
Submitted:
04 December 2023
Posted:
07 December 2023
You are already at the latest version
Abstract
Antimicrobial peptides (AMPs) are essential components of the innate immune system, present across a spectrum of organisms. Their versatility lies in their ability to effectively inhibit a wide range of pathogens including bacteria, fungi, parasites, and viruses. As the challenge of antibiotic resistance escalates and concerns surrounding antibiotic use grow, AMPs have emerged as promising alternatives. Their applications span diverse domains such as medicine, agriculture, food production, animal husbandry, and aquaculture, primarily due to their efficacy against resistant pathogens, signifying their crucial role in combating infections. Beyond their antimicrobial properties, research illuminates their intricate involvement in the immune response network. These peptides impact crucial processes including cytokine release, chemotaxis, wound healing, angiogenesis and the activation of the adaptive immune system. This comprehensive review delves into the multifaceted realm of AMPs, encompassing their classification, varied applications across industries and notably, their significance in diverse disease conditions like infections, skin diseases, cancer, oral diseases, lung diseases and cardiovascular ailments. Special attention is directed towards current advancements in clinical applications of peptide-based therapeutics, offering invaluable insights into their potential future integration within clinical settings.
Keywords:
Antimicrobial peptides
; Classification
; Disease conditions
; Therapeutic action and Clinical advancements
1. Introduction
Every member of the Animalia kingdom possesses inherent defense mechanisms against various disease-causing microorganisms such as bacteria, fungi, protozoa, and viruses. The human immune system comprises two key types: innate immunity, which is present from birth and provides natural protection against diseases; and acquired immunity, which develops after exposure to disease-causing agents. Notably, innate immunity is inherent in all multicellular organisms, necessitating no prior exposure to foreign pathogens and often being genetically predetermined. Among the integral components of this innate immune system are antimicrobial peptides, protein molecules with relatively small molecular weights that exhibit protective antimicrobial actions against a wide range of microbes including bacteria, fungi, and viruses. These evolutionarily conserved defense molecules are produced by various species such as plants, bacteria, insects and vertebrates and they are notably preserved in mammals [1]. Antimicrobial peptides function as naturally synthesized host organism defenses, akin to endogenous antibiotics, against invading pathogens. The discovery of lysozyme by Alexander Fleming in 1922 catalyzed further research and exploration into antimicrobial peptides. As per the antimicrobial peptide database (APD3), approximately 3,240 AMPs have been identified to date, and ongoing studies suggest the likelihood of discovering many more in the future [2].
Antimicrobial peptides typically manifest as small cationic molecules, ranging from 1-5 kDa in size, carrying an average positive charge of 3.32 and spanning 10-60 amino acid residues. However, there exists another category—negative charge-bearing anionic antimicrobial peptides—which possess notable quantities of glutamic acid and aspartic acid residues within their chemical structure [3,4]. These AMPs are predominantly expressed on primary barriers like the skin and mucosal epithelium of the respiratory, urinary, and gastrointestinal systems (including Paneth cells in the crypts of Lieberkühn in the small intestine), as well as on immune cells. Positioned as the first line of defense against infections, they effectively impede the colonization of pathogens within the host organism's body [5,6]. Antimicrobial peptides are stored in the granules of macrophages and other phagocytic cells, functioning to neutralize engulfed foreign pathogens by degrading their cell membranes and inducing cell death [7,8] . The majority of these peptides are characterized by their amphipathic nature, constituting small molecules with fewer than 50 amino acid units, often referred to as host defense peptides [9]. Presently, over 130 AMPs have been investigated within the human body.
Antimicrobial peptides exhibit diverse functions within the body, with their primary role being defense against invading microorganisms like bacteria, fungi, and enveloped viruses [10,11,12] . The various biological functions of AMPs are largely influenced by factors such as their molecular size, net charge, secondary or three-dimensional structure, and hydrophobicity [13,14]. Aside from their defensive actions, these peptides play crucial roles in wound healing, angiogenesis, neutralizing lipopolysaccharides and endotoxins, exhibiting chemotactic activity, and modulating the immune response [15,16,17,18]. Antimicrobial peptides engage with and penetrate bilayer lipid membranes using various mechanisms such as barrel-stave, carpet, or toroidal pore methods.
Antimicrobial peptides have emerged as alternative options to conventional antibiotics for combating various infectious diseases, showcasing broad-spectrum antimicrobial activity against both gram-positive and gram-negative bacterial species. Their bactericidal action primarily occurs through two mechanisms: direct disruption of the bacterial cell membrane or interference with intracellular biochemical processes. The positive charge of AMPs plays a pivotal role in selectively interacting with the anionic cell membrane of bacteria, while their hydrophobic segments effectively engage with the hydrophobic interior of the bacterial membrane [19,20,21]. Many AMPs not only perturb the synthesis of proteins and nucleic acids within bacteria but also hinder enzymatic reactions and generate reactive oxygen species (ROS) such as NK-18, buferin II, and lactoferricin B, initiating apoptosis in bacterial species [22,23,24].
Antimicrobial peptides play a pivotal role in preventing pathogen proliferation and biofilm development in skin lesions, thereby facilitating wound healing by modulating cellular migration, chemotaxis, angiogenesis, and cytokine release [25,26]. They serve as promising therapeutics for both infectious and non-infectious wounds [27,28]. Within the respiratory system, AMPs produced by neutrophils and airway epithelial cells aid in averting infectious conditions like pneumonia [29] . Moreover, AMPs demonstrate beneficial effects in healing corneal ulcers[30] and preventing peptic ulcers in the gastrointestinal tract while maintaining a balanced and healthy intestinal microbiota [31]. Certain AMPs exhibit potent antimicrobial properties against bacteria causing bone and joint infections without causing harm to osteoblasts [32]. In the oral cavity, AMPs synthesized by gingival epithelium act as the first line of defense against pathogens responsible for oral infections like mucositis and candidiasis [33]. Furthermore, AMPs possess the ability to internalize cells and interact with intracellular components, such as mitochondria, inducing natural cell death, thus holding promise in controlling tumor growth and preventing oncogenesis in various organs [34].
Within the human body, cathelicidins and defensins stand as extensively researched antimicrobial peptide groups. Their roles span diverse disease modulation, including gastrointestinal disorders, skin ailments, respiratory tract infections, oral diseases, cancer, and other epithelial infections. These peptides contribute significantly to bolster our immune system while providing protection against external invasive microorganisms. This review focuses on elucidating their classifications, applications across different fields, and the potential of antimicrobial peptides in preventing various diseases.
2. Classification Antimicrobial peptides (AMPs)
Owing to their natural diversity, Antimicrobial Peptides (AMPs) are classified into different categories based on source, activity, structural characteristics and amino acid rich species (Figure 1).
2.1. Classification of AMPs Based on Sources
AMPs are categorized into different types based on organisms from various phyla or classes, spanning mammals, insects, birds, amphibians, and microorganisms. Additionally, numerous AMPs have been sourced from marine animals found in the ocean.
2.1.1. Antimicrobial Peptides Derived from Mammals
Antimicrobial peptides (AMPs) are abundant across various mammals, including humans, cattle, and vertebrates. Enzymatic hydrolysis of milk and its products serves as a promising source for these peptides, often isolated from components like α-lactalbumin, β-lactoglobulin, lactoferrin, and casein. Among these, lactoferricin B (Lfcin B) stands out as a prominent AMP derived from milk [35]. In humans, AMPs are primarily classified into cathelicidins and defensins. Defensins, categorized by the arrangement of disulfide bonds, encompass α-, β-, and θ-defensins [36]. Human host defense peptides (HDPs), pivotal in guarding against microbial infections, exhibit varied expressions at different life stages. For instance, cathelicidin LL-37 is prevalent in newborns' skin, while human β defensin 2 is predominantly found in older individuals [37]. AMPs are observed in multiple bodily locations including the skin, ears, eyes, mouth, lungs, respiratory tract, intestines, and urethra. Notably, AMPs present in breast milk play a crucial role in reducing morbidity and mortality rates in newborn infants [38]. Beyond their antimicrobial role, AMPs also regulate processes such as apoptosis, wound healing, and immune activity [39].
2.1.2. Antimicrobial Peptides Derived from Amphibians
Amphibians rely on antimicrobial peptides derived from their skin secretions as a defense against pathogenic threats, potentially contributing to the preservation of their population [40]. Notably, amphibians like frogs from genera Xenopus, Silurana, Hymenochirus, and Pserdhymenochirus offer sources for AMPs such as Magainin and others found in their skin secretions [41] . Additionally, the discovery of Cancrin, the first AMP extracted from the sea amphibian Rana cancrivora, underscores the broader spectrum of AMP sources within the amphibian community [42].
2.1.3. Antimicrobial Peptides Derived from Insects
Antimicrobial peptides in insects are synthesized within blood cells and fat bodies, providing them with remarkable adaptability for survival [43]. Among these peptides, cecropin stands out as one of the most well-known families and is found in various insects like the guppy silkworm, Drosophila, and bees. Notably, cecropin A exhibits protective properties against a range of inflammatory diseases and cancers [44]. The number of AMPs varies across different insect species; for instance, invasive harlequin ladybirds (Harmonia axyridis) and black soldier flies (Hermetia illucens) possess approximately 50 AMPs, whereas the pea aphid (Acyrthosiphon pisum) lacks AMPs altogether [45]. Royal jelly produced by bees contains a peptide called jellein, displaying antimicrobial activity against numerous bacteria and fungi. Additionally, its lauric acid-conjugated form effectively regulates the growth of the kalajar parasite Leishmania major [46].
2.1.4. Antimicrobial Peptides Derived from Microorganisms
Antimicrobial peptides find their origins in microorganisms such as bacteria and fungi, with nisin and gramicidin being well-known examples obtained from species like Lactococcus lactis, Bacillus subtilis, and Bacillus brevis [47]. Biological expression systems for AMPs encompass various yeast species like Pichia pastoris, Saccharomyces cerevisiae, bacteria including E. coli and B. subtilis, as well as plants [48]. In addition to these primary sources, an increasing number of AMPs are being isolated from different parts of plants—seeds, stems, and leaves—grouped into categories like defensins, thionins, and snakins [49]. Extracting immune-related AMPs from organisms like Mytilus coruscus provides alternatives to antibiotics; for instance, myticusin beta serves as an immune-related AMP derived from this source [50]. Originating from marine sources, AMPs like pardaxin (GE33) have been used in vaccines, enhancing antitumor capabilities in mice [51].
2.2. Classification of AMPs Based on Activity
The ADP3 database categorizes antimicrobial peptides into 18 distinct groups, where antibacterial peptides constitute 60 percent, antifungal peptides account for 26 percent, antitumor peptides make up 5 percent, antiviral peptides contribute 4 percent, antiparasitic peptides encompass 3 percent, and anti-HIV peptides represent 2 percent.
2.2.1. Antibacterial Peptides
Antibacterial peptides play a crucial role in inhibiting various pathogenic bacteria found in clinical settings, food production, and aquatic products manufacturing. They effectively target bacteria such as Acinetobacter baumannii, VRE, MRSA, S. aureus, L. monocytogenes, E. coli, Salmonella, and Vibrio parahaemolyticus. Peptides like nisin, defensins, and cecropins exhibit the ability to halt the growth of both gram-positive and gram-negative bacteria. Specific antimicrobial peptides like P5 and P9 demonstrate efficacy against MRSA while also reducing cellular toxicity levels [52].
2.2.2. Antifungal Peptides
Antifungal peptides serve as a defense against various fungi and enhance the body's resilience against organisms like Aspergillus, Candida albicans, yeast, filamentous fungi, and molds in clinical, food, and agricultural contexts. Peptides such as ranatuerin, cecropins, aurein, and synthetic antifungal variants are employed to hinder the growth of Candida albicans and Aspergillus flavus, known for producing aflatoxins [53]. Notably, approximately 37 antifungal peptides have been extracted from Lactobacillus plantarum TE10, and their combination has shown a reduction in the sporulation of Aspergillus flavus in maize [54].
