
Submitted:
07 December 2023
Posted:
08 December 2023
You are already at the latest version
Abstract
Introduction: Depending on the kind of cancer and neurological disorders, there might be a direct or inverse link between neurodegeneration and cancer. The inverse interaction between cancer and Alzheimer's pathology has garnered substantial interest in medical research. While this inverse interaction between Alzheimer's pathology and cancer presents a fascinating scientific puzzle, there is still limited information and studies on the potential mechanisms involved in this inverse rela-tionship. Aim: This study aims to identify potential genes from Alzheimer's disease that may protect against cancer.
Method: The Gene Expression Omnibus (GEO) database provided the Alzheimer's pathology dataset, and GEO 2R was utilized to examine genes that were differentially expressed. The expression of upregulated Alzheimer's pathology genes in breast, colorectal, and prostate cancer was compared using VENN 2.1, colorectal, and prostate cancer was compared using VENN 2.1, and colorectal and prostate cancers were compared using VENN 2.1.
Results: Among the upregulated genes associated with Alzheimer's disease, 25 genes are shared among breast cancer, colorectal cancer, and prostate cancer, while 28 of these genes were exclusive to Alzheimer's dis-ease. Gene expression profiling interactive analysis among the shared genes revealed that AQP1, C4A, CSRP2, CRYAB, MACF1, HSPB2, and NEAT1 were downregulated in breast, colorectal, and prostate cancers. These genes have varying roles in cancer progression and suppression. Functional analysis of Alzheimer's disease-related upregulated genes revealed that C1orf56 and HIST1H2AC control cell proliferation, with HIST1H2AC playing a role in necroptosis. Additionally, C1orf56 worked as an oncogenic modifier to support DNMT3B's tumor suppressor properties.
Conclusion: Given the role of HIST1H2AC and C1orf56 in the negative control of cell proliferation and their upregulation in Alzheimer's disease, there is a need to further investigate HIST1H2AC and C1orf56 as potential tumor suppressors responsible for Alzheimer's disease and cancer inverse association.
Keywords:
Alzheimer's disease
; inverse
; breast
; colorectal cancer
; prostate
; expression
1. Introduction
Alzheimer's pathology is a neurological illness that worsens over time and mostly affects behavior, memory, and cognitive abilities. It is responsible for the development of dementia, a condition typified by a reduction in cognitive ability that impairs functioning and everyday living. Damaged brain cells and the loss of neural connections are signs of neurodegenerative brain illnesses, which severely limit a person's ability to carry out daily duties[1]. Dementia is the sixth most common cause of mortality worldwide, with a new case reported every three seconds. About 23 million people are living with dementia in the Asia-Pacific area, 8.8 million in Europe, 5.8 million in the USA, and 5.3 million in India. The global figure is as high as 50 million. By 2050, this figure is expected to increase to 152 million[2]. A global assessment of Alzheimer's reports from 2020 estimates that the overall projected cost of dementia worldwide is one trillion US dollars, and by 2030, that sum is expected to rise to two trillion US dollars [3]. One of the main characteristics of Alzheimer's disease is the culmination of abnormal protein deposits within the brain. The illness is linked to two main types of these aberrant protein accumulations: tau tangles and beta-amyloid plaques. Clusters of beta-amyloid protein fragments that accumulate between nerve cells and impair cellular function are known as beta-amyloid plaques. On the other hand, tau tangles are made of twisted protein fibers that accumulate inside nerve cells, causing them to malfunction and ultimately die [4]. Although the precise cause of Alzheimer's disease remains incompletely elucidated, it is believed to arise from genetic, environmental, and lifestyle factors contributing to its development[5,6].
A variety of symptoms are brought on by the advancement of Alzheimer's pathology and worsen with time. Memory disorders, disorientation, and trouble finishing familiar activities are common early signs. As the illness worsens, people may have altered personalities, trouble speaking, and poor judgment[7,8]. The physical alterations in the brain and the consequent reduction in cognitive function have a significant effect on the person's overall well-being. Although there is no known cure for Alzheimer's pathology, there are some interventions and therapies that can help control the symptoms and delay the illness's progression. These might include behavioral therapy and cognitive training as well as temporary pharmacological relief for cognitive symptoms and techniques to maintain memory and cognitive function. It has also been demonstrated that lifestyle changes including consistent physical activity, a balanced diet, social interaction, and mental stimulation have a good effect on general brain health [9]. Alzheimer's patients and their families, carers, and society at large face several obstacles as a result of the condition. Certain determinants are similar to both conditions, however, the likelihood of developing Alzheimer's pathology and cancer is inversely correlated. Compared to control people, Alzheimer's patients had a 61 percent decreased risk of acquiring cancer[10]
The inverse interaction between cancer and Alzheimer's is a perplexing phenomenon that has garnered substantial attention within medical research[11,12]. This intriguing observation suggests that individuals who develop Alzheimer's disease seem less likely to develop cancer. While the exact mechanisms behind this association remain unclear, several theories have been proposed to explain this phenomenon. One possible explanation is related to the role of the immune system. Cancer involves uncontrolled cell growth and division, which the immune system often recognizes as abnormal and attempts to suppress. In contrast, Alzheimer's pathology is typified by the accumulation of misfolded proteins and chronic inflammation. It is speculated that the immune response directed toward cancer cells might inadvertently help protect against Alzheimer's - related protein accumulation by stimulating immune mechanisms that target abnormal protein deposits[13]. Another theory involves the influence of genetics and specific molecular pathways. Research has identified certain genetic factors associated with Cancer and Alzheimer's disease. Variations in these genes might affect an individual's susceptibility to either condition. Additionally, cellular growth, metabolism, and energy utilization pathways could shape the interaction between Alzheimer's and cancer [13,14]. While individual studies have reported on the inverse relationship involving Alzheimer's pathology and cancer, there is a critical gap in our comprehension of the specific molecular mechanisms that drive this association[12]. The interaction linking Alzheimer's disease and cancer is not straightforward and can vary depending on the specific types of cancer and the stage of each disease. The exact nature of this association is still an active area of research, and further studies are required to fully comprehend the underlying mechanisms. Understanding why certain individuals are seemingly protected from one condition while experiencing an increased risk of the other holds immense potential for shedding light on shared pathways, regulatory networks, and cellular responses that underlie both Alzheimer's disease and cancer. To address this knowledge gap, comprehensive research is needed to unravel the intricate molecular interactions contributing to this inverse relationship. There has been few research conducted on the inverse interaction betwixt Alzheimer's disease and cancer, even though it offers an intriguing scientific challenge.
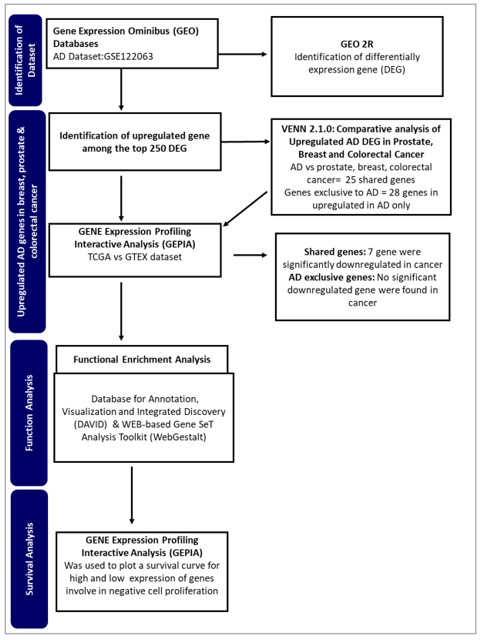
This study aims to identify potential genes from Alzheimer's disease that may be involved in the inverse interaction betwixt Alzheimer's pathology and cancer. By employing an integrated bioinformatics approach (Figure 1), the study seeks to identify genes and regulatory networks that might provide insight into the observed phenomenon.
2. Results
2.1. Differential Gene Expression
Analysis of Alzheimer's disease genes revealed differentially expressed genes between Alzheimer's disease and non-dementia controls (Figure 1). The top 250 significant differentially expressed genes were downloaded and used for further analysis.
2.2. Gene Filtration
The top 250 differentially expressed genes were filtered for Alzheimer's pathology -upregulated genes. 84 of the differential Alzheimer's pathology genes were identified to be significantly upregulated among the top 250 genes (Figure 2).
