
Submitted:
06 December 2023
Posted:
07 December 2023
You are already at the latest version
Abstract
We studied the capacity of glutathione and a specific inactivated dry yeast rich in glutathione to protect white grape must from browning compared to using sulphur dioxide. The results indicate that sulphur dioxide drastically reduces the oxygen consumption rate, protects hydroxycinnamic acids from oxidation and prevents grape must against browning even in the presence of laccase. Glutathione, pure or in the form of a specific inactivated dry yeast rich in glutathione, also reduces the oxygen consumption rate but to a lesser extent than sulphur dioxide. It favours the formation of GRP from hydroxycinnamic acids and effectively protects grape must against browning in healthy grape conditions. Nevertheless, in the presence of laccase it was not effective enough. Therefore, both forms of glutathione can be considered as interesting alternative tools to sulphur dioxide for preventing browning in white grape must, but only when the grapes are healthy.
Keywords:
Grape must
; Glutathione
; Inactivated Dry Yeasts
; Browning
; Laccase
1. Introduction
One of the major problems of the current oenology is enzymatic browning [1] especially when the grapes are affected by the filamentous fungi Botrytis cinerea [2]. Browning is an enzymatic oxidation process in several foods that causes the appearance of brown colour, which sometimes leads consumers to refuse them [3]. This problem is particularly damaging in oenology since grape must is very susceptible to browning [4].
Two enzymes belonging to the broad family of oxidoreductases called polyphenol oxidases (PPOs) are responsible for enzymatic browning: tyrosinase (EC 1.14.18.1), which is naturally present in grape berries [5], and laccase (EC 1.10.3.2), which is only present in grapes infected by epiphytic fungi, mainly Botrytis cinerea [6].
Both enzymes are multicopper oxidases that use oxygen to primarily oxidize some phenolic compounds such as hydroxycinnamic acids and other diphenols [7]. Due to the polyphenol oxidase activity, some diphenols are oxidized to diquinones, which polymerize later to form melanins [3]. These melanins are responsible for the darkening of the yellow colour in white wines (browning) and for the loss of red colour in red wines (oxidasic haze). When the grapes are affected by grey rot, the laccase activity can be much greater than that of tyrosinase in healthy grapes [8] and therefore the risk of browning is much higher. Moreover, laccase can oxidize a greater range of substrates than tyrosinase [4,6].
The most common method used in wineries to protect the grape must from browning is to add sulphur dioxide because it is a powerful inhibitor of tyrosinase and laccase [9]. Sulphur dioxide is also used in winemaking for its antimicrobial properties [10]. Giménez et al. [7] reported in a kinetic study that sulphur dioxide drastically reduces the Vmax and increases the K0.5 of laccase, which shows that this additive not only inhibits this enzyme but also decreases its affinity versus the substrates. Tyrosinase is very sensitive to sulphur dioxide and therefore small doses of this additive inactivate it. However, laccase is more resistant to sulphur dioxide and, unlike tyrosinase, can be present in wine after alcoholic fermentation [9,11].
However, the recent tendency in winemaking is to diminish and even eliminate sulphur dioxide owing to its negative effects on the environment [12] and human health [13]. Therefore, some other additives have been proposed as alternatives to sulphur dioxide to prevent browning, such as ascorbic acid [14], inert gases [15], oenological tannins [16,17] and, more recently, the use of reduced glutathione [7,18,19] or inactivated dry yeasts rich in glutathione [20,21].
Ascorbic acid consumes oxygen very quickly [17,22] making it unavailable to phenol oxidases. However, ascorbic acid generates hydrogen peroxide after consuming oxygen and consequently it can cause subsequent oxidations in the wine [4]. Therefore, the use of ascorbic acid always requires the presence of sulphur dioxide to prevent wine oxidation [23] and so it must be considered as a complement but never as an alternative.
Using inert gases minimizes the presence of oxygen and, by eliminating its substrate, oxidation caused by phenol oxidases is avoided [11,14].
Oenological tannins have been shown to be effective inhibitors of laccase activity [16,17]. Consequently, when musts and wines are supplemented with oenological tannins, they exert a protective effect on the colour of the wine.
Glutathione reduces browning because it reacts with the ortho-quinones originated by enzymatic oxidation of ortho-diphenols to produce 2-S-glutathionylcaftaric acid, commonly known as grape reaction product (GRP). In this way, glutathione traps the ortho-quinones in a colourless form and thus limits the formation of melanins [18,24].
Among all these possible alternatives to sulphur dioxide for preventing browning, glutathione is probably the most promising. The use of glutathione in grape must and wine was authorized in 2015 by the International Organization of Vine and Wine (OIV) at a maximum dosage of 20 mg/L [25,26]. Nevertheless, glutathione is not often used in its pure form due to its very high price. Therefore, using certain inactivated dry yeasts as a source of glutathione (IDY-GSH) has been much more successful as it is a more economical solution [20,21]. Using these inactivated dry yeasts with guaranteed levels of glutathione in winemaking was authorized by the OIV in 2018 [27].
In a recent article [28] our research group has studied the influence of glutathione on oxygen consumption kinetics and colour of the grape must under similar conditions that those described in this work. Since the results were very interesting, we decided to repeat the experience analyzing also hydroxycinnamic acids and GRP in order to try to better understand the mechanism by which glutathione protects the must from browning. The aim of this paper was therefore to study the mechanism by which reduced glutathione, whether pure or as commercial inactivated dry yeasts rich in glutathione, exert a protective effect against the oxidation caused by polyphenol oxidases.
2. Materials and Methods
2.1. Chemicals
Syringaldazine (purity ≥ 98%), and FeSO4・7H2O (purity ≥ 99%) were purchased from Sigma-Aldrich (Madrid, Spain). L-( +)-Tartaric acid (purity ≥ 99.5%), sodium hydroxide (purity ≥ 98%), sodium acetate (purity ≥ 99%), acetonitrile (purity ≥ 99%), Methanol (purity ≥ 99%), and CuSO4・5H2O (purity ≥ 99%) were purchased from Panreac (Barcelona, Spain). Ethanol (96% vol.) was supplied by Fisher Scientific (Madrid, Spain). L-glutathione reduced (purity ≥ 98%) was purchased from Sigma-Aldrich (Madrid, Spain). Cellulose membranes of 3.5 KDa (6.4 mL/cm) were supplied by Spectrum Laboratories, Inc (Rancho Dominguez, USA). The equipment used was as follows: a Hpand and Entris II Series Analytical Balance (Sartorius, Goettingen, Germany), a spectrophotometer UV–Vis Helios Alpha ™ (Thermo Fisher Scientific Inc., Waltham, MA, USA); a centrifuge Heraeus™ Primo™ (Thermo Fisher Scientific Inc., Waltham, MA, USA); a liquid chromatograph Agilent Series 1200 (Agilent, Germany) equipped with a photodiode array detector (G1315D) and a Zorbax Eclipse XDB C18 column (4.6 x 150mm).
2.2. Collecting and preparing the samples
Muscat of Alexandria grapes were collected in the experimental vineyard of the Rovira i Virgili University (Mas dels Frares, Constantí, Tarragona: 41°08’44.1’’N, 1°11’51.0’’E, height: 59 masl) during the 2022 vintage. The grapes were pressed in an environment saturated with nitrogen with a vertical manual press until 60% of their must was extracted and transferred in a 750 mL bottle, also saturated with nitrogen.
