
Submitted:
07 December 2023
Posted:
08 December 2023
You are already at the latest version
Abstract
Rheumatoid arthritis (RA) is an autoimmune inflammatory disease characterized by chronic synovitis and progressive destruction of cartilage and bone. RA is commonly accompanied by extra-articular comorbidities. The pathogenesis of RA and its comorbidities is complex and not completely elucidated. Assembly of NLRP3 inflammasome activates caspase-1, which induces the maturation of interleukin (IL)-1β and IL-18 and leads to cleavage of gasdermin D with promot-ing pyroptosis. Although the available therapeutic agents are effective for RA treatment, their high cost and increased infection rate are causes for concern. Recent evidence revealed the components of NLRP3 inflammasome as potential therapeutic targets in RA and its comorbidities. This nar-rative review summarizes the current research evidence regarding the pathogenic role and the therapeutic potential of NLRP3 inflammasome in RA and its comorbidities.
Areas covered. We searched the MEDLINE database using the PubMed interface and reviewed English-language literature on NLRP3 inflammasome from 2000 to 2023. Its pathogenic role and therapeutic potential in RA and its comorbidities are the focus of this review.
Expert opinion. NLRP3 inflammasome may play a critical role in both innate and adaptive im-munity, and its dysfunction contributes to the pathogenesis of RA and its comorbidities. Conse-quently, the components of NLRP3 inflammasome signaling represent promising therapeutic tar-gets, and ongoing research might lead to the development of new, effective treatments for RA and its comorbidities.
Keywords:
NLRP3 inflammasome
; comorbidities
; pathogenic player
; therapeutic potential
; rheumatoid arthritis
1. Introduction
The pathology of rheumatoid arthritis (RA) is characterized by an infiltration of macrophages, B cells, and T cells, synovial hyperplasia, and progressive destruction of cartilage and bone, with resultant joint deformities [1,2]. It affects approximately 1% of the population in developed countries [3]. The exact etiopathogenesis of RA is not fully understood, and the proposed causes include genetic factors, various infections, and immune dysregulation with excessive production of proinflammatory mediators such as cytokines and chemokines [4,5,6,7,8]. It has recently been established that dysfunction of innate and adaptive immunity is a critical etiological factor in the development and maintenance of RA [7,9,10]. The pathogenic alterations in innate and adaptive immunity are potential targets for therapeutic intervention in RA [7,9,10]. Besides the typical involvement of joints, RA is often associated with other systemic organ diseases and is complicated by comorbidities and organ dysfunction due to the chronic inflammatory process [11]. It has been estimated that up to 80% of RA patients have one or more comorbidities, resulting in a shortening of life span [12,13]. RA-related comorbidities mainly include atherosclerotic cardiovascular disease (ASCVD), osteoporosis (OP), and interstitial lung disease (ILD) [14,15,16,17,18,19,20]. The presence of comorbidities may affect RA disease activity, become a barrier to optimal disease control, and lead to an impairment in the quality of life (QoL) [21,22]. RA-related comorbidities are often sub-optimally managed, and effective treatment of RA-related comorbidities is an unmet need for rheumatologists in clinical practice.
Nucleotide-binding domain leucine-rich repeat-containing receptors (NLRs)-containing a pyrin domain (NLRPs), the major components of inflammasomes, play a pathogenic role in innate immunity and inflammation [23,24,25]. NLRP3 inflammasome, a supramolecular cytoplasmic complex, may respond to stimuli such as adenosine triphosphate (ATP) and then recruit caspase-1, which cleaves pro-IL-1β and pro-IL-18 into their active biologic forms [26,27,28]. It is established that NLRP3 Inflammasome dysregulation causes autoinflammatory diseases (AIDs) [29,30,31,32]; such dysfunction may similarly contribute to RA pathogenesis [10,33,34,35,36,37,38,39,40,41,42], although the underlying mechanisms are yet fully elucidated.
Targeting the complex pathogenic factors in RA, various emerging new agents are available for the treatment of this disease [43,44]. Recent guidelines for RA treatment rank Janus kinase inhibitors (JAKi) or targeted synthetic biologic disease-modifying anti-rheumatic drugs (tsDMARDs) in parallel with biologic DMARDs (bDMARDs) as the options for patients who are refractory to initial conventional synthetic DMARDs (csDMARDs) therapy [45,46,47]. Nevertheless, a proportion of RA patients still fail to respond to current therapies [48,49,50]. With the high cost of b/tsDMARDs and their associated increased infection risk, alternative or add-on therapeutic agents targeting immune or inflammatory responses are worth exploring.
Supporting the significance of NLRP3 inflammasome in the pathogenesis of RA and its comorbidities, increasing clinical and pre-clinical evidence has revealed the components of NLRP3 inflammasome as potential therapeutic targets in RA [51,52,53]. This review aims to summarize the current research evidence on the pathogenic role of the NLRP3 inflammasome signaling pathway and its clinical implications as the therapeutic target in RA and its comorbidities.
2. Materials and Methods
2.1. Literature Search
This review focuses on the updated research regarding the NLRP3 inflammasome as a pathogenic player and its therapeutic potential in RA. We searched the MEDLINE database using the PubMed interface and reviewed English-language literature as of 31 October 2023, from 2000 to 2023. The search keywords included pathogenesis, innate immunity, adaptive immunity, immune response, inflammation, NLRP3 inflammasome, proinflammatory cytokines, pyroptosis, IL-1β, IL-18, RA, RA-related comorbidities, ASCVD, OP, ILD, clinical implication, and therapeutic potential. Duplicates and manuscripts with incomplete data were excluded. The details of the search strategy are illustrated in Figure 1.
2.2. Study Selection
Three authors (PK Chen, KT Tang, and DY Chen) independently assessed the titles and abstracts from the search results and retrieved the relevant full-text articles. Two authors (KT Tang and DY Chen) independently evaluated the full-text articles for eligibility. Articles were selected if they 1) were probably relevant to the pathogenic role of the NLRP3 inflammasome in RA and its comorbidities, and 2) were potentially relevant to its therapeutic potential in this disease, including clinical trials, cohorts, case reports, and case-control studies.
2.3. Data Extraction
The authors extracted data from these studies electronically. Information regarding innate immunity, adaptive immunity, immune response, pathogenesis, the NLRP3 inflammasome, proinflammatory cytokines, pyroptosis, IL-1β, IL-18, clinical implication, therapeutic potential, RA, RA-related comorbidities, ASCVD, OP, or ILD was recorded from each study. The influence of relevant drugs, including small molecule inhibitors, natural products, corticosteroids, csDMARDs, bDMARDs, tsDMARDs, and targeted therapeutics for the NLRP3 inflammasome in RA and its comorbidities was also documented.
3. Etiopathogenesis of RA and Its Comorbidities
The pathogenesis of RA is multifactorial and complex, including environmental factors, genetic variables, and immune dysregulation such as inflammasome activation and cytokine-mediated inflammation [1,2,3,4,5,6,7,8,54,55]. Kolly et al. demonstrated that the components of NLRP3 inflammasome were highly expressed in the synovia of RA patients [33]. Recent evidence suggests that the percentages of CD4+ T cells with activated caspase-1 are significantly higher in RA patients compared with normal controls. Pharmacological and genetic inhibition of the DNA repair nuclease MRE11A may cause mitochondrial dysfunction in CD4+ T cells, leading to the NLRP3 inflammasome assembly, caspase-1 activation, and pyroptosis in RA CD4+ T cells [56]. Increasing evidence indicates that the NLRP3 inflammasome plays a critical role in the pathogenesis of RA [10,45,46,47,48,49,50,51,52,53,54].
Atherosclerosis is a chronic inflammatory process that leads to vascular atheromatous plaque buildup and the development of full-blown ASCVD [57]. The high ASCVD burden in RA patients [58,59] may result from a combination of traditional risk factors, disease-specific factors, chronic inflammation, genetic components, and the use of medications [60,61,62,63]. Variants of the gene encoding apolipoprotein (apo)E have been shown to be related to ASCVD risk in RA patients [64]. Among the particles of low-density lipoprotein (LDL), a lipoprotein class, the density, size, electric charge, and composition are varied. Supporting the lipid paradox hypothesis in RA [65], studies have shown an inverse correlation between RA-related inflammation and circulating levels of LDL cholesterol (LDL-C) [66,67]. Electronegative LDL, a naturally occurring LDL, exerts potent atherogenic effects in cells and animals without undergoing ex vivo oxidation [68]. Elevated circulating levels of L5, the most electronegative subfraction of LDL-C have been observed in RA patients and may be a predictor of ASCVD in this disease [67]. Besides, a high L5 percentage was significantly associated with elevated expression of the gene encoding integrin CD11c, which was linked to carotid arterial plaque formation [69].
Osteoporosis (OP) or bone fragility arises from a complex interaction of traditional risk factors and disease inflammation in RA. RA patients have an elevated risk of OP or osteoporotic fractures than healthy controls [18,70]. Using Mendelian randomization analysis, Yu et al. revealed that genetically determined RA was linked to estimated bone mineral density (eBMD) and fracture risk [71]. Similarly, osteoporosis risk could be causally increased by the presence of anti-citrullinated peptide antibodies (ACPA) in Asians [71,72]. The presence of ACPA, prolonged RA disease duration, significant exposure to corticosteroids, decreased physical activity, or history of low trauma fracture are the risk factors of OP or fragility fracture in RA [72,73].
ILD is the leading cause of mortality and the most common pulmonary manifestation of RA [20,74]. Estimated approximately 30% of RA patients have subclinical ILD as shown by high-resolution computerized tomography (HRCT) scans [75]. Although the exact pathogenesis of ILD in RA remains unclear [74,75,76,77], smoking, male gender, older age, high titers of ACPA, disease duration, and positivity of Human leukocyte antigen (HLA)-DR4 were probable risk factors for RA-ILD [78,79,80,81,82]. A study of the Western population revealed the MUC5B promoter variant rs35705950 as a genetic risk factor for developing RA-ILD, particularly in those with the usual interstitial pneumonia (UIP) pattern [83]. Shirai et al. identified rs12702634 at RPA3-UMAD1 as a risk variant for RA-ILD in the Japanese population [84]. Citrullination, a post-translational modification characterized by the conversion of arginine to citrulline, and the emergence of ACPA probably contribute to RA-ILD by releasing neutrophil extracellular traps [85]. Recently, Zhang et al. revealed a pathogenic role of IL-17 in murine pulmonary fibrosis and RA-ILD [86]. Air pollutants, such as the elements of particulate matter (PM)<2.5μm in size (PM2.5), may trigger the development of ILD in genetically susceptible patients [87].
4. NLRP3-Inflammasome
4.1. NLRP3-Inflammasome Signaling in Immune Responses and Inflammation
The innate immune system encompasses the germline-encoded pattern recognition receptors (PRRs), including Toll-like receptors (TLRs) and NLRs [35,88,89]. The NLRP3 inflammasome is a cytoplasmic protein complex with a key role in the innate immune response and inflammatory reaction. Assembly of the NLRP3 inflammasome recruits and activates caspase-1, which induces the maturation and release of proinflammatory cytokines, including IL-1β and IL-18 [26,27,28]. Activation of the NLRP3 inflammasome also leads to cleavage of gasdermin D (GSDMD) at the GSDMD-N terminus [28,90] and promotes a lytic form of cell death, pyroptosis, with pores formation in the cell membrane and release of IL-1β and IL-18 [91,92,93]. The NLRP3 inflammasome signaling may play a critical role in both innate and adaptive immunity and act as a checkpoint in innate immunity to lead to skewed adaptive immune responses in autoimmune diseases [10].
