
Submitted:
08 December 2023
Posted:
08 December 2023
You are already at the latest version
Abstract
This study explored the effects of pBD114 (porcine β-defensin 114) on inflammatory response. The results in RAW264.7 cells showed that 100 μg/mL of pBD114 significantly increased the concentrations of TNFα and IL-10, and 10 μg/mL of pBD114 significantly decreased the concen-tration of TNFα in LPS-induced RAW264.7 cell cultures at 10 ng/mL. These results suggest that pBD114 can exhibit pro-inflammatory or anti-inflammatory activities under certain conditions. RNA-seq analysis was performed to gain further insight into the effects of pBD114 on the in-flammatory response. Among pBD114 promoting RAW264.7 pro-inflammatory response, pBD114 significantly up-regulated 1170 genes and down-regulated 724 genes. KEGG enrichment found that the DEGs were significantly enriched in immune- and signal-transduction-related signaling pathways. PPI and KDA analyses revealed that Bcl10 and Bcl3 were the key genes. In addition, pBD114 significantly up-regulated 12 genes and down-regulated 38 genes in the an-ti-inflammatory response. KEGG enrichment analysis revealed that the DEGs were mainly en-riched in the "Cytokine-cytokine receptor interaction" signaling pathway, and PPI and KDA analyses showed that Stat1 and Csf2 were the key genes. The results of qRT-PCR verified those of RNA-seq. In vivo mice tests also confirmed the pro- or anti-inflammatory activity of pBD114. Although the inflammatory response is a rapid and complex physiological reaction to noxious stimuli, this study found that pBD114 plays an essential role in the inflammatory response mainly by acting on genes related to immunity, signal transduction, signaling molecules, and interactions.
Keywords:
defensins
; lipopolysaccharide
; RNA-seq
; RAW264.7
; inflammation
; mice
1. Introduction
One of the most important medical discoveries of the past two decades has been that the immune system and inflammatory processes are involved in not just a few select disorders but a wide variety of mental and physical health problems that dominate present-day morbidity and mortality worldwide[1,2,3,4]. Inflammation is part of the innate defense mechanism of the body against infectious or non-infectious etiologies [5]. A normal inflammatory response is characterized by the temporally restricted upregulation of inflammatory activity that occurs when a threat is present and that resolves once the threat has passed [6,7,8]. However, the presence of specific social, psychological, environmental, and biological factors has been linked to the prevention of resolution of acute inflammation and, in turn, the promotion of a state of low-grade, non-infective (that is, ‘sterile’) systemic chronic inflammation (SCI). When it becomes SCI, it can cause a breakdown of immune tolerance and lead to significant alterations in all tissues and organs, as well as normal cellular physiology, which can increase the risk for various non-communicable diseases in young and older individuals [8,9,10]. In humans, SCI has been linked to diseases such as cancer, heart disease, diabetes, arthritis, depression, and Alzheimer’s [11]. Similarly, increased pro-inflammatory mediators have been associated with enhanced resistance to a range of important poultry and pig pathogens in livestock animals. However, inflammation may have undesirable consequences, including potentially exacerbating tissue damage and diverting nutrients from productive purposes [12,13,14]. These detrimental effects underscore the necessity for a delicate balance in the inflammatory response - a balance that, when disrupted, can have severe implications for human and animal health.
Defensins are a family of cationic antimicrobial peptides active against various infectious microbes, including bacteria, viruses, and fungi, playing important roles as innate effectors and immune modulators in the immunological control of microbial infection [15,16,17]. They function primarily as antimicrobial peptides, eliminating potential pathogens by disrupting their cellular membranes [18]. However, their role extends beyond direct antimicrobial activity [19,20]. Defensins display substantial immunomodulatory properties in vitro and in vivo, and these features are becoming increasingly appreciated in the literature [16,21,22,23]. The immune response is a highly complex process involving multiple interconnected signaling pathways. Defensins influence the entire signaling network of the immune response, and, as a result, their effects on biological processes are also complex. The ability of defensins to influence many different cell types and pathways has implications for various immune-associated diseases [24,25,26,27,28,29,30,31]. Defensins have been described as endogenous alarmins, alerting the body to danger and responding to inflammatory signals by promoting both local innate and adaptive systemic immune responses. Lu et al. found that HD-5 up-regulates the expression of genes involved in cell survival and inflammation in an NF-kB-dependent fashion in epithelial cells. Further, they find that HD-5 has pro-apoptotic effects on intestinal epithelial cells as well as primary CD4+ T cells [32]. Similarly, hBD-2 via TLR-4 leads to the maturation of dendritic cells (DCs), which consequently exhibit Th1-polarized responses, such as the production of pro-inflammatory cytokines IL-12, IL-1α, IL-1β, and IL-6, which may counter suppressive action of microbial factors by generating more robust host inflammatory and Th1 responses [33]. HDPs efficiently attract Th17 (T helper 17 cells), which in turn secrete pro-inflammatory cytokines, IL-17A, IL-17F, IL-21, and IL-22, responsible for mounting mucosal defense against pathogenic microbes in the respiratory or intestinal tract [34]. For instance, the pro- and anti-inflammatory effects of cathelicidin LL-37 are concentration-dependent, i.e., the former is visible at >20 µg/mL, whereas the latter is at 1–5 µg/mL [35]. However, there is increasing evidence that they exert variable control on the response to danger, creating a dichotomous response that can suppress inflammation in some circumstances but exacerbate the response to danger and damage in others and, at higher levels, lead to a cytotoxic effect [36]. Koeninger et al. found that hBD2 improves disease activity indices and prevents colitis-associated weight loss in three mouse models (dextran sodium sulfate (DSS), 2,4,6-Trinitrobenzenesulfonic acid (TNBS) and T-cell transfer into immunodeficient recipient mice). Furthermore, they found that hBD2 engages with CCR2 on DCs to inhibit NF-κB activity and to promote CREB phosphorylation, thus reducing the expression of inflammatory factors [37]. Our previous studies showed that pBD114, a porcine β-defensin, endotoxin-induced inflammation and apoptosis in IECs through downregulation of two critical inflammation-associated signaling proteins, NF-kappa-B inhibitor alpha (IkB-α) and extracellular signal-regulated kinase1/2 (ERK1/2), suppresses inflammation and IEC apoptosis in mice exposed to bacterial endotoxins [38]. Similarly, pBD2 and pBD129 also exhibit anti-inflammatory activity [39,40,41]. Hence, defensins are significantly interested in understanding and controlling the inflammatory response.
Porcine defensins have been shown to have antibacterial, antiviral, antioxidant, intestinal barrier maintenance, and immunomodulatory effects [42,43,44,45,46], but it is not clear whether porcine defensins possess the ability to activate inflammatory responses. Our previous studies have shown that pBD114 modulates inflammation, barrier, and apoptosis in porcine intestinal epithelial cells [38]. Studies have shown that defensins’ functions seem to depend on the environmental stimuli, cell and tissue type, interaction with different cellular receptors, and the concentration of the peptides [47]. We supposed that pBD114 plays different inflammatory regulatory roles under different environmental stimuli. Therefore, in the present study, we investigated the effects of pBD114 on the inflammatory responses by ex vivo assays and transcriptome. Our study contributes to understand the function and mechanism of porcine defensins in balancing inflammation in host cells.
2. Results
2.1. Preparation and Basic Biological Characteristics of pBD114
The SignalP-5.0 online analysis of the signal peptide cutting site shows in Figure S1A that there is a significant signal peptide cutting site between the 22nd and 23rd amino acids. Hence, the mature peptide of pBD114 is 47 amino acids. The mature peptide of pBD114 was synthesized by artificial chemical solid-phase synthesis technology, and the purity was > 95% by high-performance liquid chromatography (Figure S1B,C), which could be applied to subsequent cell and mice assays. The antimicrobial activity of pBD114 against Staphylococcus aureus, Enterococcus faecalis, Escherichia coli, Pseudomonas aeruginosa, Salmonella typhimurium, and Klebsiella pneumoniae was explored by minimum inhibitory concentration (MIC). The MIC values of all were more than 128 μg/mL (Table S1), indicating that the antibacterial activity of pBD114 was low. The hemolytic test conducted with 4% rabbit red blood cell suspension showed that when the concentration of pBD114 was increased by 256 μg/mL, the hemolytic rate was still lower than 5%(Figure 1A), indicating that pBD114 had lower hemolytic and better safety. RAW264.7 was also used to examine the cytotoxicity of pBD114, and cell viability was measured by CCK-8 after treatment with different concentrations of pBD114. There was no cytotoxicity (Figure 1B) when the concentration of pBD114 was no more than 256 μg/mL. These results suggested that pBD114 was safe under a concentration of no more than 256 μg/mL.
2.2. Proinflammatory and Anti-Inflammatory Activity of pBD114 on Mouse Mononuclear Macrophage RAW264.7
Our previous study indicated that pBD114 was essential in maintaining intestinal epithelium homeostasis in response to various infections or diseases [38]. However, the effects of pBD114 on the inflammatory response to macrophage remains unclear. As shown in Figure 2, 100 μg/mL pBD114 co-cultured with RAW264.7 cells for 3 (Figure 2B), 6 (Figure 2C), and 12 h (Figure 2D) significantly increased the concentration of TNFα in the culture medium (P < 0.05). The concentration of IL-10 in the culture medium was increased considerably after co-culture of 100 μg/mL pBD114 with RAW264.7 cells for 6 (Figure 2G) and 12 h (Figure 2H) (P < 0.05). As shown in Figure 3, 100 and 1000 ng/mL LPS treated for 1 h (Figure 3B) could induce inflammation in mouse mononuclear macrophage RAW264.7, and significantly increase the protein concentration of inflammatory factor TNF-α in cell culture medium (P<0.05). Treatment with lipopolysaccharide (LPS) of 10, 100, and 1000 ng/mL for 2 (Figure 3C) and 4 h (Figure 3C) could significantly induce the protein concentration of inflammatory factor TNF-α in RAW264.7 cell culture medium of mouse mononuclear macrophages (P<0.05). Therefore, in the follow-up experiment, the immune activation of RAW264.7 cells was treated with 10 ng/mL LPS for 2 h. To study the inhibitory effect of pBD114 on the immune activation of RAW264.7 cells, In this study, RAW264.7 cells were co-cultured with 1, 5, 10, 20 and 50 μg/mL pBD114 and 10 ng/mL LPS for 2 h. The results indicated that 5 and 10 μg/mL pBD114 (Figure 3E) significantly inhibited the production of inflammatory cytokine TNF-α (P<0.05). The results showed that pBD114 (5 and 10 μg/mL) showed immunosuppressive activity in RAW264.7 mouse mononuclear macrophage activated by LPS.
2.3. RNA-seq Analyzed the Mechanism of pBD114 on Inflammation of Mouse Mononuclear Macrophage RAW264.7
The effects and mechanism of pBD114 on the inflammatory response to RAW246.7 were analyzed in depth by RNA-seq. 16 samples were measured using the DNBSEQ platform, with an average output of 1.17G data per sample. The average matching rate of the samples against the genome was 90.06%, and the average matching rate against the gene set was 84.86%; a total of 18,126 genes were detected. After quality filtering and trimming, 21.20 to 23.86 M clean reads were obtained, and the percentage of Q20 and Q30 base was over 97.40% and 92.22 (Table 1). The obtained clear reads were mapped with the reference genome (Genome ref. number: GCF_000001635.26_GRCm38.p6) using HISAT2 software, and the mapping rate was all higher than 88.51% (Table S2).
2.3.1. Differentially Expressed Genes
Prior to the analysis of differentially expressed genes (DEGs), a principal component (PCA) analysis was performed. PCA plots showed significant differences between the four groups (Figure 4A). From the Volcano plots of PBD114/MOCK (Figure 4B), there are 1,894 DEGs, including 1,170 up-regulated genes and 724 down-regulated genes. From the Volcano plots of PBD114LPS/LPS (Figure 4C), there are 50 DEGs, including 12 up-regulated genes and 38 down-regulated genes. The detailed data of DEGs were shown in Table S3.
