
Submitted:
06 December 2023
Posted:
07 December 2023
You are already at the latest version
Abstract
The liver health of Gilthead sea bream and European sea bass fed fishmeal-free diets including various proportions of plant proteins, insect and poultry by-product meals, was investigated through biochemical and histological analyses using a new Liver Index (LI) formula. Four isoproteic (45% DM) and isolipidic (20% DM) diets were compared: a control, plant-based diet (CV) and other three test ones where 40% of a plant protein-rich ingredient mixture was replaced by Hermetia illucens (H40) or poultry by-product (P40) meals alone or in combination (H10P30). The trials lasted 12 and 18 weeks for sea bream and sea bass, respectively. The results so far ob-tained highlighted species-specific differences in the physiological response to dietary changes. In sea bream, the biochemical and histological responses suggest favorable physiological status and liver health, with higher serum cholesterol (CHO) and triglyceride (TAG) levels and moderate hepatocyte lipid accumulation with the H10P30 diet compared to CV (p<0.05). In sea bass, all diets resulted in elevated serum TAG levels and liver lipid accumulation particularly in fish fed the P40 one (p<0.05) which resulted in the highest LI, coupled with higher frequency of severe lipid accumulation, hypertrophy, cord loss, peripheral nuclei displacement and pyknosis.
In conclusion sea bream well adapted to the test diets, while sea bass exhibited altered hepatic lipid metabolism leading to incipient liver steatosis, likely due to the high lipid content of the diets including the insect and poultry meals. The LI developed in this study proved to be a reliable tool for assessing the effects of dietary changes on liver health of sea bream and sea bass, consistent with biochemical and histological findings.
Keywords:
liver histology
; liver index
; sustainable feed
; poultry by-product meal
; Hermetia illucens meal
; plant-protein
; lipid metabolism
; fish welfare
; blood biochemistry
1. Introduction
World population is projected to reach around 8.5 billion people in 2030, and aquaculture is expected to provide approximately 53% of the world's seafood supply, significantly contributing to global food security and sustainable development [1,2,3]. Global aquaculture production of fed-species is projected to reach 58.9 million tons by 2025, with an estimated demand for 69.5 million tons of aquafeed [4]. To support this growth, the industry will need to explore innovative solutions for sourcing sustainable ingredients to increase aquafeed production [5]. Over the past two decades, the transition towards the use of plant-based ingredients to reduce dependence on fish meal and fish oil has been challenging [6,7]. Similarly, the shift towards alternative ingredients generated through the circular bioeconomy will require significant efforts to achieve a sustainable balance between food production and resources conservation, as well as to enhance climate change resilience within aquafeed production and aquaculture farming systems [8,9,10,11]. Currently, several raw materials of plant-origin and animal by-products are employed as ingredients in commercial feeds. However, high levels of substitution in feeds for carnivorous fish can lead to adverse physiological effects on digestibility and nutrient utilization, growth, metabolism, gut integrity, immune response, disease resistance, and overall health and welfare, which are still under study [12,13].
Poultry by-product and insect meals have been receiving significant attention as alternative protein sources to both fish and plant protein-rich meals. This is owing to their characteristics associated with circularity, environmental sustainability, and minimal conflict for direct human consumption. Poultry by-product meals are cheap, easily available and with a high nutritional value to carnivorous fish species being rich in crude protein with a nearly complete amino acid composition and as sources of minerals and vitamins [14,15]. Among edible insect species, processed meals derived from the black soldier fly, Hermetia illucens (Diptera order), are also rich in protein with a high biological value and an essential amino acid profile comparable to fish meal [16,17,18,19]. They also contain lipids and bioactive compounds that provide beneficial effects on fish health, by stimulating the immune system and modulating the gut microbiota [20]. Deficiencies in omega-3 fatty acids and some vitamins can be improved through the selection of high-quality feeding substrates [21]. Currently, insect meal production is still limited and expensive, but the intensification of industrialized insect production system could significantly increase volume and reduce production costs to meet the demands of the aquafeed industry [22].
To date, many studies have examined the fish meal replacement with poultry by-products [23,24,25] and insect meals [26,27,28,29,30,31,32,33,34,35,36,37]. In case of Gilthead sea bream, Sparus aurata and European sea bass, Dicentrarchus labrax, the results obtained so far provided broad support to the use of both alternative protein sources as substitutes of fish meal to develop sustainable aquafeeds.
On the other hand, only a few research studies have evaluated the partial replacement of plant protein-rich feeds with insect and poultry by-product meals, singly or combined, in low fish meal diets in sea bream [38,39] and sea bass [40]. These studies mostly investigated the dietary effects on appetite, growth performance, gut health and fish quality, while fish response in terms of liver health status have received limited attention.
Indeed, in fish as in other vertebrates, the liver plays a key role in energy metabolism and storage, synthesis of bile, proteins and hormones, other than in immune defense and detoxification [41,42]. Due to its role in metabolizing products from the digestive tract, the liver is also considered a valuable marker of nutritional pathologies [43]. Therefore, histopathology, which provides an analytical view of disease and its impact on tissues and cells, represents a valuable tool in studies on fish nutrition, particularly in feeding trials aimed at evaluating the suitability of alternative ingredients for innovative aquafeed [44,45]. Histological qualitative and semi-quantitative methods to evaluate hepatic tissue alterations have been widely applied in farmed fish to assess the effects of different diets on liver health. General morphology, lipid deposition, and glycogen distribution in the cytoplasm have been described and considered valid parameters for comparing the liver status with respect to the test diets in sea bream and sea bass [24,46,47,48]. In other studies evaluating the tolerance of these species to fish meal replacement with novel dietary ingredients, a semi-quantitative scoring system was applied, and several parameters including nucleus shape and nuclear displacement, hypertrophy, cytoplasmic lipid accumulation in hepatocytes, peripancreatic fat infiltration were evaluated using graded system scales [25,36,49,50,51,52,53]. Some studies applied the semi-quantitative Bernet's protocol [54], an organ-by-organ evaluation system originally designed to monitor the health status of fish exposed to environmental pollution [55,56,57,58,59]. This protocol was subsequently adapted to evaluate diet-related alterations [60], general health status of farmed fish [61] and the quality of their larval and juvenile stages [62]. The method used by Bernet is the subject of a recent review proposing standardization for histological evaluation to generate reliable, high-quality, and comparable data in fish histology [63].
Additionally, histological quantitative methods involving the analysis of morphometric parameters, such as hepatocyte area, hepatocyte nuclear area, and their relative ratios, as well as the area of lipid droplets, were frequently used as indicators of hepatocyte metabolic activity in several farmed fish species, including sea bream and sea bass [64,65,66,67,68,69,70].
The present study is part of a larger experiment where ten different diets including graded levels of poultry by-product meal and H. illucens meal either alone or in combination were included in a plant protein-rich, fish meal free basal formulation. Here, we specifically focused on the effects of the diets with the highest level of substitution on liver health status in Gilthead sea bream and European sea bass, the most farmed and commercially important species in the Mediterranean [71]. For this purpose, the histological semi-quantitative scoring system proposed by Bernet et al., [54] was modified by developing a new formula for calculating the Liver Index (LI) and its relative scoring scheme. Complementary blood biochemistry analysis was performed to evaluate the nutritional-metabolic status of the fish. Additionally, body condition indices and gross liver anatomy were analyzed to provide a comprehensive assessment of liver health status and evaluate its potential consequences on fish physiology in response to dietary treatments.
2. Materials and Methods
2.1. Test diets
Four test diets were formulated to be grossly isoproteic (45%), isolipidic (20%), and isoenergetic (21 MJ kg-1). A control diet rich in plant-derived ingredients (denoted CV), was designed to obtain a 90:10 weight ratio between vegetable and marine protein and a 67:33 weight ratio between vegetable and fish lipid. Weight ratio of ingredients was calculated from the crude protein and lipid contribution to the whole diet of marine and plant-based feed ingredients.
From the basal formulation above, three other test diets were prepared by replacing 40% of crude protein from the vegetable protein-rich ingredient mixture of the CV with an equal amount of partially defattened H. illucens prepupae meal (H40 diet; ProteinX™; Protix Company Dongen, Netherlands; ) and poultry by-product meal (P40 diet; Agricola Tre Valli s.r.l., Villaganzerla, Vicenza, Italy) or a combination of the two ingredients (H10P30 diet), while maintaining the same 67:33 vegetable to fish lipid ratio as in the CV diet. Marine derivatives, based on a mixture of squid meal and fish protein concentrate, were used at 5.5% concentration in all diets as feed-attractant/stimulant. Wherever necessary, pure essential amino acids were supplemented to the diets to meet the nutrient requirement of sea bream and sea bass (National Research Council (NRC), 2011). All diets were manufactured as extruded pellet (3 and 5 mm) by SPAROS Lda (Olao, Portugal) and stored in sealed plastic buckets in a cool room until used. Proximate composition analysis of the test diets was performed according to the AOAC methods [72]. All diets were analyzed for dry matter (105 °C for 24 h), ash by combustion in a muffle furnace at 500 °C for 5 h (Nabertherm L9/11/B170, Bremen, Germany), crude protein (N × 6.25) using the Kjeldahl system (K350 Buchi, Switzerland) and crude fat according to Bligh and Dyer as modified by [73]. Gross energy was determined in an adiabatic bomb calorimeter (Model Werke C2000, IKA, Staufen, Germany). The ingredients and proximate composition of the test diets are shown in Table 1.
2.2. Fish husbandry and sampling
Two experiments on sea bream and sea bass were carried out at the aquaculture facilities of the Department of Agricultural, Food, Environmental and Animal Sciences of the University of Udine.
For this study 216 sea bream (48.8 ± 8.8 g) and sea bass (44.0±5.1g) juveniles, were randomly divided into 12 fiberglass of 250 L tanks (18 individuals per tank) included in a marine recirculated aquaculture system.
Water temperature and salinity were kept at 23.0±1.0°C and 29 ppt for both sea bream and sea bass, and fish were held under a 12L:12D photoperiod. Fish were acclimated for two weeks to culture conditions and fed a commercial diet before being randomly assigned in triplicate to the four dietary treatments.
Fish were fed to satiety, twice a day for six days a week, by belt feeders. The trial lasted 12 weeks for sea bream and 18 weeks for sea bass, up to triple the respective initial body weight. All fish were sampled at the end of the experiment after 24 h fasting.
Five fish per tank of both fish species (i.e. 15 fish per dietary treatment, n= 60 fish) were rapidly anaesthetized with Tricaine methanesulfonate MS222 at a dose of 150 mg/L (PHARMAQ, Fordingbridge Hampshire, UK) and reached a deep stage of anaesthesia within 3 min. Blood samples were drawn from the caudal vein and after clotting were centrifuged at 3000 rpm for 10 min at 4 °C. Serum aliquots were deep-frozen at −80 °C until analysis according to Di Marco et al., [74].
Fish were then sacrificed with an overdose of MS222 at 300 mg/L and individual body weight (BW) and total length (TL) were recorded. Fish autopsy was performed by observing gross anatomy and the occurrence of macroscopic alterations. In particular the liver of all sampled fish was observed in situ and after dissection before being, weighted to the nearest 0.01 g. Low, medium and high degree of lipid accumulation was assigned to each liver by two operators based on gross diffusion of lipid depots, changes in color and consistency. Samples of liver were further collected from 6 specimens for each dietary treatment, fixed in Bouin’s solution for 24 h, washed and preserved in 70% ethanol for histological analysis.
Based on biometric data collection, fish condition indices were calculated as follows:
Fulton’s condition factor (K) = 100 x body weight/lengthˆ3
Hepatosomatic index (HSI) = 100 x (liver weight/body weight)
2.3. Liver histological analyses
Small liver sub-samples were dehydrated, clarified and paraffin (Bio-Optica, Milan, Italy) embedded following standardized protocols for examination of hepatic tissue morphology. For each liver, slides were prepared by mounting three sections (5μm thick), at 100 µm intervals. Two slides were stained with Haematoxylin-Eosin (H-E, Bio-Optica, Milan, Italy) for hepatocyte morphology and one slide with Periodic Acid Schiff (PAS; Bio-Optica, Milano, Italy) to highlight glycogen deposition in hepatocytes. Before the histological examination, coded slides were quickly checked under a light microscope at 100x magnification to eventually exclude those presenting technical tissue artefacts. General histo-morphology was firstly evaluated for the description of the hepatic tissue in all the specimens according to test diets. For histomorphometric analysis and histo-pathological evaluation, 9 fields (0.320596 mm2 each) were randomly selected in three liver sections under a DMLB microscope (Leica, Germany) and 9 microphotographs (216 for sea bream and 216 for sea bass, in total) were acquired with a Leica ICC50-HD camera at 200x magnification. All histological observations were performed by experienced personnel in two independent, blinded evaluations.
2.3.1. Histomorphometric analysis
Hepatocyte and nucleus area were measured (90 cells per fish) on microphotographs using the Image J 1.41 software [44] and nucleus/hepatocyte (N/H) ratio was calculated. The degree of lipid accumulation (seen as cytoplasmatic vacuoles) and nucleus position were recorded, on the same microphotograph, for each measured hepatocyte, for the classification into mild, moderate and severe lipid accumulation classes, and subsequently for the comparison of measurements as a function of dietary treatment.
2.3.2. Histopathological evaluation
For each specimen (n=6/dietary treatment), the presence/absence of 18 histopathological alterations (Table 2) was blindly evaluated in sea bream and sea bass liver according to the protocol proposed by Bernet et al., [54], modified for this study. Briefly, each histological alterations was checked in 9 randomly selected fields and classified into five reaction patterns: circulatory disturbance, regressive and progressive changes, inflammation, and tumors. For each alteration, an importance factor (w), ranging from 1 to 3 score (1 minimal, 2 moderate and 3 marked pathological importance), was assigned. An importance factor w=1 was attributed both for mild and moderate lipid accumulation characterized by increasing lipid accumulation and nucleus still centrally or para-centrally located. The score w=2 was assigned in presence of severe lipid accumulation with vacuoles completely filling the cytoplasm of the hepatocyte and consequent nucleus displacement at the periphery.
Due to the uneven distribution within the hepatic tissue, the frequency percentage of histological alterations within each of the 9 fields was also considered by assigning a grading score ranging from 0 to 6 as follows: 0 = absent; 2 = low ≤10%; 4 = medium 10–50%; 6 = high ≥50%. Intermediate values were not considered [57]. Peri-pancreatic fat infiltration was evaluated in sea bream by using the above 0-6 grading scale on three whole sections per liver at 50x magnification. This parameter was not included among the 18 histopathological alterations used for the calculation of the liver index.
2.3.3. Liver index calculation
A liver index formula was developed based on the scoring system reported by Bernet et al., [54] and modified in this study. The formula takes into account the percentage ratio between the sum of the weighted scores of observed histological alterations and the sum of the maximum achievable weighted scores, assigning a score of 6 to all examined alterations.