4.2. NLRP3-Inflammasome Activation and Regulation in RA Pathogenesis.
The dysregulation of the NLRP3 inflammasome is linked to a variety of inflammatory diseases such as RA [10,33,34,35,36,37,38,39,40,41,42,94]. Zhang et al. demonstrated that NLRP3 was highly expressed in the synovial proliferation and subchondral vasculitis areas in the paws of collagen-induced arthritis (CIA) mice compared to control mice [34]. NLRP3 mRNA levels were upregulated in the synovia of RA patients compared to OA patients [38]. Guo et al. also documented that NLRP3 inflammasome was highly activated in the synovia from RA patients and murine models [39]. Recent studies have found that pyroptosis is involved in the occurrence and progression of RA, and large amounts of IL-1β and IL-18 are present in RA patients. In RA, complement C1q and pentaxin 3 (PTX3) in monocytes synergistically promote NLRP3 inflammasome over-activation and pyroptosis [40]. Wu et al. revealed that acid-sensitive ion channel-1a mediates chondrocyte pyroptosis in arthritis by promoting NLRP3 inflammatory vesicle assembly, caspase-1 expression, and IL-1β and IL-18 release [95]. Besides, Ca2+ and cyclic AMP are two key molecular regulators of the NLRP3 inflammasome [96]. Werner et al. demonstrated that increased extracellular Ca2+ ([Ca2+]ex) could induce inflammation through promoting NLRP3 inflammasome assembly and IL-1β release [97]. They further proposed that increased [Ca2+]ex, calciprotein particles, and proinflammatory cytokines drive a vicious cycle of inflammation and bone destruction in RA [97]. Besides, the stimulation of anti-citrullinated peptide antibodies (ACPA) could activate pannexin channels with the release of ATP and promote the NLRP3 inflammasome activation and IL-1β production in RA [41]. These observations suggest that the activation of NLRP3 inflammasome signaling and the ensuing overproduction of inflammatory cytokines are key to the pathogenesis of RA [35,36,37,38,39,40,41,42,98].
4.3. The Genetic Predisposition of NLRP3 Inflammasome in RA.
Nucleotide polymorphisms occur within the regulatory region of cytokine genes, and some are associated with an altered rate of gene expression. The NLRP3 inflammasome gene polymorphisms contribute to susceptibility, disease activity, or disease severity in RA [99,100,101,102]. The genetic variants of the NLRP3 inflammasome can also affect the therapeutic response to TNF-α inhibitors in RA patients [103,104,105].
4.4. The Involvement of NLRP3 Inflammasome Activation in RA-Related Comorbidities
Elevated cholesterol and genetic predisposition may trigger the activation of NLRP3 inflammasome signaling pathway and promote the development and progression of ASCVD [106,107,108]. Karasawa et al. revealed that the crystallization of released cholesterol in the atherosclerotic plaque may activate NLRP3 inflammasome with the production of IL-1β and IL-18 [106]. In contrast, the lack of caspase-1 exhibits a protective effect against the evolution of atherosclerotic lesions, further resonating with the causative association of NLRP3 inflammasome with atherosclerosis [106]. Rhoads et al. also demonstrated that oxidized LDL (oxLDL) immune complex induced inflammasome activation through a more robust mechanism than oxLDL alone did [109]. Besides, an association study has identified the Q705K polymorphism(rs35829419) in the NLRP3 gene as a protective factor against the risk of developing infarction in females [110]. The C10X variants (rs2043211) in the CARD8 gene were related to the lower expression of CARD8 in carotid plaques in a Swedish cohort [111]. In a Chinese cohort, a variant (rs2043211) in the CARD8 gene was associated with ischemic stroke [112]. Kastbom et al. showed that genetic variants of the NLRP3 inflammasome were associated with ischemic stroke in Swedish patients with RA [113]. These observations suggest the close link between the NLRP3 inflammasome signaling pathway and ASCVD in inflammatory diseases such as RA.
Increasing evidence also supports the association between the NLRP3 inflammasome signaling pathway and OP [114,115]. In an ovariectomized (postmenopausal) OP rats’ model, inhibition of the NLRP3 could increase osteoblasts number and bone density, suggesting a pathogenic role of NLRP3 in OP [116]. The activation of NLRP3 inflammasome contributes to the maturation of downstream proinflammatory cytokines, IL-1β and IL-18, and pyroptosis [26,27,28]. Pyroptosis of osteoblasts may participate in OP pathogenesis [117]. Lei et al. revealed that IL-17 could induce pyroptosis of murine primary osteoblasts in the NLRP3-mediated pathway, which further promoted the release of IL-1β and receptor activator of nuclear factor-kappa B ligand (RANKL) and exacerbated the progression of OP [118]. Interestingly, IL-1 may also participate in the TNF-α-mediated inflammatory bone loss [119]. He et al. reported an association of IL-1β haplotype with OP susceptibility in the Chinese Han population [120]. Besides, IL-18 could upregulate the production of key osteoclastogenic mediators and increase bone loss in RA [121].
There are strong mechanistic similarities between usual interstitial pneumonitis (UIP), the most common pattern of RA-ILD, and idiopathic pulmonary fibrosis (IPF) in RA. However, Lasithiotaki et al. revealed distinct NLRP3 inflammasome activation profiles between RA-UIP and IPF [122], with significantly higher levels of IL-1β and IL-18 in bronchoalveolar lavage fluid (BALF) from RA-UIP patients compared with IPF patients. Intracellular IL-1β levels were also augmented in RA-UIP BALF cells upon NLRP3 inflammasome stimulation [122]. These findings suggest that NLRP3 inflammasome is involved in the pathogenesis of RA-ILD. Similarly, Aberrant activation of NLRP3 inflammasome is observed in scleroderma-associated ILD [123,124]. Ramos-Martinez and colleagues demonstrated enhanced activity of the NLRP3 inflammasome in the lungs of patients with anti-synthetase syndrome [125]. Hence, NLRP3 inflammasome and associated cytokines may participate in the pathogenesis of autoimmune diseases-related ILD.
5. Therapeutic Potential by Targeting NLRP3 Inflammasome
5.1. Small Molecule Inhibitors
With increasing evidence supporting the importance of NLRP3 inflammasome in RA pathogenesis [10,33,34,35,36,37,38,39,40,41,42], there has been significant interest in developing therapeutic agents targeting the components of NLRP3 inflammasome signaling [51,52,53]. One approach is to develop small molecule inhibitors that can block the activation of the NLRP3 inflammasome. As illustrated in Table 1, several compounds have been identified as inhibitors of the components of the NLRP3 inflammasome signaling pathway, including MCC950 [39,126,127], VX-765 [128], and disulfiram [129]. Guo et al. demonstrated that treatment with MCC950, a selective NLRP3 inhibitor, led to reduced joint inflammation and bone destruction in the murine RA model [39]. Another approach is to target the downstream products of NLRP3 inflammasome activation, such as the sophisticated pyroptosis pathway [129,130] and the production of pro-inflammatory cytokines. MCC950 could reduce macrophage infiltration and atherosclerotic lesion size through attenuating inflammation and pyroptosis in atherosclerosis murine models [131,132]. Li et al. reported that VX-765 could inhibit atherosclerosis in ApoE-deficient mice by modulating the pyroptosis of vascular smooth muscle cells [133]. Given the pathogenic role of IL-18 in RA-associated OP [121], IL-18BP, an antagonist of IL-18, is speculated to be effective in the management of OP [134]. Mansoori et al. similarly revealed that high serum IL-18BP was associated with a low risk of osteoporosis in postmenopausal women [134].
5.2. Natural Products
Besides the small molecule inhibitors, several natural products exhibit anti-inflammatory effects through targeting the NLRP3 inflammasome signaling pathway (Table 1). Celastrol, a natural product isolated from Tripterygium wilfordii, has displayed therapeutic potential in inflammatory diseases, such as RA. One recent study showed the attenuating effects of celastrol on parvovirus B19-NS1-induced NLRP3 inflammasome activation in macrophages [135]. Jing et al. also revealed that celastrol inhibited inflammation by inhibiting the reactive oxygen species- NF-κB-NLRP3 inflammasome axis and relieved RA symptoms [136]. Baihu-Guizhi decoction (BHGZD), a traditional Chinese-medicine-originated disease-modifying anti-rheumatic prescription, may reduce the disease activity of RA [137]. Li et al. reported that BHGZD could suppress the NLRP3 inflammasome activation and GSDMD-mediated pyroptosis by inhibiting NF-κB via TLR4/PI3K/AKT signaling in an adjuvant-induced arthritis-modified rat model [138]. Sulforaphene, extracted from radish seeds, has been demonstrated to suppress the M1 polarization of macrophages and reduce synovitis in the CIA murine model [139]. Osthole, a characteristic coumarin compound in Angelicae pubescentis radix, can improve arthritis in the CIA rat model by inhibiting inflammation and oxidative stress [140]. Jiang et al. further showed that Osthole could Inhibit NLRP3 inflammasome activation by regulating mitochondrial homeostasis [141]. Scropolioside B, isolated from a Tibetan medicine, Scrophularia dentada Royle ex Benth., could inhibit NF-κB activity, reduce NLRP3 expression, and suppress the maturation and release of IL-1β, suggesting its therapeutic potential in RA and its associated atherosclerosis [142]. Cao and colleagues demonstrated that Wedelolactone, derived from Eclipta alba, could ameliorate synovial inflammation, cardiac complication, and fibrosis by inhibiting the activation of the NF-κB/NLRP3 inflammasome pathway [143].
5.3. Disease-Modifying Anti-Rheumatic Drugs (DMARDs)
Hydroxychloroquine (HCQ), a known ion channel inhibitor, is a commonly used and effective treatment for RA. Schroeder et al. demonstrated that HCQ could inhibit Ca2+-activated K+ channels and suppress inflammasome activation [144]. Cui et al. also revealed that chloroquine inhibited the activation of NLRP3 inflammasome in a murine model of hyperuricemic nephropathy [145]. Since IL-1β, an NLRPP3 inflammasome downstream cytokine, promotes synovial inflammation in RA patients, the IL-1β receptor antagonist anakinra has been approved for active RA patients unresponsive to csDMARDs [146]. Targeting IL-1β might also have therapeutic potential in RA-associated comorbidities. The Canakinumab Antiinflammatory Thrombosis Outcome Study (CANTOS) study showed the benefit of canakinumab, an IL-1β blockade, in patients with previous myocardial infarction [147].
5.4. Janus Kinase Inhibitors (JAKi)
Janus kinase inhibitors (JAKi) exert their therapeutic effects by blocking JAK/STAT-mediated signaling implicated in RA pathogenesis. Although tofacitinib, a JAK1/JAK3 inhibitor, is effective in RA treatment [48,49,50], the underlying mechanisms of drug action remain obscure. Yang et al. demonstrated that tofacitinib could restore the balance of γδTreg/γδT17 cells in RA by inhibiting the NLRP3 inflammasome [148].
5.5. microRNAs (miRNAs) and Stem Cells
Multiple miRNAs are shown to be involved in the post-transcriptional regulated expression of NLRP3 inflammasome [149,150]. The miRNAs are short non-coding RNAs composed of approximately 20 to 24 nucleotides that mediate messenger (m)RNA cleavage, translational repression, or mRNA destabilization [151,152]. Xie et al. revealed an elevated expression of miRNA-33 as a positive regulator of the NLRP3 inflammasome in RA patients [153]. Yang et al. also showed that miRNA-30a might suppress the expression of NLRP3 inflammasome in macrophages and regulate inflammation in RA [154]. A recent study revealed that miR-223 carried by bone marrow stem cells-derived exosomes can target NLRP3 and inhibit the activation of inflammasome in macrophages and rats RA models [155]. Liao et al. also demonstrated that neutrophil-derived exosomal miR-223 could suppress the NLRP3-inflammasome signaling and IL-18 production in macrophages in an in vitro assay [156]. Besides, long non-coding RNAs (lncRNAs), the non-protein-coding transcripts greater than 200 nucleotides, have emerged as novel players in gene regulation [157,158]. The LncRNAs have been shown to be the key regulators of inflammatory responses [159]. Wang et al. reported that LncRNA MIAT (myocardial infarction-associated transcript) could downregulate IL-1β and TNF-α to suppress macrophage inflammation in the 774A.1 cell-based assay [160]. Recently, cell-based therapies using mesenchymal stem cells (MSCs) have been spotlighted as a promising strategy for the management of RA [161]. Shin et al. demonstrated that human umbilical cord blood-MSCs (hUCB-MSCs) could downregulate the activation of the NLRP3 inflammasome via a paracrine loop of IL-1β signaling in the CIA murine model [162], suggesting the therapeutic potential of hUCB-MSCs in RA treatment.
