Submitted:
04 December 2023
Posted:
05 December 2023
You are already at the latest version
Abstract
The new genus and the new edaphic species, Prokletijella montana gen. n., sp. n., from the Prokletije mountains in Montenegro is described and illustrated. This is the first genus in Leptodirini with an asymmetric median lobe of the aedeagus. The phylogenetic relationship with the genus Adelopsella Jeannel, 1908 is discussed. The genus Adelopsella is redescribed to be accurately compared with Prokletijella. The occurrence of asymmetric genitalia in Leptodirini is discussed in relation with the occurrence of such asymmetry in males and females in other tribes of Cholevinae. The geographic distribution of the two genera is discussed in connexion with their phylogenetic relationship and mapped. The synonymy of the two subspecies A. bosnica bosnica and A. bosnica jezerensis Jeannel, 1911 is confirmed.
Keywords:
Asymmetric genitalia
; Phylogeny
; Prokletije Mts.
; Taxonomy
; Zoogeography
1. Introduction
The subterranean fauna of the Balkan peninsula is one of the most diverse in the world for most of zoological groups which have colonized the subterranean habitat and especially for Coleoptera [1]. The investigation of the subterranean diversity of Coleoptera is still in progress and this paper introduces a new genus of Leptodirini, Prokletijella, living in Prokletije mountains (Montenegro), which exhibits many unusual morphological characters in the tribe. The most surprising character is the asymmetry of the aedeagus. Such an asymmetry is observed in other tribes of Cholevinae, extensively in Ptomaphagini, sparsely in Cholevini, but currently it has never been observed in Leptodirini.
Prokletijella shares many morphological characters with Adelopsella Jeannel, 1908, a genus widely distributed in Bosnia and which is recorded in this paper for the first time from Montenegro. The phylogenetic relationship of these two genera are investigated, based on detailed morphological description and redescription.
2. Materials and Methods
2.1. Samples
All specimens of the new species (eight specimens) were collected by sifting of leaf-litter in the elevation of 1000-1150 m in the context of the authors’ ongoing project titled “Investigation of the biodiversity of litter and soil dwelling beetles from Montenegro”, with research permits granted by the “Agencija za zaštitu životne sredine” (03-UPI-1220/4 and 03-UPI-1534/6) of the Republic of Montenegro.
In descriptions, all paired structures are treated singular.
2.2. Preparation for Microscopic Observations
After dissection, the aedeagus were dehydrated in 95% ethanol and then mounted in Euparal on a glass coverslip. The female genitalia were cleaned in 10% potassium hydroxyde heated for 10 min, colored with Azoblack™ , rinsed in deionized water, then mounted in dimethyl-hydantoin-formaldehyde (DMHF) on a glass coverslip. The glass slides were glued in turn on a piece of cardboard and stitched under the specimens in collections.
2.3. Picturing
Images were taken with a Keyence VHX5000 microscope equipped with a VH-Z250T lens. When necessary, multifocal layers were combined with the Helicon focus software (http://www.heliconsoft.com/heliconsoft-products/helicon-focus/).
2.4. Labels
Label data of the holotype and paratypes are quoted verbatim, with slashes (/) separating lines of text. All type specimens were provided with the following red printed label: HOLOTYPE or PARATYPE, Prokletijella montana sp. n., M. Perreau & P. Hlaváč det., 2023.
2.5. Abbreviations
The following abbreviations are used for institutions and collections:
| MNHN | Muséum national d’histoire naturelle de Paris, France. |
| NMPC | Entomologické oddélení národní ho muzea, Prague, Czech Republic. |
| PCPH | Peter Hlaváč, Czech republic. |
| PCDC | David Ceplik, Košice, Czech republic. |
| PCEQ | Eric Quéinnec, Paris, France. |
| PCMP | Michel Perreau, Paris, France. |
| PCAS | Adam Šima, Brandýs nad Labem-Stará Boleslav, Czech Republic. |
3. Phylogeny
3.1. The “Adelopsella” Genus Group
We will notice in the section devoted to the taxonomy, that the new genus shares with the genus Adelopsella characters which are not usually encountered in most of Leptodirini. Presently included in Pholeuina, the phylogenetic relationship of Adelopsella with other genera of the subtribe remains poorly understood. The monophyly of Pholeuina itself is questioned in recent molecular investigations ???. Adelopsella is generally considered as an archaic isolated genus because of the presence of a dilated first male mesotarsomere and pigmented well-defined eyes [2,3,4]. The discovery of a new genus Prokletijella which is likely phylogenetically related to Aldelopsella reveals in these two genera many other unique characters among Leptodirini. Here we list the main ones:
- First male mesotarsomere dilated. Likely plesiomorphic, in Leptodirini, it occurs only in the genus Platycholeus, but it is rather common in other tribes or subtribes of Cholevinae: Ptomaphagini, Anemadini, Catopina [5].
- Protibia sexually dimorphic. Likely apomorphic, never recorded in other genera of Leptodirini.
- Presence of two lateral apophysis to the anterior margin of the female abdominal ventrite VIII, in addition to the usual central spiculum ventrale (Figure 3f and Figure 5d). Likely apomorphic, never recorded in all Cholevinae and also in all Leiodidae, but poorly investigated. In other subfamilies of Leiodidae than Cholevinae, the morphology of female ventrite VIII was rarely studied.
- Elongated female appendicular pieces, especially gonocoxites. Likely apomorphic, currently never recorded in Leptodirini but not studied in most genera.