2.3.2. KEGG Enrichment Analysis
KEGG enrichment is based on KEGG Pathway annotation classification, functional classification of differential genes, and enrichment analysis using the phyper function in R software. By analyzing the KEGG pathway enrichment of DEGs of PBD114/MOCK, a total of 329 signaling pathways were enriched by KEGG pathway enrichment analysis, of which 95 KEGG pathways were significantly enriched (Q ≤ 0.05). The analysis of the first 20 enriched pathways showed the presence of three immune-related pathways, the Toll-like receptor signaling pathway, C-type lectin receptor signaling pathway, and IL-17 signaling pathway. It is noted that the C-type lectin receptor signaling pathway with the most significant number of DEGs was being enriched (Figure 5A). At the same time, the analysis of the first 20 enriched pathways showed the presence of four signal transduction-related pathways, the TNF signaling pathway, MAPK signaling pathway, NF-kappa B signaling pathway and MAPK signaling pathway-fly, it is noted that the MAPK signaling pathway with the most significant number of DEGs was being enriched (Figure 5A). The DEGs of PBD114LPS/LPS were analyzed by KEGG pathway enrichment, and a total of 117 signaling pathways were enriched, of which 30 KEGG pathways were significantly enriched (Q ≤ 0.05). The top 20 pathways with the most significant enrichment related to immunomodulation were selected and presented in the form of bubble plots (Figure 5B). Among them, Hematopoietic cell lineage, Th17 cell differentiation, Th1 and Th2 cell differentiation, IL-17 signaling pathway, Toll-like receptor signaling pathway, and Chemokine signaling pathway related immune-system, it is noted that the Hematopoietic cell lineage with the largest number of DEGs was being enriched. Cytokine-cytokine receptor interaction, Viral protein interaction with cytokine and cytokine receptor, Jak-STAT signaling pathway, NF-kappa B signaling pathway, Chemokine signaling pathway, and TNF signaling pathway related signal transduction, it is noted that the Cytokine-cytokine receptor interaction” with the largest number of DEGs was being enriched.
2.3.3. PPI and KDA Analysis
PPI and KDA analysis were performed to further explore the key genes of pBD114 regulating RAW264.7 inflammatory response. 303 DEGs significantly enriched by PBD114/MOCK into 28 signaling pathways related to immunity and signaling were selected for analysis, and the PPI results showed that Tnf, Kras, Jun, Src, and Mapk14 had the most interactions with other proteins (Figure 6A). Further KDA analysis identified Bcl10, Bcl3, Cflar, Il1a, Dusp1, Traf1, Map2k7, Map3k8, Irf5, and Ikbke as key driver genes (Figure 6C). These DEGs were predominantly enriched in the NF-kappa B signaling pathway, C-type lectin receptor signaling pathway, MAPK signaling pathway, and Toll-like receptor signaling pathway (Table S4). Since there were only 50 DEGs in PBD114LPS/LPS, we subjected all DEGs to PPI and DEGs analysis. The results showed that Stat1 and Cxcl10 had the most interactions with other proteins (Figure 6B). Further KDA analyses showed that Stat1, Csf2, and Il1b are key driver genes (Figure 6D ). These DEGs were mainly enriched in the C-type lectin receptor signaling pathway, MAPK signaling pathway, and Chemokine signaling pathway Table S4).
2.3.4. Validation of RNA-Seq Results via Quantitative PCR Analysis
2.4. Modulation of Inflammatory Responses in Mice by pBD114
The in vitro results of this study showed that pBD114 was able to modulate the inflammatory response of mouse mononuclear macrophage RAW264.7. The present study investigated the effects of pBD114 on the inflammatory response in mice by intraperitoneal injection of pBD114 and LPS administered for 12 h, to verify whether pBD114 could perform the same biological function in vivo. As shown in Figure 7, the weight of the mice was not affected by pBD114 and LPS (Figure 7A), but LPS significantly increased the weight of the spleen (Figure 7B) and spleen index (Figure 7C) (P < 0.05). LPS significantly increased the concentrations of TNFα, IL-6, IL-1β, and IL-10 proteins (P < 0.05) in mice serum (Figure 7D–G), pBD114 also significantly increased (P < 0.05) the concentrations of TNFα, IL-6 and IL-10 proteins in mice serum (Figure 7D,E,G), and significantly decreased the LPS-induced concentrations of IL-1β protein in mice serum (Figure 7F) (P < 0.05).
The expression of inflammation-related genes in mouse spleen was measured to investigate the effects of pBD114 on the inflammation response in mice. As shown in Figure 8, targeting mouse spleen pro-inflammatory and anti-inflammatory factor-related genes, LPS significantly increased the mRNA levels of TNFα, IL-1β and IL-10 and significantly decreased the mRNA levels of TGF-β (P < 0.05), while pBD114 also considerably increased the mRNA levels of TNFα, IL-1β, TGF-β and IL-10 (P < 0.05), and at the same time pBD114 also significantly reduced the LPS-induced increase in mRNA levels of IL-1β and IL-10 (P < 0.05). For mouse spleen chemokine-related genes, LPS significantly increased the mRNA levels of COX-2, CXCL-10 and MCP-1 (P < 0.05), while pBD114 significantly increased the mRNA levels of CXCL-10 and MCP-1 (P < 0.05), meanwhile, pBD114 also significantly decreased LPS-induced increases in the COX-2 and CXCL-10 mRNA levels (P < 0.05) and significantly increased the LPS-induced decrease in mRNA levels of csf2 (P < 0.05). Targeting inflammatory response-associated signaling pathway receptors, LPS significantly decreased the mRNA levels of CD4 and IL2Rb (P < 0.05), and pBD114 significantly decreased the mRNA levels of CD4 and significantly increased the mRNA levels of TLR-4 (P < 0.05). Targeting inflammatory response-related transcription factor-related genes, LPS significantly decreased the mRNA levels of RELA, p38, and JNK9 (P < 0.05) and significantly increased the mRNA level of JUN, and pBD114 significantly increased the mRNA of p38, JUN, and FOS (P < 0.05), as well as significantly increased the mRNA level of LPS-induced p38 (P < 0.05).
3. Discussion
Analysis of the porcine genome using bioinformatics methods in 2006 led to the first discovery of pBD114, which is expressed in the pig’s intestines, liver, spleen, lungs, and male reproductive tract. The fact that pBD114 is expressed in several pig tissues is a harbinger of a diverse range of biological functions of pBD114. Our previous study utilized recombinant pBD114 obtained from the E. coli expression system to have weak activity against E. coli DH5α [48]. In this study, pBD114 was prepared by chemical solid phase synthesis with >95% purity. Again, only weak antimicrobial activity was observed, with MIC values of 128 μg/mL against Enterococcus faecalis and Salmonella typhimurium, while all other indicator bacteria were >256 μg/mL. Increasing evidence indicates that the direct bactericidal activity of defensins in regulating the antibacterial immune response is not the only essential role of defensins in regulating host immune homeostasis [18]. The lack of anti-microbial activity of pBD114 may be compensated by immunomodulation, and some defensins have been shown to have such biological properties [49,50]. Similar to other defensins [39,48,51,52], pBD114 was not hemolytic or cytotoxic, which is also in agreement with the results of our previous study [48].
Researches on porcine defensins has shifted from anti-microbial activity to immunomodulation in recent years. In particular, many studies have been conducted on porcine β-defensin 2 (pBD2), showing that pBD2 has antioxidant, intestinal barrier-enhancing, and anti-inflammatory activities [40,41,43,53,54]. Our previous study found that pBD114 can also inhibit the expression of inflammatory factors and alleviate the inflammatory injury of intestinal epithelial cells in mice caused by LPS [2]. However, whether porcine β-defensins, like human defensins, can also activate inflammatory responses? Macrophages play a critical role in initiating, maintaining, and resolving inflammation [55,56]. The famous murine macrophage cell line, RAW 264.7, is often used to initially screen natural products for bioactivity and predict their potential effect in vivo or on primary cells. The cell line response is used to evaluate the effective bioactivity of the product [57]. Therefore, RAW264.7 is used extensively to carry out in vitro screens for immunomodulators [58]. In contrast, the biological functions of defensins are related to the environment and cell type in which they are found [47]. Therefore, we used mouse monocyte macrophage RAW264.7 to investigate the effects of pBD114 on the inflammatory responses in different physiological environments. Our results showed that under the normal physiological state of mouse monocyte macrophage RAW264.7, pBD114 significantly increased TNFα and IL-10 with a growing concentration of pBD114 and prolongation of treatment time, activated in activating the inflammatory response. However, when the inflammatory response of mouse monocyte macrophage RAW264.7 was activated by LPS and then given pBD114 treatment, pBD114 surprisingly again showed inhibition of the inflammatory response, and the concentration of anti-inflammatory active pBD114 was much lower than that of pro-inflammatory active pBD114. The results were in agreement with that of the study on LL-37, pro- and anti-inflammatory effects of cathelicidin LL-37 are concentration-dependent, i.e., the former is visible at >20 µg/mL, whereas the latter at 1–5 µg/mL [35]. The human cathelicidin LL 37 is transcribed from the CAMP gene in various cell types, including epithelial cells and many immune system cells. The effects of LL 37 are widespread, as this HDP is known to elicit a wide range of responses in a broad assortment of cell types. Similar to LL-37, pBD114 is also widely expressed in the intestine, liver, spleen, lung, and male reproductive tract of pigs[59], indicating that the effects of pBD114 are also widespread.
Previous studies have shown that defensins exert pro-inflammatory and anti-inflammatory activities through signaling pathways such as NF-κB and MAPK [60,61,62]. However, their studies lacked a comprehensive and systematic study of defensins’ pro-inflammatory and anti-inflammatory activities. RNA-seq is a technique to study the transcription and transcriptional regulation laws of all genes in a cell at the overall level, which can comprehensively and rapidly obtain the sequence information and expression information of almost all transcripts of a specific cell or tissue in a certain state, to accurately analyze the differences in gene expression, gene structure variability, analysis of the interaction network between RNAs, and other important issues in life sciences [63]. Therefore, in this study, the effects of pBD114 on the inflammatory response of mouse monocyte macrophage RAW264.7 were analyzed by RNA-seq. The results showed that pBD114 significantly altered the expression of 1894 genes, of which 117 were up-regulated, and 724 were down-regulated, in RAW264.7 under normal physiological state. The differentially expressed genes were mainly enriched in the TNF signaling pathway, MAPK signaling pathway, and NF-kappa B signaling pathway which have been identified as pivotal regulators of the initiation and resolution of inflammation [64,65]. The results suggested that pBD114 activates the inflammatory response of RAW264.7 primarily through TNF, MAPK, and NF-kappa B signaling pathways, which is consistent with previous studies’ results [60,61,62]. PPI analysis showed that Tnf, Kras, Jun, Src, and Mapk14 had the most interactions with other proteins, and KDA analysis showed that Bcl10, Bcl3, Cflar, Il1a, Dusp1, Traf1, Map2k7, Map3k8, Irf5 and Ikbke played key driver genes in the activation of the inflammatory response of RAW264.7 by pBD114. Bcl3 and Traf1 are mainly involved in the TNF signaling pathway, which participates in negative intracellular signaling [66,67]. Cflar and Bcl10 are primarily involved in the NF-kappa B signaling pathway, which participated in the B cell receptor signaling pathway and cell survival [68,69]. Map2k7, Map3k8, Dusp1, and Il1a are mainly involved in the MAPK signaling pathway, which activated ERK and JUK to promote proliferation, differentiation, and inflammation [70,71,72,73]. Irf5 and Ikbke are mainly involved in the MyD88-independent pathway, which takes part in chemotactic and antiviral effects [74,75]. The above results indicate that pBD114 activates the inflammatory response through multiple signaling pathways and key genes such as TNF, MAPK, and NF-kappa B signaling pathways.
The effects of pBD114 on the inflammatory response of LPS-activated mouse monocyte macrophage RAW264.7 were analyzed by RNA-seq. However, pBD114 altered only 50 genes (up-regulated by 12 and down-regulated by 38) in RAW264.7 with inflammatory response activated by LPS. KEGG enrichment analysis showed that these genes were mainly enriched in the Cytokine-cytokine receptor interaction, Viral protein interaction with cytokine and cytokine receptor, Hematopoietic cell lineage and Th17 cell differentiation. The PPI analysis shows that Stat1 and Cxcl10 had the most interactions with other proteins. Further KDA analyses showed that Stat1, Csf2, and Il1b play the role of key driver genes. These results suggest that the inhibition of RAW264.7-activated inflammatory responses by pBD114 is not as strong, unlike the findings for HBD3 and pBD2. There was a substantial effect of HBD3 on TLR4 activation — 5,494 genes compared with 1,779 genes that were differentially expressed between macrophages stimulated with KLA or KLA and hBD3, respectively [76]. Similarly, there were 812 DEGs between the E. coli and E. coli +pBD2 groups, including 431 significantly upregulated genes and 381 significantly downregulated genes [53].