Briefly, targeting the NLRP3 inflammasome signaling pathway represents a promising therapeutic strategy for RA and its comorbidities, and ongoing research would facilitate the development of novel and effective treatments for this disease.
6. Conclusions
With progressive insight into the pathogenesis of RA and its comorbidities, the role of the NLRP3 inflammasome is gaining importance in this disease [10,21,22,23,24,25,26,27,28,29,30]. Hence, the components of the NLRP3 inflammasome signaling pathway would be a promising therapeutic target in RA and its comorbidities [39,40,41]. Based on the available evidence, we summarized the data regarding the pivotal role of the NLRP3 inflammasome in the pathogenesis of RA and its comorbidities and its therapeutic potential in Figure 2. Hopefully, this will lead to effective novel therapies for RA and its comorbidities.
Authors’ Contributions
All authors made substantive intellectual contributions to this review and approved the final manuscript. P-KC, K-TT, and D-YC performed the literature search and appraised the selected articles. P-KC and D-YC drafted the manuscript. K-TT and D-YC revised the final manuscript.
Data Availability
No data were used to support this review article.
Conflicts of Interest Statement
All authors report no financial interests or conflicts of interest.
Funding
This research received no external funding.
References
- Smolen, J.S.; Aletaha, D.; McInnes, I.B. Rheumatoid arthritis. Lancet. 2016, 388, 2023–2038. [Google Scholar] [CrossRef]
- Gravallese, E.M.; Firestein, G.S. Rheumatoid arthritis: common origins, divergent mechanisms. N Engl J Med. 2023, 388, 529–542. [Google Scholar] [CrossRef]
- Lai, C.H.; Lai, M.S.; Lai, K.L.; Chen, H.H.; Chiu, Y.M. Nationwide population-based epidemiologic study of rheumatoid arthritis in Taiwan. Clin Exp Rheumatol. 2012, 30, 358–363. [Google Scholar]
- Kurko, J.; Besenyei, T.; Laki, J.; Glant, T.T.; Mikecz, K.; Szekanecz, Z. Genetics of rheumatoid arthritis - a comprehensive review. Clin Rev Allergy Immunol. 2013, 45, 170–179. [Google Scholar] [CrossRef]
- Viatte, S.; Plant, D.; Raychaudhuri, S. Genetics and epigenetics of rheumatoid arthritis. Nat Rev Rheumatol. 2013, 9, 141–153. [Google Scholar] [CrossRef]
- Furst, D.E.; Emery, P. Rheumatoid arthritis pathophysiology: update on emerging cytokine and cytokine-associated cell targets. Rheumatology (Oxford) 2014, 53, 1560–1569. [Google Scholar] [CrossRef]
- Scherer, H.U.; Häupl, T.; Burmester, G.R. The etiology of rheumatoid arthritis. J Autoimmun. 2020, 110, 102400. [Google Scholar] [CrossRef]
- Zhao, T.; Wei, Y.; Zhu, Y.; Xie, Z.; Hai, Q.; Li, Z.; Qin, D. Gut microbiota and rheuamtoid arthritis: from pathogenesis to novel therapeutic opportunisties. Front Immunol. 2022, 13, 1007165. [Google Scholar] [CrossRef]
- Szekanecz, Z.; McInnes, I.B.; Schett, G.; Szamosi, S.; Benkő, S.; Szűcs, G. Autoinflammation and autoimmunity across rheumatic and musculoskeletal diseases. Nat Rev Rheumatol. 2021, 17, 585–595. [Google Scholar] [CrossRef]
- Zhang, Y.; Yang, W.; Li, W.; Zhao, Y. NLRP3 inflammasome: checkpoint connecting innate and adaptive immunity in autoimmune diseases. Front Immunol. 2021, 12, 732933. [Google Scholar] [CrossRef]
- Mitrović, J.; Hrkač, S.; Tečer, J.; Golob, M.; Ljilja, Posavec, A.; Kolar, Mitrović, H.; Grgurević, L. Pathogenesis of extraarticular manifestations in rheumatoid arthritis-a comprehensive review. Biomedicines 2023, 11, 1262. [Google Scholar] [CrossRef] [PubMed]
- Emery, P.; Hajjaj-Hassouni, N.; Harigai, M.; Luo, S.F.; Kurucz, R.; Maciel, G.; Mola, E.M.; Montecucco, C.M.; McInnes, I.; Radner, H.; et al. Prevalence of comorbidities in rheumatoid arthritis and evaluation of their monitoring: results of an international, cross-sectional study (COMORA). Ann Rheum Dis. 2014, 73, 62–68. [Google Scholar]
- Gabriel, S.E.; Michaud, K. Epidemiological studies in incidence, prevalence, mortality and comorbidity of the rheumatic diseases. Arthritis Res Thera. 2009, 11, 229. [Google Scholar] [CrossRef] [PubMed]
- Symmons, D.P.; Gabriel, S.E. Epidemiology of CVD in rheumatic disease, with a focus on RA and SLE. Nat Rev Rheumatol. 2011, 7, 399–408. [Google Scholar] [CrossRef]
- Avina-Zubieta, J.A.; Thomas, J.; Sadatsafavi, M.; Lehman, A.J.; Lacaille, D. Risk of incident cardiovascular events in patients with rheumatoid arthritis: a meta-analysis of observational studies. Ann Rheum Dis 2012, 71, 1524–1529. [Google Scholar] [CrossRef]
- Rho, Y.H.; Chung, C.P.; Oeser, A.; Solus, J.; Asanuma, Y.; Sokka, T.; Pincus, T.; Raggi, P.; Gebretsadik, T.; Shintani, A.; et al. Inflammatory mediators and premature coronary atherosclerosis in rheumatoid arthritis. Arthritis Rheum 2009, 61, 1580–1585. [Google Scholar] [CrossRef] [PubMed]
- Haugeberg, G.; Uhlig, T.; Falch, J.A.; Halse, J.I.; Kvien, T.K. Bone mineral density and frequency of osteoporosis in female patients with rheumatoid arthritis. Arthritis Rheum. 2000, 43, 522–530. [Google Scholar] [CrossRef]
- Kim, S.Y.; Schneeweiss, S.; Liu, J.; Daniel, G.W.; Chang, C.L.; Garneau, K.; Solomon, D.H. Risk of osteoporotic fracture in a large population-based cohort of patients with rheumatoid arthritis. Arthritis Res Thera. 2010, 12, R154. [Google Scholar] [CrossRef] [PubMed]
- Shaw, M.; Collins, B.F.; Ho, L.A.; Raghu, G. Rheumatoid arthritis-associated lung disease. Eur Respir Rev. 2015, 24, 1–16. [Google Scholar] [CrossRef]
- Ng, K.H.; Chen, D.Y.; Lin, C.H.; Chao, W.C.; Chen, Y.M.; Chen, Y.H.; Huang, W.N.; Hsieh, T.Y.; Lai, K.L.; Tang, K.T.; et al. Risk of interstitial lung disease in patients with newly diagnosed systemic autoimmune rheumatic disease: a nationwide, population-based cohort study. Semin Arthritis Rheum. 2020, 50, 840–845. [Google Scholar] [CrossRef]
- Tymms, K.; Zochling, J.; Scott, J.; Bird, P.; Burnet, S.; de Jager, J.; Griffiths, H.; Nicholls, D.; Roberts, L.; Arnold, M.; et al. Barriers to optimal disease control for rheumatoid arthritis patients with moderate and high disease activity. Arthritis Care Res 2014, 66, 190–196. [Google Scholar] [CrossRef]
- Scott, I.C.; Machin, A.; Mallen, C.D.; Hider, S.L. The extra-articular impacts of rheumatoid arthritis: moving towards holistic care. BMC Rheumatology 2018, 2, 32. [Google Scholar] [CrossRef] [PubMed]
- Muruve, D.A.; Petrilli, V.; Zaiss, A.K.; White, L.R.; Clark, S.A.; Ross, P.J.; Tschopp, J. The inflammasome recognizes cytosolic microbial and host DNA and triggers an innate immune response. Nature 2008, 452, 103–107. [Google Scholar] [CrossRef] [PubMed]
- Martinon, F.; Gaide, O.; Petrilli, V.; Mayor, A.; Tschopp, J. NALP inflammasome: a central role in innate immunity. Semin Immunopathol. 2007, 29, 29,213–229. [Google Scholar] [CrossRef] [PubMed]
- Shaw, P.J.; McDermott, M.F.; Kanneganti, T.D. Inflammasomes and autoimmunity. Trends Mol Med. 2011, 17, 57–64. [Google Scholar] [CrossRef]
- Schroder, K.; Tschopp, J. The inflammasomes. Cell 2010, 140, 821–832. [Google Scholar] [CrossRef] [PubMed]
- Swanson, K.V.; Deng, M.; Ting, J.P.Y. The NLRP3 inflammasome: molecular activation and regulation to therapeutics. Nat Rev Immunol. 2019, 9, 477–489. [Google Scholar] [CrossRef]
- Xu, J.; Núñez, G. The NLRP3 inflammasome: activation and regulation. Trends Biochemical Sciences 2023, 48, 331–344. [Google Scholar] [CrossRef]
- Kim, M.L.; Chae, J.J.; Park, Y.H.; De Nardo, D.; Stirzaker, R.A.; Ko, H.J.; Tye, H.; Cengia, L.; DiRago, L.; Metcalf, D.; et al. Aberrant actin depolymerization triggers the pyrin inflammasome and autoinflammatory disease that is dependent on IL-18, not IL-1β. J Exp Med. 2015; 212, 927–938. [Google Scholar]
- Hsieh, C.W.; Chen, Y.M.; Lin, C.C.; Tang, K.T.; Chen, H.H.; Hung, W.T.; Lai, K.L.; Chen, D.Y. Elevated Expression of the NLRP3 Inflammasome and Its Correlation with Disease Activity in Adult-onset Still Disease. J Rheumatol. 2017, 44, 1142–1150. [Google Scholar] [CrossRef]
- Gattorno, M.; Martini, A. Beyond the NLRP3 inflammasome: autoinflammatory diseases reach adolescence. Arthritis Rheum 2013, 65, 1137–1147. [Google Scholar] [CrossRef]
- Moltrasio, C.; Romagnuolo, M.; Marzano, A.V. NLRP3 inflammasome and NLRP3-related autoinflammatory diseases: from cryopyrin function to targeted therapies. Front Immunol 2022, 13, 1007705. [Google Scholar] [CrossRef] [PubMed]