The liver index (LI) was calculated for each fish as follows:
where:
i - ith fish
sj - is the score assigned to each histological alteration (j) from 0 to 6, considering the frequency percentage within each field, as described above;
wj - is the importance factor expressing the severity of the histological alteration (j);
a - is the number of the observed fields;
sj - is the maximum score attainable for each histological alteration
This index indicates the extent and severity of liver histological alterations, enabling the conversion of qualitative observations into a quantitative value.

Table 2.
Histological alterations (j) and importance factors (w) considered for histopathological liver evaluation in sea bream and sea bass fed test diets.
Table 2.
Histological alterations (j) and importance factors (w) considered for histopathological liver evaluation in sea bream and sea bass fed test diets.
| Reaction pattern | Histological alteration (j) | Importance factor (w) |
|---|---|---|
| Circulatory disturbance | Sinusoid congestion | w= 1 |
| Blood vessel congestion | w= 1 | |
| Haemorragies | w= 2 | |
| Regressive changes | Mild and moderate lipid accumulation | w=1 |
| Severe lipid accumulation | w= 2 | |
| Peripheric nuclei | w= 1 | |
| Pycnotic nuclei | w= 2 | |
| Cord loss | w= 2 | |
| Vacuolar tissue degeneration | w= 2 | |
| Tissue necrosis | w= 3 | |
| Progressive changes | Hepatocyte hyperplasia | w= 2 |
| Hepatocytes hypertrophy | w= 2 | |
| Bile duct hypertrophy | w= 1 | |
| Inflammation | Granulocyte infiltration | w= 2 |
| MMc occurrence | w= 1 | |
| Tumour | Benign tumour | w= 2 |
| Malignant tumour | w= 3 |
2.4. Blood biochemistry analyses
A set of metabolic parameters, including glucose (GLU), triglycerides (TAG), cholesterol (CHO), total protein (TP), albumin (ALB), aspartate aminotransferase (AST and alanine aminotransferase (ALT) were analyzed on serum samples using an automated biochemistry analyser (BPC BIOSED, Rome, Italy) and commercial kits (Giesse Diagnostics, Rome, Italy) according to Cardinaletti et al. [75].
2.5. Statistical analysis
Datasets on fish size, body condition indices, hepatocyte measurements, liver index and blood chemistry parameters were analyzed by the Kruskal-Wallis H-test followed by post-hoc multiple comparisons with Bonferroni adjustment, to evaluate the effects of the dietary treatment (p<0.05). The Chi-square test was used to evaluate differences in the frequency percentage of the affected area by the histological alterations.
In this study, the focused principal components analysis (FPCA) was applied to evaluate the relationship between both biometric and biochemical parameters and liver index, this latter considered the variable of interest, as well as to examine the existing correlations among the variables themselves. The graphical visualization of FPCA analysis shows correlations as concentric circles, with the variable of interest positioned at the centre, directing the analysis. Stronger correlations are represented by smaller circles. A red circle is used to indicate statistical significance at p<0.05. Negative and positive correlations are distinguished in the graph using yellow and green dots respectively. The interpretation of existing correlations between explanatory variables follows the same criteria of classical PCA analysis. The data analysis was carried out using the R software statistical package [76].
2.6. Ethical approval
The present study was approved by the Ethics Committee of the University of Udine and authorized by the Italian Ministry of Health (n.290/2019-PR) in accordance with European legal frameworks on the protection of animals used for scientific purposes (Directive 2010/63/EU).
3. RESULTS
3.1. Biometric measurements and fish condition
All the diets were highly palatable and were well accepted. No mortality occurred throughout the trial. Growth and body condition indices of sea bream and sea bass are shown in Table 3. In sea bream, the final BW of samples specimens was significantly higher in fish fed H40, P40, and H10P30 diets compared to the controls (CV). The HSI values were similar in sea bream fed CV, H40, and P40 diets, while the HSI value was significantly higher in fish fed H10P30 diet compared to P40 dietary group. In sea bass, no significant differences were observed in final BW due to dietary treatment. The K index was significantly higher in fish fed H10P30 diet compared to P40 dietary group, while no significant differences were observed in HSI values.
Post-mortem analysis showed an overall good health status of sea bream and sea bass fed test diets. The macroscopic observation revealed a clear difference in the liver appearance between sea bream and sea bass. Sea bream showed low to medium degree of lipid accumulation in all dietary groups, while the high degree was rarely observed. Indeed the liver color was not homogeneous in sea bream specimens, ranging from dark red to light brown with a few cases showing areas of pale color. The consistency of the organ was maintained in most of the samples examined. Conversely, most of sea bass displayed a high degree of liver lipid accumulation in all dietary groups, whereas the moderate degree was observed in a few specimens and the mild degree was not found. The livers of sea bass presented diffuse areas ranging from whitish to light yellowish in color, leading to easy fragmentation in most cases.
3.2. Liver histology
In most cases, the structure of the hepatic parenchyma exhibited irregular cords arranged in two-cellular layers of hepatocytes surrounded by sinusoids. The roundish polygonal hepatocytes appeared moderately eosinophilic and displayed a clear spherical nucleus positioned centrally or peripherally, depending on the degree of lipid accumulation. The occurrence of alterations, varying in extent and severity based on diet and species, and mostly associated with lipid accumulation, is detailed in the section 3.2.2. In both species, the exocrine pancreatic tissue appeared well organized within the hepatic structure, surrounding the branches of the hepatic portal vein. Medium to high frequency percentage of peripancreatic fat infiltration was observed in sea bream (Figure 1A), without any significant difference among dietary groups.
3.2.1. Histomorphometry
In sea bream, the mean area of hepatocytes with a moderate lipid accumulation was significantly higher (165.4 µm2) than in hepatocytes with a mild lipid accumulation (97.4 µm2), while the nucleus area and the nucleus/hepatocyte ratio were significantly smaller (Table 4). Similarly, in sea bass, the mean cell area significantly increased based on mild (119.1 µm2), moderate (205.3 µm2) and severe (372.1 µm2) lipid accumulation. In this species, a significant decrease of mean nucleus area was only observed between mild and severe lipid accumulation, while a strong significant decrease was observed for the nucleus/hepatocyte ratio among all classes of lipid accumulation.
Histomorphometric differences in hepatocytes were also observed as a function of dietary treatment (Table 5). In sea bream and sea bass, the mean cell area was significantly larger in the replaced-dietary groups compared to CV group, while the mean nucleus area and nucleus/hepatocyte area ratio were significantly smaller. The largest hepatocytes were found in sea bream fed H10P30 diet (177.9±50.5 µm2) and in sea bass fed P40 diet (428.1 ± 23.5 µm2) compared to the other dietary treatments.
3.2.2. Histopathology
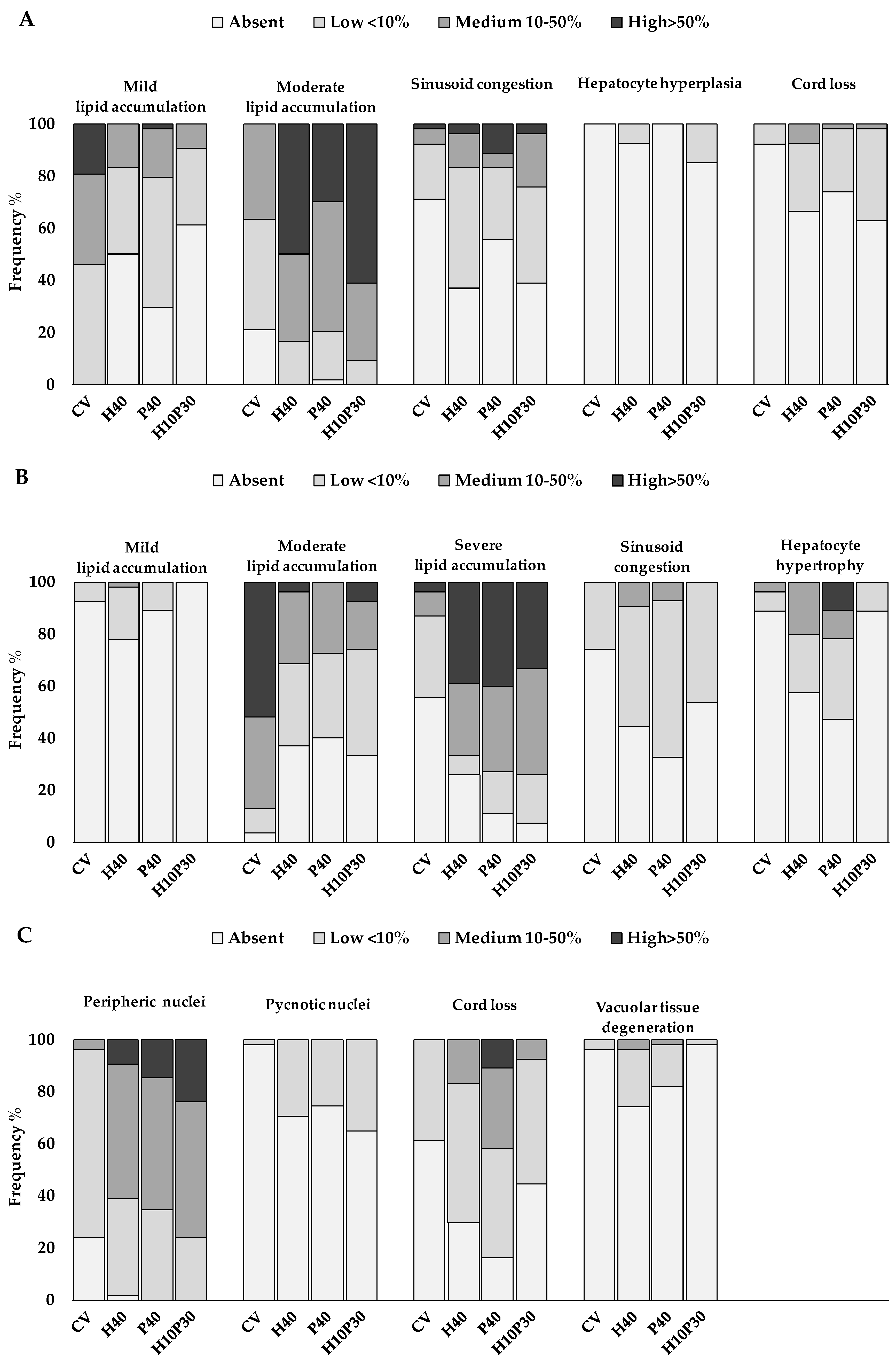
The main histological alterations observed in sea bream and sea bass livers are illustrated in Figure 1 and Figure 2, while the frequency percentage of significant histological alterations is presented in Figure 3. In sea bream, significant differences in response to dietary treatment were observed for hepatocyte lipid accumulation (p<0.001), sinusoid congestion (p<0.005), cell hyperplasia (p<0.05), and cord loss (p<0.01). In particular, mild and moderate lipid accumulation were observed in the liver of sea bream (Figure 1B,C). Moderate lipid accumulation was prevalent in fish fed replaced-diets compared to the CV diet. In particular, considering the frequency percentage of this alteration greater than 10% (i.e. sum of score 4 and 6), the frequencies observed were 83.3% in H40, 79.6% in P40, 90.7% in H10P30 compared to CV (37%) (Figure 3A). Sinusoid congestion was prevalent in sea bream fed H40 (63%), H10P30 (61.1%), and P40 (44.5%) diets (Figure 1D), compared to CV (29%). A low frequency of cell hyperplasia (14.8%) (p<0.05) and moderate cord loss (35%) were also observed, particularly in sea bream fed the H10P30 diet (Figure 1E). No significant differences were observed in blood vessel congestion (12.0%, 9.3%, 9.3%, and 16.7% in CV, H40, P40, H10P30 dietary groups) (Figure 1F) nor in granulocyte occurrence (13%, 11.3%, 14,8.%, and 5.6% in CV, H40, P40, H10P30 dietary groups) (Figure 1G). Tissue necrosis was observed in all dietary groups in a non-significant manner (Figure 1H).
Sea bass showed a worse histopathological picture compared to sea bream, in terms of number and severity of histological alterations observed (Figure 3B,C). CV group was characterized by moderate lipid accumulation (87%) (p<0.001) (Figure 2A), while fish fed test diets were mainly characterized by severe lipid accumulation (66.7%, 72.7%, 74.0% in H40, P40, H10P30 respectively) (p<0.001) (Figure 2B). Additional alterations related to severe lipid accumulation were observed in sea bass, including nucleus peripheral displacement (Figure 2C), hepatocyte hypertrophy (Figure 2D), cord loss (Figure 2E) (p<0.001), tissue vacuolar degeneration (p<0.01), accounting for 100%, 52.7%, 80.6%, and 18.2% respectively in P40 diet compared to the others. This group also presented higher frequency percentage of sinusoid congestion (52.7%) (p<0.001) (Figure 2F) and a limited frequency percentage (5.4%) of granulocytes, without significant differences among dietary groups (Figure 2G). Only a few pyknotic nuclei were observed in sea bass fed H40, P40, and H10P30 dietary groups but higher respect to the CV diet (p<0.001). Tissue necrosis occurred with low frequency and did not significantly differ among groups (Figure 2H). Bile ducts were patent in all groups and no hemorrhagic pattern was ever observed either in sea bream or sea bass liver.
Table 4.
Histomorphometric measurements of hepatocytes as a function of lipid accumulation level in sea bream and sea bass (n=90 cells per fish).
Table 4.
Histomorphometric measurements of hepatocytes as a function of lipid accumulation level in sea bream and sea bass (n=90 cells per fish).
| Lipid accumulation | Kruskal Wallis test | ||||
|---|---|---|---|---|---|
| Mild | Moderate | Severe | |||
| Sea bream | Hepatocyte area (µm2) | 97.4±13.4a | 165.4±37.9b | - | p< 0.001 |
| Nucleus area (µm2) | 16.5±1.8a | 13.0±1.2b | - | p< 0.001 | |
| N /H ratio | 0.17 ± 0.03a | 0.08 ± 0.02b | - | p< 0.001 | |
| Sea bass | Hepatocyte area (µm2) | 119.1 ±17.1a | 205.3 ± 40.1b | 372.1 ± 175.1c | p< 0.001 |
| Nucleus area (µm2) | 15.5 ± 0.36a | 14.6 ± 1.4a | 12.32 ± 1.3b | p< 0.001 | |
| N/H ratio | 0.13 ± 0.02a | 0.07 ± 0.02b | 0.04 ± 0.01c | p< 0.001 | |
Data are expressed as mean±sd. Different superscripts indicate significant differences between groups set a p< 0.05.
Table 5.
Histomorphometric measurements of hepatocytes based on dietary treatment in sea bream and sea bass (n=90 cells per fish).
Table 5.