To further verify the effects of pBD114 on inflammatory response, we carried out an in vivo assay of the effects pBD114 on inflammatory response in mice. The results showed that pBD114 could activate the inflammatory response in mice and inhibit the inflammatory response to some extent after LPS induced the inflammatory response in mice, which were consistent with the results of studies related to human defensins and porcine defensins [37,60,77]. As shown by previous findings and our own, the effect of defensins on the inflammatory response goes far beyond simply acting as an inflammatory response modulator via a singular receptor or linear signaling. It has a multifaceted effect on multiple pathways in the cell and the organism, and the effect varies in different environments. Therefore, determining the mechanisms of defensin action on cells or organisms in characteristic environments is essential for revealing defensin function.
4. Materials and Methods
4.1. Chemical Solid Phase Synthesis of pBD114
The amino acid sequence of the pBD114 precursor protein was obtained from NCBI entry number NP_001123445.1, and SignalP-5.0 analyzed the signal peptide of the pBD114 precursor protein. The amino acid sequence of the pBD114 mature peptide was obtained by removing the signal peptide, and pBD114 was prepared by GL Biochem (Shanghai)Ltd. (Shanghai, China) with chemical solid phase synthesis.
4.2. Minimal Inhibitory Concentration
The microtiter broth dilution method measured the minimal inhibitory concentration (MIC) of pBD114 [78]. Staphylococcus aureus ATCC 29213, Staphylococcus aureus ATCC 43300, Enterococcus faecalis ATCC 29212, Escherichia coli ATCC 25922, Pseudomonas aeruginosa ATCC 27853, Salmonella typhimurium ATCC 13311 and Klebsiella pneumoniae ATCC 13883 were grown to 0.4 OD600nm at 37 °C in MH. The target cell culture was diluted to 1×105 CFUs/ml with MH, respectively. 100 μL of pBD114 and 100 μL of cell suspension were added into each well. The activity of pBD114 was tested over a concentration range of 256, 128, 64, 32, 16, 8, 4, 2, 1, 0.5, and 0.25 μg/mL, and all assays were tested in triplicated. Bacterial plates were incubated at 37 °C for 16 h, and the absorption of cell culture was recorded at 600 nm. MIC was defined as the lowest concentration of peptide at which there was no change in optical density.
4.3. Hemolytic Activity Assay
The hemolytic activity of pBD114 was measured spectrophotometrically using a hemoglobin release assay [38]. Freshly pig blood was collected to prepare erythrocytes by centrifugation at 1,500 rpm for 10 min at room temperature. The erythrocytes were washed three times warmly with PBS (pH 7.2) and resuspended at 4% (v/v) in PBS (pH 7.2). 4% erythrocytes were incubated with 1, 4, 16, 64, and 256 μg/mL pBD114 for 1 h at 37℃ and then centrifuged at 1,000 rpm for 5 min. 3 mL supernatant was added to a 2 cm quartz cuvette, and the absorbance was measured at 414 nm with a UV-1100 spectrophotometer (Shanghai, China). No hemolysis and 100% hemolysis were determined in PBS and Triton X-100, respectively.
4.4. Cell Culture and Treatment
Mouse mononuclear macrophage RAW264.7 cells were cultured in 75 cm2 cell culture flask in DMEM-F12 with 10% FBS, 100 U/mL penicillin, and 100 μg/mL streptomycin. Cells were cultured in an incubator with sufficient humidity at 37 ◦C and 5% CO2. The effects of pBD114 on the inflammatory response of mouse mononuclear macrophage RAW264.7 were tested as follows:
- 1)
- Cytotoxicity of pBD114: The cells were inoculated into 96-well culture plates and cultured until the fusion reached 50%. The cells were treated with pBD114 at the final concentration of 0, 1, 4, 16, 64, and 256 μg/mL for 24 h. CCK-8 detected the cell activity of pBD114.
- 2)
- The effects of pBD114 on inflammatory response in macrophages under normal physiological conditions: Cells were inoculated into 24-well culture plates and cultured until the fusion reached 80%, then treated with 10 and 100 μg/mL pBD114 for 1, 3, 6 and 12 h. Cell culture medium was collected to detect TNF-α (R&D Systems,DY410) and IL-10 (R&D Systems,M1000B) protein concentration by ELISA.
- 3)
- Optimization of LPS treatment time and concentration: Cells were inoculated into a 24-well culture plate and cultured until the fusion reached 80%, then treated with 1, 5, 10, 20 μg/mL LPS and 50 μg/mL pBD114f for 0.5, 1, 2 and 4 h, cell culture medium was collected, and TNF-α protein concentration was detected by ELISA.
- 4)
- The effects of pBD114 on inflammatory response in macrophages under inflammatory activation: Cells were inoculated into a 24-well culture plate and cultured until the fusion reached 80%, then treated with 1, 5, 10, 20, and 50 μg/mL pBD114 and 10 ng/mL LPS for 2 h, cell culture medium was collected, and TNF-α protein concentration was detected by ELISA.
- 5)
- Samples of RNA-seq: Mouse mononuclear macrophage RAW264.7 were inoculated into a 6-well plate and cultured until the fusion reached 80%. The cells were treated with or without pBD114 for 12 h, and 10 ng/mL LPS-induced cells were simultaneously treated with or without 10 μg/mL pBD114 for 2 h. The concentration of TNF-α and IL-10 protein in the culture solution was detected by ELISA. The cells were collected for RNA-seq.
4.5. RNA-Seq and Data Analysis
Four biological replicates of each sample were submitted to BGI Genomics (The Beijing Genomics Institute) for RNA extraction, library construction, and RNA sequencing. High-throughput sequencing was performed using the DNBSEQ platform (BGI, Shenzhen, China) with 150 bp paired-end reads [79]. Data analysis was conducted using the Dr. Tom system from BGI http://report.bgi.com. Differentially expressed genes were identified based on an adjusted p-value < 0.05 and fold-change > 2. RT-qPCR further validated the results.
4.6. Animal Trials
A mouse assay was performed to investigate the effects of pBD114 on the inflammatory response in animals, a total of 40 C57BL/6 mice (7 weeks) were randomly allocated to four groups: CON, treated without pBD114 and LPS; PBD114, treated with 10 mg/kg pBD114 and without LPS (Sigma-Aldrich); LPS, treated without pBD114 and with 1 mg/kg LPS; PBD114LPS, treated with 1 mg/kg pBD114 and with 1 mg/kg LPS; pBD114 and LPS were intraperitoneally injected via 1 ml insulin syringe (Braun, Melsungen, Germany) within 200 μL PBS. After 12 h, all mice were sacrificed by carbon dioxide anesthesia, and weight, liver, and serum were collected for analysis.
4.7. Quantitative Real-Time PCR Analysis
Total RNA was extracted from the tissues of the spleen using the Trizol reagent (Takara, Takara Biomedical Technology in Beijing. China), followed by reverse transcription using the High Capacity cDNA Archive kit (Takara), according to the manufacturer’s protocols. Real-time PCR was carried out by the ABI 7500 real-time PCR system (Applied Biosystems, Waltham, MA, USA) involving SYBR Green. The primer sequences used for the mRNA determined were listed in Table S6. The relative abundance of a target gene was calculated by the 2-∆∆Ct method.
4.8 Statistical Analysis
Statistical tests were performed using GraphPad Prism 9. Results are shown as the mean ± SE. The statistical significance of data was determined using the T-test (unpaired) or One-way ANOVA and Dunnett’s multiple range test (p < 0.05). In all cases, a p-value < 0.05 was considered to be statistically significant (* p < 0.05, ** p < 0.01, *** p < 0.001).
5. Conclusions
In vitro and in vivo tests have shown that high concentrations of pBD114 activate the inflammatory response, and low concentrations inhibit the activated inflammatory response. RNA-seq analyses indicated that pBD114 activates the inflammatory response through Bcl10, Bcl3, and immune- and signal- transduction-related signaling pathways. pBD114 inhibits the activated inflammatory response through Stat1, Csf2 and “Cytokine-cytokine receptor interaction” signaling pathway. Although the inflammatory response is a rapid and complex physiological reaction to noxious stimuli, this study found that pBD114 plays an important role in the inflammatory response mainly by acting on genes related to immunity, signal transduction, signaling molecules, and interactions.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org, Figure S1: Chemical solid phase synthesis and quality inspection of pBD114; Table S1: Minimum inhibitory concentration value of pBD114 against various indicator bacteria; Table S2: Quality statistics for filtered Reads; Table S3: The results about the comparison of clean reads with reference genome; Table S4: DEGs; Table S5: KDA; Table S6: The primer sequences.
Author Contributions
Conceptualization, Guoqi Su and Jinxiu Huang; Data curation, Guoqi Su and Shan Jiang; Funding acquisition, Guoqi Su and Zuohua Liu; Methodology, Guoqi Su; Project administration, Guoqi Su; Resources, Li Chen; Supervision, Feiyun Yang; Validation, Feiyun Yang and Jinxiu Huang; Visualization, Sheng Huang; Writing – original draft, Guoqi Su; Writing – review & editing, Guoqi Su, Guixue Wang and Jinxiu Huang.
Funding
This research was funded by “Research Institutions Performance Guidance Project, grant number cstc2021jxjl80001” and “National Key Research and Development Plan, grant number 2022YFC2105003”.
Institutional Review Board Statement
The animal study protocol was approved by the Animal Ethics Committee of Chongqing Academy of Animal Science (File number: Cqaa2020010).
Informed Consent Statement
Not applicable.