- Kolly, L.; Busso, N.; Palmer, G.; Talabot-Ayer, D.; Chobaz, V.; So, A. Expression and function of the NALP3 inflammasome in rheumatoid synovium. Immunology. 2010, 129, 178–185. [Google Scholar] [CrossRef]
- Zhang, Y.; Zheng, Y.; Li, H. NLRP3 inflammasome plays an important role in the pathogenesis of collagen-induced arthritis. Mediators Inflamm. 2016, 2016, 9656270. [Google Scholar] [CrossRef] [PubMed]
- Unterberger, S.; Davies, K.A.; Rambhatla, S.B.; Sacre, S. Contribution of Toll-like receptors and the NLRP3 inflammasome in rheumatoid arthritis pathophysiology. Immuno Targets and Therapy 2021, 10, 285–298. [Google Scholar] [CrossRef]
- Yin, H.; Liu, N.; Sigdel, K.R.; Duan, L. Role of NLRP3 inflammasome in rheumatoid arthritis. Front Immunol. 2022, 13, 931690. [Google Scholar] [CrossRef]
- Murakami, T.; Nakaminami, Y.; Takahata, Y.; Hata, K.; Nishimura, R. Activation and function of NLRP3 inflammasome in bone and joint-related diseases. Int J Mol Sci. 2022, 23, 5365. [Google Scholar] [CrossRef]
- Rosengren, S.; Hoffman, H.M.; Bugbee, W.; Boyle, D.L. Expression and regulation of cryopyrin and related proteins in rheumatoid arthritis synovium. Ann Rheum Dis. 2005, 64, 708–714. [Google Scholar] [CrossRef] [PubMed]
- Guo, C.; Fu, R.; Wang, S.; Huang, Y.; Li, X.; Zhou, M.; Zhao, J.; Yang, N. NLRP3 inflammasome activation contributes to the pathogenesis of rheumatoid arthritis. Clinical and Experimental Immunology. 2018, 194, 231–243. [Google Scholar] [CrossRef]
- Wu, X.Y.; Li, K.T.; Yang, H.X.; Yang, B.; Lu, X.; Zhao, L.D.; et al. Complement C1q synergizes with PTX3 in promoting NLRP3 inflammasome over-activation and pyroptosis in rheumatoid arthritis. J Autoimmun. 2020, 106, 102336. [Google Scholar] [CrossRef]
- Dong, X.; Zhengl, Z.; Linl, P.; Fu, X.; Li, F.; Jiang, J.; Zhu, P. ACPAs promote IL-1β production in rheumatoid arthritis by activating the NLRP3 inflammasome. Cell Mol Immunol. 2020, 17, 261–271. [Google Scholar] [CrossRef]
- Werner, L.E.; Wagner, U. Calcium-sensing receptor-mediated NLRP3 inflammasome activation in rheumatoid arthritis and autoinflammation. Front Physiol. 2023, 13, 1078569. [Google Scholar] [CrossRef] [PubMed]
- Fraenkel, L.; Bathon, J.M.; England, B.R.; St Clair, E.W.; Arayssi, T.; Carandang, K.; Deane, K.D.; Genovese, M.; Huston, K.K.; Kerr, G.; et al. 2021 American College of Rheumatology guideline for the treatment of rheumatoid arthritis. Arthritis Rheumatol. 2021, 73, 1108–1123. [Google Scholar] [CrossRef]
- Tanaka, Y. Recent progress in treatments of rheumatoid arthritis: an overview of developments in biologics and small molecules, and remaining unmet needs. Rheumatology 2021, 60, vi12–vi20. [Google Scholar] [CrossRef] [PubMed]
- Smolen, J.S.; Landewé, R.B.M.; Bergstra, S.A.; Kerschbaumer, A.; Sepriano, A.; Aletaha, D.; Caporali, R.; Edwards, C.J.; Hyrich, K.L.; Pope, J.E.; et al. EULAR recommendations for the management of rheumatoid arthritis with synthetic and biological disease modifying antirheumatic drugs:2022 update. Ann Rheum Dis. 2023, 82, 3–18. [Google Scholar] [CrossRef] [PubMed]
- Nash, P.; Kerschbaumer, A.; Dörner, T.; Dougados, M.; Fleischmann, R.M.; Geissler, K.; McInnes, I.; Pope, J.E.; van der Heijde, D.; Stoffer-Marx, M.; et al. Points to consider for the treatment of immune-mediated inflammatory diseases with Janus kinase inhibitors: a consensus statement. Ann Rheum Dis. 2021, 80, 71–87. [Google Scholar] [CrossRef] [PubMed]
- Kerschbaumer, A.; Sepriano, A.; Bergstra, S.A.; Smolen, J.S.; van der Heijde, D.; Caporali, R.; Edwards, C.J.; Verschueren, P.; de, Souza, S.; Pope, J.E.; et al. Efficacy of synthetic and biological DMARDs: a systemic literature review informing the 2022 update of the EULAR recommendations for the management of rheumatoid arthritis. Ann Rheum Dis 2023, 82, 95–106. [Google Scholar] [CrossRef] [PubMed]
- Mueller, R.B.; Hasler, C.; Popp, F.; Mattow, F.; Durmisi, M.; Souza, A.; Hasler, P.; Rubbert-Roth, A.; Schulze-Koops, H.; Kempis, J.V. Effectiveness, tolerability, and safety of tofacitinib in rheumatoid arthritis: A Retrospective Analysis of Real-World Data from the St. Gallen and Aarau Cohorts. J Clin Med. 2019, 8, 1548. [Google Scholar] [CrossRef]
- Lee, E.B.; Fleischmann, R.; Hall, S.; Wilkinson, B.; Bradley, J.D.; Gruben, D.; Koncz, T.; Krishnaswami, S.; Wallenstein, G.V.; Zang, C.; et al. A. Tofacitinib versus methotrexate in rheumatoid arthritis. N Engl J Med. 2014, 370, 2377–2386. [Google Scholar] [CrossRef]
- Charles-Schoeman, C.; Burmester, G.; Nash, P.; Zerbini, C.A.; Soma, K.; Kwok, K.; Hendrikx, T.; Bananis, E.; Fleischmann, R. Efficacy and safety of tofacitinib following inadequate response to conventional synthetic or biological disease-modifying antirheumatic drugs. Ann Rheum Dis. 2016, 75, 1293–1301. [Google Scholar] [CrossRef] [PubMed]
- Mekni, N.; De Rosa, M.; Cipollina, C.; Gulotta, M.R.; De, Simone, G.; Lombino, J.; Padova, A.; Perricone, U. In silico insights towards the identification of NLRP3 druggable hot spots. Int J Mol Sci. 2019, 20, 4974. [Google Scholar] [CrossRef]
- Gao, J.; Zhang, H.; Yang, Y.; Tao, J. Therapeutic potential of targeting the NLRP3 inflammasome in rheumatod arthritis. Inflammation 2023, 46, 835–840. [Google Scholar] [CrossRef] [PubMed]
- Li, N.; Zhang, R.; Tang, M.; Zhao, M.; Jiang, X.; Cai, X.; Ye, N.; Su, K.; Peng, J.; Zhang, X.; et al. Recent progress and prospects of small molecules for NLRP3 inflammasome inhibition. J Med Chem. 2023, 66, 14447–14473. [Google Scholar] [CrossRef] [PubMed]
- Ishikawa, Y.; Terao, C. The impact of cigarette smoking on risk of rheumatoid arthritis: a narrative review. Cells. 2020, 9, 475. [Google Scholar] [CrossRef] [PubMed]
- Adami, G.; Viapiana, O.; Rossini, M.; Orsolini, G.; Bertoldo, E.; Giollo, A.; Gatti, D.; Fassio, A. Association between environmental air pollution and rheumatoid arthritis flares. Rheumatology (Oxford) 2021, 60, 4591–4597. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Shen, Y.; Jin, K.; Wen, Z.; Cao, W.; Wu, B.; Wen, R.; Tian, L.; Berry, G.J.; Goronzy, J.J.; et al. The DNA Repair Nuclease MRE11A Functions as a Mitochondrial Protector and Prevents T Cell Pyroptosis and Tissue Inflammation. Cell Metab. 2019, 30, 477–492e6. [Google Scholar] [CrossRef] [PubMed]
- Libby, P. Inflammation in Atherosclerosis. Arter. Thromb. Vasc. Biol. 2012, 32, 2045–2051. [Google Scholar] [CrossRef] [PubMed]
- Kremers, H.M.; Crowson, C.S.; Therneau, T.M.; Roger, V.L.; Gabriel, S.E. High ten-year risk of cardiovascular disease in newly diagnosed rheumatoid arthritis patients. Arthritis Rheumatol. 2008, 58, 2268–2274. [Google Scholar] [CrossRef]
- Aviňa-Zubieta, J.A.; Choi, H.K.; Sadatsafavi, M.; Etminan, M.; Esdaile, J.M.; Lacaille, D. Risk of cardiovascular mortality in patients with rheumatoid arthritis: a meta-analysis of observational studies. Arthritis Rheum 2008, 59, 1690–1697. [Google Scholar] [CrossRef] [PubMed]
- Boyer, J.F.; Gourraud, P.A.; Cantagrel, A.; Davignon, J.L.; Constantin, A. Traditional cardiovascular risk factors in rheumatoid arthritis: a meta-analysis. Joint bone spine 2011, 78, 179–183. [Google Scholar] [CrossRef]
- Im, C.H.; Kim, N.R.; Kang, J.W.; Kim, J.H.; Kang, J.Y.; Bae, G.B.; Nam, E.J.; Kang, Y.M. Inflammatory burden interacts with conventional cardiovascular risk factors for carotid plaque formation in rheumatoid arthritis. Rheumatology (Oxford) 2015, 54, 808–815. [Google Scholar] [CrossRef]
- Chester, Wasko, M.; Dasgupta, A.; Ilse, Sears, G.; Fries, J.F.; Ward, M.M. Prednisone use and risk of mortality in patients with rheumatoid arthritis: moderation by use of disease-modifying antirheumatic drugs. Arthritis Care Res. 2016, 68, 706–710. [CrossRef] [PubMed]
- Danelich, I.M.; Wright, S.S.; Lose, J.M.; Tefft, B.J.; Cicci, J.D.; Reed, B,N. Safety of nonsteroidal anti-inflammatory drugs in patients with cardiovascular disease. Pharmacotherapy 2015, 35, 520–535. [CrossRef] [PubMed]
- Maehlen, M.T.; Provan, S.A.; de Rooy, D.P.; van der Helm-van, A.H.; Krabben, A.; Saxne, T.; Lindqvist, E.; Semb, A.G.; Uhlig, T.; van, der, Heijde, D.; et al. Associations between APOE Genotypes and Disease Susceptibility, Joint Damage and Lipid Levels in Patients with Rheumatoid Arthritis. PLoS ONE 2013, 8, e60970.