Histomorphometric measurements of hepatocytes based on dietary treatment in sea bream and sea bass (n=90 cells per fish).
| Test diets | Kruskal Wallis | |||||
|---|---|---|---|---|---|---|
| CV | H40 | P40 | H10P30 | test | ||
| Sea bream | Hepatocyte area (µm2) | 109.9±26.8a | 146.5±33.1b | 135.7±38.8b | 177.9±50.5c | p< 0.001 |
| Nucleus area (µm2) | 16.2±2.4a | 13.8±1.2b | 14.1±1.9b | 12.6±1.2c | p< 0.001 | |
| N/H ratio | 0.16 ± 0.05a | 0.10 ± 0.03b | 0.12 ± 0.05b | 0.08 ± 0.03c | p< 0.001 | |
| Sea bass | Hepatocyte area (µm2) | 199.2 ±44.9a | 277.4 ± 89.6b | 428.1 ± 23.5c | 247.6 ± 56.5b | p< 0.001 |
| Nucleus area (µm2) | 15.5± 0.9a | 13.32 ± 1.6b | 11.87 ±1.0c | 13.26 ± 1.5b | p< 0.001 | |
| N/H ratio | 0.08 ± 0.02a | 0.06 ± 0.03b | 0.03 ± 0.01c | 0.06 ± 0.02b | p< 0.001 | |
Data are expressed as mean±sd. Different superscripts indicate significant differences between groups set a p< 0.05.
Figure 1.
Main histological alterations in liver of S. aurata fed test diets. A) CV diet: large areas of peri-pancreatic fat infiltration; B) H40 diet: mild hepatocyte lipid accumulation and central round nucleus (arrow); C) P40 diet: moderate hepatocyte lipid accumulation and round nucleus still centrally or para-centrally located (arrow); D) P40 diet: sinusoid congestion; E) H10P30 diet: hepatocyte hyperplasia and cord loss (arrowhead); F) H40 diet: blood vessel congestion; G) P40 diet: focus of granulocytes; H) P40 diet: tissue necrosis area. BV=blood vessel; P=exocrine pancreas; Gr=granulocytes; S=sinusoid; TN=tissue necrosis;. Haematoxylin and eosin staining.
Figure 1.
Main histological alterations in liver of S. aurata fed test diets. A) CV diet: large areas of peri-pancreatic fat infiltration; B) H40 diet: mild hepatocyte lipid accumulation and central round nucleus (arrow); C) P40 diet: moderate hepatocyte lipid accumulation and round nucleus still centrally or para-centrally located (arrow); D) P40 diet: sinusoid congestion; E) H10P30 diet: hepatocyte hyperplasia and cord loss (arrowhead); F) H40 diet: blood vessel congestion; G) P40 diet: focus of granulocytes; H) P40 diet: tissue necrosis area. BV=blood vessel; P=exocrine pancreas; Gr=granulocytes; S=sinusoid; TN=tissue necrosis;. Haematoxylin and eosin staining.
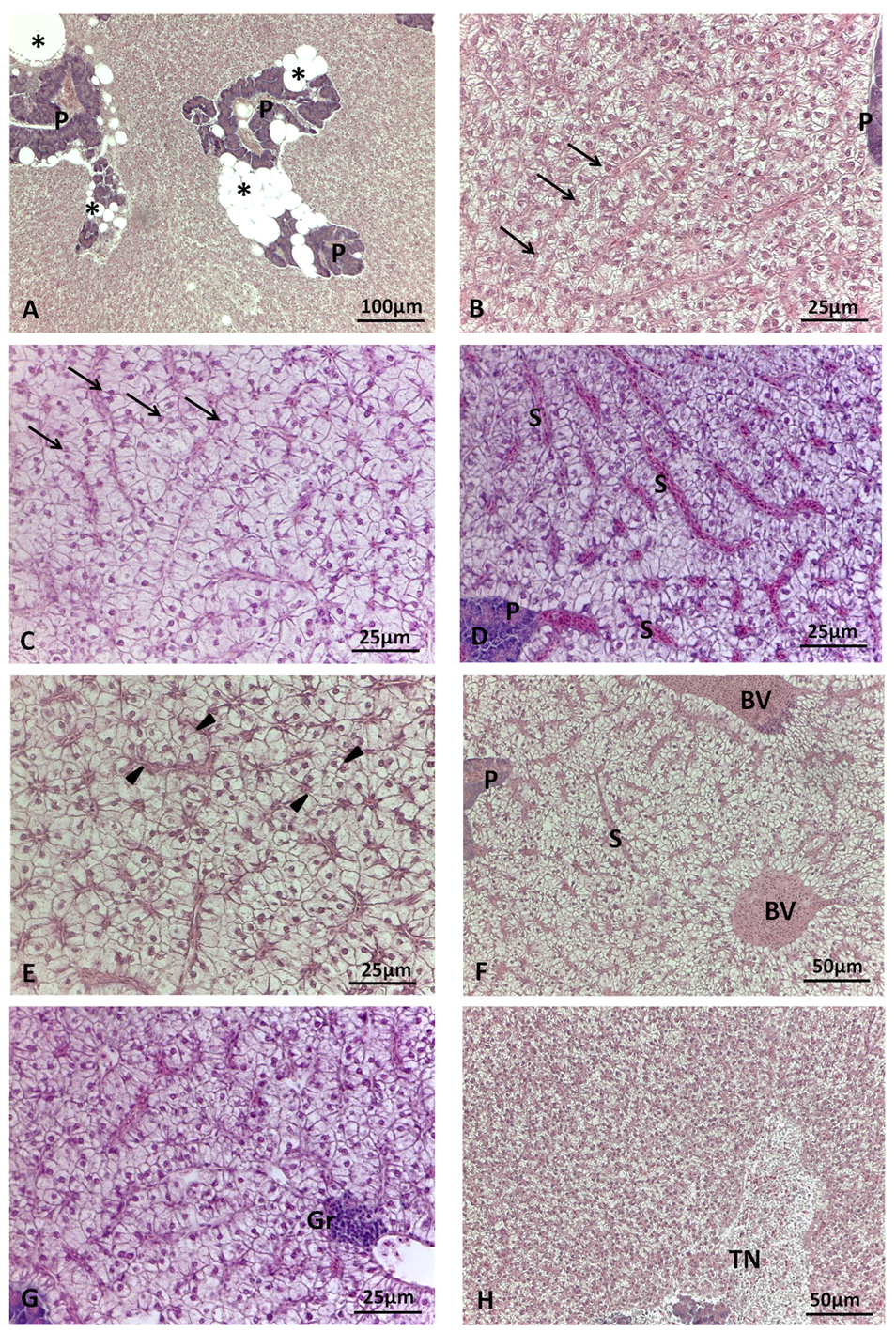
Figure 2.
Main histological alterations in liver of D. labrax fed test diets. A) CV diet: moderate hepatocyte lipid accumulation; B) P40 diet: severe hepatocyte lipid accumulation; C) P40 diet: peripheral nuclei in hepatocytes with severe lipid accumulation (arrow); D) H10P30: hepatocyte hypertrophy (circle); E) P40 diet: vacuolar degeneration (asterisk) areas and cord loss; F) H10P30 diet: sinusoid congestion; G) P40 diet: foci of granulocytes; H) P40 diet: tissue necrosis areas. S=sinusoids; Gr=granulocytes; TN=tissue necrosis. Haematoxylin and eosin staining.
Figure 2.
Main histological alterations in liver of D. labrax fed test diets. A) CV diet: moderate hepatocyte lipid accumulation; B) P40 diet: severe hepatocyte lipid accumulation; C) P40 diet: peripheral nuclei in hepatocytes with severe lipid accumulation (arrow); D) H10P30: hepatocyte hypertrophy (circle); E) P40 diet: vacuolar degeneration (asterisk) areas and cord loss; F) H10P30 diet: sinusoid congestion; G) P40 diet: foci of granulocytes; H) P40 diet: tissue necrosis areas. S=sinusoids; Gr=granulocytes; TN=tissue necrosis. Haematoxylin and eosin staining.
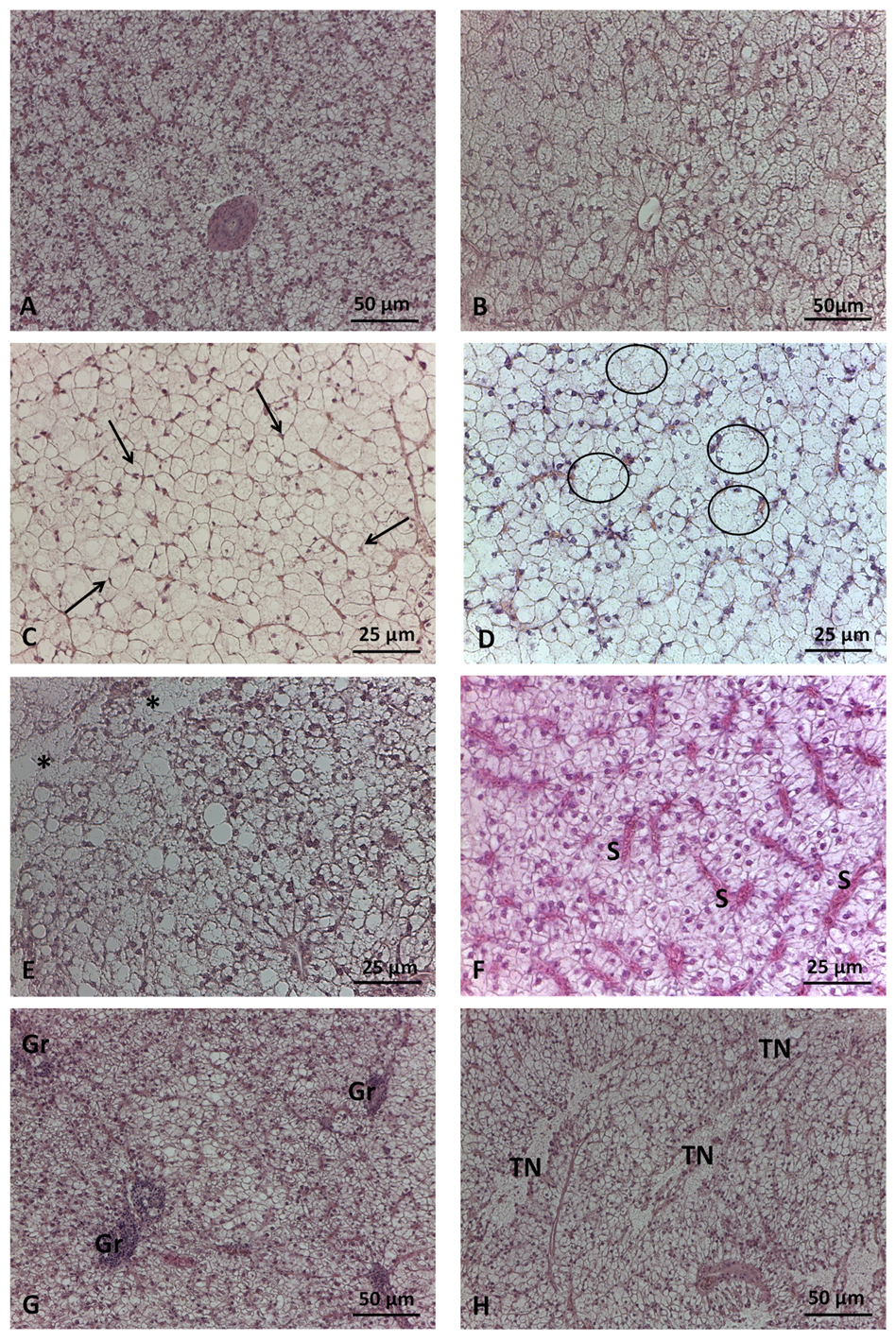
Figure 3.
Frequency percentage of significant histological alterations in liver of sea bream (A) and sea bass (B-C) fed test diets. Chi-square test p<0.05.
Figure 3.
Frequency percentage of significant histological alterations in liver of sea bream (A) and sea bass (B-C) fed test diets. Chi-square test p<0.05.
3.3. Liver index
3.3.1. Formula and scoring scheme development
The LI formula is based on the percentage ratio between the histological alterations observed and a theoretical reference condition. This reference condition is derived by assigning the maximum score of 6 to all alterations observed in the 9 fields, representing the most severe histopathological condition observable. For this reason, the formula of the liver index provides lower values compared to those calculated using the Bernet index. The two indices were compared through a regression analysis. The mathematical model that links the two indices has zero intercept, the same angular coefficient of 0.67 for both sea bream and sea bass, and the coefficient R2 ~ 1. Based on the slope factor, the LI values were recalibrated according to the scoring system suggested by Zimmerli et al. [56] to define corresponding classes, as shown in Table 6.
3.3.2. Liver health calculation
The LI values are shown in Figure 4. Dietary treatment significantly affected the LI in both sea bream (p< 0.001) and sea bass (p< 0.01). In both species the LI was significantly higher in fish fed H40, P40, and H10P30 diets compared to the CV diet (p<0.05). The LI values in sea bream varied from 5.8 in CV dietary group (Class 1) to 7.6, 7.9 and 9.2 respectively in H40, P40, and H10P30 groups (Class 2). The LI values were higher in sea bass varying from 8.4 in CV group (Class 2) to 15.5, 17.9 and 13.2 respectively in H40, P40, and H10P30 groups (Class 3), indicating a more pronounced dietary effect on liver health compared to sea bream. In particular, sea bass fed P40 diet showed the highest LI value, although it was not significantly different from H40 and H10P30 dietary groups.
3.4. Blood biochemistry
Dietary treatment significantly affected serum biochemical parameters, except AST enzyme activity, in both species (p<0.05) (Figure 5 and Figure 6). In sea bream, the serum GLU and TP levels in P40 diet were similar to those of the CV dietary group and significantly lower (p<0.05) compared to the levels measured in the H10P30 group. These parameters remained similar between sea bass fed P40 diet and the control group but were significantly higher compared to H10P30 dietary group. The serum levels of CHO and TAG showed a progressive increase in sea bream fed with H40, P40, and H10P30 diets compared to the CV diet, resulting only significantly higher in H10P30 compared to CV diet. In sea bass, these parameters were significantly higher only in fish fed the P40 diet compared to the CV (p<0.05). The lowest levels of TAG were measured in sea bass fed the H10P30 diet. Furthermore, sea bass exhibited notably higher TAG levels (700-800 mg/dl) compared to sea bream (300-400 mg/dl) irrespective of the dietary treatment. The ALT enzyme activity was significantly lower in sea bream and higher in sea bass fed H40 diet compared to CV.