2.2.3. Antiviral Peptides
Antiviral peptides exhibit diverse mechanisms by hindering virus attachment, cell membrane fusion, and disrupting viral membranes, consequently impeding viral replication [55]. Within the antiviral peptide category, there exists a subgroup focused on combating HIV. Commonly used and commercialized anti-HIV peptides encompass alpha and beta defensins, LL-37, gramicidin D, caerin 1, maximin 3, maginin 2, dermaseptin-S1, dermaseptin-S4, siamycin-I, siamycin-II, RP71955, and FuzeonTM [56]. Specific AMPs like epinecidin-1 demonstrate inhibition against the virus responsible for foot-and-mouth disease [57]. Additionally, swine intestinal AMP (SIAMP)-IBV has shown effectiveness in reducing the occurrence of infectious bronchitis virus in chicken embryos [58]. Fusion inhibitor peptides like peptide HR2, its lipid binding counterpart, temporin, and rhesus theta defensin 1 collectively act against SARS-CoV and MERS-[59,60].
2.2.4. Anti-parasitic Peptides
Anti-parasitic peptides play a crucial role in thwarting the growth and reproduction of parasites responsible for various diseases such as kalazar and malaria, impacting both humans and other organisms [61,62]. Peptides like temporins-SHD and cathelicidin impede biological processes, effectively preventing the invasion of parasitic organisms [63]. A synthetic marine AMP, epinecidin 1, disrupts the membrane of Trichomonas vaginalis [64]. Furthermore, jellein from royal jelly, along with the amino acid AMP KDEL (lysine, aspartic acid, glutamic acid, and leucine), inhibits Trichomonas vaginalis and the Leishmania donovani parasite [65] . Notably, the antiparasitic action of cyanobacterial AMPs relies on specific protein targets, differing from those of higher eukaryotic AMPs.
2.2.5. Anticancer Peptides
Anticancer peptides display multifaceted functions including the ability to eliminate tumour cells through immune or dendritic cell activation, trigger apoptosis in cancerous cells, inhibit angiogenesis, impede metastasis, and activate regulatory proteins that disrupt the transcription and translation of tumour cells [66,67]. Peptides like tritripticin, inolicidin, and puroindoline A showcase anticancer properties and contribute to countering carcinogenesis [68]. These Anticancer Peptides (ACPs) operate by balancing net charge and hydrophobicity, thereby harnessing their anticancer benefits.
Top of Form
2.3. Classification of AMPs Based on Amino Acid Rich Species
Antimicrobial peptides fall into four distinct categories:
(a) Proline-rich peptides function by targeting the bacterial inner membrane using the SbmA transporter. They gain entry into the cytoplasm, affecting ribosomes and disrupting the termination process of translation. This disturbance ultimately hampers protein synthesis in bacteria [69].
(b) Tryptophan and arginine-rich peptides impact the cell membrane by disrupting the lipid bilayer interface. They influence peptide charge and hydrogen bond interactions. Examples like tritripticin and indolicidin showcase effectiveness against gram-negative bacteria such as E. coli and Pseudomonas aeruginosa, as well as gram-positive Staphylococcus aureus.
(c) Histidine-rich peptides like HV2 enhance cell membrane permeation, leading to rupture and subsequent bacterial cell membrane death. They also exhibit anti-inflammatory properties by inhibiting the synthesis of tumour necrosis factor [70].
(d) Glycine-rich peptides like attacins and diptericins demonstrate efficacy against gram-negative bacteria, effectively eliminating them [71].
2.4. Classification of AMPs Based on Antimicrobial Peptide Structure
Antimicrobial peptides are grouped based on their structures into four classes: linear α-helical, β-sheet, linear extension, and peptides that combine both α-helix and β-sheet structures [72]. Additionally, researchers have examined various cyclic peptides and antimicrobial peptides with more intricate configurations, including lasso peptides and thioether bridged structures [73].
3. Application of Antimicrobial Peptides in Different Fields
Antimicrobial peptides (AMPs) have gained extensive applications across diverse fields like medicine, food, agriculture, animal husbandry, aquaculture etc (Figure 2). In medicine, AMPs play roles in addressing infections, wound healing, skin diseases, gastrointestinal issues, dental problems, surgical injuries, cancer treatment, respiratory tract disorders, and ophthalmology. Notably, the FDA has approved three AMPs—gramicidin, daptomycin, and colistin—for specific medical uses. Innovations include pheromone-labelled and locally triggered AMPs that exhibit improved targeting mechanisms.
In the food industry, AMPs effectively inhibit the growth of bacteria, fungi, and other microorganisms, thereby preventing food spoilage. Their resilience against acids, alkalis, and temperature fluctuations positions them as alternatives to conventional food preservatives. Examples like nisin, plylysine, and Lactic acid bacteria in dairy products showcase this efficacy. Specific AMPs such as pedocin combat Listeria monocytogenes, a common cause of meat spoilage, while enterocin aids in preserving cider, fruit and vegetable juices, and soy milk.
Additionally, AMPs prove valuable in agriculture by inhibiting plant pathogens like Aspergillus flavus in corn and peanuts, Penicillium digitatum, and Geotrichum citriaurantii in citrus, and Botrytis cinerea in strawberries. In animal farming, AMPs like SIAMP and NKLPs help prevent various diseases and infections in poultry, swine, ruminants, and fish farming. They find applications in coating implants, contact lenses, and biomedical devices. Moreover, AMPs play a role in biosensors' detection and are utilized in water purifiers [74].
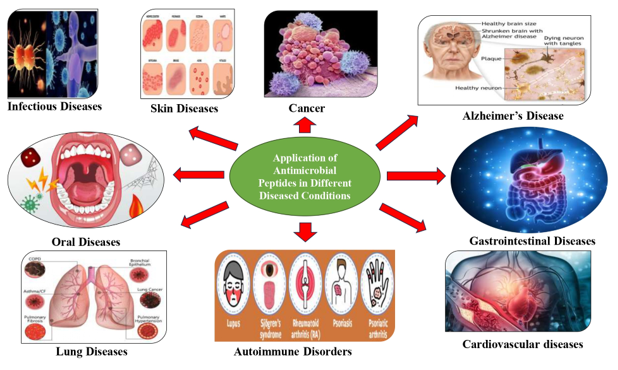
4. Application of Antimicrobial Peptides in Different Diseased Conditions
Antimicrobial peptides prove valuable in treating and preventing numerous diseases in humans and other organisms. They serve as an alternative to antibiotics in combating bacterial, viral, and fungal infections. These versatile compounds are utilized in addressing a wide array of infectious disorders, including skin infections, AIDS, lung diseases, gastrointestinal tract disturbances, oral and dental diseases, cancers, iron disorders, autoimmune diseases, and Alzheimer’s disease, both in natural and synthetic forms within clinical settings.
4.1. Role of AMPs in Infectious Diseases
Most antimicrobial peptides (AMPs) exhibit a unique ability to eliminate invasive microorganisms while causing minimal harm to host organisms. This effectiveness arises from the distinction in cell membrane structure between prokaryotes and eukaryotes. The outer membrane of gram-positive bacteria contains teichoic and lipoteichoic acid, while gram-negative bacteria possess lipopolysaccharide and phosphatidyl-glycerol, resulting in a negative charge. Positively charged AMPs interact electrostatically with this negative charge, affecting the surface structure of these bacteria. Notably, there is no interaction between AMPs and mammalian cell membranes due to the neutral net charge of mammalian membranes [75]. AMPs don't just impact the bacterial cell membrane; they also penetrate the cytoplasm, leading to alterations in numerous cellular processes such as cell wall formation, protein synthesis, nucleic acid synthesis, and enzyme activity [76].
AMPs have demonstrated a synergistic effect when combined with traditional antibiotics, enhancing the prevention of microbial infection [77]. Studies involving magainin II and cecropin A, administered either independently or with rifampicin against multi-drug-resistant Pseudomonas aeruginosa, both in vivo and in vitro, have shown significant reductions in bacterial multiplication, LPS and TNF-α secretion, as well as infection rates and mortality [78]. P5 peptide's collaboration with isepamicin in cholelithiatic patients effectively combats Pseudomonas aeruginosa [79].
AMPs also target bacterial biofilms linked to dental plaque, endocarditis, lung infections, and infections related to medical devices. For instance, Kappcin, a non-glycosylated κ-casein, has demonstrated a substantial reduction in Streptococcus mutans biofilms in the presence of zinc chloride [80]. Melimine, a non-hemolytic hybrid peptide derived from melittin and protamine, diminishes bacterial adhesion to covalently linked contact lenses [81]. Citropin 1.1, derived from the green tree frog Litoria citropa, in combination with rifampin and minocycline, exhibits increased anti-biofilm activity against S. aureus [82].
A multitude of AMPs, including magainins, temporins, defensins, cathelicidins, bacteriocins, bombinins, protonectins, abaecin, among others, effectively hinder the growth and formation of biofilms associated with various gram-positive and gram-negative bacterial infectious diseases. In conjunction with antifungal drugs like fluconazole, caspofungin, amphotericin B, clotrimazole, and flucytosine, antifungal peptides combat various fungal infections caused by Candida spp., Aspergillus, Histoplasma, Blastomyces, and Coccidioides [83].
4.2. Role of AMPs in Skin Diseases
The skin acts as the initial defense line against various microorganisms in organisms, with the epidermis forming a physical barrier providing immediate protection against infections. Within the skin, low molecular weight antimicrobial peptides (AMPs) such as α- and β-defensins, cathelicidins, S100 proteins, and ribonucleases are present in cells like mast cells, keratinocytes, eccrine and sebaceous glands, and phagocytic cells [84,85]. Several AMPs play roles in chronic inflammatory skin conditions like atopic dermatitis and psoriasis vulgaris [86]. Defensins, including α-, β-, and θ-defensins, are small cystine-rich molecules with a β-hairpin structure stabilized by three conserved disulfide bonds [87,88]. Human α-defensins -1 to -4, known as human neutrophil peptides (HNP1 to HNP4), found in neutrophils, and enteric peptides like HD-5 and HD-6 expressed in Paneth cells are among these [89].
Human defensins exhibit antibacterial effects against various gram-negative bacteria like Escherichia coli, Candida albicans, and Pseudomonas aeruginosa. For instance, hBD-3 targets multidrug-resistant gram-positive bacteria such as Staphylococcus aureus, Streptococcus pyogenes, and vancomycin-resistant Enterococcus faecium .hBD-3 contributes to wound healing, dendritic cell activation, migration, and polarization [90]. Another peptide, hBD-2, induced by the human papillomavirus, combats gram-negative bacteria like E. coli, E. faecalis, Propionibacterium acnes, and P. aeruginosa while aiding in wound repair. Dermcidin exhibits antibacterial activity against various pathogens such as E. coli, S. aureus, C. albicans, and E. faecalis [91]. RNase 7 displays antimicrobial activity against both gram-negative and gram-positive bacteria like E. coli, S. aureus, E. faecium, P. aeruginosa, MRSA, and yeast C. albicans [92].
Human cathelicidin LL-37, a versatile antimicrobial peptide, disrupts bacterial cell membranes, viral envelopes, and fungal pathogens like yeast. LL-37 also exhibits pro-apoptotic activity in airway epithelial cells and promotes angiogenesis, contributing to the wound healing process [93]. Psoriasin (S100A7), a Ca2+ binding protein found in psoriatic lesions characterized by keratinocyte differentiation and tumor formation, demonstrates antibacterial activity against E. coli and chemotactic activity for CD4+ T lymphocytes and neutrophils [94,95]. Calprotectin provides protection against E. coli, S. aureus, Staphylococcus epidermis, Klebsiella spp., and Candida albicans [96].