- Spermatheca membranous. Likely plesiomorphic, exceptional in Leptodirini where the spermatheca is generally sclerified at both ends. Observed also in Remyella [7].
Among those characters two are likely plesiomorphic and four are likely apomorphic. This suggests that the group of these two genera – which is here called “Adelopsella” genus group or phyletic lineage after Jeannel’s terminology – is monophyletic, what should be later checked by the molecular analysis.
Some other shared characters which are not exclusive of these two genera are listed in the diagnose of this group which follows.
3.2. Diagnose of the “Adelopsella” Genus Group
Body convex, of bathyscoid shape. Head with occipital carina. Eyes present, with about 6-10 ommatidia of different size. Pronotum transverse, slightly wider than elytra, encompassing the base of elytra, its lateral outlines widely rounded (Figure 1a,c). Elytron lacking parasutural stria. Posterior wings absent. Male protarsus pentamerous, female protarsus tetramerous. Male protarsus and first male mesotarsomere dilated (Figure 2b,d and Figure 4b,d). Female protarsus and mesotarsus lacking dilatation. All tarsi with two interongular setae. Protibia sexually dimorphic, in males dilated from base to apex (Figure 2b, Figure 4b), roughly parallel-sided in female (Figure 2c and Figure 4c). Protibia lacking lateral row of spines of equal length, but with sparse strong spines. Meso and metatibia lacking crown of spines of equal length. Male tergite VIII with two lateral apophysis pointing anteriorly (Figure 2d and Figure 4c); anterior margin of female abdominal ventrite VIII with two lateral outgrowths in addition to central usual spiculum ventrale (Figure 3f and Figure 5d); female gonocoxites very elongate, spermatheca membranous (Figure 3h and Figure 5e).
Based on these morphological characters, this genus group belongs to Pholeuina, but the morphological definition of the subtribes of Leptodirini after Jeannel [4] and Guéorguiev [8], is most probably not fitting to the phylogeny of Leptodirini, which needs to be investigated through molecular analyses which are presently in progress [9,10].
3.3. Identification Key of the Genera of the “Adelopsella” Genus Group
- -
- -
4. Taxonomy
4.1. Genus Prokletijella gen. n.
Type species:Prokletijella montana sp. n., here designated.
Diagnose: In addition to the diagnosis of the “Adelopsella” genus group. Body elongate, ∼2.2 times as long as wide. Eyes with about 8-10 ommatidia of different size. Pronotum ∼1.6 times as long as wide. Aedeagus elongate, slender, asymmetric in dorsal view, endophallus with two parallel phanera of spines mixed with some strong teeth (Figure 3a,b). Endophallus is partially evaginated in the two available male specimens.
Description: As the genus is presently monospecific, the detailed description is given for genus as well as species in the species description.
Etymology: Topotypic, named after the beautiful Balkanic mountains Prokletije. Gender feminine.
4.2. Prokletijella Montana sp. n.
Holotype: : MONTENEGRO, with one label: „MONTENEGRO [MNE016] / Prokletije NP, Dolina Rapojane / Oko Skakavice, 5.v.2023, sifting / 4230’42”N, 1950’08”E, 1020-1040m / P. Hlaváč, P. Baňař & R. Kolimár lgt.” [white, printed] (PCPH).
Paratypes: 1 and 1 : same data as for holotype (PCPH, PCMP); 1 and 1 : MONTENEGRO [MNE015] / Prokletije NP, Dolina Rapojane / 4230’10”N, 1949’35”E / 1115m, sifting, 5.v.2023 / P. Hlaváč, P. Baňař & R. Kolimár lgt. [white, printed] (PCPH, PCMP); 4 , MONTENEGRO [MNE014] / Prokletije NP, Dolina Rapojane / 4229’50”N, 1949’06”E / 1150m, sifting, 5.v.2023 / P. Hlaváč, P. Baňař & R. Kolimár lgt. [white, printed] (PCPH, PCMP, PCDC); 2 and 1 , MONTENEGRO 1160 m /4229’49”N 1948’59”E / Prokletije NP, Rapojana / valley, sifting, Fagetum / A. Šima leg. 4.vi.2023 [white, printed] (PCAS).
Description: Length 2.6-3.0 mm. Body elongate of bathyscioid shape, ∼2.2 times as long as wide (Figure 1a)
Head with occipital carina and with small pigmented eyes with ∼10 ommatidies (Figure 2a). Antenna as on Figure 2g. Ratios length/width of all antennomeres as well as length of each antennomere/length of scape are given in Table 1.
Pronotum transverse, ∼1.6 times as wide as long, ∼1.05 times as wide as elytra, with lateral outlines widely rounded, posterior angles shortly rounded and slightly protruding backwards; base encompassing base of elytra; outlines of lateral sides of pronotum and elytra shaping rentrant angle; maximum width at posterior quarter; punctation fine and uniform, associated to short and recumbent setation and lacking transversal strigae.
Metatergite with short and wide posterior process (Figure 2f). Mesoventral process low, not extending over metaventrite.
Elytra ∼1.40 times as long as wide; punctation fine and uniform, associated to short and recumbent setae, similar to punctation of pronotum, lacking longitudinal stria (including the parasutural one) and lacking transversal strigae; lacking posterior wings.
Male protarsus pentamerous, female protarsus tetramerous. Male protarsus and first male mesotarsomere dilated (Figure 2b,d). Female protarsus and mesotarsus lacking dilatation. All tarsi with two interongular setae. Protibia sexually dimorphic, dilated from base to apex in males (Figure 2b), roughly parallel-sided in females (Figure 2c). Protibia lacking lateral row of spines of equal length but with sparse strong spines. Meso and metatibia lacking crown of spines of equal length.