An FPCA analysis was applied to examine the relationships between both the biochemical parameters and LI in sea bream and sea bass (Figure 7). Cholesterol was significantly correlated with the LI (visualized inside the red circle) in both species. Moreover, a significant inverse correlation was evident between the LI, BW and K in sea bass. Three clusters (CL) of variables that showed positive correlations were identified in both species. The first cluster comprised CHO, GLU and TAG in sea bream, along with HSI in sea bass. In sea bream, HSI showed a strong association with the BW and TL (second cluster). Notably, in sea bass, metabolic parameters grouped in cluster 1 displayed an inverse correlation with cluster 2 variables (BW, TL, K), resulting in an opposite positioning. The third cluster consisted of TP, ALB, AST, and ALT, which exhibited a closer relationship in sea bream than in sea bass.
4. Discussion
Ensuring fish health and welfare has become a priority in modern research on fish nutrition and feeding in aquaculture for improving the sustainability of feed industry, profitability, and ethics of aquaculture production [77]. Assessing fish health and welfare is particularly significant in feeding trials that aim to test alternative diets, as sub-optimal feed formulations can impact fish physiology and adversely affect growth performance, metabolism and immune system [7,78,79,80]. Blood biochemistry is an effective method for assessing the health and welfare of farmed fish. This method is valuable both for farming management and research applied to fish nutrition and feeding, as it enables the early identification of changes in physiological parameters related to alterations in organ functions before the occurrence of pathological conditions [37,81,82]. The liver is a key metabolic organ and an important indicator of nutritional pathology [43]. Histological investigations represent the initial step in detecting potential changes associated with inflammation, degeneration, and cell death. These investigations are crucial for assessing the effects of novel diet formulations in feeding trials on liver metabolism and health [44,45,75].
This study highlighted a different response to the test diets between sea bream and sea bass in terms of biometric and blood chemistry parameters as well as liver health status. In sea bream, the dietary inclusion of alternative ingredients resulted in a significant increase in final body weight and somatic indices, indicating beneficial effects on overall fish health. These findings are supported by FPCA analysis, which reveals a positive correlation between the HSI and both BW and TL. Conversely, these effects were not observed in sea bass, where final BW remained unchanged among dietary groups, and the K factor decreased, especially in the P40 dietary group. The values of the K and HSI indices observed in this study for sea bream and sea bass are consistent with those reported in the literature for healthy fish [30,32]. Serum biochemical parameters remained within the physiological range across all dietary groups suggesting a good nutritional status in both fish species [83,84]. Major changes were observed in serum CHO and TAG levels, that progressively increased in sea bream fed H10, P40 and H10P30 diets in comparison to fish fed the CV one. In sea bass these changes were only evident in fish fed P40 diet, despite very high levels of TAG found in all dietary groups. The observed increase in serum lipid levels may be in response to changes in the relative proportion of plant protein-rich feed mixture in the diets including the test ingredients. This could possibly reflect a reduced lipotropic effect due to the concurrent lower inclusion levels of soy products [82,85,86,87]. Differences in CHO and TAG levels observed among diets H40, P40, and H10P30 can be partially explained by a higher digestible carbohydrates intake due to the different starch content in the replaced diets compared to the control one. Besides, the effect of chitin and taurine present in insect meal, may have contributed to minor differences among fish fed the replaced diets [27,31,37,38,88] as well as differences in the amino acid composition of proteins from insect and poultry by-product meals could also have affected the de novo lipid synthesis contributing to increased serum lipid levels [87,88,89]. Indeed, dietary carbohydrate levels and lipid sources as well as the proportion of saturated and unsaturated fatty acids, regulate glucose and lipid metabolism in sea bream and sea bass [90,91,92]. The positive correlation obtained from FPCA analysis among GLU, CHO, and TAG levels in both fish species indicates a relationship between carbohydrate and lipid metabolism, supporting this hypothesis. These parameters are further correlated with the K index in sea bream and with HSI in sea bass, suggesting a different metabolic status in the two fish species, favoring growth in the former and liver lipid accumulation in the latter. The histological results, which are detailed below, also provide support to this hypothesis.
According to the FPCA analysis, serum TP were only correlated with serum transaminases which showed very low values across diets. The AST enzymatic activity was unaffected by dietary treatments while minor changes were observed in ALT activity in sea bass fed H40 diet compared to the other groups. A high liver ALT activity was reported in sea bream fed the diet containing 30% H. illucens meal and interpreted as an increased amino acid catabolism for energy production [30].
Histological observations highlighted the presence of several alterations, primarily associated with lipid accumulation, with a lower number and frequency percentage in sea bream compared to sea bass, indicating an overall better liver condition in sea bream. These findings confirm the macroscopic observations, which indicated low to medium levels of hepatic lipid accumulation for sea bream and medium to high levels for sea bass. These data also appear consistent with the histomorphometric analysis. Indeed, the mean values of hepatocyte and nucleus areas as well as the N/H ratio correspond to mild and moderate hepatocyte lipid accumulation for sea bream and from mild to severe hepatocyte lipid accumulation for sea bass. Furthermore, the histomorphometric evaluation highlighted significant differences among the tested diets, consistent with the severity and frequency of the observed hepatic lipid accumulation in both fish species. In this regard, the mean hepatocyte area was significantly larger in sea bream fed H10P30 diet in agreement with the higher frequency of moderate lipid accumulation compared to fish fed H40 and P40 diets. In sea bass, the mean hepatocyte area was significantly larger in fish H40, P40, and H10P30 compared to those fed the CV diet. The largest mean area was measured in the P40 hepatocytes corresponding to the highest frequency percentage of severe lipid accumulation and vacuolar tissue degeneration. These results confirm the utility of the histomorphometric analysis in assessing and comparing the effects of different feeds on fish liver, as previously reported by Raskovic et al. [93]. However, our results are not directly comparable to those reported by other authors, as they were calculated based on different measurements of hepatocytes and nuclei, such as the diameter ratio in sea bream [68] and sea bass [66], or the maximum and minimum length of hepatocytes, considering the nucleus as arbitrary reference point in sea bream fed microalgae and poultry oils [67].
Among the 18 histopathological parameters selected for this study, the most frequently observed ones directly related to the diets were those associated with the degree of lipid accumulation, showing different frequencies among dietary groups and between the two fish species. Relative to controls, hepatocytes of sea bream fed the replaced diets, besides higher lipid accumulation, displayed also low to medium occurrence of sinusoidal congestion, hyperplasia and cord loss. Similar results were reported in the same fish species fed a diet where 50% fishmeal was replaced with poultry by-products meal [25]. Hepatic lipid accumulation is classified as a regressive histological alteration. Its reversibility was observed in sea bream fed vegetable oils, when fish were re-fed with a balanced diet, denoting the non pathological character of this liver alteration [50]. The hepatocyte hyperplasia observed in sea bream fed the diets H40 and H10P30 could in part be interpreted as an adaptive response to increased carbohydrate metabolism. The higher serum glucose levels, although not statistically significant, observed in these dietary groups compared to P40 group, provides support to this interpretation. Hepatocyte hyperplasia has also been reported in relation to a faster absorption rate of carbohydrates from pea seed meal in sea bass [47]. These histological findings, particularly in sea bream fed H10P30 diet showing the higher frequency of moderate lipid accumulation and hepatocyte hyperplasia compared to CV group, are consistent with the higher HSI values and serum lipid level. Moderate to severe peri-pancreatic fat infiltration were observed in sea bream with no significant difference among dietary groups. This seems consistent with the histological picture reported in this fish species after long-term feeding with vegetable oils [50]. On the contrary, Baeza-Ariño et al. [52] observed a significant reduction of fat infiltrations in sea bream fed a mixture of vegetable protein concentrates vs a commercial diet.
In sea bass, the hepatocyte lipid accumulation was found mostly moderate in fish fed the control diet and turned to severe in those fed the replaced diets. In the latter dietary groups, other significant associated alterations were observed, including displacement of peripheral nucleus, hepatocyte hypertrophy, cord loss, tissue vacuolar degeneration, pycnotic nuclei and sinusoid congestion, indicative of an incipient steatosis condition probably due to altered liver lipid metabolism. A worsening of the liver health condition was observed in sea bass fed the P40 diet, which exhibited the most pronounced histological alterations. These alterations appear to be slightly attenuated in fish fed the H10P30 diet consistently with lower serum TAG levels compared to P40 dietary group. Liver steatosis generally occurs when dietary lipid or energy exceeds the capacity of hepatic cells to oxidize fatty acids, resulting in increased TAG synthesis and deposition into hepatocytes [94,95]. The balance between lipogenesis and lipolysis is maintained through various transcription factors and genes, which are, in turn, subject to regulation by dietary fatty acids [67,79,96]. Steatosic liver is generally considered a para-physiological condition, commonly detectable in farmed sea bass and sea bream in relation to feeding regimes [61,97,98,99].
The results of LI analysis also highlight a major influence of the dietary treatment on liver health status in both fish species but with a different amplitude. In sea bream, slightly but significantly higher LI values were observed in fish fed H40, P40, and H10P30 diets, indicating slight modifications of normal liver architecture and morphology (class 2) compared to those fed the control diet (falling into class 1, normal liver histology), which is consistent with histomorphometric results. Higher LI values falling in class 3 and indicating the occurrence of moderate liver alterations, were found in sea bass fed replaced diets compared to fish fed CV diet which displayed an LI in class 2. In any case, these results indicate that the liver parenchyma architecture and functional condition were maintained. These findings are consistent with the biochemical and histological observations, particularly the elevated serum TAG levels and severe lipid accumulation observed in all dietary groups. The results of the FPCA highlighted a significant correlation between the LI and CHO levels (green dot inside the red circle) both in sea bream and sea bass. This suggest that changes in dietary CHO is the primary cause of the metabolic effect here observed when replacing plant proteins with insect and poultry by-product meals in the diet. However, the significant inverse correlation between LI and both final BW and K factor in sea bass (yellow dots inside the red circle) suggests a potential influence of liver condition on fish body weight and biometric parameters.
The developed histological protocol, new LI formula, and scoring scheme which enable transformation of qualitative observations into a semi-quantitative index, provided objective information on the liver health of sea bream and sea bass. Particular attention has been paid to evaluating the severity and frequency of hepatocyte lipid accumulation and the associated alterations in response to the dietary treatment. Differently from the Bernet index [54], the new LI formula and the developed scoring scheme express the observed liver health condition in relation to a theoretical reference condition, making the assessment more objective compared to the criterion used by Zimmerli et al. for defining the severity classes of the liver index [56]. Moreover, based on results of regression analysis, the new LI formula returns values equivalent to those of Bernet index and the recalibrated classes are equivalent to those developed by Zimmerli et al., [56], allowing comparison of our results with literature data.
The biochemical and histological results indicated a favorable physiological status and good liver conditions in sea bream, with a slight increase in serum CHO and TAG levels, along with moderate hepatocyte lipid accumulation especially in response to the H10P30 diet. In sea bass, elevated serum TAG levels and severe liver lipid accumulation suggest a high energy content in the replaced diets under the experimental conditions [78,100,101]. However, this condition did not influence body weight and nutritional status. High circulating lipid levels and fatty liver often occur when farmed fish are fed moderate to high-energy commercial feeds, similar to those formulated in the present study [98,102].
5. Conclusions
Overall, the results highlighted species-specific differences in the physiological response to alternative dietary protein-rich ingredients. Sea bream adapted to the replaced diets, displaying efficient utilization of dietary nutrients, which results in improved body weight gain and good liver health. Sea bass, despite the HSI values observed in this study are consistent with those reported in the literature for healthy fish, showed altered liver lipid metabolism and additional lipid accumulation when fed diets rich in H. illucens and poultry by product meals, leading to incipient hepatic steatosis. Therefore, potential risks to liver health should be not ruled out when fish are fed longer with substantial levels of processed animal proteins in the diet.
In this study, blood biochemistry and liver histomorphology has proven to be highly informative in assessing the physiological status of sea bream and sea bass in response to the tested diets. This stresses the importance of evaluating liver health for an overall assessment of fish welfare in relation to the feeding treatment. Moreover, consistently with biochemical and histological findings, the liver index here proposed has proven to be a reliable tool for assessing the liver health of sea bream and sea bass.
Author Contributions
Conceptualization, V.D., P.D.M., A.M., C.G and G.M..; methodology, M.G.F and A.M.; formal analysis, M.G.F., V.D. and P.D.M.; investigation, V.D., P.D.M., A.M., M.G., G.C., E.T., T.P. and A.L.; resources, P.D.M., V.D., A.M.., A.L., T.P. and G.C.; data curation, V.D., P.D.M., A.M., M.G.F., T.P., A.L. and G.C.; validation, V.D., P.D.M., M.G.F. and A.M.; visualization, M.G.F., A.M., V.D., P.D.M., T.P., A.L. and G.C.; writing-original draft preparation, V.D., P.D.M. A.M., M.F.G., G.C., T.P.; writing-review and editing, V.D., P.D.M., A.M.; supervision, G.M., and E.T.; project administration, G.M.; funding acquisition, E.T. All authors have read and agreed to the published version of the manuscript.
Funding
This research was supported by the SUSHIN (SUstainable fiSH feeds Innovative ingredients) project funded by Ager Agroalimentare e Ricerca Foundation. (AGER2- grant n. 2016-0112.
Institutional Review Board Statement
The present study was approved by the Ethics Committee of the University of Udine and authorized by the Italian Ministry of Health (n.290/2019-PR) in accordance with European legal frameworks on the protection of animals used for scientific purposes (Directive 2010/63/EU).
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
The authors would also like to express their thanks to the staff of the University of Udine for conducting the experiments and providing assistance for fish sampling.
Conflicts of Interest
The authors declare no conflict of interest.
References
- FAO. 2020. The State of World Fisheries and Aquaculture 2020. Sustainability in Action. FAO Rome; ISBN 978-92-5-132692-3.
- FAO. 2022. The State of World Fisheries and Aquaculture 2022. Towards Blue Transformation. FAO, Rome.; 2022; ISBN 978-92-5-136364-5.
- UN DESA. 2023. The Sustainable Development Goals Report 2023: Special Edition - July 2023.; New York, USA: UN DESA.; ISBN 978-92-1-101460-0.