2.3. Synthetic buffer
A solution of 4 g/L of L-( +)-tartaric acid, 3 mg of iron/L, in the form of FeCl3·6H2O, and 0.3 mg of copper/L in the form of CuSO4·5H2O adjusted to pH 3.5 with NaOH was used for all experiments.
2.4. Production of the laccase enzyme and measurement of enzymatic activity
Active laccase extracts were obtained from the isolate 213 strain of Botrytis cinerea following the procedure reported by Vignault et al. [17]. This laccase extract was treated with 0.16 g of PVPP/mL for 10 minutes and centrifuged at 7500 rpm for 10 minutes. The supernatant was subsequently dialyzed with 3.5 KDa cellulose membrane for two days against a 0.3 M ammonium formate solution and for two more days against distilled water. The laccase activity was determined using the syringaldazine test method [29].
2.5. Measurement of oxygen consumption kinetics
The experimental conditions were those described by Giménez et al. [28]. The assays were carried out in clear glass flasks (66 mL) with an oxygen sensor spot (PreSens Precision Sensing GmbH, order code: SP-PSt3-NAU-D5-CAF; batch number: 1203- 01_PSt3-0828-01, Regensburg, Germany) for noninvasively measuring the dissolved oxygen by luminescence (Nomasense TM O2 Trace Oxygen Analyzer, Nomacorc S.A., Thimister Clermont, Belgium).
The grape must was diluted (20% (v/v)) with the synthetic buffer mentioned above to reduce the oxygen consumption rate of the grape must so it could be monitored with greater precision, and also to uniformize the pH of the different measurements. More specifically, thirteen mL of grape must and 52 mL of buffer were added to each flask, with the different antioxidant agents. The antioxidants used were: sulphur dioxide (20 mg/L in the form of potassium disulphite, pure reduced glutathione (20 mg/L) and a commercial inactivated dry yeast (Glutastar™, Lallemand Inc, Montreal, Canada) rich in glutathione (400 mg/L). A control without anything added to it was also prepared. These assays were also performed by combining sulphur dioxide with each of the other antioxidant agents with and without the addition of 2 U of laccase/mL (enzyme units or μmol/minute). All assays were performed in triplicate.
The bottles were immediately saturated in oxygen (around 7–8 mg O2/L) by manual vigorous stirring for a few seconds. The oxygen concentration was measured [30] periodically until asymptotic behaviour was reached (around 3 h) to determine the oxygen consumption kinetics. All measurements were performed at 22 ± 2 °C. The total oxygen consumption capacity (TOCC) was determined using a mathematical model previously described by Pons-Mercadé et al. [31]. When the oxygen concentrations were below 1 mg/L or oxygen consumption attained an asymptotic behaviour, the samples were added with 50 mg of SO2/L to avoid the colour evolution. The samples were then used for colour measurements and for HPLC analysis of hydroxycinnamic acids and GRP.
2.6. Colour measurements
The intensity of the yellow colour (A420nm) and the CieL*a*b* coordinates of the samples were measured according to Ayala et al. [32]. The data were processed using the MSCV software [33]. The total colour difference (ΔEab*) was calculated using the following formula: ΔEab* = ((L*1-L*2)2 + (a*1-a*2)2 + (b*1-b*2)2)1/2, where L* is the lightness; a* is the colour’s green–red component, and b* is the colour’s blue–yellow component. ΔEab* is used to determine whether the difference between two samples can be detected visually by the human eye. Generally, it is considered that the difference is visible to the human eye when ΔEab* > 3 units [34].
2.7. Hydroxycinnamic acid and GRP analysis by HPLC-DAD
Hydroxycinnamic acids and GRP were analysed by the reversed-phase HPLC−DAD following the method described by Lago-Vanzela et al. [35]. This comprised an Agilent Series 1200 HPLC (Agilent, Waldbronn, Germany) equipped with a DAD (G1315D) coupled to an Agilent Chem Station (version B.01.03) data processing station. The samples were filtered (0.20 μm, polyester membrane, Chromafil PET 20/25, Machery-Nagel, Düren, Germany) and then injected (20 μL) into a Zorbax Eclipse XDB C18 column (4.6 x 150mm). The solvent was A [water/formic acid/acetonitrile (88.5:8.5:3, v/v/v)], B [water/formic acid/acetonitrile (41.5:8.5:50, v/v/v)] and C [water/formic acid/methanol (1.5:8.5:90, v/v/v)] and the flow rate was 0.19 mL/ min. The gradient was: from 0 to 37 min: A = 96%/B = 4%; from 37 to 51 min: A = 70%/B = 17%/C = 13%; from 51 to 57 min: A = 50%/B = 30%/C = 20%; from 57 to 64 min A = 0%/B = 50%/C = 50%. The compounds were quantified by measuring the absorbance at 320 nm with the external calibration curve of caftaric acid. The different compounds were identified according to the retention time that was previously identified with an HPLC–DAD-ESI-MS/MS analysis using the same chromatographic conditions.
2.8. Statistics
All data are expressed as the arithmetic average ± standard deviation of three replicates. One-factor analysis of variance (ANOVA F test) was carried out using the SPSS 15.0 software (SPSS Inc., Chicago, IL). Significant differences were considered when the p value was less than 0.05.
3. Results
3.1. Oxygen consumption kinetics
Figure 1A shows the oxygen consumption kinetics of the diluted grape must either supplemented or not with sulphur dioxide, glutathione and with the combination of both additives in the absence or presence of 2 U of laccase/mL.
The control diluted grape must without any additives initially consumed oxygen very quickly and then moderately until reaching values of around 2 mg O2/L after 3 hours. This behaviour is probably due to the depletion of substrates for polyphenol oxidase, which slows down the reaction rate. In contrast, when the sample was supplemented with sulphur dioxide the oxygen consumption rate (OCR) decreased drastically, confirming that this additive is a powerful inhibitor of the polyphenol oxidase (tyrosinase) present in the must of healthy grapes [9]. Supplementation with glutathione (GSH) also reduced the OCR but to a lesser extent than sulphur dioxide. This is probably because GSH reacts with the ortho-quinones formed by the action of polyphenol oxidases and thus their availability for oxidation is decreased [18,28]. Figure 1A also shows the OCR of all these samples in the presence of laccase. In general, laccase supplementation accelerated the OCR of the control and of the sample supplemented with GSH in comparison with their corresponding samples without laccase. In contrast, the samples supplemented with sulphur dioxide alone or in combination with GSH showed a similar behaviour to the corresponding samples without laccase. This confirms that sulphur dioxide is a very effective inhibitor not only of tyrosinase but also of laccase [7,10].
Figure 1B shows the OCR of the diluted grape must supplemented with inactivated dry yeasts rich in glutathione (IDY-GSH) in the absence and presence of laccase. This figure also shows the influence of sulphur dioxide and of the combination of the two treatments. In general, the observed behaviour for IDY-GSH in the absence of laccase was very similar to that reported for pure GSH. When laccase was present, the OCR of the control and especially that of IDY-GSH were also accelerated whereas the supplementation with sulphur dioxide reduced them a lot in a similar way as in the absence of laccase.