Male genitalia: male tergite VIII with two anteriorly pointed lateral apophysis (Figure 3d). Abdominal segment IX with epipleurites of unequal length (Figure 3e). Aedeagus about 1 mm long, elongate, slender, 3.75 times as long as wide in basal part, dorsally asymmetric (Figure 3a), strongly curved in basal third in lateral view (Figure 3c). Endophallus with two parallel phanera of mixed strong spines and teeth, which begin after half of length of median lobe and slightly exceeding its apex (Figure 3a,b). This could result of a partial evagination of the endophallus on the two available male specimens. Parameres shorter than median lobe, reaching distal fifth of median lobe, with single long apical seta.
Female genitalia: anterior margin of VIIIth abdominal ventrite with two lateral outgrowths additionnally to the central usual spiculum ventrale which is as long as 1/5 of largest width of ventrite. Size of lateral outgrowths variable according to specimens and frequently slightly asymmetric. Posterior margin of the ventrite VIII also variable, sometimes regularly arcuate (Figure 3f), more frequently straight or slightly sinuous at the middle (Figure 3g). Figure 3f and Figure 3g show two different specimens to illustrate the variability, but not enougth specimens are presently available to make accurate statistics. Female genital coxites very elongate, spermatheca membranous, lacking the two sclerified basal and apical capsules which are common in most species of Leptodirini (Figure 3h).
Biology: All specimens of the type serie were collected by the sifting of the forest leaf-litter in the elevation of 1000-1150 m.
Distribution: Montenegro, Prokletije Mts. Known only from the type locality.
Etymology: The name refers to its habitat in mountains.
4.3. Genus Adelopsella Jeannel
Adelopsella Jeannel, 1908: 182, type species by monotypy: Bathyscia (Bathyscia) bosnica Reitter, 1884.
Diagnostic redescription: in addition to the diagnose of the “Adelopsella” genus group. Length 2.35-2.80 mm. Head with occipital carina. Antenna pictured on Figure 4e). Ratios length/width for all antennomeres and length of each antennomere/scape are given in table 1. Body ∼1.9 times as long as wide. Pronotum transverse, ∼1.75 times as wide as long, approximately as wide as elytra, largest width at base. Posterior angles slightly protruding backwards, outlines of lateral sides of pronotum and elytra nearly continuous. Mesoventral process low, not extending over metaventrite. Metatergite with short and thick posterior process (Figure 4f). Elytra ∼1.15 times as long as wide. Male genital segment symmetric, the epipleurites of equal length. Aedeagus symmetric in dorsal view (Figure 5a), thick and strongly curved in lateral view (Figure 5b). Female spiculum ventrale as long as 1/4 of the largest width of the ventrite VIII (Figure 5d).
Discussion: The genus contains a single species: Adelopsella bosnica Reitter. The synonymy of the two subspecies bosnica and jezerensis Jeannel, 1911 has been the subject of controversies between Jeannel and Müller. First established by Jeannel [4], the author of the subspecies jezerensis himself [3], the synonymy had been challenged by Müller [11]. We had the opportunity to observe many specimens of most of localities recorded in the litterature and of several new localities and we could not find reliable differences between populations which could justify the separation in two subspecies. So we confirm here the synonymy established by Jeannel.
DistributionAdelopsella bosnica occurs in almost all mountainous territories of Bosnia and Herzegovina. It is lacking only in the narrow stripe of low altitude plains bordering the Northern frontier with Croatia. The distribution area overlaps the Western frontier with Croatia in Troglav planina and the Southern frontier with Montenegro in Maglič (new record) and Durmitor (Sušica valley). All known localities are plotted on the map of Figure 6 and listed below.
Data from litterature: Bosnia and Herzegovina: Igman planina; Treskavica planina; Doljani [12]. Bosnia and Herzegovina: Kamenica; Zavidovici; Sarajevo; Igman planina; Bjelašnica planina; Trebević; Jajce; Šator planina; Troglav planina. Croatia: Troglav planina [4]. Bosnia and Herzegovina: Ivan planina [13]. Bosnia and Herzegovina: Vitorog planina; Vlašić planina; Vranica planina. Montenegro: Durmitor, Sušica valley [11]. Bosnia and Herzegovina: Lubovčići [14].
Data from ancient collects: Bosnia and Herzegovina: Stambulčić; Vareš; Žepcě; Vlašić planina; Ivan planina; Oštrelj (PCMP).
New records: Bosnia and Herzegovina: Hranisava Planina; Vran Planina, Omercica; Tarčin, Bjelašnica Planina; Borci, Prenj planina, Boračka draga (PCPH); Zelengora, Tjientiste, sifting near Gornje Bare lake (PCEQ). Montenegro, Maglič Mts., Mratinje valley (PCPH).
5. Zoogeography
The new genus is located ∼100 km Southeastwards from Sušica valley in Durmitor which is the nearest locality of the genus Adelopsella (map of Figure 6). This gap suggests an ancient separation from an hypothetical common ancestor which would not have left traces between the two distribution areas. A morphological support of this hypothesis is the huge morphological divergence of the aedeagus.
However, the distribution area of Adelopsella bosnica is not completely circumscribed and further investigations could extend it towards the Southeast.