2.3. Cancer union Alzheimer's disease complements Genes
The upregulated Alzheimer's disease genes were compared to genes present in prostate cancer, breast cancer, and colon cancer to identify the common genes shared by Alzheimer's pathology and cancers and to reveal the genes related to Alzheimer's pathology only and not the cancers. In all, 25 upregulated Alzheimer's pathology genes were identified to be common in all the cancers (shared genes), whereas 28 upregulated genes were found to be exclusively present in Alzheimer's pathology and not the cancers (AD exclusive genes). We, therefore, hypothesized that some of these Alzheimer's disease-upregulated genes may contribute to the inverse association between cancers and Alzheimer's pathology. Nevertheless, the shared genes may also account for inverse association depending on the level of expression and function of these genes.
2.4. Expression of Alzheimer's disease upregulated genes in breast, prostate, and colon cancer
Expression patterns of common genes between Alzheimer's disease and breast, prostate, and colon cancer were determined. The Upregulated genes in Alzheimer's pathology that are also significantly downregulated in breast, prostate, and colon cancer are represented in Figure 5
2.5. Functional Analysis
The DAVID, GOTERM Biological Process analysis after functional enrichment showed that C1orf56 and HIST1H2AC are involved in regulating cell proliferation. This analysis was further confirmed using the WebGestalt KEGG pathway analysis, which showed that HIST1H2AC is involved in necroptosis.
2.6. Survival Analysis
Gene survival analysis was conducted for the two AD-upregulated genes identified to be involved in regulating cell proliferation. The hazard ratio (HR) was determined
3. Discussion
The inverse interaction between Alzheimer's disease and cancer continues to be a complex and intriguing topic within the medical community. Cancer-Alzheimer's disease paradox suggests that individuals with a previous history of cancer may have a reduced risk of progressing to Alzheimer's pathology[15]. Research into the mechanisms driving this relationship could provide valuable insights into the fundamental processes governing these two major diseases, potentially paving the way for innovative therapeutic strategies and a deeper understanding of human health and aging. In this study, we used an integrated computational approach to identify upregulated genes in Alzheimer's pathology that may also influence cancer suppression and progression. We hypothesized that these genes may potentially be associated with the inverse comorbidity betwixt Alzheimer's pathology and cancer.
Comparative gene analysis reveals a similarity of 0.1% of the upregulated genes found in Alzheimer's disease-related genes and prostate, breast, and colorectal cancer. The gene expression analysis affirms that most of these genes were distinctively downregulated in the cancers. AQP1, C4A, CSRP2, CRYAB, MACF1, HSPB2, and NEAT1 were downregulated in some cancers. However, the role of these genes in tumor suppression and promotion is context-dependent.
HspB2 is a unique member of the small HSP family predominantly expressed in skeletal and heart muscles. Traditionally, HSPS has been linked to cancer development, but HspB2 contradicts this by acting as a tumor suppressor. This protein plays an anti-cancer role by partially restoring mutant p53 activity and promoting the expression of tumor-suppressor genes RPRM, BAI-1, and TSAP6[16]. In certain tumors, like pancreatic and esophageal cancers, HSPB2 levels are reduced or absent. HSPB2 also repairs p53 transcriptional activity, leading to the inhibition of pancreatic cancer cell progression. However, its role in breast cancer is debated, with some studies showing suppression [17], while others indicating progression[18]. Given these findings, further investigations into HspB2 and its variants, especially in breast cancer, are vital to explain the role of HspB2 in cancer and provide insight into its potential as a possible tumor suppressor in Alzheimer's disease.
CSRP2 is a multifunctional protein involved in cell proliferation, differentiation, and adhesion. CSRP2 also suppresses colorectal cancer progression via the P130Cas/Rac1 signaling pathway[19]. Again, CSRP2 is thought to hinder the spread and metastasis of cancer cells in gastric cancer, whereas it plays a contrary function in breast cancer[19,20]. Therefore, the underlying mechanisms of CSRP2 function in various cancers need to be carefully examined. The protein's function in cancer is complex, and understanding its intricacies could highlight its role in Alzheimer's disease.
CRYAB suppresses NPC tumor formation in mice and affects NPC progression-associated phenotypes[21]. CRYAB inhibits migration and invasion through various mechanisms, such as inhibiting RAS activation and the Raf/MEK/ERK signaling pathway[22]. While CRYAB has tumor-suppressive properties in the nasopharyngeal, and bladder, and it also promotes angiogenesis in some cancers. However, to elucidate the role of CRYAB in cancer and Alzheimer's pathology, there is a need to investigate various forms of CRYAB present in Alzheimer's disease[22,23].
MACF1 is a multidomain cytoskeletal linker protein essential for cell migration, cytoskeletal organization, and intracellular transport. In cancer, it can both promote and inhibit progression[24,25] MACF1 can suppress cancer cell motility by maintaining cytoskeletal integrity and may interact with tumor suppressor proteins [25]. Though MACF1 has a multifaceted role in cancer, it could be responsible for the inverse association between Alzheimer's disease and cancer since it can interact with tumor suppressor proteins.
Neat1 plays an important role in the growth and specialization of the mammary gland and corpus luteum. However, its impact on cancer has resulted in contradictory findings. While some research links Neat1 to the advancement of tumors, recent studies have demonstrated its potential as a suppressor of tumor growth [26]. The tumor-suppressive actions of NEAT1 have been attributed to several molecular pathways. These include preserving pre-messenger RNAs in the cell nucleus and promoting the development of pre-mRNAs and pre-microRNAs[27,28], protein sequestration into paraspeckles and removal from chromatin[29], and controlling the expression of genes by making direct contacts with chromatin. It is s possible that NEAT1 suppresses tumors by influencing gene expression via a variety of methods simultaneously, or that its ability to operate as a tumor suppressor depends only on a subset of these processes [27,28,30]. When exposed to nutlin or Adriamycin, increased levels of NEAT1 have been shown to inhibit the development of cells in lung cancer and osteosarcoma cell lines [31]. Again, increased levels of NEAT1 expression were linked to improved survival rates in cases of nasopharyngeal cancer. Additionally, elevated NEAT1 levels were found to significantly suppress the growth of nasopharyngeal cancer cells in subcutaneous tumor experiments, functioning through a process that inhibits miR-101-3p[32]. Research indicates that point mutations and deletions in the NEAT1 promoter are rather common in cases of liver and breast cancer. Moreover, mutations found in the NEAT1 promoter in breast cancer frequently result in decreased NEAT1 production. Although it is not shown that the particular mutations seen in breast cancer patients influence the p53 binding site, the deletions often damage the p53 response element[33,34,35]. Furthermore, it was discovered that decreasing NEAT1 1 in colorectal cancer cell lines decreased invasiveness and cell proliferation[36]. While the precise process of tumor inhibition remains undefined, NEAT1 shows promise as a tumor suppressor in individuals with Alzheimer's disease, potentially playing a role in the reduced susceptibility of Alzheimer's patients to cancer.
Furthermore, the comparative analysis also showed that 28 genes are exclusively upregulated in Alzheimer's disease and cannot be found in prostate, breast, and colorectal cancer. Therefore, we hypothesized that these genes could be responsible for the inverse comorbidity observed between cancers and Alzheimer's disease. The upregulation of genes exclusively related to Alzheimer's disease could be involved in inverse comorbidity. The fact that these 28 genes are upregulated in Alzheimer's pathology but not in the mentioned cancers suggests that Alzheimer's pathology and these cancers may involve distinct molecular and cellular pathways. The processes driving Alzheimer's pathology progression might differ from those driving these cancers' progression. If these genes influence the pathology of Alzheimer's, their absence or repression in certain cancers could contribute to the observed inverse association. The molecular processes they trigger may be involved in the protective effects against the onset or progression of these cancers. To draw more concrete conclusions, it is crucial to understand the biological functions of these 28 genes, particularly their involvement in cell growth, apoptosis, DNA repair, immune response, or neural functions.
