
Submitted:
05 December 2023
Posted:
06 December 2023
You are already at the latest version
Abstract
This comprehensive review delves into the cognitive neuroscience of hypnosis and variations in hypnotizability by examining research employing functional magnetic resonance imaging (fMRI), positron emission tomography (PET), and electroencephalography (EEG) methods. Key focus areas include functional brain imaging correlations in hypnosis, EEG band oscillations as indicators of hypnotic states, alterations in EEG functional connectivity during hypnosis and wakefulness, drawing critical conclusions, and suggesting future research directions.
The reviewed functional connectivity findings support the notion that disruptions in the functional integration between different components of the executive control network during hypnosis may correspond to altered subjective appraisals of the agency during the hypnotic response, as per dissociated and cold control theories of hypnosis. Building upon existing EEG results, a promising exploration avenue involves investigating how frontal lobes' integrated functioning with other cortical regions influences hypnosis and individual differences in hypnotizability during both waking and hypnotic states. Future studies investigating the effects of hypnosis on brain function should prioritize examining distinctive activation patterns across various neural networks. A practical approach entails simultaneously analyzing the causal relationships in functional connectivity linked to cognitive, physiological, and behavioral responses.
Keywords:
Functional neuroimaging
; EEG oscillations
; functional connectivity
; hypnosis
; hypnotizability
; resting
; cognitive neuroscience.
1. Introduction
A peculiar characteristic of the human brain lies in its ability to transform endogenous mental representations into perceptual states. The construction of perceptual states involves a top-down dynamic interplay between sensory processing, memory systems, attentional mechanisms, and higher-order cognitive functions. This complex orchestration allows us to construct a coherent and meaningful perception of the external world based on our internal mental representations. Such top-down modulation process is amplified by hypnosis, a social interaction in which, in the classic instance, the participant responds to verbal suggestions for imaginative experiences conveyed by the hypnotist involving distortions in the environment awareness, e.g., conscious perception and memory, and the participant's sense of agency [1,2,3,4,5]. In hypnosis, a special rapport between the hypnotized person and the hypnotist and the absorption of the hypnotist's voice is essential. During the hypnotic procedure, the subject became deeply absorbed and focused on the hypnotist's voice with disconnection from extraneous stimuli and the letting go of thoughts [2,6]. According to Reyher [7], hypnosis research can be split into "intrinsic" and "instrumental" main approaches. Intrinsic research attempts to understand the phenomena of hypnosis itself and to disclose the components of hypnotic responding.
In contrast, instrumental research uses hypnosis as a device to study a variety of psychological processes, including a dynamic interplay between sensory processing, memory systems, attentional mechanisms, and higher-order cognitive functions (involved, e.g., in functional amnesia, functional visual disorders, paranoia, false memories) and facilitating research in other fields (for details see [8]). By adopting the intrinsic approach, Cardeña and Spiegel [9] suggested three main components for hypnosis: absorption, dissociation, and hypnotic suggestibility. The term absorption refers to the degree to which one participant is intensely focused on a mental experience, while dissociation concerns the mental disengagement from the external environment [9,10,11]. Hypnotic suggestibility (or hypnotizability) refers to the individual ability to respond to hypnotic suggestions involving self-orientation and automaticity, i.e., suggested responses are experienced as being produced involuntarily and effortlessly [6,11,12]. Hypnotizability varies among people, with some individuals being more quickly and deeply hypnotizable while others may have more difficulty responding to suggestions. Experiential hypnotizability, i.e., a measure of experiential involvement in hypnotic suggestions, has been associated with the tendency to experience alterations in consciousness as boundary lessness, self-transcendence, and absorption [13]. Hypnotizability is generally considered stable across the lifespan [14]. Still, it has also been reported that it is not a stable trait and can be modified [15,16]. It varies during a single day [17,18] and between sessions [19]. However, none specifies the neural processes underlying behavioral responses to hypnotic suggestions, and little research has been focused on the specific factors that optimize responsiveness to hypnotic suggestions [20]. Lynn and collaborators [21] reported an impressive increase in hypnotic responsiveness in low hypnotizable subjects (LHs) after an 8-week training geared to increase positive expectancies about hypnosis (e.g., de-mythologize hypnosis, increase motivation and rapport, encourage imagination, teach successful strategies, manipulating performance standards). The use of subject-oriented hypnotizability training is devoted to reduce state anxiety in LHs that consequently enhances hypnotic responsiveness necessary for potential clinical treatment. These findings raise the question of whether there are differences in brain activity between people with nature high hypnotizable (HH) individuals and those trained HH ones, which can be a new research topic for future research.
Over the past two decades, extensive research into the neural aspects of hypnosis and hypnotic responsiveness has yielded tangible evidence of objective changes in the brain resulting from hypnosis. However, the findings from various studies appear to be in conflict and have sparked controversy, as detailed in reviews by Landry et al. [22] and Vanhaudenhuyse et al. [6]. Dissociated and Cold Control theories have resulted as the most used models to explain experimental hypnosis data. These theoretical models served to carry out well-designed experiments and formulate testable hypotheses (e.g., [23,24]). The Dissociated Control Theory proposes that hypnosis can be explained by a breakdown in communication between executive and monitoring systems [25,26]. In contrast, the Cold Control Theory suggests that hypnosis primarily involves unconscious executive control [27]. According to this perspective, hypnotic experiences arise from the interference of meta-representations in the selection and execution of responses, resulting in incorrect higher-order assessments of intentions for thoughts and actions [23].
Research on the neural underpinnings of agency connects executive control to the retrospective monitoring of the fluency between intentions and actions and the resultant sense of control [28,29]. While the executive system effectively responds to suggestions, hypnotized individuals' perception of a lack of ownership over their thoughts and actions is attributed to inaccurate metacognitive representations [30].
However, it is essential to note that these theories often settle on the traditional suggestion effect (expressly, the disruption of agency) and, therefore, may have limited explanatory scope. According to Terhune [31], it is improbable that a single mechanism can comprehensively elucidate the entire spectrum of hypnotic phenomena. Hypnosis is a multifaceted process encompassing absorption, embodied relaxation, alterations of self-perception, and changes in agency, and none of these elements individually constitute the entire spectrum of hypnosis phenomena. Nevertheless, neural findings, on the whole, confirmed the complexity of the hypnosis phenomena and indicated that the neural mechanisms of hypnosis and hypnotizability are not fully understood [32]. For instance, even though the relevance of hypnosis as an altered state of consciousness has been questioned and hypnotic suggestions have been consistently and efficaciously used to treat clinical pain [33,34,35], the specific top-down cognitive processes underlying responsiveness to hypnotic suggestions are still poorly explored and explained [5,36]. Barnier and Nash [37] have suggested that the lack of a widely accepted definition of hypnosis is one cause of the limited research on hypnosis and the scarce knowledge of mechanisms characterizing hypnosis. Hypnosis is a complex phenomenon embodying several elements, such as interpersonal interaction, suggestion, relaxation, focused attention, concentration, imagination, mental peace, altered perception of the environment, amnesia, change in emotional perception, disengagement of the discursive and critical analytical reasoning [38]. This complexity makes it difficult to determine whether a specific treatment should be classified as hypnosis. Thus, researchers and clinicians need to specify the definition and reference model of hypnosis they use in their work. Recently, Jensen and colleagues [36] have suggested several critical recommendations for a research agenda for the next decade to solve this question. They mainly recommend that the hypnosis researcher follow several specific suggestions for driving forward hypnosis research. These authors underlined as essential (1) the use of data sharing, (2) redirecting resources away from studies comparing state and non-state hypnosis models to neuro-clinical studies evaluating the efficacy of hypnotic treatments in the influence of Central nervous system processes, (3) the neurophysiological underpinnings of hypnotic phenomena. According to these authors, the pursuit of the signature of a supposed hypnotic state has distracted researchers away from testing more essential hypotheses and thus perceived as unproductive, time-consuming researcher resources. Hypnosis research might be better-spent testing hypotheses focused on decoding cognitive functions and neurophysiological mechanisms activated by hypnotic suggestions and responsible for individual differences in hypnotizability [39]. Our improved understanding of neurophysiological mechanisms underlying hypnosis and hypnotizability could consequently strengthen our knowledge of essential brain functions underpinning mental processes. Although scant neuroscience research has focused on the specific elements modulating responsiveness to hypnotic suggestions [20,40], recent MRI work by DeSouza and colleagues has provided a significant positive association between GABA concentration in the anterior cingulate cortex (ACC) and hypnotizability, indicating a putative neurobiological basis of individual differences in hypnotizability [41].
Additionally, this study found a negative relationship between glutamate and the absorptive ability of the participants [41]. Moreover, further work by the Spiegel group has highlighted an inverse relationship between trait hypnotizability and perseveration, an executive function that utilizes brain regions of the executive control and the salience systems. These findings suggest shared cognitive mechanisms of hypnotizability with executive functioning and information salience [42] and contribute to our understanding of the role of hypnotizability on treatment response in a psychotherapeutic context.
Interestingly, in line with previous findings, Santarcangelo and collaborators conceptualized hypnotizability as an individual trait responsible for relative variability in postural control and highlighted hypnotizability as being involved in constructing individual sensorimotor selves. The model reserved a leading role of the cerebellum in hypnotic responding and suggested the involuntariness in hypnotic responding to sensorimotor suggestions as a real effect rather than merely experienced by an HH individual [43,44]. This new conceptualization of hypnotizability seems very interesting and deserves further validation and extensions.
The present review paper aims to provide an overview of the cognitive neuroscience of hypnosis with emphasis on three related questions: (i) functional neuroimaging correlates of hypnosis and hypnotizability; (j) the electroencephalography (EEG) oscillations and their associations with hypnosis and hypnotizability; (k) the EEG connectivity findings as a signature of hypnosis and hypnotizability. The review is based on all the research/review papers providing reliable statistical information derived from PsycArticles, MEDLINE, Scopus, and Science Citation Index (years 1962-2023; key words: “hypnotic susceptibility or hypnotizability and EEG”, “hypnosis or hypnotizability and EEG oscillations and functional connectivity”, “hypnosis or hypnotizability and functional connectivity”, “hypnosis and EEG neuroimaging”, “hypnosis or hypnotizability and functional neuroimaging”, “hypnosis and EEG and resting state or hypnosis and fMRI and resting state”,). The review allows selecting one hundred-eight-two studies. The results were reported considering separately individual differences in cognitive domains: fantasy proneness, cognitive and physiological flexibility, focused attention, divided, attention, executive functions, memory, and visuospatial abilities.
2. Functional neuroimaging correlates of hypnosis and hypnotizability
Several neuroimaging studies have demonstrated that hypnosis induces plastic changes in neuronal activity (e.g., [22,38,45,46,47,48,49,50,51,52,53]. For instance, a positron emission tomography (PET) study by Maquet et al. (1999) revealed that subjects, during hypnosis, experienced significant activations in a complex neural network spanning prefrontal, cingulate, precentral, parietal, and occipital cortices while listening to pleasant autobiographical memories. Rainville et al. [51] employed PET to show that hypnotic relaxation increased occipital regional cerebral blood flow (rCBF), reduced cortical arousal, and decreased cross-modality suppression. Conversely, heightened mental absorption during hypnosis correlated with rCBF increases in a distributed network involving cortical and subcortical structures associated with the brain's attentional system. Egner et al. [45] used event-related fMRI and EEG coherence measures to link individual differences in hypnotizability to the efficiency of the frontal attention system. They found that during Stroop task performance in hypnosis, there was a functional dissociation of conflict monitoring and cognitive control processes. Muzur [54], in his review, indicates the central role of the prefrontal cortex in triggering most or even all hypnotic phenomena. Hallsband [38], using a PET scan, investigated the neural mechanisms of encoding and retrieval of high-imagery words in highly hypnotizable individuals under hypnosis and in the waking state. Encoding under hypnosis was associated with more pronounced bilateral activations in the occipital cortex and the prefrontal areas compared to learning in the waking state. Word-pair retrieval, learned under hypnosis, produced activations in the occipital lobe and cerebellum. While hypnosis hasn't been found to enhance learning and memory in healthy individuals, Lindeløv et al. [55] reported that targeted hypnotic suggestions effectively improved working memory performance in brain-lesioned patients.
Perceptual changes can also occur during hypnosis. Kosslyn et al. [49] conducted a PET study in which they demonstrated that color areas in the brain were activated under hypnosis when subjects were asked to perceive color, whether or not they were shown the color or a grey-scale stimulus. These findings highlight how changes in subjective experience during hypnosis can be reflected in brain function among highly hypnotizable subjects. Later, Halsband and collaborators [2,56], using fMRI and Granger causality mapping, confirmed previous findings that hypnosis induces activation in color processing areas when subjects are suggested to perceive color while viewing grayscale stimuli and indicate that visual illusion in hypnosis induces changes in effective connectivity among fusiform gyrus, anterior cingulate cortex, and parietal areas.
Studies on resting activity following hypnosis showed increased brain activity in the anterior part of the default mode network (DMN) during rest in HH individuals. The DMN comprises active brain regions without goal-directed activity (Figure 1). It involves the posterior cingulate cortex and precuneus, medial prefrontal and pregenual cingulate cortices, temporoparietal regions, and medial temporal lobes and is implicated in episodic memory retrieval, self-reflection, mental imagery, and stream-of-consciousness processing [57,58,59]. HH subjects showed a more pronounced selective reduction of resting state activity in the medial prefrontal cortex (i.e., the anterior parts of the default mode circuit) than LHs [9].