Data Availability Statement
All data can be requested from authors.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Furman, D.; Chang, J.; Lartigue, L.; Bolen, C.R.; Haddad, F.; Gaudilliere, B.; Ganio, E.A.; Fragiadakis, G.K.; Spitzer, M.H.; Douchet, I.; et al. Expression of specific inflammasome gene modules stratifies older individuals into two extreme clinical and immunological states. Nat Med 2017, 23, 174–184. [Google Scholar] [CrossRef] [PubMed]
- Netea, M.G.; Balkwill, F.; Chonchol, M.; Cominelli, F.; Donath, M.Y.; Giamarellos-Bourboulis, E.J.; Golenbock, D.; Gresnigt, M.S.; Heneka, M.T.; Hoffman, H.M.; et al. A guiding map for inflammation. Nat Immunol 2017, 18, 826–831. [Google Scholar] [CrossRef] [PubMed]
- Slavich, G.M. Understanding inflammation, its regulation, and relevance for health: A top scientific and public priority. Brain Behav Immun 2015, 45, 13–14. [Google Scholar] [CrossRef]
- Bennett, J.M.; Reeves, G.; Billman, G.E.; Sturmberg, J.P. Inflammation–Nature’s Way to Efficiently Respond to All Types of Challenges: Implications for Understanding and Managing “the Epidemic” of Chronic Diseases. Front Med 2018, 5. [Google Scholar] [CrossRef] [PubMed]
- Ferrero-Miliani, L.; Nielsen, O.H.; Andersen, P.S.; Girardin, S.E. Chronic inflammation: Importance of NOD2 and NALP3 in interleukin-1beta generation. Clin Exp Immunol 2007, 147, 227–235. [Google Scholar] [CrossRef] [PubMed]
- Kotas, Maya E.; Medzhitov, R. Homeostasis, Inflammation, and Disease Susceptibility. Cell 2015, 160, 816–827. [CrossRef]
- Straub, R.H. The brain and immune system prompt energy shortage in chronic inflammation and ageing. Nat Rev Rheumatol 2017, 13, 743–751. [Google Scholar] [CrossRef] [PubMed]
- Fullerton, J.N.; Gilroy, D.W. Resolution of inflammation: A new therapeutic frontier. Nat Rev Drug Discov 2016, 15, 551–567. [Google Scholar] [CrossRef] [PubMed]
- Straub, R.H.; Cutolo, M.; Pacifici, R. Evolutionary medicine and bone loss in chronic inflammatory diseases—A theory of inflammation-related osteopenia. Semin Arthritis Rheum 2015, 45, 220–228. [Google Scholar] [CrossRef]
- Miller, A.H.; Raison, C.L. The role of inflammation in depression: From evolutionary imperative to modern treatment target. Nat Rev Immunol 2016, 16, 22–34. [Google Scholar] [CrossRef]
- Furman, D.; Campisi, J.; Verdin, E.; Carrera-Bastos, P.; Targ, S.; Franceschi, C.; Ferrucci, L.; Gilroy, D.W.; Fasano, A.; Miller, G.W.; et al. Chronic inflammation in the etiology of disease across the life span. Nat Med 2019, 25, 1822–1832. [Google Scholar] [CrossRef] [PubMed]
- Broom, L.J.; Kogut, M.H. Inflammation: Friend or foe for animal production? Poult Sci 2018, 97, 510–514. [Google Scholar] [CrossRef] [PubMed]
- Chastant, S.; Saint-Dizier, M. Inflammation: Friend or foe of bovine reproduction? Anim Reprod 2019, 16, 539–547. [Google Scholar] [CrossRef] [PubMed]
- Adams, J.R.G.; Mehat, J.; La Ragione, R.; Behboudi, S. Preventing bacterial disease in poultry in the post-antibiotic era: A case for innate immunity modulation as an alternative to antibiotic use. Front Immunol 2023, 14, 1205869. [Google Scholar] [CrossRef] [PubMed]
- Gao, X.; Ding, J.; Liao, C.; Xu, J.; Liu, X.; Lu, W. Defensins: The natural peptide antibiotic. Adv Drug Deliv Rev 2021, 179, 114008. [Google Scholar] [CrossRef]
- Xu, D.; Lu, W. Defensins: A Double-Edged Sword in Host Immunity. Front Immunol 2020, 11. [Google Scholar] [CrossRef] [PubMed]
- Solanki, S.S.; Singh, P.; Kashyap, P.; Sansi, M.S.; Ali, S.A. Promising role of defensins peptides as therapeutics to combat against viral infection. Microb Pathog 2021, 155, 104930. [Google Scholar] [CrossRef] [PubMed]
- Fu, J.; Zong, X.; Jin, M.; Min, J.; Wang, F.; Wang, Y. Mechanisms and regulation of defensins in host defense. Signal Transduct Target Ther 2023, 8, 300. [Google Scholar] [CrossRef]
- Contreras, G.; Shirdel, I.; Braun, M.S.; Wink, M. Defensins: Transcriptional regulation and function beyond antimicrobial activity. Dev Comp Immunol 2020, 104, 103556. [Google Scholar] [CrossRef]
- Hancock, R.E.W.; Alford, M.A.; Haney, E.F. Antibiofilm activity of host defence peptides: Complexity provides opportunities. Nat Rev Microbiol 2021, 19, 786–797. [Google Scholar] [CrossRef]
- Hilchie, A.L.; Wuerth, K.; Hancock, R.E.W. Immune modulation by multifaceted cationic host defense (antimicrobial) peptides. Nat Chem Biol 2013, 9, 761–768. [Google Scholar] [CrossRef] [PubMed]
- Lu, W. Antimicrobial peptides. Semin Cell Dev Biol 2019, 88, 105–106. [Google Scholar] [CrossRef]
- Brook, M.; Tomlinson, G.H.; Miles, K.; Smith, R.W.P.; Rossi, A.G.; Hiemstra, P.S.; van ’t Wout, E.F.A.; Dean, J.L.E.; Gray, N.K.; Lu, W.; et al. Neutrophil-derived alpha defensins control inflammation by inhibiting macrophage mRNA translation. Proc Natl Acad Sci 2016, 113, 4350–4355. [Google Scholar] [CrossRef]
- Hancock, R.E.W.; Haney, E.F.; Gill, E.E. The immunology of host defence peptides: Beyond antimicrobial activity. Nat Rev Immunol 2016, 16, 321–334. [Google Scholar] [CrossRef] [PubMed]
- Chen, Q.; Yang, Y.; Hou, J.; Shu, Q.; Yin, Y.; Fu, W.; Han, F.; Hou, T.; Zeng, C.; Nemeth, E.; et al. Increased gene copy number of DEFA1/DEFA3 worsens sepsis by inducing endothelial pyroptosis. Proc Natl Acad Sci 2019, 116, 3161–3170. [Google Scholar] [CrossRef]
- Higazi, M.; Abdeen, S.; Abu-Fanne, R.; Heyman, S.N.; Masarwy, A.; Bdeir, K.; Maraga, E.; Cines, D.B.; Higazi, A.A.-R. Opposing effects of HNP1 (α-defensin-1) on plasma cholesterol and atherogenesis. PLoS ONE 2020, 15, e0231582. [Google Scholar] [CrossRef]
- Abu-Fanne, R.; Stepanova, V.; Litvinov, R.I.; Abdeen, S.; Bdeir, K.; Higazi, M.; Maraga, E.; Nagaswami, C.; Mukhitov, A.R.; Weisel, J.W.; et al. Neutrophil α-defensins promote thrombosis in vivo by altering fibrin formation, structure, and stability. Blood 2019, 133, 481–493. [Google Scholar] [CrossRef] [PubMed]
- Antikainen, A.A.V.; Sandholm, N.; Trégouët, D.-A.; Charmet, R.; McKnight, A.J.; Ahluwalia, T.S.; Syreeni, A.; Valo, E.; Forsblom, C.; Gordin, D.; et al. Genome-wide association study on coronary artery disease in type 1 diabetes suggests beta-defensin 127 as a risk locus. Cardiovasc Res 2020, 117, 600–612. [Google Scholar] [CrossRef]
- Chen, Q.; Yang, Y.; Pan, Y.; Shen, L.; Zhang, Y.; Zheng, F.; Shu, Q.; Fang, X. Human Neutrophil Defensins Disrupt Liver Interendothelial Junctions and Aggravate Sepsis. Mediators Inflamm 2022, 2022, 7659282. [Google Scholar] [CrossRef]
- Miani, M.; Le Naour, J.; Waeckel-Enée, E.; Verma, S.c.; Straube, M.; Emond, P.; Ryffel, B.; van Endert, P.; Sokol, H.; Diana, J. Gut Microbiota-Stimulated Innate Lymphoid Cells Support β-Defensin 14 Expression in Pancreatic Endocrine Cells, Preventing Autoimmune Diabetes. Cell Metab 2018, 28, 557–572.e556. [Google Scholar] [CrossRef]
- Kolbinger, F.; Loesche, C.; Valentin, M.-A.; Jiang, X.; Cheng, Y.; Jarvis, P.; Peters, T.; Calonder, C.; Bruin, G.; Polus, F.; et al. β-Defensin 2 is a responsive biomarker of IL-17A–driven skin pathology in patients with psoriasis. J Allergy Clin Immunol 2017, 139, 923–932.e928. [Google Scholar] [CrossRef]
- Lu, W.; de Leeuw, E. Pro-inflammatory and pro-apoptotic properties of Human Defensin 5. Biochem Biophys Res Commun 2013, 436, 557–562. [Google Scholar] [CrossRef]
- Biragyn, A.; Ruffini, P.A.; Leifer, C.A.; Klyushnenkova, E.; Shakhov, A.; Chertov, O.; Shirakawa, A.K.; Farber, J.M.; Segal, D.M.; Oppenheim, J.J.; et al. Toll-like receptor 4-dependent activation of dendritic cells by beta-defensin 2. Science 2002, 298, 1025–1029. [Google Scholar] [CrossRef] [PubMed]
- Prasad, S.V.; Fiedoruk, K.; Daniluk, T.; Piktel, E.; Bucki, R. Expression and Function of Host Defense Peptides at Inflammation Sites. Int J Mol Sci 2019, 21. [Google Scholar] [CrossRef]
- Lai, Y.; Gallo, R.L. AMPed up immunity: How antimicrobial peptides have multiple roles in immune defense. Trends Immunol 2009, 30, 131–141. [Google Scholar] [CrossRef] [PubMed]
- Shelley, J.R.; Davidson, D.J.; Dorin, J.R. The Dichotomous Responses Driven by β-Defensins. Front Immunol 2020, 11. [Google Scholar] [CrossRef]
- Koeninger, L.; Armbruster, N.S.; Brinch, K.S.; Kjaerulf, S.; Andersen, B.; Langnau, C.; Autenrieth, S.E.; Schneidawind, D.; Stange, E.F.; Malek, N.P.; et al. Human β-Defensin 2 Mediated Immune Modulation as Treatment for Experimental Colitis. Front Immunol 2020, 11. [Google Scholar] [CrossRef]
- Su, G.; Luo, Y.; Chen, D.; Yu, B.; He, J. NF-κB-dependent induction of porcine β-defensin 114 regulates intestinal epithelium homeostasis. Int J Biol Macromol 2021, 192, 241–249. [Google Scholar] [CrossRef]
- Xie, K.; Su, G.; Chen, D.; Yu, B.; Huang, Z.; Yu, J.; Zheng, P.; Luo, Y.; Yan, H.; Li, H.; et al. The immunomodulatory function of the porcine β-defensin 129: Alleviate inflammatory response induced by LPS in IPEC-J2 cells. Int J Biol Macromol 2021, 188, 473–481. [Google Scholar] [CrossRef] [PubMed]
- Jin, X.; Li, Q.-h.; Sun, J.; Zhang, M.; Xiang, Y.-q. Porcine β-defensin-2 alleviates AFB1-induced intestinal mucosal injury by inhibiting oxidative stress and apoptosis. Ecotoxicol Environ Saf 2023, 262, 115161. [Google Scholar] [CrossRef]
- Zhang, K.; Lian, S.; Shen, X.; Zhao, X.; Zhao, W.; Li, C. Recombinant porcine beta defensin 2 alleviates inflammatory responses induced by Escherichia coli in IPEC-J2 cells. Int J Biol Macromol 2022, 208, 890–900. [Google Scholar] [CrossRef] [PubMed]
- Jin, X.; Li, Q.H.; Sun, J.; Zhang, M.; Xiang, Y.Q. Porcine β-defensin-2 alleviates AFB1-induced intestinal mucosal injury by inhibiting oxidative stress and apoptosis. Ecotoxicol Environ Saf 2023, 262, 115161. [Google Scholar] [CrossRef]
