
Submitted:
06 December 2023
Posted:
07 December 2023
You are already at the latest version
Abstract
Woody and non-woody plant species conservation is one of the ecosystem services provided by agroforestry (AF) systems across the agricultural landscapes. Little attention has been paid in as-sessing the plant diversity conservation of AF systems. The study was, thus, aimed to investigate plant species diversity, structure, and composition of three AF systems in Gedeo zone of south eastern Rift–valley Agricultural landscape of Ethiopia. The study was conducted in three agroforestry systems, namely, enset based, coffee–enset based (C-E based AF) and coffee-fruit trees–enset based (C-Ft-E based AF) agroforestry systems. Twenty farms representative of each AF system were randomly selected and inventory of the floristic diversity was employed in a 10×10 meter sample plot per farm. A total of 52 perennial woody and non–woody plant species belonging to 30 families were recorded. Of all species identified 33 (63.5%) were native, of which two species were registered as endemic. The highest proportion of native species was recorded in enset based AF (93.3%) and the least were in C–Ft–E based AF (59%). According to IUCN Red List and local criteria, 13 species were recorded as of interest for conservation in all AF systems. The woody species Prunus africana was identified as both vulnerable by IUCN Red List and rare for 25% of species that least occurred. The Shannon diversity index and richness showed that C–Ft–E based AF systems were significantly different from the two remaining AF systems. However, the species abundance and evenness did not show significant differences between the three AF systems. In general, retaining such number of woody and non–woody perennial plant species richness under AF systems of the present study underlines their potential for biodiversity conservation.
Keywords:
plant diversity
; indigenous agroforestry system
; coffee
; enset
; native species
1. Introduction
Deforestation and forest degradation has been a common issue and continued as a challenge in many parts of the world despite some initiatives to reforest these degraded areas [1]. The consequences of such resource degradation exert and exacerbate a significant negative impact on our globe and thus harming human lives for instance biodiversity loss, global warming and climate change [2]. To reduce such negative impacts adoption of Agroforestry (AF) system could be one option. Before mentioning the benefits of adopting AF system it is good to define what AF system mean. Although different authors tried to define the definition given by ICRAF was the most comprehensive and clear. Therefore, Agroforestry is defined as “an ecologically based natural resource management system that integrates trees (for fibre, food and energy) with crop and/or animal on farms with aim of diversifying and sustaining income and production while maintaining ecosystem services” [3].
As reported by several studies, AF systems have multiple contributions to biodiversity conservation through: arranging and providing additional supportive habitats for species that do not tolerate high levels of disturbance [4]; by conserving gene pools of native tree species in fragmented landscapes and conserving and enhancing biodiversity [5,6,7]; playing an important role in increasing microbial, avian and faunal diversities [8]; soil conservation and allowing water to recharge, thereby preventing habitat degradation and habitat loss; protecting against the pressure on forest degradation and deforestation in the surrounding natural habitat; and construction of a corridor and stepping stones for perseverance of floral and faunal species through connecting different fragmented habitats in the landscape [4,9,10]. Moreover, AF systems can help to create corridors of habitat that can connect fragmented areas of natural habitat. This can help to reduce the impact of habitat fragmentation and improve the movement of species [11]. In general, the native and non-native perennial woody and non–woody species could provide several functions: economic benefit such as source of firewood, timber, wood for different purposes (local construction, farm implements, and house-hold utensils), fodder, food, medicine; environmental benefits such as erosion control and soil fertility improvements and finally ecological improvements such as biodiversity conservation [12,13].
Even-though AF systems have the above mentioned multiple benefits there are also challenges associated to effective implementation of the systems. For instance: 1. Limited access to land: In many parts of the world, land is a scarce resource, making it difficult for farmers to implement AF systems. 2. Lack of knowledge and training: Many farmers lack the knowledge and skills needed to implement AF systems, making it difficult to adopt the technology. 3. Limited access to financial resources: AF systems require significant investments in terms of land, labor, and capital: Many farmers lack access to the necessary financial resources. 4. Pest and disease management: Pests and diseases can limit the success of AF systems, as they can cause damage to crops and trees. 5. Climate change: Climate change can affect the success of AF systems, as it can cause changes in temperature, precipitation, and other environmental factors. 6. Socio-economic factors: Socio-economic factors such as poverty, gender, and cultural norms can limit the success of AF systems. Therefore, if these problems are well addressed the AF systems could be implemented effectively.
Different AF systems show different diversity status based on their richness, abundance and frequency of plant species [12]. Considering the number of plant species as measure to categorize the species richness status in different AF systems of tropical and sub-tropical countries, indigenous AF systems have the highest number of species and followed by coffee systems, tree–crop systems and cocoa systems. Different management practices in each AF system may result in difference in species richness among these AF systems [14]. The four tropical AF systems with the highest recorded number of plant species are: (1) homegardens in west Java, Indonesia, (2) homegarden of Chagga, at the border of Tanzania and Kenya, (3) trees on agricultural land on Mount Kenya, and (4) traditional homegardens, in South west Bangladesh [15,16,17].
There are also some studies conducted in Ethiopia focusing on the contribution of AF systems as refuge for plants species. The reports indicated that there are between 17 (mainly in fruit tree AF systems) and 429 (in various AF systems) plant species growing and therefore, the systems are very important in biodiversity conservation of the native species [14]. Species richness in Ethiopia showed a variation among the different AF practices and regions of the country. For instance, the highest plant species richness was reported in southern Ethiopia ranging from 50–198 plant species [18,19,20] followed by south-west Ethiopia with 149 plant species [21], Central Ethiopia from 27–114 plant species [22,23,24] and the least was recorded in north Ethiopia with 17–40 plant species [25,26]. Reports specifically from south-eastern Ethiopia revealed that the indigenous AF systems of the region are rich in plant species diversity. Accordingly, [23] reported that 90 woody species including native tree species such as Juniperus procera Hochst. ex Endl., Olea europaea subsp. cuspidata (Wall. ex G. Don) Cif., Podocarpus falcatus (Thunb.) R. Br. ex Mirb., Acacia tortilis (Forssk.) Hayne, Acacia etbaica Schweinf., and Hagenia abyssinica J.F. Gmel.
AF practice in Gedeo zone of southern Ethiopia is known as ancient and indigenous in nature. The Gedeo agroforestry system has been in place for centuries, and is an important part of the local culture and economy [27]. Gedeo agroforests are also a well–known land-use system and are believed to have self-sustaining and self-regulating attributes compared to other land-use system in the area [28]. The systems are characterized by mixture of trees, shrubs, and annuals that grow in a diverse form and looks like intact and continuous vegetation cover [28]. Due to this reason, the practice is known to be an exemplary land-use system and ideal AF demonstration site in the region.
Previous studies have been conducted [13,28,29,30,31,33,34] on management of indigenous AF practices, component interaction, diversity and composition, carbon stocks and additional ecosystems services of AF systems in Gedeo zone and other locations. However, studies dealt with the contribution of enset based, coffee–enset (C–E) based and coffee–fruit tree-enset (C–Ft–E) based AF systems to biodiversity conservation in the study area are found to be limited. To fill the gap in scientific knowledge about the function and structure of these systems, and their exact extent more attention is needed.
The overall objective of this study was to assess the contribution of enset based, coffee–enset (C–E) based and coffee–fruit tree–enset (C–Ft–E) based AF systems to biodiversity conservation. The specific objectives were (i) to assess and evaluate the woody and non woody plant species richness in three indigenous AF systems (ii) to compare woody and non–woody perennial plants composition, structure and diversity among three AF systems (iii) to identify endemic native woody plant species and species of conservation concern (iv) to assess the effect of diameter at breast height (DBH) and stem number on basal area of the AF systems. Research questions were initiated to answer: a) Could species diversity differ among the three AF systems owing different management system? b) Do the AF systems conserve significant number of woody and non–woody plant species? c) How do indigenous agroforestry systems contribute to maintain native, endemic species and species of conservation concern? The findings of the present study will help to managers, conservationists and scientists to better understand the functional complexity of multistrata AF systems and the diversity status of the species that require local genetic diversity conservation.
2. Materials and Methods
2.1. Study area and sites
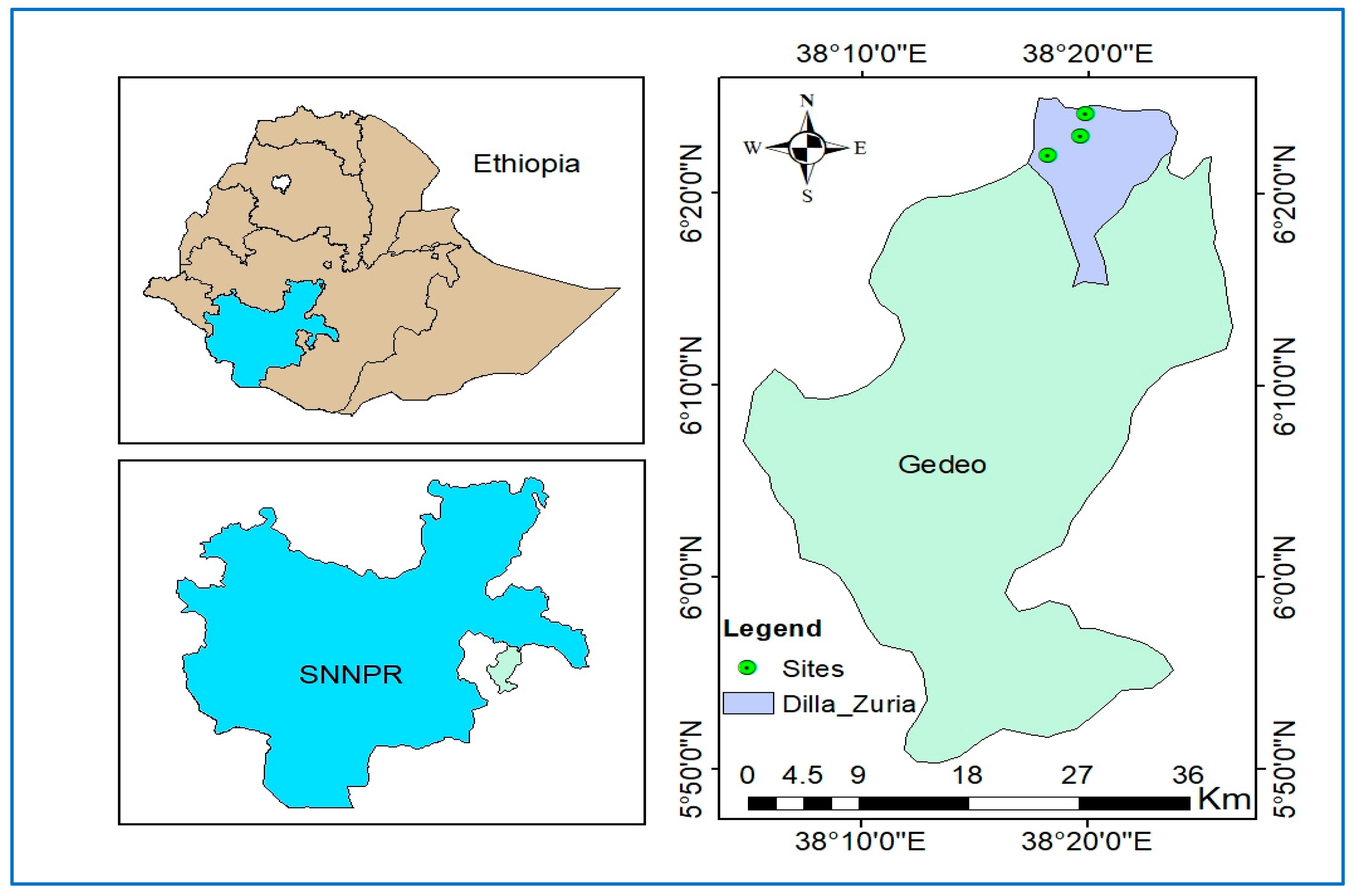
The study was conducted in the Gedeo zone of the Southern Nations, Nationalities, and Peoples’ Regional State of Ethiopia (SNNPRs), more specifically in the Dilla Zuria district (Figure 1). Geographically, it lies between 38° 03’ 02” and 38° 18’ 59” E longitude and 5° 50’ 26” and 6° 12’ 48” N latitude. The research area’s elevation varies from 1544 to 1830 meters above sea level. The mean monthly air temperature ranges from 13 to 28 0C, while the annual rainfall ranges from 1127 to 1624 mm, according to meteorological data collected over a period of nine years [35]. The Gedeo zone has a total area of 13,4700 ha, which is divided into various land-use types, including agricultural land (94.5%), AF land used for perennial and annual crops, grassland (1.4%), wetland (0.8%), natural forest (0.5%), plantations (0.1%), and others (pasture land, residential) (2.7%) [36]. According to the traditional agro climatic categorization, the zone has a mid-altitude climate (Dega), which makes up 37% of the total area, a subtropical climate (Weynadega), which makes up 62% of the total area, and a hot tropical climate (Kolla) for the remaining fraction of area. The soil type is nitosol having a deep, reddish-brown color with relatively high organic matter content. Theses soils are well-drained and fertile [37]. The texture of the soil is predominantly clay. The main economic activity of farmers in the study area is AF based food and cash crops production. AF is the primary source of income for farmers in the study area where coffee and fruit production is predominate ones. Livestock husbandry is insignificant in the system except few attempts in fattening of oxen, goats, and sheep. The three main AF systems practiced in the study area are 1) enset based AF system (mainly practiced in Sisota site), 2) coffee–enset based AF system (practiced in Golla site), and 3) coffee–fruit trees–enset based AF system (practiced in Chichu site).
Figure 1.
Locations of the study sites in the Dilla Zuria district of the Gedeo zone, which lies between 38° 03’ 02” and 38° 18’ 59” E longitude and 5° 50’ 26” and 6° 12’ 48” N latitude, in Ethiopia.
Figure 1.
Locations of the study sites in the Dilla Zuria district of the Gedeo zone, which lies between 38° 03’ 02” and 38° 18’ 59” E longitude and 5° 50’ 26” and 6° 12’ 48” N latitude, in Ethiopia.