There is experimental evidence that suggestions within the context of hypnosis can induce dynamic changes in brain activity [32,40,60,61]. It has been observed that during hypnosis, successful suggestions provided to modify the sensory and emotional components of a given stimulus can enhance activity in the brain regions involved in the suggested sensory and emotional changes in which the modulation of functional connectivity between the ACC and the involved different brain regions is relevant [2,62]. For example, the effectiveness of hypnosis in reducing pain sensation has been well documented [46,51,62,63], and the anterior midcingulate cortex has been suggested as the brain region mediating hypnosis-induced analgesia. The findings mentioned above indicate that pain reduction under hypnosis involves enhanced functional connectivities between the midcingulate cortex and brain regions as insular, pregenual, frontal, and pre-supplementary motor area (SMA) as well as the brainstem, thalamus, and basal ganglia. These findings align with those reported in a recent pain study [64] and indicated that midcingulate cortex and insular cortex may serve as a network hub that integrates information to create the perception of pain and or at least significantly contribute to its modulation.
In a recent comprehensive review of fMRI studies, Landry and his research team [22] have offered a framework for exploring the neural correlates of hypnotic phenomena. They brought attention to the salience network (SN), executive control network (ECN), and default-mode network (DMN) as pivotal networks involved in hypnotizability, the induction of hypnosis, and the response to hypnotic suggestions (Figure 1). The SN encompasses regions such as the dorsal anterior cingulate cortex (dACC), frontoinsular cortices, precuneus posterior cingulate cortex (PCC), anterior insula (Figure 1), amygdala, and ventral striatum. This review proposed that the SN's primary function is to detect, integrate, and filter relevant interoceptive, autonomic, and emotional information [59,64]. Additionally, the ECN is central in maintaining focused attention on relevant information, deploying mental strategies to generate a reliable hypnotic response, anticipating and preparing for the hypnotic response, and assessing the subjective sense of agency. Components of the ECN include the dorsolateral prefrontal cortex (DLPFC) and lateral parietal cortices, which are critical for selecting and retaining in working memory the relevant information needed for preparing actions [59,64]. Landry and his colleagues have suggested that functions associated with the ECN could potentially contribute to generating and maintaining mental imagery, which is a valuable aspect of hypnosis. Consequently, this neural pattern is likely to influence the activity of the DMN, which may also be involved in generating mental imagery [65,66].
A decrease in activity in the frontal part of the DMN corresponds to a reduction in self-related or internally directed thoughts [9,67]. Changes in ECN activity during hypnosis may also correspond to shifts in subjective evaluations of one's agency during the hypnotic experience, as proposed by the Dissociated and Cold Control theories of hypnosis [68,69]. Research has shown that specific hypnotic suggestions can induce a credible, compelling delusion with features strikingly similar to clinical cases, e.g., mirrored self-misidentification [70]. This research delves into how hypnotic procedures affect belief formation in healthy subjects, demonstrating their ability to replicate certain aspects of monothematic delusions observed in schizophrenia patients [71]. Moreover, it has been observed that being in hypnosis alone can induce several monothematic delusional beliefs, as hypnosis impairs belief evaluation since research has shown that a hypnotic induction diminishes the ability of HHs to differentiate between suggested and actual events [72,73].
The SN plays a crucial role in processing signals from both internal and external sources to regulate how the brain responds to the detection and perception of sensory, interoceptive, and affective stimuli [74,75]. It is important to note that the activity of internal and external awareness networks has been recognized as inversely correlated [76,77,78]. The internal awareness network is primarily activated by stimuli related to oneself, while the external awareness network responds to the stimuli associated with the environment [76,77,78]. The internal awareness network involves midline brain regions such as the medial prefrontal cortex, anterior cingulate, and posterior cingulate cortex. In contrast, the external awareness network mainly engages bilateral frontal and parietal areas. The SN is activated when individuals experience hypnotic absorption and focus on specific mental experiences [11,79]; its activity may reflect the monitoring of attentional focus, prioritizing relevant signals while filtering out irrelevant ones from conscious awareness [79]. Consistent with this view, one study found that the self-reported level of hypnotic absorption correlated with the activity in two central nodes of the SN, the anterior insula and cingulate cortices [51]. Further, fMRI research [80] suggests that the significantly higher grey matter (GM) volume in the medial frontal cortex and ACC and lower connectivity in the DMN during hypnosis facilitate experiences of greater hypnotic depth. Also, greater GM volume in the left temporal-occipital cortex was associated with greater hypnotizability.
Furthermore, another study reported increased connectivity between the DMN and ECN following hypnotic induction [48].
Evidence demonstrates that the SN mediates the ECN and DMN [58,81,82,83]. Changes in the SN during hypnosis appear to correspond to profound shifts in awareness of internal and external events [84,85]. Additionally, the SN may play a coordinating role in the observed alterations in the dynamics of the ECN and DMN often observed during hypnosis [9,86,87]. Functional changes in the SN during hypnosis are closely associated with changes in awareness of both internal and external events [84,85].
In sum, hypnosis relates to reduced DMN activity, likely to reflect decreased self-referential thoughts and increased functional connectivity between the ECN and the SN [22]. This holistic perspective on explaining hypnosis is supported by substantial evidence linking ECN to the cognitive processes that underlie cognitive flexibility [88]. Nevertheless, Landry et al.'s [22] findings, derived from a meta-analysis using the Activation Likelihood Estimation method, did not support this view. Instead, the results showed that hypnosis correlates with activation of the lingual gyrus (a brain region involved in higher-order visual processing and mental imagery). In contrast, other brain regions did not display a robust pattern of association across studies [22]. However, Jiang and colleagues [48] have reported intriguing findings regarding brain activity during hypnosis. The study disclosed reduced activity in the dorsal anterior cingulate cortex (dACC) and increased connectivity between the dorsolateral prefrontal cortex (DLPFC) within the executive control network (ECN) and the insula in the salience network (SN).
Conversely, they observed a decrease in connectivity between the ECN (DLPFC) and the DMN, including the posterior cingulate cortex (PCC) [48]. Comparing these findings with the ones mentioned above reported by DeSouza et al. [41] that higher GABA concentrations in the ACC are linked to increased hypnotizability, it appears particularly intriguing that the same dACC regions have shown reduced activity during hypnosis in HHs [48]. However, in a recent review, De Benedittis [89] suggests that the role of the DLPFC in hypnosis may depend on various factors, including the type of suggestion given, which could explain the observed variations in dACC activity.
While there is a consensus regarding decreased DMN functional connectivity during hypnosis, changes in connectivity patterns are debated. Evidence suggests that hypnosis reduces activity in the dACC, increases functional connectivity between the dorsolateral prefrontal cortex (DLPFC, in the ECN) and the insula in the SN, and reduces connectivity between the ECN (DLPFC) and the DMN (PCC) [48]. Those reported by Hoeft et al. [90] align with these findings, indicating that HHs exhibited more pronounced functional connectivity between the left DLPFC and the SN.
In contrast, Demertzi et al. [87] found reduced connectivity in the posterior midline and parahippocampal structures of the DMN and increased connectivity in the lateral parietal and middle frontal areas during hypnosis. Additionally, in a study evaluating pain perception under hypnosis, Demertzi and her team [84] aimed to quantify the relationship between external and internal awareness when individuals enter a modified subjective state induced by hypnosis. Their findings demonstrated increased self-oriented processing concurrently with a heightened disconnection from the external environment. Furthermore, brain components responsible for self-awareness and external awareness were less negatively correlated during hypnosis than when individuals were at rest [84]. In a broader sense, hypnosis appears to alter resting-state fMRI networks by diminishing "extrinsic" lateral frontoparietal cortical connectivity, which might reflect a decrease in sensory awareness. In this altered state, the default mode network (DMN) exhibits increased connectivity in its lateral parietal and middle frontal regions but reduced connectivity in its posterior midline and parahippocampal structures [87]. However, it is worth noting that other studies have reported contradictory findings. Specifically, Lipari and colleagues [90] observed enhanced activity in the posterior regions of the DMN and decreased metabolic activity in the anterior DMN areas. The authors also observed DMN and complex neural network changes in non-DMN areas, including prefrontal, precentral, cingulate, parietal, and occipital areas. These findings were confirmed by a study by Oakley and Halligan [4]. According to these authors, hypnosis is associated with significant modulation of activity and connectivity that is not limited to the DMN, depending on the depth of hypnosis, the type of mental content, and emotional involvement.
For a critical discussion on divergent study findings regarding the connectivity patterns among regions of interest, see the review by Landry and colleagues [5,22].
Despite the heterogeneity among these investigations, the studies mentioned above have produced several consistent results that support participants' subjective experiences and implicate brain regions and networks associated with those phenomena. However, it is crucial to acknowledge the challenges of generalizing these results due to the diverse imaging techniques employed and the wide-ranging applications of hypnosis in different contexts. It remains an important task to extend these research findings to construct theories of hypnotic suggestion or to utilize this research for highlighting the neural mechanisms underlying responses to suggestion. Consequently, it can be challenging for research to disentangle neurophysiological patterns specific to the induction process from those associated with the actual suggestion, which is particularly crucial given findings indicating changes in the anterior cingulate cortex after induction alone [48] and in response to various suggestions [5,22,91]. Many challenges can be overcome by standardizing induction procedures and suggestion formulations across studies and using a more standardized neuroimaging method to investigate brain activity in and out of hypnosis.
3. EEG oscillations and their associations with hypnotizability and hypnosis
The EEG's oscillatory activity is traditionally defined as the superposition of oscillations falling within distinct frequency bands. The primary frequency bands of normative EEG oscillatory activity in adulthood are named delta (1– 4 Hz), theta (4– 8 Hz), alpha (8– 13 Hz), beta (13– 30 Hz), and gamma (> 35 Hz). The EEG oscillations are widely believed to play a fundamental role in brain functioning, mainly facilitating communication between different brain regions and supporting associative processes. The frequency mentioned above oscillations are linked in a complex way to various aspects of brain functions, encompassing sensory processing, perception, motor control, and cognitive activities such as attention, learning, memory, and emotional processing. These oscillations essentially serve as a functional code for the brain [92,93,94,95].
Oscillatory activity is typically quantified by its frequency (i.e., the rate of repetition of the oscillatory event), amplitude or power (i.e., the magnitude of the oscillatory signal at a given frequency), and phase (a measure of its position in time relative to a pre-defined cycle or close to a reference oscillation at the same frequency).
The EEG recordings are widely employed in current hypnosis research [32,60,69,96,97] since both EEG oscillations and the associated event-related potentials (ERPs) provide a framework for the neural syntax of complex mental activity.
Early investigations into EEG patterns during hypnosis reported an increased occurrence of occipital alpha waves in HHs compared to LHs [98,99,100,101,102,103]. Later studies have reported increased alpha activity in HHs during hypnosis [104], as well as after the hypnotic induction procedure [105,106,107]. However, findings from later studies failed to detect an increase in alpha activity with hypnosis [108,109].
The most consistent relationship between EEG activity and hypnosis is reported in the theta band (4-8 Hz) (for a more detailed review, see [40,60,97]). Several studies have reported increased spectral power in the EEG-theta band during hypnosis [105,109,110,111,112]. Crawford and collaborators [113] reported that HHs, compared to LHs, had significantly more significant hemispheric asymmetries (right greater than left) in the parietal region for all frequency bands usually associated with sustained attentional processing, in high-theta (5.5-7.45 Hz), high-alpha (11.5-13.45 Hz), and beta activity (16.5 and 25 Hz). Later, De Pascalis et al. [111] found that only HHs exhibited a higher low-band theta (4-6 Hz) amplitude in bilateral frontal and right posterior areas and a smaller alpha (8.25-10 Hz) amplitude bilaterally in the frontal cortex. Other observations of increased theta activity, particularly in HH individuals, during hypnotic inductions and suggestions[109,114] have been reported. Additionally, HHs tend to exhibit higher baseline theta activity than LHs during hypnosis and waking conditions [109,112,115,116].
There is also evidence of increased gamma activity (above 35 Hz) under hypnosis [117,118]. In an early study, Ulett and colleagues [103] measured a decrease in theta and an increase in alpha, beta, and gamma (40 Hz) activity in the right occipital cortex during hypnotic induction. Further, De Pascalis and colleagues [119,120] reported that HHs exhibited greater 40-Hz EEG amplitude density during emotional states than LHs in nonhypnotic and hypnotic conditions. However, these results were not confirmed by Crawford and coworkers' [113] observations, although these authors reported a significant beta power increase in the right parietal region. Furter, Schnyer, and Allen [121] found enhanced relative gamma band power (36-44 Hz) in HH participants experiencing recognition amnesia, suggesting that gamma power is associated with hypnotic amnesia phenomena.
Notably, the most pronounced differences in EEG patterns related to hypnotizability have been reported in the theta spectrum. However, these differences are primarily observed between HHs and LHs, excluding medium hypnotizable individuals (MHs). Crawford [122] proposed a dynamic neuropsychophysiological model of hypnosis involving the activation of the frontal-limbic attentional system. This model posits that attentional and disattentional processes are crucial in experiencing hypnosis, with low theta (3-6 Hz) and high theta (6-8 Hz) rhythms linked to these processes. Accordingly, HHs are thought to possess superior attentional filtering abilities compared to LH individuals, and these differences are reflected in underlying brain dynamics.
Sabourin et al. [109] observed that during hypnosis, both LHs and HHs increased mean theta power (4-7.75 Hz), indicating intensified attentional processes and imagery enhancement. Further research consistently found that HHs tend to exhibit more significant slow-wave theta activity than LHs, both at baseline and during hypnosis, and both groups showed an increase in slow-wave activity after hypnotic inductions [107,110]. However, in the Graffin et al. [105] study, the HHs had higher theta power (3.9-8 Hz) than LHs, mainly in frontal and temporal areas during baseline periods before hypnotic induction. In contrast, the induction of hypnosis produced a decrease in theta activity in HHs. It increased in LHs, particularly in parietal and occipital areas, whereas alpha activity increased across all participant sites.