- Myasoedova, E.; Crowson, C.S.; Kremers, H.M.; Roger, V.L.; Fitz-Gibbon, P.D.; Therneau, T.M.; Gabriel, S.E. Lipid paradox in rheumatoid arthritis: the impact of serum lipid measures and systemic inflammation on the risk of cardiovascular disease. Ann. Rheum. Dis. 2011, 70, 482–487. [Google Scholar] [CrossRef] [PubMed]
- Chang, C.T.; Shen, M.Y.; Lee, A.S.; Wang, C.C.; Chen, W.Y.; Chang, C.M.; Chang, K.C.; Stancel, N.; Chen, C.H. Electronegative low-density lipoprotein increases the risk of ischemic lower-extremity peripheral artery disease in uremia patients on maintenance hemodialysis. Sci. Rep. 2017, 7, 4654. [Google Scholar] [CrossRef] [PubMed]
- Chang, C.Y.; Chen, C.H.; Chen, Y.M.; Hsieh, T.Y.; Li, J.P.; Shen, M.Y.; Lan, J.L.; Chen, D.Y. Association between Negatively Charged Low-Density Lipoprotein L5 and Subclinical Atherosclerosis in Rheumatoid Arthritis Patients. J. Clin. Med. 2019, 8, 177. [Google Scholar] [CrossRef] [PubMed]
- Tang, D.; Lu, J.; Walterscheid, J.P.; Chen, H.H.; Engler, D.A.; Sawamura, T.; Chang, P.Y.; Safi, H.J.; Yang, C.Y.; Chen, C.H. Electronegative LDL circulating in smokers impairs endothelial progenitor cell differentiation by inhibiting Akt phosphorylation via LOX-1. J. Lipid Res. 2008, 49, 33–47. [Google Scholar] [CrossRef] [PubMed]
- Chang, C.K.; Chen, P.K.; Lan, J.L.; Chang, S.H.; Hsieh, T.Y.; Liao, P.J.; Chen, C.H.; Chen, D.Y. Association of Electronegative LDL with Macrophage Foam Cell Formation and CD11c Expression in Rheumatoid Arthritis Patients. Int. J. Mol. Sci. 2020, 21, 5883. [Google Scholar] [CrossRef] [PubMed]
- Van, Staa, T.P.; Geusens, P.; Bijlsma, J.W.; Leufkens, H.G.; Cooper, C. Clinical assessment of the long-term risk of fracture in patients with rheumatoid arthritis. Arthritis Rheum. 2006, 54, 3104–3112. [CrossRef]
- Yu, X.H.; Yang, Y.Q.; Cao, R.R.; Cai, M.K.; Zhang, L.; Deng, F.Y.; Lei, S.F. Rheumatoid arthritis and osteoporosis: shared genetic effect, pleiotropy and causality. Human Molecular Genetics. 2021, 30, 1932–1940. [Google Scholar] [CrossRef]
- Hecht, C.; Englbrecht, M.; Rech, J.; Schmidt, S.; Araujo, E.; Engelke, K.; Finzel, S.; Schett, G. Additive Effect of Anti-Citrullinated Protein Antibodies and Rheumatoid Factor on Bone Erosions in Patients with RA. Ann. Rheum. Dis. 2015, 74, 2151. [Google Scholar] [CrossRef] [PubMed]
- Baker, R.; Narla, R.; Baker, J.F.; Wysham, K.D. Risk factors for osteoporosis and fractures in rheumatoid arthritis. Best Pract Res Clin Rheumatol. 2022, 36, 101773. [Google Scholar] [CrossRef] [PubMed]
- Kadura, S.; Raghu, G. Rheumatoid arthritis-interstitial lung disease: manifestations and current concepts in pathogenesis and management. Eur. Respir. Rev. 2021, 30, 210011. [Google Scholar] [CrossRef] [PubMed]
- Castelino, F.V.; Varga, J. Interstitial lung disease in connective tissue diseases: evolving concepts of pathogenesis and management. Arthritis Res Thera. 2010, 12, 213. [Google Scholar] [CrossRef] [PubMed]
- Tsuchiya, Y.; Takayanagi, N.; Sugiura, H.; Miyahara, Y.; Tokunaga, D.; Kawabata, Y.; Sugita, Y. Lung diseases directly associated with rheumatoid arthritis and their relationship to outcome. Eur Respir J. 2011, 37, 1411–1417. [Google Scholar] [CrossRef] [PubMed]
- Paulin, F.; Doyle, T.J.; Fletcher, E.A.; Ascherman, D.P.; Rosas, I.O. Rheumatoid arthritis-associated interstitial lung disease and idiopathic pulmonary fibrosis: shared mechanistic and phenotypic traits suggest overlapping disease mechanisms. Rev Investing Clin. 2015, 67, 280–286. [Google Scholar]
- Johnson, C. Recent advances in the pathogenesis, prediction, and management of rheumatoid arthritis-associated interstitial lung disease. Curr Opin Rheumatol. 2017, 29, 254–259. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; Zhang, J.; Lau, J.; Wang, S.; Taneja, V.; Matteson, E.L.; Vassallo, R. Mechanisms of lung disease development in rheumatoid arthritis. Nat Rev Rheumatol. 2019, 15, 581–596. [Google Scholar] [CrossRef] [PubMed]
- Paulin, F.; Doyle, T.J.; Fletcher, E.A.; Ascherman, D.P.; Rosas, I.O. Rheumatoid arthritis-associated interstitial lung disease and idiopathic pulmonary fibrosis: shared mechanistic and phenotypic traits suggest overlapping disease mechanisms. Rev Invest Clin. 2015, 67, 280–286. [Google Scholar]
- Oka, S.; Furukawa, H.; Shimada, K.; Sugii, S.; Hashimoto, A.; Komiya, A.; Fukui, N.; Suda, A.; Tsunoda, S.; Ito, S.; et al. Association of Human Leukocyte Antigen Alleles with Chronic Lung Diseases in Rheumatoid Arthritis. Rheumatology 2016, 55, 1301–1307. [Google Scholar] [CrossRef]
- Juge, P.A.; Borie, R.; Kannengiesser, C.; Gazal, S.; Revy, P.; Wemeau-Stervinou, L.; Debray, M.P.; Ottaviani, S.; Marchand-Adam, S.; Nathan, N.; et al. AB0007 Shared Genetic Predisposition in Rheumatoid Arthritis–Interstitial Lung Disease and Familial Pulmonary Fibrosis. Ann. Rheum. Dis. 2017, 76, 1049. [Google Scholar]
- Joo, Y.B.; Ahn, S.M.; Bang, S.Y.; Park, Y.; Hong, S.J.; Lee, Y.; Cho, S.K.; Choi, C.B.; Sung, Y.K.; Kim, T.H.; et al. MUC5B promoter variant rs35705950, rare but significant susceptibility locus in rheumatoid arthritis-interstitial lung disease with usual interstitial pneumonia in Asian populations. RMD Open 2022, 8, e002790. [Google Scholar] [CrossRef] [PubMed]
- Shirai, Y.; Honda, S.; Ikari, K.; Kanai, M.; Takeda, Y.; Kamatani, Y.; Morisaki, T.; Tanaka, E.; Kumanogoh, A.; Harigai, M.; et al. Association of the RPA3-UMAD1 locus with interstitial lung diseases complicated with rheumatoid arthritis in Japanese. Ann Rheum Dis. 2020, 79, 1305–1309. [Google Scholar] [CrossRef]
- Khandpur, R.; Carmona-Rivera, C.; Vivekanandan-Giri, A.; Gizinski, A.; Yalavarthi, S.; Knight, J.S.; Friday, S.; Li, S.; Patel, R.M.; Subramanian, V.; et al. NETs Are a Source of Citrullinated Autoantigens and Stimulate Inflammatory Responses in Rheumatoid Arthritis. Sci. Transl. Med. 2013, 5, 178ra40. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Wang, D.; Wang, L.; Wang, S.; Roden, A.C.; Zhao, H.; Li, X.; Prakash, Y.S.; Matteson, E.L.; Tschumperlin, D.J.; et al. Profibrotic effect of IL-17A and elevated IL-17RA in idiopathic pulmonary fibrosis and rheumatoid arthritis-associated lung disease support a direct role for IL-17A/IL-17RA in human fibrotic interstitial lung disease. Am J Physiol Lung Cell Mol Physiol. 2019, 316, L487–L497. [Google Scholar] [CrossRef] [PubMed]
- Zhao, N.; Al-Aly, Z.; Zheng, B.; van, Donkelaar, A.; Martin, R.V.; Pineau, C.A.; Bernatsky, S. Fine particles matter components and interstitial lung disease in rheumatoid arthritis. Eur Respir J. 2022, 60, 2102149. [CrossRef]
- Dolasia, K.; Bisht, M.K.; Pradhan, G.; Udgata, A.; Mukhopadhyay, S. TLRs/NLRs: shaping the landscape of host immunity. Int Rev Immunol. 2018, 37, 3–19. [Google Scholar] [CrossRef]
- Jung, J.Y.; Suh, C.H.; Kim, H.A. The role of damage-associated molecular pattern for pathogenesis and biomarkers in adult-onset Still’s disease. Expert Rev Mol Diagn. 2019, 19, 459–468. [Google Scholar] [CrossRef] [PubMed]
- Heilig, R.; Dick, M.S.; Sborgi, L.; Meunier, E.; Hiller, S.; Broz, P. The gasdermin-D pore acts a conduit for IL-1β secretion in mice. Eur J Immunol. 2018, 48, 584–592. [Google Scholar] [CrossRef]
- Bergsbaken, T.; Fink, S.L, Cookson BT. Pyroptosis: host cell death and inflammation. Nat Rev Microbiol. 2009, 7, 99–109. [CrossRef]
- Sollberger, G.; Strittmatter, G.E.; Garstkiewicz, M.; Sand, J.; Beer, H.D. Caspase-1: the inflammasome and beyond. Innate Immun. 2014, 20, 115–125. [Google Scholar] [CrossRef] [PubMed]
- Vande, Walle, L.; Lamkanfi, M. Pyroptosis. Curr Biol. 2016, 26, R568–R72.
- Li, Z.; Guo, J.; Bi, L. Role of the NLRP3 inflammasome in autoimmune diseases. Biomedicine Pharmacotherapy 2020, 130, 110542. [CrossRef]
- Wu, X.; Ren, G.; Zhou, R.; Ge, J.; Chen, F.H. The role of Ca (2+) in acid-sensing ion channel 1a-mediated chondrocyte pyroptosis in rat adjuvant arthritis. Lab Invest. 2019, 99, 499–513. [Google Scholar] [CrossRef] [PubMed]
- Lee, G.S.; Subramanian, N.; Kim, A.I.; Aksentijevich, I.; Goldbach-Mansky, R.; Sacks, D.B.; Germain, R.N.; Kastner, D.L.; Chae, J.J. The calcium-sensing receptor regulates the NLRP3 inflammasome through Ca2+ and cAMP. Nature 2012, 492, 123–127. [Google Scholar] [CrossRef] [PubMed]
- Werner, L.E.; Wagner, U. Calcium-sensing receptor-mediated NLRP3 inflammasome activation in rheumatoid arthritis and autoinflammation. Front Physiol. 2023, 13, 1078569. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Q.; Wang, X.; Xu, X.; Hu, L.; Zhou, G.; Liu, R.; Yang, G.; Cui, D. Inflammasomes in rheumatoid arthritis: a pilot study. BMC Rheumatol. 2023, 7, 39. [Google Scholar] [CrossRef] [PubMed]
- Kastbom, A.; Verma, D.; Eriksson, P.; Skogh, T.; Wingren, G.; Söderkvist, P. Genetic variations in proteins of the cryopyrine inflammasome influences the susceptibility and severity of rheumatoid arthritis (the Swedish TIRA project). Rheumatology (Oxford) 2008, 47, 415–417. [Google Scholar] [CrossRef] [PubMed]
- Jenko, B.; Praprotnik, S.; Tomsic, M.; Dolzan, V. NLRP3 and CARD8 polymorphisms influence higher disease activity in rheumatoid arthritis. J Med Biochem. 2016, 35, 319–323. [Google Scholar] [CrossRef]
- Yang, Z.; Cao, J.; Yang, Q.; Zhang, Y.; Han, L. NLRP3 p. Q705K and CARD8 p.C10X single nucleotide polymorphisms are not associated with susceptibility to rheumatoid arthritis: a meta-analysis. Int J Rheum Dis. 2017, 20, 1481–1491. [CrossRef]
- Li, R.N.; Ou, T.T.; Lin, C.H.; Lin, Y.Z.; Fang, T.J.; Chen, Y.J.; Tseng, C.C.; Sung, W.Y.; Wu, C.C.; Yen, J.H. NLRP3 gene polymorphisms in rheumatoid arthritis and primary Sjögren’s syndrome patients. Diagnostics (Basel). 2023, 13, 206. [Google Scholar] [CrossRef] [PubMed]
- Sode, J.; Vogel, U.; Bank, S.; Andersen, P.S.; Thomsen, M.K.; Hetland, M.L.; Locht, H.; Heegaard, N.H.; Andersen, V. Anti-TNF treatment response in rheumatoid arthritis patients is associated with genetic variation in the NLRP3-inflammasome. PLOS ONE. 2014, 9, e10036. [Google Scholar] [CrossRef] [PubMed]
- Mathews, R.J.; Robinson, J.I.; Battellino, M.; Wong, C.; Taylor, J.C; Biologics in Rheumatoid Arthritis Genetics and Genomics Study Syndicate (BRAGGSS).; Eyre, S.; Churchman, S.M.; Wilson, A.G.; Isaacs, J.D, et al. Evidence of NLRP3-inflammasome activation in rheumatoid arthritis (RA); genetic variants within the NLRP3-inflammasome complex in relation to susceptibility to RA and response to anti-TNF treatment. Ann Rheum Dis. 2014, 73, 1202–1210.