- Tacon, A.G.J.; Metian, M.; McNevin, A.A. Future Feeds: Suggested Guidelines for Sustainable Development. Reviews in Fisheries Science & Aquaculture 2022, 30, 135–142. [Google Scholar] [CrossRef]
- Hua, K.; Cobcroft, J.M.; Cole, A.; Condon, K.; Jerry, D.R.; Mangott, A.; Praeger, C.; Vucko, M.J.; Zeng, C.; Zenger, K.; et al. The Future of Aquatic Protein: Implications for Protein Sources in Aquaculture Diets. One Earth 2019, 1, 316–329. [Google Scholar] [CrossRef]
- Gatlin, D.M.; Barrows, F.T.; Brown, P.; Dabrowski, K.; Gaylord, T.G.; Hardy, R.W.; Herman, E.; Hu, G.; Krogdahl, Å.; Nelson, R.; et al. Expanding the Utilization of Sustainable Plant Products in Aquafeeds: A Review. Aquaculture Res 2007, 38, 551–579. [Google Scholar] [CrossRef]
- Glencross, B.D.; Baily, J.; Berntssen, M.H.G.; Hardy, R.; MacKenzie, S.; Tocher, D.R. Risk Assessment of the Use of Alternative Animal and Plant Raw Material Resources in Aquaculture Feeds. Reviews in Aquaculture 2020, 12, 703–758. [Google Scholar] [CrossRef]
- Cottrell, R.S.; Blanchard, J.L.; Halpern, B.S.; Metian, M.; Froehlich, H.E. Global Adoption of Novel Aquaculture Feeds Could Substantially Reduce Forage Fish Demand by 2030. Nat Food 2020, 1, 301–308. [Google Scholar] [CrossRef]
- Colombo, S.M.; Roy, K.; Mraz, J.; Wan, A.H.L.; Davies, S.J.; Tibbetts, S.M.; Øverland, M.; Francis, D.S.; Rocker, M.M.; Gasco, L.; et al. Towards Achieving Circularity and Sustainability in Feeds for Farmed Blue Foods. Reviews in Aquaculture 2023, 15, 1115–1141. [Google Scholar] [CrossRef]
- Cao, L.; Halpern, B.S.; Troell, M.; Short, R.; Zeng, C.; Jiang, Z.; Liu, Y.; Zou, C.; Liu, C.; Liu, S.; et al. Vulnerability of Blue Foods to Human-Induced Environmental Change. Nat Sustain 2023. [Google Scholar] [CrossRef]
- Van Riel, A.; Nederlof, M.A.J.; Chary, K.; Wiegertjes, G.F.; De Boer, I.J.M. Feed-food Competition in Global Aquaculture: Current Trends and Prospects. Reviews in Aquaculture 2023, 15, 1142–1158. [Google Scholar] [CrossRef]
- Colombo, S.M. Physiological Considerations in Shifting Carnivorous Fishes to Plant-Based Diets. In Fish Physiology; Elsevier, 2020; Vol. 38, pp. 53–82 ISBN 978-0-12-820794-9.
- Aragão, C.; Gonçalves, A.T.; Costas, B.; Azeredo, R.; Xavier, M.J.; Engrola, S. Alternative Proteins for Fish Diets: Implications beyond Growth. Animals 2022, 12, 1211. [Google Scholar] [CrossRef]
- Meeker, D.L. Essential Rendering: All about the Animal by-Products Industry; National Renderers Association : Fats and Proteins Research Foundation : Animal Protein Producers Industry: Alexandria, Va., 2006; ISBN 978-0-9654660-3-5.
- Lim, C.; Webster, C.D.; Lee, C.-S. Alternative Protein Sources in Aquaculture Diets; Haworth press: New York, 2008; ISBN 978-1-56022-148-7.
- Nogales-Mérida, S.; Gobbi, P.; Józefiak, D.; Mazurkiewicz, J.; Dudek, K.; Rawski, M.; Kierończyk, B.; Józefiak, A. Insect Meals in Fish Nutrition. Reviews in Aquaculture 2019, 11, 1080–1103. [Google Scholar] [CrossRef]
- Hawkey, K.J.; Lopez-Viso, C.; Brameld, J.M.; Parr, T.; Salter, A.M. Insects: A Potential Source of Protein and Other Nutrients for Feed and Food. Annu. Rev. Anim. Biosci. 2021, 9, 333–354. [Google Scholar] [CrossRef] [PubMed]
- Alfiko, Y.; Xie, D.; Astuti, R.T.; Wong, J.; Wang, L. Insects as a Feed Ingredient for Fish Culture: Status and Trends. Aquaculture and Fisheries 2022, 7, 166–178. [Google Scholar] [CrossRef]
- Gougbedji, A.; Detilleux, J.; Lalèyè, P.; Francis, F.; Caparros Megido, R. Can Insect Meal Replace Fishmeal? A Meta-Analysis of the Effects of Black Soldier Fly on Fish Growth Performances and Nutritional Values. Animals 2022, 12, 1700. [Google Scholar] [CrossRef] [PubMed]
- Hossain, Md.S.; Small, B.C.; Hardy, R. Insect Lipid in Fish Nutrition: Recent Knowledge and Future Application in Aquaculture. Reviews in Aquaculture 2023, 15, 1664–1685. [Google Scholar] [CrossRef]
- Arena, R.; Manuguerra, S.; Curcuraci, E.; Cusimano, M.; Lo Monaco, D.; Di Bella, C.; Santulli, A.; Messina, C.M. Fisheries and Aquaculture By-Products Modulate Growth, Body Composition, and Omega-3 Polyunsaturated Fatty Acid Content in Black Soldier Fly (Hermetia illucens) Larvae. Front. Anim. Sci. 2023, 4, 1204767. [Google Scholar] [CrossRef]
- Maroušek, J.; Strunecký, O.; Maroušková, A. Insect Rearing on Biowaste Represents a Competitive Advantage for Fish Farming. Reviews in Aquaculture 2023, 15, 965–975. [Google Scholar] [CrossRef]
- Karapanagiotidis, I.T.; Psofakis, P.; Mente, E.; Malandrakis, E.; Golomazou, E. Effect of Fishmeal Replacement by Poultry By-Product Meal on Growth Performance, Proximate Composition, Digestive Enzyme Activity, Haematological Parameters and Gene Expression of Gilthead Seabream (Sparus aurata). Aquacult Nutr 2019, 25, 3–14. [Google Scholar] [CrossRef]
- Sabbagh, M.; Schiavone, R.; Brizzi, G.; Sicuro, B.; Zilli, L.; Vilella, S. Poultry By-Product Meal as an Alternative to Fish Meal in the Juvenile Gilthead Seabream (Sparus aurata) Diet. Aquaculture 2019, 511, 734220. [Google Scholar] [CrossRef]
- Psofakis, P.; Meziti, A.; Berillis, P.; Mente, E.; Kormas, K.A.; Karapanagiotidis, I.T. Effects of Dietary Fishmeal Replacement by Poultry By-Product Meal and Hydrolyzed Feather Meal on Liver and Intestinal Histomorphology and on Intestinal Microbiota of Gilthead Seabream (Sparus aurata). Applied Sciences 2021, 11, 8806. [Google Scholar] [CrossRef]
- Magalhães, R.; Sánchez-López, A.; Leal, R.S.; Martínez-Llorens, S.; Oliva-Teles, A.; Peres, H. Black Soldier Fly (Hermetia illucens) Pre-Pupae Meal as a Fish Meal Replacement in Diets for European Seabass (Dicentrarchus labrax). Aquaculture 2017, 476, 79–85. [Google Scholar] [CrossRef]
- Abdel-Tawwab, M.; Khalil, R.H.; Metwally, A.A.; Shakweer, M.S.; Khallaf, M.A.; Abdel-Latif, H.M.R. Effects of Black Soldier Fly (Hermetia illucens L.) Larvae Meal on Growth Performance, Organs-Somatic Indices, Body Composition, and Hemato-Biochemical Variables of European Sea Bass, Dicentrarchus labrax. Aquaculture 2020, 522, 735136. [Google Scholar] [CrossRef]
- Karapanagiotidis, I.T.; Neofytou, M.C.; Asimaki, A.; Daskalopoulou, E.; Psofakis, P.; Mente, E.; Rumbos, C.I.; Athanassiou, C.G. Fishmeal Replacement by Full-Fat and Defatted Hermetia Illucens Prepupae Meal in the Diet of Gilthead Seabream (Sparus aurata). Sustainability 2023, 15, 786. [Google Scholar] [CrossRef]
- Fabrikov, D.; Vargas-García, M.D.C.; Barroso, F.G.; Sánchez-Muros, M.J.; Cacua Ortíz, S.M.; Morales, A.E.; Cardenete, G.; Tomás-Almenar, C.; Melenchón, F. Effect on Intermediary Metabolism and Digestive Parameters of the High Substitution of Fishmeal with Insect Meal in Sparus Aurata Feed. Insects 2021, 12, 965. [Google Scholar] [CrossRef] [PubMed]
- Mastoraki, M.; Katsika, L.; Enes, P.; Guerreiro, I.; Kotzamanis, Y.P.; Gasco, L.; Chatzifotis, S.; Antonopoulou, E. Insect Meals in Feeds for Juvenile Gilthead Seabream (Sparus aurata): Effects on Growth, Blood Chemistry, Hepatic Metabolic Enzymes, Body Composition and Nutrient Utilization. Aquaculture 2022, 561, 738674. [Google Scholar] [CrossRef]
- Mastoraki, M.; Panteli, N.; Kotzamanis, Y.P.; Gasco, L.; Antonopoulou, E.; Chatzifotis, S. Nutrient Digestibility of Diets Containing Five Different Insect Meals in Gilthead Sea Bream (Sparus aurata) and European Sea Bass (Dicentrarchus labrax). Animal Feed Science and Technology 2022, 292, 115425. [Google Scholar] [CrossRef]
- Mastoraki, M.; Mollá Ferrándiz, P.; Vardali, S.C.; Kontodimas, D.C.; Kotzamanis, Y.P.; Gasco, L.; Chatzifotis, S.; Antonopoulou, E. A Comparative Study on the Effect of Fish Meal Substitution with Three Different Insect Meals on Growth, Body Composition and Metabolism of European Sea Bass (Dicentrarchus labrax L.). Aquaculture 2020, 528, 735511. [Google Scholar] [CrossRef]
- Bousdras, T.; Feidantsis, K.; Panteli, N.; Chatzifotis, S.; Piccolo, G.; Gasco, L.; Gai, F.; Antonopoulou, E. Dietary Tenebrio Molitor Larvae Meal Inclusion Exerts Tissue-Specific Effects on Cellular, Metabolic, and Antioxidant Status in European Sea Bass (Dicentrarchus labrax) and Gilthead Seabream (Sparus aurata). Aquaculture Nutrition 2022, 2022, 1–20. [Google Scholar] [CrossRef]
- Gai, F.; Cusimano, G.M.; Maricchiolo, G.; Caccamo, L.; Caimi, C.; Macchi, E.; Meola, M.; Perdichizzi, A.; Tartarisco, G.; Gasco, L. Defatted Black Soldier Fly Meal in Diet for Grow-Out Gilthead Seabream (Sparus aurata L. 1758): Effects on Growth Performance, Gill Cortisol Level, Digestive Enzyme Activities, and Intestinal Histological Structure. Aquaculture Research 2023, 2023, 1–18. [Google Scholar] [CrossRef]
- Di Rosa, A.R.; Caccamo, L.; Pansera, L.; Oteri, M.; Chiofalo, B.; Maricchiolo, G. Influence of Hermetia illucens Larvae Meal Dietary Inclusion on Growth Performance, Gut Histological Traits and Stress Parameters in Sparus Aurata. Animals 2023, 13, 339. [Google Scholar] [CrossRef]
- Anedda, R.; Melis, R.; Palomba, A.; Vitangeli, I.; Biosa, G.; Braca, A.; Antonini, M.; Moroni, F.; Rimoldi, S.; Terova, G.; et al. Balanced Replacement of Fish Meal with Hermetia illucens Meal Allows Efficient Hepatic Nutrient Metabolism and Increases Fillet Lipid Quality in Gilthead Sea Bream (Sparus Aurata). Aquaculture 2023, 576, 739862. [Google Scholar] [CrossRef]
- Basto, A.; Valente, L.M.P.; Sousa, V.; Conde-Sieira, M.; Soengas, J.L. Total Fishmeal Replacement by Defatted Tenebrio Molitor Larvae Meal Induces Alterations in Intermediary Metabolism of European Sea Bass (Dicentrarchus labrax). Journal of Animal Science 2023, 101, skad040. [Google Scholar] [CrossRef]
- Randazzo, B.; Zarantoniello, M.; Cardinaletti, G.; Cerri, R.; Giorgini, E.; Belloni, A.; Contò, M.; Tibaldi, E.; Olivotto, I. Hermetia illucens and Poultry By-Product Meals as Alternatives to Plant Protein Sources in Gilthead Seabream (Sparus aurata) Diet: A Multidisciplinary Study on Fish Gut Status. Animals 2021, 11, 677. [Google Scholar] [CrossRef]
- Pulido-Rodriguez, L.F.; Cardinaletti, G.; Secci, G.; Randazzo, B.; Bruni, L.; Cerri, R.; Olivotto, I.; Tibaldi, E.; Parisi, G. Appetite Regulation, Growth Performances and Fish Quality Are Modulated by Alternative Dietary Protein Ingredients in Gilthead Sea Bream (Sparus aurata) Culture. Animals 2021, 11, 1919. [Google Scholar] [CrossRef]
- Pleić, I.L.; Bušelić, I.; Messina, M.; Hrabar, J.; Žuvić, L.; Talijančić, I.; Žužul, I.; Pavelin, T.; Anđelić, I.; Pleadin, J.; et al. A Plant-Based Diet Supplemented with Hermetia illucens Alone or in Combination with Poultry by-Product Meal: One Step Closer to Sustainable Aquafeeds for European Seabass. J Animal Sci Biotechnol 2022, 13, 77. [Google Scholar] [CrossRef] [PubMed]
- Bakke, A.M.; Glover, C.; Krogdahl, Å. Feeding, Digestion and Absorption of Nutrients. In Fish Physiology; Elsevier, 2010; Vol. 30, pp. 57–110 ISBN 978-0-12-374982-6.
- Bruslé, J.; Gonzàlez I Anadon, G. The Structure and Function of Fish Liver. In Fish Morphology; Datta Munshi, J.S., Dutta, H.M., Eds.; Routledge, 2017; pp. 77–93 ISBN 978-0-203-75599-0.
- Roberts, R.J. Nutritional Pathology. In Fish Nutrition; Elsevier, 2022; pp. 823–855 ISBN 978-0-12-819587-1.