4.3. Role of AMPs in Oral Diseases
Different oral diseases, including dental caries, periodontal issues, mucosal infections, and oral cancer, stem from various causes and microbes. Dental caries result from localized demineralization due to acid accumulation during bacterial fermentation of leftover food particles [97]. Secondary conditions like pulp and periapical diseases often involve bacteria like Enterococcus faecalis, which forms biofilms in root canals leading to apical periodontitis [98]. Natural and synthetic antimicrobial peptides (AMPs) play roles in inhibiting pathogenic microbes, aiding enamel remineralization, and pulp tissue healing. They also act as biomarkers indicating caries risk. Human β-defensins 1, 2, and 3 expressed in pulp and odontoblasts combat Streptococcus mutans and E. faecalis by preventing their association with host cells [99,100]. Histatin-5 and cathelicidin LL37 also exhibit antibacterial activities against Streptococcus mutans [101,102]. Synthetic AMPs derived from hBD-3, such as HBD3-C15 and D1-23, inhibit Streptococcus gordonii and S. mutans, preventing their biofilm formation [103,104]. Peptides like VSL2, DGL13K, PR39, DJK-5, IDR-1002, and C16G2 show efficacy against harmful effects of E. faecalis and S. mutans [71,105,106].
Periodontal diseases often involve bacteria like Porphyromonas gingivalis, Fusobacterium nucleatum, and Prevotella intermedia. hBD-2 displays inhibitory action against these pathogens, curbing periodontal inflammation similar to minocycline at comparable concentrations [107]. Cathelicidin LL37 helps suppress inflammation caused by P. gingivalis [108]. AMPs derived from humans, cattle, sheep, and pigs—such as P-113, hBD3-C15, BMAP28, PMAP23, SMAP28, SMAP29, and Nal-P-113—show broad-spectrum antimicrobial effects [109].
Oral mucosal diseases like oral candidiasis often stem from autoimmune inflammatory conditions and fungal attacks. AMPs like Histatin 5, HNP-1, hBD-2, hBD-3, LL37, and Lactoferricin B safeguard the oral environment against Candida albicans fungal infections. Salivary LL37 displays antiviral activity against Kaposi's sarcoma-associated herpes virus [110]. hBD-1 is effective in suppressing oral squamous cell cancer and benign tumors in salivary glands [111].
4.4. Role of AMPs in Lung Diseases
The respiratory pathway, being directly exposed to external air during inhalation, remains consistently vulnerable to microbial exposure. The innate immune system of the lungs comprises epithelial and myeloid cells like alveolar epithelial cells, neutrophils, and dendritic cells, which actively combat microbial colonization. These cells express primary antimicrobial peptides (AMPs) of the respiratory tract, particularly defensins and cathelicidins. Epithelial cells, monocytes, macrophages, and dendritic cells express hBD-1 and hBD-2 [112,113]. During microbial infections, hBD-2, hBD-3, and hBD-4 are significantly induced and abundantly expressed [114]. Bacterial exposure, such as to Pseudomonas aeruginosa, Haemophilus influenza, Legionella pneumophila, and Streptococcus pneumoniae, activates hBD-2 and hBD-3 in respiratory epithelium [115]. Various mycobacterial strains facilitate the expression of hBD-2 [116]. LL37 demonstrates antimicrobial activity against both gram-negative and gram-positive bacteria. α-defensins HNP-1 and HNP-2, along with LL-37, exhibit heightened levels in chronic obstructive pulmonary disease [117]. The antimicrobial efficacy of defensins is sensitive to salt, and excessive mucus production in the respiratory airways can affect the potency of LL-37 [118].
Top of Form
4.5. Role of AMPs in Gastrointestinal Diseases
Antimicrobial peptides function as internal antibiotics, combating the health effects instigated by fungi, protozoa, viruses, and bacteria within the gastrointestinal (GI) tract from the oral cavity to the anal region. Helicobacter pylori-induced peptic ulcers trigger the production of hBD-2 and cathelicidin LL-37 by the GI tract's epithelial cells [119,120]. Chemokines CCL14 and CCL15, expressed in the intestinal lining, display potent antibacterial actions [121].
In liver, gallbladder, and bile duct issues like hepatolithiasis or biliary obstruction, cholangitis activates the induction and dominant expression of AMPs hBD-2 and hBD-1 [122]. α-defensins HD-5 and HD-6 are notably present in the human small intestine [123]. HD-5 expression decreases during Salmonella typhimurium infections [124]. Proteins like ATG16L1 aid in invading bacteria degradation, while X-box binding protein 1 poses a risk factor for Crohn's disease and ulcerative colitis [125]. Bacteria such as E. coli and Campylobacter jejuni, along with cytokines, stimulate hBD-2 [126]. Cathelicidin impedes the growth and colonization of Citrobacter rodentium [127]. Polymorphonuclear cells secrete lysozyme in high concentrations in ulcerative colitis [128]. Lactoferritin in milk binds to the lipopolysaccharide layer of the bacterial cell wall, rupturing the cell, depriving microbial pathogens of iron needed for their growth, and forming toxic peroxide inside the cell, ultimately killing it [129]. Hepcidin, significantly elevated in ulcerative colitis and Crohn’s disease, inhibits the growth of Salmonella and Mycobacteria by restricting iron availability to them, showcasing antimicrobial properties [130]. Coprisin from the Korean dung beetle helps in curbing the mucosal inflammation caused by C. difficile. Synthetic glycopeptides like telavancin and dalbavancin exert antibacterial action against C. difficile and colitis associated with its contamination [131].
4.6. Role of AMPs in Autoimmune Disorders
Autoimmune diseases like psoriasis, type 1 diabetes, and rheumatoid arthritis often exhibit elevated levels of various antimicrobial peptides. In psoriasis, LL-37 directly activates T cells as an autoantigen [132]. Human α-defensin and LL-37 are significantly expressed in the synovial fluid of joints under the influence of TNF-α and interleukin 6. The macrocyclic peptide RTD-1 aids in alleviating painful symptoms and joint discomfort in arthritis patients [133]. Mouse β-defensin-14 diminishes inflammation in the central nervous system (CNS) and curbs cytokines and cytotoxic T cells, the causative agents in encephalomyelitis.
4.7. Role of AMPs in Alzheimer’s Disease
Alzheimer's disease can stem from various causes, among which microbial infections in the central nervous system leading to sepsis and meningitis are considered significant factors. Microorganisms like Chlamydia, Helicobacter pylori, Borrelia spirochetes, herpes simplex virus, and human immuno-deficiency virus have been linked to infections affecting both outer and inner brain regions [133,134]. Some microorganisms like E. coli, herpes simplex virus, Cryptococcus neoformans, and Chlamydia pneumoniae contribute to the formation of amyloid plaques, a hallmark of Alzheimer's disease [135]. Certain antimicrobial peptides, when in their β-sheet configuration, disrupt cell membranes by aggregating on the membrane surface, causing leakage of cell contents [136]. AMPs exhibit immunostimulatory effects by neutralizing lipopolysaccharides, boosting TNF-α and interleukin-8 levels, and aiding in brain tissue wound healing [137]. The Aβ1-42 peptide, when self-aggregated into β-sheet structures, can act as pore-forming agents with antimicrobial properties [138].
4.8. Role of AMPs in Cardiovascular diseases
Cardiovascular diseases, particularly characterized by arterial wall thickening called atherosclerosis, arise due to various risk factors that trigger inflammation and oxidative stress within the vessel walls. The progression of atherosclerosis and coronary heart disease (CHD) often correlates with alterations in the expression of numerous proteins. Certain antimicrobial peptides, like CRAMP and cathelicidin LL-37, serve as biomarkers linked to the risk of cardiovascular disease. LL-37 levels have been observed to decrease at thrombosis sites. CRAMP plays a role in activating Akt and ERK1/2, facilitating FoxO3a phosphorylation and nuclear export. This action aids in safeguarding the heart by reducing cardiomyocyte apoptosis, thereby potentially decreasing the risk of myocardial infarction. Notably, the serum levels of LL-37 appear to be lower in individuals affected by cardiovascular conditions [139].
4.9. Role of AMPs in Cancer
Anticancer peptides differentiate between malignant and normal cells due to variations in cell membrane composition. Malignant cells possess a higher negative charge on their membranes, owing to the presence of anionic molecules like sialylated gangliosides, phosphatidylserine (PS), heparin sulfate, and O-glycosylated mucins. Anticancer peptides selectively target tumor cell membranes, causing pore formation, rapid disruption, altering ion channels, and enhancing permeation [140].
Various antimicrobial peptides exhibit anticancer properties, such as Na-D1, Polybia-MP1, Parasporin, m2386, and LTX-315. Na-D1, derived from ornamental tobacco Nicotiana alata, is a cationic peptide that interacts with phosphatidylinositol 4,5-bisphosphate on the plasma membrane of cancer cells, leading to their destruction. MP1 modifies tumor cell membranes by creating pores. These AMPs internalize within cells and engage with intracellular organelles like mitochondria, triggering programmed cell death. Peptide m2386, extracted from the lactic acid bacterium Lactobacillus casei ATCC334, induces tumor cell apoptosis. Parasporin, derived from Bacillus thuringiensis, and peptides from hexokinase-II expedite cell apoptosis as well [141].

Table 1.
Expression level and proposed functions of antimicrobial peptides/proteins in various human inflammatory diseases.
Table 1.
Expression level and proposed functions of antimicrobial peptides/proteins in various human inflammatory diseases.
| Disease state | Peptides | Expression levels and potential functions | References |
|---|---|---|---|
| Skin inflammatory diseases | |||
| Psoriasis | LL-37, defensins | Overexpressed, absence of S. aureus | [142] |
| Atopic dermatitis | LL-37, defensins | Downregulated, presence of S. aureus | [142] |
| Lupus, erythrematous, and contact dermatitis | LL-37 | Increased | [143] |
| Acne vulgaris | MX-594 AN | Inhibits P. acne | [144] |
| Granulysin | Kills P. acne, anti-inflammatory action | [145] | |
| Respiratory diseases | |||
| Cystic fibrosis | LL-37, β-defensins |
Reduced antimicrobial activity due to salt accumulation | [146] |
| Periodontal disease | Defensins | Reduced in saliva of patients with oral candidiasis | [147] |
| LL-37 | Absent in patients with congenital neutropenia | [148] | |
| Histatin 5 | Protects periodontium from bacterial infection and prevents biofilm formation | [149,150] | |
| Inflammatory bowel disease | |||
| Crohn’s disease | HD5 and HD6 | Deficient expression in Paneth cells | [151] |
| HD5 and HD6 | Reduced in CD patients with Nod2 mutation | [151] | |
| LL-37 | Expression is altered | [151] | |
| Ulcerative colitis | HD5, 6; hBD2–4 | Upregulated in patients with UC | [152] |
| Cancer Magainin | II Toxic | effect against canc er cell lines melanoma, breast and lung cancer, lymphoma, and leukemia | [153,154] |
| Insect cecropins | Lyse tumour cells | [155] | |
| Bovine lactoferrin | Inhibits lung and liver metastasis of murine melanomas and lymphomas and cytotoxic toward neuroblastoma cells | [156,157] | |
| Atherosclerosis | Defensins | Involved in lipoprotein metabolism, exhibit anti-fibrolytic activity and regulate angiogenesis | [158,159,160] |
| LL-37 | Increased expression in human lesions | [161] | |
| Inflammatory articular joints | hBD-3, LL-37 | Upregulated in osteoarthritis | [162] |
5. Clinical development of antimicrobial peptides
The current focus lies in leveraging the knowledge of antimicrobial peptides (AMPs) to craft and advance beneficial drugs. Biotechnology firms have embarked on the development of new peptide-based compounds, showcasing distinct and potentially enhanced resistance profiles compared to earlier antibiotics. A recent comprehensive review detailing AMP-based therapeutics showcased various peptides in preclinical or clinical trials. Here are some noteworthy examples:
Plectasin (Novozymes), a defensin peptide, exhibits potent microbicidal activity against antibiotic-resistant bacteria linked to diseases like pneumonia [163]. Unlike other AMPs in clinical trials, plectasin displays tolerance at high doses and effectiveness in treating systemic infections. Its development as a therapeutic agent is progressing into the preclinical phase, with expectations for clinical approval within 8–10 years.