- Awni, A.A.; Hamed, Z.O.; Abdul-Hassan, Abbas, A.; Abdulamir, A.S. Effect of NLRP3 inflammasome genes polymorphisms on disease susceptibility and response to TNF-α inhibitors in Iraqi patients with rheumatoid arthritis. Heliyon 2023, 9, e16814. [CrossRef] [PubMed]
- Karasawa, T.; Takahashi, M. Role of NLRP3 inflammasome in atherosclerosis. J Atheroscler Thromb. 2017, 24, 443–451. [Google Scholar] [CrossRef] [PubMed]
- Baragetti, A.; Catapano, A.L.; Magni, P. Multifactorial activation of NLRP3 inflammasome: relevance for a precision approach to atherosclerotic cardiovascular risk and disease. Int. J. Mol. Sci. 2020, 21, 4459. [Google Scholar] [CrossRef] [PubMed]
- Duewell, P.; Kono, H.; Rayner, K.J.; Sirois, C.M.; Vladimer, G.; Bauernfeind, F.G.; Abela, G.S.; Franchi, L.; Nuñez, G.; Schnurr, M.; et al. NLRP3 inflammasomes are required for atherogenesis and activated by cholesterol crystals. Nature 2010, 464, 1357–1361. [Google Scholar] [CrossRef] [PubMed]
- Rhoads, J.P.; Lukens, J.R.; Wilhelm, A.J.; Moore, J.L.; Mendez-Fernandez, Y.; Kanneganti, T.D.; Major, A.S. Oxidized Low-Density Lipoprotein Immune Complex Priming of the Nlrp3 Inflammasome Involves TLR and FcγR Cooperation and Is Dependent on CARD9. J Immunol. 2017, 198, 2105–2114. [Google Scholar] [CrossRef] [PubMed]
- Varghese, G.P.; Fransén, K.; Hurtig-Wennlöf, A.; Bengtsson, T.; Jansson, J.H.; Sirsjö, A. Q705K variant in NLRP3 gene confers protection against myocardial infarction in female individuals. Biomed. Rep. 2013, 1, 879–882. [Google Scholar] [CrossRef]
- Paramel, G.V.; Folkersen, L.; Strawbridge, R.J.; Elmabsout, A.A.; Särndahl, E.; Lundman, P.; Jansson, J.H.; Hansson, G.K.; Sirsjö, A.; Fransén, K. CARD8 gene encoding a protein of innate immunity is expressed in human atherosclerosis and associated with markers of inflammation. Clin Sci. 2013, 125, 401–407. [Google Scholar] [CrossRef]
- Bai, Y.; Nie, S.; Jiang, G.; Zhou, Y.; Zhou, M.; Zhao, Y.; Li, S.; Wang, F.; Lv, Q.; Huang, Y.; et al. Regulation of CARD8 expression by ANRIL and association of CARD8 single nucleotide polymorphism rs2043211 (p.C10X) with ischemic stroke. Stroke 2014, 45, 383–388. [Google Scholar] [CrossRef]
- García-Bermúdez, M.; López-Mejías, R.; González-Juanatey, C.; Corrales, A.; Castañeda, S.; Ortiz, A.M.; Miranda-Filloy, J.A.; Gómez-Vaquero, C.; Fernández-Gutiérrez, B.; Balsa, A.; et al. CARD8 rs2043211 (p.C10X) polymorphism is not associated with disease susceptibility or cardiovascular events in Spanish rheumatoid arthritis patients. DNA Cell Biol. 2013, 32, 28–33. [Google Scholar] [CrossRef]
- Murakami, T.; Nakaminami, Y.; Takahata, Y.; Hata, K.; Nishimura, R. Activation and function of NLRP3 inflammasome in bone and joint-related diseases. Int J Mol Sci. 2022, 23, 5365. [Google Scholar] [CrossRef] [PubMed]
- Yu, C.; Zhang, C.; Kuang, Z.; Zheng, Q. The Role of NLRP3 Inflammasome Activities in Bone Diseases and Vascular Calcification. Inflammation 2021, 44, 434–449. [Google Scholar] [CrossRef] [PubMed]
- Xu, L.; Shen, L.; Yu, X.; Li, P.; Wang, Q.; Li, C. Effects of irisin on osteoblast apoptosis and osteoporosis in postmenopausal osteoporosis rats through upregulating Nrf2 and inhibiting NLRP3 inflammasome. Exp Ther Med. 2020, 19, 1084–1090. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Wang, H.; Feng, C.; Li, C.; Li, Z.; He, J.; Tu, C. The regulatory role and therapeutic application of pyroptosis in musculoskeletal diseases. Cell Death Discovery 2022, 8, 492. [Google Scholar] [CrossRef]
- Lei, L.; Sun, J.; Han, J.; Jiang, X.; Wang, Z.; Chen, L. Interleukin-17 induces pyroptosis in osteoblasts through the NLRP3 inflammasome pathway in vitro. Int Immunopharmacol. 2021, 96, 107781. [Google Scholar] [CrossRef]
- Polzer, K.; Joosten, L.; Gasser, J.; Distler, J.H.; Ruiz, G.; Baum, W.; Redlich, K.; Bobacz, K.; Smolen, J.S.; van, den, Berg, W.; et al. Interleukin-1 is essential for systemic inflammatory bone loss. Ann Rheum Dis. 2010, 69, 284–290.
- He, Z.; Sun, Y.; Wu, J.; Xiong, Z.; Zhang, S.; Liu, J.; Liu, Y.; Li, H.; Jin, T.; Yang, Y.; et al. Evaluation of genetic variants in IL-1B and its interaction with the predisposition of osteoporosis in the northwestern Chinese Han population. J Gene Med. 2020, 22, e3214. [Google Scholar] [CrossRef]
- Zhang, W.; Cong, X.L.; Qin, Y.H.; He, Z.W.; He, D.Y.; Dai, S.M. IL-18 upregulates the production of key regulators of osteoclastogenesis from fibroblast-like synoviocytes in rheumatoid arthritis. Inflammation 2013, 36, 103–109. [Google Scholar] [CrossRef]
- Lasithiotaki, I.; Giannarakis, I.; Tsitoura, E.; Samara, K.D.; Margaritopoulos, G.A.; Choulaki, C.; Vasarmidi, E.; Tzanakis, N.; Voloudaki, A.; Sidiropoulos, P.; et al. NLRP3 inflammasome expression in idiopathic pulmonary fibrosis and rheumatoid lung. Eur Respir J. 2016, 47, 910–918. [Google Scholar] [CrossRef] [PubMed]
- Artlett, C. M., Sassi-Gaha, S., Rieger, J. L., Boesteanu, A. C., Feghali-Bostwick, C. A., and Katsikis, P. D. The inflammasome activating caspase 1 mediates fibrosis and myofibroblast differentiation in systemic sclerosis. Arthritis Rheum. 2011, 63, 3563–3574. [CrossRef] [PubMed]
- Woo, S.; Gandhi, S.; Ghincea, A.; Saber, T; Lee, C.J.; Ryu, C. Targeting the NLRP3 inflammasome and associated cytokines in scleroderma associated interstitial lung disease. Front Cell Dev Biol. 2023, 11, 1254904.
- Ramos-Martinez, E.; Vega-Sánchez, A.E.; Pérez-Rubio, G.; Mejia, M.; Buendía-Roldán, I.; González-Pérez, M.I.; Mateos-Toledo H,N.; Andrade, W.A.; Falfán-Valencia, R.; Rojas-Serrano, J. Enhanced activity of the NLRP3 inflammasome in the lung of patients with anti-synthetase syndrome. Cells 2023, 12, 60.
- Mangan, M.S.J.; Olhava, E.J.; Roush, W.R.; Seidel, H.M.; Glick, G.D.; Latz, E. Targeting the NLRP3 inflammasome in inflammatory diseases. Nature Reviews Drug Discovery 2018, 17, 588–606. [Google Scholar] [CrossRef] [PubMed]
- Coll, R.C.; Hill, J.R.; Day, C.J.; Zamoshnikova, A.; Boucher, D.; Massey, N.L.; Chitty, J.L.; Fraser, J.A.; Jennings, M.P.; Robertson, A.A.B.; et al. MCC950 directly targets the NLRP3 ATP- hydrolysis motif for inflammasome inhibition. Nature Chemical Biology 2019, 15, 556–559. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Zheng, Y. Effects and mechanisms of potent caspase-1 inhibitor VX765 treatment on collagen induced arthritis in mice. Clinical and Experimental Rheumatology 2016, 34, 111–118. [Google Scholar] [PubMed]
- Hu, J.J.; Liu, X.; Xia, S.; Zhang, Z.; Zhang, Y.; Zhao, J.; Ruan, J.; Luo, X.; Lou, X.; Bai, Y.; et al. FDA-approved disulfiram inhibits pyroptosis by blocking gasdermin D pore formation. Nature Immunology 2020, 21, 736–745. [Google Scholar] [CrossRef]
- You, R.; He, X.; Zeng, Z.; Zhan, Y.; Xiao, Y.; Xiao, R. Pyroptosis and its role in autoimmune diseases: a potential therapeutic target. Front Immunol. 2022, 13, 841732. [Google Scholar] [CrossRef]
- van, der, Heijden, T.; Kritikou, E.; Venema, W.; van, Duijn, J.; van, Santbrink, P.J.; Slütter, B.; Foks, A.C.; Bot, I.; Kuiper, J. NLRP3 inflammasome inhibition by MCC950 reduces atherosclerotic lesion development in apolipoprotein E-deficient mice-brief report. Arterioscler. Thromb. Vasc. Biol. 2017, 37, 1457–1461.