- Sirri, R.; Sarli, G.; Bianco, C.; Bonaldo, A.; Gatta, P.P.; Fontanillas, R.; De Vico, G.; Carella, F.; Brachelente, C.; Parma, L.; et al. Retrospective Study of Pathology-Based Investigative Techniques for the Assessment of Diet-Induced Changes in Liver and Intestine of Flatfish. Italian Journal of Animal Science 2018, 17, 518–529. [Google Scholar] [CrossRef]
- Rašković, B.; Berillis, P. Special Issue on the Histopathology of Aquatic Animals. Applied Sciences 2022, 12, 971. [Google Scholar] [CrossRef]
- Caballero, M.J.; López-Calero, G.; Socorro, J.; Roo, F.J.; Izquierdo, M.S.; Férnandez, A.J. Combined Effect of Lipid Level and Fish Meal Quality on Liver Histology of Gilthead Seabream (Sparus aurata). Aquaculture 1999, 179, 277–290. [Google Scholar] [CrossRef]
- Russell, P.M.; Davies, S.J.; Gouveia, A.; Tekinay, A.A. Influence of Dietary Starch Source on Liver Morphology in Juvenile Cultured European Sea Bass (Dicentrarchus labrax L.): Dietary Starch in Sea Bass Diets. Aquaculture Research 2001, 32, 306–314. [Google Scholar] [CrossRef]
- Benedito-Palos, L.; Navarro, J.C.; Sitjà-Bobadilla, A.; Gordon Bell, J.; Kaushik, S.; Pérez-Sánchez, J. High Levels of Vegetable Oils in Plant Protein-Rich Diets Fed to Gilthead Sea Bream (Sparus aurata L.): Growth Performance, Muscle Fatty Acid Profiles and Histological Alterations of Target Tissues. Br J Nutr 2008, 100, 992–1003. [Google Scholar] [CrossRef]
- Figueiredo-Silva, A.; Rocha, E.; Dias, J.; Silva, P.; Rema, P.; Gomes, E.; Valente, L.M.P. Partial Replacement of Fish Oil by Soybean Oil on Lipid Distribution and Liver Histology in European Sea Bass (Dicentrarchus labrax) and Rainbow Trout (Oncorhynchus Mykiss) Juveniles. Aquac Nutrition 2005, 11, 147–155. [Google Scholar] [CrossRef]
- Caballero, M.J.; Izquierdo, M.S.; Kjørsvik, E.; Fernández, A.J.; Rosenlund, G. Histological Alterations in the Liver of Sea Bream, Sparus aurata L., Caused by Short- or Long-term Feeding with Vegetable Oils. Recovery of Normal Morphology after Feeding Fish Oil as the Sole Lipid Source. Journal of Fish Diseases 2004, 27, 531–541. [Google Scholar] [CrossRef]
- Kokou, F.; Sarropoulou, E.; Cotou, E.; Rigos, G.; Henry, M.; Alexis, M.; Kentouri, M. Effects of Fish Meal Replacement by a Soybean Protein on Growth, Histology, Selected Immune and Oxidative Status Markers of Gilthead Sea Bream, Sparus aurata. J. World Aquaculture Soc 2015, 46, 115–128. [Google Scholar] [CrossRef]
- Baeza-Ariño, R.; Martínez-Llorens, S.; Nogales-Mérida, S.; Jover-Cerda, M.; Tomás-Vidal, A. Study of Liver and Gut Alterations in Sea Bream, Sparus aurata L., Fed a Mixture of Vegetable Protein Concentrates. Aquac Res 2016, 47, 460–471. [Google Scholar] [CrossRef]
- Saleh, N.E. Assessment of Sesame Meal as a Soybean Meal Replacement in European Sea Bass (Dicentrarchus labrax) Diets Based on Aspects of Growth, Amino Acid Profiles, Haematology, Intestinal and Hepatic Integrity and Macroelement Contents. Fish Physiol Biochem 2020, 46, 861–879. [Google Scholar] [CrossRef] [PubMed]
- Bernet, D.; Schmidt, H.; Meier, W.; Burkhardt-Holm, P.; Wahli, T. Histopathology in Fish: Proposal for a Protocol to Assess Aquatic Pollution. J Fish Diseases 1999, 22, 25–34. [Google Scholar] [CrossRef]
- Bernet, D.; Schmidt-Posthaus, H.; Wahli, T.; Burkhardt-Holm, P. Evaluation of Two Monitoring Approaches to Assess Effects of Waste Water Disposal on Histological Alterations in Fish. Hydrobiologia 2004, 524, 53–66. [Google Scholar] [CrossRef]
- Zimmerli, S.; Bernet, D.; Burkhardt-Holm, P.; Schmidt-Posthaus, H.; Vonlanthen, P.; Wahli, T.; Segner, H. Assessment of Fish Health Status in Four Swiss Rivers Showing a Decline of Brown Trout Catches. Aquat. Sci. 2007, 69, 11–25. [Google Scholar] [CrossRef]
- Van Dyk, J.C.; Cochrane, M.J.; Wagenaar, G.M. Liver Histopathology of the Sharptooth Catfish Clarias Gariepinus as a Biomarker of Aquatic Pollution. Chemosphere 2012, 87, 301–311. [Google Scholar] [CrossRef]
- Beegam, A.; Lopes, M.; Fernandes, T.; Jose, J.; Barreto, A.; Oliveira, M.; Soares, A.M.V.M.; Trindade, T.; Thomas, S.; Pereira, M.L. Multiorgan Histopathological Changes in the Juvenile Seabream Sparus aurata as a Biomarker for Zinc Oxide Particles Toxicity. Environ Sci Pollut Res 2020, 27, 30907–30917. [Google Scholar] [CrossRef]
- Santos, R.M.B.; Monteiro, S.M.V.; Cortes, R.M.V.; Pacheco, F.A.L.; Fernandes, L.F.S. Seasonal Differences in Water Pollution and Liver Histopathology of Iberian Barbel (Luciobarbus Bocagei) and Douro Nase (Pseudochondrostoma Duriense) in an Agricultural Watershed. Water 2022, 14, 444. [Google Scholar] [CrossRef]
- Rašković, B.; Jarić, I.; Koko, V.; Spasić, M.; Dulić, Z.; Marković, Z.; Poleksić, V. Histopathological Indicators: A Useful Fish Health Monitoring Tool in Common Carp (Cyprinus Carpio Linnaeus, 1758) Culture. Open Life Sciences 2013, 8, 975–985. [Google Scholar] [CrossRef]
- Saraiva, A.; Costa, J.; Serrão, J.; Cruz, C.; Eiras, J.C. A Histology-Based Fish Health Assessment of Farmed Seabass (Dicentrarchus labrax L.). Aquaculture 2015, 448, 375–381. [Google Scholar] [CrossRef]
- Pacorig, V.; Galeotti, M.; Beraldo, P. Multiparametric Semi-Quantitative Scoring System for the Histological Evaluation of Marine Fish Larval and Juvenile Quality. Aquaculture Reports 2022, 26, 101285. [Google Scholar] [CrossRef]
- Rey, A.L.; Asín, J.; Ruiz Zarzuela, I.; Luján, L.; Iregui, C.A.; De Blas, I. A Proposal of Standardization for Histopathological Lesions to Characterize Fish Diseases. Reviews in Aquaculture 2020, 12, 2304–2315. [Google Scholar] [CrossRef]
- Raskovic, B.; Stankovic, M.; Markovic, Z.; Poleksic, V. Histological Methods in the Assessment of Different Feed Effects on Liver and Intestine of Fish. J Agric Sci BGD 2011, 56, 87–100. [Google Scholar] [CrossRef]
- Mandrioli, L.; Sirri, R.; Gatta, P.P.; Morandi, F.; Sarli, G.; Parma, L.; Fontanillas, R.; Bonaldo, A. Histomorphologic Hepatic Features and Growth Performances of Juvenile Senegalese Sole (Solea Senegalensis) Fed Isogenertic Practical Diets with Variable Protein/Lipid Levels: Histomorphologic Hepatic Features and Growth Performances in Senegalese Sole. Journal of Applied Ichthyology 2012, 28, 628–632. [Google Scholar] [CrossRef]
- Wassef, E.A.; Wahbi, O.M.; Saqr, E.M.; Saleh, N.E. Response of European Seabass (Dicentrarchus labrax) to Canola Oil Diets: Effect on Growth Performance, Fish Health and Liver and Intestine Histomorphology. Aquacult Int 2016, 24, 1073–1088. [Google Scholar] [CrossRef]
- Carvalho, M.; Montero, D.; Torrecillas, S.; Castro, P.; Zamorano, M.J.; Izquierdo, M. Hepatic Biochemical, Morphological and Molecular Effects of Feeding Microalgae and Poultry Oils to Gilthead Sea Bream (Sparus Aurata). Aquaculture 2021, 532, 736073. [Google Scholar] [CrossRef]
- Tefal, E.; Jauralde, I.; Tomás-Vidal, A.; Martínez-Llorens, S.; Peñaranda, D.S.; Jover-Cerdá, M. New Organic Raw Materials for Gilthead Seabream (Sparus aurata) Feeding and the Effects on Growth, Nutritive Parameters, Digestibility, and Histology. Fishes 2023, 8, 330. [Google Scholar] [CrossRef]
- Valaroutsou, E.; Voudanta, E.; Mente, E.; Berillis, P. A Microscope and Image Analysis Study of the Liver and Exocrine Pancreas of Sea Bream Sparus aurata Fed Different Diets. Int. J. Zool. Res. 2013, 3, 55–58. [Google Scholar] [CrossRef]
- Bilen, A.M.; Bilen, S. Effect of Diet on the Fatty Acids Composition of Cultured Sea Bass (Dicentrarchus labrax) Liver Tissues and Histology Compared with Wild Sea Bass Caught in Eagean Sea. Mar. Sci. Tech. Bull. 2013, 2, 15–21. [Google Scholar] [CrossRef]
- FAO FishStatJ. 2023. FishStatJ - Software for Fishery and Aquaculture Statistical Time Series. FAO, Statistics and Information Service, FAO Fisheries Department, Fishery Information, Data and Statistics Unit, Rome, Italy. Https://Www.Fao.Org/Fishery/Statistics/Software/Fishstatj/En.
- Official Methods of Analysis of AOAC International; Latimer, G.W., AOAC International, Eds.; 20th Edition.; AOAC International: Gaithersburg, Md, 2016; ISBN 978-0-935584-87-5.
- Burja, A.M.; Armenta, R.E.; Radianingtyas, H.; Barrow, C.J. Evaluation of Fatty Acid Extraction Methods for Thraustochytrium Sp. ONC-T18. J. Agric. Food Chem. 2007, 55, 4795–4801. [Google Scholar] [CrossRef]
- Di Marco, P.; Priori, A.; Finoia, M.G.; Massari, A.; Mandich, A.; Marino, G. Physiological Responses of European Sea Bass Dicentrarchus labrax to Different Stocking Densities and Acute Stress Challenge. Aquaculture 2008, 275, 319–328. [Google Scholar] [CrossRef]
- Cardinaletti, G.; Di Marco, P.; Daniso, E.; Messina, M.; Donadelli, V.; Finoia, M.G.; Petochi, T.; Fava, F.; Faccenda, F.; Contò, M.; et al. Growth and Welfare of Rainbow Trout (Oncorhynchus Mykiss) in Response to Graded Levels of Insect and Poultry By-Product Meals in Fishmeal-Free Diets. Animals 2022, 12, 1698. [Google Scholar] [CrossRef] [PubMed]
- R Core Team (2017) R: A Language and Environment for Statistical Computing. https://www.R-project.org/.
- Franks, B.; Ewell, C.; Jacquet, J. Animal Welfare Risks of Global Aquaculture. Sci. Adv. 2021, 7, eabg0677. [Google Scholar] [CrossRef] [PubMed]
- Oliva-Teles, A.; Enes, P.; Peres, H. Replacing Fishmeal and Fish Oil in Industrial Aquafeeds for Carnivorous Fish. In Feed and Feeding Practices in Aquaculture; Elsevier, 2015; pp. 203–233 ISBN 978-0-08-100506-4.
- Panserat, S.; Marandel, L.; Seiliez, I.; Skiba-Cassy, S. New Insights on Intermediary Metabolism for a Better Understanding of Nutrition in Teleosts. Annu. Rev. Anim. Biosci. 2019, 7, 195–220. [Google Scholar] [CrossRef] [PubMed]
- Ciji, A.; Akhtar, M.S. Stress Management in Aquaculture: A Review of Dietary Interventions. Reviews in Aquaculture 2021, 13, 2190–2247. [Google Scholar] [CrossRef]
- Ballester-Lozano, G.F.; Benedito-Palos, L.; Estensoro, I.; Sitjà-Bobadilla, A.; Kaushik, S.; Pérez-Sánchez, J. Comprehensive Biometric, Biochemical and Histopathological Assessment of Nutrient Deficiencies in Gilthead Sea Bream Fed Semi-Purified Diets. Br J Nutr 2015, 114, 713–726. [Google Scholar] [CrossRef]
- Bonvini, E.; Bonaldo, A.; Mandrioli, L.; Sirri, R.; Dondi, F.; Bianco, C.; Fontanillas, R.; Mongile, F.; Gatta, P.P.; Parma, L. Effects of Feeding Low Fishmeal Diets with Increasing Soybean Meal Levels on Growth, Gut Histology and Plasma Biochemistry of Sea Bass. Animal 2018, 12, 923–930. [Google Scholar] [CrossRef]
- Peres, H.; Santos, S.; Oliva-Teles, A. Selected Plasma Biochemistry Parameters in Gilthead Seabream ( Sparus aurata ) Juveniles. J. Appl. Ichthyol. 2013, 29, 630–636. [Google Scholar] [CrossRef]
- Peres, H.; Santos, S.; Oliva-Teles, A. Blood Chemistry Profile as Indicator of Nutritional Status in European Seabass (Dicentrarchus labrax). Fish Physiol Biochem 2014, 40, 1339–1347. [Google Scholar] [CrossRef]
- Couto, A.; Kortner, T.M.; Penn, M.; Bakke, A.M.; Krogdahl, Å.; Oliva-Teles, A. Effects of Dietary Soy Saponins and Phytosterols on Gilthead Sea Bream (Sparus aurata) during the on-Growing Period. Animal Feed Science and Technology 2014, 198, 203–214. [Google Scholar] [CrossRef]
- Kaushik, S.J.; Covès, D.; Dutto, G.; Blanc, D. Almost Total Replacement of Fish Meal by Plant Protein Sources in the Diet of a Marine Teleost, the European Seabass, Dicentrarchus labrax. Aquaculture 2004, 230, 391–404. [Google Scholar] [CrossRef]
- Dias, J.; Alvarez, M.J.; Arzel, J.; Corraze, G.; Diez, A.; Bautista, J.M.; Kaushik, S.J. Dietary Protein Source Affects Lipid Metabolism in the European Seabass (Dicentrarchus labrax). Comparative Biochemistry and Physiology Part A: Molecular & Integrative Physiology 2005, 142, 19–31. [Google Scholar] [CrossRef] [PubMed]
- Ekmann, K.S.; Dalsgaard, J.; Holm, J.; Campbell, P.J.; Skov, P.V. Effects of Dietary Energy Density and Digestible Protein:Energy Ratio on de Novo Lipid Synthesis from Dietary Protein in Gilthead Sea Bream (Sparus aurata) Quantified with Stable Isotopes. Br J Nutr 2013, 110, 1771–1781. [Google Scholar] [CrossRef]
- Messina, M.; Piccolo, G.; Tulli, F.; Messina, C.M.; Cardinaletti, G.; Tibaldi, E. Lipid Composition and Metabolism of European Sea Bass (Dicentrarchus labrax L.) Fed Diets Containing Wheat Gluten and Legume Meals as Substitutes for Fish Meal. Aquaculture 2013, 376–379, 6–14. [Google Scholar] [CrossRef]
- Castro, C.; Corraze, G.; Pérez-Jiménez, A.; Larroquet, L.; Cluzeaud, M.; Panserat, S.; Oliva-Teles, A. Dietary Carbohydrate and Lipid Source Affect Cholesterol Metabolism of European Sea Bass (Dicentrarchus labrax) Juveniles. Br J Nutr 2015, 114, 1143–1156. [Google Scholar] [CrossRef]
- Castro, C.; Corraze, G.; Firmino-Diógenes, A.; Larroquet, L.; Panserat, S.; Oliva-Teles, A. Regulation of Glucose and Lipid Metabolism by Dietary Carbohydrate Levels and Lipid Sources in Gilthead Sea Bream Juveniles. Br J Nutr 2016, 116, 19–34. [Google Scholar] [CrossRef]
- Torrecillas, S.; Robaina, L.; Caballero, M.J.; Montero, D.; Calandra, G.; Mompel, D.; Karalazos, V.; Kaushik, S.; Izquierdo, M.S. Combined Replacement of Fishmeal and Fish Oil in European Sea Bass (Dicentrarchus labrax): Production Performance, Tissue Composition and Liver Morphology. Aquaculture 2017, 474, 101–112. [Google Scholar] [CrossRef]
- Raskovic, B.; Stankovic, M.; Markovic, Z.; Poleksic, V. Histological Methods in the Assessment of Different Feed Effects on Liver and Intestine of Fish. J Agric Sci BGD 2011, 56, 87–100. [Google Scholar] [CrossRef]
- Spisni, E.; Tugnoli, M.; Ponticelli, A.; Mordenti, T.; Tomasi, V. Hepatic Steatosis in Artificially Fed Marine Teleosts. Journal of Fish Diseases 1998, 21, 177–184. [Google Scholar] [CrossRef] [PubMed]
- Montero, D.; Izquierdo, M. Welfare and Health of Fish Fed Vegetable Oils as Alternative Lipid Sources to Fish Oil. In Fish Oil Replacement and Alternative Lipid Sources in Aquaculture Feeds; Turchini, G., Ng, W.-K., Tocher, D., Eds.; CRC Press, 2010; pp. 439–485 ISBN 978-1-4398-0862-7.