Functional analysis of the biological processes of these genes revealed that HIST1H2AC is involved in necroptosis, a form of regulated cell death morphologically and mechanistically distinct from apoptosis. Unlike apoptosis, which is a form of programmed cell death generally considered non-inflammatory and involves the activation of caspases, necroptosis resembles necrotic cell death and is often associated with inflammation. As a member of the histone H2A family, the HIST1H2AC gene is essential to the structural arrangement of chromatin, which can affect a number of DNA-related functions, including transcription, replication, and repair. The expression levels of histone modifications have a significant impact on how genes and cellular pathways are regulated [37]. In the context of uterine cervical preneoplastic lesions, studies have shown a trend in the expression of the HIST1H2AC gene. Specifically, there has been a progressive downregulation of HIST1H2AC in keratinocytes that are positive for Human papillomavirus (HPV). HPV is a well-established causative agent for cervical cancer, with certain high-risk strains being more associated with malignancy than others. Though the expression of HIST1H2AC has not been investigated in various types of cancers, this observation suggests a potential correlation between decreased HIST1H2AC expression and advancing cellular abnormalities. Therefore, we postulate that the upregulation of HIST1H2AC may be a factor for the inverse correlation between cancer and Alzheimer's pathology. HIST1H2AC upregulation could influence chromatin structure, gene accessibility, and, consequently, cellular growth and differentiation in Alzheimer's pathologic patients. However, further studies on its exact role, mechanism of downregulation, and broader implications of HIST1H2AC could provide valuable insights into cancer biology and potential therapeutic strategies.
Biological process analysis reveals that HIST1H2AC is involved in negative cell proliferation and chromatin assembling, this further confirms the involvement of HIST1H2AC in necroptosis. Given the role of HIST1H2AC regulation of cell proliferation and its upregulation in Alzheimer's disease, there is a need to further investigate HIST1H2AC as a potential tumor suppressor responsible for Alzheimer's disease and cancer inverse association. Furthermore, the biological process also revealed that C1orf56 is involved in the regulation of cell proliferation. C1orf56 was involved in the control of cellular proliferation and served as an oncogenic modifier contributing to the tumor suppressor function of DNMT3B. Dnmt3b affects tumor progression [38]. This shows that genes involved in Alzheimer's disease complexities continue to be explored. Characterizing these genes may also provide insight into their role in the inverse association between cancer and Alzheimer's disease.
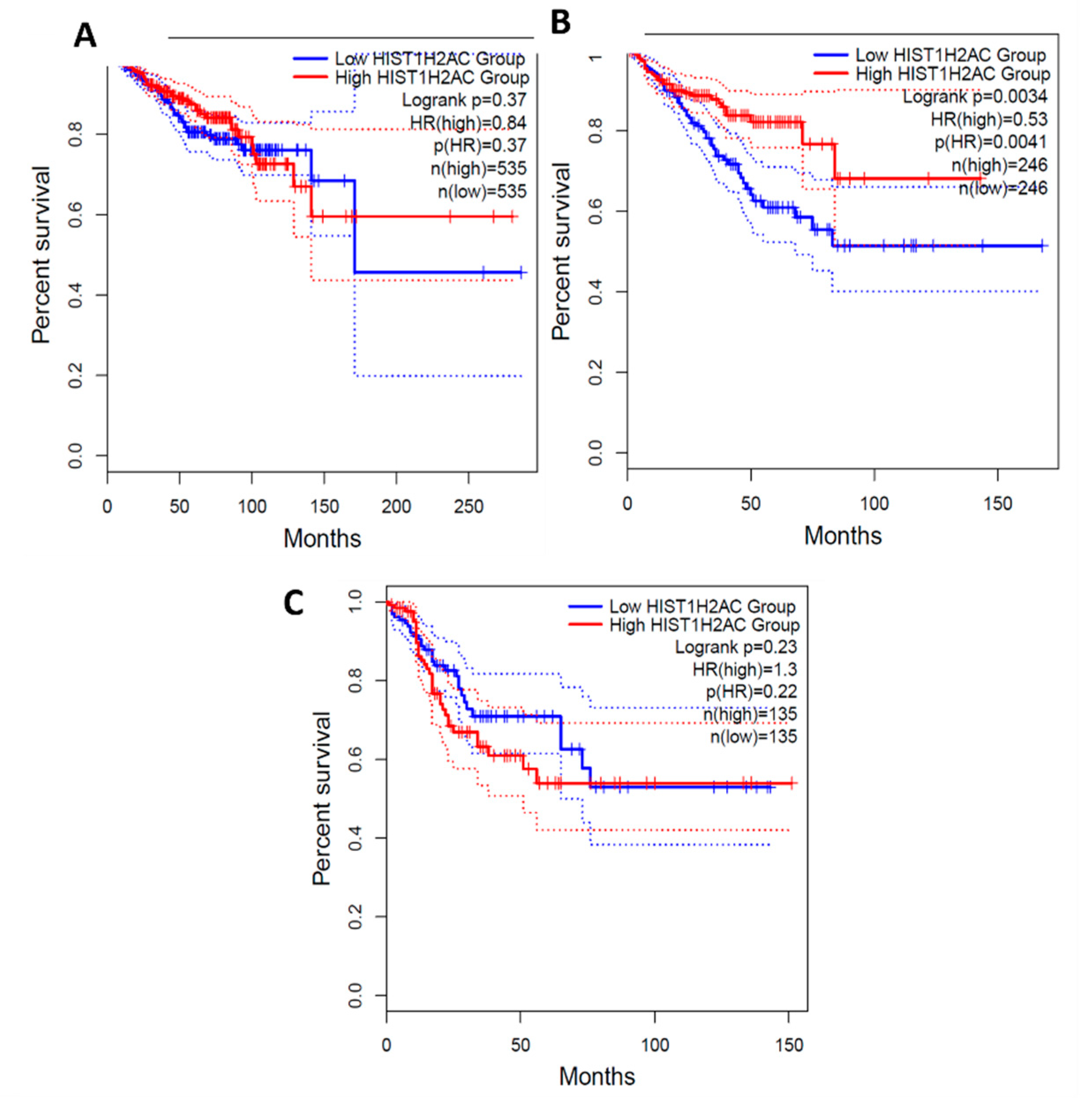
From the survival analysis, high expression of C1orf56 in the breast and prostate did show a significant improvement in survival with a hazard ratio of 1.1 and 1.2 however there was an observable difference in colorectal cancer patient survival with high expression of C1orf56. This indicates that C1orf56 could potentially suppress colorectal cancer progression and improve survival. Though the survival analysis did not give a consistent pattern in all cells, there is an indication that the role of these genes needs to be further elucidated. For HIST1H2AC survival analysis, high expression of HIST1H2AC was associated with improvement in breast and prostate cancer, however contrary to C1orf56, HIST1H2AC did not have an observable effect on improving patient survival. This indicates that different factors may work together to reduce the risk of cancer development in AD patients.
In conclusion, there is a need to further investigate HIST1H2AC and C1orf56 as potential tumor suppressors responsible for the inverse interaction betwixt Alzheimer's disease and cancer given the role of HIST1H2AC and C1orf56 in negative regulation of cell proliferation and their upregulation in Alzheimer's pathology. The inverse interaction betwixt Alzheimer's disease and cancer may depend on various factors, such as the upregulation of potential tumor suppressors whose roles are not clearly defined and the upregulation of genes involved in cell proliferation. From this study, it is also apparent that there is limited information on the role of most of the upregulated Alzheimer's disease genes in cancer; hence, further research is needed into some of these genes and their interacting pathways.
4. Materials and Methods
4.1. Identification and retrieval of dataset
We accessed the gene expression microarray data associated with Alzheimer's disease using the Gene Expression Omnibus (GEO) at the National Center for Biotechnology Information (NCBI) (http://www.ncbi.nlm.nih.gov/geo/). The specific dataset identified by the accession number GSE122063 was extracted for our research. GSE122063 is comprised of Agilent-039494 SurePrint G3 Human GE v2 8x60K Microarray data featuring gene expression profiles from the frontal and temporal cortex. Through the University of Michigan Brain Bank, these profiles were gathered from people who had Alzheimer's disease (AD), vascular dementia (VaD), and non-demented controls (Control). In the autopsied hemisphere, there were no infarcts seen in the controls or Alzheimer's disease patients. Expression profiles of control and Alzheimer's pathology were only selected for our study. The dataset used for this study contains 56 separate microarray expression profiles from 12 Alzheimer's pathology samples and 44 separate microarray expression profiles from 10 non-demented controls[39]. Meanwhile, cancer-related targets were obtained from the Genecards Database (https://www.genecards.org/). Genecard integrates data from more than 190 sources to provide a disease-centric, gene-centric, and pathway-centric database with interconnections and rich annotations. All annotated and predicted human genes involved in breast, prostate, and colorectal cancer were identified and used for this study.