Despite these observations, some recent studies have not found significant power changes in EEG frequency bands during hypnosis, questioning the hypothesis of theta rhythm as a definitive neurophysiological signature of hypnosis (e.g., [123,124]). Terhune and colleagues [125] also reported increased alpha2 (10.5-12 Hz) power during hypnosis but no significant differences in other EEG bands. Even studies comparing HHs to HHs in the waking state have shown mixed results, with some studies reporting higher theta power in HHs [6,105,125] and others reporting no theta differences between the groups [126].
Williams and Gruzelier [107] reported increases in alpha power (7.5-13.5 Hz) in HHs at posterior regions during the transition from pre-hypnosis to hypnosis conditions, with decreases observed after hypnosis. The reverse pattern was seen in LHs. Schnyer and Allen [121] reported that HHs exhibited a greater density in the 36-44 Hz frequency band during posthypnosis recognition amnesia, indicating their enhanced ability to maintain focused attention outside hypnosis. Later, De Pascalis et al. [111] reported higher 40 Hz EEG amplitudes in HHs during resting hypnosis conditions compared to LHs. Additionally, Croft et al. [127] found that gamma power (32–100 Hz) predicted pain ratings in non-hypnosis control conditions but not during hypnosis and hypnotic analgesia in HHs, suggesting that hypnosis may alter the relationship between gamma power and pain perception.
In our recent study, we tested the influences of hypnotizability, contextual factors, and EEG alpha on placebo analgesia [12] using multiple regression and mediation analyses. The authors found that in waking conditions, the enhancement of relative left-parietal alpha2 power directly influenced the enhancement in pain reduction and, indirectly, through the mediating positive effect of involuntariness in placebo responding. In contrast, following hypnosis, the enhanced individual left-temporoparietal upper-alpha ("alpha2") power did not directly influence pain reduction. However, the indirect mediation effect was significant through the increased involuntariness in placebo analgesia responses. Overall, this study suggests that placebo analgesia during waking and hypnosis involves different processes of top-down regulation.
Some research suggests that enhanced theta oscillatory power may be the most significant oscillatory band associated with hypnosis responding and individual differences in hypnotizability [60]. However, further research is needed to establish theta band rhythm as a definitive neural marker of hypnotizability [5]. While previous research has suggested increased alpha activity in HHs compared to LHs, along with alpha enhancements following post-hypnotic procedures [104,105], hypnosis’ effects on alpha activity have been considered inconsistent [128]. Research on the effects of hypnosis on beta, delta, and gamma activity has been few, making it challenging to formulate specific hypotheses regarding hypnosis's impact on bandwidth activities other than theta [60]. Additionally, gamma power during hypnosis has shown results in both directions, probably dependent on contextual differences and delivered hypnotic suggestions as well as on the fact that the phenomenon of theta-gamma coupling may be at work, with increases or decreases in gamma activity depending upon waxing and waning phases of theta oscillations [93]. However, more recently, hypnosis research is giving significant emphasis on phase synchrony measures in the gamma band since there is experimental evidence that gamma oscillations may play a key role in various aspects of brain function, including information processing, perception, attention, memory, emotion, and pain [129,130,131,132]. The HHs tend to experience more significant pain relief than LHs in response to hypnosis, and the first also showed significant reductions in somatosensory event-related phase-ordered gamma oscillations to the obstructive hallucination of stimulus perception during hypnosis [129].
Further, gamma oscillations bind information across different brain regions, resulting in a unified perceptual and cognitive experience [133]. Again, some studies have reported that gamma oscillations are particularly prominent during states of heightened consciousness, such as meditation and the experience of lucid dreaming [134]. However, in other studies, gamma power during hypnosis has shown mixed findings. For example, in the survey by Cardena and colleagues, it is reported that hypnosis increases gamma power in the brain [135], while in some other studies, it is shown that hypnotic induction decreases gamma power [136,137].
Hypnosis theories often have emphasized a top-down mental process within frontal networks involving attention, executive control, and cognitive monitoring in which HH individuals can direct their attention more efficiently, leading to flexible control over their attentional focus [138]. Nevertheless, we still do not fully understand the nature of gamma power fluctuations during hypnosis. According to Jensen et al. [118], the different results across these studies are likely influenced by factors such as the specific hypnotic induction used, the cortical regions under investigation, and the methodologies employed to measure gamma activity. In addition, Lynn and colleagues [139] noted methodological limitations in EEG research, which hinder definitive interpretations of findings. The absence of consistent replications may also contribute to inconsistent results, preventing conclusive statements from being made.
4. EEG Connectivity in Hypnosis and Hypnotizability
4.1. EEG Functional Connectivity
The brain's electrical activity results from dynamic interactions among distributed neural networks, displaying transient and quasi-stationary processes. The statistical dependency of physiological time series recorded from different brain areas, known as "functional connectivity," encompasses synchronous oscillatory activity crucial for neural coordination across various cell assemblies involved in multiple systems and representations [140,141]. It is widely accepted that neural synchrony is pivotal in integrating information essential for perception, cognition, emotion, and the representation of consciousness aspects such as body ownership, self-consciousness, and identity [142,143]. One prevalent assumption in hypnosis-suggestion research is that induced alterations in experiential content are associated with distinct changes in functional connectivity. EEG functional connectivity and fMRI research have provided consistent findings showing that HH individuals exhibit distinctive patterns of neural network connectivity.
Measures of dependence between multiple time series, whether linear (coherence, COH) or nonlinear (phase synchronization), can be expressed as combinations of lagged and instantaneous dependence. These measures, with values ranging from 0 to 1, indicate independence when they are zero. They are defined in the frequency domain and apply to stationary and non-stationary time series. These measures find utility in various fields, including neurophysiology, where they assess the connectivity of electric neuronal activity across different brain regions. However, it is essential to note that any measure of dependence in this context may include non-physiological contributions from volume conduction and limited spatial resolution [144]. A conventional measure of functional connectivity between two cortical regions is EEG coherence, believed to reflect the strength of interconnections between cortical areas [145]. EEG coherence between pairs of scalp locations offers valuable insights into brain states, indicating the competition between functional segregation and integration in brain dynamics. However, scalp-recorded EEG coherence has limitations, as even focal brain activity generates widespread EEG voltage patterns.
Conversely, functional connectivity measures based on fMRI and PET have limited temporal resolution and provide only indirect measures of cortical oscillatory activity. Despite its relatively poor temporal resolution, EEG coherence offers a measure of phase synchrony between two-time series, indicating the degree of functional connectivity within the cortex. However, classic EEG-derived functional connectivity measures suffer from volume conduction issues, where spatially separate electrode sites may appear functionally connected despite the absence of information flow [146]. Autoregressive models and Granger causality analysis have been suggested to improve temporal resolution and directionality estimation of information flow (for a review [147]) but do not address the volume conduction problem entirely. Consequently, several synchronization measures have been developed to estimate functional connectivity while controlling for volume conduction, including the imaginary component of coherency (iCOH), phase lag index (PLI) [148], weighted Phase Lag Index (wPLI) [149,150], and source space analysis including LORETA functional connectivity measures of phase synchronization [144,151]. All the PLIs mentioned above are considered robust estimates of the effective phase coupling between two signals because these indexes ignore phase lags of zero since the instantaneous couplings reflect the effects of volume conduction rather than any accurate coupling. These indexes vary between 0 and 1, indicating the extent to which two signals have a phase coupling, with higher values indicating stronger coupling between two brain region signals.
4.2. EEG Functional Connectivity findings under hypnosis
Gruzelier [152] has proposed a working neurophysiological model of hypnosis in which HHs under hypnosis are characterized by a reduced upper alpha band coherence between the left frontal and medial electrode pairs. In contrast, increased upper alpha coherence between the same electrode pairs characterized the LHs. The decreased coherence in HHs was interpreted as a diminished synchronization within left frontal brain activity during hypnosis.
Advanced research by Egner et al. in 2005 [45] used a combination of event-related fMRI and EEG coherence recordings with the Stroop task to test predictions from the dissociation theory that hypnosis dissociates executive control and monitoring processes (see [26,153]). These authors evaluated neural activity in the Stroop task after hypnotic induction without task-specific suggestions. The fMRI results revealed that conflict-related anterior cingulate cortex (ACC) activity interacted with hypnosis and hypnotizability. HH participants displayed increased conflict-related neural activity in the hypnosis condition compared to baseline, contrasting with LH subjects. Interestingly, activity in the dorsolateral prefrontal cortex (DLPFC), associated with attention and cognitive control, did not differ between groups and conditions.
Additionally, HH subjects exhibited a decrease in EEG gamma band coherence between frontal midline and left lateral scalp sites after hypnosis, suggesting a link between hypnotic susceptibility and the efficiency of the frontal attention system. These results indicate that hypnosis involves a functional dissociation of conflict monitoring and cognitive control processes. The findings mentioned above align with those reported by White and colleagues [124] of a decreased beta EEG coherence between medial frontal and lateral left prefrontal regions in HH during hypnosis induced by virtual reality. In contrast, LHs showed increased coherence in the same areas. Parallel findings are reported using a functional connectivity measure by Fingelkurts and colleagues in 2007 [154] in a single virtuoso participant after a "neutral" hypnotic induction examined functional connectivity. They highlighted disrupted synchrony between anterior sites across various frequency bands and a disturbance in the left frontal cortex's local and long-range functional cortical connectivity. In addition, Miltner and Weiss [155] reported decreased EEG coherence in the gamma band during hypnotic analgesia between somatosensory and frontal cortical regions, suggesting a disruption in coherent large-scale cortical oscillations. Later, Jamieson et al. [96] proved that amnesia suggestions in hypnosis directly influence memory performance. These authors conducted an EEG connectivity study focusing on hypnotic amnesia for faces as a paradigm to test if changes in topographic patterns of upper-alpha (10–12 Hz) EEG oscillations selectively inhibit the recall of memories during hypnotic amnesia (evaluated by using an old-new recognition paradigm), as the mechanism of hypnotic dissociation. Behavioral results for face recognition accuracy showed that the used paradigm elicits failures in recognizing recently presented face stimuli in the HH participants in response to a hypnotic amnesia suggestion to forget these stimuli.
Most importantly, retrieval of these stimuli is restored after the cancellation of the amnesia suggestion to a level indistinguishable from that of a group of LH participants. For previously seen faces that were wrongly identified, compared to new faces correctly identified, the upper-alpha connectivity significantly increased between right BA34 (parahippocampal gyrus) and right BAs 7, 20, and 22 in the same condition. Since integration between these regions is considered essential for recall of recent faces, during amnesia, the enhanced upper-alpha lagged connectivity was suggested to inhibit recall lying in the functional integration of these regions. Interestingly, these patterns disappeared after the reversal of amnesia suggestion. This study indicated that the inability to recognize old faces in response to the amnesia suggestion, and only this condition, is linked to significant increases in evoked upper alpha (hence functional inhibition) in the right superior parietal lobule. This region provides top-down control for face recollection. In the same condition, upper alpha phase synchrony uniquely increased between right BA7 and other functionally specific areas required for the memory of recent faces (right parahippocampal gyrus BA34, right fusiform gyrus BA20, and right middle temporal gyrus BA22).
More recently, Keshmiri et al. [156] recorded EEG in HHs and LHs to quantify in response to hypnotic induction suggestions the Differential Entropy (i.e., the average amount of variation in information in the frequency band of interest) to assess differences in information content in theta, alpha, and beta frequency bands. Results indicate that higher hypnotizability is linked to lower variability in the frequency bands mentioned above. Additionally, higher hypnotizability correlates with increased functional connectivity in parietal and parieto-occipital regions for theta and alpha, while beta shows no significant change. These helpful connectivity findings were similar to those obtained using iCOH measures reported by Jamieson and Burgess [123] between pre-hypnosis and hypnosis. However, they also extended the observed effects from neutral hypnosis to hypnotic suggestions.
Further, in terms of differential entropy changes to hypnotic suggestions, this study suggests a central role of theta activity in the transfer of information between the hippocampus and the neocortex [157] and alpha activity in information processing and transfer of information between functionally connected brain regions. These findings pointed to the engagement of the executive attentional network [11,158] during hypnotic experiences. This view found further evidence in the requirement of the attentional processes for selective enhancement of target-stimulus processing as well as inhibition of competing processes and responses [48,159,160,161].
In sum, the findings mentioned above reported reduced gamma band connectivity after hypnosis between executive control regions in the left frontal cortex and other monitoring and control regions within the frontal lobe [45,155], suggesting the operation of a breakdown in the functional integration between different components of executive control network. These findings fit well with the general model proposed by Jamieson and Woody [69], in which breakdowns in the functional integration between different components of executive control networks account for core features in the phenomenology of the hypnotic condition. Alternatively, these findings can also be explained as dissociation or shifts in subjective evaluations of one's agency during the hypnotic experience, as proposed by the Cold Control theory of hypnosis [27]. The Cold Control theory suggests that hypnosis primarily involves a change in metacognition, where hypnotic responses occur due to a lack of awareness regarding the intentions that drive cognitive or sensorimotor actions, i.e., the activation of sensorimotor network (SMN, Figure 1). For instance, stiffness in a suggested rigid arm might result from intentionally contracting opposing muscles without being conscious of that intent.
Similarly, a visual, auditory, tactile, or taste hallucination arises from imagining content without being aware of the intention behind that imagination. Research has shown that disrupting the activity of the left dorsolateral prefrontal cortex (DLPFC) can enhance hypnotic responsiveness, as seen in a study using repetitive transcranial magnetic stimulation (rTMS) [68]. However, it is worth noting that in the first experiment reported by Coltheart et al. [162], which was an exact replication of Dienes and Hutton's study [68], the authors did not find any significant change in hypnotic responsiveness after applying rTMS to the left DLPFC. However, in a second experiment by the same authors, wherein hypnotic response was measured objectively, they observed increased hypnotic responsiveness with right-sided DLPFC stimulation. Nevertheless, in a more recent study, Faerman and colleagues [42] have provided results in support of previous Dienes and Hutton findings [68]. These authors using an original personalized and targeted neuroimaging-guided transcranial magnetic stimulation (TMS, with a continuous theta-burst) to the left-dorsolateral prefrontal cortex (L-DLPFC), obtained a temporary increase of hypnotizability and subjective experience of hypnosis. In another recent study by Perri and Di Filippo [163], a unilateral inhibitory transcranial direct current stimulation (tDCS) over the left DLPFC was observed to enhance the experience of hypnotizability by 15.4% and altered a few dimensions of consciousness, such as self-awareness and absorption.