- Geay, F.; Ferraresso, S.; Zambonino-Infante, J.L.; Bargelloni, L.; Quentel, C.; Vandeputte, M.; Kaushik, S.; Cahu, C.L.; Mazurais, D. Effects of the Total Replacement of Fish-Based Diet with Plant-Based Diet on the Hepatic Transcriptome of Two European Sea Bass (Dicentrarchus labrax) Half-Sibfamilies Showing Different Growth Rates with the Plant-Based Diet. BMC Genomics 2011, 12, 522. [Google Scholar] [CrossRef] [PubMed]
- Coz-Rakovac, R.; Strunjak-Perovic1, I.; Hacmanjek, M.; Popovic, N.T.; Lipej, Z.; Sostaric, B. Blood Chemistry and Histological Properties of Wild and Cultured Sea Bass (Dicentrarchus labrax) in the North Adriatic Sea. Vet Res Commun 2005, 29, 677–687. [Google Scholar] [CrossRef] [PubMed]
- Di Marco, P.; Petochi, T.; Marino, G.; Priori, A.; Finoia, M.G.; Tomassetti, P.; Porrello, S.; Giorgi, G.; Lupi, P.; Bonelli, A.; et al. Insights into Organic Farming of European Sea Bass Dicentrarchus labrax and Gilthead Sea Bream Sparus aurata through the Assessment of Environmental Impact, Growth Performance, Fish Welfare and Product Quality. Aquaculture 2017, 471, 92–105. [Google Scholar] [CrossRef]
- Tina, P.; Kević, N.; Restović, I.; Bočina, I. Histological and Biochemical Features of the Digestive System in the Cage-Reared Gilthead Sea Bream (Sparus aurata). Int J Biotechnol Recent Adv 2018, 2, 51–56. [Google Scholar] [CrossRef]
- Peres, H.; Oliva-Teles, A. Effect of Dietary Lipid Level on Growth Performance and Feed Utilization by European Sea Bass Juveniles (Dicentrarchus labrax). Aquaculture 1999, 179, 325–334. [Google Scholar] [CrossRef]
- Dias, J.; Alvarez, M.J.; Diez, A.; Arzel, J.; Corraze, G.; Bautista, J.M.; Kaushik, S.J. Regulation of Hepatic Lipogenesis by Dietary Protein/Energy in Juvenile European Seabass (Dicentrarchus labrax). Aquaculture 1998, 161, 169–186. [Google Scholar] [CrossRef]
- Marchi, A.; Bonaldo, A.; Di Biase, A.; Cerri, R.; Scicchitano, D.; Nanetti, E.; Candela, M.; Picone, G.; Capozzi, F.; Dondi, F.; et al. Towards a Free Wild-Caught Fishmeal, Fish Oil and Soy Protein in European Sea Bass Diet Using by-Products from Fishery and Aquaculture. Aquaculture 2023, 573, 739571. [Google Scholar] [CrossRef]
Figure 4.
Liver index of sea bream and sea bass fed test diets. Data are expressed as mean±sd. Different superscripts indicate significant differences among groups within the same species (p< 0.05).
Figure 4.
Liver index of sea bream and sea bass fed test diets. Data are expressed as mean±sd. Different superscripts indicate significant differences among groups within the same species (p< 0.05).
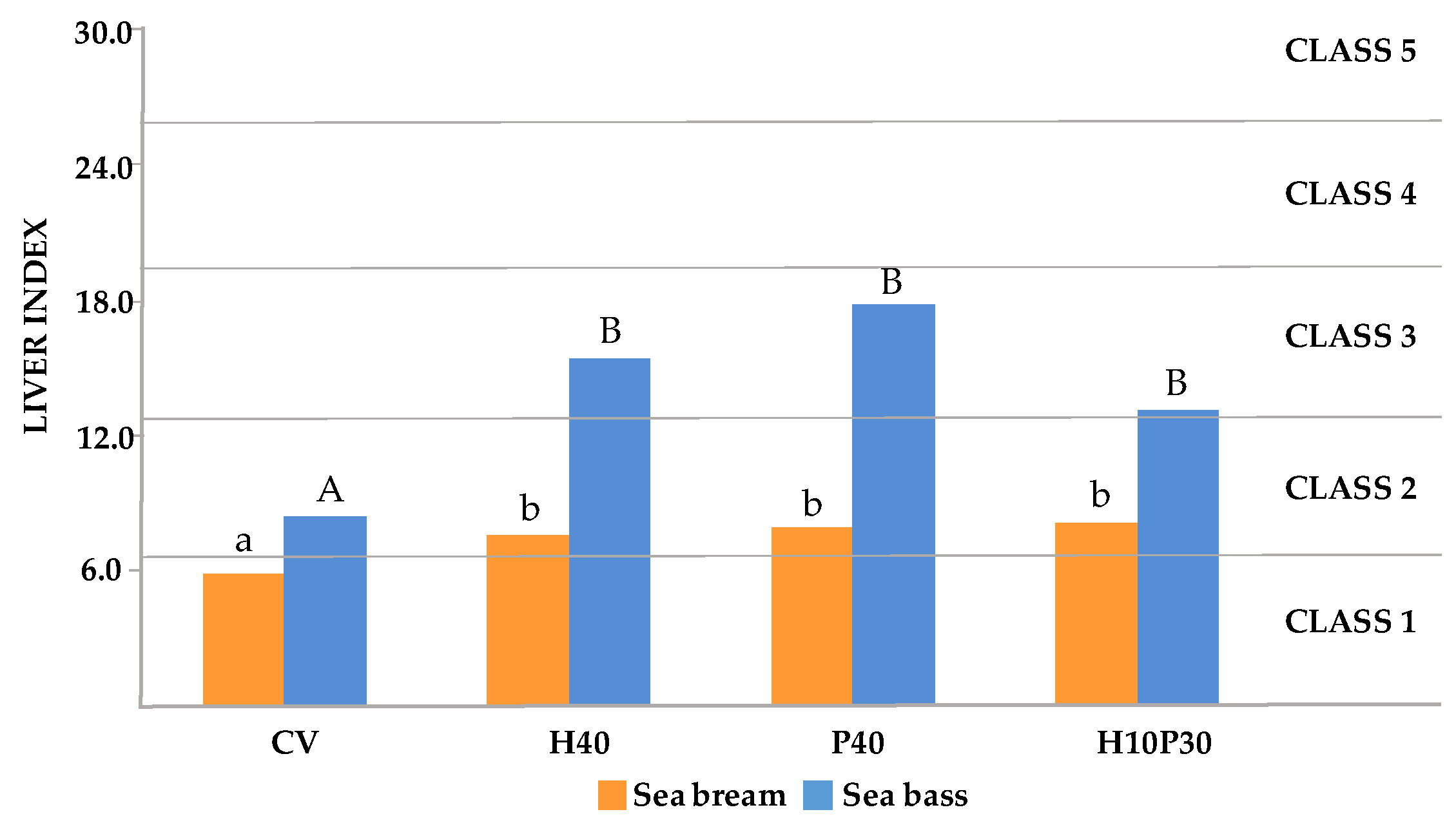
Figure 5.
Box plot of serum biochemical parameters levels in sea bream fed test diets. Different superscripts indicate significant differences among dietary treatments. Data expressed are median, lower and upper quartiles, minimum, and maximum values. GLU=glucose; TAG=triglycerides; CHO=cholesterol, TP=Total protein; AST=aspartate aminotransferase; ALT=alanine aminotransferase.
Figure 5.
Box plot of serum biochemical parameters levels in sea bream fed test diets. Different superscripts indicate significant differences among dietary treatments. Data expressed are median, lower and upper quartiles, minimum, and maximum values. GLU=glucose; TAG=triglycerides; CHO=cholesterol, TP=Total protein; AST=aspartate aminotransferase; ALT=alanine aminotransferase.
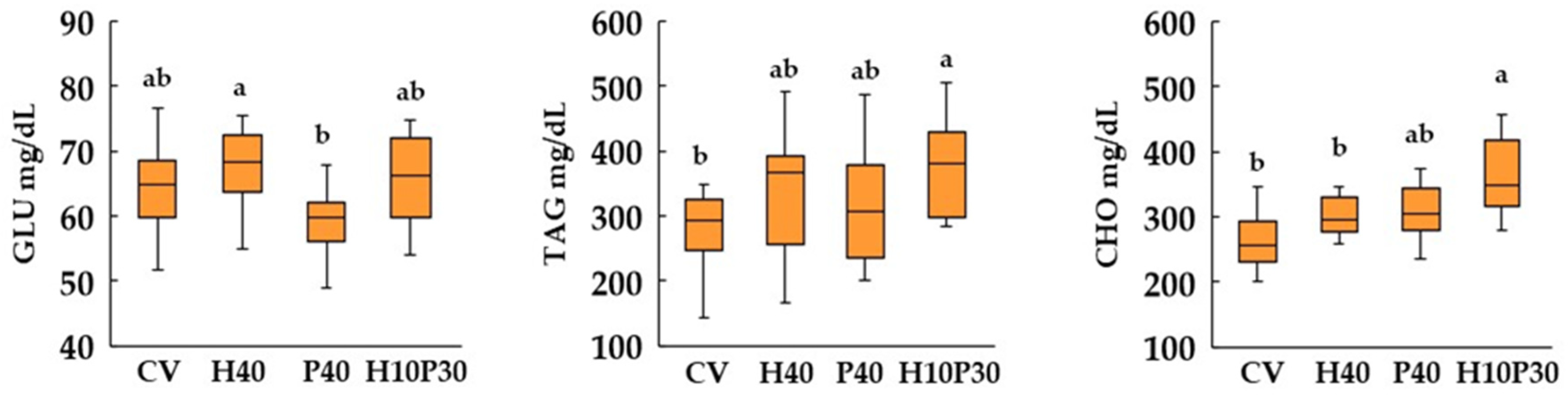
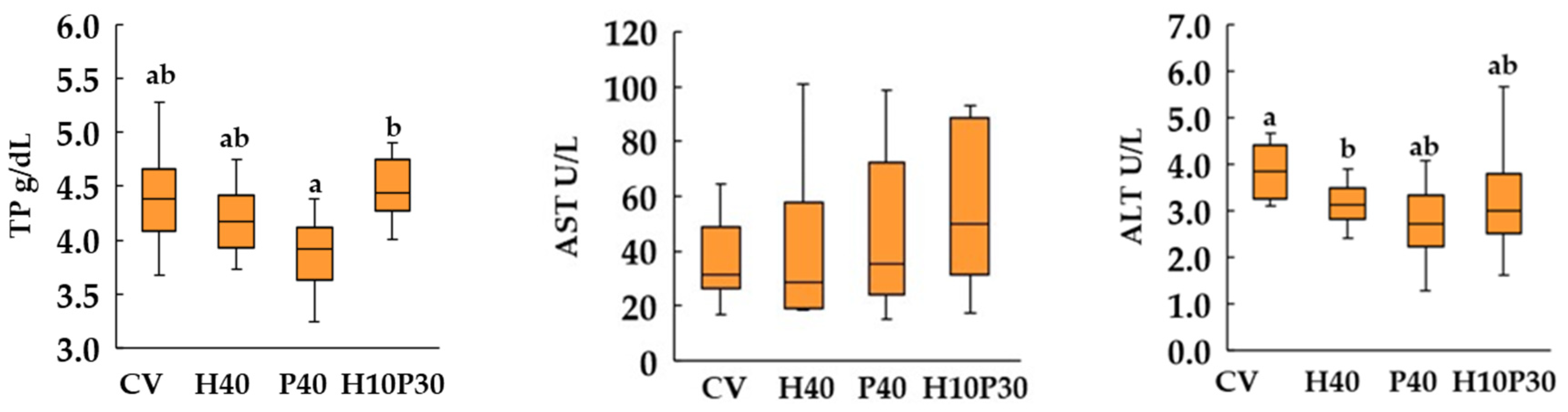
Figure 6.
Box plot of serum biochemical parameters levels in sea bass fed test diets. Different superscripts indicate significant differences among dietary treatments. Data expressed are median, lower and upper quartiles, minimum, and maximum values. GLU=glucose; TAG=triglycerides; CHO=cholesterol, TP=Total protein; AST=aspartate aminotransferase; ALT=alanine aminotransferase.
Figure 6.
Box plot of serum biochemical parameters levels in sea bass fed test diets. Different superscripts indicate significant differences among dietary treatments. Data expressed are median, lower and upper quartiles, minimum, and maximum values. GLU=glucose; TAG=triglycerides; CHO=cholesterol, TP=Total protein; AST=aspartate aminotransferase; ALT=alanine aminotransferase.
