, hitchhiking and background selection. Genetics 1997, 147, 915–925. [Google Scholar] [CrossRef] [PubMed]
- Geller, J.; Meyer, C.; Parker, M.; Hawk, H. Redesign of PCR primers for mitochondrial cytochrome c oxidase subunit I for marine invertebrates and application in all–taxa biotic surveys. Molecular Ecology Resources 2013, 13, 851–861. [Google Scholar] [CrossRef] [PubMed]
- Grygier, M. J. An endoparasitic Lamippid Copepod in Acanella from the North Atlantic. Crustaceana 1983, 45, 176–182. [Google Scholar] [CrossRef]
- Grygier, M. J. Two new lamippid copepods parasitic on gorgonians from Hawaii and the Bahamas. Proceedings of the Biological Society of Washington 1980, 93, 662–673. [Google Scholar]
- Guindon, S.; Dufayard, J. F.; Lefort, V.; Anisimova, M.; Hordijk, W.; Gascuel, O. New algorithms and methods to estimate maximum–likelihood phylogenies: assessing the performance of PhyML 3.0. 0. Systematic Biology 2010, 59, 307–321. [Google Scholar] [CrossRef]
- Harvell, C. D.; Markel, S.; Jordan–Dahlgren, E.; Raymundo, L. J.; Rosenberg, E.; Smith, G. W.; Willis, B. L.; Weil, E. Coral disease, environmental drivers and the balance between coral and microbial associates. Oceanography 2007, 20, 36–59. [Google Scholar] [CrossRef]
- Hoeksema, B. W.; Reimer, J. D.; Vonk, R. Editorial: biodiversity of Caribbean coral reefs (with a focus on the Dutch Caribbean). Marine Biodiversity 2017, 47, 1–10. [Google Scholar] [CrossRef]
- Humes, A. G. Lamippe concinna sp. n., a copepod parasitic in a West African pennatulid coelenterate. Parasitology.
- Huys, R.; Llewellyn–Hughes, J.; Olson, P. D.; Nagasawa, K. Small subunit rDNA and Bayesian inference reveal Pectenophilus ornatus (Copepoda incertae sedis) as highly transformed Mytilicolidae, and support assignment of Chondracanthidae and Xarifiidae to Lichomolgoidea (Cyclopoida). Biological Journal of the Linnean Society 2006, 87, 403–425. [Google Scholar] [CrossRef]
- iNaturalist. Available from https://www.inaturalist.org. Accessed 15th 22. 20 December.
- Ivanenko, V. N.; Hoeksema, B. W.; Mudrova, S. V.; Nikitin, M. A.; Martínez, A.; Rimskaya–Korsakova, N. N.; Berumen, M. L.; Fontaneto, D. Lack of host specificity of copepod crustaceans associated with mushroom corals in the Red Sea. Molecular Phylogenetics and Evolution 2018, 127, 770–780. [Google Scholar] [CrossRef] [PubMed]
- Ivanenko, V. N.; Nikitin, M. A.; Hoeksema, B. W. Multiple purple spots in the Caribbean Sea fan Gorgonia ventalina caused by parasitic copepods at St. Eustatius, Dutch Caribbean. Marine Biodiversity 2017, 47, 79–80. [Google Scholar] [CrossRef]
- Ivanenko, V. N.; Ferrari, F. D. Redescription of adults and description of copepodid development of Dermatomyzon nigripes (Brady, Robertson, 1876) and of Asterocheres lilljeborgi Boeck, 1859 (Copepoda: Siphonostomatoida: Asterocheridae). Proceedings of the Biological Society of Washington 2003, 116, 661–691. [Google Scholar]
- Ivanenko, V. N.; Ferrari, F. D.; Smurov, A. V. Nauplii and copepodids of Scottomyzon gibberum (Copepoda: Siphonostomatoida: Scottomyzotidae, a new family), a symbiont of Asterias rubens (Asteroidea). Proceedings of the Biological Society of Washington 2001, 114, 237–61. [Google Scholar]
- Ivanenko, V. N.; Defaye, D. A new and primitive genus and species of deep–sea Tegastidae (Crustacea, Copepoda, Harpacticoida) from the Mid–Atlantic Ridge, 37°N (Azores Triple Junction, Lucky Strike). Cahiers de Biologie Marine 2004, 45, 255–268. [Google Scholar]
- Jossart, Q.; De Ridder, C.; Lessios, H. A.; Bauwens, M.; Sébastien, M.; Thierry, R.; Rémi, A. W.; Bruno, D. Highly contrasted population genetic structures in a host–parasite pair in the Caribbean Sea. Ecology and Evolution 2017, 7, 9267–9280. [Google Scholar] [CrossRef]
- Katoh, K.; Rozewicki, J.; Yamada, K. D. MAFFT online service: multiple sequence alignment, interactive sequence choice and visualization. Briefings in Bioinformatics 2017, bbx108, 1–7. [Google Scholar] [CrossRef]
- Kearse, M.; Moir, R.; Wilson, A.; Stones–Havas, S.; Cheung, M.; Sturrock, S.; Drummond, A. Geneious Basic: an integrated and extendable desktop software platform for the organization and analysis of sequence data. Bioinformatics 2012, 28, 1647–1649. [Google Scholar] [CrossRef]
- Kim, K.; Rypien, K. Aspergillosis of Caribbean Sea Fan Corals, Gorgonia spp. In Diseases of Coral; 2015, 236–241.