P-113 (Dermegen), a 12-amino acid fragment derived from histatin 5, demonstrates anti-candidal activity comparable to its parent form. Currently in phase I/II clinical trials, it's used as a mouth rinse for treating plaques and gingivitis. Human model data highlight P-113's efficacy against gingivitis and plaques [164]. MBI-226 (Migenix), an Idolicidin analogue in phase III clinical trials, aims to treat catheter-related bloodstream infections [165]. MX-594AN (Migenix), an antimicrobial cationic peptide, is developed as a topical treatment for mild to moderate acne vulgaris. Clinical trials demonstrate its efficacy against various acne lesions, and it's also being tested for topical treatment of rosacea. Phase II trials are projected for completion in 2007.PG-1 protegrin (Intrabiotics) is in phase III clinical trials for treating peritoneal infections caused by P. aeruginosa, S. aureus, and methicillin-resistant S. aureus. rBPI-21 (Xoma), a recombinant form of human bactericidal/permeability-increasing (BPI) protein, is in phase II/III clinical trials for treating meningococcaemia and Crohn’s disease. Heliomycin (EntoMed), a peptide derived from insects with antifungal activity, is undergoing preclinical trials.
While numerous clinical trials have underscored the broad therapeutic potential of AMP-based drugs, these agents are still in early stages of technological refinement, with several challenges ahead to overcome.
6. Conclusion and Future perspectives
Pharmaceutical companies are eagerly exploring the development of novel peptides as a promising new avenue for medications. However, these antimicrobial peptides (AMPs) face inherent challenges due to their peptidic nature. These hurdles include the high costs associated with manufacturing, their short half-lives within the body, the potential loss of activity under physiological conditions, application-related issues, risks of unwanted systemic reactions like aggregation or immunoreactivity, and the possibility of disrupting normal bacterial flora. Moreover, several unresolved matters remain, such as the absence of standardized assessment techniques, incomplete understanding of molecular regulation mechanisms, difficulties in targeted delivery to specific sites, concerns regarding tolerance and toxicity, and the necessity for ideal peptide characteristics such as high tolerance, resistance to degradation, and suitability for various administration methods.
Despite these obstacles, ongoing research aims to shed light on the broader roles of AMPs beyond their antimicrobial activities. This exploration includes understanding their functions in complex diseases, which could potentially position these peptides as prototypes for innovative drugs, sensors, or biomarkers for early disease detection and prevention. Comprehensive knowledge of AMPs may eventually enable researchers to create drugs that modulate the expression of specific AMPs, effectively ameliorating particular disease conditions. Gene therapy involving AMPs could revolutionize treatments for various inflammatory and infectious diseases if strategies for efficient gene delivery are optimized. However, these aspirations face significant challenges related to the properties of AMPs that need to be addressed, such as their cationic nature, which may lead to interactions with anionic components of host cells. An ideal peptide drug would be one that is well-tolerated at high doses, resistant to degradation, and suitable for administration via various routes, including topical and intravenous methods. In conclusion, focused research on these issues is poised to provide crucial insights into the roles AMPs play in complex diseases, potentially laying the groundwork for their effective use in innovative drug development, disease management, and early disease detection strategies.
Author Contributions
First and corresponding author did planning, designing, writing and editing work of manuscript, another first author contributed manuscript content. second author did total editing of whole manuscript.
Conflicts of Interest
The authors report no conflict of interest. The authors are responsible for the content and writing of the paper.
References
- Lehrer, R.I.; Ganz, T. Antimicrobial peptides in mammalian and insect host defence. Current Opinion in Immunology 1999, 11, 23–27. [Google Scholar] [CrossRef]
- Huan, Y.; Kong, Q.; Mou, H.; Yi, H. Antimicrobial peptides: classification, design, application and research progress in multiple fields. Frontiers in microbiology 2020, 11, 2559. [Google Scholar] [CrossRef] [PubMed]
- Malkoski, M.; Dashper, S.G.; O'Brien-Simpson, N.M.; Talbo, G.H.; Macris, M.; Cross, K.J.; Reynolds, E.C. Kappacin, a novel antibacterial peptide from bovine milk. Antimicrobial agents and chemotherapy 2001, 45, 2309–2315. [Google Scholar] [CrossRef]
- Schittek, B.; Hipfel, R.; Sauer, B.; Bauer, J.; Kalbacher, H.; Stevanovic, S.; Schirle, M.; Schroeder, K.; Blin, N.; Meier, F.; Rassner, G. Dermcidin: a novel human antibiotic peptide secreted by sweat glands. Nature immunology 2001, 2, 1133–1137. [Google Scholar] [CrossRef]
- Bulet, P.; Stocklin, R.; Menin, L. Antimicrobial peptides from invertebrates to vertebrates. Immunological Reviews 2004, 198, 169–184. [Google Scholar] [CrossRef]
- Wang, G. Human antimicrobial peptides and proteins. Pharmaceuticals 2014, 7, 545–594. [Google Scholar] [CrossRef] [PubMed]
- Ganz, T. Defensins and host defense. Science 1999, 286, 420–421. [Google Scholar] [CrossRef]
- Zasloff, M. Antimicrobial peptides of multicellular organisms. Nature 2002, 415, 389–395. [Google Scholar] [CrossRef]
- Lai, A.C.; Chen, F.Z. Comparison of a new Eulerian model with a modified Lagrangian approach for particle distribution and deposition indoors. Atmospheric Environment. 2007, 41, 5249–5256. [Google Scholar] [CrossRef]
- McKelvey, J.A.; Yang, M.; Jiang, Y.; Zhang, S. Salmonella enterica serovar enteritidis antimicrobial peptide resistance genes aid in defense against chicken innate immunity, fecal shedding, and egg deposition. Infection and Immunity 2014, 82, 5185–5202. [Google Scholar] [CrossRef] [PubMed]
- Yang, M.; Zhang, C.; Zhang, X.; Zhang, M.Z.; Rottinghaus, G.E.; Zhang, S. Structure-function analysis of Avian beta-defensin-6 and beta-defensin- 12: role of charge and disulfide bridges. BMC Microbiol 2016, 16, 210. [Google Scholar] [CrossRef]
- Yang, M.; Zhang, C.; Zhang, M.Z.; Zhang, S. Beta-defensin derived cationic antimicrobial peptides with potent killing activity against gram negative and gram positive bacteria. BMC Microbiology 2018, 18, 54. [Google Scholar] [CrossRef]
- Wimley, W.C. Describing the mechanism of antimicrobial peptide action with the interfacial activity model. ACS Chemical Biology 2010, 5, 905–917. [Google Scholar] [CrossRef]
- Yang, M.; Zhang, C.; Zhang, M.Z.; Zhang, S. (2017). Novel synthetic analogues of avian beta-defensin-12: the role of charge, hydrophobicity, and disulfide bridges in biological functions. BMC Microbiology 2017, 17, 43. [Google Scholar] [CrossRef]
- Yang, D.; Biragyn, A.; Kwak, L.W.; Oppenheim, J.J. Mammalian defensins in immunity: more than just microbicidal. Trends in Immunololgy 2002, 23, 291–296. [Google Scholar] [CrossRef] [PubMed]
- Zaiou, M. Multifunctional antimicrobial peptides: therapeutic targets in several human diseases. Journal of Molecular Medicines 2007, 85, 317–329. [Google Scholar] [CrossRef]
- Yang, M. Avian β-Defensins as Antimicrobial and Immunomodulatory Agents 2020. Available online at: https://hdl.handle.net/10355/68953.
- Zhang, C.; Yang, M.; Ericsson, A.C. Antimicrobial peptides: potential application in liver cancer. Frontiers in Microbiology 2019, 10, 1257. [Google Scholar] [CrossRef]
- Shai, Y. Mode of action of membrane active antimicrobial peptides. Peptide Science 2002, 66, 236–248. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Z.; Vasil, A.I.; Hale, J.D.; Hancock, R.W.E.; Vasil, M.L.; Hodges, R.S. Effects of net charge and the number of positively charged residues on the biological activity of amphipathic α-helical cationic antimicrobial peptides. Journal of Peptide Science 2002, 90, 369–383. [Google Scholar] [CrossRef]
- Huang, Y.; Huang, J.; Chen, Y. Alpha-helical cationic antimicrobial peptides: relationships of structure and function. Protein Cell 2010, 1, 143–152. [Google Scholar] [CrossRef] [PubMed]
- Park, C.B.; Kim, H.S.; Kim, S.C. Mechanism of action of the antimicrobial peptide buforin II: buforin II kills microorganisms by penetrating the cell membrane and inhibiting cellular functions, Biochemical Biophysical Research Communication 1998, 244, 253–257. [Google Scholar] [CrossRef] [PubMed]