2.1.1. Three indigenous agroforestry systems as focus of the study
- Enset based agroforestry
Enset based AF system is predominantly practiced in Central, south-western, and southern Ethiopia [38,39]. Depending on the distribution of other plant species, enset (Ensete ventricosum) known as false banana, belong to Musaceae family could cover 60–70% of the total area in this system. This perennial species is local and has been tamed as one of staple food crops in south, central and southwestern Ethiopia. The edibile parts of the enset plant include pseudo stem and underground corm. The growth habit of the plant could be either monocrop plantation or mixed with other crops with a rotation period of 3 to 15 years [40,41]. An estimated area of enset cover in Ethiopia to nearly 300,000 ha, yielding approximately 4.4 million tons per annum and feeding approximately 20% of the total population of the country [42,43,44].
Enset not only provides economic benefits to the farmers but it also gives a significant environmental service. For instance micro–climate amelioration, addition of nutrients through litterfall and hence improving the soil fertility, protection of the soil from water erosion and runoff hazard are some of the environmental services. One of the good things related with nutrient addition to the soil is, when enset is harvested most of the biomass remains in the system. This is because the leaves and all the other inedible part of the enset corm are returned directly to the soil [45,46].
Figure 2.
Enset based AF system of South-eastern Rift-valley Landscapes, Ethiopia.

- Coffee–enset based agroforestry (C–E based AF)
One of the traditional agroforestry homegardens in Southern Ethiopia is the C–E based AF system [39]. The technology can be effectively used at elevations of 1,500 to 2,300 meters above sea level. It is suggested that the temperature and moisture levels at this height will be favorable for these agroforestry techniques. In Southern Ethiopia’s agroforestry systems, the two dominant native perennial crops, notably coffee and enset, collectively occupy more than 60% of the available land [33]. Enset is a staple food crop and coffee serves as cash crop [33]. According to the National Bank of Ethiopia report the share of coffee in the country’s total merchandise export revenue is 25.1% [47]. The majority of export revenue comes from coffee which means coffee is playing a great role in getting hard currency for the country [48]. Due to their significant socioeconomic and ecological advantages, enset and coffee could be considered as “key-stone” species for the region’s farming system.
C–E based AF systems harbor several native woody species (Cordia africana, Millettia forginea, roots (ginger, sweet potato) and annual crops (maize) favorably growing in intimate association with enset and coffee. The perennial woody species are growing in spatial and vertical configurations in this AF system. Generally, these indigenous C-E based AF are all inclusive farm systems from which households get most of their subsistence as well as cash needs [18]. The average size of C–E based AF as homegarden in Southern Ethiopia is estimated to approximately 0.7 hectares per farmer. These systems can support a population of 500-1000 persons per km2 [49]. Like other AF systems, this system also maintains high species diversity which combines crops, trees and animals having different uses and production cycles.
- Coffee–fruit tree –Enset based agroforestry(C–Ft–E based AF)
Components of this AF system include multifunctional tree and shrub species, coffee (C. arabica L.), enset, several fruits, annual crops, vegetables, medicinal plants, and animal species. The majority of this sort of indigenous AF system is made up of coffee, fruit trees, and enset, with the remainder consisting of vegetables and spices [50]. For each species, the percentage of coffee, fruit trees, and enset is anticipated to range between 20 and 25. In C–Ft–E based AF system the fruit trees (e.g., Persea americana Mill., Mangifera indica L., Casimiroa edulis Lal Llave and Lex.), coffee and enset are shaded by tree species such as Cordia africana, Milletia ferruginea, Ficus Vasta and Ficus sur. Herbaceous plants such as Zea mays L., Musa spp., Brassica oleracea L., and Ipomoea batatas (L.) Lam make up the understory [51]. In one season of the year, four to six plant species often functioned as a source of revenue and dietary variety. These indigenous C–Ft–E based AF systems are all inclusive farm systems from which households get almost all their subsistence as well as cash needs [18]. Communities practicing this type of AF system were self–sufficient in wood for energy and generated 47% and 45% of their annual income from fruit trees and coffee, respectively [50].
The illustration of the coffee–enset based AF system and coffee–fruit tree–enset based AF system is displayed below.
Figure 3.
Coffee–enset based AF system of South-eastern Rift-valley Landscapes, Ethiopia.

Figure 4.
Coffee–fruit tree–enset based AF system of South-eastern Rift-valley Landscapes, Ethiopia.
Figure 4.
Coffee–fruit tree–enset based AF system of South-eastern Rift-valley Landscapes, Ethiopia.

2.2. Methods
2.2.1. Sampling design and data collection
We compared three different AF types’ floristic content, structure, and diversity. This was done to allow for a more accurate assessment of the relative capacity of agroforests with different crop combinations to conserve biodiversity along altitudinal gradients. A total of 60 AF farms (20 farms from each AF) were randomly chosen from lists of farms along the altitudinal gradients. Additionally, information on the site history, height, slope, GPS location, AF type, and age of each AF farm were recorded.
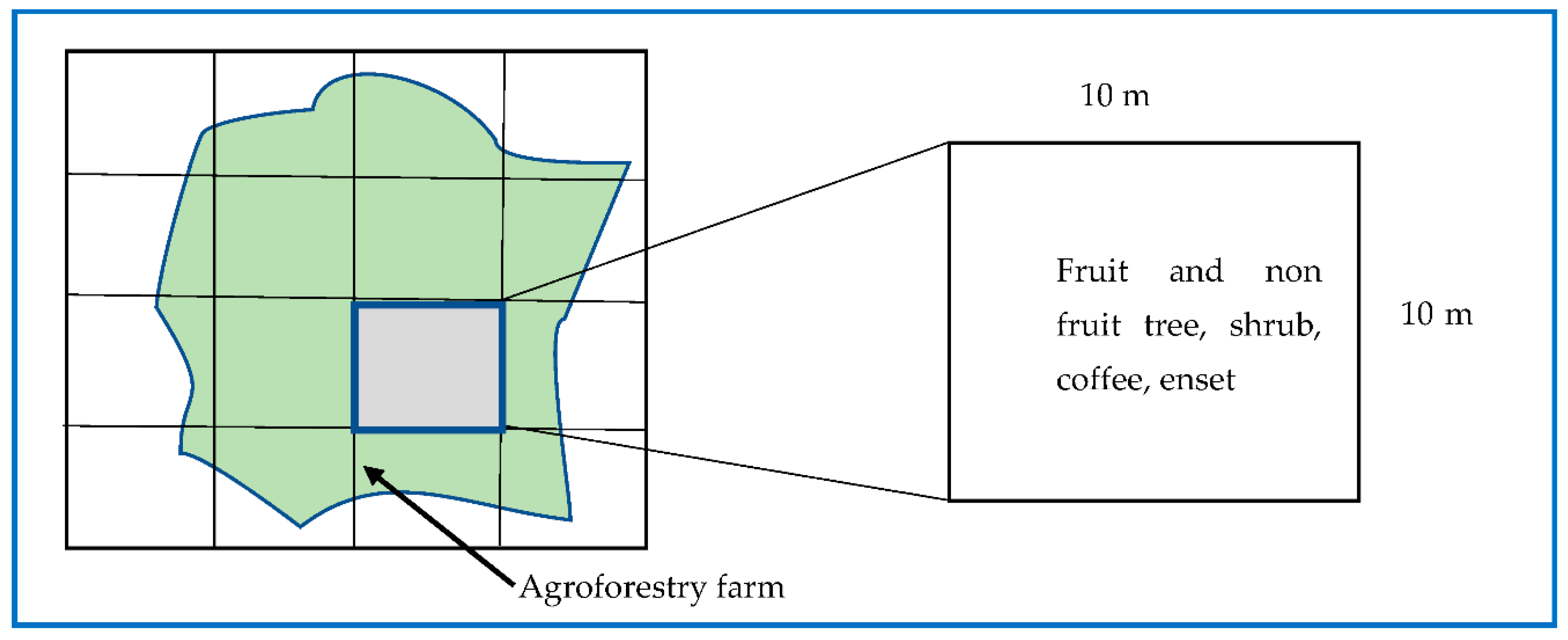
A nested quadrat with 10×10 m size was established in each AF farm for the inventory of trees/shrubs, coffee and enset. Ocular estimate was first employed to divide the farm into equal sections in order to find the location of a quadrat’s center. Second, a number was given to every component. Third, a data collecting plot was chosen using a random number generator. The size of the quadrats and sampling size coincide with recommended practice in literature for similar agroforestry farms by [14]. In some cases the size of the quadrant might occupy the whole farm. Because of the small size of some farms, cost, and time-related issues, the size of the quadrant was limited to 100 m2.
Figure 5.
Sample plot layout for inventory of trees, shrubs, coffee and enset plants (10×10 m) See 2.2.2 for detail.
Figure 5.
Sample plot layout for inventory of trees, shrubs, coffee and enset plants (10×10 m) See 2.2.2 for detail.
2.2.1. Sampling design and data collection
To assess the plant species diversity and composition an inventory of all the trees (fruit and non-fruit), coffee and enset plants was carried out. Inventory of non-fruit trees and shrubs, fruit trees, enset and coffee plants were done for the 60 farms (20 farms from each of the three AF systems). Inventory of species which exist out of the 60 sample plots but still in the selected study sites were also carried out to assess the total number of perennial woody and non–woody plant species. Measurements such as diameter at breast height (DBH, cm ±0.1), total height (h, m ±0.1) of all trees and shrubs (single and multi-stemmed) having a breast height diameter ≥ 2.5 cm and height ≥ 1.5 m were employed in the 10×10 m sample plots on each AF farm [13]. For coffee plants (in enset–coffee and C–Ft–E based AF systems), the stem diameter at stump height (40 cm), d40, was also measured. For enset based AF systems the basal diameter of the pseudostem (height of 10 cm, d10) of plants one year old or older was measured. Stem diameter (d, d10 and d40) was measured using Caliper in two perpendicular directions and the average value was used. In the studied AF systems biodiversity analysis was done by counting all woody species above 20 cm height and species were identified by using identification keys and local informants [14]. The reason why we used 20 cm as minimum height is because plant species above this height are easily identifiable and countable.
2.3. Data analysis
2.3.1. Stand characteristics of plant species and diversity analysis
Tree and stand characteristics were calculated for each agroforestry sample plot in the three studied AF systems. Parameters such as relative frequency, relative abundance, relative dominance, DBH, height, basal area (BA) and stem numbers of the perennial woody and non–woody plant species were determined. One way ANOVA was carried out to test differences among the three AF systems followed by Post hoc testing by means of the LSD test (Fisher’s LSD test). Diversity indices (Shannon diversity index, Pielou’s evenness index and Margalef’s richness index) were also calculated. This comparison helped to evaluate the mean differences among one AF systems [52]. Levene’s test was also conducted to check homogeneity of variances. For assessing the relationship between some parameters regression analysis was employed. For multi-stemmed plants mainly in the case of coffee plant (2 to 12 stems per plant) and Mango plant (2–4), each stem was measured and the equivalent diameter of the plant was calculated as the square root of the sum of squared diameters of all stems per plant [53].
where de (cm) = equivalent diameter at breast height, de40 (cm) = equivalent diameter at 40 cm height, di= sum of all squared diameters up to the ith stem
The Shannon diversity index [54,55], Pielou’s evenness index (J) and Margalef’s richness index (Dmg) [56] were calculated for each plot. Sorensen’s similarity coefficient determined the similarity/dissimilarity between AF systems. Species richness and abundance followed by Mann–Whitney U–test for multiple comparisons. The following formulas were used for the diversity indices.
where, H’ = Shannon Diversity Index, Pi = the abundance of ith species expressed as a proportion of total cover [54,56]. As [13] mentioned, the Shannon diversity index is more preferred to know plant species diversity due to its sensitivity to sample size. It also gives more weight in assessing rare plant species.
where: J= Pielou’s evenness (Equitability) [57], H’= Shannon diversity index and H’max= ln S where “S” is the number of species. J has values between 0 and 1.0, where 1.0 represents a situation in which all species are equally abundant.
where DMg is Margalef’s richness index, S = species richness, N= the total number of individuals in the plot
where: Ss= Sorensen’s similarity coefficient, a=number of species common to both samples, b=number of species in sample 1 but not in 2, c=number of species in sample 2 but not in 1
Another very crucial index which is used to investigate the structural role of each plant species in the sampling plots is the important value index percentage (IVI %). It was calculated using the percentage of relative abundance (RA), relative dominance (RD.) and relative frequency (RF). Therefore, to investigate the importance value index (IVI %) of each species we used the following formulas.
where
IVI (%) = Relative abundance + Relative dominance + Relative frequency,
2.3.2. Analysis of species conservation concern
Assessing the status of species conservation concern in order to sustainably maintain the plant species in the studied AF systems is crucial. Species conservation concern (rare, threatened, vulnerable, least concern) of each studied AF types was identified and then recorded their level of conservation. Geographical distribution, habitat preference and population size are the main factors to take into account and then to classify if species are rare [58]. To analyze species conservation concerns in our study three approaches were used: (i) those woody or non-woody species retained in the different agroforestry systems and listed as of least concern, threatened/vulnerable by IUCN Red Lists [59,60]; (ii) 25% of species that have the least occurrence in each AF type [56]; and (iii) based on local criteria [61,62]. The local criteria might be based on information from published and unpublished documents. Under this species conservation concern categorization approach classes made by the Woody Biomass Inventory and Strategic Planning Project were used [61]. The above study has covered about 60% of the area in Ethiopia and the classification includes those species with a population density below 100,000 individuals in the country.
To test for differences in stand structure, diversity and richness a one-way ANOVA followed by post–hoc testing (Fisher’s LSD test) was used. Levene’s test was also conducted to check homogeneity of variances. Linear regression analysis was performed to analyze the relationship between some parameters.
All statistical analysis were done using Statistical Package for Social Sciences–IBM SPSS version 26 (SPSS Inc. 2019) and Microsoft Window Excel (2016).
3. Results and Discussion
3.1. Plant diversity and conservation in indigenous agroforestry systems
3.1.1. Perennial plant species composition
Our results show that ‘Dilla Zuria’ AF systems harbor perennial woody and non-woody plant species diversity. This underlines conservation of these biological resources should not be restricted to forest areas alone since there have been massive encroachment and pressure by anthropogenic factors [63]. A total of 52 perennial woody and non–woody plant species belonging to 30 families were recorded (Table A1). Out of this number 31 plant species were recorded from the 60 inventoried sample plots while the remaining 21 plant species were recorded out of the 60 sample plots. The highest number of species richness was recorded in C–Ft–E based AF system (22 species) (Table A2) whereas the lowest number was recorded in enset based AF system (15 species) (Table A3). Similar results were found in Gedeo Zone of Southern Ethiopia where enset based AF system had the lowest species richness (26 species compared to the remaining two systems [13].