- Kimes, N. E.; Van Nostrand, J. D.; Weil, E.; Zhou, J.; Morris, P. J. Microbial functional structure of Montastraea faveolata, an important Caribbean reef–building coral, differs between healthy and Caribbean yellow–band diseased colonies. Environmental Microbiology 2010, 12, 541–561. [Google Scholar] [CrossRef] [PubMed]
- Knowlton, N.; Rohwer, F. Multispecies microbial mutualisms on coral reefs: the host as a habitat. The American Naturalist 2003, 162, 51–62. [Google Scholar] [CrossRef] [PubMed]
- Korein, E.; Vega–Rodriguez, M.; Metz Estrella, T. Developing recommendations for coral disease management in Puerto Rico using key informant interviews and participatory mapping. Ocean and Coastal Management 2023, 236, 106488. [Google Scholar] [CrossRef]
- Korzhavina, O.A.; Grishina, D.Y.; Chen, X.; Fontaneto, D.; Ivanenko, V.N. Diving into diversity: copepod crustaceans in octocoral association. Diversity 2023, 15, 1140. [Google Scholar] [CrossRef]
- Korzhavina, O.A.; Hoeksema, B.W.; Ivanenko, V.N. A review of Caribbean Copepoda associated with reef–dwelling cnidarians, echinoderms, and sponges. Contributions to Zoology 2019, 88, 297–349. [Google Scholar] [CrossRef]
- Korzhavina, O.A.; Reimer, J.D.; Ehrlich, H.; Ivanenko, V.N. Global diversity and distribution of Lamippidae copepods symbiotic on Octocorallia. Symbiosis 2021, 83, 265–277. [Google Scholar] [CrossRef]
- Kuraku, S.; Zmasek, C. M.; Nishimura, O.; Katoh, K. aLeaves facilitates on–demand exploration of metazoan gene family trees on MAFFT sequence alignment server with enhanced interactivity. Nucleic Acids Research 2013, 41, W22–W28. [Google Scholar] [CrossRef] [PubMed]
- Lanfear, R.; Calcott, B.; Ho, S. Y.; Guindon, S. PartitionFinder: combined selection of partitioning schemes and substitution models for phylogenetic analyses. Molecular Biology and Evolution 2012, 29, 1695–1701. [Google Scholar] [CrossRef] [PubMed]
- Lanfear, R.; Frandsen, P. B.; Wright, A. M.; Senfeld, T.; Calcott, B. PartitionFinder 2: new methods for selecting partitioned models of evolution for molecular and morphological phylogenetic analyses. Molecular Biology and Evolution 2016, 34, 772–773. [Google Scholar] [CrossRef]
- Laubier, L. Lamippe (Lamippe) bouligandi sp. nov., copépode parasite d'Octocoralliaire de la Mer du Labrador. Crustaceana 1972, 22, 285–293. [Google Scholar] [CrossRef]
- Leigh, J. W.; Bryant, D. POPART: full–feature software for haplotype network construction. Methods in Ecology and Evolution 2015, 6, 1110–1116. [Google Scholar] [CrossRef]
- Lesser, M. P.; Bythell, J. C.; Gates, R. D.; Johnstone, R. W.; Hoegh–Guldberg, O. Are infectious diseases really killing corals? Alternative interpretations of the experimental and ecological data. Journal of Experimental Marine Biology and Ecology 2007, 34, 36–44. [Google Scholar] [CrossRef]
- Martin, S. W.; Meek, A. H.; Willerberg, P. Veterinary epidemiology, principles and methods. Iowa State University Press, Ames 1987, p. 343.
- McFadden, C. S.; France, S. C.; Sánchez, J. A.; Alderslade, P. A molecular phylogenetic analysis of the Octocorallia (Cnidaria: Anthozoa) based on mitochondrial protein–coding sequences. Molecular Phylogenetics and Evolution 2006, 41, 513–527. [Google Scholar] [CrossRef]
- McFadden, C. S.; Sánchez, J. A.; France, S. C. Molecular phylogenetic insights into the evolution of Octocorallia: a review. Integrative and Comparative Biology 2010, 50, 389–410. [Google Scholar] [CrossRef]
- Medlin, L.; Elwood, H. J.; Stickel, S.; Sogin, M. L. The characterization of enzymatically amplified eukaryotic 16S–like rRNA–coding regions. Gene 1988, 71, 491–499. [Google Scholar] [CrossRef]
- Mikhailov, K.V.; Ivanenko, V.N. Lack of reproducibility of molecular phylogenetic analysis of Cyclopoida. Molecular Phylogenetics and Evolution 2019, 139, 106574. [Google Scholar] [CrossRef] [PubMed]
- Michels, J.; Appel, E.; Gorb, S.N. Functional diversity of resilin in Arthropoda. Beilstein J Nanotechnol. 2016, 1, 1241–1259. [Google Scholar] [CrossRef] [PubMed]
- Mikhailov, K.V.; Ivanenko, V.N. Low support values and lack of reproducibility of molecular phylogenetic analysis of Copepoda orders. Arthropoda Selecta 2021, 30, 39–42. [Google Scholar] [CrossRef]
- Montano, S.; Maggioni, D.; Liguori, G. , Morpho–molecular traits of Indo–Pacific and Caribbean Halofolliculina ciliate infections. Coral Reefs 2020, 39, 375–386. [Google Scholar] [CrossRef]