Additionally, Franz and co-workers [164] re-analyzed their-own previously published ERP data [165] to investigate whether hypnotic suggestions of a visual obstruction would influence the connectivity of the ECN by modulating the information processing in posterior attentional and lower-level brain areas of visual processing. Data were obtained from a visual oddball paradigm composed of target, standard, and distractor stimuli during hypnosis and a control condition wherein participants were asked to count the rare targets on a video screen. During hypnosis, participants received suggestions that a wooden board in front of their eyes would obstruct their view of the screen. The time-variant partial directed coherence (PDC; MultiVariate AutoRegressive Models, frequency range 1–30 Hz) was computed to measure functional connectivity between electrodes. This analysis revealed reduced directed information flow from parietal attentional to frontal executive sources while processing target stimuli. These results are in line with previous research by Gruzelier [166,167], Fingelkurts et al. [154], and Terhune et al. [168], who also observed hypofrontality and inhibition in the left hemisphere during hypnosis and provided evidence that suggestions of visual obstruction are associated with reduced information flow from the frontal to parietal regions implicated in top-down control.
4.3. EEG Functional Connectivity during rest condition
Isotani and colleagues [169] applied low-resolution electromagnetic tomography analysis (LORETA) tools for seven frequency bands (full band from 1.5 Hz to 30 Hz) in HHs and LHs during a 2-min eyes-closed resting EEG-recording preceding a hypnotic induction. The authors reported that Fast Fourier Transform (FFT) Dipole Approximation analysis had a significantly more posterior and more left source gravity center for theta (6.5–8 Hz) in HHs, whereas in these subjects, beta-1 (12.5–18 Hz) and beta-2 (18.5–21 Hz) frequencies were localized more posteriorly; LORETA source localization method in LHs showed a cortical anteriorization of beta-1 and beta-2. Finally, Global Dimensional Complexity in the whole band was higher in HHs. Thus, Isotani and colleagues indicated that in a hypnotic context, before hypnosis is induced, the HHs and LHs can be in different brain electric states with the most pronounced posterior brain activations in HHs, whereas anteriorization of brain activation patterns in LHs. In their single case study, Fingelkurts and co-workers [154] observed significant alteration, although to different extents in five frequency bands (delta, theta, alpha, beta, and gamma) during pure hypnosis (i.e., with no suggestions after induction) in a very HH participant (virtuoso). These changes remained stable even a year later. Using an Index of Structural Synchrony (ISS), the study provided evidence of local and long-distance functional connectivity changes within the cortex. The authors discovered a disruption in the functional synchronization among neural assemblies in the left frontal cortex. This finding aligns with previous research by Gruzelier [166], who also observed hypofrontality and inhibition in the left hemisphere during hypnosis. In a later study, Terhune and colleagues [168] recorded the EEG in HH and LH participants during the eyes-closed resting state in control and hypnosis conditions. Synchronization was assessed using the phase lag index (PLI), a phase synchrony measure that controls for volume conduction artifacts [148]. They observed that HHs, compared to LHs, reliably experienced a more significant dissociation state and a lower frontal-parietal phase synchrony in the alpha2 frequency band (10.5–12Hz) during hypnosis, indicating that high hypnotic suggestibility may be linked to functional disruption of the frontoparietal network. These findings are consistent with previous ones of greater posterior upper alpha power in HHs [107].
The above-reported results may reflect a shift from an anterior to a posterior neurophysiological mode, as observed in patients with schizophrenia [170]. Resting state studies on hypnosis have traditionally neglected individual differences in spontaneous perceptual states [171,172], and thus, despite quite convergent results, interpretation of these results is speculative. Preliminary research addressed this issue and observed that global functional connectivity following a minimal, hypnotic induction was negatively associated with different dimensions of consciousness, including anomalous perceptual states [135]. Cumulatively, these studies seem to suggest that HH individuals exhibit reduced frontal connectivity, although further research is needed to clarify the oscillatory specificity of these effects.
Cardeña et al. [135] recorded resting EEG during hypnosis by adopting a neurophenomenological approach to investigate neutral hypnosis (involving no specific suggestion other than to go into hypnosis). They found topographic variability in beta and gamma bands to be positively related to hypnotic depth reports amongst HHs, a finding seen consistent with the position reported in previous studies that hypnosis is associated with increased 40 Hz gamma activity, particularly among HHs (e.g., [173]). Following the neutral hypnotic induction, they observed that global functional connectivity was negatively associated with different dimensions of consciousness, including anomalous perceptual states [135].
Hypnosis resting state research by Jamieson and Burgess [123] showed increased posterior connectivity (iCOH, but not COH) from a pre-hypnosis to hypnosis condition in the theta band (4–7.9Hz) and decreased anterior connectivity in the beta1 band (13–19.9Hz), with a focus on a frontocentral and an occipital hub, that was greater in HHs compared to LHs. Interestingly, in HHs compared to LHs, the hypnotic induction elicited a qualitative change in the organization of specific control systems within the brain, seen as a potential indicator of the hypothesized hypnotic state. These changes indicated a qualitative shift in brain organization in HH participants following hypnosis, like the ones mentioned above reported by Isotani and collaborators [169] during eyes-closed resting in a hypnotic context and by Egner and colleagues [45], which reported reduced frontal EEG-gamma connectivity (COH) during a Stroop task performance following hypnosis induction in HHs. These findings support a link between hypnotic susceptibility and the efficiency of the frontal attention system with hypnotic conditions involving a functional dissociation between conflict monitoring and cognitive control processes.
Li and collaborators in a resting state EEG study [174] observed altered resting state EEG coherence in individuals undergoing hypnosis treatment for nicotine addiction. The EEG was recorded during two eyes-closed resting periods, one during a waking baseline and the other after hypnotic induction. Hypnosis produced increases in delta and theta coherence and reduced alpha and beta coherence, suggesting that hypnotic induction yields alterations in consciousness. Interestingly, this study's delta coherence between specific brain regions predicted reductions in cigarette craving [174]. More recently, Panda and colleagues [175] conducted a study comparing neutral hypnosis to an eyes-closed waking-rest condition in HH individuals. They used the weighted phase lag index (wPLI) to assess functional connectivity between brain regions and employed graph theory analyses to examine brain network topology in terms of both segregation and integration. The findings of the study revealed several key insights. During hypnosis, the authors observed decreased brain connectivity in the alpha (8–11.75 Hz) and beta-2 (20–29.75 Hz) bands, particularly in the midline and frontal-midline regions. This reduced connectivity was associated with a reduction in external awareness and a sense of dissociation from the surrounding environment. In addition, they reported an increase in delta (1–3.75 Hz) band connectivity in frontal and frontoparietal regions during hypnosis, seen as reflecting a heightened state of dissociation. Finally, bilaterally in frontal and right parietal electrodes, the authors observed increased short-range network segregation connectivity in delta and alpha bands and increased long-range integration connectivity in the beta-2 band. This modified connectivity patterns of increased short-range network segregation and long-range brain network integration was seen as reflecting a modification of the internal and external awareness for a more efficient cognitive processing and lower rates of mind-wandering during hypnosis.
Overall, findings from the above studies indicate that HH individuals exhibit reduced frontal connectivity across various frequency bands during hypnosis. Further research is required to clarify the specificity of these effects. Some of the mentioned studies suggest that variability in hypnotic suggestibility is associated with inter-individual differences in theta and gamma functional connectivity.
Our recent study tested the influences of hypnotizability, contextual factors, and EEG alpha (7.5 – 12 Hz) on placebo analgesia [12]. Using multiple regression and mediation analyses, we found: (1) during waking that the increase in the relative individual left-parietal alpha2 power, induced by the suggestive effect of a placebo analgesic cream, directly influenced the enhancement in pain reduction and, indirectly, through the mediating positive effect of involuntariness; (2) during hypnosis the relation in (1) was disrupted, however, the enhancement of individual left-temporoparietal alpha2 power, through the mediation of involuntariness, continued to influence pain reduction. The findings during waking were in line with the original ones [176] and indicated that enhanced alpha2 power may serve as a direct objective measure of the subjective reduction of tonic pain in response to placebo analgesia treatment. In addition, mediation analyses indicate that placebo analgesia during waking and hypnosis involves different processes of top-down regulation."
5. Conclusion and future directions
The report discusses the neurophysiological aspects of hypnosis by summarizing functional connectivity findings derived from studies using fMRI and EEG methodology.
Functional neuroimaging methods, such as PET and fMRI scannings, have advanced our understanding of the cognitive neuroscience of hypnosis, providing insights into the neurophysiological mechanisms underlying hypnosis, hypnotic suggestions, and hypnotizability [9,49,80,84,87,177,178]. These studies have revealed that the effects of hypnotic suggestions differ from those of imagination and have identified similar cortical activation patterns between suggested experiences and corresponding perceptual states. Researchers have also explored the mechanisms underlying the perceived involuntariness of ideomotor suggestions [179,180] and offered new models of hypnosis, one suggesting a unified theory of hypnosis and meditation states within the interoceptive predictive coding approach as an alternative view of self-regulation [85], and another suggesting a pivotal role of the cerebellum in individual differences in hypnotizability [43].
It is noteworthy, from the above-mentioned resting fMRI and structural MRI studies, that HHs, following a hypnotic induction, the hypnotized HH person during rest, relative to LH one, can reduce the anterior default mode network (DMN) activity (anterior cingulate, medial, and superior frontal gyri bilaterally) without increasing activity in other cortical regions [9]. This finding indicates that neutral hypnosis is not like a resting state, given that the DMN is a network of brain regions more active during rest than during goal-directed tasks and plays a crucial role in self-related processing, monitoring, and mind wandering. Additionally, the state of attentional absorption during a hypnotic induction has been associated with reduced DMN activity and increased prefrontal attentional system activity [86]. Moreover, research using functional connectivity derived from fMRI and EEG methods has underscored the complexity of neural mechanisms during hypnosis. Specifically, HH individuals, who usually feel the most hypnotized, have shown the highest connectivity between the bilateral DLPFC (i.e., ECN) and ipsilateral insula (i.e., SN) during hypnosis compared to rest, while for LHs, there are no differences. This enhanced connectivity during hypnosis is accompanied by reduced connectivity within DMN regions, contributing to altered agency and self-consciousness [48,181]. The intricate relationship between salience network (SN), executive control network (ECN), and DMN further delineates the neural correlates of hypnotizability. Some authors have reported the uncoupling of connectivity between the ECN and the DMN during hypnosis [48].
In contrast, others have found the contrary [9]. Spiegel and colleagues' work explained the dissociation between ECN and DMN in response to hypnotic induction as an engagement in the hypnotic state and the associated detachment from internal mental processes such as mind wandering and self-reflection. This explanation reinforces the idea of resting hypnosis as a different state of consciousness rather than a reduced level of arousal [80].
According to Demertzi and colleagues' conceptualization [84], hypnosis alters resting-state fMRI networks by diminishing "extrinsic" lateral frontoparietal cortical connectivity, which might reflect a decreased sensory awareness. In this altered state, the default mode network (DMN) exhibits increased connectivity in its lateral parietal and middle frontal regions but reduced connectivity in its posterior midline and parahippocampal structures [84,87]. However, other studies reported that connectivity in hypnosis is not limited to the DMN, depending on the depth of hypnosis and the ongoing emotional involvement and mental content, making this conceptualization questionable [90,182].
Nevertheless, a further assessment of neuroimaging assays of hypnosis using the Activation Likelihood Estimation (ALE) method by Landry et al.'s [22] findings did not align with this perspective. Instead, the study demonstrated a stronger correlation between hypnotic responses and activation of the lingual gyrus, suggesting a potential association with mental imagery.
Finally, demonstrating that noninvasive neuromodulation methods (TMS, tDCS) in humans can increase individual hypnotizability [42,68,163] opens new therapeutic interventions and offers potential pathways for enhancing hypnotic responsiveness in clinical populations.
Although neuroimaging studies on hypnosis have enhanced the knowledge of neural processes during hypnosis and individual differences in hypnotizability, there is no full convergence among revised neuroimaging findings. It is difficult to generalize the results due to the lack of comparability among various imaging techniques employed, the different measures of hypnotizability, and the different hypnotic phenomena studied.
EEG investigations during hypnosis reveal distinct patterns in frequency bands, notably with variations in alpha, theta, and gamma oscillations, and emphasize their association with attentional processes and integration of information [32,40,97], although increased theta oscillations to hypnosis responding is the most reliable finding [60]. More support for this hypothesis is necessary for understanding the mechanisms underpinning hypnosis. Moreover, research employing EEG functional connectivity has revealed that hypnosis leads to substantial connectivity changes across the spectrum of EEG oscillations [97]. Functional connectivity studies, including both EEG and fMRI recordings [45], disclosed that individual differences in hypnotizability are linked with the efficiency of the frontal attention system and that a functional dissociation of conflict monitoring and cognitive control processes characterizes the hypnotized condition. These results align with the experience of involuntariness in hypnotic responding and amnesia, suggesting reduced conflict-related activity in the anterior cingulate cortex during hypnosis. Considering that changes in ECN activity during hypnosis may correspond to shifts in subjective evaluations of one's agency during the hypnotic experience, these observations are consistent with existing models of hypnosis of Dissociated and Cold Control conceptualizations of hypnosis [68,69]. Future studies could further explore the phenomenology of hypnosis, particularly examining how the perceived reality or externality of a hallucinated image relates directly to the lack of awareness regarding the intention behind it.