4.2. Differentially expressed gene identification
We performed the differentially expressed genes (DEGs) analysis of the human microarray dataset by comparing gene expression profiles from Alzheimer's disease samples and the gene expression profiles control samples from persons without dementia. The built-in R software, GEO2R, within the Gene Expression Omnibus, was used to analyze the expression of genes. With the interactive web application GEO2R, users may compare several sample groups within a GEO Series. To provide high-throughput genomic data processing capabilities, GEO2R uses a number of R packages from the Bioconductor project, an open-source software project that uses the R programming language. The research used the Linear Models for Microarray (Limma) program in Bioconductor. To identify the statistically significant differentially expressed genes (DEGs) for both datasets, a minimum absolute base-2 log Fold Change (logFC) value of 1.0 and an FDR-adjusted p-value threshold of 0.05 were set, respectively. Cross-comparative research then revealed the important DEGs that the brain and blood have in common.
4.3. Upregulated Alzheimer's disease genes
Finding the elevated genes in Alzheimer's disease was important because the study focused on the condition. Thus, genes associated with Alzheimer's disease that were substantially expressed in comparison to the control and had a log fold change more than 0 at a p-value threshold of 0.05 were chosen and employed in subsequent research.
4.4. Identification of upregulated Alzheimer's disease gene that overlaps in Prostate, Breast, and Colorectal cancer
Venny 2.1.0 was used to conduct a common target screening for elevated Alzheimer's disease genes in colorectal, prostate, and breast malignancies. A Venn diagram with shared objectives was produced. The Gene Expression Profiling Interactive Analysis website was used to assess targets associated with cancer and Alzheimer's disease that were overlapping and elevated (GEPIA2).
To assess the expression of the Alzheimer's disease elevated genes in breast, prostate, and colorectal cancers, the Gene Expression Profiling Interactive Analysis (GEPIA2) web server (http://gepia2.cancer-pku.cn/, accessed on July 5, 2023) was utilized. Gene expression data from the Genotype-Tissue Expression (GTEx) project, which provides expression data for normal tissues, and The Cancer Genome Atlas (TCGA), which contains expression data from tumor tissues, are both included in GEPIA. The thresholds for statistical significance were |Log2FC| > 1 and p-value < 0.01.
4.5. Functional Enrichment Analyses and Gene Orthology
The Database for Annotation, Visualization, and Integrated Discovery (DAVID) (http://david.ncifcrf.gov; version 6.8) is an accessible platform that combines biological information and analytical resources to annotate genes and pathways functionally. GO is a computational tool to label genes and study their involvement in various biological processes. Additionally, the WEB-based Gene Set Analysis Toolkit (WebGestalt) is a repository for assessing pertinent signaling pathways in extensive molecular datasets resulting from high-throughput experimental methods. In this study, DAVID facilitated the GO enrichment analysis of the Differentially Expressed Genes (DEGs) in terms of their biological processes, whereas WebGestalt was utilized to conduct KEGG pathway enrichment analysis, elucidating the roles of the DEGs in cell signaling pathways.
4.6. Survival Analysis
The Gene Expression Profiling Interactive Analysis (GEPIA2) web server function for survival analysis was used to determine the survival pattern for high and low expression of C1orf56 and HIST1H2AC in breast, prostate and colorectal cancer data from The Cancer Genome Atlas (TCGA), which contains expression data from tumor tissues. The Confidence interval was set at 95%. GEPIA uses the Cox Proportional-Hazards Model to determine the Hazard Ratio of survival.
Author Contributions
Conceptualization, B.K.A, K.F, J.W.; Methodology and data analysis, B.K.A, K.F.; Data Validation, J.W., T.L.; writing—original draft preparation, K.F., B.K.A, S.K.; writing—review and editing, T.L., S.K., J.W., B.K.A.; funding acquisition, J.W.
Funding
The project was partially supported by the National Natural Science Foundation of China (32161143021, 81271410) and Henan Natural Science Foundation of China (182300410313).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
No new data were created.
Acknowledgments
Author would like to Acknowledge Irene Anyomi for her guidance in retrieving the Alzheimer's disease dataset.
Conflicts of Interest
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
References
- Kumar, A. , Sidhu, J., Goyal, A., Tsao, J.W. Alzheimer Disease. W. Alzheimer Disease. StatPearls 2022, 1–27. [Google Scholar]
- 2020 Alzheimer’s disease facts and figures. Alzheimer’s & Dementia 2020, 16, 391–460.
- Siddappaji, K.K.; Gopal, S. Molecular mechanisms in Alzheimer’s disease and the impact of physical exercise with advancements in therapeutic approaches. AIMS Neurosci 2021, 8, 357. [Google Scholar] [CrossRef]
- Alzheimer’s disease - Causes - NHS. Available online: https://www.nhs.uk/conditions/alzheimers-disease/causes/ (accessed on 25 August 2023).
- Livingston, G.; Huntley, J.; Sommerlad, A.; et al. Dementia prevention, intervention, and care: 2020 report of the Lancet Commission. The Lancet 2020, 396, 413–446. [Google Scholar] [CrossRef]
- Raulin, A.C.; Doss, S.V.; Trottier, Z.A.; Ikezu, T.C.; Bu, G.; Liu, C.C. ApoE in Alzheimer’s disease: pathophysiology and therapeutic strategies. Molecular Neurodegeneration 2022, 17, 1–26. [Google Scholar] [CrossRef]
- Collins, M.E. Occupational Therapists’ Experience with Assistive Technology in Provision of Service to Clients with Alzheimer’s Disease and Related Dementias. Phys Occup Ther Geriatr 2018, 36, 179–188. [Google Scholar] [CrossRef]
- Dementia. Available online: https://www.asha.org/practice-portal/clinical-topics/dementia/ (accessed on 25 August 2023).