Figure 1 gives an idea of the OCR but cannot be used to establish statistically significant differences within the different experimental groups. Therefore, a previously reported kinetic model [31] was applied to statistically compare the results of this graph and calculate the total oxygen consumption (TOC) of the different samples. This model consists of representing the inverse of consumed oxygen versus the inverse of elapsed time. Using this mathematical model, the following equation can be established.
1/[O2] = A/t + B
In which [O2] is the concentration of consumed oxygen, A is the slope of the regression line, B is the point of intersection with the ordinate axis and t is the time. After clearing up the oxygen in the equation, the TOC can be calculated as the limit when time tends towards infinity.
Figure 2A shows the calculated TOC of the diluted grape must supplemented or not with sulphur dioxide, GSH and with the combination of the two additives in the absence or presence of 2 U laccase/mL.
This graph confirms that the TOC of the samples supplemented with sulphur dioxide, alone or combined with GSH, was significantly lower than their corresponding controls. Supplementation with GSH also significantly reduced the TOC but to a lesser extent than sulphur dioxide. No significant differences were found in the TOC between samples with and without supplementation with laccase. This indicates that laccase increased the initial OCR but not the total consumption of oxygen.
The supplementation with IDY-GSH produced very similar results to that of GSH in the absence of laccase since it also reduced the TOC significantly in comparison with the control (Figure 2B). However, in the presence of laccase, IDY-GSH did not reduce the TOC, which indicates that in these conditions it is not as effective as pure GSH for protecting grape must against oxidation.
3.2. Hydroxycinnamic acids and GRP
Figure 3A shows the hydroxycinnamic acids and GRP concentrations of the original grape must and the samples supplemented or not with sulphur dioxide, GSH and the combination of the two additives after oxygen consumption.
In the absence of laccase, the original grape must was very rich in hydroxycinnamic acids, especially in caftaric acid, whereas the concentration of GRP was practically nil. These data confirm that the grape must was very well protected against oxidation. In the absence of sulphur dioxide, the hydroxycinnamic acid concentration of the control grape must drastically decreased after the oxygen consumption without the appearance of GRP. This indicates that nearly all the hydroxycinnamic acids were oxidized to the corresponding ortho-quinones. The very low concentration of GRP would indicate that the original grape must was very poor in GSH. In contrast, when the grape must was supplemented with sulphur dioxide, the hydroxycinnamic acid concentration remained at similar levels to those in the original grape must, which confirms that this additive really inhibits the action of tyrosinase. This result is in accordance with the observed reduction in the oxygen consumption caused by sulphur dioxide previously described (Figure 1 and Figure 2). In contrast, hydroxycinnamic acids completely disappeared when the grape must was supplemented with GSH in the absence of sulphur dioxide, and the concentration of GRP increased very significantly. It therefore appears that almost all hydroxycinnamic acids were oxidized to ortho-quinones that were immediately converted to GRP upon reaction with GSH. When GSH and sulphur dioxide were added together some of the hydroxycinnamic acids were conserved and some GRP was formed. These results are more difficult to interpret since it seems that sulphur dioxide has inhibited the action of tyrosinase but to a lesser extent than when only sulphur dioxide was added because some of the hydroxycinnamic acids have been transformed into GRP.
Very similar results were obtained when the samples were supplemented with 2 U of laccase. The only remarkable difference was that the levels of GRP of the samples supplemented with GSH were lower, probably because laccase, unlike tyrosinase, can oxidize GRP [18].
Figure 3 B shows the hydroxycinnamic acids and GRP concentrations of the original grape must and the samples supplemented or not with sulphur dioxide, IDY-GSH and the combination of the two additives. In this case, the original grape must contained GRP and the level of total hydroxycinnamic acids was somewhat lower than in the previous experiment. These data would indicate that the grape must has undergone some oxidation since GRP is formed by adding GSH to the ortho-quinone formed by oxidation of caftaric acid [18,24]. When no sulphur dioxide was added, the GRP concentration was slightly but significantly lower than in the original grape must and the concentration of hydroxycinnamic acids was very low, which would indicate that tyrosinase has nearly completely oxidized these compounds. In contrast, when sulphur dioxide was present the levels of GRP and hydroxycinnamic acids were similar to those of the original grape must, which indicates that this additive exerted a protective effect against oxidation. When the grape must was supplemented with IDY-GSH, the GRP concentration was similar to that of the original grape must and the concentration of hydroxycinnamic acids was significantly lower than in the original grape must but higher than in the grape must without sulphur dioxide. Finally, the levels of GRP and hydroxycinnamic acids of the grape must supplemented with IDY-GSH and sulphur dioxide were very similar to those of the grape must supplemented only with sulphur dioxide. In general, in the presence of laccase the levels of GRP and hydroxycinnamic acids were lower than in its absence, confirming that this polyphenol oxidase is very effective at oxidizing these compounds [4,6,8].
3.3. Colour parameters
Figure 4A shows the absorbance at 420 nm (A420nm) and the CIEL*a*b* blue–yellow component (b*) of the samples supplemented with sulphur dioxide, GSH and their combination in the absence and presence of laccase as indicators of browning intensity.
The addition of SO2 exerted an evident protecting effect against browning because the values of A420 and b* were significantly lower than in the control sample, confirming again that this additive effectively avoids this problem [4,9,10]. The supplementation with GSH, alone or combined with sulphur dioxide, also significantly decreased A420 and b* with respect to the control, confirming that this antioxidant really protects grape must against browning even in the absence of sulphur dioxide.
In the presence of laccase, although the samples supplemented with sulphur dioxide and GSH showed significantly lower values of A420 and b* than the control, all of them had significantly higher values of the two parameters than their equivalent samples without laccase. These data confirm the high oxidative capacity of this polyphenol oxidase. It is also clear that it is more difficult to completely protect grapes affected by grey rot against browning, even using sulphur dioxide.
Figure 4B shows the equivalent results using IDY-GSH instead of pure GSH, and it can be seen that the results are very similar. Sulphur dioxide and IDY-GSH, alone or combined with sulphur dioxide, exerted a significant protective effect against browning in the absence of laccase; however, this protection was not so effective when the samples were supplemented with this enzyme. It seems, therefore, that sulphur dioxide and also GSH, pure or in the form of IDY-GSH, can prevent must from browning in healthy grapes but their effect is not efficient enough when the grapes are affected by the presence of laccase.
Figure 5 shows the total colour difference (ΔEab*) between the different samples.
It is generally accepted that when the ΔEab* between two samples of wine is lower than three units the human eye cannot distinguish between them [34]. The sample supplemented with sulphur dioxide was used as the reference to compare with all the other samples since this is the most usual procedure for protecting grape must from browning. Consequently, when the ΔEab* between one sample and this control reference is greater or lower than three units, it indicates whether the browning intensity can or cannot be perceived by the human eye.