Finally, studies investigating resting-state functional connectivity during hypnosis and its correlates of hypnotizability consistently suggest reduced frontal connectivity in HHs compared to LHs during hypnosis, particularly in the alpha and beta bands. Moreover, increased posterior connectivity in the theta band and decreased anterior connectivity in the beta1 band in HHs point towards a qualitative shift in brain organization, possibly indicating hypnosis.
Notably, findings from a very recent hypnosis study [175] on HHs indicated decreased brain connectivity in the alpha and beta-2 bands during hypnosis correlated with reduced external awareness and heightened dissociation. These changes were associated with increased short-range network segregation connectivity in the delta and alpha bands and increased long-range integration connectivity in the beta-2 band, underlining shifts in brain network topology.
This comprehensive understanding of hypnosis and hypnotizability, derived from a multitude of neuroimaging methodologies, not only disclosed the regions most involved in neutral hypnosis but also emphasized the need for standardized approaches and further research to comprehend the complex neural functional connections among these regions underpinning hypnosis. Integrating neuroimaging and neuromodulation techniques provides exciting prospects for unraveling the multifaceted landscape of hypnosis and its potential therapeutic applications in cognitive neuroscience.
Collectively, the evidence suggests that HH individuals during hypnosis demonstrate reduced frontal connectivity across various frequency bands, indicating potential neural markers of hypnotic suggestibility. However, further investigation is required to clarify the specificity and reliability of these oscillatory effects. Moreover, theta oscillatory power has been proposed as a predictor of hypnotic analgesia, necessitating more in-depth exploration. Recent research has also shed light on the different processes involved in top-down regulation during placebo analgesia in waking and hypnotic states, emphasizing the distinct neural mechanisms underlying pain reduction in response to suggestion and hypnotic induction.
According to Kihlstrom [108], scientific developments in this field will demand more intricate experimental frameworks to delve into the specific queries hypnosis uniquely addresses—those tied to how consciousness monitors and controls. Hypnosis unveils two critical facets: the splitting of awareness and the sensation of involuntary action.
Consider hypnotic analgesia or posthypnotic amnesia, where individuals remain oblivious to events typically within their conscious reach; sensory anesthesias render them unaware of certain sensory stimuli, while negative hallucinations block specific stimuli from their sensory field. Meanwhile, in ideomotor responses and positive hallucinations, individuals experience suggested scenarios without perceiving themselves as actively conjuring the mental images. These nuances highlight the complexities hypnosis introduces to our understanding of consciousness.
Acknowledgments
No specific funding was obtained to support this research
Conflicts of Interest
The author declare no conflict of interest
References
- Kihlstrom, J.F. The domain of hypnosis, revisited. The Oxford handbook of hypnosis: Theory, research and practice 2008, 21–52. [Google Scholar]
- Halsband, U.; Mueller, S.; Hinterberger, T.; Strickner, S. Plasticity changes in the brain in hypnosis and meditation. Contemporary Hypnosis 2009, 26, 194–215. [Google Scholar] [CrossRef]
- Kihlstrom, J.F.; Glisky, M.L.; McGovern, S.; Rapcsak, S.Z.; Mennemeier, M.S. Hypnosis in the right hemisphere. Cortex 2013, 49, 393–399. [Google Scholar] [CrossRef] [PubMed]
- Oakley, D.A.; Halligan, P.W. Hypnotic suggestion: opportunities for cognitive neuroscience. Nature Reviews Neuroscience 2013, 14, 565–576. [Google Scholar] [CrossRef]
- Terhune, D.B.; Cleeremans, A.; Raz, A.; Lynn, S.J. Hypnosis and top-down regulation of consciousness. Neuroscience & Biobehavioral Reviews 2017, 81, 59–74. [Google Scholar] [CrossRef]
- Vanhaudenhuyse, A.; Laureys, S.; Faymonville, M.E. Neurophysiology of hypnosis. Neurophysiologie Clinique/Clinical Neurophysiology 2014, 44, 343–353. [Google Scholar] [CrossRef] [PubMed]
- Reyher, J. A paradigm for determining the clinical relevance of hypnotically induced psychopathology. Psychological Bulletin 1962, 59, 344–352. [Google Scholar] [CrossRef]
- Cox, R.E.; Bryant, R.A. Advances in hypnosis research: methods, designs and contributions of intrinsic and instrumental hypnosis. The Oxford handbook of hypnosis: Theory, research, and practice 2008, 2008, 311–336. [Google Scholar]
- McGeown, W.J.; Mazzoni, G.; Venneri, A.; Kirsch, I. Hypnotic induction decreases anterior default mode activity. Consciousness and Cognition 2009, 18, 848–855. [Google Scholar] [CrossRef]
- Oakley, D.A.; Halligan, P.W. Hypnotic suggestion and cognitive neuroscience. Trends in cognitive sciences 2009, 13, 264–270. [Google Scholar] [CrossRef]
- Rainville, P.; Price, D.D. Hypnosis Phenomenology and the Neurobiology of Consciousness. International Journal of Clinical and Experimental Hypnosis 2003, 51, 105–129. [Google Scholar] [CrossRef]
- De Pascalis, V.; Scacchia, P.; Vecchio, A. Influences of hypnotic suggestibility, contextual factors, and EEG alpha on placebo analgesia. American Journal of Clinical Hypnosis 2021, 63, 302–328. [Google Scholar] [CrossRef]
- Cardeña, E.; Terhune, D.B. Hypnotizability, personality traits, and the propensity to experience alterations of consciousness. Psychology of Consciousness: Theory, Research, and Practice 2014, 1, 292–307. [Google Scholar] [CrossRef]
- Piccione, C.; Hilgard, E.R.; Zimbardo, P.G. On the degree of stability of measured hypnotizability over a 25-year period. Journal of Personality and Social Psychology 1989, 56, 289–295. [Google Scholar] [CrossRef] [PubMed]
- Gorassini, D.R.; Spanos, N.P. A social-cognitive skills approach to the successful modification of hypnotic susceptibility. Journal of Personality and Social Psychology 1986, 50, 1004–1012. [Google Scholar] [CrossRef]
- Gorassini, D.R.; Spanos, N.P. The Carleton Skill Training Program for modifying hypnotic suggestibility: Original version and variations. In Clinical hypnosis and self-regulation: Cognitive-behavioral perspectives; American Psychological Association: Washington, DC, US, 1999; pp. 141–177. [Google Scholar] [CrossRef]
- Rossi, E.L. Hypnosis and Ultradian Cycles: A New State(s) Theory of Hypnosis? American Journal of Clinical Hypnosis 1982, 25, 21–32. [Google Scholar] [CrossRef] [PubMed]
- Green, J.P.; Smith, R.J.; Kromer, M. Diurnal Variations in Hypnotic Responsiveness: Is There an Optimal Time to Be Hypnotized? International Journal of Clinical and Experimental Hypnosis 2015, 63, 171–181. [Google Scholar] [CrossRef]
- Fassler, O.; Lynn, S.J.; Knox, J. Is hypnotic suggestibility a stable trait? Consciousness and Cognition 2008, 17, 240–253. [Google Scholar] [CrossRef] [PubMed]
- Lynn, S.J.; Maxwell, R.; Green, J.P. The hypnotic induction in the broad scheme of hypnosis: A sociocognitive perspective. Hypnotic Induction 2017, 109–130. [Google Scholar] [CrossRef]
- Lynn, S.J. Hypnotic reponsiveness can be modified after hypnotic training. In Proceedings of Oral presentation at XXI world congress of medical and clinical hypnosis 2018.
- Landry, M.; Lifshitz, M.; Raz, A. Brain correlates of hypnosis: A systematic review and meta-analytic exploration. Neuroscience & Biobehavioral Reviews 2017, 81, 75–98. [Google Scholar] [CrossRef]
- Dienes, Z.; Beran, M.; Brandl, J.L.; Perner, J.; Proust, J. Is hypnotic responding the strategic relinquishment of metacognition. Foundations of metacognition 2012, 267–277. [Google Scholar]
- Jaffer, U.; Jamieson, G.A. Hypnosis and the neuroscience of cognitive and affective control. Australian Journal of Clinical & Experimental Hypnosis 2012, 40. [Google Scholar]
- Bowers, K.S. Imagination and Dissociation in Hypnotic Responding. International Journal of Clinical and Experimental Hypnosis 1992, 40, 253–275. [Google Scholar] [CrossRef]
- Woody, E.Z.; Sadler, P. Dissociation theories of hypnosis. In The Oxford Handbook of Hypnosis: Theory, Research, and Practice, A, N.M.a.B., Ed. Oxford University Press Inc.: New York, 2008; pp. 81-110.
- Dienes, Z.; Perner, J. Executive control without conscious awareness: The cold control theory of hypnosis. In Hypnosis and conscious states: The cognitive neuroscience perspective; Jamieson, G.A., Ed.; Oxford University Press: NY, 2007; pp. 293–314. [Google Scholar]
- Chambon, V.; Wenke, D.; Fleming, S.M.; Prinz, W.; Haggard, P. An Online Neural Substrate for Sense of Agency. Cerebral Cortex 2013, 23, 1031–1037. [Google Scholar] [CrossRef]
- Haggard, P.; Chambon, V. Sense of agency. Current biology 2012, 22, R390–R392. [Google Scholar] [CrossRef]
- Lush, P.; Naish, P.; Dienes, Z. Metacognition of intentions in mindfulness and hypnosis. Neuroscience of Consciousness 2016, 2016. [Google Scholar] [CrossRef]
- Terhune, D.B. Discrete response patterns in the upper range of hypnotic suggestibility: A latent profile analysis. Consciousness and Cognition 2015, 33, 334–341. [Google Scholar] [CrossRef]
- Halsband, U.; Gerhard Wolf, T. Functional Changes in Brain Activity After Hypnosis: Neurobiological Mechanisms and Application to Patients with a Specific Phobia—Limitations and Future Directions. International Journal of Clinical and Experimental Hypnosis 2019, 67, 449–474. [Google Scholar] [CrossRef]
- Jensen, M.P.; Patterson, D.R. Hypnotic approaches for chronic pain management: Clinical implications of recent research findings. American Psychologist 2014, 69, 167–177. [Google Scholar] [CrossRef] [PubMed]
- Facco, E.; Bacci, C.; Zanette, G. Hypnosis as sole anesthesia for oral surgery: The egg of Columbus. The Journal of the American Dental Association 2021, 152, 756–762. [Google Scholar] [CrossRef] [PubMed]
- Faymonville, M.; Fissette, J.; Mambourg, P.; Roediger, L.; Joris, J.; Lamy, M. Hypnosis as adjunct therapy in conscious sedation for plastic surgery. Regional anesthesia 1995, 20, 145–145. [Google Scholar]
- Jensen, M.P.; Jamieson, G.A.; Lutz, A.; Mazzoni, G.; McGeown, W.J.; Santarcangelo, E.L.; Demertzi, A.; De Pascalis, V.; Bányai, É.I.; Rominger, C. , et al. New directions in hypnosis research: strategies for advancing the cognitive and clinical neuroscience of hypnosis. Neuroscience of Consciousness 2017, 2017. [Google Scholar] [CrossRef]
- Barnier, A.J.; Nash, M.R. 1 Introduction: a roadmap for explanation, a working definition. In The Oxford Handbook of Hypnosis: Theory, Research, and Practice, Barnier, A.J., Nash, M.R., Eds. Oxford University Press: 2008. [CrossRef]
- Halsband, U. Learning in trance: Functional brain imaging studies and neuropsychology. Journal of Physiology-Paris 2006, 99, 470–482. [Google Scholar] [CrossRef] [PubMed]
- Woody, E.Z.; McConkey, K.M. What we don’t know about the Brain and Hypnosis, but need to: A View from the Buckhorn Inn. International Journal of Clinical and Experimental Hypnosis 2003, 51, 309–338. [Google Scholar] [CrossRef] [PubMed]
- Halsband, U.; Wolf, T.G. Current neuroscientific research database findings of brain activity changes after hypnosis. American Journal of Clinical Hypnosis 2021, 63, 372–388. [Google Scholar] [CrossRef] [PubMed]
- DeSouza, D.D.; Stimpson, K.H.; Baltusis, L.; Sacchet, M.D.; Gu, M.; Hurd, R.; Wu, H.; Yeomans, D.C.; Willliams, N.; Spiegel, D. Association between Anterior Cingulate Neurochemical Concentration and Individual Differences in Hypnotizability. Cerebral Cortex 2020, 30, 3644–3654. [Google Scholar] [CrossRef]
- Faerman, A.; Spiegel, D. Shared cognitive mechanisms of hypnotizability with executive functioning and information salience. Scientific Reports 2021, 11, 5704. [Google Scholar] [CrossRef] [PubMed]
- Santarcangelo, E.L. New views of hypnotizability. Frontiers in behavioral neuroscience 2014, 8, 224. [Google Scholar] [CrossRef]
- Santarcangelo, E.L.; Scattina, E. Responding to Sensorimotor Suggestions: From Endothelial Nitric Oxide to the Functional Equivalence Between Imagery and Perception. International Journal of Clinical and Experimental Hypnosis 2019, 67, 394–407. [Google Scholar] [CrossRef]
- Egner, T.; Jamieson, G.; Gruzelier, J. Hypnosis decouples cognitive control from conflict monitoring processes of the frontal lobe. NeuroImage 2005, 27, 969–978. [Google Scholar] [CrossRef]
- Faymonville, M.E.; Laureys, S.; Degueldre, C.; Del Fiore, G.; Luxen, A.; Franck, G.; Lamy, M.; Maquet, P. Neural mechanisms of antinociceptive effects of hypnosis. ANESTHESIOLOGY-PHILADELPHIA THEN HAGERSTOWN- 2000, 92, 1257–1267. [Google Scholar] [CrossRef] [PubMed]
- Grond, M.; Pawlik, G.; Walter, H.; Lesch, O.M.; Heiss, W.-D. Hypnotic catalepsy-induced changes of regional cerebral glucose metabolism. Psychiatry Research: Neuroimaging 1995, 61, 173–179. [Google Scholar] [CrossRef] [PubMed]
- Jiang, H.; White, M.P.; Greicius, M.D.; Waelde, L.C.; Spiegel, D. Brain Activity and Functional Connectivity Associated with Hypnosis. Cerebral Cortex 2017, 27, 4083–4093. [Google Scholar] [CrossRef]
- Kosslyn, S.M.; Thompson, W.L.; Costantini-Ferrando, M.F.; Alpert, N.M.; Spiegel, D. Hypnotic Visual Illusion Alters Color Processing in the Brain. American Journal of Psychiatry 2000, 157, 1279–1284. [Google Scholar] [CrossRef] [PubMed]
- Maquet, P.; Faymonville, M.E.; Degueldre, C.; Delfiore, G.; Franck, G.; Luxen, A.; Lamy, M. Functional neuroanatomy of hypnotic state. Biological Psychiatry 1999, 45, 327–333. [Google Scholar] [CrossRef]
- Rainville, P. Brain mechanisms of pain affect and pain modulation. Current opinion in neurobiology 2002, 12, 195–204. [Google Scholar] [CrossRef]
- Rainville, P.; Hofbauer, R.K.; Paus, T.; Duncan, G.H.; Bushnell, M.C.; Price, D.D. Cerebral mechanisms of hypnotic induction and suggestion. J Cogn Neurosci 1999, 11, 110–125. [Google Scholar] [CrossRef]
- Szechtman, H.; Woody, E.; Bowers, K.S.; Nahmias, C. Where the imaginal appears real: A positron emission tomography study of auditory hallucinations. Proceedings of the National Academy of Sciences 1998, 95, 1956–1960. [Google Scholar] [CrossRef]
- Muzur, A. Hypnosis and modern frontal-lobe concepts–a sketch for a review and an invitation to one particularly promising field. Collegium antropologicum 2006, 30, 205–211. [Google Scholar]
- Lindeløv, J.K.; Overgaard, R.; Overgaard, M. Improving working memory performance in brain-injured patients using hypnotic suggestion. Brain 2017, 140, 1100–1106. [Google Scholar] [CrossRef]
- Otto, T. Effective connectivity changes in hypnotic visual illusion.-A study carried out by Otto T, Halsband, U Goebel R. Master Thesis, University of Maastricht, 2007.