- Yiannopoulou, K.G.; Papageorgiou, S.G. Current and Future Treatments in Alzheimer Disease: An Update. J Cent Nerv Syst Dis. 2020. [Google Scholar] [CrossRef] [PubMed]
- Zabłocka, A.; Kazana, W.; Sochocka, M.; Stańczykiewicz, B.; Janusz, M.; Leszek, J.; Orzechowska, B. Inverse Correlation Between Alzheimer’s Disease and Cancer: Short Overview. Mol Neurobiol 2021, 58, 6335. [Google Scholar] [CrossRef]
- Li, J.M.; Liu, C.; Hu, X.; Cai, Y.; Ma, C.; Luo, X.G. Yan, X.X. Inverse correlation between Alzheimer’s disease and cancer: implication for a strong impact of regenerative propensity on neurodegeneration? BMC Neurol 2014, 14, 211. [Google Scholar] [CrossRef]
- Roe, C.M.; Behrens, M.I.; Xiong, C.; Miller, J.P.; Morris, J.C. Alzheimer disease and cancer. Neurology 2005, 64, 895–898. [Google Scholar] [CrossRef]
- Zabłocka, A.; Kazana, W.; Sochocka, M.; Stańczykiewicz, B.; Janusz, M.; Leszek, J.; Orzechowska, B. Inverse Correlation Between Alzheimer’s Disease and Cancer: Short Overview. Mol Neurobiol 2021, 58, 6335. [Google Scholar] [CrossRef] [PubMed]
- Behrens, M.I.; Lendon, C.; Roe, C.M. A common biological mechanism in cancer and Alzheimer’s disease? Curr Alzheimer Res 2009, 6, 196. [Google Scholar] [CrossRef]
- Zhang, D.D.; Ou, Y.N.; Yang, L.; Ma, Y.H.; Tan, L.; Feng, J.F.; Cheng, W.; Yu, J.T. Investigating the association between cancer and dementia risk: a longitudinal cohort study. Alzheimers Res Ther 2022, 14, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Yu, Z.; Wang, H.; Fang, Y.; Lu, L.; Li, M.; Yan, B.; Nie, Y.; Teng, C. Molecular chaperone HspB2 inhibited pancreatic cancer cell proliferation via activating p53 downstream gene RPRM, BAI1, and TSAP6. J Cell Biochem 2020, 121, 2318–2329. [Google Scholar] [CrossRef]
- Zoppino, F.C.M.; Guerrero-Gimenez, M.E.; Castro, G.N.; Ciocca, D.R. Comprehensive transcriptomic analysis of heat shock proteins in the molecular subtypes of human breast cancer. BMC Cancer 2018, 18, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Sklirou, A.D.; Gianniou, D.D.; Karousi, P.; Cheimonidi, C.; Papachristopoulou, G.; Kontos, C.K.; Scorilas, A.; Trougakos, I.P. High mRNA Expression Levels of Heat Shock Protein Family B Member 2 (HSPB2) Are Associated with Breast Cancer Patients’ Relapse and Poor Survival. Int J Mol Sci. 2022. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Long, X.; Duan, S.; Liu, X.; Chen, J.; Lan, J.; Liu, X.; Huang, W.; Geng, J.; Zhou, J. CSRP2 suppresses colorectal cancer progression via p130Cas/Rac1 axis-meditated ERK, PAK, and HIPPO signaling pathways. Theranostics 2020, 10, 11063–11079. [Google Scholar] [CrossRef]
- Wang, J.; Guan, X.; Zhang, Y.; et al. Exosomal miR-27a Derived from Gastric Cancer Cells Regulates the Transformation of Fibroblasts into Cancer-Associated Fibroblasts. Cellular Physiology and Biochemistry 2018, 49, 869–883. [Google Scholar] [CrossRef]
- Huang, Z.; Cheng, Y.; Chiu, P.M.; et al. Tumor suppressor Alpha B-crystallin (CRYAB) associates with the cadherin/catenin adherens junction and impairs NPC progression-associated properties. Oncogene 2012, 31, 3709–3720. [Google Scholar] [CrossRef]
- Ruan, H.; Li, Y.; Wang, X.; Sun, B.; Fang, W.; Jiang, S.; Liang, C. CRYAB inhibits migration and invasion of bladder cancer cells through the PI3K/AKT and ERK pathways. Jpn J Clin Oncol 2020, 50, 254–260. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.; Ruan, Q.; Han, S.; Xi, L.; Jiang, W.; Jiang, H.; Ostrov, D.A.; Cai, J. Discovery of structure-based small molecular inhibitor of αB-crystallin against basal-like/triple-negative breast cancer development in vitro and in vivo. Breast Cancer Res Treat 2014, 145, 45–59. [Google Scholar] [CrossRef]
- Miao, Z.; Ali, A.; Hu, L.; Zhao, F.; Yin, C.; Chen, C.; Yang, T.; Qian, A. Microtubule actin cross-linking factor 1, a novel potential target in cancer. Cancer Sci 2017, 108, 1953–1958. [Google Scholar] [CrossRef]
- Zhao, W.; Qian, H.; Zhang, R.; Gao, X.; Gou, X. MicroRNA targeting microtubule cross-linked protein (MACF1) would suppress the invasion and metastasis of malignant tumor. Med Hypotheses 2017, 104, 25–29. [Google Scholar] [CrossRef]
- Mello, S.S.; Attardi, L.D. Neat-en-ing up our understanding of p53 pathways in tumor suppression. Cell Cycle 2018, 17, 1527–1535. [Google Scholar] [CrossRef]
- Jiang, L.; Shao, C.; Wu, Q.J.; et al. NEAT1 Scaffolds RNA Binding Proteins and the Microprocessor to Globally Enhance Pri-miRNA Processing. Nat Struct Mol Biol 2017, 24, 816. [Google Scholar] [CrossRef]
- Cooper, D.R.; Carter, G.; Li, P.; Patel, R.; Watson, J.E.; Patel, N.A. Long Non-Coding RNA NEAT1 Associates with SRp40 to Temporally Regulate PPARγ2 Splicing during Adipogenesis in 3T3-L1 Cells. Genes 2014, 5, 1050. [Google Scholar] [CrossRef]
- Hirose, T.; Virnicchi, G.; Tanigawa, A.; et al. A Highlights from MBoC Selection: NEAT1 long noncoding RNA regulates transcription via protein sequestration within subnuclear bodies. Mol Biol Cell 2014, 25, 169. [Google Scholar] [CrossRef]
- West, J.A.; Davis, C.P.; Sunwoo, H.; Simon, M.D.; Sadreyev, R.I.; Wang, P.I.; Tolstorukov, M.Y.; Kingston, R.E. The long noncoding RNAs NEAT1 and MALAT1 bind active chromatin sites. Mol Cell 2014, 55, 791. [Google Scholar] [CrossRef]
- Idogawa, M.; Ohashi, T.; Sasaki, Y.; Nakase, H.; Tokino, T. Long non-coding RNA NEAT1 is a transcriptional target of p53 and modulates p53-induced transactivation and tumor-suppressor function. Int J Cancer 2017, 140, 2785–2791. [Google Scholar] [CrossRef]
- Wang, Y.; Wang, C.; Chen, C.; Wu, F.; Shen, P.; Zhang, P.; He, G.; Li, X. Long non-coding RNA NEAT1 regulates epithelial membrane protein 2 expression to repress nasopharyngeal carcinoma migration and irradiation-resistance through miR-101-3p as a competing endogenous RNA mechanism. Oncotarget 2017, 8, 70156. [Google Scholar] [CrossRef]
- Nakagawa, H.; Fujita, M.; Fujimoto, A. Genome sequencing analysis of liver cancer for precision medicine. Semin Cancer Biol 2019, 55, 120–127. [Google Scholar] [CrossRef]
- Fujimoto, A.; Furuta, M.; Totoki, Y.; et al. Whole-genome mutational landscape and characterization of noncoding and structural mutations in liver cancer. Nat Genet 2016, 48, 500–509. [Google Scholar] [CrossRef]
- Rheinbay, E.; Parasuraman, P.; Grimsby, J.; et al. Recurrent and functional regulatory mutations in breast cancer. Nature 2017, 547, 55–60. [Google Scholar] [CrossRef]
- Wu, Y.; Yang, L.; Zhao, J.; et al. Nuclear-enriched abundant transcript 1 as a diagnostic and prognostic biomarker in colorectal cancer. Mol Cancer 2015, 14, 191. [Google Scholar] [CrossRef]
- Cohen, I.; Porȩba, E.; Kamieniarz, K.; Schneider, R. Histone Modifiers in Cancer: Friends or Foes? Genes Cancer 2011, 2, 631. [Google Scholar] [CrossRef]
- C1orf56 chromosome 1 open reading frame 56 [Homo sapiens (human)] - Gene - NCBI. Available online: https://www.ncbi.nlm.nih.gov/gene/54964 (accessed on 21 October 2023).
- McKay, E.C.; Beck, J.S.; Khoo, S.K.; Dykema, K.J.; Cottingham, S.L.; Winn, M.E.; Paulson, H.L.; Lieberman, A.P.; Counts, S.E. Peri-Infarct Upregulation of the Oxytocin Receptor in Vascular Dementia. J Neuropathol Exp Neurol 2019, 78, 436. [Google Scholar] [CrossRef]
Figure 1.
A flowchart of the approach used in this study.