The results clearly show the protective effect of the various antioxidants against browning. Figure 5A shows the ΔEab* between the samples supplemented with GSH, alone or in combination with sulphur dioxide, and the reference supplemented only with sulphur dioxide. In conditions of healthy grape must (absence of laccase) the ΔEab* of the control sample was clearly higher than three units and this value was much higher when laccase was present in the media. These data confirm that without sulphur dioxide the grape musts develop browning to a sufficient extent to be perceived by the human eye. In contrast, supplementation with GSH, alone or in combination with sulphur dioxide, allowed ΔEab* values to be kept below three units, which confirms that this antioxidant exerts a real protective effect.
However, when the samples were also supplemented with laccase, supplementation with GSH and even with sulphur dioxide was not effective enough to protect the samples against browning because the ΔEab* values were above three units.
When IDY-GSH was used instead of GSH (Figure 5B), the ΔEab* values showed a similar trend to those obtained for pure GSH (Figure 5A). IDY-GSH protected against browning (ΔEab* < 3 units) when no laccase was added; however, its protective effect was not enough when laccase was added (ΔEab* > 3 units).
Figure 6 shows the pictures of the different samples at the end of the experiment.
In these pictures, it can be clearly perceived that the control sample without any addition has undergone a clear browning process. In contrast, the supplementation with sulphur dioxide, GSH, IDY-GSH and the combination of sulphur dioxide with these two last additives really protect against browning in the absence of laccase since no visual differences in the browning intensity of the samples can be perceived by potential consumers. However, in the presence of laccase the protective effect of GSH and IDY-GSH are not enough to prevent browning.
4. Conclusions
In this work, we analysed how supplementation with sulphur dioxide, glutathione and a specific inactivated dry yeast rich in glutathione, influences oxygen consumption kinetics, hydroxycinnamic acids and GRP concentrations, as well as browning intensity in two situations. The first condition was grape must from healthy grapes, while the second was grape must supplemented with the enzyme laccase to reproduce what happens when the grapes are affected by grey rot. The results show that sulphur dioxide drastically reduces the oxygen consumption rate, protects hydroxycinnamic acids from oxidation and prevents grape must from browning even in the presence of laccase. The supplementation with glutathione, pure or in the form of a specific inactivated dry yeast rich in glutathione, also reduces the oxygen consumption rate but to a lesser extent than sulphur dioxide, it favours the formation of GRP from hydroxycinnamic acids, and effectively protects grape must against browning in conditions of healthy grapes. However, in the presence of laccase, glutathione was not effective enough to prevent browning. It can be concluded therefore that glutathione and the specific inactivated dry yeast rich in glutathione can be considered adequate alternative tools to sulphur dioxide for preventing browning in white grape must in conditions of healthy grapes.
Author Contributions
M.B.: methodology, investigation, writing – original draft, data curation; P.G.: methodology, research; A.J.B: methodology, research; I.S.C.: methodology, research; J.G.: methodology, research; J.M.H.: supervision; N.S.: supervision; M.G.: supervision; J.P.N.: supervision; S.G.A.: supervision; J.M.C.: supervision. F.Z.: supervision, methodology, research, writing - original draft, data curation.
Funding
This research was funded by the Spanish Ministry of Science and Innovation, Interministerial Commission for Science and Technology (CICYT - projects RTI2018-095658-B-C33 and PID2022-139868OB-C33).
Data Availability Statement
Data are contained within the article
Acknowledgments
The authors are thankful for the financial support provided by the European Union’s Horizon 2020 research and innovation programme under the Marie Skłodowska-Curie grant agreement No. 945413. They also thank Josep M. Alsina, Josep Anguera and Pedro Cabanillas for the support in the vineyard and winery.
Conflicts of Interest
Two of the authors, José M. Heras and Nathalie Sieczkowski, work for the company Lallemand which manufacture the used inactivated dry yeast Glutastar™ used in this experimentation. The other authors declare no conflict of interest.
References
- Li, H.; Guo, A.; Wang, H. Mechanisms of oxidative browning of wine. Food Chem. 2008, 108, 1–13. [Google Scholar] [CrossRef]
- Gancel, A.L.; Vignault, A.; Pilard, E.; Miramont, C.; Jourdes, M.; Fermaud, M.; Roudet, J.; Zamora, F.; Teissedre, P.L. Impact of added oenological tannins on red wine quality to counteract Botrytis infection in merlot grapes. Oeno One 2021, 55, 381–402. [Google Scholar] [CrossRef]
- Friedman, M. ; Food browning and its prevention: an overview. J. Agric. Food Chem. 1966, 44, 631–653. [Google Scholar] [CrossRef]
- Oliveira, C.M.; Ferreira, A.C.; De Freitas, V.; Silva, A.M. Oxidation mechanisms occurring in wines. Food Res. Int. 2011, 44, 1115–1126. [Google Scholar] [CrossRef]
- Fronk, P.; Hartmann, H.; Bauer, M.; Solem, E.; Jaenicke, E.; Tenzer, S.; Decker, H. Polyphenoloxidase from Riesling and Dornfelder wine grapes (Vitis vinifera) is a tyrosinase. Food Chem. 2015, 183, 49–57. [Google Scholar] [CrossRef]
- Claus, H.; Sabel, A.; König, H. Wine phenols and laccase: an ambivalent relationship. In Wine - Phenolic Composition, Classification and Health Benefits; Rayess, Y.E., Ed.; Nova Science Publishers: New York, NY, USA, 2014; pp. 155–185. [Google Scholar]
- Giménez, P.; Anguela, S.; Just-Borras, A.; Pons-Mercadé, P.; Vignault, A.; Canals, J.M.; Teissedre, P.L.; Zamora, F. Development of a synthetic model to study browning caused by laccase activity from Botrytis cinerea. LWT - Food Sci. Technol. 2022, 154, 112871. [Google Scholar] [CrossRef]
- Steel, C.C.; Blackman, J.W.; Schmidtke, L.M. Grapevine bunch rots: Impacts on wine composition, quality, and potential procedures for the removal of wine faults. J. Agric. Food Chem. 2013, 61, 5189–5206. [Google Scholar] [CrossRef] [PubMed]
- Ribéreau-Gayon, P.; Glories, Y.; Maujean, A.; Dubordieu, D. The use of sulphur dioxide in must and wine treatment. In Handbook of Enology: The Microbiology of Wine and Vinifications, 2nd ed.; John Wiley & Sons: Chichester, 2006; Volume 2, pp. 281–304. [Google Scholar]
- Ough, C.S.; Crowell, E.A. Use of sulphur dioxide in winemaking. J. Food Sci. 1987, 52, 386–389. [Google Scholar] [CrossRef]
- Du Toit, W.J.; Marais, J.; Pretorius, I.S.; du Toit, M. Oxygen in must and wine: A review. South African J. Enol. Vitic. 2006, 27, 76–94. [Google Scholar] [CrossRef]
- Stockley, C.S. Wine and health: sulfur dioxide and the wine consumer. Aust. N.Z. Grapegrow. Winemak. 2005, 501, 73–76. [Google Scholar]
- Vally, H.; Misso, N.L.A. Adverse reactions to the sulphite additives. Gastroenterol. Hepatol. Bed Bench 2012, 5, 16–23. [Google Scholar] [PubMed]
- Ribéreau-Gayon, P.; Glories, Y.; Maujean, A.; Dubordieu, D. Products and methods complementing the effect of sulphur dioxide. In Handbook of Enology: The Microbiology of Wine and Vinifications, 2nd ed.; John Wiley & Sons: Chichester, 2006; Volume 2, pp. 223–240. [Google Scholar]
- Martinez, M.V.; Whitaker, J.R. The biochemistry and control of enzymatic browning. Trends Food. Sci. Technol. 1995, 6, 195–200. [Google Scholar] [CrossRef]
- Vignault, A.; Pascual, O.; Jourdes, M.; Moine, V.; Fermaud, M.; Roudet, J.; Canals, J.M.; Teissedre, P.L.; Zamora, F. Impact of enological tannins on laccase activity. Oeno One 2019, 53, 27–38. [Google Scholar] [CrossRef]
- Vignault, A.; Gombau, J.; Jourdes, M.; Moine, V.; Canals, J.M.; Fermaud, M.; Roudet, J.; Zamora, F.; Teissedre, P.L. Oenological tannins to prevent Botrytis cinerea damage in grapes and musts: Kinetics and electrophoresis characterization of laccase. Food Chem. 2020, 316, 126334. [Google Scholar] [CrossRef] [PubMed]
- Kritzinger, E.C.; Bauer, F.F.; du Toit, W.J. Role of glutathione in winemaking: A review. J. Agric. Food Chem. 2013, 61, 269–277. [Google Scholar] [CrossRef] [PubMed]
- Zimdars, S. Role of glutathione and Botrytis cinerea laccase activities in wine quality. Cuvillier Verlag, Gotinga, 2020.