- Buchner, R.; Andrews-Hanna, J.; Schacter, D. The brain’s default network. Annals of the New York Academy ofSciences 2008, 1124, 1–38. [Google Scholar] [CrossRef]
- Sridharan, D.; Levitin, D.J.; Menon, V. A critical role for the right fronto-insular cortex in switching between central-executive and default-mode networks. Proceedings of the National Academy of Sciences 2008, 105, 12569–12574. [Google Scholar] [CrossRef]
- Fox, M.D.; Raichle, M.E. Spontaneous fluctuations in brain activity observed with functional magnetic resonance imaging. Nature Reviews Neuroscience 2007, 8, 700–711. [Google Scholar] [CrossRef] [PubMed]
- Jensen, M.P.; Adachi, T.; Hakimian, S. Brain Oscillations, Hypnosis, and Hypnotizability. American Journal of Clinical Hypnosis 2015, 57, 230–253. [Google Scholar] [CrossRef] [PubMed]
- Wolf, T.G.; Faerber, K.A.; Rummel, C.; Halsband, U.; Campus, G. Functional Changes in Brain Activity Using Hypnosis: A Systematic Review. Brain Sciences 2022, 12, 108. [Google Scholar] [CrossRef]
- Faymonville, M.E.; Boly, M.; Laureys, S. Functional neuroanatomy of the hypnotic state. Journal of Physiology-Paris 2006, 99, 463–469. [Google Scholar] [CrossRef] [PubMed]
- Rainville, P.; Carrier, B.t.; Hofbauer, R.K.; Bushnell, M.C.; Duncan, G.H. Dissociation of sensory and affective dimensions of pain using hypnotic modulation. Pain 1999, 82, 159–171. [Google Scholar] [CrossRef]
- Greicius, M.D.; Srivastava, G.; Reiss, A.L.; Menon, V. Default-mode network activity distinguishes Alzheimer's disease from healthy aging: evidence from functional MRI. Proceedings of the National Academy of Sciences 2004, 101, 4637–4642. [Google Scholar] [CrossRef] [PubMed]
- Daselaar, S.M.; Porat, Y.; Huijbers, W.; Pennartz, C.M.A. Modality-specific and modality-independent components of the human imagery system. NeuroImage 2010, 52, 677–685. [Google Scholar] [CrossRef]
- Zvyagintsev, M.; Clemens, B.; Chechko, N.; Mathiak, K.A.; Sack, A.T.; Mathiak, K. Brain networks underlying mental imagery of auditory and visual information. European Journal of Neuroscience 2013, 37, 1421–1434. [Google Scholar] [CrossRef]
- Lynn, S.J.; Laurence, J.-R.; Kirsch, I. Hypnosis, Suggestion, and Suggestibility: An Integrative Model. American Journal of Clinical Hypnosis 2015, 57, 314–329. [Google Scholar] [CrossRef]
- Dienes, Z.; Hutton, S. Understanding hypnosis metacognitively: rTMS applied to left DLPFC increases hypnotic suggestibility. Cortex 2013, 49, 386–392. [Google Scholar] [CrossRef]
- Jamieson, G.A.; Woody, E. Dissociated control as a paradigm for cognitive neuroscience research and theorizing in hypnosis. In Hypnosis and conscious states: The cognitive neuroscience perspective; Jamieson, G.A., Ed.; Oxford University press: NY, 2007; pp. 111–129. [Google Scholar]
- Connors, M.H. Hypnosis and belief: A review of hypnotic delusions. Consciousness and Cognition 2015, 36, 27–43. [Google Scholar] [CrossRef] [PubMed]
- Meiron, O.; Lavidor, M. Prefrontal oscillatory stimulation modulates access to cognitive control references in retrospective metacognitive commentary. Clinical Neurophysiology 2014, 125, 77–82. [Google Scholar] [CrossRef]
- Bryant, R.A.; Mallard, D. Seeing is believing: The reality of hypnotic hallucinations. Consciousness and Cognition 2003, 12, 219–230. [Google Scholar] [CrossRef] [PubMed]
- Barnier, A.J.; Cox, R.E.; O'Connor, A.; Coltheart, M.; Langdon, R.; Breen, N.; Turner, M. Developing hypnotic analogues of clinical delusions: Mirrored-self misidentification. Cognitive Neuropsychiatry 2008, 13, 406–430. [Google Scholar] [CrossRef] [PubMed]
- Uddin, L.Q. Salience processing and insular cortical function and dysfunction. Nature Reviews Neuroscience 2015, 16, 55–61. [Google Scholar] [CrossRef]
- Uddin, L.Q. Chapter 3—functions of the salience network. In Salience Network of the Human Brain; Uddin, L.Q., Ed.; Academic Press: San Diego, 2016; pp. 11–16. [Google Scholar]
- Smallwood, J.; Brown, K.; Baird, B.; Schooler, J.W. Cooperation between the default mode network and the frontal–parietal network in the production of an internal train of thought. Brain Research 2012, 1428, 60–70. [Google Scholar] [CrossRef] [PubMed]
- Demertzi, A.; Soddu, A.; Laureys, S. Consciousness supporting networks. Current Opinion in Neurobiology 2013, 23, 239–244. [Google Scholar] [CrossRef]
- Vanhaudenhuyse, A.; Demertzi, A.; Schabus, M.; Noirhomme, Q.; Bredart, S.; Boly, M.; Phillips, C.; Soddu, A.; Luxen, A.; Moonen, G. , et al. Two Distinct Neuronal Networks Mediate the Awareness of Environment and of Self. Journal of Cognitive Neuroscience 2011, 23, 570–578. [Google Scholar] [CrossRef]
- Tellegen, A.; Atkinson, G. Openness to absorbing and self-altering experiences ("absorption"), a trait related to hypnotic susceptibility. Journal of Abnormal Psychology 1974, 83, 268–277. [Google Scholar] [CrossRef] [PubMed]
- McGeown, W.J.; Mazzoni, G.; Vannucci, M.; Venneri, A. Structural and functional correlates of hypnotic depth and suggestibility. Psychiatry Research: Neuroimaging 2015, 231, 151–159. [Google Scholar] [CrossRef] [PubMed]
- Deshpande, G.; Santhanam, P.; Hu, X. Instantaneous and causal connectivity in resting state brain networks derived from functional MRI data. NeuroImage 2011, 54, 1043–1052. [Google Scholar] [CrossRef] [PubMed]
- Di, X.; Biswal, B.B. Dynamic brain functional connectivity modulated by resting-state networks. Brain Structure and Function 2015, 220, 37–46. [Google Scholar] [CrossRef]
- Gu, H.; Hu, Y.; Chen, X.; He, Y.; Yang, Y. Regional excitation-inhibition balance predicts default-mode network deactivation via functional connectivity. NeuroImage 2019, 185, 388–397. [Google Scholar] [CrossRef]
- Demertzi, A.; Vanhaudenhuyse, A.; Noirhomme, Q.; Faymonville, M.-E.; Laureys, S. Hypnosis modulates behavioural measures and subjective ratings about external and internal awareness. Journal of Physiology-Paris 2015, 109, 173–179. [Google Scholar] [CrossRef]
- Jamieson, G.A. A unified theory of hypnosis and meditation states: The interoceptive predictive coding approach. In Hypnosis and Meditation: Towards an Integrative Science of Conscious Planes; Raz, A., Lifshitz, M., Eds.; Oxford University Press: New York, NY, 2016; pp. 313–342. [Google Scholar]
- Deeley, Q.; Oakley, D.A.; Toone, B.; Giampietro, V.; Brammer, M.J.; Williams, S.C.R.; Halligan, P.W. Modulating the Default Mode Network Using Hypnosis. International Journal of Clinical and Experimental Hypnosis 2012, 60, 206–228. [Google Scholar] [CrossRef] [PubMed]
- Demertzi, A.; Soddu, A.; Faymonville, M.E.; Bahri, M.A.; Gosseries, O.; Vanhaudenhuyse, A.; Phillips, C.; Maquet, P.; Noirhomme, Q.; Luxen, A., et al. Chapter 20 - Hypnotic modulation of resting state fMRI default mode and extrinsic network connectivity. In Progress in Brain Research, Van Someren, E.J.W., Van Der Werf, Y.D., Roelfsema, P.R., Mansvelder, H.D., Lopes Da Silva, F.H., Eds. Elsevier: 2011; Vol. 193, pp. 309-322.
- D'Esposito, M.; Postle, B.R. The Cognitive Neuroscience of Working Memory. Annual Review of Psychology 2015, 66, 115–142. [Google Scholar] [CrossRef]
- De Benedittis, G. Neural Mechanisms of Hypnosis and Meditation-Induced Analgesia: A Narrative Review. International Journal of Clinical and Experimental Hypnosis 2021, 69, 363–382. [Google Scholar] [CrossRef]
- Lipari, S.; Baglio, F.; Griffanti, L.; Mendozzi, L.; Garegnani, M.; Motta, A.; Cecconi, P.; Pugnetti, L. Altered and asymmetric default mode network activity in a “hypnotic virtuoso”: An fMRI and EEG study. Consciousness and Cognition 2012, 21, 393–400. [Google Scholar] [CrossRef]
- Landry, M.; Raz, A. Hypnosis and Imaging of the Living Human Brain. American Journal of Clinical Hypnosis 2015, 57, 285–313. [Google Scholar] [CrossRef]
- Güntekin, B.; Başar, E. A review of brain oscillations in perception of faces and emotional pictures. Neuropsychologia 2014, 58, 33–51. [Google Scholar] [CrossRef] [PubMed]
- Buzsáki, G. Rhythms of The Brain Oxford University Press Inc.: NewYork 2006.
- Cohen, M.X. Analyzing neural time series data: theory and practice; MIT press: 2014.
- Keil, A.; Thigpen, N. From Neural Oscillations to Cognitive Processes. In The Oxford Handbook of EEG Frequency, Gable, P.A., Miller, M.W., Bernat, E.M., Eds. Oxford University Press: 2022; pp. 40-64. [CrossRef]
- Jamieson, G.A.; Kittenis, M.D.; Tivadar, R.I.; Evans, I.D. Inhibition of retrieval in hypnotic amnesia: dissociation by upper-alpha gating. Neuroscience of consciousness 2017, 2017, nix005. [Google Scholar] [CrossRef]
- De Pascalis, V. EEG oscillatory activity concomitant with hypnosis and hypnotizability. In International Handbook of Clinical Hypnosis, Julie H. Linden, G.D.B., Laurence I. Sugarman and Katalin Varga (Editors), Ed. Routledge: London In press.
- Bakan, P. Hypnotizability, Laterality of Eye-Movements and Functional Brain Asymmetry. Perceptual and Motor Skills 1969, 28, 927–932. [Google Scholar] [CrossRef]
- Edmonston, W.E.; Grotevant, W.R. Hypnosis and Alpha Density. American Journal of Clinical Hypnosis 1975, 17, 221–232. [Google Scholar] [CrossRef] [PubMed]
- Engstrom, D.R.; London, P.; Hart, J.T. Hypnotic Susceptibility increased by EEG Alpha Training. Nature 1970, 227, 1261–1262. [Google Scholar] [CrossRef]
- London, P.; Hart, J.T.; Leibovitz, M.P.; McDevitt, R.A. The psychophysiology of hypnotic susceptibility. In Proceedings of Psychophysiological Mechanisms of Hypnosis: An International Symposium sponsored by the International Brain Research Organization and the Centre de Recherche, Institut de Psychiatrie La Rochefoucauld, Paris; pp. 151-172.