- Li, Q.; Zhang, M.; Sun, J.; Li, Y.; Zu, S.; Xiang, Y.; Jin, X. Porcine β-defensin-2 alleviates aflatoxin B1 induced intestinal mucosal damage via ROS-Erk(1/2) signaling pathway. Sci Total Environ 2023, 905, 167201. [Google Scholar] [CrossRef]
- Huang, J.; Liu, X.; Sun, Y.; Huang, C.; Wang, A.; Xu, J.; Zhou, H.; Li, L.; Zhou, R. Porcine β-defensin 2 confers enhanced resistance to swine flu infection in transgenic pigs and alleviates swine influenza virus-induced apoptosis possibly through interacting with host SLC25A4. Antiviral Res 2022, 201, 105292. [Google Scholar] [CrossRef]
- Huang, C.; Yang, X.; Huang, J.; Liu, X.; Yang, X.; Jin, H.; Huang, Q.; Li, L.; Zhou, R. Porcine Beta-Defensin 2 Provides Protection Against Bacterial Infection by a Direct Bactericidal Activity and Alleviates Inflammation via Interference With the TLR4/NF-κB Pathway. Front Immunol 2019, 10, 1673. [Google Scholar] [CrossRef]
- Veldhuizen, E.J.; Rijnders, M.; Claassen, E.A.; van Dijk, A.; Haagsman, H.P. Porcine beta-defensin 2 displays broad antimicrobial activity against pathogenic intestinal bacteria. Mol Immunol 2008, 45, 386–394. [Google Scholar] [CrossRef]
- Mookherjee, N.; Anderson, M.A.; Haagsman, H.P.; Davidson, D.J. Antimicrobial host defence peptides: Functions and clinical potential. Nat Rev Drug Discov 2020, 19, 311–332. [Google Scholar] [CrossRef] [PubMed]
- Su, G.; Xie, K.; Chen, D.; Yu, B.; Huang, Z.; Luo, Y.; Mao, X.; Zheng, P.; Yu, J.; Luo, J.; et al. Differential expression, molecular cloning, and characterization of porcine beta defensin 114. J Anim Sci Biotechnol 2019, 10, 60. [Google Scholar] [CrossRef] [PubMed]
- Yu, L.-t.; Xiao, Y.-P.; Li, J.-J.; Ran, J.-S.; Yin, L.-Q.; Liu, Y.-P.; Zhang, L. Molecular characterization of a novel ovodefensin gene in chickens. Gene 2018, 678, 233–240. [Google Scholar] [CrossRef]
- Yang, D.; Chen, Q.; Chertov, O.; Oppenheim, J.J. Human neutrophil defensins selectively chemoattract naive T and immature dendritic cells. J Leukoc Biol 2000, 68, 9–14. [Google Scholar] [CrossRef]
- Corrales-García, L.L.; Serrano-Carreón, L.; Corzo, G. Improving the heterologous expression of human β-defensin 2 (HBD2) using an experimental design. Protein Expr Purif 2020, 167, 105539. [Google Scholar] [CrossRef] [PubMed]
- Fan, K.; An, Y.; Wang, Z.; Yin, W.; Sun, N.; Sun, Y.; Khan, A.; Li, H. Antibacterial Activity of Recombinant Porcine β-Defensin 2. Pak Vet J 2019, 39, 2074–7764. [Google Scholar] [CrossRef]
- Lian, S.; Lin, X.; Zhan, F.; Shen, X.; Liang, Y.; Li, C. Transcriptome Analysis Reveals the Multiple Functions of pBD2 in IPEC-J2 Cells against E. coli. Int J Mol Sci 2022, 23, 9754. [Google Scholar] [CrossRef] [PubMed]
- Huang, C.; Sun, Y.; Qiu, X.; Huang, J.; Wang, A.; Zhang, Q.; Pang, S.; Huang, Q.; Zhou, R.; Li, L. The Intracellular Interaction of Porcine beta-Defensin 2 with VASH1 Alleviates Inflammation via Akt Signaling Pathway. J Immunol 2022, 208, 2795–2805. [Google Scholar] [CrossRef] [PubMed]
- Oishi, Y.; Manabe, I. Macrophages in inflammation, repair and regeneration. Int Immunol 2018, 30, 511–528. [Google Scholar] [CrossRef] [PubMed]
- Fujiwara, N.; Kobayashi, K. Macrophages in inflammation. Curr Drug Targets Inflamm Allergy 2005, 4, 281–286. [Google Scholar] [CrossRef] [PubMed]
- Merly, L.; Smith, S.L. Murine RAW 264.7 cell line as an immune target: Are we missing something? Immunopharmacol Immunotoxicol 2017, 39, 55–58. [Google Scholar] [CrossRef] [PubMed]
- Elisia, I.; Pae, H.B.; Lam, V.; Cederberg, R.; Hofs, E.; Krystal, G. Comparison of RAW264.7, human whole blood and PBMC assays to screen for immunomodulators. J Immunol Methods 2018, 452, 26–31. [Google Scholar] [CrossRef]
- Sang, Y.; Patil, A.A.; Zhang, G.; Ross, C.R.; Blecha, F. Bioinformatic and expression analysis of novel porcine β-defensins. Mamm Genome 2006, 17, 332–339. [Google Scholar] [CrossRef]
- Huang, C.; Yang, X.; Huang, J.; Liu, X.; Yang, X.; Jin, H.; Huang, Q.; Li, L.; Zhou, R. Porcine Beta-Defensin 2 Provides Protection Against Bacterial Infection by a Direct Bactericidal Activity and Alleviates Inflammation via Interference With the TLR4/NF-κB Pathway. Front Immunol 2019, 10. [Google Scholar] [CrossRef]
- Han, F.; Zhang, H.; Xia, X.; Xiong, H.; Song, D.; Zong, X.; Wang, Y. Porcine β-Defensin 2 Attenuates Inflammation and Mucosal Lesions in Dextran Sodium Sulfate–Induced Colitis. J Immunol Methods 2015, 194, 1882–1893. [Google Scholar] [CrossRef] [PubMed]
- Hemshekhar, M.; Choi, K.G.; Mookherjee, N. Host Defense Peptide LL-37-Mediated Chemoattractant Properties, but Not Anti-Inflammatory Cytokine IL-1RA Production, Is Selectively Controlled by Cdc42 Rho GTPase via G Protein-Coupled Receptors and JNK Mitogen-Activated Protein Kinase. Front Immunol 2018, 9, 1871. [Google Scholar] [CrossRef] [PubMed]
- Stark, R.; Grzelak, M.; Hadfield, J. RNA sequencing: The teenage years. Nat Rev Genet 2019, 20, 631–656. [Google Scholar] [CrossRef] [PubMed]
- van Loo, G.; Bertrand, M.J.M. Death by TNF: A road to inflammation. Nat Rev Immunol 2023, 23, 289–303. [Google Scholar] [CrossRef]
- Zhao, H.; Wu, L.; Yan, G.; Chen, Y.; Zhou, M.; Wu, Y.; Li, Y. Inflammation and tumor progression: Signaling pathways and targeted intervention. Signal Transduct Target Ther 2021, 6, 263. [Google Scholar] [CrossRef] [PubMed]
- Tang, W.; Wang, H.; Ha, H.L.; Tassi, I.; Bhardwaj, R.; Claudio, E.; Siebenlist, U. The B-cell tumor promoter Bcl-3 suppresses inflammation-associated colon tumorigenesis in epithelial cells. Oncogene 2016, 35, 6203–6211. [Google Scholar] [CrossRef] [PubMed]
- Anto Michel, N.; Colberg, C.; Buscher, K.; Sommer, B.; Pramod, A.B.; Ehinger, E.; Dufner, B.; Hoppe, N.; Pfeiffer, K.; Marchini, T.; et al. Inflammatory Pathways Regulated by Tumor Necrosis Receptor-Associated Factor 1 Protect From Metabolic Consequences in Diet-Induced Obesity. Circ Res 2018, 122, 693–700. [Google Scholar] [CrossRef] [PubMed]
- Lawson, K.A.; Sousa, C.M.; Zhang, X.; Kim, E.; Akthar, R.; Caumanns, J.J.; Yao, Y.; Mikolajewicz, N.; Ross, C.; Brown, K.R.; et al. Functional genomic landscape of cancer-intrinsic evasion of killing by T cells. Nature 2020, 586, 120–126. [Google Scholar] [CrossRef] [PubMed]
- Markó, L.; Park, J.K.; Henke, N.; Rong, S.; Balogh, A.; Klamer, S.; Bartolomaeus, H.; Wilck, N.; Ruland, J.; Forslund, S.K.; et al. B-cell lymphoma/leukaemia 10 and angiotensin II-induced kidney injury. Cardiovasc Res 2020, 116, 1059–1070. [Google Scholar] [CrossRef]
- Lao, Y.; Yang, K.; Wang, Z.; Sun, X.; Zou, Q.; Yu, X.; Cheng, J.; Tong, X.; Yeh, E.T.H.; Yang, J.; et al. DeSUMOylation of MKK7 kinase by the SUMO2/3 protease SENP3 potentiates lipopolysaccharide-induced inflammatory signaling in macrophages. J Biol Chem 2018, 293, 3965–3980. [Google Scholar] [CrossRef]
- Gong, J.; Fang, C.; Zhang, P.; Wang, P.X.; Qiu, Y.; Shen, L.J.; Zhang, L.; Zhu, X.Y.; Tian, S.; Li, F.; et al. Tumor Progression Locus 2 in Hepatocytes Potentiates Both Liver and Systemic Metabolic Disorders in Mice. Hepatology 2019, 69, 524–544. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Liu, S. LncRNA GAS5 suppresses inflammatory responses and apoptosis of alveolar epithelial cells by targeting miR-429/DUSP1. Exp Mol Pathol 2020, 113, 104357. [Google Scholar] [CrossRef] [PubMed]
- Dagvadorj, J.; Mikulska-Ruminska, K.; Tumurkhuu, G.; Ratsimandresy, R.A.; Carriere, J.; Andres, A.M.; Marek-Iannucci, S.; Song, Y.; Chen, S.; Lane, M.; et al. Recruitment of pro-IL-1alpha to mitochondrial cardiolipin, via shared LC3 binding domain, inhibits mitophagy and drives maximal NLRP3 activation. Proc Natl Acad Sci U S A 2021, 118. [Google Scholar] [CrossRef]
- Corbin, A.L.; Gomez-Vazquez, M.; Berthold, D.L.; Attar, M.; Arnold, I.C.; Powrie, F.M.; Sansom, S.N.; Udalova, I.A. IRF5 guides monocytes toward an inflammatory CD11c(+) macrophage phenotype and promotes intestinal inflammation. Sci Immunol 2020, 5. [Google Scholar] [CrossRef]
- Gerbino, V.; Kaunga, E.; Ye, J.; Canzio, D.; O’Keeffe, S.; Rudnick, N.D.; Guarnieri, P.; Lutz, C.M.; Maniatis, T. The Loss of TBK1 Kinase Activity in Motor Neurons or in All Cell Types Differentially Impacts ALS Disease Progression in SOD1 Mice. Neuron 2020, 106, 789–805.e785. [Google Scholar] [CrossRef] [PubMed]
- Semple, F.; MacPherson, H.; Webb, S.; Cox, S.L.; Mallin, L.J.; Tyrrell, C.; Grimes, G.R.; Semple, C.A.; Nix, M.A.; Millhauser, G.L.; et al. Human β-defensin 3 affects the activity of pro-inflammatory pathways associated with MyD88 and TRIF. Eur J Immunol 2011, 41, 3291–3300. [Google Scholar] [CrossRef] [PubMed]
- Xie, K.; Xie, H.; Su, G.; Chen, D.; Yu, B.; Mao, X.; Huang, Z.; Yu, J.; Luo, J.; Zheng, P.; et al. β-Defensin 129 Attenuates Bacterial Endotoxin-Induced Inflammation and Intestinal Epithelial Cell Apoptosis. Front Immunol 2019, 10, 2333. [Google Scholar] [CrossRef]
- Wiegand, I.; Hilpert, K.; Hancock, R.E. Agar and broth dilution methods to determine the minimal inhibitory concentration (MIC) of antimicrobial substances. Nat Protoc 2008, 3, 163–175. [Google Scholar] [CrossRef]
- Wen, P.; He, J.; Zhang, Q.; Qi, H.; Zhang, A.; Liu, D.; Sun, Q.; Wang, Y.; Li, Q.; Wang, W.; et al. SET Domain Group 703 Regulates Planthopper Resistance by Suppressing the Expression of Defense-Related Genes. Int J Mol Sci 2023, 24, 13003. [Google Scholar] [CrossRef]
Figure 1.
Hemolytic (A) and Cytotoxicity (B) of pBD114.
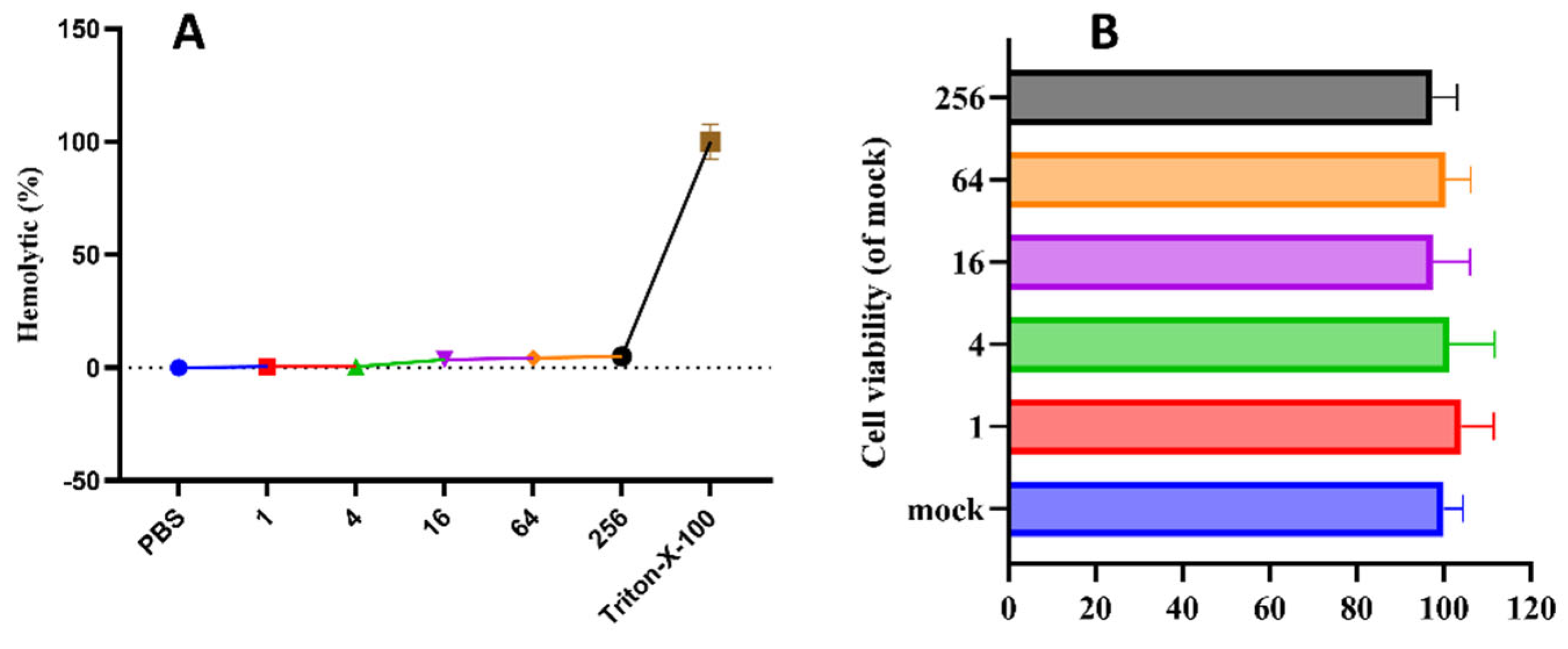
Figure 2.