- Yan, J.; Wang, K.; Dang, W.; Chen, R.; Xie, J.; Zhang, B. Two hits are better than one: membrane-active and DNA binding-related double-action mechanism of NK-18, a novel antimicrobial peptide derived from mammalian NK-Lysin. Antimicrobial Agents and Chemotherapy 2013, 57, 220–228. [Google Scholar] [CrossRef] [PubMed]
- Lee, B.; Hwang, J.S.; Lee, D.G. Antibacterial action of lactoferricin B like peptide against Escherichia coli: reactive oxygen species-induced apoptosis-like death. Journal of applied microbiology 2020, 129, 287–295. [Google Scholar] [CrossRef]
- Ramos, R.; Silva, J.P.; Rodrigues, A.C.; Costa, R.; Guardão, L.; Schmitt, F. Wound healing activity of the human antimicrobial peptide LL37. Peptides 2011, 32, 1469–1476. [Google Scholar] [CrossRef]
- Mangoni, M.L.; McDermott, A.M.; Zasloff, M. Antimicrobial peptides and wound healing: biological and therapeutic considerations. Experimental Dermatology 2016, 25, 167–173. [Google Scholar] [CrossRef] [PubMed]
- Sørensen, O.E. Antimicrobial peptides in cutaneous wound healing. Antimicrobial Peptides 2016, 1–15. [Google Scholar] [CrossRef]
- Pfalzgraff, A.; Brandenburg, K.; Weindl, G. Antimicrobial peptides and their therapeutic potential for bacterial skin infections and wounds. Frontiers in Pharmacology 2018, 9, 281. [Google Scholar] [CrossRef]
- Majewski, K.; Kozłowska, E.; Żelechowska, E.P.; Brzezińska-Błaszczyk, A. Serum concentrations of antimicrobial peptide cathelicidin LL-37 in patients with bacterial lung infections. Central Europian Journal of Immunology 2018, 43, 453–457. [Google Scholar] [CrossRef]
- Griffith, G.L.; Kasus-Jacobi, A.; Pereira, H.A. Bioactive antimicrobial peptides as therapeutics for corneal wounds and infections. Adv Wound Care (New Rochelle) 2017, 6, 175–190. [Google Scholar] [CrossRef]
- Rivas-Santiago, B.; Schwander, S.K.; Sarabia, C.; Diamond, G.; Klein-Patel, M.E.; Hernandez-Pando, R.; Ellner, J.J.; Sada, E. Human {beta}-defensin 2 is expressed and associated with Mycobacterium tuberculosis during infection of human alveolar epithelial cells. Infection and Immunity 2005, 73, 4505–4511. [Google Scholar] [CrossRef]
- Bormann, N.; Koliszak, A.; Kasper, S.; Schoen, L.; Hilpert, K.; Volkmer, R. A short artificial antimicrobial peptide shows potential to prevent or treat bone infections. International Journal of Scientific Reports 2017, 7, 1506. [Google Scholar] [CrossRef] [PubMed]
- Donnelly, J.P.; Bellm, L.A.; Epstein, J.B.; Sonis, S.T.; Symonds, R.P. Antimicrobial therapy to prevent or treat oral mucositis. The Lancet Infectious Diseases 2003, 3, 405–412. [Google Scholar] [CrossRef]
- Deslouches, B.; Di, Y.P. Antimicrobial peptides with selective antitumor mechanisms: prospect for anticancer applications. OncoTargets and Therapy 2017, 8, 46635. [Google Scholar] [CrossRef] [PubMed]
- Sibel Akalın, A. Dairy-derived antimicrobial peptides: action mechanisms,pharmaceutical uses and production proposals. Trends in Food Science and Technology 2014, 36, 79–95. [Google Scholar] [CrossRef]
- Reddy, K.V.R.; Yedery, R.D.; Aranha, C. Antimicrobial peptides:premises and promises. International Journal of Antimicrobial Agents 2004, 24, 536–547. [Google Scholar] [CrossRef] [PubMed]
- Gsehwandtner, M.; Zhong, S.; Tschachler, A.; Miltz, V.; Karner, S.; Elbe-Burger, A. Fetal human keratinocytes produce large amounts of antimicrobial peptides: involvement of histone-methylation process. Journal of Investigative Dermatology 2014, 134, 2192–2201. [Google Scholar] [CrossRef] [PubMed]
- Field, C.J. The immunological components of human milk and their effect on immune development in infants. Journal of Nutrition 2005, 135, 1–4. [Google Scholar] [CrossRef]
- Wang, J.; Chou, S.; Xu, L.; Zhu, X.; Dong, N.; Shan, A. High specific selectivity and membrane-active mechanism of the synthetic centrosymmetric α-helical peptides with Gly-Gly pairs. Scientific Reports 2015, 5, 15963. [Google Scholar] [CrossRef]
- Rollins-Smith, L.A. The role of amphibian antimicrobial peptides in protection of amphibians from pathogens linked to global amphibian declines. Biochimica et Biophysica Acta Biomembranes 2009, 1788, 1593–1599. [Google Scholar] [CrossRef] [PubMed]
- Conlon, J.M.; Mechkarska, M. Host-defence peptides with therapeutic potential from skin secretions of frogs from the family pipidae. Pharmaceuticals 2014, 7, 58–77. [Google Scholar] [CrossRef]
- Lu, Y.; Ma, Y.; Wang, X.; Liang, J.; Zhang, C.; Zhang, K. The first antimicrobial peptide from sea amphibian. Molecular Immunology 2008, 45, 678–681. [Google Scholar] [CrossRef] [PubMed]
- Vilcinskas, A. Evolutionary plasticity of insect immunity. Journal of Insect Physiology 2013, 59, 123–129. [Google Scholar] [CrossRef] [PubMed]
- Dutta, P.; Sahu, R.K.; Dey, T.; Lahkar, M.D.; Manna, P.; Kalita, J. Beneficial role of insect derived bioactive components against inflammation and its associated complications (colitis and arthritis) and cancer. Chemico-Biological Interactions 2019, 313, 108824. [Google Scholar] [CrossRef] [PubMed]
- Shelomi, M.; Jacobs, C.; Vilcinskas, A.; Vogel, H. The unique antimicrobial peptide repertoire of stick insects. Dev. Compar. Immunol. 2020, 103, 103471. [Google Scholar] [CrossRef]
- Zahedifard, F.; Lee, H.; No, J.H.; Salimi, M.; Seyed, N.; Asoodeh, A. Comparative study of different forms of jellein antimicrobial peptide on leishmania parasite. Experimental Parasitology 2020, 209, 107823. [Google Scholar] [CrossRef] [PubMed]
- Cao, J.; de la Fuente-Nuez, C.; Ou, R.W.; Torres, M.D.T.; Pande, S.G.; Sinskey, A.J. Yeast based synthetic biology platform for antimicrobial peptide production. ACS Synthetic Biology 2018, 896–902. [Google Scholar] [CrossRef]
- Parachin, N.S.; Mulder, K.C.; Viana, A.A.B.; Dias, S.C.; Franco, O.L. Expression systems for heterologous production of antimicrobial peptides. Peptides 2002, 38, 446–456. [Google Scholar] [CrossRef]
- Tang, S.-S.; Prodhan, Z.H.; Biswas, S.K.; Le, C.-F.; Sekaran, S.D. Antimicrobial peptides from different plant sources: isolation, characterisation, and purification. Phytochemistry 2018, 154, 94–105. [Google Scholar] [CrossRef] [PubMed]
- Oh, R.; Lee, M.J.; Kim, Y.-O.; Nam, B.-H.; Kong, H.J.; Kim, J.-W. Myticusin-beta, antimicrobial peptide from the marine bivalve. Mytilus coruscus. Fish and Shellfish Immunology 2020, 99, 342–352. [Google Scholar] [CrossRef]
- Huang, H.N.; Rajanbabu, V.; Pan, C.Y.; Chan, Y.L.; Wu, C.J.; Chen, J.Y. A cancer vaccine based on the marine antimicrobial peptide pardaxin (GE33) for control of bladder associated tumors. Biomaterials 2013, 34, 10151–10159. [Google Scholar] [CrossRef]
- Li, C.; Zhu, C.; Ren, B.; Yin, X.; Shim, S.H.; Gao, Y. Two optimized antimicrobial peptides with therapeutic potential for clinical antibiotic resistant Staphylococcus aureus. European journal of medicinal chemistry 2019, 183, 111686. [Google Scholar] [CrossRef] [PubMed]
- Madanchi, H.; Shoushtari, M.; Kashani, H.H.; and Sardari, S. Antimicrobial peptides of the vaginal innate immunity and their role in the fight against sexually transmitted diseases. New Microbes New Infections 2020, 34, 100627. [Google Scholar] [CrossRef] [PubMed]
- Muhialdin, B.J.; Algboory, H.L.; Kadum, H.; Mohammed, N.K.; Saari, N.; Hassan, Z. Antifungal activity determination for the peptides generated by Lactobacillus plantarum TE10 against Aspergillus flavus in maize seeds. Food Control 2020, 109, 106898. [Google Scholar] [CrossRef]
- Jung, Y.; Kong, B.; Moon, S.; Yu, S.H.; Chung, J.; Ban, C. Envelope-deforming antiviral peptide derived from influenza virus M2 protein. Biochemical and Biophysical Research Communications 2019, 517, 507–512. [Google Scholar] [CrossRef] [PubMed]
- Mahlapuu, M.; Hakansson, J.; Ringstad, L.; Bjorn, C. Antimicrobial peptides: an emerging category of therapeutic agents. Frontiers in Cellular and Infection Microbiology 2016, 6, 194. [Google Scholar] [CrossRef]
- Sun, Q.; Wang, K.; She, R.; Ma, W.; Peng, F.; Jin, H. Swine intestine antimicrobial peptides inhibit infectious bronchitis virus infectivity in chick embryos. Poultry Science 2010, 89, 464–469. [Google Scholar] [CrossRef]
- Huang, H.N.; Pan, C.Y.; Chen, J.Y. Grouper (Epinephelus coioides) antimicrobial peptide epinecidin-1 exhibits antiviral activity against foot-and-mouth disease virus in vitro. Peptides 2018, 106, 91–95. [Google Scholar] [CrossRef] [PubMed]
- Wohlford-Lenane, C.L.; Meyerholz, D.K.; Perlman, S.; Zhou, H.; Tran, D.; Selsted, M.E. Rhesus theta-defensin prevents death in a mouse model of severe acute respiratory syndrome coronavirus pulmonary disease. Journal of Virology 2009, 83, 11385–11390. [Google Scholar] [CrossRef]
- Marimuthu, S.K.; Nagarajan, K.; Perumal, S.K.; Palanisamy, S.; Subbiah, L. (2019). Insilico alpha-helical structural recognition of temporin antimicrobial peptides and its interactions with middle-east respiratory syndrome corona virus. International Journal of Peptide Research and Therapeutics 2019, 26, 1473–1483. [Google Scholar] [CrossRef]