6. Biology
Prokletijella has been collected by sifing litter in a doline (Rapojane Dolina). Some specimens of Adelopsella have also been collected by sifting litter as those from Boračka draga on Prenj planina or from Gornje Bare lake in Zelengora. Other specimens have been collected under large stones as specimens from Tarčin in Bjelašnica Planina or in MSS as specimens from Omercica in Vran Planina. A picture of a biotope in which Adelopsella has been collected is visible in [14].
These observations suggest that Adelopsella and Prokletijella live in similar biotopes, which are primarily MSS, but also biotopes peripheral to MSS: underside of large stones, litter...
7. The Asymmetry of Genitalia
The asymmetry of male genitalia occurs in many families of Coleoptera, in Leiodidae especially in the subfamily Cholevinae [15,16]. Generally, the asymmetry is directional, only one of the two chiral forms occurs. More rarely, both chiral forms occur in the same species, what is (improperly) called antisymmetry. In Coleoptera such antisymmetry has been observed in Ptinidae [17] and Lathriidae [18], but may be more widespread and overlooked in many cases since it needs to proceed to the dissection of many specimens to be revealed, which is rarely done (Orousset pers. comm.).
In Leiodidae, no case of antisymmetry of the aedeagus has ever been observed, but asymmetry occurs more or less frequently according to tribes [5,16]. All species of Ptomaphagini have an asymmetric aedeagus, the apical orifice being opened on the left side or the right side according to genera [5,19]. The asymmetry of the aedeagus occurs more rarely in Cholevini where it is observed only in the genera Nargus [5,20], Apterocatops [21] and Sciodrepoides [5,16]. Prokletijella is the first genus in Leptodirini with an asymmetric aedeagus.
Moreover, the asymmetry occurs also in the female terminalia, at the level of the abdominal ventrite VIII. Few observations have been devoted to an eventual correlation between asymmetric male and female genitalia in Coleoptera. The question is poorly investigated, but it occurs probably frequently. In Prokletijella, the amplitude of the asymmetry is variable in the few known specimens. This variation should be carefully investigated when further sampling will be available. But the coexistence of an asymmetric aedeagus and an asymmetric female ventrite VIII is observed also in Sciodrepoides fumatus Spence where the female spiculum ventrale is turned to the right (Figure 7a,b,c) and also in Apterocatops ennogyojya Harusawa where the spiculum ventrale is turned to the left [21] p. 213, Figure 6h,i,j,k. This correlation is not systematic in Apterocatops since Apterocatops kiimontanus Harusawa has an asymmetric aedeagus, but seemingly a symmetric female ventrite VIII [21] p. 215, Figs. 7h,i,j,k. The female ventite VIII of the two other species of Apterocatops – which have also an asymmetric aedeagus – has not been described nor illustrated in the original description and no information is available. In Nargus of the subgenus Eunargus also, with a strongly asymmetric aedeagus, the female ventrite VIII is symmetric but weakly protruding, nearly imperceptible [20] p. 275, Figure 4D. In Ptomaphagini where all species have an asymmetric aedeagus, the spiculum ventrale is completely absent.
Author Contributions
Resources, P.H.; all other topics, M.P. and P.H.
Funding
This research received no external funding.
Acknowledgments
We are grateful to Jan Růžička (Prague, Cezch Republic) and Jean Orousset (Paris, France) for fruitful discussions.
Conflicts of Interest
The author declare no conflict of interest.
References
- Hlaváč, P.; Perreau, M.; Čeplík, D. The subterranean beetles of Balkan Peninsula, Czech University of Life Sciences, Faculty of Forestry and Wood Sciences, Department of Forest Protection and Entomology: Praha. 2017; 267 pp.
- Jeannel, R. Adelopsella, nouveau genre oculé de la tribu des Bathysciini (Col.). Bull. Soc. entomol. Fr., 1908 13, 182-185.
- Jeannel, R. Révision des Bathysciinae (Coléoptères Silphides). Morphologie; distribution géographique, systématique. Arch. Zool. exp. gén. 1911 47(1), 1-641.
- Jeannel, R. Monographie des Bathysciinae. Arch. Zool. exp. gén. 1924 3(1), 1-436.
- Jeannel, R. Monographie des Catopidae. Mém. Mus. natl. Hist. nat. (n. s.) 1936 1(1), 1-433.
- Newton, A.F. Phylogenetic problems, current classification and generic catalogue of world Leiodidae (including Cholevinae), In Phylogeny and evolution of subterranean and endogean Cholevidae (=Leiodidae Cholevinae), proceedings of XX international congress of Entomology, Firenze, 1996. Atti Mus. reg. Sci nat. Torino 1998, 41-178.
- Njunjić, I.; Schilthuizen, M.; Pavićević, D.; Perreau, M. Further clarifications to the systematics of cave beetles Remyella and Rozajella (Coleoptera: Leiodidae: Cholevinae: Leptodirini). Arthropod syst. Phylogeny 2017 75(1), 141-158.
- Guéorguiev, V.B. Recherches sur la taxonomie, la classification et la phylogénie des Bathysciinae (Coleoptera Catopide). Razprave Sazu 1976 19(4), 91-129.
- Ribera, I.; Fresneda, J.; Bucur, R.; Izquierdo, A.; Vogler, A.P.; Salgado, J.M.; Cieslak, A. Ancient origin of a Western Mediterranean radiation of subterranean beetles. BMC evolutionary biology 2010 10(29, 1-14.