- Zeng, W.; Wu, D.; Sun, Y.; Suo, Y.; Yu, Q.; Zeng, M.; Gao, Q.; Yu, B.; Jiang, X.; Wang, Y. The selective NLRP3 inhibitor MCC950 hinders atherosclerosis development by attenuating inflammation and pyroptosis in macrophages. Sci. Rep. 2021, 11, 19305. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Niu, X.; Xu, H.; Li, Q.; Meng, L.; He, M.; Zhang, J.; Zhang, Z.; Zhang, Z. VX-765 attenuates atherosclerosis in ApoE deficient mice by modulating VSMCs pyroptosis. Exp. Cell Res. 2020, 389, 111847. [Google Scholar] [CrossRef]
- Mansoori, M.N.; Shukla, P.; Kakaji, M.; Tyagi, A.M.; Srivastava, K.; Shukla, M.; Dixit, M.; Kureel, J.; Gupta, S.; Singh, D. IL-18BP is decreased in osteoporotic women: prevents inflammasome mediated IL-18 activation and reduces Th17 differentiation. Sci Rep. 2016, 6, 33680. [Google Scholar] [CrossRef]
- Huang, C.L.; Chen, D.Y.; Tzang, C.C.; Lin, J.W.; Tzang, B.S.; Hsu, T.C. Celastrol attenuates human parvovirus B19 NS1-induced NLRP3 inflammasome activation in macrophages. Mol Med Rep. 2023, 28, 193. [Google Scholar] [CrossRef] [PubMed]
- Jing, M.; Yang, J.; Zhang, L.; Liu, J.; Xu, S.; Wang, M.; Zhang, L.; Sun, Y.; Yan, W.; Hou, G.; et al. Celastrol inhibits rheumatoid arthritis through the ROS-NF-κB-NLRP3 inflammasome axis. Int Immunopharmacol. 2021, 98, 107879. [Google Scholar] [CrossRef]
- Wu, D. Efficacy of Baihu-Guizhi Decoction Combined With Western Medicine in Treating Rheumatoid Arthritis. Chin J Urban Rural Enterp Hyg. 2021, 36, 158–160. [Google Scholar]
- Li, W.; Wang, K.; Liu, Y.; Wu, H.; He, Y.; Li, C.; Wang, Q.; Su, X.; Yan, S.; Su, W.; et al. A novel drug combination of mangiferin and cinnamic acid alleviates rheumatoid arthritis by inhibiting TLR4/NF-κB/NLRP3 activation-induced pyroptosis. Front Immunol. 2022, 13, 912933. [Google Scholar] [CrossRef]
- Ye, Q.; Yan, T.; Shen, J.; Shi, X.; Luo, F.; Ren, Y. Sulforaphene targets NLRP3 inflammasome to suppress M1 polarization of macrophages and inflammatory response in rheumatoid arthritis. J Biochem Mol Toxicol. 2023, 37, e23362. [Google Scholar] [CrossRef] [PubMed]
- Xu, R.; Liu, Z.; Hou, J.; Huang, T.; Yang, M. Osthole improves colllagen-induced arthritis in a rat model through inhibiting inflammation and cellular stress. Cell Mol Biol Lett. 2018, 23, 19. [Google Scholar] [CrossRef]
- Jiang, X.; Lu, Z.; Zhang, Q.; Yu, J.; Han, D.; Liu, J.; Li, P.; Li, F. Osthole: a potential AMPK agonist that inhibits NLRP3 inflammasome by regulating mitochondrial homeostasis for combating rheumatoid arthritis. Phytomemdicine 2023, 110, 154640. [Google Scholar] [CrossRef]
- Zhu, T.; Zhang, L.; Ling, S.; Duan, J.; Qian, F.; Li, Y.; Xu, J.W. Scropolioside B inhibits IL-1β and cytokines expression through NF-kB and inflammasome NLRP3 pathways. Mediators Inflammation 2014, 2014, 819053. [Google Scholar] [CrossRef] [PubMed]
- Cao, J.; Ning, Y.; Ningm, X.; Zhang, H. Wedelolactone ameliorate synovial inflammation and cardiac complications in a murine model of collagen-induced arthritis by inhibiting NF-κB/NLRP3 inflammasome activation. Folia Histochemica et Cytobiologica 2022, 60, 301–310. [Google Scholar] [CrossRef] [PubMed]
- Eugenia, Schroeder, M.; Russo, S.; Costa, C.; Hori, J.; Tiscornia, I.; Bollati-Fogolín, M, Zamboni DS, Ferreira G, Cairoli E, Hill M. Pro-inflammatory Ca++-activated K+ channels are inhibited by hydroxychloroquine. Sci Rep. 2017, 7, 1892.
- Cui, J.; Hong, P.; Li, Z.; Lin, J.; Wu, X.; Nie, K.; Zhang, X.; Wan, J. Chloroquine inhibits NLRP3 inflammasomes activation and alleviates renal fibrosis in mouse model of hyperuricemic nephropathy with aggravation by a high-fat-diet. Int Immunopharmacol. 2023, 120, 110353. [Google Scholar] [CrossRef] [PubMed]
- Mertens, M.; Singh, J.A. Anakinra for rheumatoid arthritis. Cochrane Database Systematic Review. 2009, 21, CD005121. [Google Scholar] [CrossRef] [PubMed]
- Ridker, P.M.; Everett, B.M.; Thuren, T.; MacFadyen, J.G.; Chang, W.H.; Ballantyne, C.; Fonseca, F.; Nicolau, J.; Koenig, W.; Anker, S.D.; et al. Antiinflammatory therapy with canakinumab for atherosclerotic disease. N. Engl. J. Med. 2017, 377, 1119–1131. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Zhan, N.; Jin, Y.; Ling, H.; Xiao, C.; Xie, Z.; Zhong, H.; Yu, X.; Tang, R.; Ma, J.; et al. Tofacitinib restores the balance of γδTreg/γδT17 cells in rheumatoid arthritis by inhibiting the NLRP3 inflammasome. Theranostics 2021, 11, 1445–1457. [Google Scholar] [CrossRef] [PubMed]
- Tezcan, G.; Martynova, E.V.; Gilazieva, Z.E.; McIntyre, A.; Rizvanov, A.A.; Khaiboullina, S.F. MicroRNA post-transcriptional regulation of the NLRP3 inflammasome in immunopathologies. Front Pharm. 2019, 10, 451. [Google Scholar] [CrossRef] [PubMed]
- Xue, Z.; Xi, Q.; Liu, H.; Guo, X.; Zhang, J.; Zhang, Z.; Li, Y.; Yang, G.; Zhou, D.; Yang, H.; et al. miR-21 promotes NLRP3 inflammasome activation to mediate pyroptosis and endotoxic shock. Cell Death Dis. 2019, 10, 461. [Google Scholar] [CrossRef]
- Krol, J.; Loedige, I.; Filipowicz, W. The widespread regulation of microRNA biogenesis, function, and decay. Nat Rev Genet. 2010, 11, 597–610. [Google Scholar] [CrossRef]
- Wei, B.; Pei, G. MicroRNAs: critical regulators in Th17 cells and players in diseases. Cell Mol Immunol. 2010, 7, 175–181. [Google Scholar] [CrossRef]
- Xie, Q.; Wei, M.; Zhang, B.; Kang, X.; Liu, D.; Zheng, W.; Pan, X.; Quan, Y.; Liao, D.; Shen, J. MicroRNA-33 regulates the NLRP3 inflammasome signaling pathway in macrophages. Mol Med Rep 2018, 17, 3318–3327. [Google Scholar] [CrossRef]
- Yang, Q.; Zhao, W.; Chen, Y.; Chen, Y.; Shi, J.; Qin, R.; Wang, H.; Wang, R.; Yuan, H.; Sun, W. RelA/MicroRNA-30a/NLRP3 signal axis is involved in rheumatoid arthritis via regulating NLRP3 inflammsome in macrophages. Cell Death and Disease 2021, 12, 1060. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.; Lu, D.; Ma, W.; Liu, J.; Ning, Q.; Tang, F.; Li, L. miR-223 in exosomes from bone marrow mesenchymal stem cells ameliorates rheumatoid arthritis via downregulation of NLRP3 expression in macrophages. Molecular Immunology 2022, 143, 68–76. [Google Scholar] [CrossRef] [PubMed]
- Liao, T.L.; Chen, Y.M.; Tang, K.T.; Chen, P.K.; Liu, H.J.; Chen, D.Y. MicroRNA-223 inhibits IL-18-mediated neutrophil extracellular trap through regulating calcium influx in adult-onset Still’s disease. Sci Rep. 2021, 11, 15676. [Google Scholar] [CrossRef]
- Guttman, M.; Amit, I.; Garber, M.; French, C.; Lin, M.F.; Feldser, D.; Huarte, M.; Zuk, O.; Carey, B.W.; Cassady, J.P.; et al. Chromatin signature reveals over a thousand highly conserved large non-coding RNAs in mammals. Nature. 2009, 458, 223–227. [Google Scholar] [CrossRef]
- Geisler, S.; Coller, J. RNA in unexpected places: long non-coding RNA functions in diverse cellular contexts. Nat Rev Mol Cell Biol. 2013, 14, 699–712. [Google Scholar] [CrossRef] [PubMed]
- Mathy, N.W.; Chen, X.M. Long non-coding RNAs (lncRNAs) and their transcriptional control of inflammatory responses. J Biol Chem. 2017, 292, 12375–12382. [Google Scholar] [CrossRef]
- Wang, Z.; Kun, Y.; Lei, Z.; Dawei, W.; Lin, P.; Jibo, W. LncRNA MIAT downregulates IL-1β, TNF-α to suppress macrophage inflammation but is suppressed by ATP-induced NLRP3 inflammasome activation. Cell Cycle 2021, 20, 194–203. [Google Scholar] [CrossRef]
- Sarsenova, M.; Issabekova, A.; Abisheva, S.; Rutskaya-Moroshan, K.; Ogay, V.; Saparov, A. Mesenchymmal stem cell-based therapy for rheumatoid arthritis. Int J Mol Sci. 2021, 22, 11592. [Google Scholar] [CrossRef]
- Shin, T.H.; Kim, H,S.; Kang, T.W.; Lee, B.C.; Lee, H.Y.; Kim, Y.J.; Shin, J.H.; Seo, Y.; Won Choi S.; Lee, S.; et al. Human umbilical cord blood-stem cells direct macrophage polarization and block inflammasome activation to alleviate rheuamatoid arthritis. Cell Death and Disease 2016, 7, e2524. [CrossRef] [PubMed]
Figure 1.
The flow diagram of the literature selection process [Search conducted on 31 October 2023]. Duplicates and manuscripts with incomplete data have been excluded. RA: rheumatoid arthritis; NLRP3: nucleotide-binding domain leucine-rich repeat-containing receptors (NLRs)-containing a pyrin domain-3; ASCVD: atherosclerotic cardiovascular disease; OP: osteoporosis; ILD: interstitial lung disease.
Figure 1.
The flow diagram of the literature selection process [Search conducted on 31 October 2023]. Duplicates and manuscripts with incomplete data have been excluded. RA: rheumatoid arthritis; NLRP3: nucleotide-binding domain leucine-rich repeat-containing receptors (NLRs)-containing a pyrin domain-3; ASCVD: atherosclerotic cardiovascular disease; OP: osteoporosis; ILD: interstitial lung disease.
Figure 2.
The proposed model for the pathogenic role of NLRP3 inflammasome signaling pathway in RA and its clinical implications as the therapeutic potential. Several ligands that bind to TNFRs and TLRs can activate nuclear factor (NF)-κB. As a transcription factor, activated NF-κB can translocate into the nucleus and thereby activate the expression of NLRP3 and pro-IL-1β. As an endogenous ligand, anti-citrullinated peptide antibodies (ACPA), can promote NF-κB phosphorylation through binding to TNFRs and TLRs. In the second signal, extracellular ATP can bind to P2X7 and thereby lead to K+ efflux and extracellular Ca++ influx, which activate the NLRP3 inflammasome with overproduction of mature form of IL-1β and IL-18. The NLRP3 inflammasome activation also leads to the cleavage of gasdermin D, which promotes pyroptosis with the formation of pores in the cell membrane and the release of IL-1β and IL-18. ACPA can also activate the pannexin channel, resulting in ATP secretion and NLRP3 inflammation activation. Besides, In RA monocytes, complement C1q and pentaxin 3 (PTX3) synergistically activate the NLRP3 inflammasome and pyroptosis. Several compounds have been identified as inhibitors of the components of the NLRP3 inflammasome signaling pathway. MCC950, VX-765, osthole, and sulforaphane can inhibit the activation of the NLRP3 inflammasome. Tofacitinib, one of JAKi, may restore the balance of γδTreg/γδT17 cells in RA by inhibiting the NLRP3 inflammasome. Disulfiram inhibits GSDMD and thereby blocks pyroptosis and the release of IL-1β and IL-18. Anakinra, an IL-1β receptor antagonist, blocks the effects of NLRP3 inflammasome downstream cytokine. Among natural products, both Baihu-Guizhi decoction (BHGZD) and celastrol can inhibit NLRP3 activation by blocking the NF-κB pathway. HCQ could inhibit Ca2+-activated K+ channels and then impair inflammasome activation. The microRNA-30a and miR-223 inhibit the expression of NLRP3 and inflammasome activation. RA: rheumatoid arthritis; TNFα: tumor necrosis factor-α; TNFRs: TNFα receptors; TLRs: Toll-like receptors; ATP: adenosine triphosphate; PTX3: pentaxin 3; GSDMD: gasdermin D; IL: interleukin; ASCVD: atherosclerotic cardiovascular disease; OP: osteoporosis; ILD: interstitial lung disease.