The cumulative species richness in the current study sites was within the range of woody and non-woody species recorded in AF systems of southern Ethiopia (50–120 plant species) [18,19,20,64] and in central Ethiopia is 27–114 species [22,23,24]. However, the species richness of the present study was higher than in northern Ethiopia (17–40) species [25,26]. In addition, the results of this study showed a higher richness over three AF practices compared to Wolayta zone of southern Ethiopia with 32 woody species belonging to 19 families [65]; 39 woody species belonging to 25 plant families recorded by [66] in the same district of south-eastern rift-valley landscapes but in different sites than the current study. Similarly, species richness of the present study was higher than reported in coffee-based AF system, eastern Uganda (50 woody species; [67].
Higher plant species richness than the studied AF systems was also recorded in different parts of the country as well as in other tropical countries: 55 woody species for traditional AF practices of Dellomenna district of south-eastern Ethiopia [68], 58 wood species for Gedeo Zone of Southern Ethiopia [13], 69 for the compound farms of Nigeria [69], 77 woody species for Kandy in Srilanka [70], 83 species for Nicaragua, 100 species for Yem special district of southern Ethiopia [63] 129 species for Kerala in India [71], 168 species for Peruvian Amazon [72] and 179 species for west Java [73], 289 woody plants from sub-urban areas in Sri Lanka [74] and 459 tree and shrub species around Mt. Kenya in central and eastern Kenya [75]. The higher species richness in these study sites might be related to the scale of areal coverage included in the study and the range of agro-climatic zones. Because, some authors argued that the wider the scale of the study in terms of areal coverage [39] and altitudinal range [76], the better is the probability of getting more additional woody and non-woody plant species adapted to different agro–ecology. The distance from natural forest areas can also greatly influence the species richness of AF systems. For instance, studies of AF Systems in a biodiversity hotspot region of northeast India have shown that tree species richness with the proximity to natural forests, and the tree species are closely similar [77]. AF systems that are structurally complex and vegetatively diverse are thought to have the ability to improve landscape connectedness, lessen edge effects, and preserve relatively high levels of species richness [10]. [39] reported that the variation in plant species richness in different study areas could be also related with the difference in site characteristics (farm size, altitude), management strategy of the practitioner and socioeconomic factors. [34,78] stated that farmers’ preferences for the trees and shrubs they choose to plant for various purposes may have an impact on the diversity in species richness in a given AF system. In general, the high woody species richness under AF systems of the present study underlines their potential for biodiversity conservation.
Out of the 30 families recorded in the studied agroforestry systems, three families had higher number of species: Fabaceae (represented by 5 species), Myrtaceae (4) and Euphorbiaceae (3). Francoaceae, Rhizophoraceae, Rubiaceae, Anacardiaceae, Lauraceae, Boraginaceae, Rhamnaceae, Asteraceae, Dracaenacea, Caricaceae, Annonaceae, Solanaceae, Cupressaceae, Salicaceae and Phyllanthaceae however were only represented by one species (Table A1). In general, the small numbers of families (10%) were represented by 5, 4 and 3 species while the majority of families (50%) were represented by a single species. The remaining 40% of families were represented by two species. The highest number of perennial woody or non-woody plant species in this study was represented by the family Fabaceae. Similar studies conducted by [13,63,65] also found that the family Fabaceae in different AF systems scored higher number of species compared to other families. The assessment regarding the origin of the woody and non–woody species across the three AF systems showed that 33 of 52 species (63.5%) were native while the remaining 19 of 52 species (36.5%) were non-native.
The highest native perennial woody and non-woody plant species number was registered in enset based AF system with 14 out of 15 species (93.3 %) while the lowest was in C–Ft–E based AF system with 13 out of 22 species (59%) (Table A2, Table A3 and Table A4). This might be due to practitioners established these AF systems by selectively thinning suppressed tree natural forests. It has been also a common practice to deliberately kept native trees for the purpose of shading for coffee or/and soil fertility and other ecosystem services. For instance, Millettia ferruginea (Hochst.) Baker and Cordia africana Lam. have been used as shade for coffee because of their less dense crown and scattered branches. In addition, the practitioners believed that M. ferruginea (Hochst.) Baker has the ability to improve soil fertility and enhance the productivity of crop and vegetables that grow beneath them [79]. The average native plant species percentage in the present study (63.5%) was higher than in a study reported from 402 homegardens in six regions in south-western Bangladesh with 247 out of 419 (59%) [16]. However, average native plant species percentage in the current study was lower than three indigenous AF systems in Gedeo zone of Southern Ethiopia where 50 out of 58 species (86%) were reported [13].
However, in the case of C–Ft–E based AF systems the plots were dominated by non-native fruit species such as Persea americana, Musa acuminata, Psidium guajava, Carica papaya and Mangifera indica. The dominance of non-native species in this type of AF might be due to the high number of fruit tree species which were introduced by development missionaries and domesticated for lower altitude areas [20]. In lower altitude areas warmer temperatures are mostly reflected. This type of weather condition might speed up litter decomposition and thus improve soil fertility, which favors the growth of a variety of plants. Introduction of these non-native species might affect the existence of native species implying that they could be replaced by non-native ones due to shortage of space for proper growing. These non-native species may also be attractive for the farmers because of their values for consumption and the market. In general, maintaining such considerable number of tree and shrub species in the studied AF systems, both native and non-native in origin implies a great role of these indigenous AF systems in the conservation of plant genetic resources. The role of AF systems in maintaining substantial number of plant species was also reported by [80,81] who conducted a research in homegarden AF systems of west Java and east Usambara of Indonesia.
3.1.2. Plant species endemism and conservation concern
Agroforestry systems can play a great role in conservation of plant species by providing a variety of habitats for different species. Agroforestry systems can be designed to provide a range of different microclimates, soil types, and other environmental conditions, as well as to provide habitat for wildlife, and promote natural regeneration of native species, which can help to restore plant diversity in an area [7,82]. Farmers who adopt AF instead of monocultures have less risk of pest and disease attack due to the diversity of plant species in the system, frequent disturbance through crop rotation, tillage and burning and minimizing the population of pathogens and their dispersal [83]. In the current study the role of the three AF systems in conservation of native and endemic perennial woody and non–woody species was assessed. According to the results M. ferruginea (Hochst.) Baker and Erythrina brucei were some of the common woody species found across the studied AF systems being registered as native and endemic. However, it has been argued that the species distinctiveness expressed in terms of their presence as rare species or endemic species of AF systems is low compared to forest areas because of anthropogenic drivers [10]. The reason for a small number of endemic woody and non-woody perennial plant species in the present study might therefore be related with anthropogenic activities such as removal of native trees and replacing with some cash crops and non-native fruit trees.
Species conservation concern of AF systems is also one of the important issues to deal with. As the inventory from the three AF systems showed, a total of 13 species were listed as species of conservation concern according to the IUCN Red Lists and local criteria. M. ferruginea, Erythrina brucei, Dracaena steudneri, Senna siamea, Trichilia dregeana, Melia azedarach L., Azadirachta indica var., Albizia grandibracteata Taub., Bridelia micrantha (Hochst.) Baill. were listed under the least concern by IUCN red lists [60]. Rhamnus prinoides was listed as both rare for 25% of species that least occurred [56] and as least number of individuals (\100,000 individuals in the country) as per local criteria [61]. Prunus africana was listed as both vulnerable by IUCN Red Lists [60] and rare for 25% of species that least occurred [56]. Albizia gummifera and Ficus vasta were listed as rare for 25% of species that least occurred [56]. The number of species listed under IUCN Red List in the present study (10 out of 52) were higher than reported in south-western Bangladesh (6 out of 419) [16]. In terms of proportion from the total species, the number of Red List species in the current study (25%) was by far higher than the reported in South-western Bangladesh (1.4%). This difference might be due to the physiogeographic situation (distinct type of landscape, landforms, rock type, and evolutionary history of Ethiopia and thus situated in East African highland) as compared to Bangladesh.
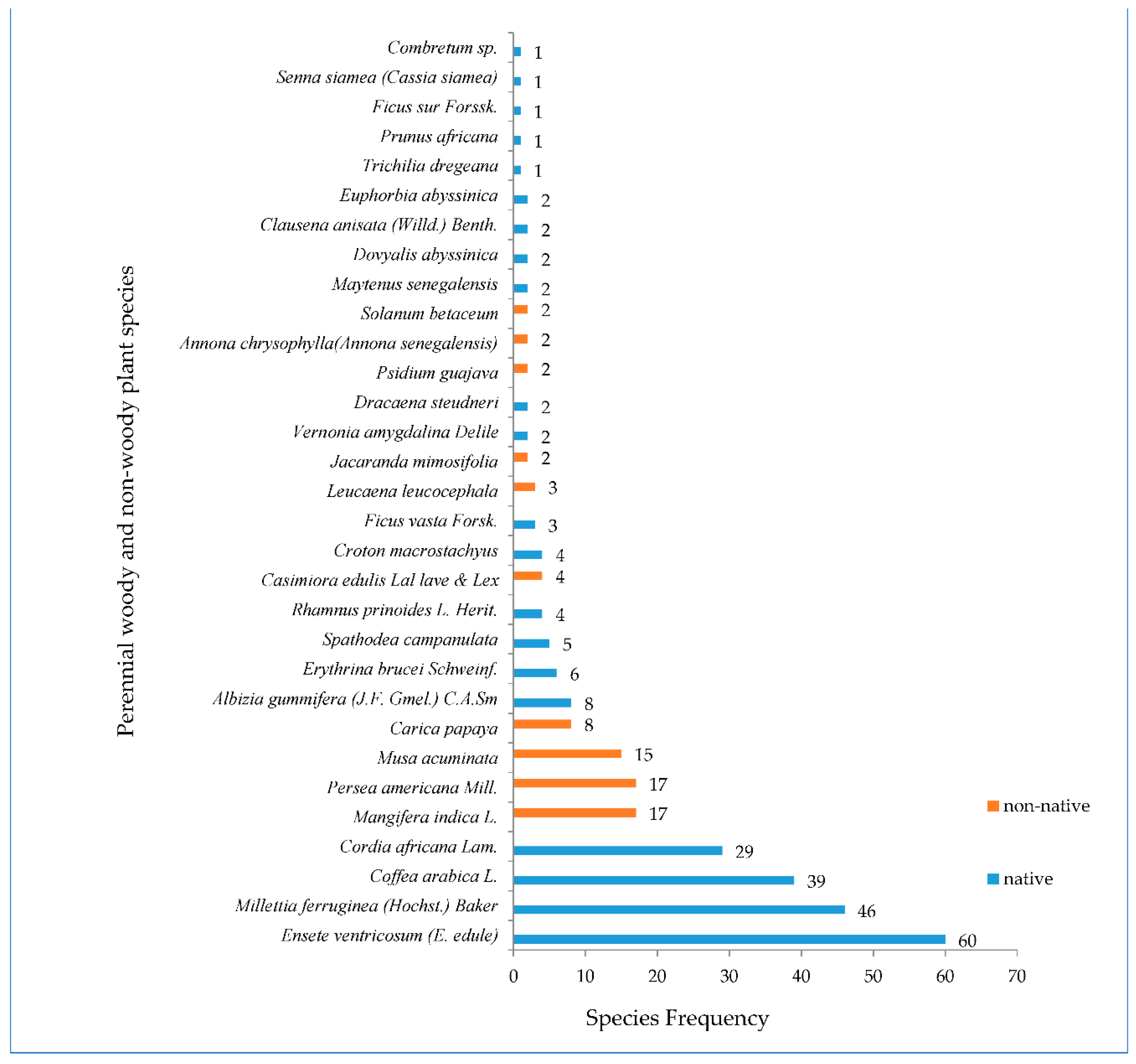
The assessment of species in terms of rarity showed that the occurrence of five native species was very limited in certain sample plots. Woody species such as Combretum sp., P. africana, Ficus sur Forssk, S. siamea (C. siamea) and T. dregeana occurred only in one sample plot (Figure 6), implying that these species are rare, demand conservation and need to be maintained by practitioners.
3.1.3. Plant species frequency and important value index
Out of the 31 plant species the four were the most frequent across the inventoried farms. E. ventricosum was the most frequent plant species occurring in 60 sample plots and was followed by M. ferruginea (in 46 sample plots), Coffea arabica L. (in 39 sample plots) and C. africana (in 29 sample plots) (Figure 6). A study conducted in similar zone but under different site conditions reported that C. arabica, C. africana and M. ferruginea were the most frequent perennial woody species [66]. On the other hand, 5 woody plant depicts the result reported by [68] who mentioned plant species with a greater economic or/and ecological value were found to be more frequently distributed across the smallholdings. M. ferruginea also showed higher frequency mainly due to the special characteristics of the tree species such as serving as a shade for C. Arabica. It is better adapted in the area, and propagation and management of the species is easy [13]. It further has the ability to improve soil fertility and thus increase productivity of crops and vegetables planted beneath [79]. The third most frequent species was C. arabica. The reason might be related to its economic importance and income source for the household enhancing the livelihood [28,39].
Figure 6.
Frequency of perennial woody and non-woody plant species across the three AF systems (60 sample plots) of the study sites, south eastern rift–valley landscapes, Ethiopia.
Figure 6.
Frequency of perennial woody and non-woody plant species across the three AF systems (60 sample plots) of the study sites, south eastern rift–valley landscapes, Ethiopia.
The important value index (IVI %) of each plant species in the studied AF systems was calculated to determine the significance of each individual species in the system. According to the results, five plant species with the highest important value index in C–Ft–E based indigenous AF system were E. ventricosum, Coffea arabica L., M. acuminata, M. indica L. and P. americana Mill respectively (Table 1). In comparison to the first and third AF systems the enset based AF system exhibits a higher IVI% for Enset species. Besides the lead species, the system includes four more different tree species which are important multipurpose species, namely M. ferruginea (Hochst.) Baker, C. africana Lam., Erythrina brucei Schweinf and Croton macrostachyus. In the C–E based AF system there is besides the lead species coffee and enset a certain species overlap with the second AF system.