- Morgan, A.H.; Macdonald, H.; Hilgard, E.R. EEG Alpha: Lateral Asymmetry Related to Task, and Hypnotizability. Psychophysiology 1974, 11, 275–282. [Google Scholar] [CrossRef] [PubMed]
- Ulett, G.A.; Akpinar, S.; Itil, T.M. Quantitative EEG analysis during hypnosis. Electroencephalography and Clinical Neurophysiology 1972, 33, 361–368. [Google Scholar] [CrossRef]
- De Pascalis, V.; Palumbo, G. EEG Alpha Asymmetry: Task Difficulty and Hypnotizability. Perceptual and Motor Skills 1986, 62, 139–150. [Google Scholar] [CrossRef]
- Graffin, N.F.; Ray, W.J.; Lundy, R. EEG concomitants of hypnosis and hypnotic susceptibility. Journal of Abnormal Psychology 1995, 104, 123–131. [Google Scholar] [CrossRef]
- MacLeod-Morgan, C. Hypnotic susceptibility, EEG theta and alpha waves, and hemispheric specificity. In Hypnosis 1979, G. D. Burrows, D.R.C., & L. Dennerstein, Ed. Elsevier: North Holland: Amsterdam, 1979.
- Williams, J.D.; Gruzelier, J.H. Differentiation of hypnosis and relaxation by analysis of narrow band theta and alpha frequencies. International Journal of Clinical and Experimental Hypnosis 2001, 49, 185–206. [Google Scholar] [CrossRef] [PubMed]
- Kihlstrom, J.F. Neuro-hypnotism: Prospects for hypnosis and neuroscience. Cortex 2013, 49, 365–374. [Google Scholar] [CrossRef] [PubMed]
- Sabourin, M.E.; Cutcomb, S.D.; Crawford, H.J.; Pribram, K. EEG correlates of hypnotic susceptibility and hypnotic trance: spectral analysis and coherence. International Journal of Psychophysiology 1990, 10, 125–142. [Google Scholar] [CrossRef] [PubMed]
- Crawford, H.J. Cognitive and psychophysiological correlates of hypnotic responsiveness and hypnosis. In Creative Mastery in Hypnosis and Hypnoanalysis: a Festschrift for Erika Fromm, P, F.M.L.a.B.D., Ed. Plenum Press: New York, 1990; pp. 47-54.
- De Pascalis, V.; Ray, W.J.; Tranquillo, I.; D'Amico, D. EEG activity and heart rate during recall of emotional events in hypnosis: relationships with hypnotizability and suggestibility. International Journal of Psychophysiology 1998, 29, 255–275. [Google Scholar] [CrossRef] [PubMed]
- Tebecis, A.K.; Provins, K.A.; Farnbach, R.W.; Pentony, P. Hypnosis and the EEG: A quantitative investigation. Journal of Nervous and Mental Disease 1975, 161, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Crawford, H.J.; Clarke, S.W.; Kitner-Triolo, M. Self-generated happy and sad emotions in low and highly hypnotizable persons during waking and hypnosis: laterality and regional EEG activity differences. International Journal of Psychophysiology 1996, 24, 239–266. [Google Scholar] [CrossRef]
- Jensen, M.P.; Sherlin, L.H.; Askew, R.L.; Fregni, F.; Witkop, G.; Gianas, A.; Howe, J.D.; Hakimian, S. Effects of non-pharmacological pain treatments on brain states. Clinical Neurophysiology 2013, 124, 2016–2024. [Google Scholar] [CrossRef]
- Freeman, R.; Barabasz, A.; Barabasz, M.; Warner, D. Hypnosis and Distraction Differ in Their Effects on Cold Pressor Pain. American Journal of Clinical Hypnosis 2000, 43, 137–148. [Google Scholar] [CrossRef]
- Galbraith, G.C.; London, P.; Leibovitz, M.P.; Cooper, L.M.; Hart, J.T. EEG and hypnotic susceptibility. Journal of Comparative and Physiological Psychology 1970, 72, 125–131. [Google Scholar] [CrossRef] [PubMed]
- De Benedittis, G. Neural mechanisms of hypnosis and meditation. Journal of Physiology-Paris 2015, 109, 152–164. [Google Scholar] [CrossRef] [PubMed]
- Jensen, M.P.; Adachi, T.; Tomé-Pires, C.; Lee, J.; Osman, Z.J.; Miró, J. Mechanisms of Hypnosis: Toward the Development of a Biopsychosocial Model. International Journal of Clinical and Experimental Hypnosis 2015, 63, 34–75. [Google Scholar] [CrossRef] [PubMed]
- De Pascalis, V.; Marucci, F.S.; Penna, P.M.; Pessa, E. Hemispheric activity of 40 Hz EEG during recall of emotional events: differences between low and high hypnotizables. International Journal of Psychophysiology 1987, 5, 167–180. [Google Scholar] [CrossRef] [PubMed]
- De Pascalis, V.; Marucci, F.S.; Penna, P.M. 40-Hz EEG asymmetry during recall of emotional events in waking and hypnosis: differences between low and high hypnotizables. International Journal of Psychophysiology 1989, 7, 85–96. [Google Scholar] [CrossRef] [PubMed]
- Schnyer, D.M.; Allen, J.J. Attention-related electroencephalographic and event-related potential predictors of responsiveness to suggested posthypnotic amnesia. International Journal of Clinical and Experimental Hypnosis 1995, 43, 295–315. [Google Scholar] [CrossRef] [PubMed]
- Crawford, H.J. Brain dynamics and hypnosis: attentional and disattentional processes. International Journal of Clinical and Experimental Hypnosis 1994, 42, 204–232. [Google Scholar] [CrossRef] [PubMed]
- Jamieson, G.A.; Burgess, A.P. Hypnotic induction is followed by state-like changes in the organization of EEG functional connectivity in the theta and beta frequency bands in high-hypnotically susceptible individuals. Frontiers in Human Neuroscience 2014, 8. [Google Scholar] [CrossRef] [PubMed]
- White, D.; Ciorciari, J.; Carbis, C.; Liley, D. EEG Correlates of Virtual Reality Hypnosis. International Journal of Clinical and Experimental Hypnosis 2009, 57, 94–116. [Google Scholar] [CrossRef]
- Kirenskaya, A.V.; Novototsky-Vlasov, V.Y.; Zvonikov, V.M. Waking EEG Spectral Power and Coherence Differences Between High and Low Hypnotizable Subjects. International Journal of Clinical and Experimental Hypnosis 2011, 59, 441–453. [Google Scholar] [CrossRef]
- De Pascalis, V. Psychophysiological correlates of hypnosis and hypnotic susceptibility. International Journal of Clinical and Experimental Hypnosis 1999, 47, 117–143. [Google Scholar] [CrossRef]
- Croft, R.J.; Williams, J.D.; Haenschel, C.; Gruzelier, J.H. Pain perception, hypnosis and 40 Hz oscillations. International Journal of Psychophysiology 2002, 46, 101–108. [Google Scholar] [CrossRef] [PubMed]
- Crawford, H.J.; Gruzelier, J.H. A midstream view of the neuropsychophysiology of hypnosis: Recent research and future directions. In Contemporary hypnosis research; Guilford Press: New York, NY, US, 1992; pp. 227–266. [Google Scholar]
- De Pascalis, V.; Cacace, I.; Massicolle, F. Perception and modulation of pain in waking and hypnosis: functional significance of phase-ordered gamma oscillations. Pain 2004, 112, 27–36. [Google Scholar] [CrossRef]
- Başar, E. Brain Function and Oscillations: II. Integrative Brain Function. Neurophysiology and Cognitive Processes; Springer-Verlag: Heidelberg, 1999. [Google Scholar]
- Başar, E.; Schürmann, M.; Başar-Eroglu, C.; Demiralp, T. Selectively distributed gamma band system of the brain. International Journal of Psychophysiology 2001, 39, 129–135. [Google Scholar] [CrossRef]
- Karakaş, S.; Başar, E. Early gamma response is sensory in origin: a conclusion based on cross-comparison of results from multiple experimental paradigms. International Journal of Psychophysiology 1998, 31, 13–31. [Google Scholar] [CrossRef] [PubMed]
- Buzsaki, G.; Draguhn, A. Neuronal oscillations in cortical networks. science 2004, 304, 1926–1929. [Google Scholar] [CrossRef] [PubMed]
- Lutz, A.; Greischar, L.L.; Rawlings, N.B.; Ricard, M.; Davidson, R.J. Long-term meditators self-induce high-amplitude gamma synchrony during mental practice. Proceedings of the national Academy of Sciences 2004, 101, 16369–16373. [Google Scholar] [CrossRef] [PubMed]
- Cardeña, E.; Jönsson, P.; Terhune, D.B.; Marcusson-Clavertz, D. The neurophenomenology of neutral hypnosis. Cortex 2013, 49, 375–385. [Google Scholar] [CrossRef]
- Hiltunen, S.; Karevaara, M.; Virta, M.; Makkonen, T.; Kallio, S.; Paavilainen, P. No evidence for theta power as a marker of hypnotic state in highly hypnotizable subjects. Heliyon 2021, 7. [Google Scholar] [CrossRef]
- Hinterberger, T.; Schöner, J.; Halsband, U. Analysis of Electrophysiological State Patterns and Changes During Hypnosis Induction. International Journal of Clinical and Experimental Hypnosis 2011, 59, 165–179. [Google Scholar] [CrossRef]
- Raz, A. Hypnosis: A twilight zone of the top-down variety: Few have never heard of hypnosis but most know little about the potential of this mind–body regulation technique for advancing science. Trends in cognitive sciences 2011, 15, 555–557. [Google Scholar] [CrossRef]
- Lynn, S.J.; Kirsch, I.; Knox, J.; Fassler, O.; Lilienfeld, S.O. Hypnosis and neuroscience: implications for the altered state debate. In Hypnosis and conscious states: The cognitive neuroscience perspective; Jamieson, G.A., Ed.; Oxford University Press: NY, 2007; pp. 145–165. [Google Scholar]
- Fingelkurts, A.A.; Fingelkurts, A.A.; Kähkönen, S. Functional connectivity in the brain—is it an elusive concept? Neuroscience & Biobehavioral Reviews 2005, 28, 827–836. [Google Scholar] [CrossRef]
- Lee, L.; Harrison, L.M.; Mechelli, A. A report of the functional connectivity workshop, Dusseldorf 2002. NeuroImage 2003, 19, 457–465. [Google Scholar] [CrossRef]
- Varela, F.; Lachaux, J.-P.; Rodriguez, E.; Martinerie, J. The brainweb: Phase synchronization and large-scale integration. Nature Reviews Neuroscience 2001, 2, 229–239. [Google Scholar] [CrossRef]
- Tsakiris, M.; Hesse, M.D.; Boy, C.; Haggard, P.; Fink, G.R. Neural Signatures of Body Ownership: A Sensory Network for Bodily Self-Consciousness. Cerebral Cortex 2007, 17, 2235–2244. [Google Scholar] [CrossRef]
- Pascual-Marqui, R.D. Instantaneous and lagged measurements of linear and nonlinear dependence between groups of multivariate time series: frequency decomposition. arXiv 2007, arXiv:0711.1455. [Google Scholar]
- Bullock, T.H.; McClune, M.C.; Enright, J.T. Are the electroencephalograms mainly rhythmic? Assessment of periodicity in wide-band time series. Neuroscience 2003, 121, 233–252. [Google Scholar] [CrossRef]
- Burgess, A. On the contribution of neurophysiology to hypnosis research: current state and future directions. In Hypnosis and conscious states: The cognitive neuroscience perspective; Jamieson, G.A., Ed.; Oxford University Press: NY, 2007; pp. 195–219. [Google Scholar]