- Mangoni, M.L.; Saugar, J.M.; Dellisanti, M.; Barra, D.; Simmaco, M.; Rivas, L. Temporins, small antimicrobial peptides with leishmanicidal activity. Journal of Biological Chemistry 2005, 280, 984–990. [Google Scholar] [CrossRef]
- Rhaiem, R.B.; Houimel, M. Targeting Leishmania major parasite with peptides derived from a combinatorial phage display library. Acta Tropica 2016, 159, 11–19. [Google Scholar] [CrossRef] [PubMed]
- Abbassi, F.; Raja, Z.; Oury, B.; Gazanion, E.; Piesse, C.; Sereno, D. Antibacterial and leishmanicidal activities of temporin-SHd, a 17-residue long membrane-damaging peptide. Biochimie 2013, 95, 388–399. [Google Scholar] [CrossRef] [PubMed]
- Neshani, A.; Zare, H.; Akbari, E.M.R.; Khaledi, A.; and Ghazvini, K. Epinecidin-1, a highly potent marine antimicrobial peptide with anticancer and immunomodulatory activities. BMC Pharmacology and Toxicology 2019, 20–33. [Google Scholar] [CrossRef] [PubMed]
- Cao, L.; Jiang, W.; Cao, S.; Zhao, P.; Liu, J.; Dong, H. In vitro leishmanicidal activity of antimicrobial peptide KDEL against Leishmania tarentolae. Acta Biochimica et Biophysica Sinica 2019, 51, 1286–1292. [Google Scholar] [CrossRef] [PubMed]
- Wu, D.; Gao, Y.; Qi, Y.; Chen, L.; Ma, Y.; Li, Y. Peptide-based cancer therapy: opportunity and challenge. Cancer Letters 2014, 351, 13–22. [Google Scholar] [CrossRef] [PubMed]
- Ma, R.; Wong, S.W.; Ge, L.; Shaw, C.; Siu, S.W.I.; and Kwok, H.F. In vitro and MD simulation study to explore physicochemical parameters for antibacterial peptide to become potent anticancer peptide. Molecular Theory Oncolytics 2020, 16, 7–19. [Google Scholar] [CrossRef]
- Arias, M.; Haney, E.F.; Hilchie, A.L.; Corcoran, J.A.; Hyndman, M.E.; Hancock, R.E.W. Selective anticancer activity of synthetic peptides derived from the host defence peptide tritrpticin. Biochimica et Biophysica Acta - Biomembranes 2020, 1862, 183228. [Google Scholar] [CrossRef] [PubMed]
- Seefeldt, A.C.; Nguyen, F.; Antunes, S.; Pérébaskine, N.; Graf, M.; Arenz, S. The proline-rich antimicrobial peptide Onc112 inhibits translation by blocking and destabilizing the initiation complex. Nature Structural and Molecular Biology 2015, 22, 470–475. [Google Scholar] [CrossRef] [PubMed]
- Dong, N.; Wang, C.; Zhang, T.; Zhang, L.; Xue, C.; Feng, X. Bioactivity and bactericidal mechanism of histidine rich β-hairpin peptide against gram negative bacteria. The International Journal of Molecular Sciences 2019, 20, 3954. [Google Scholar] [CrossRef]
- Wang, D.; Shen, Y.; Ma, J.; Hancock, R.E.W.; Haapasalo, M. Antibiofilm effect of denantiomeric peptide alone and combined with edta in vitro. Journal of Endodontics 2017, 43, 1862–1867. [Google Scholar] [CrossRef]
- Lei, J.; Sun, I.; Huang, S.; Zhu, C.; Li, P.; He, J. The antimicrobial peptides and their potential clinical applications. American journal of translational research 2019, 11, 3919–3931. [Google Scholar] [PubMed]
- Koehbach, J.; Craik, D.I. The vast structural diversity of antimicrobial peptides. Trends in Pharmacological Sciences 2019, 40, 517–528. [Google Scholar] [CrossRef] [PubMed]
- Mazurkiewicz-Pisarek, A.; Baran, J.; Ciach, T. Antimicrobial Peptides: Challenging Journey to the Pharmaceutical, Biomedical, and Cosmeceutical Use. International Journal of Molecular Sciences 2023, 24, 9031. [Google Scholar] [CrossRef]
- Teixeira, V.; Feio, M.J.; Bastos, M. Role of lipids in the interaction of antimicrobial peptides with membranes. Progress in Lipid Research 2012, 51, 149–177. [Google Scholar] [CrossRef] [PubMed]
- Subbalakshmi, C.; Sitaram, N. FEMS Microbiology Letters 1998, 160, 91. [CrossRef]
- Barriere, S.L. Bacterial resistance to beta-lactams, and its prevention with combination antimicrobial therapy. Pharmacotherapy 1992, 12, 397–402. [Google Scholar] [CrossRef] [PubMed]
- Cirioni, O.; Silvestri, C.; Ghiselli, R.; Orlando, F.; Riva, A.; Mocchegiani, F.; Chiodi, L.; Castelletti, S.; Gabrielli, E.; Saba, V.; et al. Protective effects of the combination of alpha-helical antimicrobial peptides and rifampicin in three rat models of Pseudomonas aeruginosa infection. The Journal of Antimicrobial Chemotherapy 2008, 62, 1332–1338. [Google Scholar] [CrossRef] [PubMed]
- Jeong, N.; Kim, J.Y.; Park, S.C.; Lee, J.K.; Gopal, R.; Yoo, S.; Son, B.K.; Hahm, J.S.; Park, Y.; Hahm, K.S. Antibiotic and synergistic effect of Leu-Lys rich peptide against antibiotic resistant microorganisms isolated from patients with cholelithiasis. Biochemical and Biophysical Research Communications 2010, 399, 581–586. [Google Scholar] [CrossRef] [PubMed]
- Porat, Y.; Marynka, K.; Tam, A.; Steinberg, D.; Mor, A. Acyl-substituted dermaseptin S4 derivatives with improved bactericidal properties, including on oral microflora. Antimicrobial Agents and Chemotheapy 2005, 50, 4153–4160. [Google Scholar] [CrossRef]
- Willcox, M.D.; Hume, E.B.; Aliwarga, Y.; Kumar, N.; Cole, N. A novel cationic-peptide coating for the prevention of microbial colonization on contact lenses. Journal Applied Microbiology 2008, 105, 1817–1825. [Google Scholar] [CrossRef]
- Cirioni, O.; Giacometti, A.; Ghiselli, R.; Kamysz, W.; Orlando, F.; Mocchegiani, F.; Silvestri, C.; Licci, A.; Chiodi, L.; Lukasiak, J.; Saba, V.; Scalise, G. Citropin 1.1-treated central venous catheters improve the efficacy of hydrophobic antibiotics in the treatment of experimental staphylococcal catheter-related infection. Peptides 2006, 27, 1210–1216. [Google Scholar] [CrossRef] [PubMed]
- Sumiyoshi, M.; Miyazaki, T.; Makau, J.N.; Mizuta, S.; Tanaka, Y.; Ishikawa, T.; Makimura, K.; Hirayama, T.; Takazono, T.; Saijo, T. Scientific Reports, 2020; 10, 17745. [CrossRef]
- Ali, R.S.; Falconer, A.; Ikram, M. Expression of the peptide antibiotics human beta defensin-1 and human beta defensin-2 in normal human skin. Journal of Investigative Dermatology. 2001, 117, 106–111. [Google Scholar] [CrossRef]
- Rieg, S.; Garbe, C.; Sauer, B. Dermcidin is constitutively produced by eccrine sweat glands and is not induced in epidermal cells under inflammatory skin conditions. British Journal of Dermatology 2004, 151, 534–539. [Google Scholar] [CrossRef]
- Harder, J.; Dressel, S.; Wittersheim, M. Enhanced expression and secretion of antimicrobial peptides in atopic dermatitis and after superfiial skin injury. Journal of Investigative Dermatology 2010, 130, 1355–1364. [Google Scholar] [CrossRef] [PubMed]
- White, S.H.; Wimley, W.C.; Selsted, M.E. Structure, function, and membrane integration of defensins. Current Opinion in Structural Biology 1995, 5, 521–527. [Google Scholar] [CrossRef] [PubMed]
- Lehrer, R.I.; Ganz, T. Defensins of vertebrate animals. Current Opinion in Immunology 2002, 14, 96–102. [Google Scholar] [CrossRef] [PubMed]
- Ganz, T.; Selsted, M.E.; Szklarek, D. Defensins. Natural peptide antibiotics of human neutrophils. Journal of Clinical Investigation 1985, 76, 1427–1435. [Google Scholar] [CrossRef]
- Ferris, L.K.; Mburu, Y.K.; Mathers, A.R. Human beta defensin 3 induces maturation of human Langerhans cell like dendritic cells: an antimicrobial peptide that functions as an endogenous adjuvant. Journal of Investigative Dermatology 2013, 133, 460–468. [Google Scholar] [CrossRef]
- Rieg, S.; Steffn, H.; Seeber, S. Deficiency of dermcidin derived antimicrobial peptides in sweat of patients with atopic dermatitis correlates with an impaired innate defense of human skin in vivo. Journal of Immunology 2005, 174, 8003–8010. [Google Scholar] [CrossRef]
- Zhang, J.; Dyer, K.D.; Rosenberg, H.F. Human RNase 7: a new cationic ribonuclease of the RNase A superfamily. Nucleic Acids Research 2003, 31, 602–607. [Google Scholar] [CrossRef]
- Koczulla, R.; von Degenfeld, G.; Kupatt, C. An angiogenicrole for the human peptide antibiotic LL-37/hCAP-18. Journal of Clinical Investigation 2003, 111, 1665–1672. [Google Scholar] [CrossRef] [PubMed]
- Alowami, S.; Qing, G.; Emberley, E. Psoriasin (S100A7) expression is altered during skin tumorigenesis. BMC Dermatol 2003, 3, 1. [Google Scholar] [CrossRef] [PubMed]
- Jinquan, T.; Vorum, H.; Larsen, C.G. Psoriasin: a novel chemotactic protein. Journal of Investigative Dermatology 1996, 107, 5–10. [Google Scholar] [CrossRef] [PubMed]
- Abtin, A.; Eckhart, L.; Gläser, R. The antimicrobial heterodimer S100A8/S100A9 (calprotectin) is upregulated by bacterial flgellin in human epidermal keratinocytes. Journal of Investigative Dermatology 2010, 130, 2423–2430. [Google Scholar] [CrossRef] [PubMed]
- Selwitz, R.H.; Ismail, A.I.; Pitts, N.B. (Dental caries). Lancet 2007, 369, 51–59. [Google Scholar] [CrossRef] [PubMed]
- Gomes, B.P.; Pinheiro, E.T.; Jacinto, R.C.; Zaia, A.A.; Ferraz, C.C.; Souza-Filho, F.J. Microbial analysis of canals of root-filled teeth with periapical lesions using polymerase chain reaction. The Journal of Endodontics 2008, 34, 537–540. [Google Scholar] [CrossRef] [PubMed]