The IVI% of E. ventricosum was recorded the highest across the three AF systems. This was due to the species showed a high relative frequency, relative abundance and relative dominance in each AF system. The variation in IVI% for various woody or/and non–woody species among the AF systems might be related to farmers’ species preference, growth performance and original stocking density of the species in the sample quadrants [65].

Table 1.
Woody and non woody plant species with the highest important value index in each of the three indigenous AF systems in South-eastern rift-valley landscapes, Ethiopia.
Table 1.
Woody and non woody plant species with the highest important value index in each of the three indigenous AF systems in South-eastern rift-valley landscapes, Ethiopia.
| Agroforestry system | Species Scientific name | Important value index (IVI %) |
|
Enset based AF system |
Ensete ventricosum (Welw. Cheesman) | 204.6 |
| Millettia ferruginea (Hochst.) Baker | 40.9 | |
| Cordia africana Lam. | 22.2 | |
| Erythrina brucei Schweinf. | 6.1 | |
| Croton macrostachyus | 4.0 | |
|
C-E based AF system |
Ensete ventricosum (Welw. Cheesman) | 159.2 |
| Coffea arabica L. | 56.3 | |
| Millettia ferruginea (Hochst.) Baker | 23.9 | |
| Cordia africana Lam. | 21.3 | |
| Albizia gummifera (J.F. Gmel.) C.A.Sm | 6.7 | |
|
C-Ft-E based AF system |
Ensete ventricosum (Welw. Cheesman) | 103.4 |
| Coffea arabica L. | 46.7 | |
| Musa acuminata | 42.3 | |
| Mangifera indica L. | 24.1 | |
| Persea americana Mill. | 21.6 |
Enset based AF system: enset based agroforestry system; C–E based AF system: coffee–enset based agroforestry system; C–Ft–E based AF: coffee–fruit tree–enset based AF system.
Under C–Ft–E based AF system the majority of the plant species with the highest IVI% were non-native species specifically fruit trees whereas in enset based and C–E based AF systems all the species with highest IVI% were the native ones. These results coincide with the report on perennial plant species composition (3.1.1) of this study which found a higher percentage of native species under enset based and C–E based AF system while the C–Ft–E based AF system had lower numbers of native species.
3.1.4. Stand structure, diversity and richness status of agroforestry systems
Considering the stem number, basal area, height and DBH of only enset species in the three AF systems, enset based AF system showed the highest whereas the lowest value for these parameters was found in C–Ft–E based AF system (Table 2). The mean stem number, BA and DBH were higher in C–Ft–E based AF system whereas mean height for the woody species was higher in enset based AF system (Table 3). The least mean values of stem number and basal area for only woody species were recorded in enset based AF system (Table 3). The computation of these four parameters was also carried out for the mixture of woody and enset species for each AF system. Based on the results the highest mean stem density was recorded in C–Ft–E based AF system (71.2 stems) whereas the least was in enset based AF system (44.6 stems) (Table 4). The highest average basal area, height and DBH of the perennial woody species and enset together was recorded in enset based AF system; 317.7 m2 ha-1, 4.6 m, 26.7 cm respectively (Table 4). The combined mean perennial woody species stem density (20.8 per 100 m2 or 2,083 stems ha–1 when extrapolated to hectare basis) for all 60 farm plots in the present study was much higher than the reported values by [13] who found stem density of woody species in enset based AF (625 stems ha-1), C–E based AF (1,240 ha–1) and fruit–coffee AF systems (1,505 ha–1) of South-eastern Ethiopia. Similarly, [39] and [84] reported 636 stems ha-1 in the enset–coffee–Maize AF systems in southern Ethiopia and 1,833 stems ha-1 in homegardens of west Java which is below mean values of the present study. Greater difference in stem density of our results and those reported by other authors might be due to the farm owners in the current study have better silvicultural management of the system. Proper lopping, pollarding, pruning and stratified layer planting are some of the good silvicultural management activities practiced by the farm owners. Having stratified layers in the studied AF systems allows growing of different trees/shrubs in some space so that they could have less competition for the above and belowground resources.
The mean stem density, basal area, height and DBH of enset species for enset based AF system was significantly different at (P<0.05) from C–Ft–E based AF system (Table 2). In addition, enset based AF system showed significant difference from C–E based AF only for mean basal area (Table 2). The computed values of only perennial woody species in the three AF systems showed that C–Ft–E based AF system was significantly different at (P<0.05) from enset based AF for their mean stem density, basal area and DBH (Table 3). However, the mean height of only woody species in enset based AF was significantly different from both C-Ft-E based and C–E based AF systems (Table 3). Under Table 4, the computation for their mean stem density, BA, height and DBH were conducted for all the woody and enset species as mixture for the three AF systems. C–Ft–E based AF system was significantly different (P<0.05) from enset based in terms of mean stem density, BA and DBH. In addition, C–Ft–E based and enset based AF system were significantly different from C–E based AF system in terms of BA and DBH. However, height of the species did not significantly differ among the AF systems (Table 4). The stem density of plant species in AF systems is related to ecological (altitude, rainfall and temperature) [39] and socioeconomic conditions (marketing, size of land holding) [39,84]. The highest mean BA and mean DBH of plant species was found in enset based AF system with 317.7 m2 ha–1 and 26.7 cm respectively. The lowest mean BA and mean DBH of species was found in C–Ft–E based AF with 149.2 m2 ha–1 and 15.7 cm respectively.
The mean basal area and DBH of all species significantly differed (P<0.05) between the three agroforestry systems (Table 4). According to the computed mean basal area values for each species, E. ventricosum (58.6%), Musa acuminate (11.8%) and C. arabica (7.0%) in C-Ft-E based (Table A4) and E. ventricosum (96.1%), C. africana (1.4%) and C. macrostachyus (0.4%) in enset based (Table A2) had the highest relative dominance. Under C–E based AF system E. ventricosum (89.8%), C. africana (3.8%) and C. arabica (2.7%) showed highest relative dominance (Table A3). The share of native plant species in terms of relative dominance was 80%, 99.9% and 99.5% in C–Ft–E based, enset based and C–E based AF systems respectively. The results revealed that native species almost fully dominate the horizontal space especially in enset based and C–E based AF systems. The average stem number (2083.3 stems ha-1) and BA (29 m2 ha-1) of perennial woody species recorded in the present study was higher than reported in other indigenous fruit-coffee AF systems of south eastern Ethiopia with 1505 stems ha-1 and BA of 11.7 m2 ha-1 [13], in coffee based agroforests in Guinea between 1,071 and 1,239 stems ha-1 and BA of 22.15 m2 ha-1 [85] and in Cocoa agroforest and mixed food agroforest in south eastern Ghana with 125 stems ha-1 and BA of 8.4 m2 ha-1 [86]. The greater difference in stem number and basal area of this investigation and those reported by other authors might be related to the tendency of the farmers to maintain more native trees from previous forest land and planting of more coffee and non-native fruit trees.
Table 2.
Mean stem number, basal area (BA), height, and diameter at breast height (DBH) of only enset species for each AF system, followed by SE in parenthesis.
Table 2.
Mean stem number, basal area (BA), height, and diameter at breast height (DBH) of only enset species for each AF system, followed by SE in parenthesis.
| Agroforestry system | n | Stem number (No/100 m2) | BA (m2 ha-1) |
Height (m) |
DBH (cm) |
| Enset based AF | 20 | 34.7(2.7)(b) | 306.4(28.8) (b) | 4.4(0.2) (b) | 31.0(1.7) (b) |
| C–E based AF | 20 | 29.3(2.8)(b) | 207.0(15.1) (c) | 4.1(0.2) (b) | 28.8(1.8) (ab) |
| C–Ft–E based AF | 20 | 13.1(2.0)(a) | 81.2(9.3) (a) | 3.6(0.2) (a) | 24.2(1.4) (a) |
| P–value | <0.05 | <0.05 | <0.05 | <0.05 |
C–E based AF: coffee–enset based AF; C–Ft–E based AF: coffee–fruit tree–enset based AF.
Table 3.
Mean stem number, basal area (BA), height and diameter at breast height (DBH) of only woody plant species for each AF system, followed by SE in parenthesis.
Table 3.
Mean stem number, basal area (BA), height and diameter at breast height (DBH) of only woody plant species for each AF system, followed by SE in parenthesis.
| Agroforestry system | n | Stem number (No/100 m2) | BA (m2 ha-1) |
Height (m) |
DBH (cm) |
| Enset based AF | 20 | 9.3(1.7)(b) | 11.3(2.5) (b) | 6.0(0.8) (b) | 11.2(1.3) (a) |
| C–E based AF | 20 | 22.0(1.1)(c) | 21.9(4.1) (bc) | 3.6(0.2) (ac) | 8.1(0.3) (b) |
| C–Ft–E based AF | 20 | 31.2(3.5)(a) | 53.8(10.4) (a) | 4.2(0.2) (a) | 11.8(0.5) (a) |
| P–value | <0.05 | <0.05 | <0.05 | <0.05 |
C–E based AF: coffee–enset based AF; C–Ft–E based AF: coffee–fruit tree–enset based AF.
Table 4.
Mean stem number, basal area (BA), height and diameter at breast height (DBH) of perennial woody and enset species together for each AF system, followed by SE in parenthesis.
Table 4.
Mean stem number, basal area (BA), height and diameter at breast height (DBH) of perennial woody and enset species together for each AF system, followed by SE in parenthesis.
| Agroforestry system | N | Stem number (No/100 m2) | BA (m2 ha-1) |
Height (m) |
DBH (cm) |
| Enset based AF | 20 | 46.9(3.0)(b) | 317.7(28.1) (b) | 4.6(0.1) (a) | 26.7(1.5) (b) |
| C–E based AF | 20 | 53.8(2.6)(b) | 228.5(14.8) (c) | 4.3(0.2) (a) | 18.9(0.7) (c) |
| C–Ft–E based AF | 20 | 71.2(3.2)(a) | 149.2(17.6) (a) | 4.3(0.1) (a) | 15.7(0.7) (a) |
| P–value | <0.05 | <0.05 | NS | <0.05 |
Note: similar letter shows not significant difference and different letters indicate significance differences between groups according to LSD multiple test (Fisher LSD test) at P<0.05; NS: not significant. C–E based AF: coffee–enset based AF; C–Ft–E based AF: coffee–fruit tree–enset based AF.
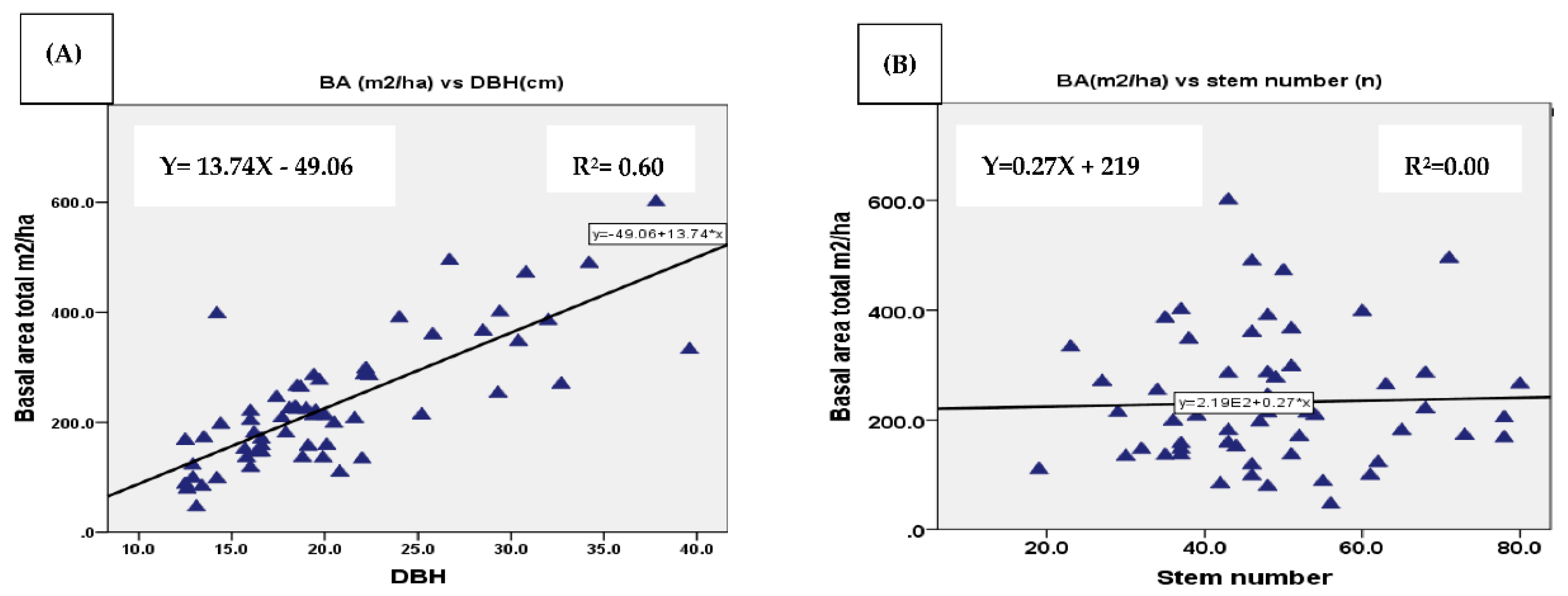
The relationship between BA and DBH; between BA and stem number was assessed using regression analysis. The assessment was conducted for enset species, woody species separately, and mixture of woody and enset species in the three AF systems. As the results displayed in the regression graph the correlation between BA and DBH by separate computing of woody species and enset was from very low to low respectively (Figure 7A and Figure 8A). The mean BA of enset and woody species separately within the AF systems was somehow affected by stem number although the correlation was low with values of r2 =0.31 and r2 =0.23 respectively (Figure 7B and Figure 8B). The mean BA was increased with increasing mean DBH with a correlation of (r2 =0.6) for mixture of woody and enset species (Figure 9A). However, no correlation was found between BA and stem number (Figure 9B). The reason might be mixing of woody and enset which increases the density of the stand and thus the possibility of smaller diameter of enset species.
Diversity indices such as Shannon diversity index, Margalef’s richness index and Pielou’s Eveness index helped us to analyze and evaluate the relationships of species distributed among the three studied AF systems. According to the results, C–E based AF system showed higher species abundance (51.3) whereas enset based AF system showed lower (44.6) The highest diversity index and richness was, however, observed in C–Ft–E based AF systems whereas the lowest was in enset based AF systems.