- Bahut, F.; Romanet, R.; Sieczkowski, N.; Schmitt-Kopplin, P.; Nikolantonaki, M.; Gougeon, R.D. Antioxidant activity from inactivated yeast: Expanding knowledge beyond the glutathione-related oxidative stability of wine. Food Chem. 2020, 325, 126941. [Google Scholar] [CrossRef]
- Gabrielli, M.; Aleixandre-Tudo, J.L.; Kilmartin, P.A.; Sieczkowski, N.; du Toit, W.J. Additions of glutathione or specific glutathione-rich dry inactivated yeast preparation (DYP) to sauvignon blanc must: Effect on wine chemical and sensory composition. South African J. Enol. Vitic. 2017, 38, 18–28. [Google Scholar] [CrossRef]
- Pascual, O.; Vignault, A.; Gombau, J.; Navarro, M.; Gómez-Alonso, S.; García-Romero, E.; Canals, J.M.; Hermosín-Gutíerrez, I.; Teissedre, P.L.; Zamora, F. Oxygen consumption rates by different oenological tannins in a model wine solution. Food Chem. 2017, 234, 26–32. [Google Scholar] [CrossRef] [PubMed]
- Bradshaw, M.P.; Prenzler, P.D.; Scollary, G.R. Ascorbic acid-induced browning of (+)-catechin in a model wine system. J. Agric. Food Chem. 2001, 49, 934–939. [Google Scholar] [CrossRef]
- Singleton, V.L.; Salgues, M.; Zaya, J.; Trousdale, E. Caftaric acid disappearance and conversion to products of enzymic oxidation in grape must and wine. Am. J. Enol. Vitic. 1985, 36, 50–56. [Google Scholar] [CrossRef]
- OIV, Resolution OIV-OENO 445-2015, Treatment of must with glutathione. 2015. Available online: https://www.oiv.int/public/medias/1686/oiv-oeno-445-2015-en.pdf (accessed on 9 July 2023).
- OIV, Resolution OIV-OENO 446-2015, Treatment of wine with glutathione. 2015. Available online: https://www.oiv.int/public/medias/1687/oiv-oeno-446-2015-en.pdf (accessed on 9 July 2023).
- OIV, Resolution OIV-OENO 603-2018, Monograph on inactivated yeasts with guaranteed glutathione levels. 2018. Available online: https://www.oiv.int/public/medias/6444/oiv-oeno-603-2018-en.pdf (accessed on 9 July 2023).
- Giménez, P.; Just-Borras, A.; Pons-Mercadé, P.; Gombau, J.; Heras, J.M.; Sieczkowski, N.; Canals, J.M.; Zamora, F. Biotechnological tools for reducing the use of sulphur dioxide in white grape must and preventing enzymatic browning: glutathione; inactivated dry yeasts rich in glutathione; and bioprotection with Metschnikowia pulcherrima. Eur. Food Res. Technol. 2023, 249, 1491–1501. [Google Scholar] [CrossRef]
- Grassin, C.; Dubourdieu, D. Quantitative determination of Botrytis laccase in musts and wines by the syringaldazine test. J. Sci. Food Agric. 1989, 48, 369–376. [Google Scholar] [CrossRef]
- Diéval, J.B.; Vidal, S.; Aagaard, O. Measurement of the oxygen transmission rate of co-extruded wine bottle closures using a luminescence-based technique. Packag. Technol. Sci. 2011, 24, 375–385. [Google Scholar] [CrossRef]
- Pons-Mercadé, P.; Anguela, S.; Giménez, P.; Heras, J.M.; Sieczkowski, N.; Rozès, N.; Canals, J.M.; Zamora, F. Measuring the oxygen consumption rate of some inactivated dry yeasts: Comparison with other common wine antioxidants. Oeno One 2021, 55, 147–158. [Google Scholar] [CrossRef]
- Ayala, F.; Echavarri, J.F.; Negueruela, A.I. A new simplified method for measuring the color of wines. ii. white wines and brandies. Am. J. Enol. Vitic. 1997, 48, 364–369. [Google Scholar] [CrossRef]
- MSCVes.zip. 2013. Available online: http://www.unirioja.es/color/descargas.shtml (accessed on 9 July 2023).
- Martínez, J.A.; Melgosa, M.; Pérez, M.M.; Hita, E.; Negueruela, A.I. Visual and Instrumental Color Evaluation in Red Wines. Food Sci. Technol. Int. 2001, 7, 439–444. [Google Scholar] [CrossRef]
- Lago-Vanzela, E.S.; Da-Silva, R.; Gomes, E.; García-Romero, E.; Hermosín-Gutiérrez, I. Phenolic composition of the Brazilian seedless table grape varieties BRS clara and BRS morena. J. Agric. Food Chem. 2011, 59, 8314–8323. [Google Scholar] [CrossRef]
Figure 1.
Oxygen consumption kinetics. Results are expressed as mean ± standard deviation of three replicates. GSH: Glutathione; IDY-GSH: Specific inactivated dry yeasts rich in glutathione.
Figure 1.
Oxygen consumption kinetics. Results are expressed as mean ± standard deviation of three replicates. GSH: Glutathione; IDY-GSH: Specific inactivated dry yeasts rich in glutathione.