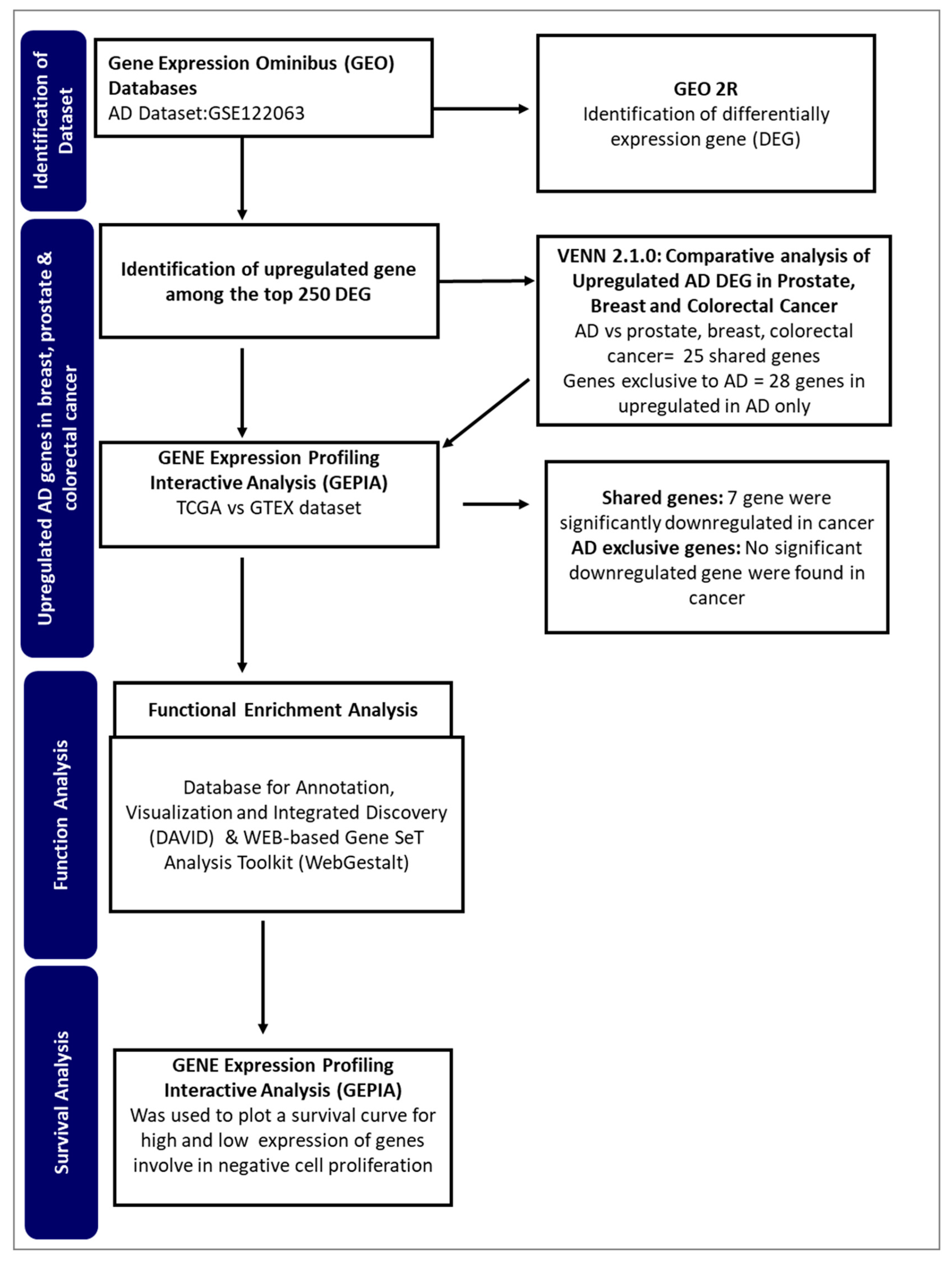
Figure 2.
Differentially expressed genes of Alzheimer's disease. The differential expressed genes of Alzheimer's pathology were presented using a Volcano plot. The red color represents upregulated genes, whereas the blue color represents downregulated genes.
Figure 2.
Differentially expressed genes of Alzheimer's disease. The differential expressed genes of Alzheimer's pathology were presented using a Volcano plot. The red color represents upregulated genes, whereas the blue color represents downregulated genes.
Figure 3.
Upregulated Alzheimer's disease-responsive genes among the top 250 DEG.

Figure 4.
Alzheimer's disease gene distribution in prostate, breast, and colon cancer.
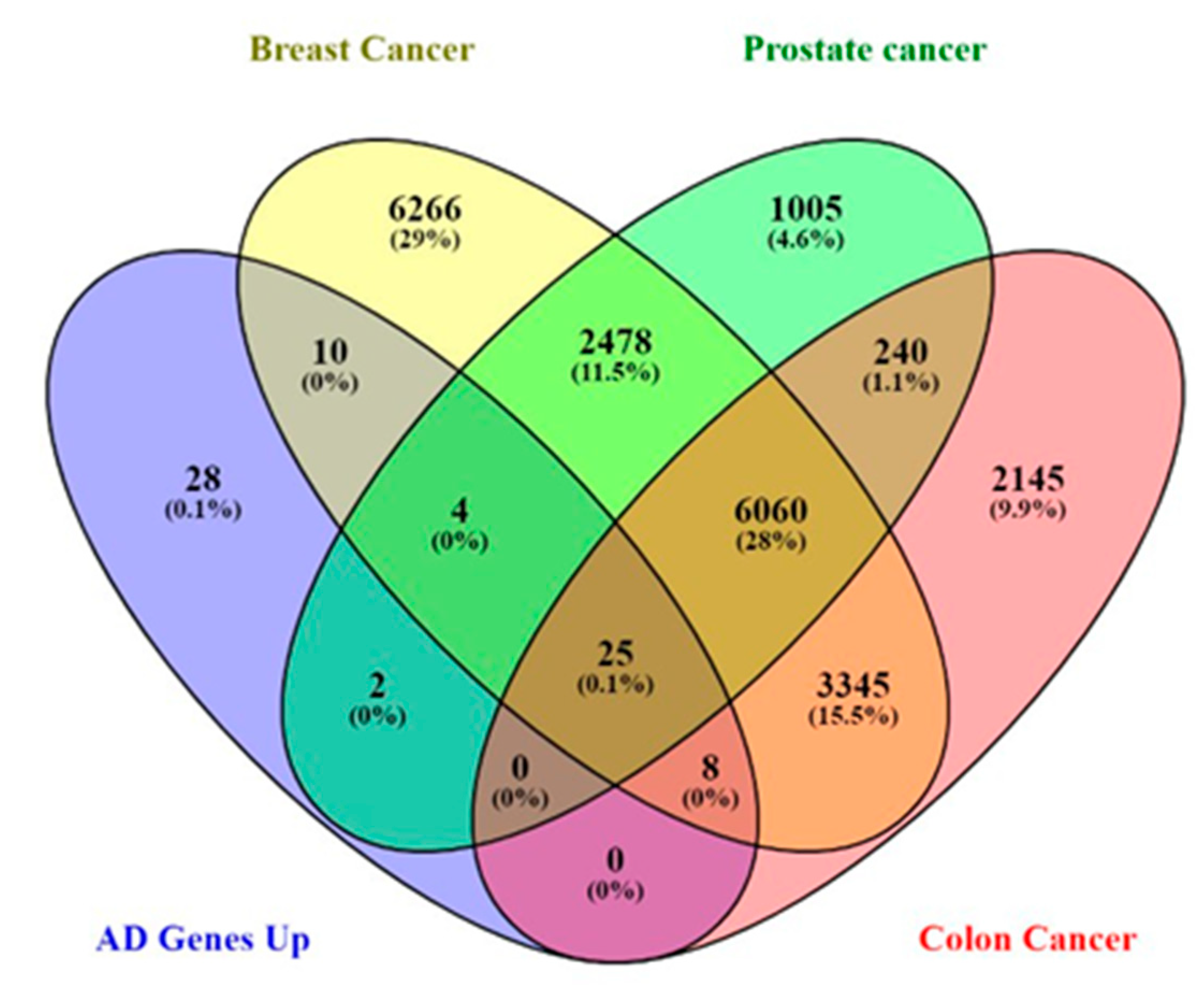
Figure 5.
Expression of Shared Alzheimer's disease upregulated genes in breast, prostate, and colon cancer using GEPIA. Red represents expression in tumor samples; black represents expression in normal samples, BRCA: breast invasive carcinoma, COAD: Colon adenocarcinoma, PRAD: Prostate adenocarcinoma: tumor, N: normal, * p-value < 0.01, |Log2FC| > 1.
Figure 5.
Expression of Shared Alzheimer's disease upregulated genes in breast, prostate, and colon cancer using GEPIA. Red represents expression in tumor samples; black represents expression in normal samples, BRCA: breast invasive carcinoma, COAD: Colon adenocarcinoma, PRAD: Prostate adenocarcinoma: tumor, N: normal, * p-value < 0.01, |Log2FC| > 1.