- Njunjić, I.; Perrard, A.; Hendriks, K.; Schilthuizen, M.; Merckx, V.; Perreau, M.; Baylac, M.; Deharveng, L. Comprehensive evolutionary analysis of the Anthroherpon radiation (Coleoptera, Leiodidae, Leptodirini). Plos One 2018 13(6), e019836. [CrossRef]
- Müller, G. Nuovi silfidi cavernicoli della Balcania e osservazioni su specie giá descritte. Atti Mus. civ. Stor. nat. Trieste 1937 13(4), 105-117. 1937.
- Sahlberg, J. Coleoptera balcanica quae mensibus Octobri et Decembri 1903 atque Martis et Aprili 1906 in peninsula balcanica collegerunt John Sahlberg et Unio Saalas. Ofvers. fin. vetensk.-soc. Förh. 1913 55[1912-1913](15), 1-108.
- Fagniez, C. Contribution à l’étude des Bathysciinae. Misc. entomol. 1927 30[1926](3), 17-25.
- Faille, A.; Bourdeau, C.; Fresneda, J. Ignacio Ribera (9.III.1963-15.IV.2020). Ann. Soc. entomol. Fr. 2021 57(2), 185-187.
- Schilthuizen, M. Something gone awry: unsolved mysteries in the evolution of asymmetric animal genitalia. Anim. Biol., 2013 63, 1-20.
- Schilthuizen, M.; de Jong, P.; van Beek, R.; Hoogenboom, T.; zu Schlochtern, M.M. The evolution of asymmetric genitalia in Coleoptera. Philos. trans. B, 2016 371, 20150400. [CrossRef]
- Orousset, J. Chiralité et antisymétrie chez les Ernobiinae et les Dorcatominae (Coleoptera Ptinidae). L’entomologiste, 2022 78(5), 322-335.
- Orousset, J.; Reisdorf, P. Chiralité et antisymétrie des genitalia mâles chez Corticarina truncatella (Mannerheim, 1844) (Coleoptera Latridiidae). L’entomologiste, 2015 71(4), 197-201.
- Jeannel, R. L’édéage, initiation aux recherches sur la systématique des coléoptères. Editions du Muséum, Paris, France, 1955; pp 1-155.
- Wang, C.-B.; Růžička J.; Zhou H.-Z. Nargus (Eunargus) celli sp. nov. (Coleoptera: Leiodidae: Cholevinae: Cholevini), a new species from China. Zootaxa 2015 4012(3), 570–580. [CrossRef]
- Harusawa, K. Descriptions of two new species of the genus Apterocatops Miyama (Coleoptera: Leiodidae: Cholevinae) from the Kii peninsula, central Japan. Entomol. Rev. Japan, 2005 60(2), 207-217.
Figure 1.
Habitus. (a) Prokletijella montana n. sp., dorsal view; (b) P. montana n. sp., ventral view; (c) Adelopsella bosnica, dorsal view.
Figure 1.
Habitus. (a) Prokletijella montana n. sp., dorsal view; (b) P. montana n. sp., ventral view; (c) Adelopsella bosnica, dorsal view.

Figure 2.
Prokletijella montanasp. n. (a) detail of the head with reduced eye (b) male protibia and protarsus; (c) female protibia and protarsus; (d) male mesotibia and mesotarsus; (e) antenna; (f) metatergite.
Figure 2.
Prokletijella montanasp. n. (a) detail of the head with reduced eye (b) male protibia and protarsus; (c) female protibia and protarsus; (d) male mesotibia and mesotarsus; (e) antenna; (f) metatergite.

Figure 3.
Prokletijella montanasp. n. genital structures. (a) aedeagus, dorsal view; (b) detail of the apex of the median lobe (dorsal view); (c) aedeagus, lateral view; (d) male abdominal tergite VIII; (e) male abdominal segment IX; (f) female abdominal ventrite VIII, dorsal view; (g) female abdominal segment IX.
Figure 3.
Prokletijella montanasp. n. genital structures. (a) aedeagus, dorsal view; (b) detail of the apex of the median lobe (dorsal view); (c) aedeagus, lateral view; (d) male abdominal tergite VIII; (e) male abdominal segment IX; (f) female abdominal ventrite VIII, dorsal view; (g) female abdominal segment IX.

Figure 4.
Adelopsella bosnica. (a) detail of the head with reduced eye; (b) male protibia and protarsus; (c) female protibia and protarsus; (d) male mesotibia and mesotarsus; (e) antenna; (f) metatergite.
Figure 4.
Adelopsella bosnica. (a) detail of the head with reduced eye; (b) male protibia and protarsus; (c) female protibia and protarsus; (d) male mesotibia and mesotarsus; (e) antenna; (f) metatergite.