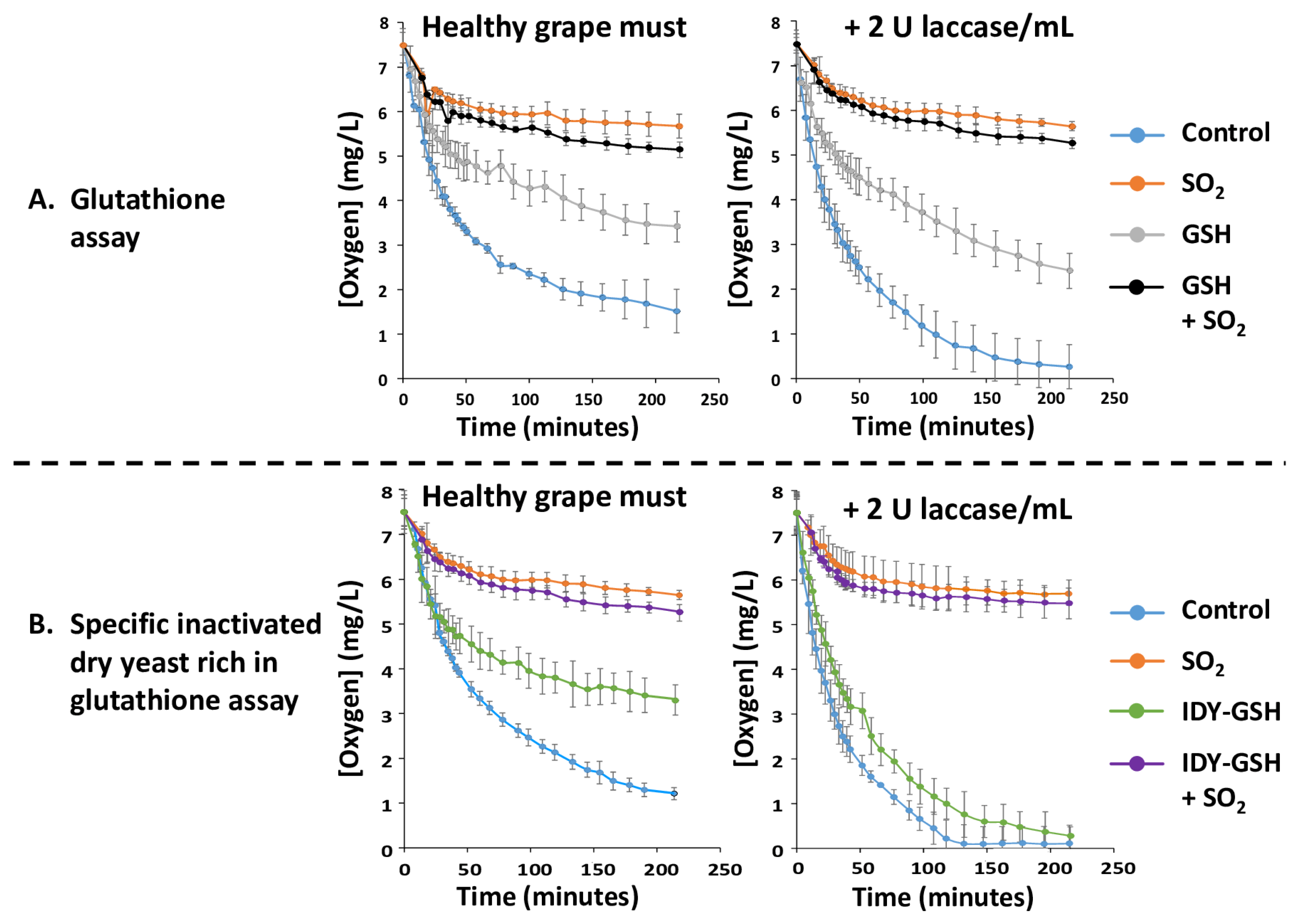
Figure 2.
Total oxygen consumption (TOC). Results are expressed as mean ± standard deviation of three replicates. TOC: Total oxygen consumption; Glutathione; IDY-GSH: Specific inactivated dry yeasts rich in glutathione. Different letters indicate the existence of significant differences (p < 0.05).
Figure 2.
Total oxygen consumption (TOC). Results are expressed as mean ± standard deviation of three replicates. TOC: Total oxygen consumption; Glutathione; IDY-GSH: Specific inactivated dry yeasts rich in glutathione. Different letters indicate the existence of significant differences (p < 0.05).
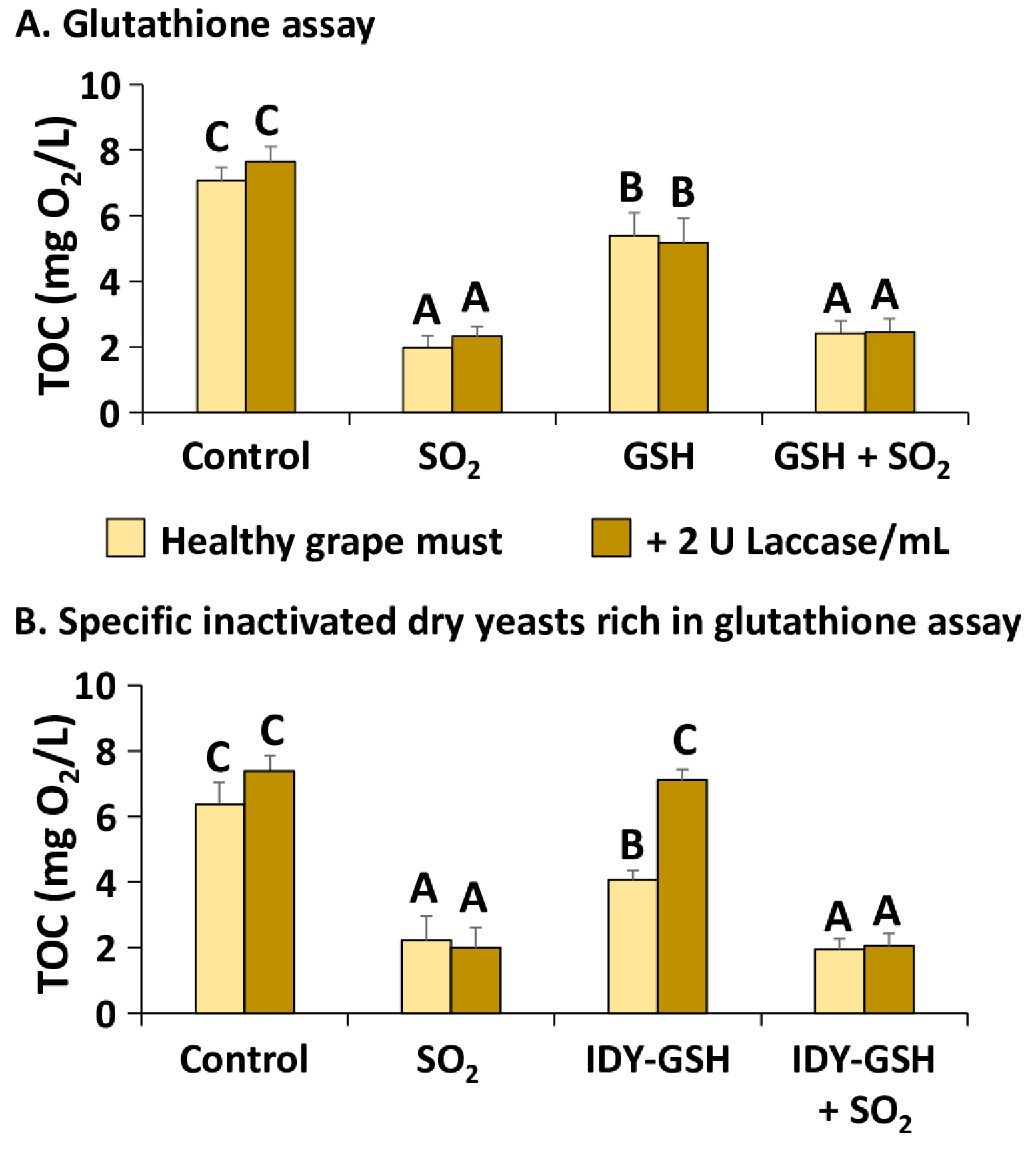
Figure 3.