Figure 9.
Relationship between diameter at breast height (DBH) and basal area (A); stem number and basal area (B) for the woody and enset species of the three studied AF systems.
Figure 9.
Relationship between diameter at breast height (DBH) and basal area (A); stem number and basal area (B) for the woody and enset species of the three studied AF systems.
Species mean abundance and Pielou’s Eveness index values between the AF systems were not significantly different (Table 5). Shannon diversity and Margalef’s richness index of species was significantly different (P<0.05) between the three AF systems. The high richness index in C–Ft–E based AF might be related with proximity to main roads [38,39] and favorable environmental conditions like temperature [13]. For instance, the temperature in the C–Ft–E based AF system (>25 o C) was slightly warmer compared to the remaining two systems with 13-28 o C. This warmer weather condition coupled with high amount of rainfall might bring favorable condition for plants to survive easily and grow faster. This might motivate the practitioners to incorporate more woody and non–woody species (in our case mainly high value fruit trees and coffee, enset and other native species) to get more benefits out of them. [20] also reported greater species richness in C–Ft–E based AF might be related with the incorporation of various native and non–native woody species along a vertical stratum.
The Shannon diversity index values in the present study for C–Ft–E based (1.1) and C–E based AF (1.0) were comparable with studies conducted in enset–coffee–Maize–Chat AF (1.15) in Sidama region of Southern Ethiopia [39] and in Kerala homegarden agroforestry (1.2) in India [71]. However, our results were lower than values reported by [68] in traditional AF practices (2.2) of the Dellomenna district of South-eastern Ethiopia; [87] in homegarden AF (1.8) of Southern Gonder of Ethiopia [65] in homegarden AF (2.23) of Wolayitta Zone, Ethiopia. Lower diversity index values of the present study compared to above mentioned reports was due to the medium evenness index values across all small holdings of the investigated AF plots. This implies species diversity is to be affected by abundance and equitability of the species within the sample plots. The results of this study in terms of Pielou’s Eveness index (with a mean value of 0.6) were also comparable with other results: in homegarden AF (0.6) of similar study zone like ours but with different study sites [66], in enset–coffee–Maize–Chat–Pineapple AF (0.55) in Sidama area of Southern Ethiopia [18] and in homegardens of Kerala (from 0.24 to 0.71) of Southern India of Southern India [71].
Table 5.
Mean ±SD. of woody and non-woody plant species abundance, Shannon diversity index (H′), Margalef’s richness index (Dmg) and Pielou’s evenness (J) of study plots under the three AF systems.
Table 5.
Mean ±SD. of woody and non-woody plant species abundance, Shannon diversity index (H′), Margalef’s richness index (Dmg) and Pielou’s evenness (J) of study plots under the three AF systems.
| Agroforestry system | N | Abundance Per 100 m2 |
Shannon diversity index | Margalef’s richness index | Pielou’s Eveness index |
| Enset based AF | 20 | 44.6(3.0) (a) | 0.7±0.2(b) | 0.6±0.2(b) | 0.6±0.1(a) |
| C–E based AF | 20 | 51.3(2.6) (a) | 1.0±0.1(c) | 1.0±0.3(c) | 0.6±0.1(a) |
| C–Ft–E based AF | 20 | 48.5(3.2) (a) | 1.1±0.2(a) | 1.2±0.3(a) | 0.6±0.1(a) |
| P–value | NS | <0.05 | <0.05 | NS |
Note: Same letter shows not significant difference and different letters indicate significance differences between groups according to LSD multiple test (Fisher LSD test) at P<0.05; NS: not significant. C–E based AF: coffee-enset based AF; C–Ft–E based AF: coffee–fruit tree–enset based AF.
The equitability of the woody species was almost the same across all the AF systems. The result of one-way ANOVA followed by post-hoc testing (Fisher’s LSD test) (n=20) showed that the difference in mean Pielou’s Eveness index between the studied AF systems was not significant. The mean Pielou’s Eveness index value of 0.6 implies a situation in which species are moderately distributed in each plot of the AF systems or in other words the relative homogeneity of the species in the sample plots was 60% of the maximum possible even population across all smallholdings. According to the analysis of Sorensen’s similarity index for the three AF systems, highest species similarity was observed between C–Ft–E based AF and C-E based AF with a value of 67% (14 woody species out of 28) while the least was between C–Ft–E based AF and enset based AF with a value of 48% (9 woody species out of 28). The species similarity between enset based AF and C–E based AF was a little higher than the later relatively with a value of 57 % (10 woody species out of 25).
3.1.5. Relationship of altitude with species richness and species abundance
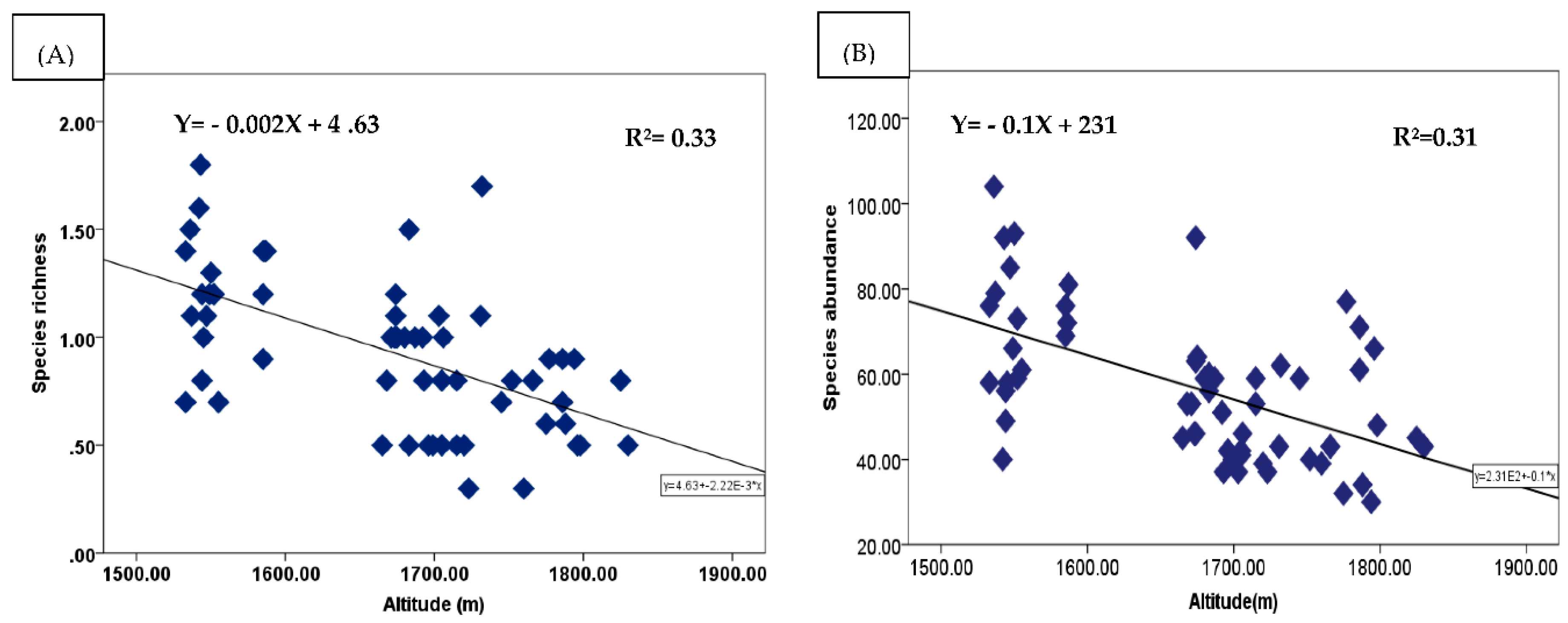
Altitude is one of the important variables that could exert an effect on habitat quality and thus influence species richness, composition and diversity. This is because, altitude affects changes in the availability of relevant resources for plant growth for instance heat and water [66,88]. Regression analysis was performed to evaluate the relationship between altitude versus species richness and abundance. The graphs representing all the three studied AF systems are displayed in Figure 10A and 10B. Our results showed that both the mean Margalef’s species richness index and abundance were decreasing as altitude increased. The correlation between mean species richness and altitude was r2=0.33 whereas the correlation between mean species abundance and altitude was r2=0.31 (Figure 10B). From the results we could understand that altitude was more related to species richness than species abundance although both have very low correlation values.
The results of the present study are in line with other reports, such as [89], who observed a decreasing trend of plant species richness as altitude increases in Northeastern Tibetan Plateau China. Further [66] and [13] found similar trends in indigenous homegarden AF of southern Ethiopia. However, contradicting results were reported by [39,90] in which an increasing trend of species richness with increasing altitude. These studies were conducted in Qinghai-Tibetan Plateau China and Southern Ethiopia respectively. The reason for increasing species richness with altitude in these studies might be related with different factors. For instance, the study conducted by the first author included all altitudinal ranges (from 320-5200 m asl). Therefore, increasing species richness with altitude was because in the higher altitudes impact of livestock in destructing plant species is very low compared to the lower altitudes. The second author also reported an increasing enset species richness with altitude but not for all species. This increase might be due to enset species prefer an altitudinal range between 2000 and 2500 m asl. Whether to get an increasing or decreasing trend of species richness with altitude is greatly affected by the scale of the study for instance altitudinal range [76]. The author articulated, if a survey of the entire altitudinal gradient is conducted the pattern show a hump shape, implying that an increasing trend of species richness up to a certain altitude range was observed and then started to decrease. But, if the survey is conducted in a narrow scale of altitudinal gradient, the pattern changes progressively to a monotically decreasing trend of species richness with increasing altitude. Therefore, from the above idea, we could understand that the relationship between species richness and altitude could be either negative or positive based on the situations.
Figure 10.
Relationship between Altitude and Margalef’s species richness index (A), Altitude and species abundance (B) under the three studied AF systems.
Figure 10.
Relationship between Altitude and Margalef’s species richness index (A), Altitude and species abundance (B) under the three studied AF systems.
Species richness and abundance could also be affected by other factors such as educational level of land owner, land ownership, slope and extension access [25] and farm size [18,25]. As farmers become aware, well educated, own bigger land size and get better access to extension service, their tendency to grow more trees becomes high. In addition, the possibility of incorporating diverse fruit and non–fruit trees would be higher. Edaphic factors such as soil conditions could positively or negatively affect species richness and evenness [91]. For instance, a study conducted by [92] revealed that species richness was negatively correlated with phosphorus availability and species evenness was negatively correlated with the ratio of organic carbon to total nitrogen in soil.
4. Conclusion and Recommendations
Agricultural landscapes which integrate AF systems showed a great potential for maintaining woody and non-woody species diversity as forest lands do. The cumulative species richness in the current study for woody and non–woody species was in a comparable range as reported for other AF systems of Southern and Central Ethiopia. The higher percentage of native plant species in the present study is an indicator of the sustainability of the system in conserving the plant species. The dominance of non-native fruit tree species in C–Ft–E based AF could be a result of global preference and marketing of these fruit species. If the dominance continues to the other AF systems it could be a threat for the maintenance of native species for the future. The studied AF systems have conserved 13 species, which are categorized conservation concern according to the IUCN Red List and local criteria. This implied how strong AF systems are able to serve as a refuge for species. In general, this study offers an opportunity to evaluate the status of the AF systems and provide scientific information on conservation potential of the systems for endemic native species. It also further contributes to the development of a national policy concerning the conservation of biodiversity. Some native tree species were found to be rare although they are dominant in other areas. For instance, species such as Trichilia dregeana, Ficus sur Forssk., Prunes africana and Combretum species were some of them. Therefore, a special conservation priority coupled with wise utilization of native plant species should be implemented by the community to maintain their presence. The governmental, non–governmental organizations and other concerned stakeholders should promote different AF practices to conserve native woody and non-woody plant species through circa situm conservation. Since the current study was conducted in a limited AF sites, further research on broader scale is needed to explore the potential of the systems to accommodate further native and endangered native taxa.
Author Contributions
H.M.T. generated the idea and study design, collected data, carried out the laboratory work, data analysis, and write up. Janine Oettel and Katharina Lapin reviewed the manuscript. M.N. supported in study design, read and reviewed the manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
Austrian Partnership Program in Higher Education and Research Development (APPEAR) and International Foundation for Science (IFS Grant. #D/6183–1) supported this research as part of a Ph.D. study.
Data Availability Statement
Datasets are attached in separate file.
Acknowledgments
We are grateful to the Austrian Partnership Program in Higher Education and Research Development (APPEAR) for giving H.M.T. a scholarship award to pursue study in Austria. H.M.T. acknowledges financial support from the International Foundation for Science (IFS Grant #D/6183–1) and Federal Research and Training Center for Forests, Natural Hazards and Landscape, Department of Biodiversity and Nature Conservation, Vienna, Austria. H.M.T. is grateful to Mesele Negash for his indispensable support from the beginning to the end, and also extends gratitude to Oettel Janine and Katharina Lapin for their valuable review. The University of Natural Resource and Life Sciences (BOKU), Institute of Forest Ecology, Austria, and Hawassa University, Wondo Genet College of Forestry and Natural Resources, Ethiopia, are also acknowledged for offering the postgraduate study opportunity. We are also indebted to Gedeo zone experts and the farmers for allowing us to visit and carry out this study on their farms and for providing marvelous experiences.
Conflicts of Interest
We declare that there are no conflicts of interest.
Appendix A
Table A1.
List of perennial woody and non–woody plant species recorded across the three studied AF systems of our study sites.
Table A1.