The proinflammatory activity of pBD114 on macrophage RAW264.7. RAW264.7 were treated with 0, 1, 10 and 100 μg/mL pBD114 for 1, 3, 6 and 12 h, the concentration of TNFα (A: 1 h, B: 3 h, C: 6 h, D: 12 h) and IL-10 (E: 1 h, F: 3 h, G: 6 h, H: 12 h) in the supernatant of cell culture were detected by ELISA.
Figure 2.
The proinflammatory activity of pBD114 on macrophage RAW264.7. RAW264.7 were treated with 0, 1, 10 and 100 μg/mL pBD114 for 1, 3, 6 and 12 h, the concentration of TNFα (A: 1 h, B: 3 h, C: 6 h, D: 12 h) and IL-10 (E: 1 h, F: 3 h, G: 6 h, H: 12 h) in the supernatant of cell culture were detected by ELISA.
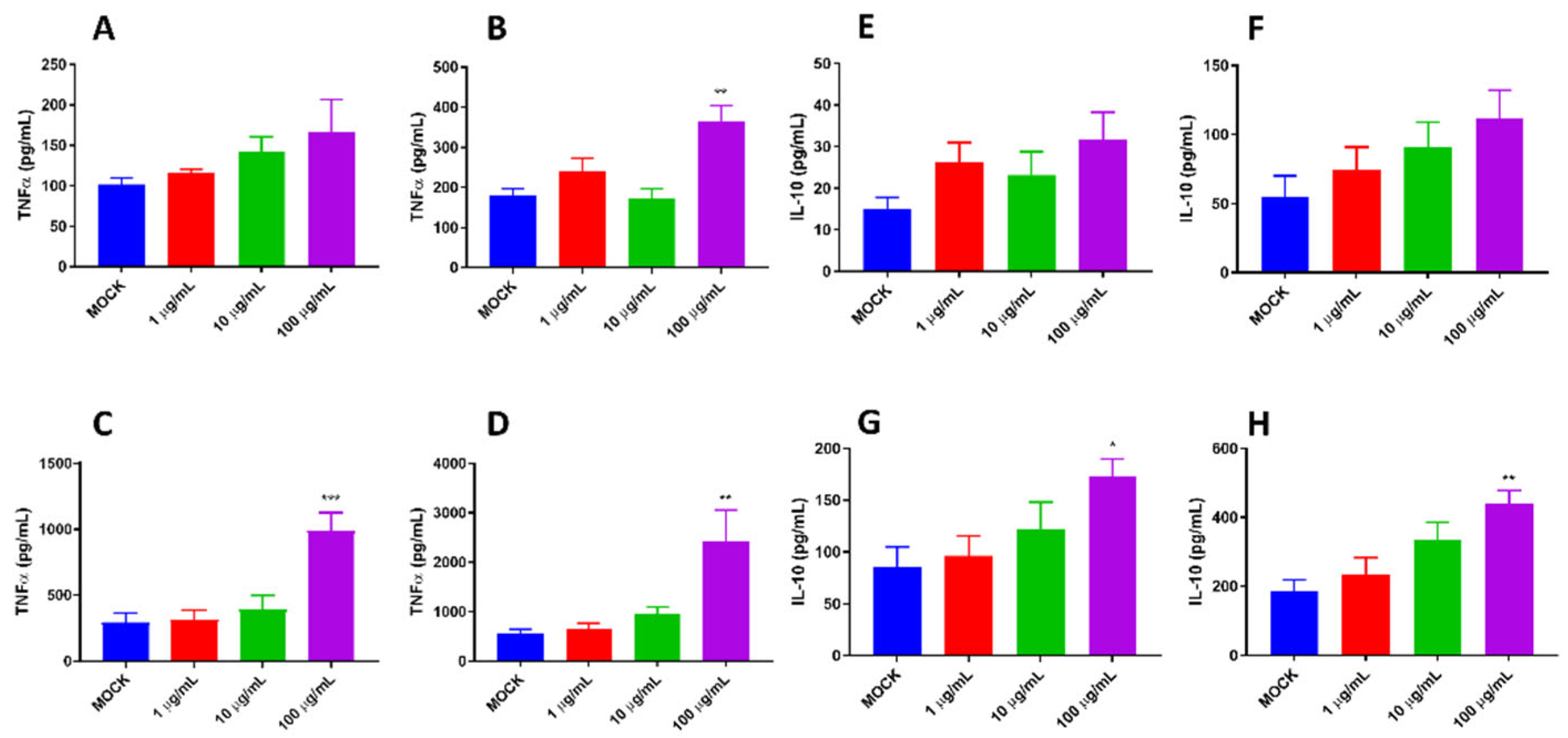
Figure 3.
Anti-inflammatory activity of pBD114 on macrophage RAW264.7. RAW264.7 were treated with 0, 10, 100, and 1000 ng/mL LPS for 0.5, 1, 2, and 4 h, the concentration of TNFα (A: 0.5 h, B: 1 h, C: 2 h, D: 4 h) in the supernatant of cell culture were detected by ELISA. RAW264.7 cells were co-cultured with 1, 5, 10, 20, and 50 μg/mL pBD114 and 10 ng/mL LPS for 2 h, and the concentration of TNFα (E) in the supernatant of cell culture was detected by ELISA.
Figure 3.
Anti-inflammatory activity of pBD114 on macrophage RAW264.7. RAW264.7 were treated with 0, 10, 100, and 1000 ng/mL LPS for 0.5, 1, 2, and 4 h, the concentration of TNFα (A: 0.5 h, B: 1 h, C: 2 h, D: 4 h) in the supernatant of cell culture were detected by ELISA. RAW264.7 cells were co-cultured with 1, 5, 10, 20, and 50 μg/mL pBD114 and 10 ng/mL LPS for 2 h, and the concentration of TNFα (E) in the supernatant of cell culture was detected by ELISA.
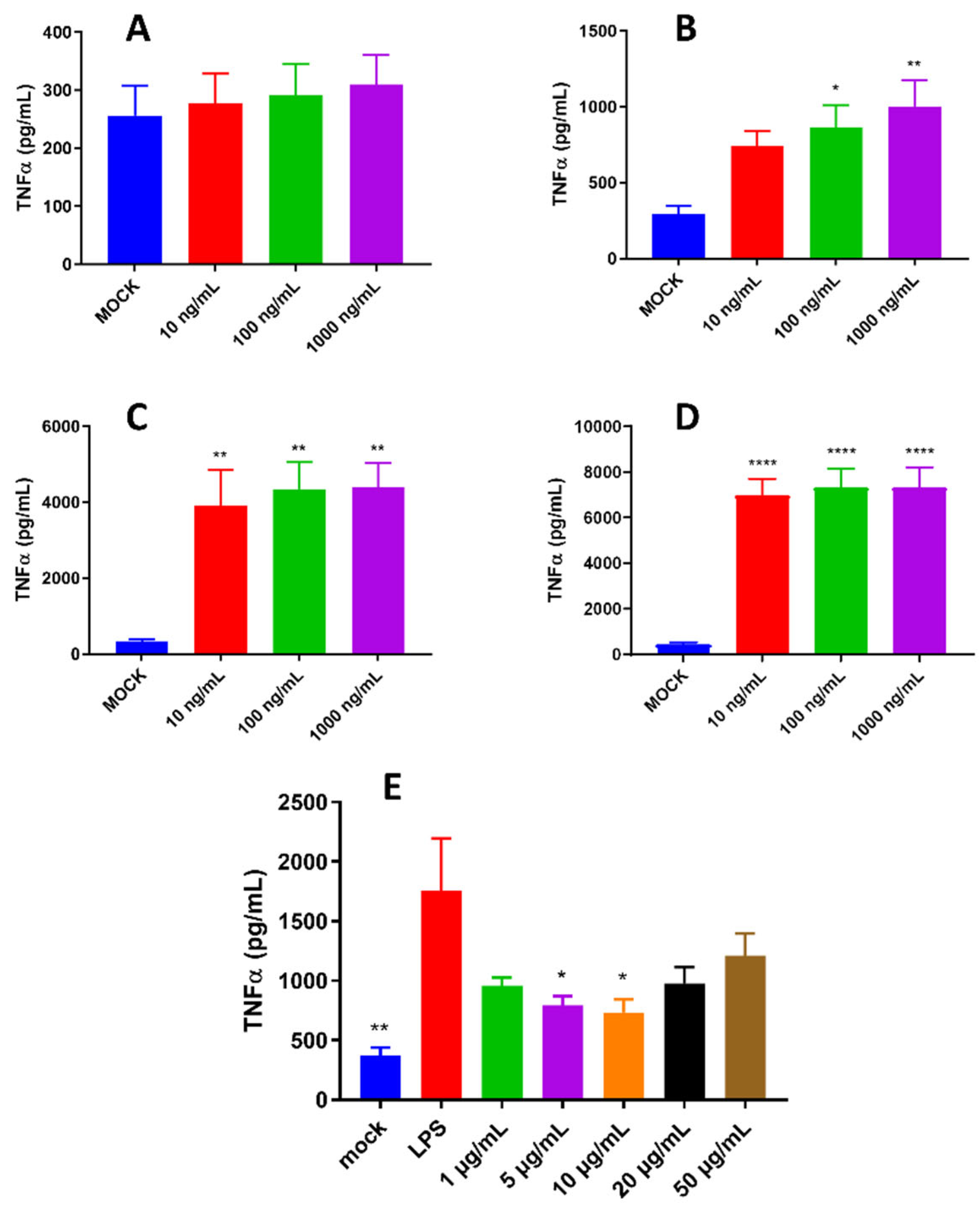
Figure 4.
Analysis of differential expression genes (DEGs). (A) Principal component analysis (PCA). The X and Y axes represent the new dataset corresponding to the principal components obtained after dimensionality reduction of the sample expressions, which is used to represent the disparity between the samples; the values in parentheses in the labels of the axes represent the percentage of the overall variance explained by the corresponding principal components. Dots represent each sample, and the same color represents the same sample group. pv denotes the proportion of variance, and SD denotes the standard deviation. Volcano plot of PBD114/MOCK (B) and PBD114LPS/LPS (C). The X-axis represents log2-transformed difference multiplicity values, and the Y-axis represents -log10-transformed significance values. Red represents up-regulated DEG, blue represents down-regulated DEGs, and gray represents non-DEG.
Figure 4.
Analysis of differential expression genes (DEGs). (A) Principal component analysis (PCA). The X and Y axes represent the new dataset corresponding to the principal components obtained after dimensionality reduction of the sample expressions, which is used to represent the disparity between the samples; the values in parentheses in the labels of the axes represent the percentage of the overall variance explained by the corresponding principal components. Dots represent each sample, and the same color represents the same sample group. pv denotes the proportion of variance, and SD denotes the standard deviation. Volcano plot of PBD114/MOCK (B) and PBD114LPS/LPS (C). The X-axis represents log2-transformed difference multiplicity values, and the Y-axis represents -log10-transformed significance values. Red represents up-regulated DEG, blue represents down-regulated DEGs, and gray represents non-DEG.