- Petes, L. E.; Harvell, C. D.; Peters, E. C.; Webb, M. A. H.; Mullen, K. M. Pathogens compromise reproduction and induce melanization in Caribbean Sea fans. Marine Ecology Progress Series 2003, 264, 167–171. [Google Scholar] [CrossRef]
- Porco, D.; Rougerie, R.; Deharveng, L.; Hebert, P. Coupling non–destructive DNA extraction and voucher retrieval for small soft–bodied Arthropods in a high–throughput context: the example of Collembola. Molecular Ecology Resources 2010, 10, 942–945. [Google Scholar] [CrossRef] [PubMed]
- Rambaut, A.; Drummond, A. J.; Xie, D.; Baele, G.; Suchard, M. A. Posterior summarization in Bayesian phylogenetics using Tracer 1.7. 7. Systematic Biology 2018, 67, 901–904. [Google Scholar] [CrossRef] [PubMed]
- Ritchie, K. B. Regulation of microbial populations by mucus–associated bacteria. Marine Ecology Progress Series 2006, 322, 1–14. [Google Scholar] [CrossRef]
- Rocha, L. A.; Rocha, C. R.; Robertson, D. R.; Bowen, B. W. Comparative phylogeography of Atlantic reef fishes indicates both origin and accumulation of diversity in the Caribbean. BMC Evolutionary Biology 2008, 8, 157. [Google Scholar] [CrossRef]
- Rogers, C. S. Words matter: recommendations for clarifying coral disease nomenclature and terminology. Diseases of Aquatic Organisms 2010, 91, 167–175. [Google Scholar] [CrossRef] [PubMed]
- Ronquist, F.; Teslenko, M.; Van Der Mark, P.; Ayres, D. L.; Darling, A.; Höhna, S.; Larget, B.; Liu, L.; Suchard, M. A.; Huelsenbeck, J. P. MrBayes 3.2: efficient Bayesian phylogenetic inference and model choice across a large model space. Systematic Biology 2012, 61, 539–542. [Google Scholar] [CrossRef] [PubMed]
- Rosenberg, E.; Ben–Haim, Y. Microbial diseases of corals and global warming. Environmental Microbiology 2002, 4, 318–326. [Google Scholar] [CrossRef] [PubMed]
- Ruiz–Moreno, D.; Willis, B. L.; Page, A. C.; Weil, E.; Croquer, A.; Vargas–Angel, B.; Jordan–Garza, A. G.; Jordán–Dahlgren, E.; Raymundo, L.; Harvell, C. D. Global coral disease prevalence associated with sea temperature anomalies and local factors. Diseases of Aquatic Organisms 2012, 100, 249–261. [Google Scholar] [CrossRef] [PubMed]
- Sanchez, J. A.; Aguilar, C.; Dorado, D.; Manrique, N. Phenotypic plasticity and morphological integration in a marine modular invertebrate. BMC Evolutionary Biology 2007, 7, 122. [Google Scholar] [CrossRef]
- Sánchez, J. A.; McFadden, C. S.; France, S. C.; Lasker, H. R. Molecular phylogenetic analyses of shallow–water Caribbean octocorals. Marine Biology 2003, 142, 975–987. [Google Scholar] [CrossRef]
- Sanchez, J. A.; Wirshing, H.H. A field key to the identification of tropical western Atlantic zooxanthellate octocorals (Octocorallia: Cnidaria). Caribbean Journal of Science 2005, 41, 508–522. [Google Scholar]
- Shelyakin, P. V.; Garushyants, S. K.; Nikitin, M. A.; Mudrova, S. V.; Berumen, M.; Speksnijder, A. G. C. L.; Hoeksema, B. W.; Fontaneto, D.; Gelfand, M. S.; Ivanenko, V. N. Microbiomes of gall–inducing copepod crustaceans from the corals Stylophora pistillata (Scleractinia) and Gorgonia ventalina (Alcyonacea). Scientific Reports 2018, 8, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Smith, G. W.; Weil, E. Aspergillosis of gorgonians. In Coral Health and Disease; Rosenberg, E., Loya, Y., Eds.; Springer: New York, 2004. [Google Scholar]
- Tajima, F. Statistical Method for Testing the Neutral Mutation Hypothesis by DNA Polymorphism. Genetics Society of America 1989, 123, 585–595. [Google Scholar] [CrossRef] [PubMed]
- Taylor, M. S.; Hellberg, M. E. Comparative phylogeography in a genus of coral reef fishes: biogeographic and genetic concordance in the Caribbean. Molecular Ecology 2006, 15, 695–707. [Google Scholar] [CrossRef] [PubMed]
- Tracy, A. M.; Weil, E.; Harvell, C. D. Octocoral co–infection as a balance between host immunity and host environment. Oecologia 2018, 186, 743. [Google Scholar] [CrossRef] [PubMed]
- Trifinopoulos, J.; Nguyen, L. T.; von Haeseler, A.; Minh, B. Q. Nucleic Acids Research 2016, 44 (W1), W232–W235.
- van de Water, J. A. J. M.; Allemand, D.; Ferrier–Pagès, C. Host–microbe interactions in octocoral holobionts – recent advances and perspectives. Microbiome 2018, 6, 64. [Google Scholar] [CrossRef]
- Weil, E.; Hooten, A. J. Underwater cards for assessing coral health on Caribbean reefs. GEF–CRTR Program, Center for Marine Sciences: University of Queensland, Brisbane, 2008.
- Weil, E. Coral reef diseases in the wider Caribbean. In Coral Health and Disease; Rosenberg, E., Loya, Y., Eds.; Springer, New York, 2004, 35–68.