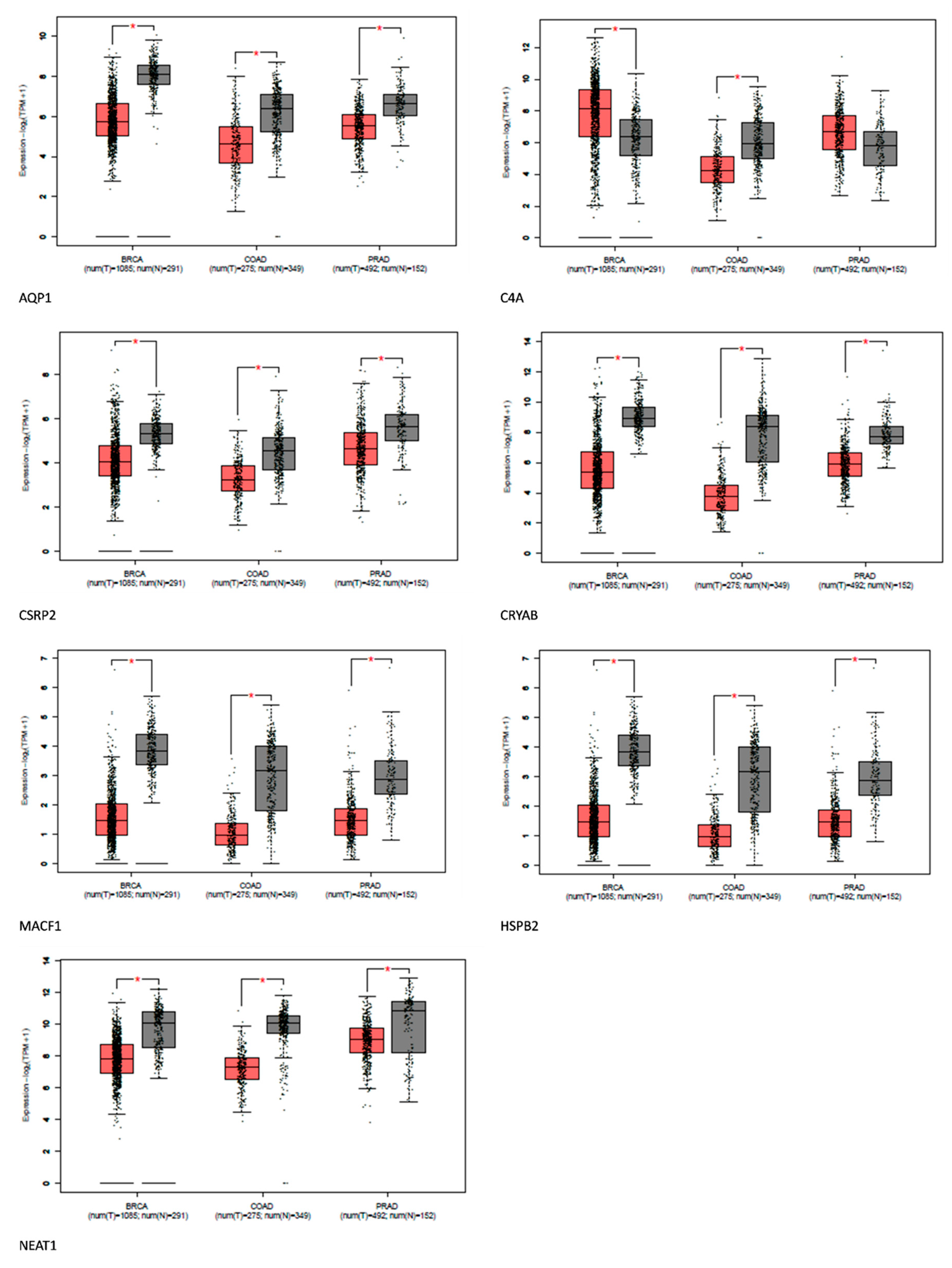
Figure 6.
Expression of AD-exclusive upregulated genes in breast, prostate, and colon cancer using GEPIA. Red represents expression in tumor samples; black represents expression in normal samples, BRCA: breast invasive carcinoma, COAD: Colon adenocarcinoma, PRAD: Prostate adenocarcinoma: tumor, N: normal, * p-value < 0.01, |Log2FC| > 1.
Figure 6.
Expression of AD-exclusive upregulated genes in breast, prostate, and colon cancer using GEPIA. Red represents expression in tumor samples; black represents expression in normal samples, BRCA: breast invasive carcinoma, COAD: Colon adenocarcinoma, PRAD: Prostate adenocarcinoma: tumor, N: normal, * p-value < 0.01, |Log2FC| > 1.
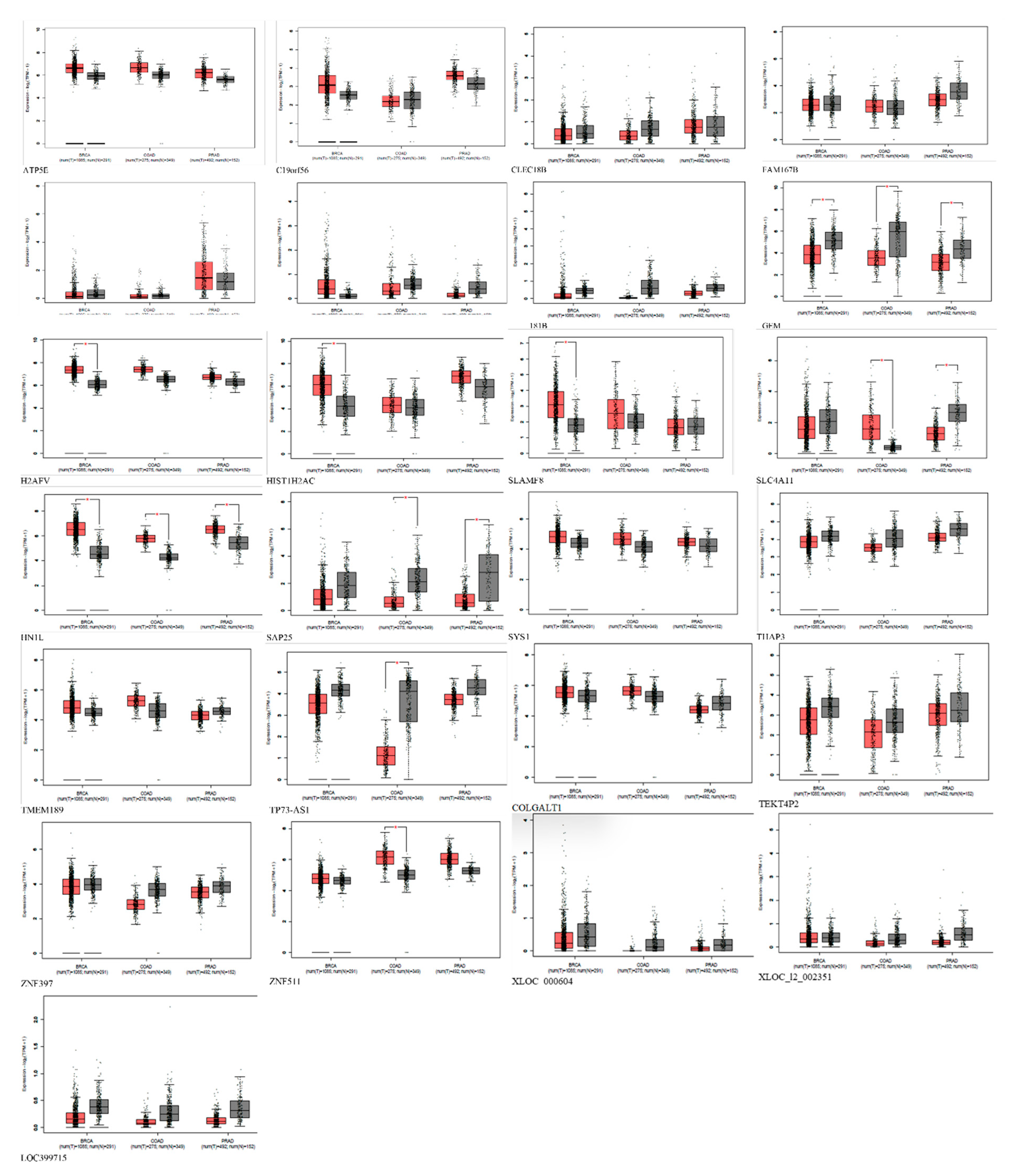
Figure 7.