- Paris, S.; Wolgin, M.; Kielbassa, A.M.; Pries, A.; Zakrzewicz, A. Gene expression of human beta-defensins in healthy and inflamed human dental pulps. Journal of Endodontics 2009, 35, 520–523. [Google Scholar] [CrossRef]
- Lee, S.H.; Baek, D.H. Antibacterial and neutralizing effect of human beta-defensins on Enterococcus faecalis and Enterococcus faecalis lipoteichoic acid. The Journal of Endodontics 2012, 38, 351–356. [Google Scholar] [CrossRef] [PubMed]
- Krzysciak, W.; Jurczak, A.; Piatkowski, J.; Koscielniak, D.; Gregorczyk-Maga, I.; Kolodziej, I.; Papiez, M.A.; Olczak-Kowalczyk, D. Effect of histatin-5 and lysozyme on the ability of Streptococcus mutans to form biofilms in in vitro conditions. Postępy Higieny i Medycyny Doświadczalnej 2015, 69, 1056–1066. [Google Scholar]
- Guo, Y.J.; Zhang, B.; Feng, X.S.; Ren, H.X.; Xu, J.R. Human cathelicidin ll-37 enhance the antibiofilm effect of egcg on Streptococcus mutans. BMC Oral Health 2016, 16, 101. [Google Scholar] [CrossRef]
- Kreling, P.F.; Aida, K.L.; Massunari, L.; Caiaffa, K.S.; Percinoto, C.; Bedran, T.B.; Spolidorio, D.M.; Abuna, G.F.; Cilli, E.M.; Duque, C. Cytotoxicity and the effect of cationic peptide fragments against cariogenic bacteria under planktonic and biofilm conditions. Biofouling 2016, 32, 995–1006. [Google Scholar] [CrossRef] [PubMed]
- Ahn, K.B.; Kim, A.R.; Kum, K.Y.; Yun, C.H.; Han, S.H. The synthetic human beta-defensin-3 c15 peptide exhibits antimicrobial activity against Streptococcus mutans, both alone and in combination with dental disinfectants. Journal of Microbiology 2017, 55, 830–836. [Google Scholar] [CrossRef] [PubMed]
- Winfred, S.B.; Meiyazagan, G.; Panda, J.J.; Nagendrababu, V.; Deivanayagam, K.; Chauhan, V.S.; Venkatraman, G. Antimicrobial activity of cationic peptides in endodontic procedures. Europian Journal of Dentistry 2014, 8, 254–260. [Google Scholar] [CrossRef] [PubMed]
- Sousa, M.G.C.; Xavier, P.D.; Cantuaria, A.P.C.; Porcino, R.A.; Almeida, J.A.; Franco, O.L.; Rezende, T.M.B. Host defense peptide idr-1002 associated with ciprofloxacin as a new antimicrobial and immunomodulatory strategy for dental pulp revascularization therapy. Microbial Pathogens 2020, 104634. [Google Scholar] [CrossRef] [PubMed]
- Ouhara, K.; Komatsuzawa, H.; Yamada, S.; Shiba, H.; Fujiwara, T.; Ohara, M.; Sayama, K.; Hashimoto, K.; Kurihara, H.; Sugai, M. Susceptibilities of periodontopathogenic and cariogenic bacteria to antibacterial peptides, {beta}-defensins and ll37, produced by human epithelial cells. Journal of Antimicroial Chemotherapy 2005, 55, 888–896. [Google Scholar] [CrossRef]
- Into, T.; Inomata, M.; Shibata, K.; Murakami, Y. Effect of the antimicrobial peptide ll-37 on toll-like receptors 2-, 3- and 4-triggered expression of il-6, il-8 and cxcl10 in human gingival fibroblasts. Cell Immunol 2010, 264, 104–109. [Google Scholar] [CrossRef]
- Brogden, K.A.; Nordholm, G.; Ackermann, M. Antimicrobial activity of cathelicidins bmap28, smap28, smap29, and pmap23 against Pasteurella multocida is more broad-spectrum than host species specific. Veterinary microbiology 2007, 119, 76–81. [Google Scholar] [CrossRef] [PubMed]
- Brice, D.C.; Toth, Z.; Diamond, G. LL-37 disrupts the kaposi's sarcoma-associated herpesvirus envelope and inhibits infection in oral epithelial cells. Antiviral Research 2018, 158, 25–33. [Google Scholar] [CrossRef]
- Winter, J.; Pantelis, A.; Reich, R.; Martini, M.; Kraus, D.; Jepsen, S.; Allam, J.P.; Novak, N.; Wenghoefer, M. Human beta-defensin-1, -2, and -3 exhibit opposite effects on oral squamous cell carcinoma cell proliferation. Cancer Investigation 2011, 29, 196–201. [Google Scholar] [CrossRef]
- Goldman, M.J.; Anderson, G.M.; Stolzenberg, E.D.; Kari, U.P.; Zasloff, M.; Wilson, J.M. Human beta-defensin-1 is a salt-sensitive antibiotic in lung that is inactivated in cystic fibrosis. Cell 1997, 88, 553–560. [Google Scholar] [CrossRef]
- Duits, L.A.; Ravensbergen, B.; Rademaker, M.; Hiemstra, P.S.; Nibbering, P.H. Expression of beta-defensin 1 and 2 mRNA by human monocytes, macrophages and dendritic cells. Journal of Immunological Sciences 2002, 106, 517–525. [Google Scholar] [CrossRef]
- Hess, C.; Herr, C.; Beisswenger, C.; Zakharkina, T.; Schmid, R.M.; Bals, R. Myeloid RelA regulates pulmonary host defense networks. European Respiratory Journal 2010, 35, 343–352. [Google Scholar] [CrossRef]
- Scharf, S.; Vardarova, K.; Lang, F.; Schmeck, B.; Opitz, B.; Flieger, A.; Heuner, K.; Hippenstiel, S.; Suttorp, N.; N’Guessan, P.D. Legionella pneumophila induces human beta defensin-3 in pulmonary cells. Respiratory Research 2010, 11, 93. [Google Scholar] [CrossRef]
- Rivas-Santiago, B.; Serrano, C.J.; Enciso-Moreno, J.A. Susceptibility to infectious diseases based on antimicrobial peptide production. Infection and Immunity 2009, 77, 4690–4695. [Google Scholar] [CrossRef]
- Merkel, D.; Rist, W.; Seither, P.; Weith, A.; Lenter, M.C. Proteomic study of human bronchoalveolar lavage fluids from smokers with chronic obstructive pulmonary disease by combining surface-enhanced laser desorption/ionization-mass spectrometry profiling with mass spectrometric protein identification. Proteomics 2005, 5, 2972–2980. [Google Scholar] [CrossRef]
- Felgentreff, K.; Beisswenger, C.; Griese, M.; Gulder, T.; Bringmann, G.; Bals, R. The antimicrobial peptide cathelicidin interacts with airway mucus. Peptides 2006, 27, 3100–3106. [Google Scholar] [CrossRef]
- Hase, K.; Murakami, M.; Iimura, M. Expression of LL-37 by human gastric epithelial cells as a potential host defense mechanism against Helicobacter pylori. Gastroenterology 2003, 125, 1613–1625. [Google Scholar] [CrossRef]
- Grubman, A.; Kaparakis, M.; Viala, J. The innate immune molecule, NOD1, regulates direct killing of Helicobacter pylori by antimicrobial peptides. Cellular Microbiology 2010, 12, 626–639. [Google Scholar] [CrossRef]
- Kotarsky, K.; Sitnik, K.M.; Stenstad, H. A novel role for constitutively expressed epithelial-derived chemokines as antibacterial peptides in the intestinal mucosa. Mucosal immunology 2010, 3, 40–48. [Google Scholar] [CrossRef]
- Harada, K.; Ohba, K.; Ozaki, S. Peptide antibiotic human beta-defensin-1 and -2 contribute to antimicrobial defense of the intrahepatic biliary tree. Hepatology, 2004; 40, 925–925. [Google Scholar] [CrossRef]
- Wehkamp, J.; Chu, H.; Shen, B. Paneth cell antimicrobial peptides: topographical distribution and quantification in human gastrointestinal tissues. FEBS Letters 2006, 580, 5344–5350. [Google Scholar] [CrossRef]
- Bevins, C.L.; Nita, H.; Salzman, N.H.; Ghosh, D.; Huttner, K.M. Human defensin-5 (HD-5) transgenic mice: paneth cell expression and protection from lethal Salmonella typhimurium infection. World Journal of Gastroenterology 2002, 122, 169. [Google Scholar]
- Kaser, A.; Lee, A.-H.; Franke, A. XBP1 links ER stress to intestinal inflammation and confers genetic risk for human inflammatory bowel disease. Cell 2008, 134, 743–756. [Google Scholar] [CrossRef]
- Zilbauer, M.; Dorrell, N.; Boughan, P.K. Intestinal innate immunity to Campylobacter jejuni results in induction of bactericidal human beta-defensins 2 and 3. Infection and Immunity 2005, 73, 7281–7289. [Google Scholar] [CrossRef]
- Limura, M.; Gallo, R.L.; Hase, K.; Miyamoto, Y.; Eckmann, L.; Kagnoff, M.F. Cathelicidin mediates innate intestinal defense against colonization with epithelial adherent bacterial pathogens. Journal of Immunology 2005, 174, 4901–4907. [Google Scholar] [CrossRef]
- Fahlgren, A.; Hammarstrom, S.; Danielsson, A.; Hammarstrom, M.L. Increased expression of antimicrobial peptides and lysozyme in colonic epithelial cells of patients with ulcerative colitis. Clinical and Experimental Immunology 2003, 131, 90–101. [Google Scholar] [CrossRef]
- Kanyshkova, T.G.; Buneva, V.N.; Nevinsky, G.A. Lactoferrin and its biological functions. Biochemistry (Mosc) 2001, 66, 1–7. [Google Scholar] [CrossRef]
- Collins, H.L. Withholding iron as a cellular defence mechanism--friend or foe? The European Journal of Immunology 2008, 38, 1803–1806. [Google Scholar] [CrossRef]
- Van Bambeke, F. Glycopeptides and glycodepsipeptides in clinical development: a comparativereview of their antibacterial spectrum, pharmacokinetics and clinical efficacy. Current Opinion in Investigational Drugs 2006, 7, 740–749. [Google Scholar]
- Lande, R.; Botti, E.; Jandus, C.; Dojcinovic, D.; Fanelli, G.; Conrad, C. The antimicrobial peptide LL37 is a T-cell autoantigen in psoriasis. Nature Communications 2014, 5, 5621. [Google Scholar] [CrossRef]
- Itzhaki, R.F.; Wozniak, M.A.; Appelt, D.M.; Balin, B.J. Infiltration of the brain by pathogens causes Alzheimer’s disease. Neurobiol Aging 2004, 25, 619–627. [Google Scholar] [CrossRef]