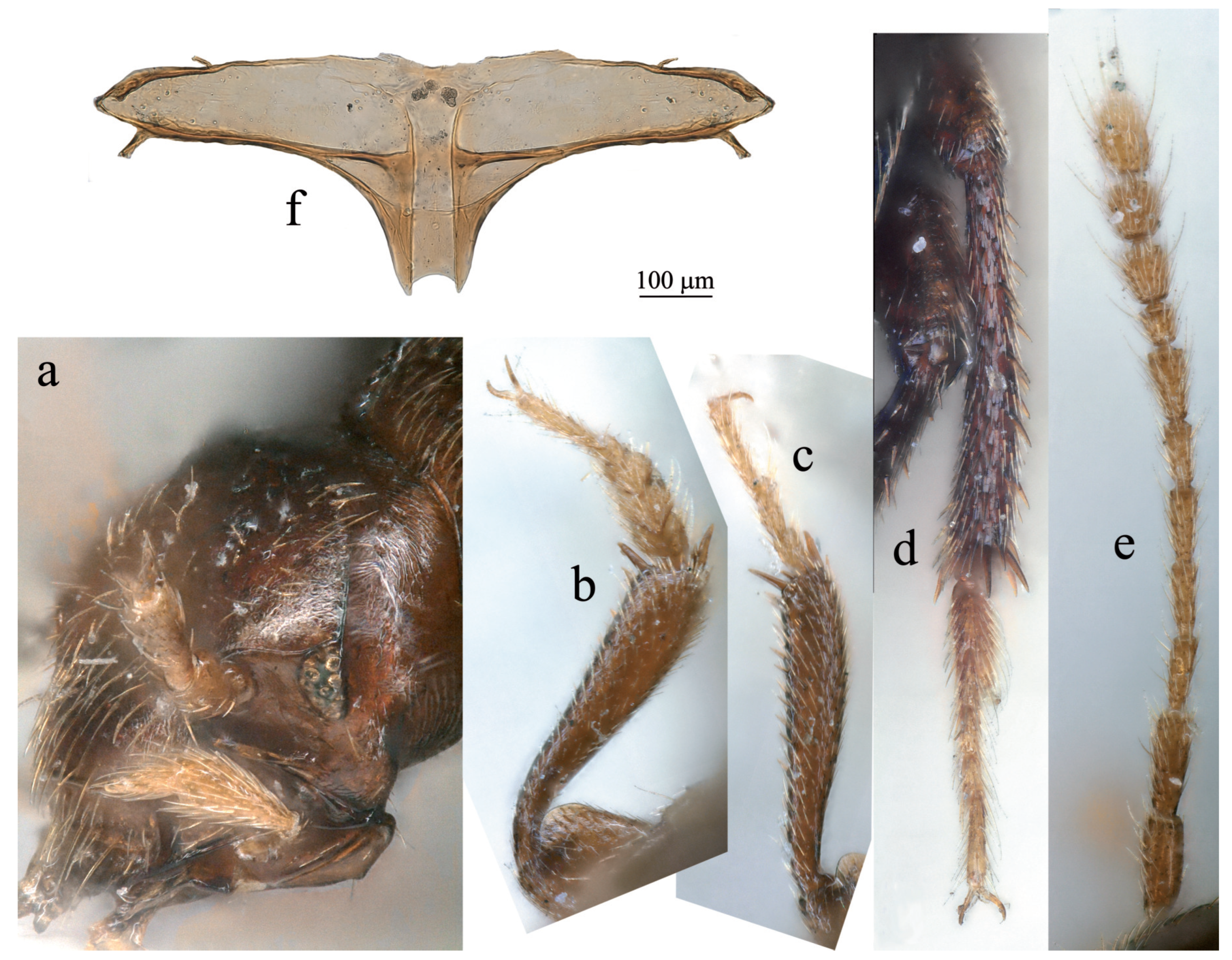
Figure 5.
Adelopsella bosnica, genital structures. (a) aedeagus, dorsal view; (b) aedeagus, lateral view; (c) male tergite VIII; (d) female abdominal ventrite VIII; (e) female abdominal segment IX.
Figure 5.
Adelopsella bosnica, genital structures. (a) aedeagus, dorsal