Gene Ontology Biological Process. Biological process of C1orf56 (Figure 4A), biological process of HIST1H2AC (Figure 4B).
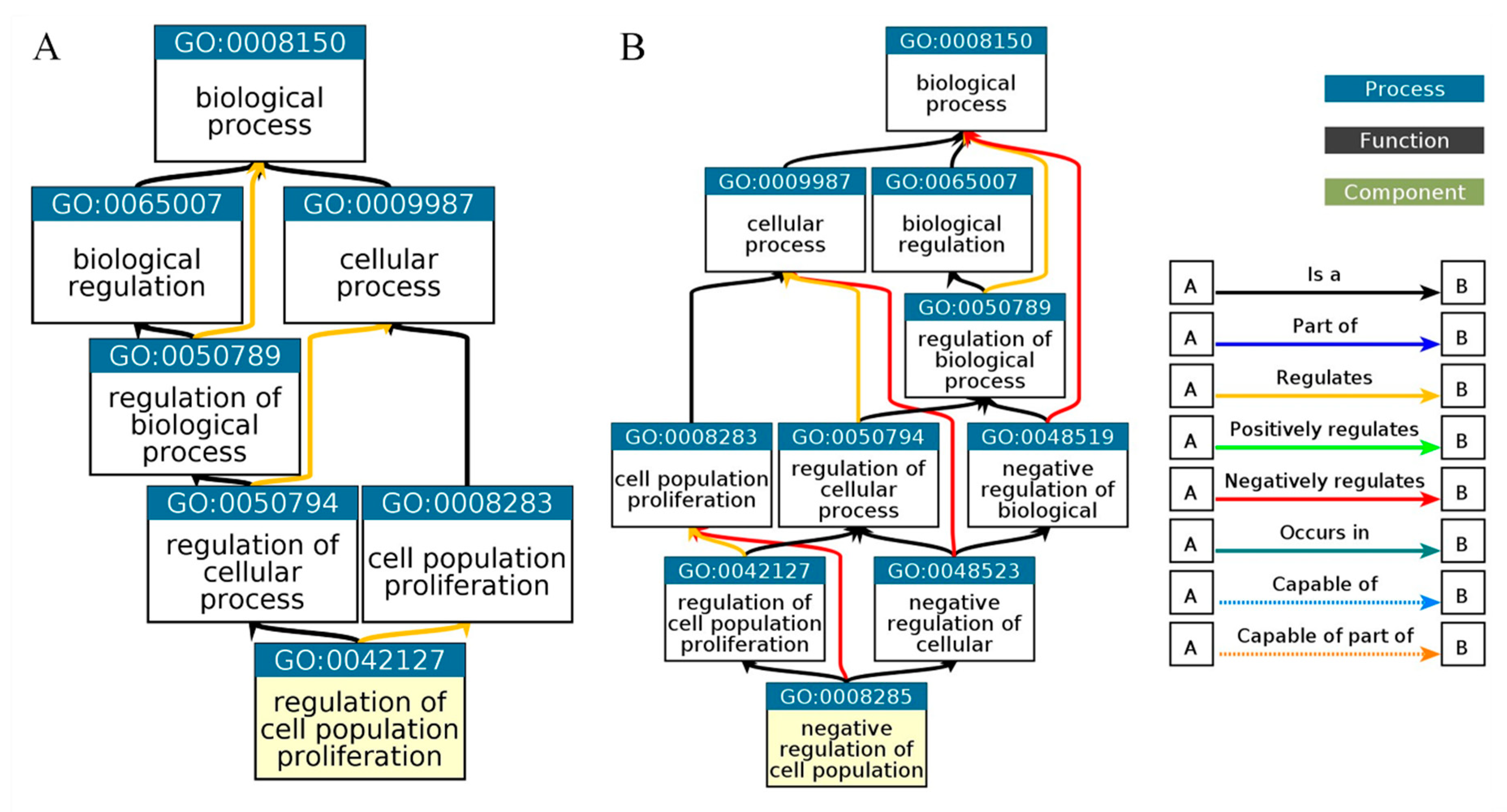
Figure 8.
Enrichment pathway for WebGestalt KEGG. Necroptosis was identified as a pathway for HIST1H2AC.
Figure 8.
Enrichment pathway for WebGestalt KEGG. Necroptosis was identified as a pathway for HIST1H2AC.
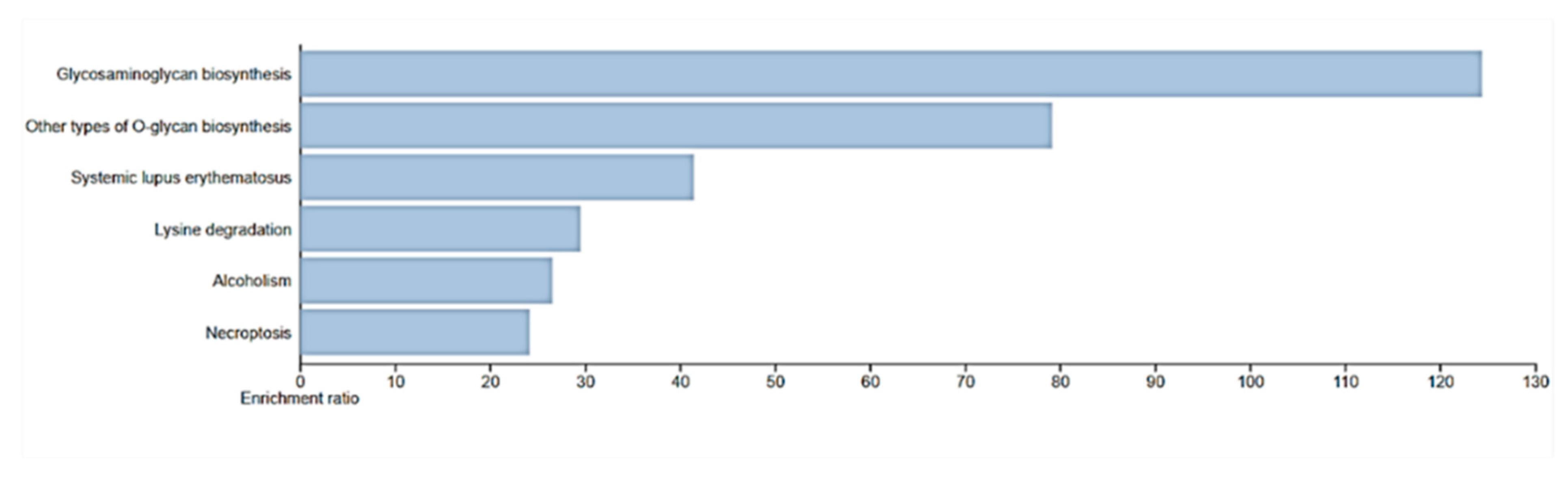
Figure 9.
Survival Analysis of C1orf56 Expression in Breast, prostate, and Colorectal cancer. 1.1, 1.2 and 0.91 hazard ratio (HR) was determined for breast, prostate, and colorectal cancer respectively. A: BRCA- Breast cancer, B: PRAD- Prostate Cancer, C: COAD- Colorectal Cancer.
Figure 9.
Survival Analysis of C1orf56 Expression in Breast, prostate, and Colorectal cancer. 1.1, 1.2 and 0.91 hazard ratio (HR) was determined for breast, prostate, and colorectal cancer respectively. A: BRCA- Breast cancer, B: PRAD- Prostate Cancer, C: COAD- Colorectal Cancer.
Figure 10.
Survival Analysis of HIST1H2AC Expression in Breast, prostate and Colorectal cancer.0.84, 0.53 and 1.3 Hazard ratio was determined for breast, prostate and colorectal cancer respectively. A: BRCA- Breast cancer, B: PRAD- Prostate Cancer, C: COAD- Colorectal Cancer.
Figure 10.
Survival Analysis of HIST1H2AC Expression in Breast, prostate and Colorectal cancer.0.84, 0.53 and 1.3 Hazard ratio was determined for breast, prostate and colorectal cancer respectively. A: BRCA- Breast cancer, B: PRAD- Prostate Cancer, C: COAD- Colorectal Cancer.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