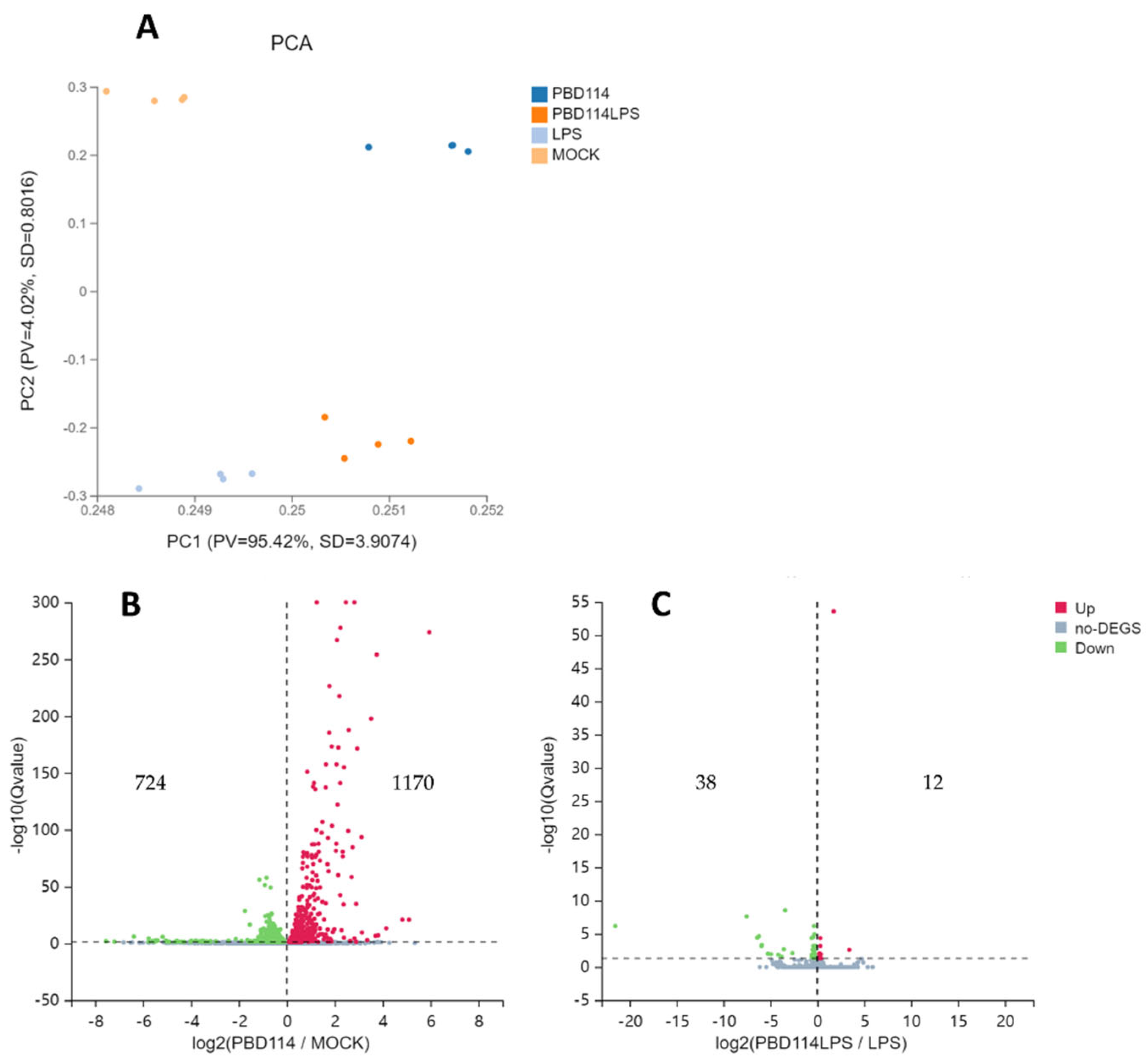
Figure 5.
KEGG enrichment analysis of differential expression genes (DEGs) for PBD114/MOCK (A) and PBD114LPS/LPS (B). The X-axis is the enrichment ratio [the ratio of the number of genes annotated to an entry in a selected gene set to the total number of genes annotated to that entry in the species, calculated as Rich Ratio = Term Candidate Gene Num / Term Gene Num]. The Y-axis is the KEGG Pathway, and the size of the bubbles indicates the number of genes annotated to a certain KEGG. The size of the bubble indicates the number of genes annotated to a KEGG pathway. The color represents the enrichment significance value (Qvalue or Pvalue, see the legend for details). The redder the color, the smaller the significance value.
Figure 5.
KEGG enrichment analysis of differential expression genes (DEGs) for PBD114/MOCK (A) and PBD114LPS/LPS (B). The X-axis is the enrichment ratio [the ratio of the number of genes annotated to an entry in a selected gene set to the total number of genes annotated to that entry in the species, calculated as Rich Ratio = Term Candidate Gene Num / Term Gene Num]. The Y-axis is the KEGG Pathway, and the size of the bubbles indicates the number of genes annotated to a certain KEGG. The size of the bubble indicates the number of genes annotated to a KEGG pathway. The color represents the enrichment significance value (Qvalue or Pvalue, see the legend for details). The redder the color, the smaller the significance value.
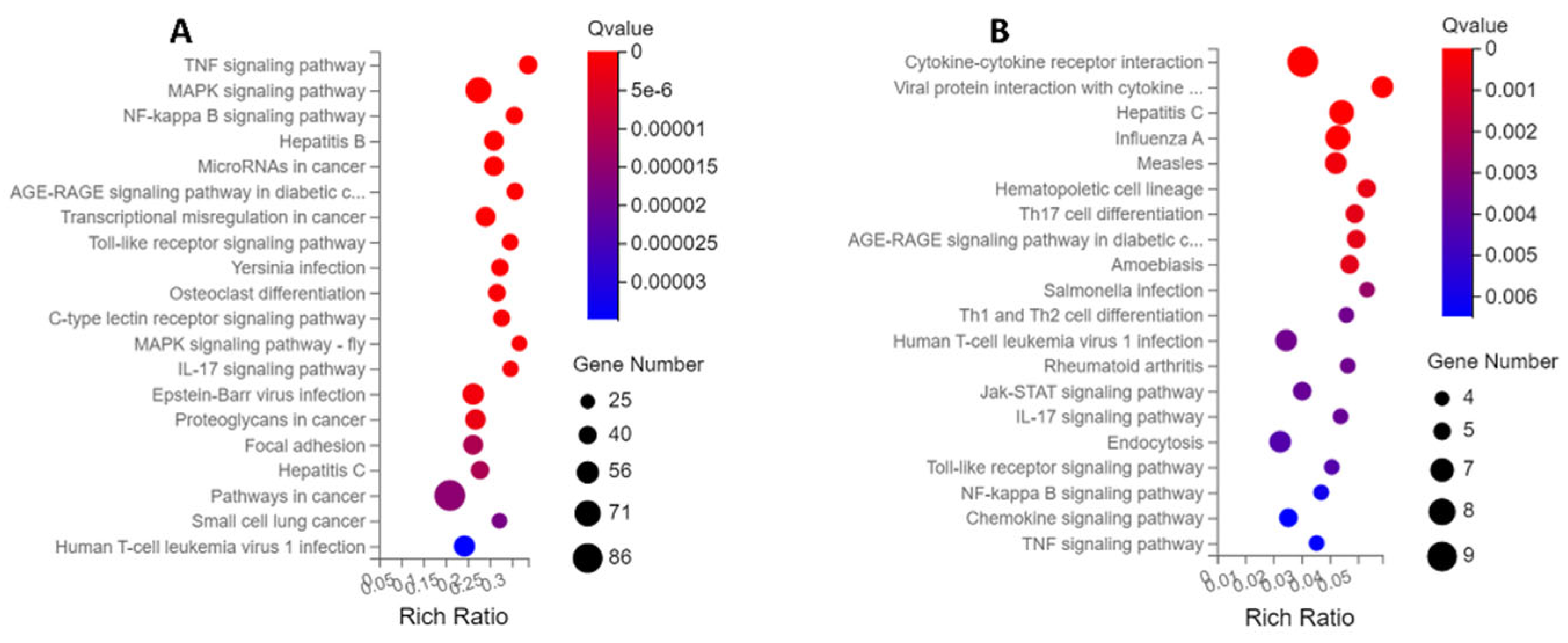
Figure 6.
PPI and KDA analysis of differential expression genes (DEGs) for PBD114/MOCK (A and C) and PBD114LPS/LPS (B and D).
Figure 6.
PPI and KDA analysis of differential expression genes (DEGs) for PBD114/MOCK (A and C) and PBD114LPS/LPS (B and D).
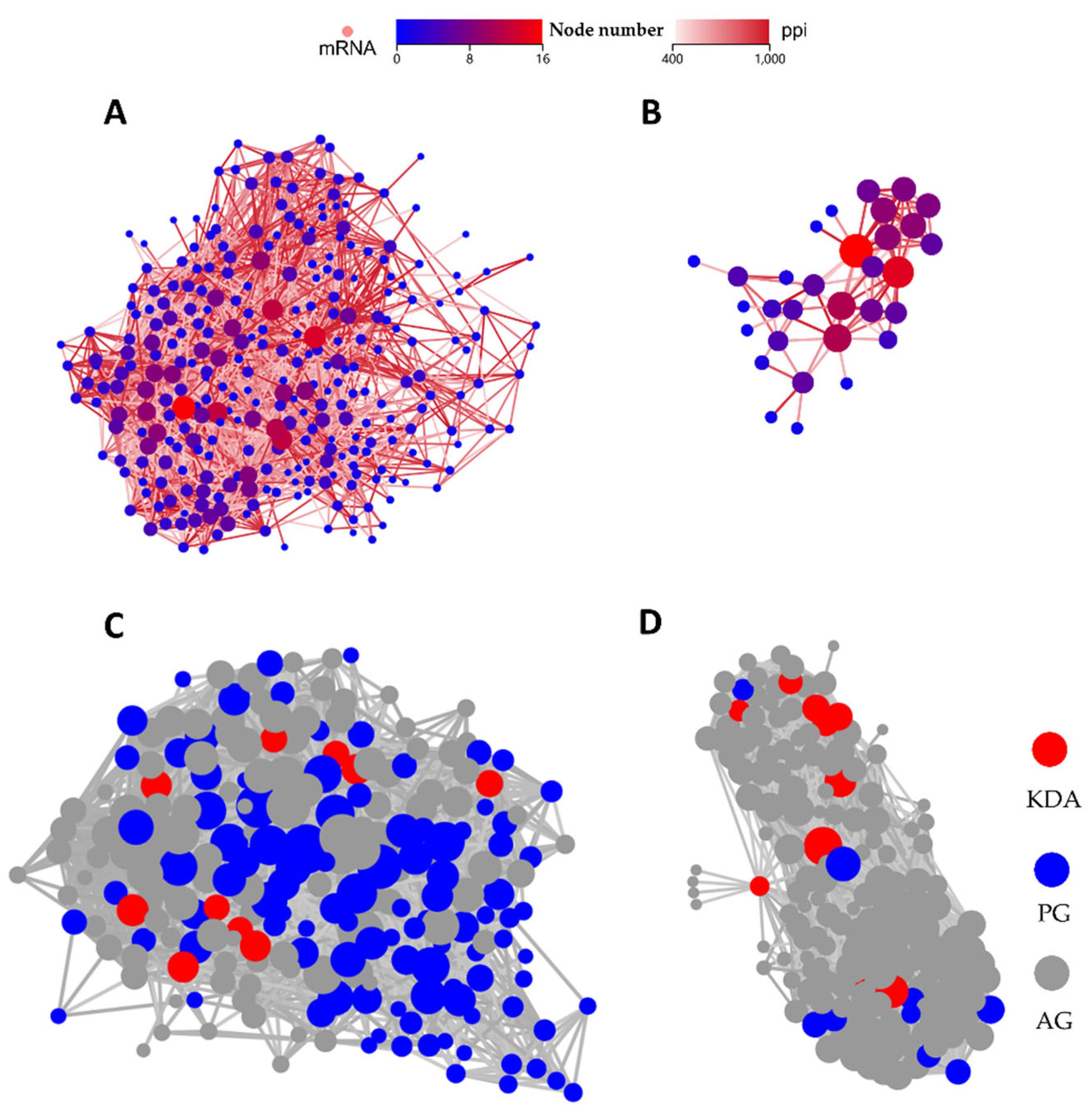
Figure 7.
Validation of RNA-seq (A) data using real-time qRT-PCR (B).