- Weil, E.; Rogers, C. S. Coral reef diseases in the Atlantic–Caribbean. In Coral Reefs: An Ecosystem in Transition; Dubinsky, Z., Stambler, N., Eds.; Springer, 2011, 465–491.
- Weil, E.; Rogers, C. S.; Croquer, A. Octocoral diseases in a changing ocean. In Marine Animal Forests: The Ecology of Benthic Biodiversity Hotspots; Rossi, S., Bramanti, L., Gori, A., Orejas Saco del Valle, C., Eds.; Springer International Publishing, 2017, 1–55.
- Williams, E. H. Jr.; Bunkley–Williams, L. Life Cycle and Life History Strategies of Parasitic Crustacea. Parasitic Crustacea 2019, 3, 179–266. [Google Scholar]
- Williams, J. D.; Anchaluisa, B.; Boyko, C. B.; McDaniel, N. Description of a new endoparasitic copepod genus and species (Lamippidae) that induces gall formation in leaves of the sea pen Ptilosarcus gurneyi (Octocorallia) from British Columbia. Marine Biodiversity 2018, 48, 1–11. [Google Scholar] [CrossRef]
- Work, T. M.; Aeby, G. S. Systematically describing gross lesions in corals. Diseases of Aquatic Organisms 2006, 70, 155–160. [Google Scholar] [CrossRef]
Figure 1.
(a) Sampling localities are distributed over three ecoregions in the Caribbean Sea (details in Table 1;): (b) Cuba in the Greater Antilles; (c) Bonaire and Curaçao in the Southern Caribbean, and (d) St. Eustatius in the Eastern Caribbean.
Figure 1.
(a) Sampling localities are distributed over three ecoregions in the Caribbean Sea (details in Table 1;): (b) Cuba in the Greater Antilles; (c) Bonaire and Curaçao in the Southern Caribbean, and (d) St. Eustatius in the Eastern Caribbean.
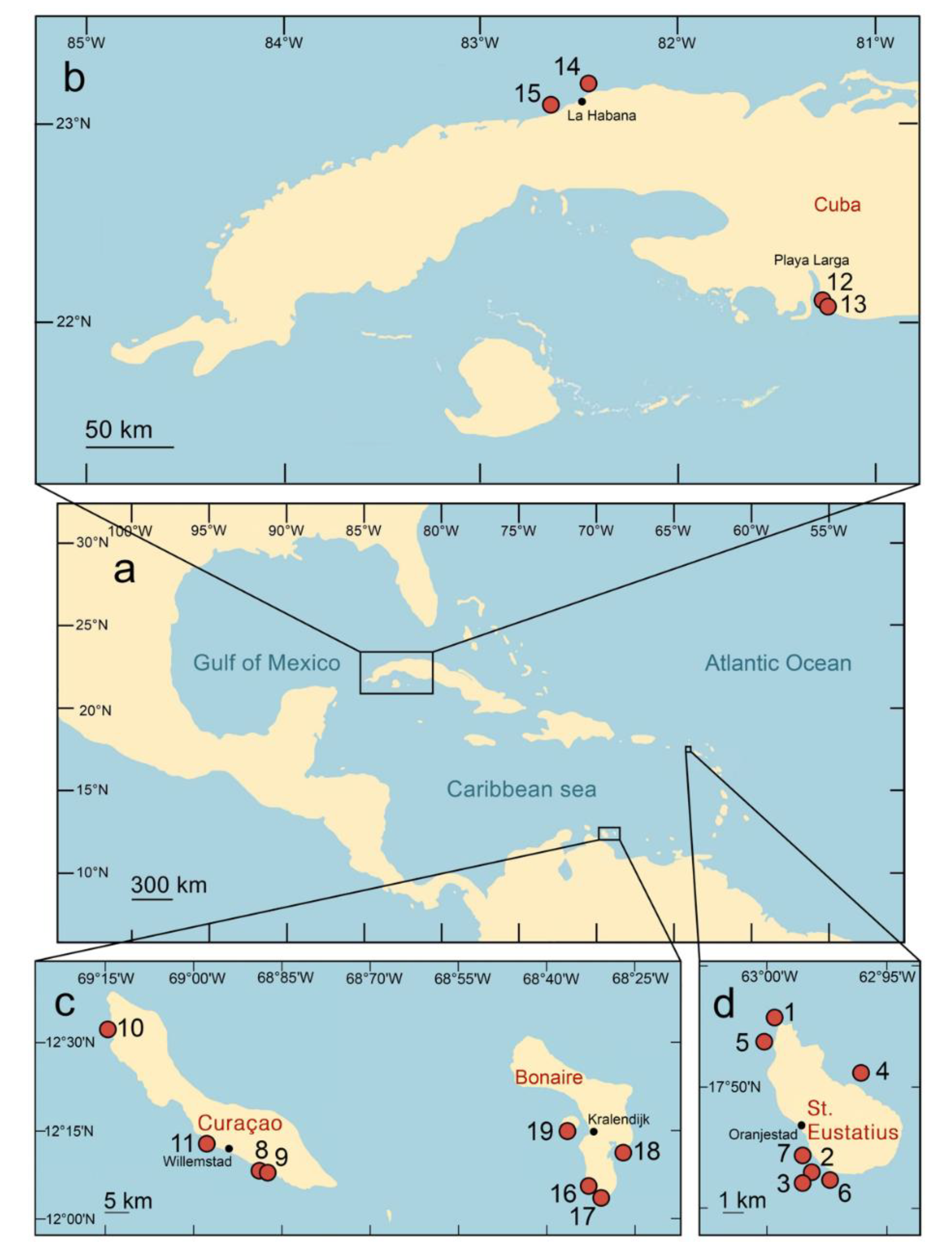
Figure 2.