List of perennial woody and non–woody plant species recorded across the three studied AF systems of our study sites.
| Number | vernacular name | Scientific name | Family | Origin |
| 1 | Gorbe | Albizia gummifera (J.F. Gmel.) C.A.Sm | Fabaceae | Native |
| 2 | Geshita | Annona chrysophylla Bojer | Annonaceae | Non-native |
| 3 | Buno | Coffea arabica L. | Rubiaceae | Native |
| 4 | Papaya | Carica papaya | Caricaceae | Non-native |
| 5 | Abukere | Casimiroa edulis Lal lave and Lex | Rutaceae | Non-native |
| 6 | Godere | Clausena anisata (Willd.) Benth. | Rutaceae | Native |
| 7 | NI | Combretum sp. | Combretaceae | Native |
| 8 | Wedesa | Cordia africana Lam. | Boraginaceae | Native |
| 9 | Mokonisa | Croton macrostachyus | Euphorbiaceae | Native |
| 10 | NI | Dovyalis abyssinica | Salicaceae | Native |
| 11 | Cho’e | Dracaena steudneri Schweinf. Ex Engl. | Dracaenaceae | Native |
| 12 | Ensete | Ensete ventricosum (Welw. Cheesman) | Musaceae | Native |
| 13 | Welale/Gedogna | Erythrina brucei Schweinf. | Leguminosae | Native |
| 14 | Kulkal | Euphorbia abyssinica | Euphorbiaceae | Native |
| 15 | wagela | Ficus sur Forssk. | Moraceae | Native |
| 16 | Kilto | Ficus vasta Forsk. | Moraceae | Native |
| 17 | NI | Jacaranda mimosifolia | Bignoniaceae | Non-native |
| 18 | Lusina | Leucaena leucocephala | Mimosoideae | Non-native |
| 19 | Mango | Mangifera indica L. | Anacardiaceae | Non-native |
| 20 | Kobo/gulo | Maytenus senegalensis | Celastraceae | Native |
| 21 | Tatato | Millettia ferruginea (Hochst.) Baker | Leguminosae | Native |
| 22 | Muse | Musa acuminata | Musaceae | Non-native |
| 23 | Avocato | Persea americana Mill. | Lauraceae | Non-native |
| 24 | Gorbe | Prunus africana | Rosaceae | Native |
| 25 | Sholla | Psidium guajava L. | Myrtaceae | Non-native |
| 26 | Gesho | Rhamnus prinoides L. Herit. | Rhamnaceae | Native |
| 27 | NI | Senna siamea (Cassia siamea) | Fabaceae | Native |
| 28 | NI | Spathodea campanulata | Bignoniaceae | Native |
| 29 | Timatim zaf | Solanum betaceum | Solanaceae | Non-native |
| 30 | NI | Trichilia dregeana | Meliaceae | Native |
| 31 | Hebicha | Vernonia amygdalina Delile | Asteraceae | Native |
| List of perennial woody andnon–woody plant species recorded out of the study plots | ||||
| 32 | NI | Albizia grandibracteata Taub. | Fabaceae | Native |
| 33 | NI | Azadirachta indica var. | Meliaceae | Non-native |
| 34 | Tibero/Sessa | Bersama abyssinica Fresen | Francoaceae | Native |
| 35 | Yebelo | Bridelia micrantha (Hochst.) Baill. | Phyllanthaceae | Native |
| 36 | Tilo | Cassipourea malosana (Baker) Alst | Rhizophoraceae | Native |
| 37 | Chate | Catha edulis (Vahl) Forssk. ex Endl. | Celastraceae | Native |
| 38 | Motokomo | Celtis africana N.L. Burm | Ulmaceae | Native |
| 39 | Motokomo | Celtis sp. | Ulmaceae | Native |
| 40 | Lomie | Citrus limon (L.) Osbeck | Rutaceae | Non-native |
| 41 | Birtukan | Citrus sinensis (L.) Osbeck | Rutaceae | Non-native |
| 42 | NI | Cupressus lusitanica | Cupressaceae | Non-native |
| 43 | Bahirzaf | Eucalyptus camaldulensis | Myrtaceae | Non-native |
| 44 | Bahirzaf | Eucalyptus globules Labill. | Myrtaceae | Non-native |
| 45 | Bahirzaf | Eucalyptus grandis | Myrtaceae | Non-native |
| 46 | NI | Grevillea robusta | Proteaceae | Non-native |
| 47 | NI | Faidherbia albida | Fabaceae | Native |
| 48 | Kilto | Ficus elastica Roxb. | Moraceae | Native |
| 49 | NI | Hagenia abyssinica | Rosaceae | Native |
| 50 | NI | Melia azedarach L. | Meliaceae | Non-native |
| 51 | NI | Ricinus communis | Euphorbiaceae | Native |
| 52 | NI | Sesbania sesban | Fabaceae | Non-native |
Table A2.
List of perennial woody and non–woody plant species and their important value index under enset based indigenous AF, South eastern rift–valley landscapes, Ethiopia.
Table A2.
List of perennial woody and non–woody plant species and their important value index under enset based indigenous AF, South eastern rift–valley landscapes, Ethiopia.
| Scientific name | Family |
Fre n |
RF (%) |
Tot Dom |
RD (%) |
AB |
RA (%) |
IVI (%) |
| Albizia gummifera (J.F. Gmel.) C.A.Sm | Fabaceae | 2.0 | 3.0 | 0.2 | 0.3 | 6.0 | 0.6 | 3.9 |
| Combretum sp. | Combretaceae | 1.0 | 1.5 | 0.0 | 0.0 | 4.0 | 0.4 | 1.9 |
| Cordia africana Lam. | Boraginaceae | 11.0 | 16.7 | 0.9 | 1.4 | 39.0 | 4.1 | 22.2 |
| Crot macrostachyus | Euphorbiacee | 2.0 | 3.0 | 0.2 | 0.4 | 6.0 | 0.6 | 4.0 |
| Dovyalis abyssinica | Salicaceae | 1.0 | 1.5 | 0.0 | 0.0 | 3.0 | 0.3 | 1.9 |
| Dracaena steudneri Schweinf. ex Engl | Dracaenaceae | 1.0 | 1.5 | 0.0 | 0.0 | 11.0 | 1.2 | 2.7 |
| Ensete ventricosum (Welw. Cheesman) | Musaceae | 20.0 | 30.3 | 61.3 | 96.1 | 743.0 | 78.2 | 204.6 |
| Erythrina brucei Schweinf. | Leguminosae | 3.0 | 4.5 | 0.0 | 0.1 | 14.0 | 1.5 | 6.1 |
| Maytenus senegalensis | Celastraceae | 1.0 | 1.5 | 0.0 | 0.0 | 1.0 | 0.1 | 1.6 |
| Millettia ferruginea (Hochst.) Baker | Leguminosae | 19.0 | 28.8 | 0.9 | 1.3 | 102.0 | 10.7 | 40.9 |
| Prunus africana | Rosaceae | 1.0 | 1.5 | 0.0 | 0.0 | 4.0 | 0.4 | 1.9 |
| Rhamnus prinoides L. Herit. | Rhamnaceae | 1.0 | 1.5 | 0.0 | 0.0 | 6.0 | 0.6 | 2.1 |
| Senna siamea (Cassia siamea) | Fabaceae | 1.0 | 1.5 | 0.0 | 0.0 | 4.0 | 0.4 | 2.0 |
| Solanum betaceum | Solanaceae | 1.0 | 1.5 | 0.0 | 0.0 | 3.0 | 0.3 | 1.9 |
| Vernonia amygdalina Delile | Asteraceae | 1.0 | 1.5 | 0.2 | 0.3 | 4.0 | 0.4 | 2.2 |
Fre: frequency; RF: relative frequency; Tot Dom: total dominance; RD: relative dominance; AB: abundance; RA: relative abundance; IVI: important value index.
Table A3.
List of perennial woody andnon–woody plant species and their important value index under C–E based indigenous AF system, South eastern rift–valley landscapes, Ethiopia.
Table A3.
List of perennial woody andnon–woody plant species and their important value index under C–E based indigenous AF system, South eastern rift–valley landscapes, Ethiopia.
| Scientific name | Family |
Fre n |
RF (%) |
Tot Dom |
RD (%) |
AB |
RA (%) |
IVI (%) |
| Albizia gummifera (J.F. Gmel.) C.A.Sm | Fabaceae | 2.0 | 3.0 | 0.2 | 0.3 | 6.0 | 0.6 | 3.9 |
| Combretum sp. | Combretaceae | 1.0 | 1.5 | 0.0 | 0.0 | 4.0 | 0.4 | 1.9 |
| Cordia africana Lam. | Boraginaceae | 11.0 | 16.7 | 0.9 | 1.4 | 39.0 | 4.1 | 22.2 |
| Crot macrostachyus | Euphorbiacee | 2.0 | 3.0 | 0.2 | 0.4 | 6.0 | 0.6 | 4.0 |
| Dovyalis abyssinica | Salicaceae | 1.0 | 1.5 | 0.0 | 0.0 | 3.0 | 0.3 | 1.9 |
| Dracaena steudneri Schweinf. ex Engl | Dracaenaceae | 1.0 | 1.5 | 0.0 | 0.0 | 11.0 | 1.2 | 2.7 |
| Ensete ventricosum (Welw. Cheesman) | Musaceae | 20.0 | 30.3 | 61.3 | 96.1 | 743.0 | 78.2 | 204.6 |
| Erythrina brucei Schweinf. | Leguminosae | 3.0 | 4.5 | 0.0 | 0.1 | 14.0 | 1.5 | 6.1 |
| Maytenus senegalensis | Celastraceae | 1.0 | 1.5 | 0.0 | 0.0 | 1.0 | 0.1 | 1.6 |
| Millettia ferruginea (Hochst.) Baker | Leguminosae | 19.0 | 28.8 | 0.9 | 1.3 | 102.0 | 10.7 | 40.9 |
| Prunus africana | Rosaceae | 1.0 | 1.5 | 0.0 | 0.0 | 4.0 | 0.4 | 1.9 |
| Rhamnus prinoides L. Herit. | Rhamnaceae | 1.0 | 1.5 | 0.0 | 0.0 | 6.0 | 0.6 | 2.1 |
| Senna siamea (Cassia siamea) | Fabaceae | 1.0 | 1.5 | 0.0 | 0.0 | 4.0 | 0.4 | 2.0 |
| Solanum betaceum | Solanaceae | 1.0 | 1.5 | 0.0 | 0.0 | 3.0 | 0.3 | 1.9 |
| Vernonia amygdalina Delile | Asteraceae | 1.0 | 1.5 | 0.2 | 0.3 | 4.0 | 0.4 | 2.2 |
Fre: frequency; RF: relative frequency; Tot Dom: total dominance; RD: relative dominance; AB: abundance; RA: relative abundance; IVI: important value index.
Table A4.
List of perennial woody and non–woody plant species and their important value index under C–Ft–E based AF system, South-eastern rift–valley landscapes, Ethiopia.
Table A4.
List of perennial woody and non–woody plant species and their important value index under C–Ft–E based AF system, South-eastern rift–valley landscapes, Ethiopia.
| Scientific name | Family |
Fre
n |
RF
(%) |
Tot
Dom |
RD
(%) |
AB |
RA
(%) |
IVI
(%) |
| Albizia gummifera (J.F. Gmel.) C.A.Sm | Fabaceae | 1 | 0.8 | 0.0 | 0.0 | 4.0 | 0.3 | 1.1 |
| Annona chrysophylla | Annonaceae | 2 | 1.5 | 0.0 | 0.2 | 22.0 | 1.8 | 3.5 |
| Casimiroa edulis Lal lave and Lex | Rutaceae | 4 | 3.1 | 0.0 | 0.0 | 11.0 | 0.9 | 4.0 |
| Carica papaya | Caricaceae | 8 | 6.2 | 0.1 | 0.2 | 22.0 | 1.8 | 8.2 |
| Coffea arabica L. | Rubiaceae | 19 | 14.6 | 2.0 | 7.0 | 310.0 | 25.1 | 46.7 |
| Cordia africana Lam. | Boraginaceae | 5 | 3.8 | 0.1 | 0.5 | 16.0 | 1.3 | 5.7 |
| Ensete ventricosum ( Welw. Cheesman) | Musaceae | 20 | 15.4 | 16.9 | 58.6 | 363.0 | 29.4 | 103.4 |
| Erythrina brucei Schweinf. | Leguminosae | 1 | 0.8 | 0.0 | 0.2 | 2.0 | 0.2 | 1.1 |
| Ficus sur Forssk. | Moraceae | 1 | 0.75 | 1.5 | 5.15 | 1 | 0.1 | 6.0 |
| Ficus vasta Forsk. | Moraceae | 1 | 0.75 | 1.5 | 5.15 | 1.0 | 0.1 | 6.0 |
| Leucaena leucocephala | Mimosoideae | 1 | 0.8 | 0.0 | 0.0 | 2.0 | 0.2 | 0.9 |
| Mangifera indica L. | Anacardiaceae | 16 | 12.3 | 0.9 | 3.1 | 108.0 | 8.8 | 24.1 |
| Maytenus senegalensis | Celastraceae | 1 | 0.8 | 0.0 | 0.0 | 2.0 | 0.2 | 0.9 |
| Millettia ferruginea (Hochst.) Baker | Leguminosae | 12 | 9.2 | 0.6 | 2.1 | 64.0 | 5.2 | 16.6 |
| Musa acuminata | Musaceae | 15 | 11.5 | 3.4 | 11.8 | 234.0 | 19.0 | 42.3 |
| Persea americana Mill. | Lauraceae | 16 | 12.3 | 1.3 | 4.5 | 59.0 | 4.8 | 21.6 |
| Prunus africana | Rosaceae | 0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| Psidium guajava | Myrtaceae | 1 | 0.8 | 0.0 | 0.0 | 1.0 | 0.1 | 0.9 |
| Rhamnus prinoides L. Herit. | Rhamnaceae | 1 | 0.8 | 0.0 | 0.0 | 4.0 | 0.3 | 1.1 |
| Solanum betaceum | Solanaceae | 1 | 0.8 | 0.0 | 0.0 | 1.0 | 0.1 | 0.9 |
| Spathodea campanulata | Bignoniaceae | 3 | 2.3 | 0.4 | 1.5 | 5.0 | 0.4 | 4.2 |
| Trichilia dregeana | Meliaceae | 1 | 0.8 | 0.0 | 0.0 | 1.0 | 0.1 | 0.9 |
Fre: frequency; RF: relative frequency; Tot Dom: total dominance; RD: relative dominance; AB: abundance; RA: relative abundance; IVI: important value index.
References
- Kenrick, P. and Crane, P. R. (1997) ‘The origin and early evolution of plants on land’, Nature, 389(6646), pp. 33–39. [CrossRef]
- Parmesan, C. and Yohe, G. (2003) ‘A globally coherent fingerprint of climate change impacts across natural systems.’, Nature, 421(6918), pp. 37–42. [CrossRef]
- ICRAF (2000) ‘Paths to prosperity through agroforestry’. ICRAF’s corporate strategy, 2001– 2010. Nairobi: International Centre for Research in Agroforestry.