- Pereda, E.; Quiroga, R.Q.; Bhattacharya, J. Nonlinear multivariate analysis of neurophysiological signals. Progress in Neurobiology 2005, 77, 1–37. [Google Scholar] [CrossRef]
- Stam, C.J.; Nolte, G.; Daffertshofer, A. Phase lag index: Assessment of functional connectivity from multi channel EEG and MEG with diminished bias from common sources. Human Brain Mapping 2007, 28, 1178–1193. [Google Scholar] [CrossRef]
- Hardmeier, M.; Hatz, F.; Bousleiman, H.; Schindler, C.; Stam, C.J.; Fuhr, P. Reproducibility of functional connectivity and graph measures based on the phase lag index (PLI) and weighted phase lag index (wPLI) derived from high resolution EEG. PloS one 2014, 9, e108648. [Google Scholar] [CrossRef]
- Vinck, M.; Oostenveld, R.; van Wingerden, M.; Battaglia, F.; Pennartz, C.M.A. An improved index of phase-synchronization for electrophysiological data in the presence of volume-conduction, noise and sample-size bias. NeuroImage 2011, 55, 1548–1565. [Google Scholar] [CrossRef] [PubMed]
- Pascual-Marqui, R.D.; Lehmann, D.; Koukkou, M.; Kochi, K.; Anderer, P.; Saletu, B.; Tanaka, H.; Hirata, K.; John, E.R.; Prichep, L. Assessing interactions in the brain with exact low-resolution electromagnetic tomography. Philosophical Transactions of the Royal Society of London A: Mathematical, Physical and Engineering Sciences 2011, 369, 3768–3784. [Google Scholar] [CrossRef] [PubMed]
- Gruzelier, J.H. A working model of the neurophysiology of hypnosis: a review of evidence. Contemporary Hypnosis 1998, 15, 3–21. [Google Scholar] [CrossRef]
- Woody, E.Z.; Farvolden, P. Dissociation in Hypnosis and Frontal Executive Function. American Journal of Clinical Hypnosis 1998, 40, 206–216. [Google Scholar] [CrossRef] [PubMed]
- Fingelkurts, A.A.; Fingelkurts, A.A.; Kallio, S.; Revonsuo, A. Cortex functional connectivity as a neurophysiological correlate of hypnosis: An EEG case study. Neuropsychologia 2007, 45, 1452–1462. [Google Scholar] [CrossRef] [PubMed]
- Miltner, W.H.; Weiss, T. Cortical mechanisms of hypnotic pain control. In Hypnosis and conscious states: The cognitive neuroscience perspective; Jamieson, G.A., Ed.; Oxford University Press: NY, 2007; pp. 51–66. [Google Scholar]
- Keshmiri, S.; Alimardani, M.; Shiomi, M.; Sumioka, H.; Ishiguro, H.; Hiraki, K. Higher hypnotic suggestibility is associated with the lower EEG signal variability in theta, alpha, and beta frequency bands. PLOS ONE 2020, 15, e0230853. [Google Scholar] [CrossRef]
- Buzsáki, G. The Hippocampo-Neocortical Dialogue. Cerebral Cortex 1996, 6, 81–92. [Google Scholar] [CrossRef]
- Bell, V.; Oakley, D.A.; W., H.P.; Deeley, Q. Dissociation in hysteria and hypnosis: evidence from cognitive neuroscience. Journal of Neurology, Neurosurgery Psychiatry 2011, 82, 332. [CrossRef]
- Posner, M.I.; Dehaene, S. Attentional networks. Trends in neurosciences 1994, 17, 75–79. [Google Scholar] [CrossRef]
- Botvinick, M.; Nystrom, L.E.; Fissell, K.; Carter, C.S.; Cohen, J.D. Conflict monitoring versus selection-for-action in anterior cingulate cortex. Nature 1999, 402, 179–181. [Google Scholar] [CrossRef]
- Carter, C.S.; Macdonald, A.M.; Botvinick, M.; Ross, L.L.; Stenger, V.A.; Noll, D.; Cohen, J.D. Parsing executive processes: strategic vs. evaluative functions of the anterior cingulate cortex. Proceedings of the National Academy of Sciences 2000, 97, 1944–1948. [Google Scholar] [CrossRef] [PubMed]
- Coltheart, M.; Cox, R.; Sowman, P.; Morgan, H.; Barnier, A.; Langdon, R.; Connaughton, E.; Teichmann, L.; Williams, N.; Polito, V. Belief, delusion, hypnosis, and the right dorsolateral prefrontal cortex: A transcranial magnetic stimulation study. Cortex 2018, 101, 234–248. [Google Scholar] [CrossRef]
- Perri, R.L.; Di Filippo, G. Alteration of hypnotic experience following transcranial electrical stimulation of the left prefrontal cortex. International Journal of Clinical and Health Psychology 2023, 23, 100346. [Google Scholar] [CrossRef]
- Franz, M.; Schmidt, B.; Hecht, H.; Naumann, E.; Miltner, W.H.R. Suggested visual blockade during hypnosis: Top-down modulation of stimulus processing in a visual oddball task. PLOS ONE 2021, 16, e0257380. [Google Scholar] [CrossRef]
- Schmidt, B.; Hecht, H.; Naumann, E.; Miltner, W.H.R. The Power of mind: Blocking visual perception by hypnosis. Scientific Reports 2017, 7, 4889. [Google Scholar] [CrossRef]
- Gruzelier, J.H. Redefining hypnosis: theory, methods and integration. Contemporary Hypnosis 2000, 17, 51–70. [Google Scholar] [CrossRef]
- Gruzelier, J.H. Frontal functions, connectivity and neural efficiency underpinning hypnosis and hypnotic susceptibility. Contemporary Hypnosis 2006, 23, 15–32. [Google Scholar] [CrossRef]
- Terhune, D.B.; Cardeña, E.; Lindgren, M. Differential frontal-parietal phase synchrony during hypnosis as a function of hypnotic suggestibility. Psychophysiology 2011, 48, 1444–1447. [Google Scholar] [CrossRef]
- Isotani, T.; Lehmann, D.; Pascual-Marqui, R.D.; Kochi, K.; Wackermann, J.; Saito, N.; Yagyu, T.; Kinoshita, T.; Sasada, K. EEG Source Localization and Global Dimensional Complexity in High- and Low- Hypnotizable Subjects: A Pilot Study. Neuropsychobiology 2001, 44, 192–198. [Google Scholar] [CrossRef] [PubMed]
- Holt, D.J.; Cassidy, B.S.; Andrews-Hanna, J.R.; Lee, S.M.; Coombs, G.; Goff, D.C.; Gabrieli, J.D.; Moran, J.M. An Anterior-to-Posterior Shift in Midline Cortical Activity in Schizophrenia During Self-Reflection. Biological Psychiatry 2011, 69, 415–423. [Google Scholar] [CrossRef] [PubMed]
- Cardeña, E. The phenomenology of deep hypnosis: quiescent and physically active. International Journal of Clinical and Experimental Hypnosis 2005, 53, 37–59. [Google Scholar] [CrossRef]
- Pekala, R.J.; Kumar, V. An empirical-phenomenological approach to quantifying consciousness and states of consciousness: With particular reference to understanding the nature of hypnosis. In Hypnosis and conscious states: The cognitive neuroscience perspective, Jamieson, G.A., Ed. Oxford University Press: NY, 2007; pp. 167-194.
- De Pascalis, V. Phase-ordered gamma oscillations and the modulation of hypnotic experience. In Hypnosis and conscious states: The cognitive neuroscience perspective, Jamieson, G.A., Ed. Oxford University Press: NY, 2007; pp. 67-89.
- Li, X.; Ma, R.; Pang, L.; Lv, W.; Xie, Y.; Chen, Y.; Zhang, P.; Chen, J.; Wu, Q.; Cui, G. , et al. Delta coherence in resting-state EEG predicts the reduction in cigarette craving after hypnotic aversion suggestions. Scientific Reports 2017, 7, 2430. [Google Scholar] [CrossRef] [PubMed]
- Panda, R.; Vanhaudenhuyse, A.; Piarulli, A.; Annen, J.; Demertzi, A.; Alnagger, N.; Chennu, S.; Laureys, S.; Faymonville, M.-E.; Gosseries, O. Altered Brain Connectivity and Network Topological Organization in a Non-ordinary State of Consciousness Induced by Hypnosis. Journal of Cognitive Neuroscience 2023, 35, 1394–1409. [Google Scholar] [CrossRef]
- Li, L.; Wang, H.; Ke, X.; Liu, X.; Yuan, Y.; Zhang, D.; Xiong, D.; Qiu, Y. Placebo analgesia changes alpha oscillations induced by tonic muscle pain: EEG frequency analysis including data during pain evaluation. Frontiers in computational neuroscience 2016, 10. [Google Scholar] [CrossRef]
- Derbyshire, S.W.G.; Whalley, M.G.; Stenger, V.A.; Oakley, D.A. Cerebral activation during hypnotically induced and imagined pain. NeuroImage 2004, 23, 392–401. [Google Scholar] [CrossRef]
- Cojan, Y.; Waber, L.; Schwartz, S.; Rossier, L.; Forster, A.; Vuilleumier, P. The brain under self-control: modulation of inhibitory and monitoring cortical networks during hypnotic paralysis. Neuron 2009, 62, 862–875. [Google Scholar] [CrossRef] [PubMed]
- Blakemore, S.J.; Oakley, D.A.; Frith, C.D. Delusions of alien control in the normal brain. Neuropsychologia 2003, 41, 1058–1067. [Google Scholar] [CrossRef]
- Deeley, Q.; Walsh, E.; Oakley, D.A.; Bell, V.; Koppel, C.; Mehta, M.A.; Halligan, P.W. Using Hypnotic Suggestion to Model Loss of Control and Awareness of Movements: An Exploratory fMRI Study. PLOS ONE 2013, 8, e78324. [Google Scholar] [CrossRef] [PubMed]
- Hoeft, F.; Gabrieli, J.D.E.; Whitfield-Gabrieli, S.; Haas, B.W.; Bammer, R.; Menon, V.; Spiegel, D. Functional Brain Basis of Hypnotizability. Archives of General Psychiatry 2012, 69, 1064–1072. [Google Scholar] [CrossRef]
- Oakley, D.A. Hypnosis and Conversion Hysteria: A Unifying Model. Cognitive Neuropsychiatry 1999, 4, 243–265. [Google Scholar] [CrossRef]
Figure 1.
Most reported studies used models to select spatial maps for functional connectivity. The following brain regions constitute four neural networks: default mode network (DMN, upper-left quadrant); salience network (SN, upper-right quadrant); executive control network (ECN, bottom-left quadrant); sensorimotor network (SMN, bottom-right quadrant). Abbreviations: SFG, superior frontal gyrus; LPC, lateral parietal cortex; ITG, inferior temporal gyrus; MTG, middle temporal gyrus; PHG, parahippocampal gyrus; IFG, inferior frontal gyrus; Ins, insula; MFG, middle frontal gyrus; SPL, superior parietal lobe; ACC, anterior cingulate cortex; OrbG, orbital gyrus; PreCu, precuneus; PCC, posterior cingulate cortex; LG, lingual gyrus; SMA, supplementary motor area; dACC, dorsal anterior cingulate cortex.
Figure 1.
Most reported studies used models to select spatial maps for functional connectivity. The following brain regions constitute four neural networks: default mode network (DMN, upper-left quadrant); salience network (SN, upper-right quadrant); executive control network (ECN, bottom-left quadrant); sensorimotor network (SMN, bottom-right quadrant). Abbreviations: SFG, superior frontal gyrus; LPC, lateral parietal cortex; ITG, inferior temporal gyrus; MTG, middle temporal gyrus; PHG, parahippocampal gyrus; IFG, inferior frontal gyrus; Ins, insula; MFG, middle frontal gyrus; SPL, superior parietal lobe; ACC, anterior cingulate cortex; OrbG, orbital gyrus; PreCu, precuneus; PCC, posterior cingulate cortex; LG, lingual gyrus; SMA, supplementary motor area; dACC, dorsal anterior cingulate cortex.
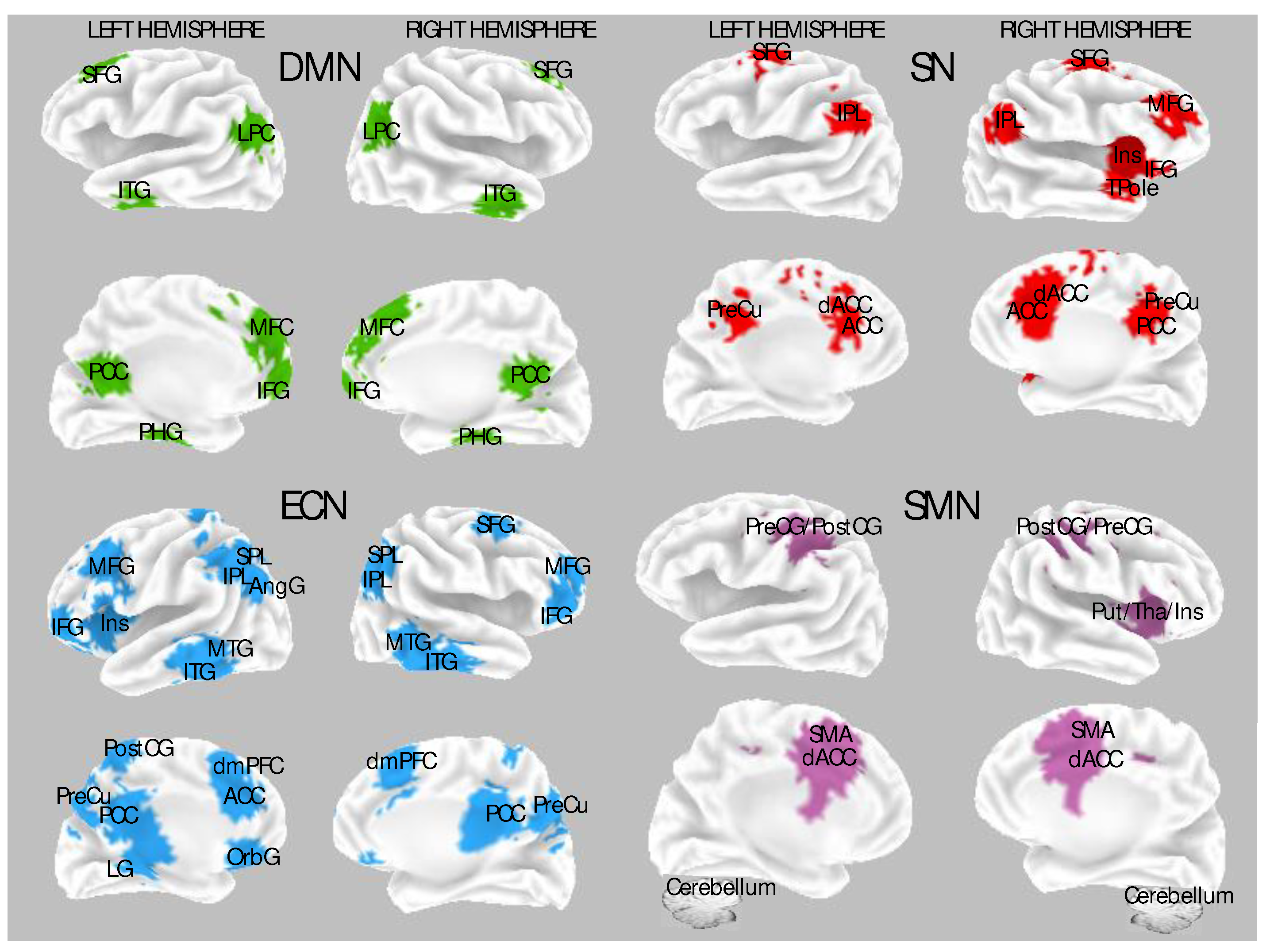
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