a, b – the sea fan Gorgonia ventalina with purple galls containing Sphaerippe spp. copepods (Lamippidae); c – spherical female of Sphaerippe sp.; d – elongated male of Sphaerippe sp.; e, f– embryonic nauplii in a capsule formed by the sea fan tissues; L1, L2 – legs 1-2; a1 – antennule, a2 antenna, cr – caudal ramus, o – oral opening, sc – sclerite of the sea fan, nv – nauplius, ventral vew, nd – nauplius dorsal view. Scale bars; a – 25 cm, b – 5 cm, c–f – 100 µm, 50 µm, 50 µm, and 20 µm, respectively.
Figure 2.
a, b – the sea fan Gorgonia ventalina with purple galls containing Sphaerippe spp. copepods (Lamippidae); c – spherical female of Sphaerippe sp.; d – elongated male of Sphaerippe sp.; e, f– embryonic nauplii in a capsule formed by the sea fan tissues; L1, L2 – legs 1-2; a1 – antennule, a2 antenna, cr – caudal ramus, o – oral opening, sc – sclerite of the sea fan, nv – nauplius, ventral vew, nd – nauplius dorsal view. Scale bars; a – 25 cm, b – 5 cm, c–f – 100 µm, 50 µm, 50 µm, and 20 µm, respectively.
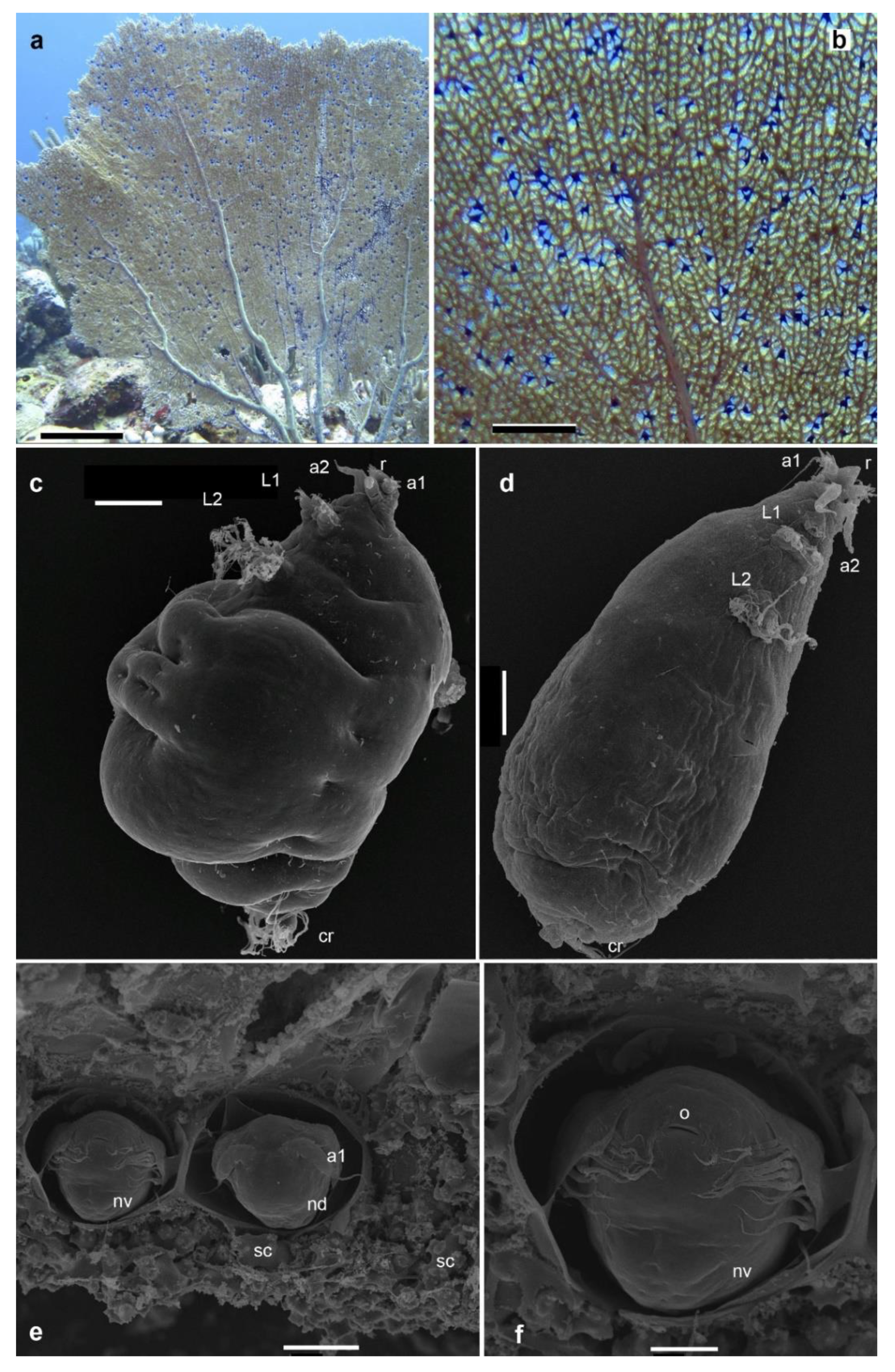
Figure 3.