Hydroxycinnamic acids and GRP. Results are expressed as mean ± standard deviation of three replicates. GSH: Glutathione; IDY-GSH: Specific inactivated dry yeasts rich in glutathione. Different letters indicate the existence of significant differences (p < 0.05).
Figure 3.
Hydroxycinnamic acids and GRP. Results are expressed as mean ± standard deviation of three replicates. GSH: Glutathione; IDY-GSH: Specific inactivated dry yeasts rich in glutathione. Different letters indicate the existence of significant differences (p < 0.05).
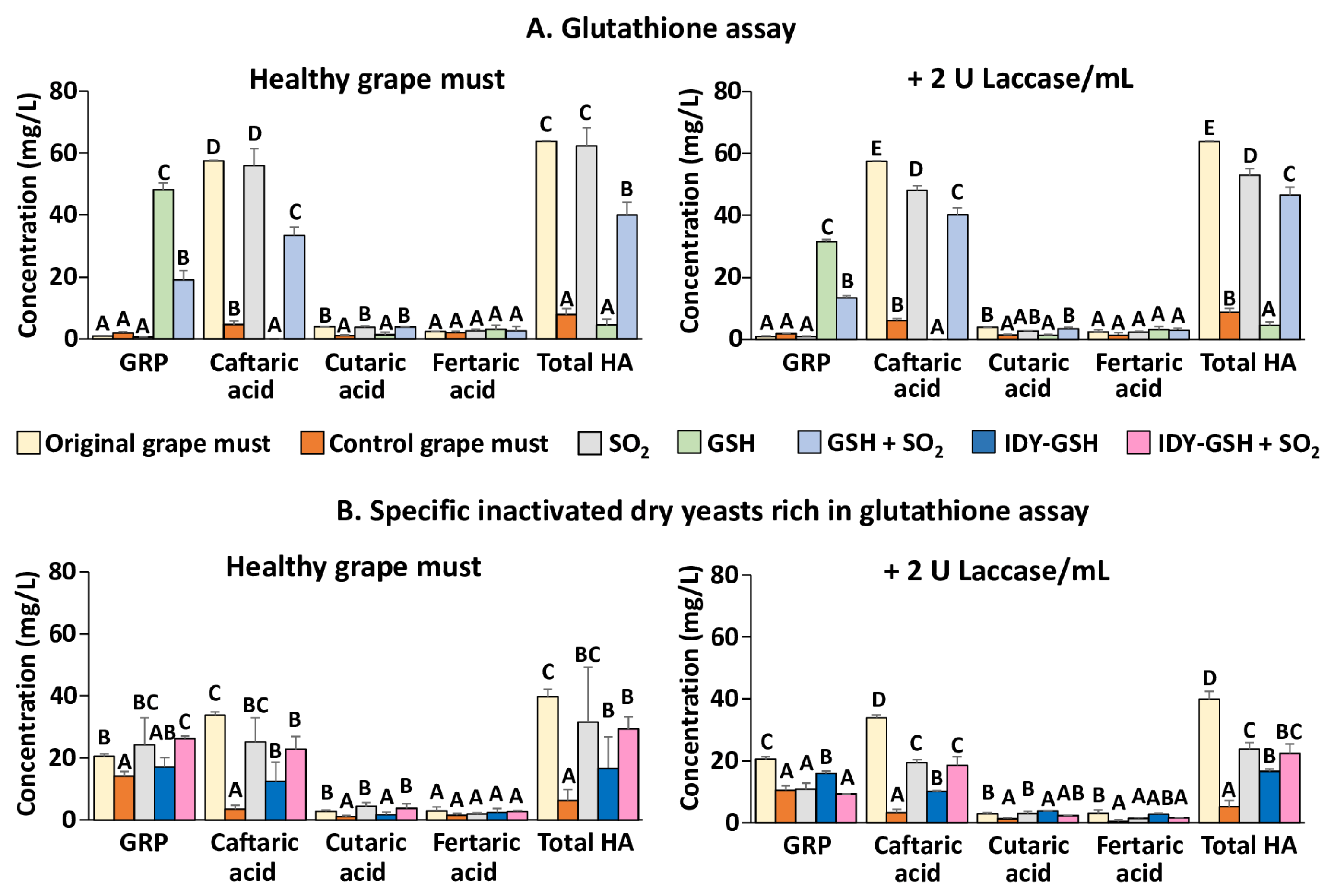
Figure 4.
Browning intensity. Results are expressed as mean ± standard deviation of three replicates. A420nm: Absorbance at 420 nm; b*: CIEL*a*b* colour’s blue–yellow component; GSH: Glutathione; IDY-GSH: Specific inactivated dry yeasts rich in glutathione. Different letters indicate the existence of significant differences (p < 0.05).
Figure 4.
Browning intensity. Results are expressed as mean ± standard deviation of three replicates. A420nm: Absorbance at 420 nm; b*: CIEL*a*b* colour’s blue–yellow component; GSH: Glutathione; IDY-GSH: Specific inactivated dry yeasts rich in glutathione. Different letters indicate the existence of significant differences (p < 0.05).