- Jose, S. (2009) ‘Agroforestry for ecosystem services and environmental benefits: An overview’, Agroforestry Systems, 76(1), pp. 1–10. [CrossRef]
- Das, T. and Das, A. K. (2005) ‘Inventorying plant biodiversity in homegardens: A case study in Barak Valley, Assam, North East India’, Current Science 89(1). pp.155-163.
- Harvey, C. A. and Gonzalez -villalobos, J. A. (2007) ‘Agroforestry systems conserve species-rich but modified assemblages of tropical birds and bats’, Biodiversity and Conservation, 16(8), pp. 2257-2292. [CrossRef]
- Jose, S. (2012) ‘Agroforestry for conserving and enhancing biodiversity’, Agroforestry Systems, 85(1), 1–8. [CrossRef]
- Gillespie, A. R., Miller, B. K. and Johnson, K. D. (1995) ‘Effects of ground cover on tree survival and growth in filter strips of the Cornbelt Region of the midwestern US’, Agriculture, Ecosystems and Environment, 53(3), pp. 263–270. [CrossRef]
- McNeely, J. A. and Schroth, G. (2006) ‘Agroforestry and Biodiversity Conservation – Traditional Practices, Present Dynamics, and Lessons for the Future’, Biodiversity and Conservation, 15(2), pp. 549–554. [CrossRef]
- Bhagwat, S.A., Willis, K.J., Birks, H.J.B. and Whittaker, R.J., (2008) ‘Agroforestry: a refuge for tropical biodiversity?’ Trends in ecology and evolution, 23(5), pp.261-267. [CrossRef]
- Haggar, J., Pons, D., Saenz, L., & Vides, M. (2019). Contribution of agroforestry systems to sustaining biodiversity in fragmented forest landscapes. Agriculture, Ecosystems & Environment, 283, 106567. [CrossRef]
- Kumar, B. M. and Nair, P. K. R. (2004) ‘The enigma of tropical homegardens’, in Agroforestry Systems, 61-62(1-3), pp.135–152. [CrossRef]
- Negash, M., Yirdaw, E., and Luukkanen, O. (2012) ‘Potential of indigenous multistrata agroforests for maintaining native floristic diversity in the south-eastern Rift-Valley escarpment, Ethiopia’, Agroforestry Systems, 85(1), pp. 9-28. [CrossRef]
- Negash, M. (2013) ‘The indigenous agroforestry systems of the south-eastern Rift–Valley escarpment, Ethiopia: Their biodiversity, carbon stocks, and litterfall’, Viikki Tropical Resources Institute (VITRI) PhD Dissertation, University of Helsinki, Finland. 62 p.
- Hemp, A. (2006) ‘The Banana forests of Kilimanjaro: Biodiversity and conservation of the Chagga homegardens’, Biodiversity and Conservation, 15(4), pp. 1193-1217. [CrossRef]
- Kabir, M. E. and Webb, E. L. (2009) ‘Household and homegarden characteristics in southwestern Bangladesh’, Agroforestry Systems, 75(2), pp. 129–145.. [CrossRef]
- Kehlenbeck, K., Kindt, R., Sinclair, F. L., Simons, A. J. and Jamnadass, R. (2011) ‘Non-native tree species displace indigenous ones on farms at intermediate altitudes around Mount Kenya’, Agroforestry Systems, 83(2), pp. 133-147. [CrossRef]
- Abebe, T., Wiersum, K.F., Bongers, F.J.J.M. and Sterck, F. (2006) ‘Diversity and dynamics in homegardens of southern Ethiopia’, In Tropical Homegardens (pp. 123-142). Springer, Dordrecht. [CrossRef]
- Tamrat, S., (2011) ‘Study of useful plants in and around gate Uduma (traditional Gedeo Homegardens) in Kochere Woreda of Gedeo zone, SNNPR, Ethiopia: An ethnobotanical approach’, PhD thesis, Addis Ababa University, Ethiopia, 144 p.
- Negash, M. and Achalu, N. (2008) ‘History of indigenous agroforestry in Gedeo, southern Ethiopia, Based on local community interviews: vegetation diversity and structure in the land use systems. Ethiopian Journal of Natural Resources 10(1):31−52.
- Woldeyes, F. (2011) ‘Homegardens and spices of Basketo and Kafa (Southwest Ethiopia): Plant diversity, product valorization and implications to biodiversity conservation’, PhD thesis, Addis Ababa University, Ethiopia, 222 p.
- Duguma, L. A. and Hager, H. (2010) ‘Woody plants diversity and possession, and their future prospects in small-scale tree and shrub growing in agricultural landscapes in central highlands of Ethiopia’, Small-scale Forestry, 9(2), pp. 153–174. [CrossRef]
- Mengesha, B. (2010) ‘Alternative technologies for sustainable agricultural production and agroecosystem conservation in arsi highlands, Southeastern Ethiopia’, PhD thesis.Addis Ababa University, Ethiopia. 198 p.
- Kebede, T.M. 2010. Homegardens agrobiodiversity conservation in Sebeta-Hawas Woreda, southwestern Shewa Zone of Oromia Region, Ethiopia. M.Sc. thesis. Addis Ababa University, Ethiopia. 78 p.
- Fentahun, M. and Hager, H. (2009) ‘Integration of indigenous wild woody perennial edible fruit bearing species in the agricultural landscapes of Amhara region , Ethiopia’, Agroforestry Systems, 78(1), pp. 79–95. [CrossRef]
- Haileselasie, T. H. and Gebrehiwot, M. T. (2012) ‘Agroforestry practices and flora composition in backyards in Hiwane , Hintalo Wejerat of Tigray , Northern Ethiopia’, International Journal of Biodiversity and Conservation, 4(7). pp. 294-303. [CrossRef]
- Degefa, S. (2016). Home garden agroforestry practices in the Gedeo zone, Ethiopia: a sustainable land management system for socio-ecological benefits. Socio-ecological production landscapes and seascapes (SEPLS) in Africa, 28.
- Kanshie, T. K. (2002) ‘Five thousand years of sustainability? : a case study on Gedeo land use (Southern Ethiopia)’, PhD Dissertation, Wageningen University, Netherlands. 295 p.
- Tesfay, H.M.; Negash, M.; Godbold, D.L.; Hager, H. (2022) ‘Assessing Carbon Pools of Three Indigenous Agroforestry Systems in the Southeastern Rift–Valley Landscapes, Ethiopia’, Sustainability, 14, 4716. [CrossRef]
- Asfaw, Z. and Agren. IG (2007) ‘Farmers’ local knowledge and topsoil properties of agroforestry practices in Sidama, Southern Ethiopia’, Agroforestry Systems, 71(1), pp. 35-48. [CrossRef]
- Negash, M. (2007) ‘Trees management and livelihoods in Gedeo’s agroforests, Ethiopia’, Forests, trees and livelihoods, 17(2), pp. 157–168. [CrossRef]
- Negash, M., Yirdaw, E., and Luukkanen, O. (2012) ‘Potential of indigenous multistrata agroforests for maintaining native floristic diversity in the south-eastern Rift–Valley escarpment, Ethiopia’, Agroforestry Systems, 85(1), pp. 9-28. [CrossRef]
- Abebe, T. (2013) ‘Determinants of crop diversity and composition in enset-coffee agroforestry homegardens of Southern Ethiopia’, Journal of Agriculture and Rural Development in the Tropics and Subtropics, 114(1), pp.29-38.
- Seta, T. and Demissew, S. (2014) ‘Diversity and standing carbon stocks of native agroforestry trees in Wenago district , Ethiopia’, Journal of Emerging Trends in Engineering and Applied Sciences (JETEAS), 5(7), pp. 125–132.
- National Metreology Agency (2019) ‘Climatic data of South nations and nationalities peoples regional state’, Hawassa, Ethiopia.
- Mebrate, B.T. (2007) ‘Agroforestry practices in Gedeo Zone, Ethiopia: A geographical analysis’, PhD dissertation, Panjab University, India. 188 p.
- Food and agriculture organization (FAO) (2010) ‘Global forest resource assessment’, Main report no. 163.FAO (Food and Agriculture Organization of the United Nations), Rome.
- Asfaw, Z. (2003) ‘Tree species diversity, topsoil conditions and arbuscular mycorrhizal association in the Sidama traditional agroforestry land use, Southern Ethiopia’, PhD Dissertation, Swedish University of Agricultural Sciences, Department of Forest management and products, SLU. Acta Universitatis Sueciae. Silverstria, 263 p.
- Abebe, T. (2005) ‘Diversity in Homegarden Agroforestry Systems of Southern Ethiopia’, PhD thesis, Wageningen University, the Netherlands. ISBN 90-8504-163-5; 143 pp.
- Brandt, S., Spring, A., Hiebsch, C., McCabe, J.T., Tabogie, E., Diro, M., Wolde-michael, G., Yntiso, G., Shigeta, M. and Tesfaye, S. (1997) ‘The “Tree against Hunger.” enset-Based Agricultural Systems in Ethiopia’. New York: American Association for the Advancement of Science, pp. 55.
- Zewdie, S., Fetene, M. and Olsson, M. (2008) ‘Fine root vertical distribution and temporal dynamics in mature stands of two enset (Ensete ventricosum Welw Cheesman) clones’, Plant and Soil, 305(1–2), pp. 227–236. [CrossRef]
- Shank, R. and Ertiro, C. (1996) ‘A linear model for predicting enset plant yield and assessment of Kocho production in Ethiopia’, World Food Program, Ministry of Agriculture, Southern Nation Nationalities, People Regional State, UNDP Emergencies Unit for Ethiopia, Addis Ababa, 62 p.
- Tsegaye, A. and Struik, P. C. (2001) ‘enset (Ensete ventricosum (Welw. Cheesman) kocho yield under different crop establishment methods as compared to yields of other carbohydrate-rich food crops’, Netherlands Journal of Agricultural Science, 49(1), pp. 81–94.. [CrossRef]
- Negash, A. and Niehof, A. (2004) ‘The significance of enset culture and biodiversity for rural household food and livelihood security in southwestern Ethiopia ’, Agriculture and Human Values, 21(1), pp. 61-71.
- Negash, A. (2001) ‘Diversity and conservation of enset (Ensete ventricosum Welw . Cheesman ) and its relation to household food and livelihood security in South-western Ethiopia’, PhD dissertation. Wageningen University, The Netherlands. 247p.
- Tsegaye, A. (2002) ‘On indigenous production, genetic diversity and crop ecology of enset (Ensete ventricosum (Welw.) Cheesman)’, PhD Thesis, Wageningen University, Netherlands. 198 p.
- NBE (National Bank of Ethiopia) annual report. Last accessed January 2023: https://nbebank.com/wp-content/uploads/pdf/annualbulletin/Annual%20Report%202020-2021/2020-21%20Annual%20Report.pdf.
- Labouisse, J.P., Bellachew, B., Kotecha, S. and Bertrand, B. (2008) ‘Current status of coffee (Coffea arabica L.) genetic resources in Ethiopia: implications for conservation’, Genetic Resources and Crop Evolution, 55(7), pp. 1079–1093. [CrossRef]
- Abebe, T., Wiersum, K.F. and Bongers, F., (2010) ‘Spatial and temporal variation in crop diversity in agroforestry homegardens of southern Ethiopia’, Agroforestry systems, 78(3), pp. 309–322. [CrossRef]
- Asfaw, Z. Mulata, Y. Assefa, B. Abebe, T. Duna, S. Mulugeta, G. Mebrahten, H. and Kassa, H. (2015) ‘Enhancing the role of Forestry in Building Climate Resilient Green Economy in Ethiopia: Strategy for scalling up effective forest management practices in Southern Nations, Nationalities and Peoples Regional State with particular an emphasis on agroforestry’, Center for International Forestry Research (CIFOR), 66 p.
- Mulugeta, G. and Mabrate, A. (2017) ‘Production and ecological potentials of Gedeo’s indigenous agroforestry practices in Southern Ethiopia’, Journal of Resources Development and Management 30(1), pp. 68–76.
- Dytham, C. (2003) ‘Choosing and using statistics: a biologist’s guide’, 2nd edition. Blackwell, Oxford.
- Snowdon, P., Raison, J., Keith, H., Ritson, P., Grierson, P., Adams, M., Montagu., K., Bi HQ, Burrows, W. and Eamus, D. (2002) ‘Protocol for sampling tree and stand biomass’, National Carbon accounting System, Technical report no. 31. Australian Greenhouse Office, Canberra.
- Shannon, C.E. and Weaver, W. (1949) ‘The mathematical theory of communication’, University of Illinois Press, Urbana, Illinois.117 p.
- Kent, M. and Coker, K. (1992) ‘Vegetation description and analysis; A practical approach’, Belhaven Press, London, 363p.
- Magurran, A.E. (2004) ‘Measuring biological diversity’, Blackwell Sciences, Oxford.
- Pielou, E.C. (1969) ‘An Introduction to Mathematical Ecology’, Wiley, New York. 286 p.
- Caiafa, A. N., and Martins, F. R. (2010) ‘Forms of rarity of tree species in the southern Brazilian Atlantic rainforest’. Biodiversity and Conservation, 19(9), 2597–2618. [CrossRef]
- Edwards, S. and Kelbessa, E. (1999) ‘Forest genetic resources of Ethiopia: status and proposed actions. In: Edwards S, Demissie A, Bekele T and Haase G (eds), The national forest resources conservations strategy development workshop’, Proceedings of national workshop from 21–22, June 1999 held in Addis Ababa. Institute of Biodiversity Conservation and Research (IBCR), GTZ, Addis Ababa, pp 101–133.
- Vivero, L.J., Kelbessa, E. and Demissew, S. (2005) ‘The red list of endemic trees and shrubs of Ethiopia and Eritrea’, Fauna and Flora International, Cambridge, 23 p.
- Bekele, T., Haase, G. and Soromessa, T. (1999) ‘Forest genetic resources of Ethiopia: status and proposed actions. In: Edwards S, Demissie A, Bekele T and Haase G (Eds) The national forest resources conservations strategy development workshop’, Proceedings of national workshop from 21–22, June 1999 held in Addis Ababa. Institute of Biodiversity Conservation and Research (IBCR), GTZ, Addis Ababa, pp 39-48.
- Gebremariam, A.H., Bekele, M. and Ridgewell, A. (2009) ‘Small and medium forest enterprises in Ethiopia’, IIED Small and Medium Forest Enterprise Series No. 26. FARM-Africa and International Institute for Environment and Development, London. 52 p.