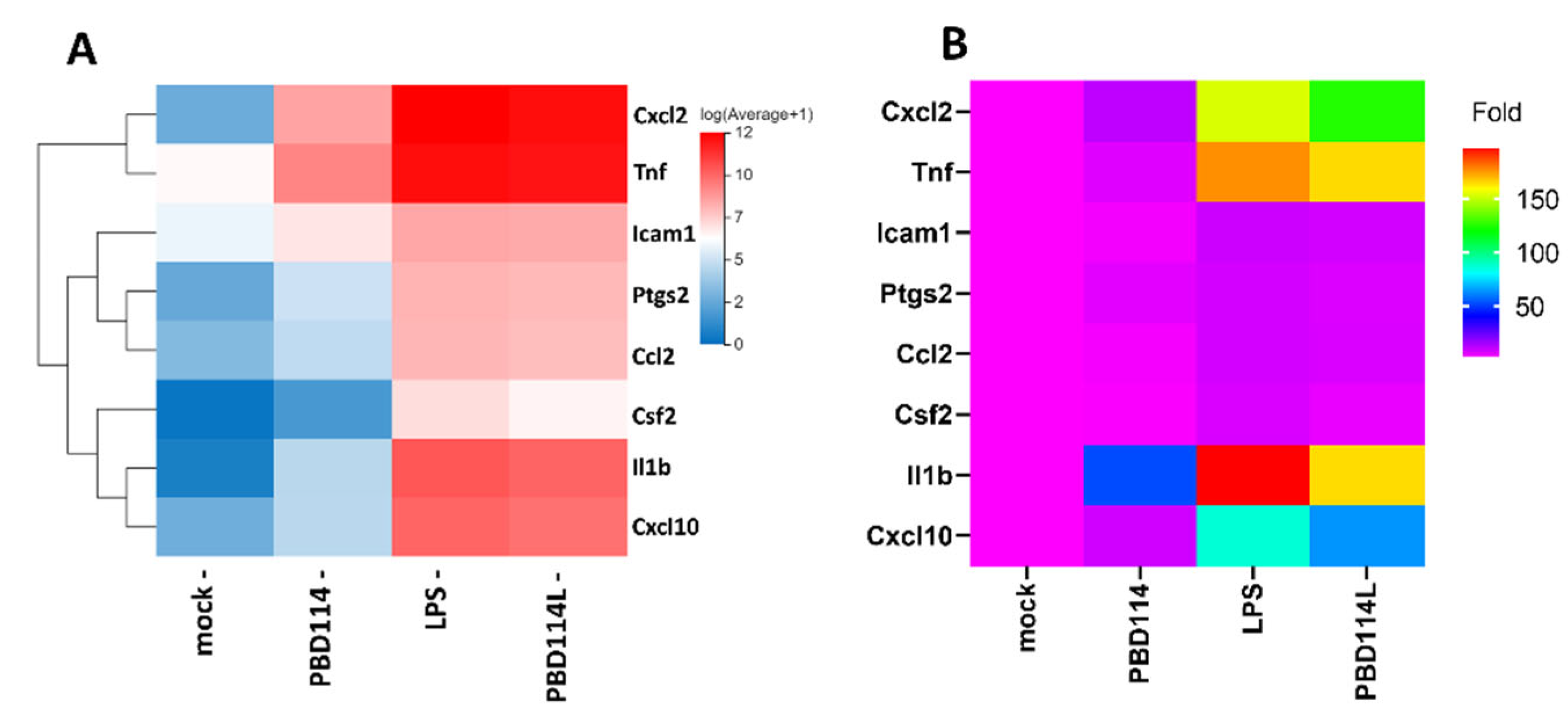
Figure 8.
Effects of pBD114 on spleen and serum inflammatory factors in mice. The intraperitoneal injection of LPS (5.0 mg/kg) and pBD114 (5.0 mg/kg) were administered in mice for 12 h. Body weight (A), spleen weight (B), and spleen index (C) were calculated, and the concentrations of TNFα (D), IL-6 (E), IL-1β (F), and IL-10 (G) proteins in mice serum were measured by ELISA.
Figure 8.
Effects of pBD114 on spleen and serum inflammatory factors in mice. The intraperitoneal injection of LPS (5.0 mg/kg) and pBD114 (5.0 mg/kg) were administered in mice for 12 h. Body weight (A), spleen weight (B), and spleen index (C) were calculated, and the concentrations of TNFα (D), IL-6 (E), IL-1β (F), and IL-10 (G) proteins in mice serum were measured by ELISA.
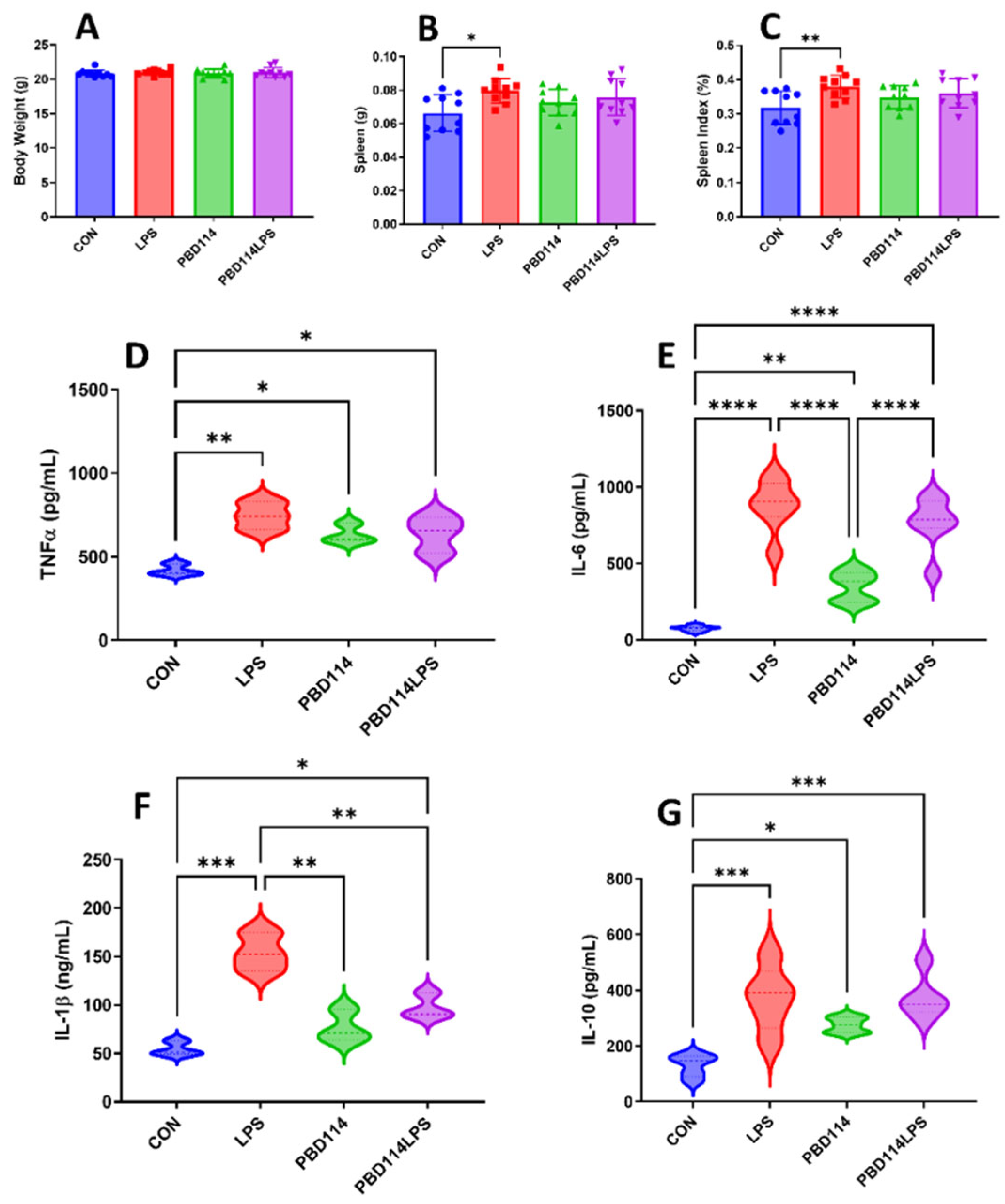
Figure 9.
Effects of pBD114 on the gene expression level of the spleen in mice.
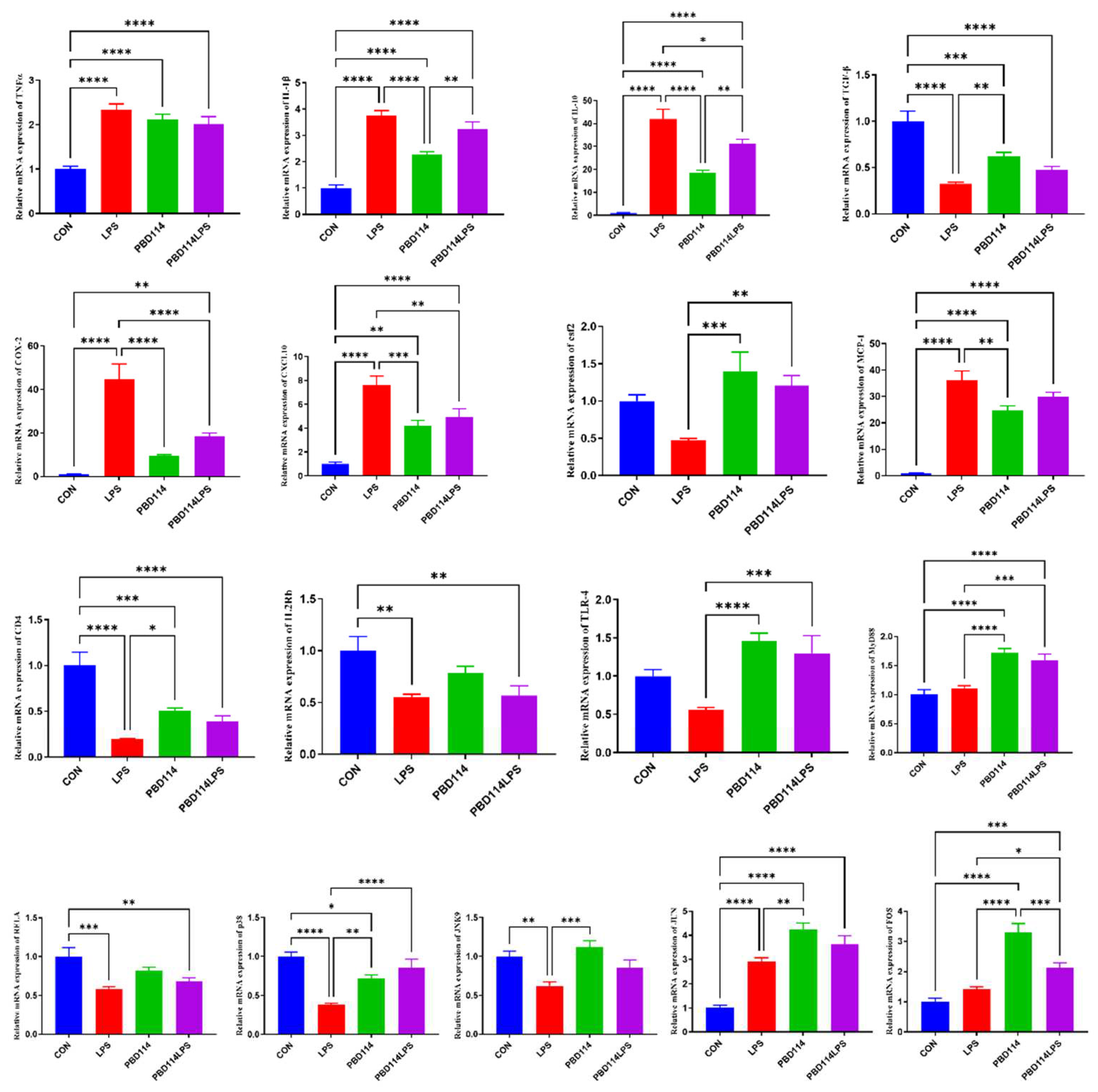

Table 1.
Quality control information for transcriptome samples.
| Sample | Total Raw Reads, M | Total Clean Reads, M | Total Clean Bases, Gb | Clean Reads Q20, % | Clean Reads Q30, % |
|---|---|---|---|---|---|
| LPS-1 | 23.92 | 23.83 | 1.19 | 97.82 | 93.66 |
| LPS-2 | 23.92 | 23.83 | 1.19 | 97.86 | 93.73 |
| LPS-3 | 23.92 | 23.82 | 1.19 | 97.87 | 93.78 |
| LPS-4 | 22.24 | 22.15 | 1.11 | 97.76 | 93.47 |
| MOCK-1 | 23.92 | 23.83 | 1.19 | 97.77 | 93.53 |
| MOCK-2 | 23.92 | 23.83 | 1.19 | 97.83 | 93.72 |
| MOCK-3 | 23.92 | 23.81 | 1.19 | 97.81 | 93.60 |
| MOCK-4 | 23.92 | 23.83 | 1.19 | 97.84 | 93.75 |
| PBD114-1 | 23.92 | 23.70 | 1.18 | 97.70 | 93.28 |
| PBD114-2 | 23.92 | 23.67 | 1.18 | 97.47 | 92.69 |
| PBD114-3 | 23.92 | 23.66 | 1.18 | 97.77 | 93.52 |
| PBD114-4 | 23.92 | 23.69 | 1.18 | 97.47 | 92.73 |
| PBD114LPS-1 | 23.92 | 23.86 | 1.19 | 97.40 | 92.22 |
| PBD114LPS-2 | 23.92 | 23.85 | 1.19 | 97.46 | 92.40 |
| PBD114LPS-3 | 22.91 | 22.84 | 1.14 | 97.59 | 92.69 |
| PBD114LPS-4 | 21.26 | 21.20 | 1.06 | 98.08 | 93.98 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