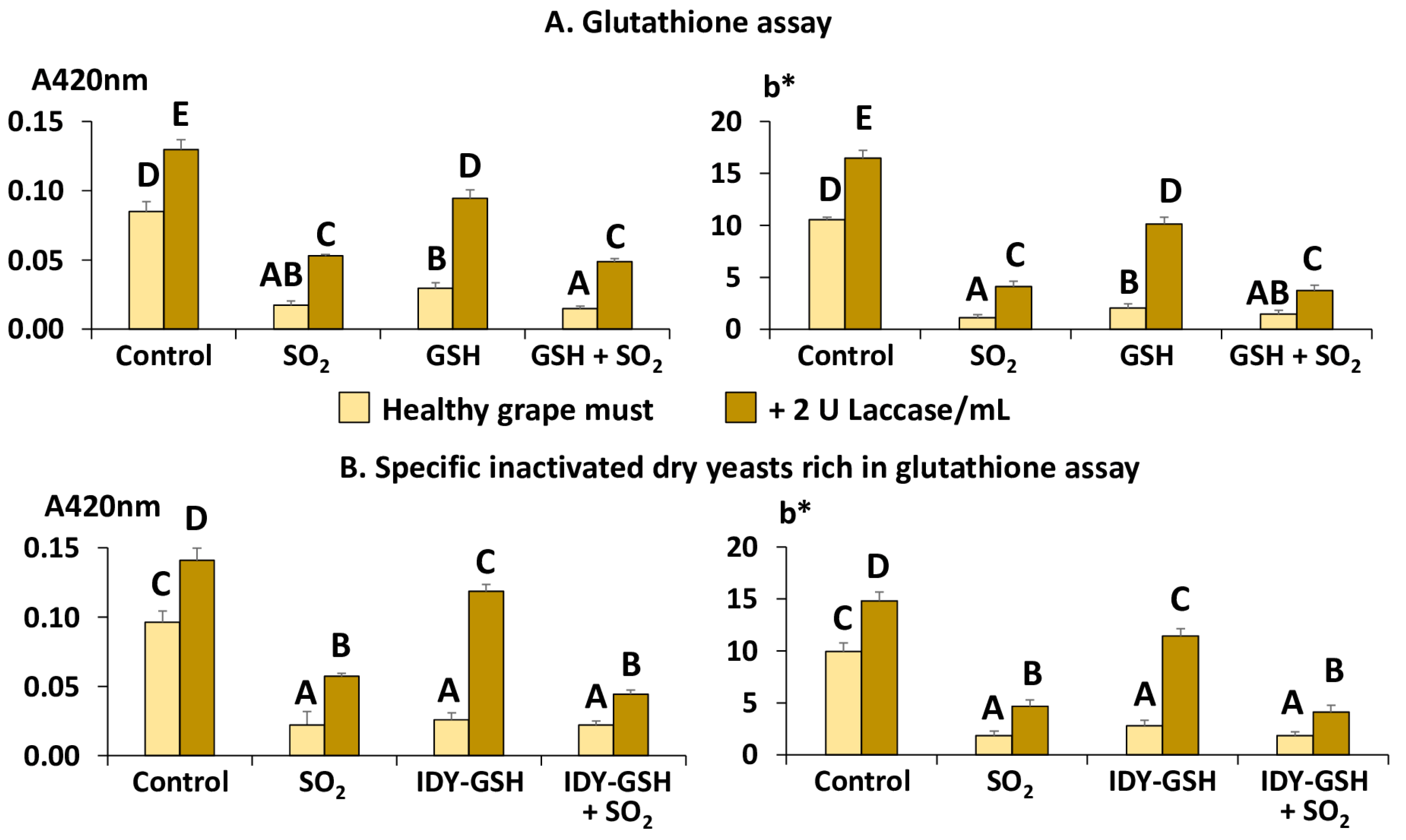
Figure 5.
Total difference of colour (ΔEab*). Results are expressed as mean ± standard deviation of three replicates. GSH: Glutathione; IDY-GSH: Specific inactivated dry yeasts rich in glutathione.
Figure 5.
Total difference of colour (ΔEab*). Results are expressed as mean ± standard deviation of three replicates. GSH: Glutathione; IDY-GSH: Specific inactivated dry yeasts rich in glutathione.
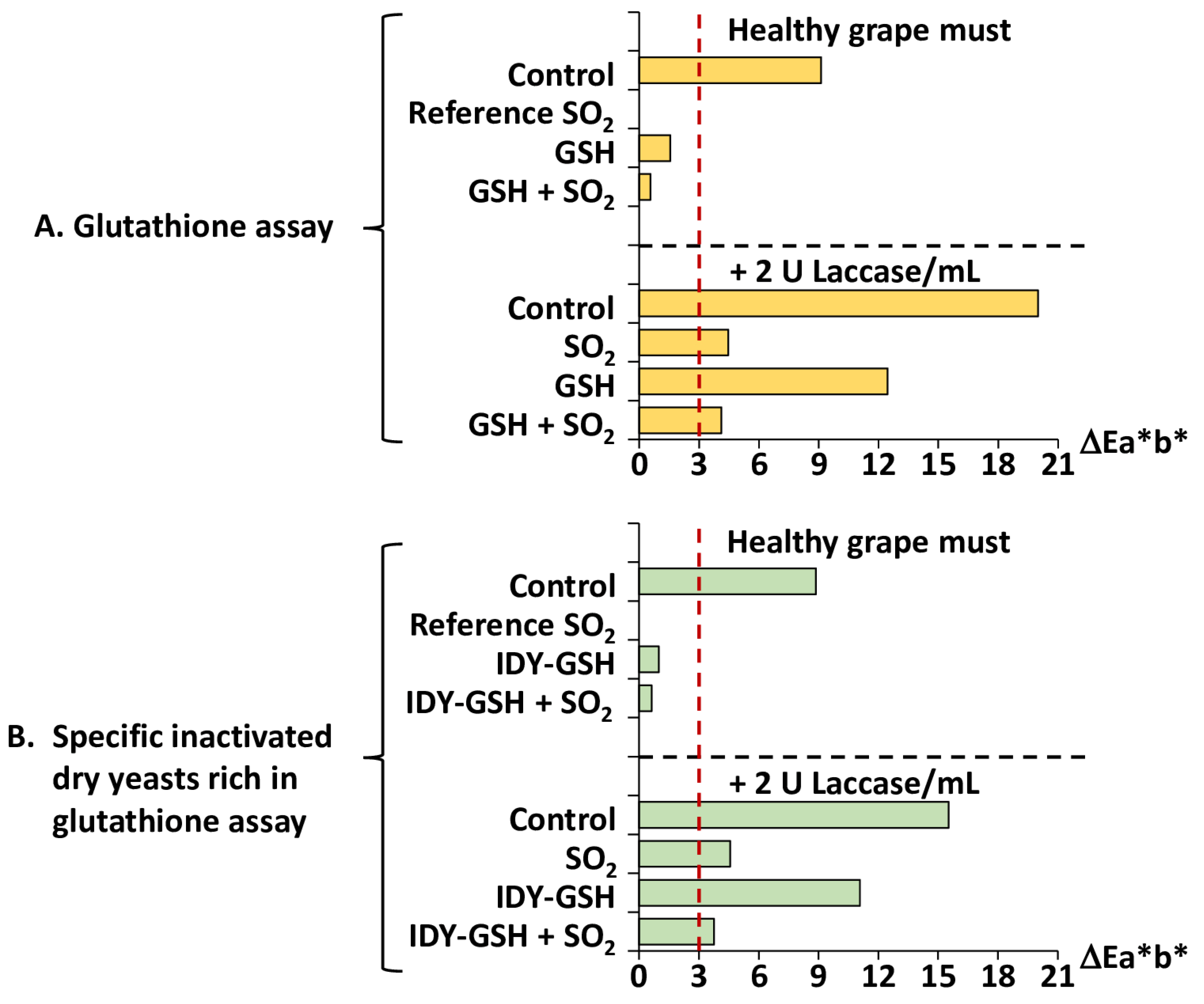
Figure 6.
Influence of laccase, SO2, glutathione and a specific inactivated dry yeast rich on glutathione on the final colour of the samples. GSH: Glutathione; IDY-GSH: Specific inactivated dry yeasts rich in glutathione.
Figure 6.
Influence of laccase, SO2, glutathione and a specific inactivated dry yeast rich on glutathione on the final colour of the samples. GSH: Glutathione; IDY-GSH: Specific inactivated dry yeasts rich in glutathione.

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