view; (b) aedeagus, lateral view; (c) male tergite VIII; (d) female abdominal ventrite VIII; (e) female abdominal segment IX.
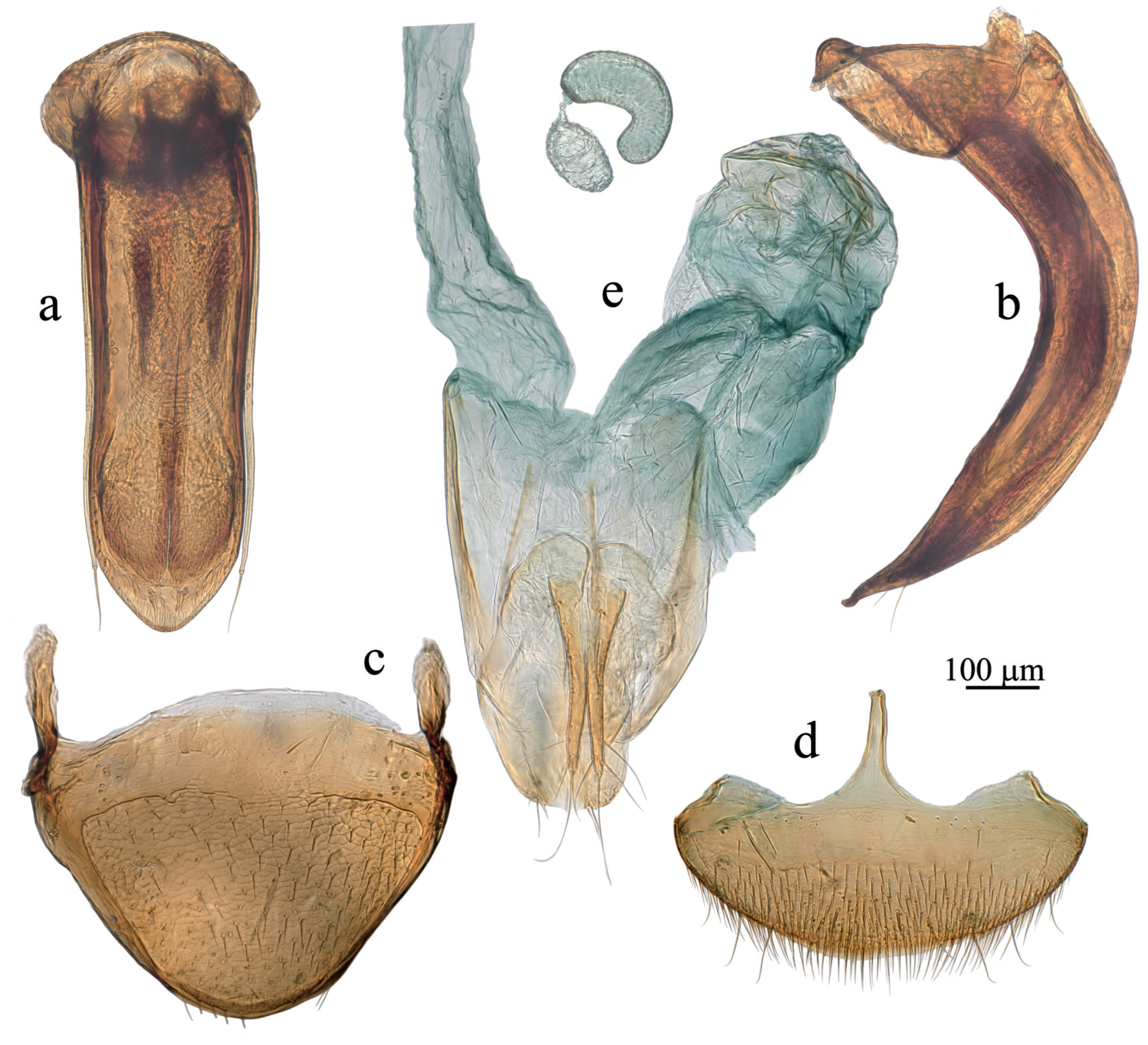
Figure 6.
Distribution map of Adelopsella bosnica and Prokletijella montana.
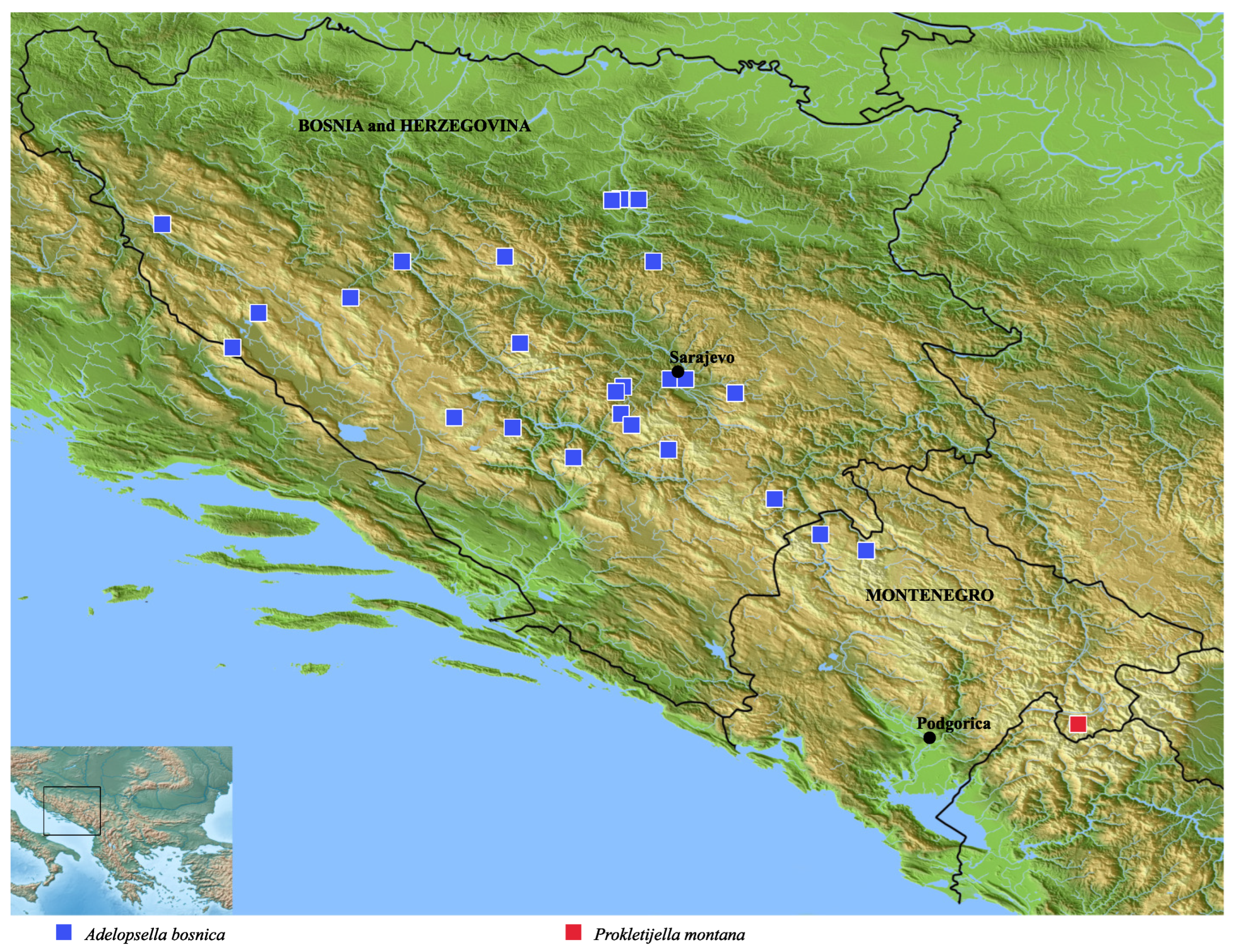
Figure 7.