Bayesian inference phylogenetic trees based on COI (a) and ITS2 (b) alignments of lamippid copepods including Sphaerippe spp. The nucleotide evolution model used was GTR + G + I, and the numbers at the nodes represent Bayesian posterior probabilities. Species delimitation results indicated by color bars on the right. For additional details, including geographic coordinates, please refer to Table 1.
Figure 3.
Bayesian inference phylogenetic trees based on COI (a) and ITS2 (b) alignments of lamippid copepods including Sphaerippe spp. The nucleotide evolution model used was GTR + G + I, and the numbers at the nodes represent Bayesian posterior probabilities. Species delimitation results indicated by color bars on the right. For additional details, including geographic coordinates, please refer to Table 1.
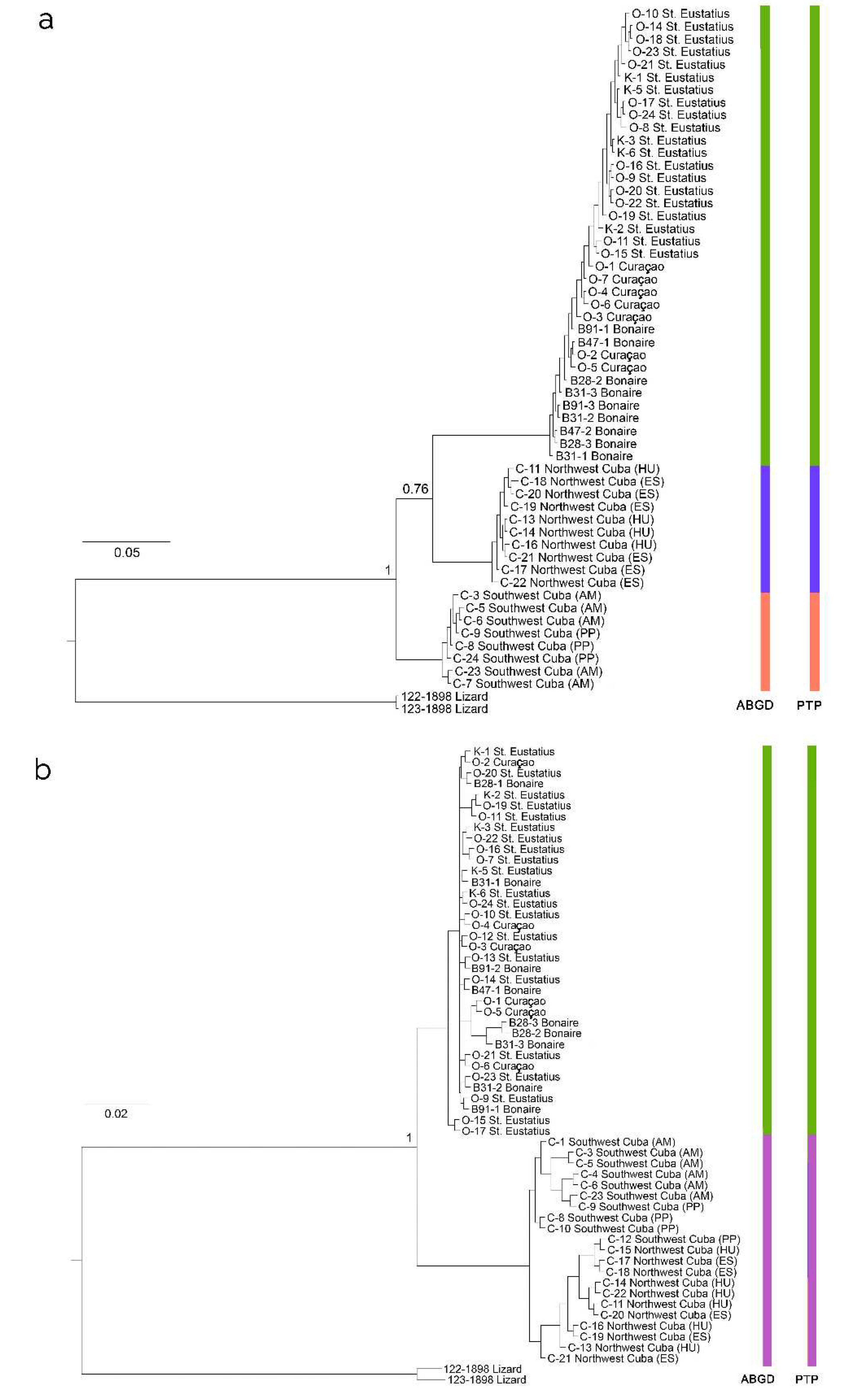
Figure 4.
Median-Joining Networks of COI and ITS2 haplotypes in Sphaerippe spp. The representation of haplotype frequency is reflected in the size of the circles, while the notches on connecting lines indicate the number of nucleotide substitutions between haplotypes. Colours distinguish the geographic origins of specimens: Yellow signifies individuals collected from St. Eustatius, light and dark green represent Bonaire and Curaçao, orange and pink denote Southwest Cuba, and blue and purple are Northwest Cuba.
Figure 4.