- Kassa, K., Abebe, T. and Ewnetu, Z. (2015) ‘Diversity, density and management of trees in different agro-forestry practices of Yem Special District, Southern Ethiopia’, Ethiopian Journal of Science, 38(1). pp. 1-16.
- Asfaw, Z. and Woldu, Z. (1997) ‘Crop association of home-gardens in Wolayta and Gurage in southern Ethiopia. Ethiopian Journal of Science, 20(1), pp. 73-90.
- Bajigo, A. and Tadesse, M. (2015) ‘Woody species diversity of traditional agroforestry practices in Gununo Watershed in Wolayitta Zone, Ethiopia. Forest Research 4(4). pp.155. [CrossRef]
- Tefera, Y., Abebe, W. and Teferi, B. (2016) ‘Woody plants species diversity of home garden agroforestry in three agroecological Zones of Dilla Zuria District, Gedeo Zone, Southern Ethiopia’, International Journal of Fauna and Biological Studies, 3(3). pp. 98-106.
- Negawo, W.J. and Beyene, D.N. (2017) ‘The Role of coffee Based Agroforestry System in Tree Diversity Conservation in Eastern Uganda’, Journal of Landscape Ecology, 10(2), pp. 1-18. [CrossRef]
- Molla, A., and Kewessa, G. (2015) ‘Woody species diversity in traditional agroforestry practices of Dellomenna District, Southeastern Ethiopia: Implication for maintaining native woody species’, International Journal of Biodiversity, 2015(3), pp.1-13. [CrossRef]
- Okafor, J. C. and Fernandes, E. C. M. (1987) ‘Compound farms of southeastern Nigeria’, Agroforestry Systems, 5(2), pp. 153–168. [CrossRef]
- Perera, A. H. and Rajapakse, R. M. N. (1991) ‘A baseline study of Kandyan Forest Gardens of Sri Lanka: Structure, composition and utilization’, Forest Ecology and Management, 45(1-4), pp. 269–280. [CrossRef]
- Kumar, M.B., George, S. J. and Chinnamani, S. (1994) ‘Diversity, structure and standing stock of wood in the homegardens of Kerala in peninsular India’, Agroforestry Systems, 25(3), pp. 243–262. [CrossRef]
- Padoch, C., & de Jong, W. (1991) ‘The house gardens of Santa Rosa: Diversity and variability in an Amazonian agricultural system’, Economic Botany, 45(2), pp. 166–175. [CrossRef]
- Soemarwoto, O. (1987) ‘Homegadens: a traditional agrforestry system with a promising future. In: Steppler H, Nair PKR (eds) Agroforestry: a decade of development. International Council for Research in Agroforestry (ICRAF), Nairobi, pp 157–170.
- Kumari, M.S., Kansuntisukmongkol, K. and Brockelman, W.Y., (2009) ‘ Plant diversity in home gardens and its contribution to household economy in suburban areas in Sri Lanka’. Environment and Natural Resources Journal, 7(2), pp.12-29.
- Oginosako, Z., Simitu, P., Orwa, C., and Mathenge, S. (2006) ‘Are they competing or compensating on farm?: Status of indigenous and non-native tree species in a wide range of agro-ecological zones of Eastern and Central Kenya, Surrounding Mt. Kenya’, Results of vegetation, farmer, and nursery Surveys (No. 16). World Agroforestry Centre, Nairobi, Kenya. 45 p.
- Nogués-Bravo, D., Araújo, M. B., Romdal, T. and Rahbek, C. (2008) ‘Scale effects and human impact on the elevational species richness gradients’, Nature, 453(7192), pp. 216-219. [CrossRef]
- Singh, M., Kumar, A., & Sharma, M. (2021). ‘Conservation of plant diversity in agroforestry systems in a biodiversity hotspot region of northeast India’. Agricultural Research, 10(4), 569-581.
- O’Neill, G. A., Dawson, I., Sotelo-Montes, C., Guarino, L., Guariguata, M., Current, D. and Weber, J. C. (2001) ‘Strategies for genetic conservation of trees in the Peruvian Amazon basin’, Biodiversity and Conservation, 10(6), pp. 837–850. [CrossRef]
- Hailu, T., Negash, L., and Olsson, M. (2000) ‘Millettia ferruginea from southern Ethiopia: Impacts on soil fertility and growth of maize’, Agroforestry Systems, 48(1), pp. 9–24. [CrossRef]
- Michon, G., Bompard, J., Hecketsweiler, P. and Ducatillion, C. (1983) ‘Tropical forest architectural analysis as applied to agroforests in the humid tropics: The example of traditional village-agroforests in West Java’, Agroforestry Systems, 1(2), pp.117–129. [CrossRef]
- Kessy, J.F. (1998) ‘Conservation and Utilization of Natural Resources in the East Usambara Forest Reserves: Conventional Views and Local Perseptives’, PhD thesis, Wageningen University, The Netherlands. ISBN: 90-5485-809-5;168 p.
- Berhanu, A. and Asfaw, Z. (2014) ‘The role of home gardens for conservation and sustainable utilization of plant biodiversity of Ethiopia. In: Girma, A. and Wube, T. (eds). Coffee production, variety and trading ways to maximize Ethiopia’s benefits. Proceedings of 24th annual conference of the biological society of Ethiopia. pp. 81-35.
- Schroth, G., Krauss, U., Gasparotto, L. J. A. D., Duarte Aguilar, J. A., & Vohland, K. (2000) ‘Pests and diseases in agroforestry systems of the humid tropics’. Agroforestry systems, 50, 199-241.
- Jensen, M. (1993b) ‘Productivity and nutrient cycling of a Javanese homegarden’, Agroforestry Systems, 24(2), pp.187–201. [CrossRef]
- Correia, M., Diabaté, M., Beavogui, P., Guilavogui, K., Lamanda, N. and de Foresta, H. (2010) ‘Conserving forest tree diversity in Guinée Forestière (Guinea, West Africa): the role of coffee-based agroforests’, Biodiversity and Conservation, 19(6), pp. 1725–1747. [CrossRef]
- Asase, A. and Tetteh, D. A. (2010) ‘The role of complex agroforestry systems in the conservation of forest tree diversity and structure in Southeastern Ghana’, Agroforestry Systems, 79(3), pp. 355-368. [CrossRef]
- Abreha, A. and Gebrekidan, W. (2014) ‘Woody plant inventory and diversity in traditional agroforestry of selected peasant association of South Gonder Zone, North West Ethiopia’, Journal of Environment and Earth Science, 4(15). pp.1-16.
- Körner, Ch. (2000) ‘Why are there global gradients in species richness? Mountains might hold the answer’, Correspondence, 15(12). pp.513-514.
- Wang, W., Wang, Q., Li, S. and Wang, G. (2006) ‘Distribution and Species Diversity of Plant Communities along Transect on the Northeastern Tibetan Plateau’, Biodiversity and Conservation, 15(5), pp. 1811-1828. [CrossRef]
- Shimono, A., Zhou, H., Shen, H., Hirota, M., Ohtsuka, T. and Tang, Y. (2010) ‘Patterns of plant diversity at high altitudes on the Qinghai-Tibetan Plateau’, Journal of Plant Ecology, 3(1), pp. 1-7. [CrossRef]
- Luzuriaga, A.L. and Escudero, A. (2011) ‘What Factors Affect Diversity and Species Composition of Endangered Tumbesian Dry Forests in Southern Ecuador’? Biotropica, 43(1) pp. 15–22. [CrossRef]
- Ma, M. (2005) ‘Species richness vs evenness: independent relationship and different responses to edaphic factors’, Oikos 111(1), pp. 192-198. [CrossRef]
Figure 7.
Relationship between DBH and BA (A); stem number and BA (B) for the enset species of the three studied AF systems.
Figure 7.
Relationship between DBH and BA (A); stem number and BA (B) for the enset species of the three studied AF systems.
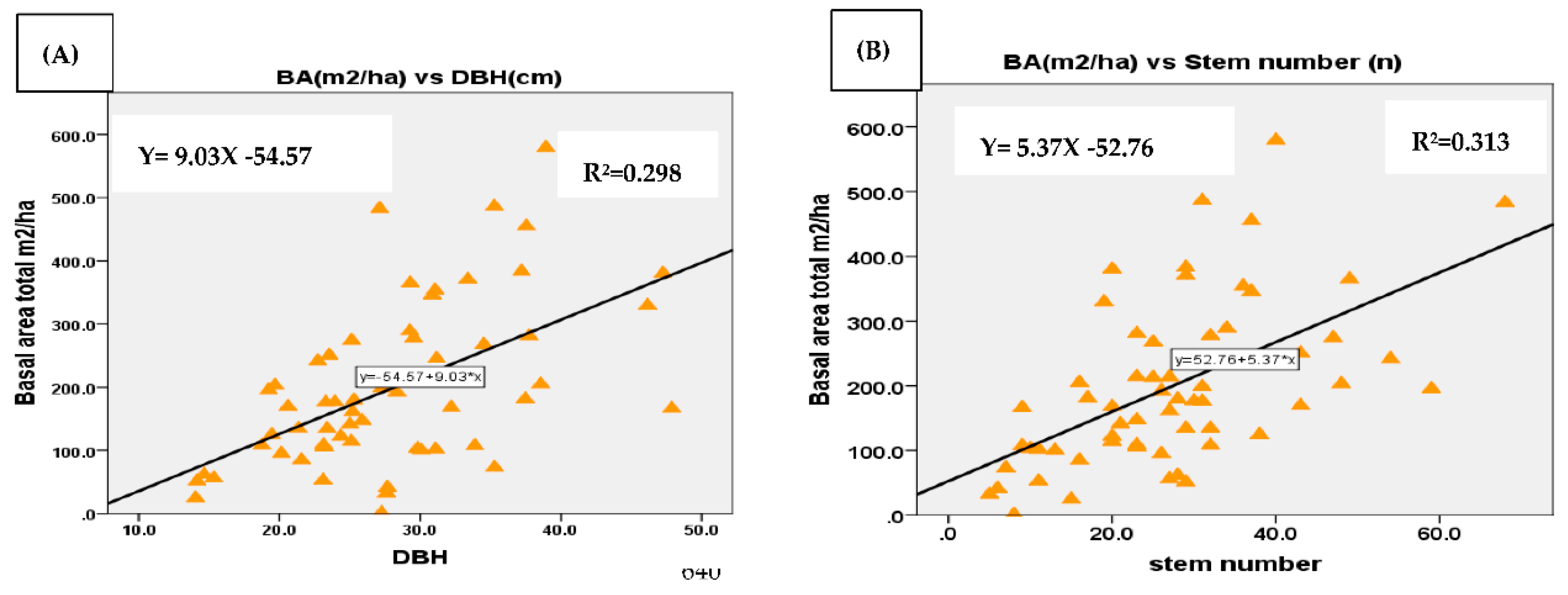
Figure 8.
Relationship between basal area and diameter at breast height (DBH) (A); basal area and stem number (B) for the woody species of the three studied AF systems.
Figure 8.
Relationship between basal area and diameter at breast height (DBH) (A); basal area and stem number (B) for the woody species of the three studied AF systems.
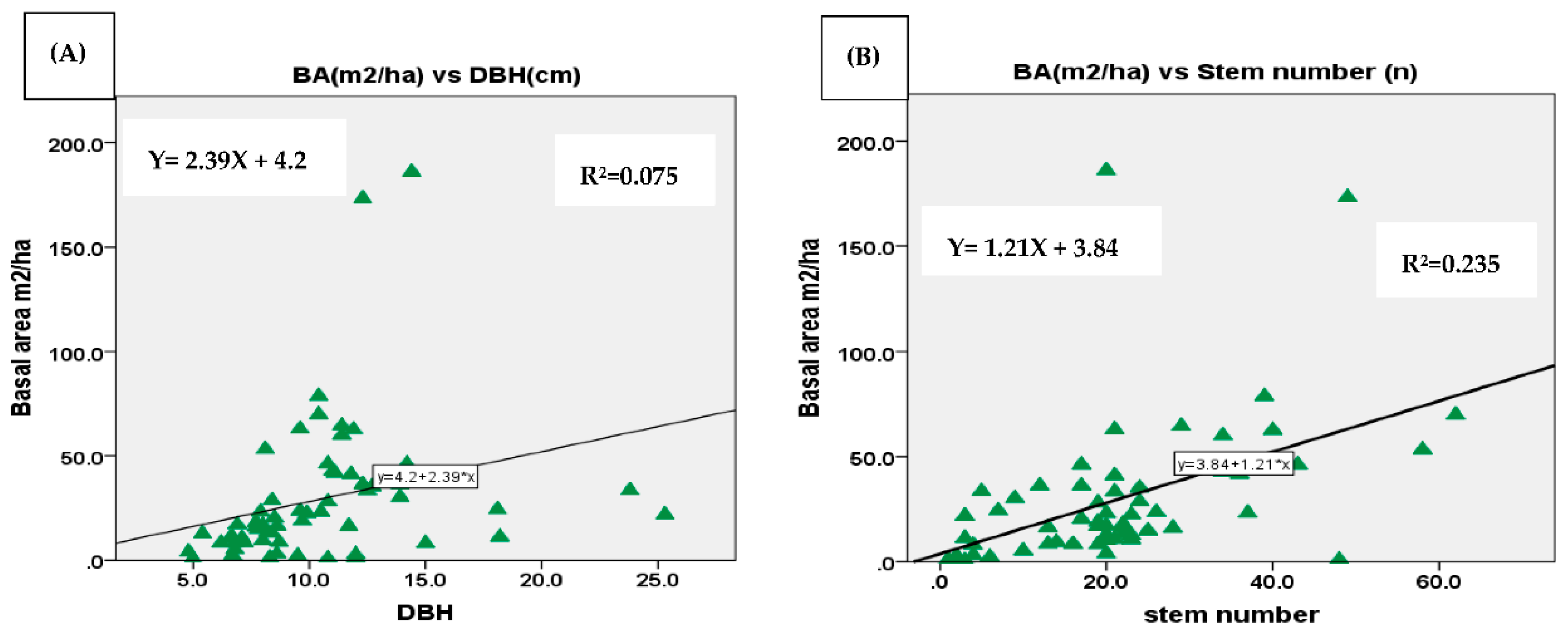
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