- Carter, C. Alzheimer’s disease: APP, gamma secretase, APOE, CLU, CR1, PICALM, ABCA7, BIN1, CD2AP, CD33, EPHA1, and MS4A2, and their relationships with Herpes simplex, C. pneumoniae, other suspect pathogens, and the immune system. International Journal of Alzheimer's Disease Research 2011, 1–34. [Google Scholar] [CrossRef]
- Sparks, S.P.; Steffen, M.J.; Smith, C.; Jicha, G.; Ebersole, J.L.; Abner, E. Serum antibodies to periodontal pathogens are a risk factor for Alzheimer’s disease. Alzheimers and Dementia 2012, 8, 196–203. [Google Scholar] [CrossRef]
- Kourie, J.I.; Shorthouse, A.A. Properties of cytotoxic peptide-formed ion channels. American Journal of Physiology-Cell Physiology 2000, 278, C1063–1087. [Google Scholar] [CrossRef]
- Su, Y.; Zhang, K.; Schluesener, H. Antimicrobial peptides in the brain. Archivum Immunologiae et Therapiae Experimentalis 2010, 58, 365–377. [Google Scholar] [CrossRef]
- Kagan, B.L.; Jang, H.; Capone, R.; Teran Arce, F.; Ramachandran, S.; Lal, R. Antimicrobial properties of amyloid peptides. Molecular Pharmaceutics 2011, 9, 708–717. [Google Scholar] [CrossRef]
- Edfeldt, K.; Agerberth, B.; Rottenberg, M.E.; Gudmundsson, G.H.; Wang, X.B.; Mandal, K.; Xu, Q.; Yan, Z.Q. Involvement of the Antimicrobial Peptide LL-37 in Human Atherosclerosis. Arteriosclerosis, Thrombosis, and Vascular Biology 2006, 26, 1551–1557. [Google Scholar] [CrossRef]
- Baxter, A.A.; Lay, F.T.; Poon, I.K.; Kvansakul, M.; Hulett, M.D. Tumor cell membranetargeting cationic antimicrobial peptides: novel insights into mechanisms of action and therapeutic prospects. Cellular and Molecular Life Sciences 2017, 74, 3809–3825. [Google Scholar] [CrossRef]
- Tsai, T.L.; Li, A.C.; Chen, Y.C.; Liao, Y.S.; Lin, T.H. Antimicrobial peptide m2163 or m2386 identified from Lactobacillus casei ATCC 334 can trigger apoptosis in the human colorectal cancer cell line SW480. Tumor Biology 2015, 36, 3775–3789. [Google Scholar] [CrossRef]
- Ong, P.Y.; Ohtake, T.; Brandt, C.; Strickland, I.; Boguniewicz, M.; Ganz, T.; Gallo, R.L.; Leung, D.Y. Endogenous Antimicrobial Peptides and Skin Infections in Atopic Dermatitis. New England Journal of Medicine 2002, 347, 1151–1160. [Google Scholar] [CrossRef]
- Frohm, M.; Agerberth, B.; Ahangari, G.; Ståhle-Bäckdahl, M.; Lidén, S.; Wigzell, H.; Gudmundsson, G.H. The Expression of the Gene Coding for the Antibacterial Peptide LL-37 Is Induced in Human Keratinocytes during Inflammatory Disorders. Journal of Biological Chemistry 1997, 272, 15258–15263. [Google Scholar] [CrossRef]
- Friedland, H.D.; Sharp, D.D.; Robinson, J.R. Double-blind, randomized, vehicle-controlled study to asses the safety and efficacy of MBI 594AN in the treatment of acne vulgaris. Abstracts of the 61st annual meeting of American Academy of Dermatology 2003, 61, 22. [Google Scholar]
- McInturff, J.E.; Wang, S.J.; Machleidt, T.; Richard, L.T.; Oren, A.; Hertz, C.J.; Krutzik, S.R.; Hart, S.; Zeh, K.; Anderson, D.H.; Gallo, R.L.; Modlin, R.L.; Kim, J. Granulysin-Derived Peptides Demonstrate Antimicrobial and Anti-Inflammatory Effects Against Propionibacterium acnes. Journal of Investigative Dermatology 2005, 125, 256–263. [Google Scholar] [CrossRef]
- Boman, H.G. Innate immunity and the normal microflora. Immunological Reviews 2000, 173, 5–16. [Google Scholar] [CrossRef]
- Tanida, T.; Okamoto, T.; Okamoto, A.; Wang, H.; Hamada, T.; Ueta, E.; Osaki, T. Decreased excretion of antimicrobial proteins and peptides in saliva of patients with oral candidiasis. Journal of Oral Pathology Medicine 2003, 32, 586–594. [Google Scholar] [CrossRef]
- Pütsep, K.; Carlsson, G.; Boman, H.G.; Andersson, M. Deficiency of antibacterial peptides in patients with morbus Kostmann: an observation study. Lancet 2002, 360, 1144–1149. [Google Scholar] [CrossRef]
- Gusman, H.; Travis, J.; Helmerhorst, E.J.; Potempa, J.; Troxler, R.F.; Oppenheim, F.G. Salivary Histatin 5 Is an Inhibitor of Both Host and Bacterial Enzymes Implicated in Periodontal Disease. Infection and Immunity 2001, 69, 1402–1408. [Google Scholar] [CrossRef]
- Yoshinari, M.; Kato, T.; Matsuzaka, K.; Hayakawa, T.; Inoue, T.; Oda, Y.; Okuda, K.; Shimono, M. Adsorption behavior of antimicrobial peptide histatin 5 on PMMA. Journal of Biomedical Materials Research Part B: Applied Biomaterials 2005, 77B, 47–54. [Google Scholar] [CrossRef]
- Wehkamp, J.; Salzman, N.H.; Porter, E.; Nuding, S.; Weichenthal, M.; Petras, R.E.; Shen, B.; Schaeffeler, E.; Schwab, M.; Linzmeier, R.; Feathers, R.W.; Chu, H.; Lima, H.; Fellermann, K.; Ganz, T.; Stange, E.F.; Bevins, C.L. Reduced Paneth cell α-defensins in ileal Crohn’s disease. Proceedings of the National Academy of Sciences 2005, 102, 18129–18134. [Google Scholar] [CrossRef]
- Fahlgren, A.; Hammarström, S.; Danielsson; Hammarström, M. L. β-Defensin-3 and -4 in intestinal epithelial cells display increased mRNA expression in ulcerative colitis. Clinical and Experimental Immunology 2004, 137, 379–385. [Google Scholar] [CrossRef]
- Baker, M.A.; Maloy, W.L.; Zasloff, M.; Jacob, L.S. Anticancer efficacy of magainin2 and analogue peptides. Cancer Research and Treatment 1993, 53, 3052–3057. [Google Scholar]
- Sloballe, P.W.; Lee Maloy, W.; Myrga, M.L.; Jacob, L.S.; Herlyn, M. Experimental local therapy of human melanoma with lytic magainin peptides. International Journal of Cancer 1995, 60, 280–284. [Google Scholar] [CrossRef]
- Moore, A.J.; Devine, D.A.; Bibby, M.C. Preliminary experimental anticancer activity of cecropins. Peptide Research 1994, 7, 265–269. [Google Scholar]
- Yoo, Y.; Watanabe, S.; Watanabe, R.; Hata, K.; Shimazaki, K.; Azuma, I. Bovine Lactoferrin and Lactoferricin, a Peptide Derived from Bovine Lactoferrin, Inhibit Tumor Metastasis in Mice. Japanese Journal of Cancer Research 1997, 88, 184–190. [Google Scholar] [CrossRef]
- Eliassen, L.T.; Berge, G.; Leknessund, A.; Wikman, M.; Lindin, I.; Løkke, C.; Ponthan, F.; Johnsen, J.I.; Sveinbjørnsson, B.; Kogner, P.; Flægstad, T.; Rekdal, Y. The antimicrobial peptide, lactoferricin B, is cytotoxic to neuroblastoma cells in vitro and inhibits xenograft growth in vivo. International Journal of Cancer 2006, 119, 493–500. [Google Scholar] [CrossRef]
- Higazi, A.A.R.; Lavi, E.; Bdeir, K.; Ulrich, A.M.; Jamieson, D.G.; Rader, D.J.; Usher, D.C.; Kane, W.; Ganz, T.; Cines, D.B. Defensin Stimulates the Binding of Lipoprotein (a) to Human Vascular Endothelial and Smooth Muscle Cells. Blood 1997, 89, 4290–4298. [Google Scholar] [CrossRef]
- Higazi, A.A.; Nassar, T.; Ganz, T.; Rader, D.J.; Udassin, R.; Bdeir, K.; Hiss, E.; Sachais, B.S.; Williams, K.J.; Leitersdorf, E.; Cines, D.B. The alpha-defensins stimulate proteoglycan-dependent catabolism of low-density lipoprotein by vascular cells: a new class of inflammatory apolipoprotein and a possible contributor to atherogenesis. Blood, 2000; 96, 1393–1398. [Google Scholar] [CrossRef]
- Chavakis, T.; Cines, D.B.; Rhee, J.S.; Liang, O.D.; Schubert, U.; Hammes, H.P.; Higazi, A.A.R.; Nawroth, P.P.; Preissner, K.T.; Bdeir, K. Regulation of neovascularization by human neutrophil peptides (α-defensins): a link between inflammation and angiogenesis. Federation of American Societies for Experimental Biology 2004, 18, 1306–1308. [Google Scholar] [CrossRef]
- Edfelt, K.; Agerberth, B.; Rottenberg, M.E.; Gudmundsson, G.H.; Wang, X.B.; Mandal, K. Involvement of the antimicrobial peptide LL-37 in human atherosclerosis. Arteriosclerosis, Thrombosis, and Vascular Biology 2006, 26, 1551–1557. [Google Scholar] [CrossRef]
- Varoga, D.; Pufe, T.; Mentlein, R.; Kohrs, S.; Grohmann, S.; Tillmann, B.; Hassenpflug, J.; Paulsen, F. Expression and regulation of antimicrobial peptides in articular joints. Annals of Anatomy - Anatomischer Anzeiger 2005, 187, 499–508. [Google Scholar] [CrossRef]
- Mygind, P.H.; Fischer, R.L.; Schnorr, K.M.; Hansen, M.T.; Sönksen, C.P.; Ludvigsen, S.; Raventós, D.; Buskov, S.; Christensen, B.; De Maria, L.; Taboureau, O.; Yaver, D.; Elvig-Jørgensen, S.G.; Sørensen, M.V.; Christensen, B.E.; Kjærulff, S.; Frimodt-Moller, N.; Lehrer, R.I.; Zasloff, M.; Kristensen, H.H. Plectasin is a peptide antibiotic with therapeutic potential from a saprophytic fungus. Nature 2005, 437, 975–980. [Google Scholar] [CrossRef]
- Paquette, D.W.; Simpson, D.M.; Friden, P.; Braman, V.; Williams, R.C. Safety and clinical effects of topical histatin gels in humans with experimental gingivitis. Journal of Clinical Periodontology 2002, 29, 1051–1058. [Google Scholar] [CrossRef]
- Review Editor, B. Micrologix Licenses Anti-Infective to Fujisawa. Pharma Deals Review 2002, (26). [CrossRef]
Figure 1.
Classification of Antimicrobial Peptides.
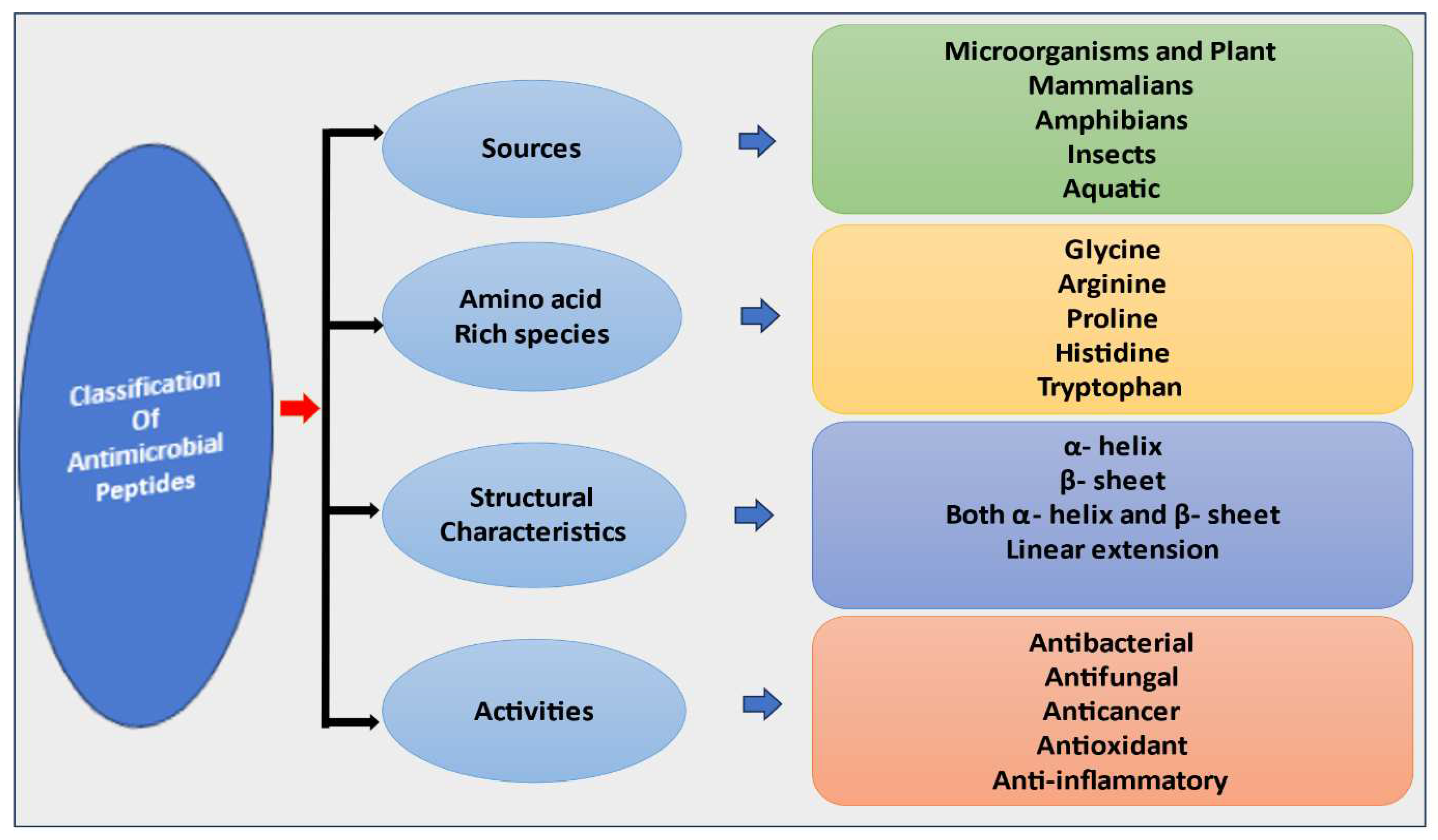
Figure 2.
Application of antimicrobial peptides in different fields.
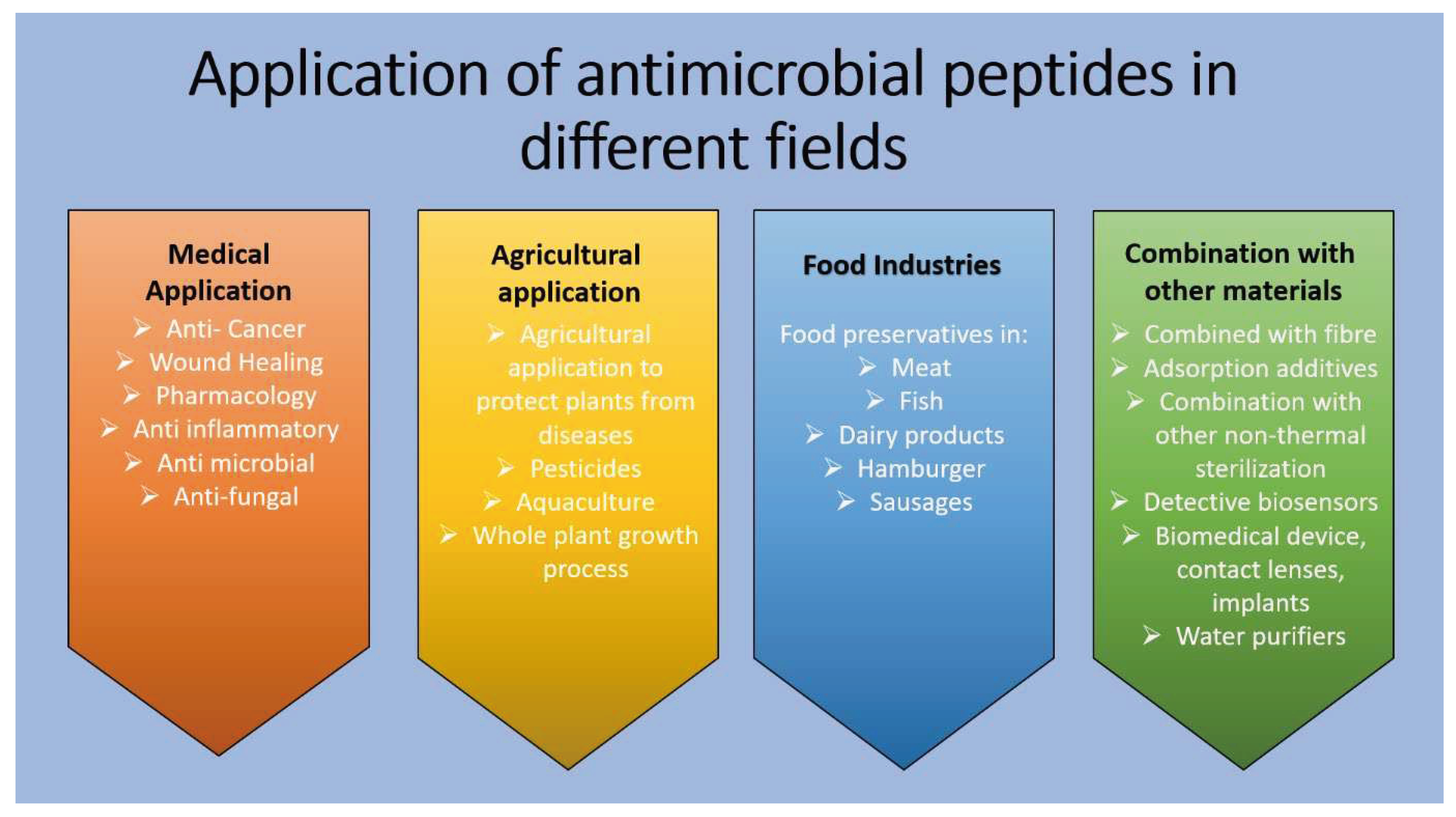
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