Male and female genital structures of Sciodrepoides fumatus. (a) aedeagus, dorsal view; (b) aedeagus, lateral view; (c) female abdominal ventrite VIII, dorsal view.
Figure 7.
Male and female genital structures of Sciodrepoides fumatus. (a) aedeagus, dorsal view; (b) aedeagus, lateral view; (c) female abdominal ventrite VIII, dorsal view.
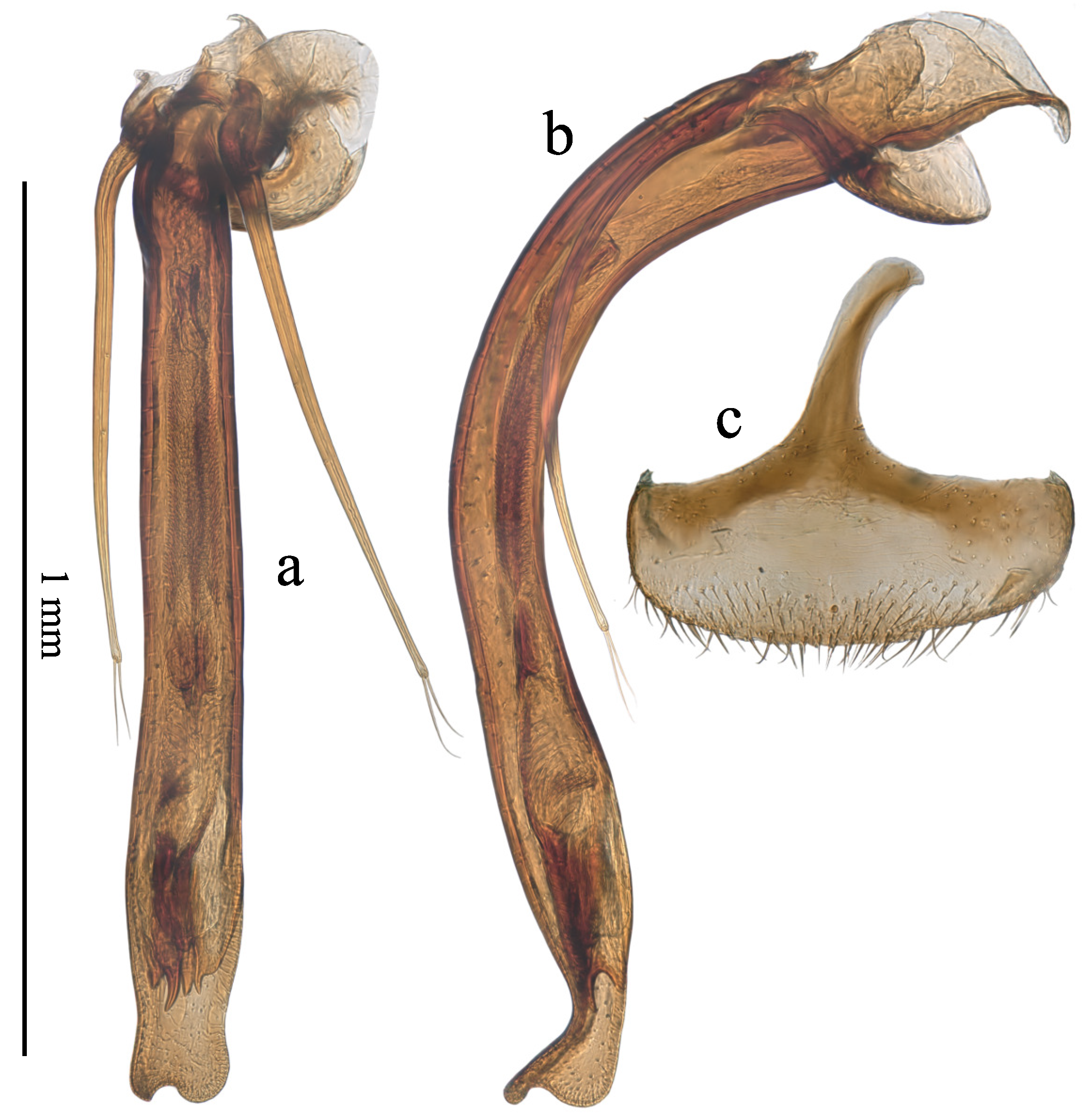

Table 1.
The ratios length/width and length/length of scape of all antennomeres.
| ]2*P. montana | length/width | 2.07 | 3.00 | 2.47 | 2.65 | 2.22 | 1.94 | 2.10 | 1.29 | 1.39 | 1.35 | 1.55 | ||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| length n/length 1 | 1.00 | 1.00 | 0.70 | 0.75 | 0.67 | 0.58 | 0.70 | 0.37 | 0.53 | 0.58 | 0.75 | |||
| ]2*A. bosnica | length/width | 2.47 | 3.21 | 2.33 | 1.47 | 2.15 | 1.86 | 1.88 | 1.62 | 1.61 | 1.09 | 1.91 | ||
| length n/length 1 | 1.00 | 0.96 | 0.60 | 0.53 | 0.60 | 0.55 | 0.68 | 0.45 | 0.62 | 0.53 | 0.89 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