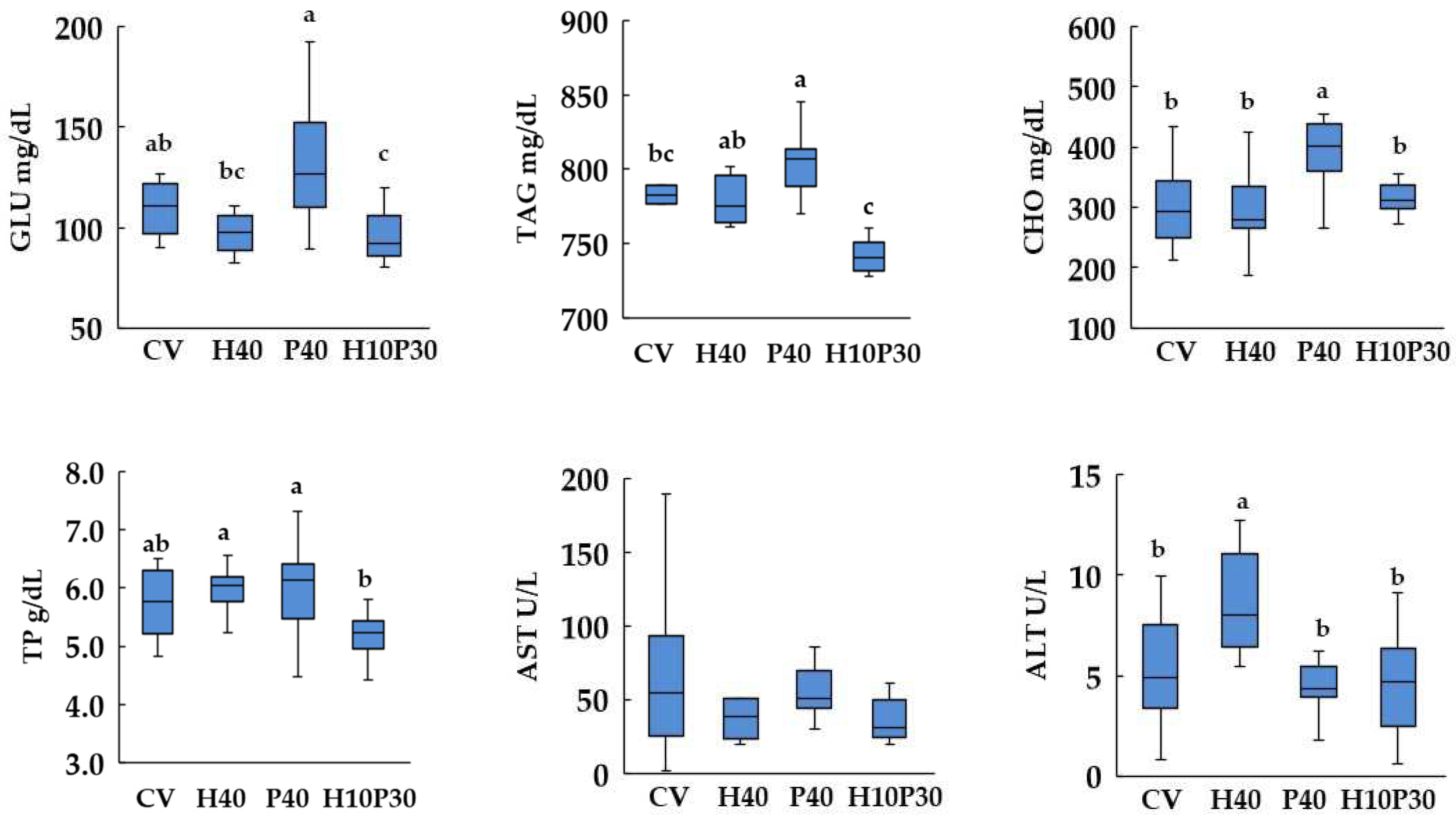
Figure 7.
Plots of FPCA analysis on the liver index, biometric, and biochemical parameters in sea bream and sea bass. GLU=glucose; TAG=triglycerides; CHO=cholesterol, TP=Total protein; AST=aspartate aminotransferase; ALT=alanine aminotransferase. Parameters inside the red circle are correlated to the Liver index (LI). Green dot= positive correlation; yellow dot= negative correlation.
Figure 7.
Plots of FPCA analysis on the liver index, biometric, and biochemical parameters in sea bream and sea bass. GLU=glucose; TAG=triglycerides; CHO=cholesterol, TP=Total protein; AST=aspartate aminotransferase; ALT=alanine aminotransferase. Parameters inside the red circle are correlated to the Liver index (LI). Green dot= positive correlation; yellow dot= negative correlation.
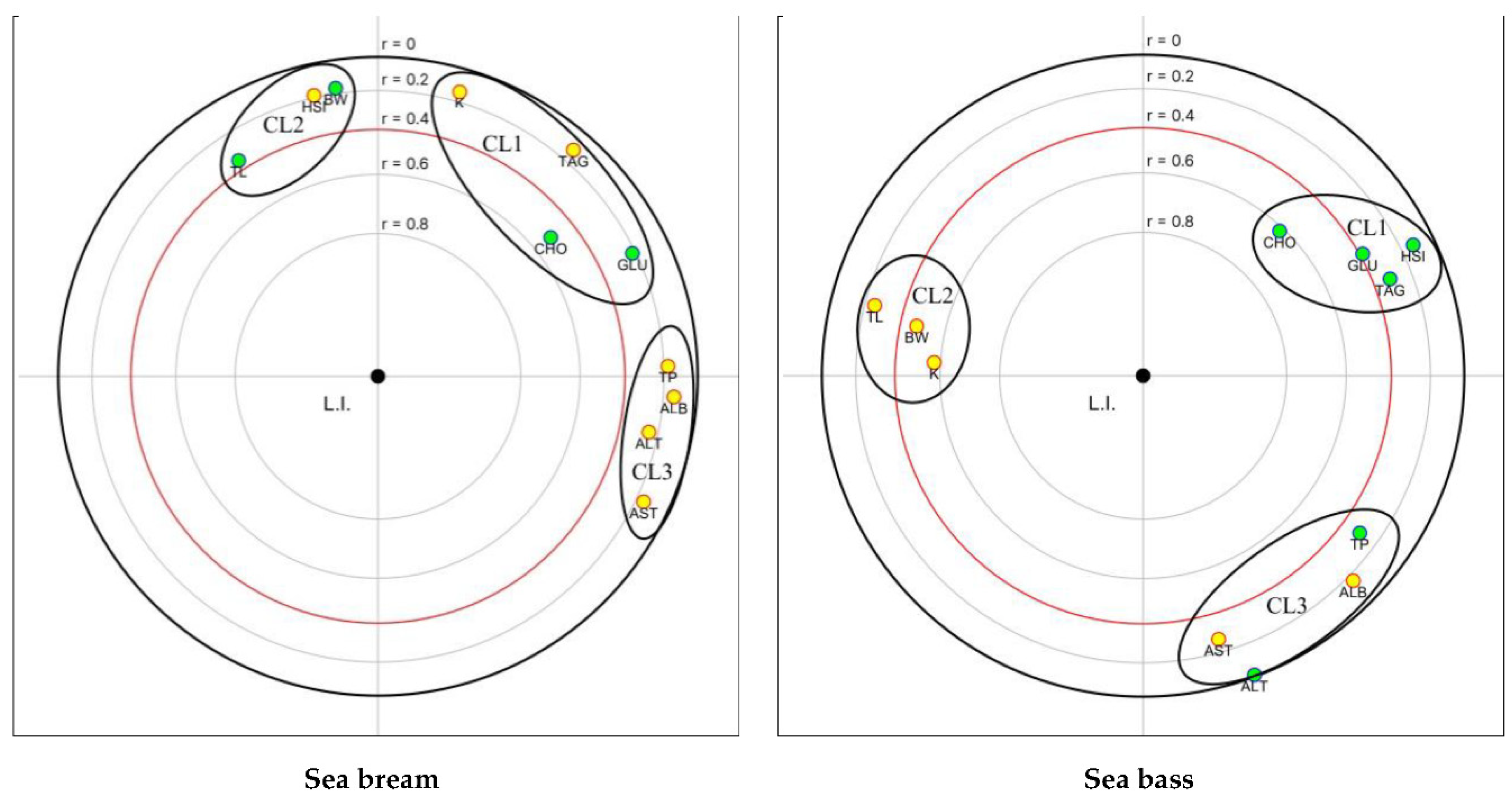
Table 1.
Ingredient (g 100 g-1) and proximate (% as fed) composition of the test diets.
| Ingredient composition (g 100 g-1) | CV | H40 | P40 | H10P30 |
|---|---|---|---|---|
| Vegetable protein mix 1 | 69.0 | 36.6 | 35.4 | 35.4 |
| Hermetia illucens meal 2 | - | 32.4 | - | 8.1 |
| Poultry by-product meal 3 | - | - | 27.5 | 20.6 |
| Feeding stimulants 4 | 5.5 | 5.5 | 5.5 | 5.5 |
| Wheat meal* | 0.4 | 4.5 | 5.6 | 5.5 |
| Whole pea* | 3.0 | 6.0 | 9.0 | 8.8 |
| Fish oil 5 | 6.2 | 6.2 | 6.2 | 6.2 |
| Vegetable oil mix 6 | 11.4 | 5.4 | 8.2 | 7.4 |
| Vit. & Min. Premix 7 | 0.4 | 0.4 | 0.4 | 0.4 |
| Sodium phosphate | 1.6 | 1.0 | 0.3 | 0.2 |
| L-Lysine 8 | 0.5 | 0.2 | 0.1 | 0.1 |
| DL-Methionine 9 | 0.5 | 0.3 | 0.3 | 0.3 |
| Celite | 1.5 | 1.5 | 1.5 | 1.5 |
|
Proximate composition (% as fed): |
||||
| Moisture | 6.7 | 6.0 | 7.1 | 8.7 |
| Protein (Nx6.25) | 45.0 | 45.2 | 45.1 | 45.1 |
| Total lipid | 20.4 | 20.4 | 20.3 | 20.4 |
| Ash | 5.8 | 6.6 | 7.8 | 7.7 |
| Chitin # | 0.02 | 1.54 | 0.02 | 0.40 |
| Energy (MJ/kg) | 21.5 | 21.0 | 21.7 | 21.5 |
1 Vegetable-protein sources mixture (% composition): dehulled, toasted soybean meal, 39; soy protein concentrate-Soycomil, 20; maize gluten, 18; wheat gluten, 15, rapeseed meal, 8. 2 ProteinX™, Protix, Dongen, The Netherlands 3 Poultry by-product meal from Azienda Agricola Tre Valli; Verona, Italy. 4 Feeding stimulants g/100 diet: CPSP90- Sopropeche, France, 3.5; Squid meal, 2.0. 5 Fish oil: Sopropêche France 6 Vegetable oil mixture % composition: rapeseed oil, 56; linseed oil, 26; palm oil, 18 7 Supplying per kg of vitamin supplement: Vit. A, 4,000,000 IU; Vit D3, 850,000 IU; Vit. K3, 5,000 mg; Vit.B1, 4,000 mg; Vit. B2, 10,000 mg: Vit B3, 15,000 mg; Vit. B5, 35,000 mg; Vit B6, 5,000 mg, Vit. B9, 3,000 mg; Vit. B12, 50 mg, Biotin, 350 mg; Choline, 600 mg; Inositol, 150,000 mg. Supplying per kg of mineral supplement: Ca, 77,000 mg; Cu, 2,500 mg; Fe, 30,000 mg; I, 750 mg; Se, 10,000 mg; Zn, 25 mg. 8 L-lysine sulphate, 99%; Ajinomoto EUROLYSINE S.A.S; France 9 DL-Methionine: 99%; EVONIK Nutrition & Care GmbH; Germany * Wherever not specified the ingredients composing the diets were obtained from local providers by Sparos Lda. # Estimated based on chitin content of the ingredients used (squid meal, 0.9% and Hermetia illucens meal 4.69%).
Table 3.
Total length, live weight and body condition indices of sea bream and sea bass fed test diets over 12 and 18 weeks, respectively.
Table 3.
Total length, live weight and body condition indices of sea bream and sea bass fed test diets over 12 and 18 weeks, respectively.
| Test diets | Kruskal Wallis test | |||||
|---|---|---|---|---|---|---|
| CV | H40 | P40 | H10P30 | |||
| Sea bream | TL(cm) | 21.9 ± 0.6 | 22.2 ± 0.8 | 22.3 ± 0.6 | 22.3 ± 0.3 | n.s. |
| BW final (g) | 175.22 ± 17.11b | 188.83 ± 24.78a | 190.27 ± 15.70a | 192.65 ± 9.51a | p< 0.05 | |
| K (cm3) −1 | 1.67 ± 0.12 | 1.72 ± 0.13 | 1.70 ± 0.06 | 1.74 ± 0.07 | n.s. | |
| HSI (%) | 1.29 ± 0.27ab | 1.34 ± 0.14ab | 1.27 ± 0.18b | 1.50 ± 0.19a | p< 0.01 | |
| Sea bass | TL (cm) | 22.8 ± 1.02 | 23.3 ± 0.94 | 23.2 ± 1.38 | 23.5 ± 0.85 | n.s. |
| BW final (g) | 160.7 ± 28.2 | 159.3 ± 20.4 | 154.5 ± 27.9 | 171.2 ± 23.9 | n.s. | |
| K (cm3) −1 | 1.31 ± 0.09a | 1.26 ± 0.08ab | 1.22 ± 0.04b | 1.32 ± 0.06a | p < 0.001 | |
| HSI (%) | 1.80 ± 0.29 | 1.60 ± 0.17 | 1.67 ± 0.32 | 1.52 ± 0.32 | ns | |
Data are expressed as mean±sd. Different superscripts indicate significant differences between dietary treatments set at p< 0.05.
Table 6.
Scoring scheme of liver index developed for sea bream and sea bass and a comparison with the scheme proposed by Zimmerli et al., [56].
Table 6.
Scoring scheme of liver index developed for sea bream and sea bass and a comparison with the scheme proposed by Zimmerli et al., [56].
| Scoring scheme (present study) |
Scoring scheme (Zimmerli et al. 2007) |
Description | |
|---|---|---|---|
| Class 1 | ≤ 6.5 | < 10 | Normal liver structure |
| Class 2 | 6.6-13.0 | 11-20 | Normal liver structure with slight histological alterations |
| Class 3 | 13.1 – 19.5 | 21-30 | Normal liver structure with moderate histological alterations |
| Class 4 | 19.6 – 25.9 | 31-40 | Pronounced alterations of liver structure |
| Class 5 | > 26 | >40 | Severe alterations of liver structure |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