Median-Joining Networks of COI and ITS2 haplotypes in Sphaerippe spp. The representation of haplotype frequency is reflected in the size of the circles, while the notches on connecting lines indicate the number of nucleotide substitutions between haplotypes. Colours distinguish the geographic origins of specimens: Yellow signifies individuals collected from St. Eustatius, light and dark green represent Bonaire and Curaçao, orange and pink denote Southwest Cuba, and blue and purple are Northwest Cuba.
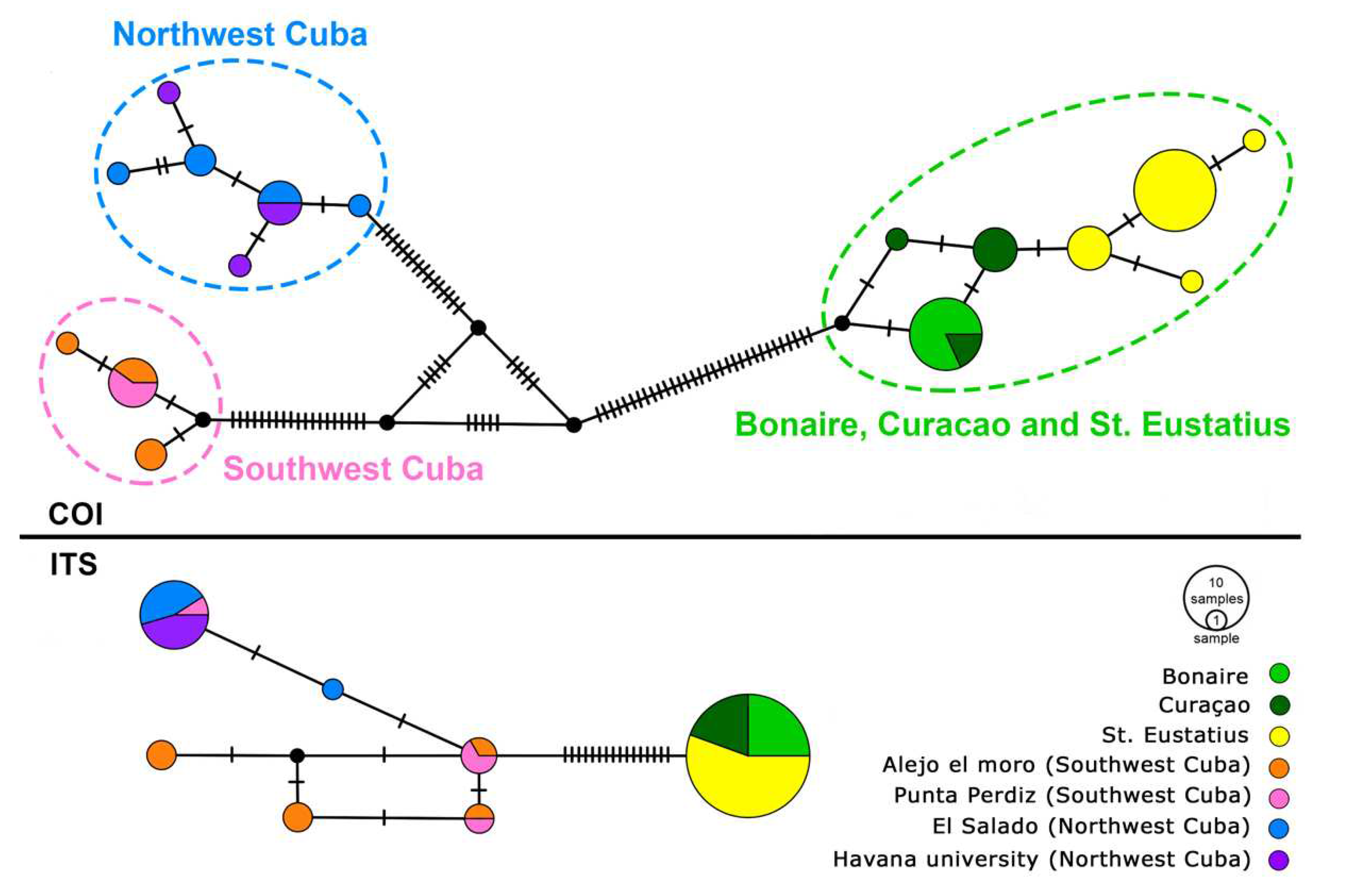

Table 1.
Localities (Figure 1), studied specimens, and sequence availability of gall-causing copepod of Sphaerippe spp. from Gorgonia ventalina in three ecoregions: the Greater Antilles (Cuba), the Eastern Caribbean (St. Eustatius) and the Southern Caribbean (Bonaire and Curaçao).
Table 1.
Localities (Figure 1), studied specimens, and sequence availability of gall-causing copepod of Sphaerippe spp. from Gorgonia ventalina in three ecoregions: the Greater Antilles (Cuba), the Eastern Caribbean (St. Eustatius) and the Southern Caribbean (Bonaire and Curaçao).