Figure 2.
The proposed model for the pathogenic role of NLRP3 inflammasome signaling pathway in RA and its clinical implications as the therapeutic potential. Several ligands that bind to TNFRs and TLRs can activate nuclear factor (NF)-κB. As a transcription factor, activated NF-κB can translocate into the nucleus and thereby activate the expression of NLRP3 and pro-IL-1β. As an endogenous ligand, anti-citrullinated peptide antibodies (ACPA), can promote NF-κB phosphorylation through binding to TNFRs and TLRs. In the second signal, extracellular ATP can bind to P2X7 and thereby lead to K+ efflux and extracellular Ca++ influx, which activate the NLRP3 inflammasome with overproduction of mature form of IL-1β and IL-18. The NLRP3 inflammasome activation also leads to the cleavage of gasdermin D, which promotes pyroptosis with the formation of pores in the cell membrane and the release of IL-1β and IL-18. ACPA can also activate the pannexin channel, resulting in ATP secretion and NLRP3 inflammation activation. Besides, In RA monocytes, complement C1q and pentaxin 3 (PTX3) synergistically activate the NLRP3 inflammasome and pyroptosis. Several compounds have been identified as inhibitors of the components of the NLRP3 inflammasome signaling pathway. MCC950, VX-765, osthole, and sulforaphane can inhibit the activation of the NLRP3 inflammasome. Tofacitinib, one of JAKi, may restore the balance of γδTreg/γδT17 cells in RA by inhibiting the NLRP3 inflammasome. Disulfiram inhibits GSDMD and thereby blocks pyroptosis and the release of IL-1β and IL-18. Anakinra, an IL-1β receptor antagonist, blocks the effects of NLRP3 inflammasome downstream cytokine. Among natural products, both Baihu-Guizhi decoction (BHGZD) and celastrol can inhibit NLRP3 activation by blocking the NF-κB pathway. HCQ could inhibit Ca2+-activated K+ channels and then impair inflammasome activation. The microRNA-30a and miR-223 inhibit the expression of NLRP3 and inflammasome activation. RA: rheumatoid arthritis; TNFα: tumor necrosis factor-α; TNFRs: TNFα receptors; TLRs: Toll-like receptors; ATP: adenosine triphosphate; PTX3: pentaxin 3; GSDMD: gasdermin D; IL: interleukin; ASCVD: atherosclerotic cardiovascular disease; OP: osteoporosis; ILD: interstitial lung disease.
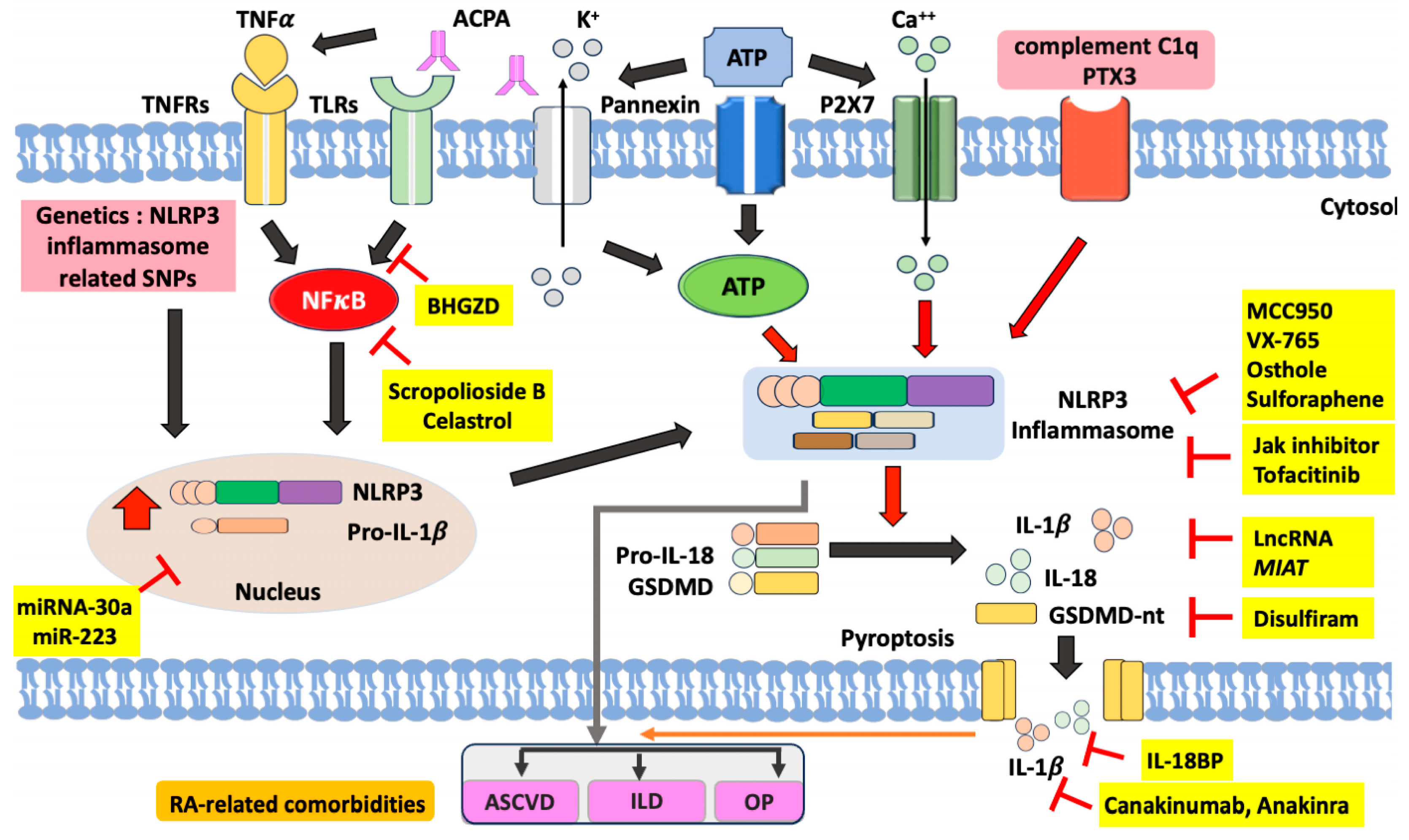

Table 1.
The potential inhibitors of the components of NLRP3 inflammasome in rheumatoid arthritis (RA).
Table 1.
The potential inhibitors of the components of NLRP3 inflammasome in rheumatoid arthritis (RA).
| Agents | Targets | Experimental model and mechanism | Diseases | References | ||
|---|---|---|---|---|---|---|
| Small molecule inhibitors | ||||||
| MCC950 | The NACHT domain of NLRP3 inflammasome | Block ASC oligomerization, inhibit inflammation (1) Reduce synovitis and cartilage erosion by inhibiting NLRP3 and caspae-1 activation in CIA model. (2) Elevated liver enzymes in phase II clinical trial. | RA, ASCVD, OP, ILD | [126,127,131,132] | ||
| VX-765 | Caspase-1 | Ameliorate the severity and progression of synovitis in CIA murine model. | RA, ASCVD | [128,133] | ||
| Disulfiram | GSDMD | Inhibits pyroptosis and inflammatory cytokine release in both canonical and noncanonical inflammasome pathways. | RA and associated OP | [129] | ||
| IL-18BP | IL-18 binding protein | Reduces Th17 cells with resultant inhibition of osteoclastogenesis and induces osteoblasts formation. | RA and associated OP | [134] | ||
| Natural products | ||||||
| Celastrol (isolated from Tripterygium wilfordii) | Inhibit the ROS-NF-κB-NLRP3 inflammasome axis. |
|
RA | [135,136] | ||
| Baihu-Guizhi decoction (BHGZD) | Inhibit TLR4/NF-κB/NLRP3 activation- induced pyroptosis. | Reduce synovitis as well as bone erosion and alleviate disease activity through inhibiting NF-κB via TLR4/PI3K/AKT signaling to suppress the NLRP3 inflammasome activation and GSDMD-mediated pyroptosis in AIA-modified rat model | RA | [138] | ||
| Sulforaphene (extracted from radish seeds) | NLRP3 | Suppress the M1 polarization of macrophages and reduce synovitis in CIA murine model | RA | [139] | ||
| Osthole (extracted from Angelicae pubescentis radix) | AMPK agonist | Inhibit NLRP3 inflammasome activation by regulating mitochondrial homeostasis in CIA rat model. | RA | [141] | ||
| Scropolioside B (isolated from Scrophularia dentada Royle ex Benth.) | NF-κB and the NLRP3 inflammasome pathway | Inhibit NF-κB activity, reduce NLRP3 expression, and suppress the maturation as well as the release of IL-1β. | RA and associated atherosclerosis | [142] | ||
| Wedelolactone, derived from Eclipta alba | NF-κB and the NLRP3 inflammasome | Ameliorate synovitis and cardiac complications via inhibiting the activation of NF-κB/NLRP3 inflammasome pathway | RA and cardiac complication | [143] | ||
| Disease-modifying anti-rheumatic drugs (DMARDs) | ||||||
| Hydroxychloroquine or chloroquine | The second signal of NLRP3 activation |
|
RA and associated ASCVD | [144,145] | ||
| Anakinra, a biological DMARDs | IL-1β receptor antagonist | Inhibit the NLRP3 inflammasome downstream cytokine, IL-1β, in RA patients. | RA | [146] | ||
| Canakinumab, a biological DMARDs | Monoclonal antibody targeting IL-1β | Reduce the rates of recurrent ASCVD, including myocardial infarction and stroke | RA and associated ASCVD | [147] | ||
| Tofacitinib, a Janus kinase 1/3 inhibitor | NLRP3 inflammasome | Restore the balance of γδTreg/γδT17 cells by inhibiting NLRP3 inflammasome in CIA model | RA | [148] | ||
| Epigenetic regulators | ||||||
| MiRNA-33 inhibitor | NLRP3 and caspase-1 | MiR-33 impairs mitochondrial oxygen consumption rate with increasing ROS, and then upregulates NLRP3 inflammasome expression in macrophages in CIA mice model | RA | [153] | ||
| MiRNA-30a | NLRP3 | MiRNA-30a inhibits the NLRP3 inflammasome activation, reduce synovitis, and bone damage in TNFα-transgenic C57BL/6 mice model. | RA | [154] | ||
| MiRNA-223 | NLRP3 | MiRNA-223 from BMSCs-derived exosomes inhibits NLRP3 activation and the release of IL-β, TNF-α, and IL-18 in RAW264.7 cells by luciferase reporter assay & rescue experiment | RA and associated ASCVD | [155] | ||
| LncRNA MIAT | IL-1β | LncRNA MIAT inhibited the expression of IL-1β, TNF-α and suppressed macrophage inflammation in J774A.1 cell-based assay. | RA | [160] | ||
| Allogenic mesenchymal stem cells | ||||||
| hUCB-MSCs | NLRP3 inflammasome | Downregulate the activation of NLRP3 inflammasome via a paracrine loop of IL-1β signaling in CIA murine model. | RA | [162] | ||
NLRP3: nucleotide-binding domain leucine-rich repeat-containing receptors (NLRs)-containing a pyrin domain-3; NACHT: nucleotide-binding domain; TLR4: Toll-like receptor 4; PI3K: phosphatidylinositol 3-kinase; AIA: adjuvant-induced arthritis; CIA: collagen-induced arthritis; CFA: complete Freund’s adjuvant; hUCB-MSCs: human umbilical cord blood mesenchymal stem cells; BMSCs: bone marrow-derived stem cells; ATP: adenosine triphosphate; ROS: reactive oxygen species; GSDMD: gasdermin-D; IL-1: interleukin-1; Treg: regulatory T cells; TNF-α: tumor necrosis factor-α; MiRNA: microRNA.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