| Locality name | Coordinates | Date of sampling | Collector(s) | Name of specimens | Depth, m | Coral | Copepods |
|---|---|---|---|---|---|---|---|
| Gibraltar, St. Eustatius, (Fig. 1, point 1) | 17°31'36.5"N 62°59'57.5"W | 12.06.2015 | V.N.I. | Statia15-99 | 5-20 | + | + |
| Anchor Point North, St. Eustatius (Fig. 1, point 2) | 17°27'50.0"N 62°59'15.7"W | 17.06.2015 | V.N.I. | Statia15-134 | 15-20 | + | |
| Statia15-135 | 15-20 | + | |||||
| Anchor Reef, St. Eustatius (Fig. 1, point 3) | 17°27'44.8"N 62°59'07.7"W | 18.06.2015 | V.N.I. | Statia15-141 | 15.6 | + | + |
| Statia15-142 | 15.6 | + | + | ||||
| English Quarter, St. Eustatius (Fig. 1, point 4) | 17°30'18.2"N 62°57'46.3"W | 19.06.2015 | V.N.I. | Statia15-146 | 17.3 | + | |
| Twin Sisters, St. Eustatius (Fig. 1, point 5) | 17°30'59.6"N 63°00'10.8"W | 22.06.2015 | V.N.I. | Statia15-163 | 13.8 | + | |
| Blund Shoal, St. Eustatius (Fig. 1, point 6) | 17°27'52.6"N 62°58'38.7"W | 26.06.2015 | V.N.I. | Statia15-170 | 5.9 | + | |
| Gallows Bay, St. Eustatius (Fig. 1, point 7) | 17°28'30.3"N 62°59'10.3"W | 27.06.2015 | V.N.I. | Statia15-173 | 13.8 | + | |
| Statia15-174 | 2-3 | + | |||||
| Director's Bay, Curaçao, (Fig. 1, point 8) | 12°03'59"N 68°51'38"W | 13.06.2017 | V.N.I. | Cur17-39 | 4.1 | + | + |
| Tugboat 2, Curaçao (Fig. 1, point 9) | 12°04'05"N, 68°51'44"W | 19.06.2017 | V.N.I. | Cur17-81 | 5.2-5.5 | + | |
| Playa Lagun, Curaçao (Fig. 1, point 10) | 12°19'02"N, 69°09'09"W | 20.06.2017 | V.N.I. | Cur17-88 | 4.9 | + | |
| Buoy 1, Curaçao (Fig. 1, point 11) | 12°07'23"N, 68°58'14"W | 21.06.2017 | V.N.I. | Cur17-96 | 8.2 | + | |
| Alejo el Moro, Cuba (Fig. 1, point 12) | 22°06'54.99"N 81°06'58.96"W | 04.02.2019 | V.N.I., O.A.K. | Cuba19-1 | 7.0 | + | + |
| Cuba19-2 | 8.5 | + | + | ||||
| Cuba19-3 | 4.5 | + | + | ||||
| Punta Perdiz, Cuba (Fig. 1, point 13) | 22°06'29.65"N 81°06'49.42"W | 04.02.2019 | V.N.I., O.A.K. | Cuba19-5 | 4.8-5.0 | + | + |
| Coast near Havana University, Cuba (Fig. 1, point 14) | 23°07'38.75"N82°25'21.68"W | 07.02.2019 | V.N.I., O.A.K. | Cuba19-21 | 11.6 | + | |
| Cuba19-22 | 8.5 | + | |||||
| Cuba19-23 | 11 | + | + | ||||
| Cuba19-25 | 8.1-8.2 | + | + | ||||
| El Salado, Cuba (Fig. 1, point 15) | 23°02'20.33"N 82°36'18.55"W | 08.02.2019 | V.N.I., O.A.K. | Cuba19-27 | 13.8 | + | + |
| Cuba19-28 | 10.0 | + | + | ||||
| Cuba19-30 | 12.6 | + | |||||
| Cuba19-32 | 8.3 | + | + | ||||
| Red Beryl, Bonaire (Fig. 1, point 16) | 12° 2' 49.14"N 68° 16' 4.38"W | 28.10.2019 | V.N.I. | Bonaire19-28 | 5 | + | + |
| Red Slave, Bonaire (Fig. 1, point 17) | 12° 1' 36.3"N 68° 15' 4.74"W | 29.10.2019 | V.N.I. | Bonaire19-31 | 14 | + | + |
| Cai (outside of lagoon), Bonaire (Fig. 1, point 18) | 12° 6' 10.98"N 68° 13' 19.98"W | 31.10.2019 | V.N.I. | Bonaire19-47 | 11 | + | + |
| Klein Bonaire: South Bay, Bonaire (Fig. 1, point 19) | 12° 9' 0.06"N 68° 19' 14.04"W | 08.11.2019 | V.N.I. | Bonaire19-91 | 3 | + | + |
Table 2.
Results of test on geographical isolation.
| Species of sample | Localities | Gene | Number of sequences | Nucleotide diversity | Tajima’s D | Fu’s F |
|---|---|---|---|---|---|---|
| Sphaerippe spp. | Bonaire, Curaçao, St. Eustatius | COI | 36 | 0.00264 | 0.30709 | -0.850 |
| Sphaerippe spp. | Northwest Cuba | COI | 10 | 0.00250 | -1.49280 | -2.563 |
| Sphaerippe spp. | Southwest Cuba | COI | 8 | 0.00168 | -0.17740 | 0.390 |
| Sphaerippe spp. | Bonaire, Curaçao, St. Eustatius | ITS2 | 36 | All sequences are identical | ||
| Sphaerippe spp. | Cuba | ITS2 | 21 | 0.00635 | 1.03432 | -0.378 |
| Gorgonia ventalina (Linnaeus 1758) | Caribbean region | ITS2 | All sequences are identical | |||
| Gorgonia ventalina (Linnaeus 1758) | Caribbean region | msh1 | 25 | 0.00538 | -1.92207 | 2.449 |
* Bold attribute in the column of Tajima’s D statistic means that this value is significant.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



