
Submitted:
08 December 2023
Posted:
11 December 2023
You are already at the latest version
Abstract
The genetic engineering of plants often relies on the use of antibiotic or herbicide resistance genes for initial selection of primary transgenic events. Nevertheless, the commercial release of genetically modified crops containing any marker gene encounters several challenges stemming from the lack of consumer acceptance. Development of strategies enabled the generation of marker-free transgenic plants is a possible alternative to deal with public concerns over the safety of biotech crops. Leveraging the high in vitro regeneration and transformation capabilities of potato, this study examined the factors associated with the development of transgenic plants without presence of selective substances in medium. To compare the antibiotic-assisted with the antibiotic-free transformation, nodal explants from five highly regenerative potato cultivars were inoculated with the Agrobacterium strain AGL0 carrying the pBIN-mGFP5-ER (35SGFP/nosNPTII) plasmid. The efficiency of transient expression and the formation of chimeric and completely transgenic shoots were assessed through vital GFP observations. Under non selective conditions, the regeneration efficiency of stable non-chimeric transgenic shoots varied from 0.9 (‘Chicago’) to 2.7% (#12-36-42). In order to streamline transformation protocol, a specific timeline for collecting transgenic plantlets was established for each analyzed cultivar. This timeline strategy was implemented to generate transgenic potato plants, which demonstrated the suppression of homologous SteIF4Es family genes as a result of the successful introduction of the marker-free hpRNAi construct.
Keywords:
Solanum tuberosum L.
; antibiotic-free
; selectable marker gene
; gene silencing
; translation initiation factor eIF4E
1. Introduction
Genetic engineering has significantly contributed to the fundamental and practical understanding of plant biology and genetics over the past 45 years. Notably, potato was one of the first plant species to successfully undergo genetic transformation [1]. Since then numerous publications have detailed the generation of diverse transgenic potato plants carrying of a wide array of engineered traits [2,3,4]. Potato is an important food crop, which is cultivated not only for direct human consumption, but also as animal feed and industrial crop for the production of alcohol, biofuel, starch, and other substances. It stands as the 4th major staple crop globally, following maize, wheat, and rice, and is cultivated in over 150 countries. Faced with various production challenges, breeders are constantly making efforts to develop new varieties adapted to local agricultural needs in order to increase productivity while reducing costs [5]. The breeding of potato is traditionally achieved through sexual hybridization between related species. However, the limiting factor in potato breeding is the long generation period, which makes it difficult to effectively introduce new traits into commercial varieties using traditional approaches to crossing. For this reason, potato genetic engineering is an important area of research, focusing on improving traits that are not easily modified by conventional breeding [2,3].
The generation of transgenic plants in any species hinges on the efficient selection of transgenic tissues, typically facilitated by various marker genes that confer resistance to selective substances, such as antibiotics or herbicides. However, the presence of antibiotic or herbicide resistance genes in transgenic plants intended for commercial use has raised significant consumer concerns regarding health risks and biosafety. This complicates the regulatory processes overseeing the release of biotech crops [6]. Thus, the development of marker-free transgenic plants stands is one of the options to mitigate perceived risks and reduce public concerns
In addressing the issue of marker gene presence, various strategies have emerged over the past decades. These include approaches that involve the direct removal of selective sequences from the transformation process or the controlled elimination of marker genes from the plant genome after the marker-assistant selection of transgenic plants is completed. The methods of removal include co-transformation, site-specific recombination, transposition, and the utilization of double right border-binary vectors or multi auto-transformation vectors [7,8,9,10]. In co-transformation, vectors are carried two separate T-DNAs: one containing the gene of interest and the other encoding a selection marker gene, both of which are introduced into the plant genome simultaneously. If insertions occur at unlinked genomic loci, they can independently segregate in subsequent generations, yielding marker-free segregants [10]. Similarly, segregation is necessary when transgenic plants are transformed with the double right border twin T-DNA vector, featuring an additional copy of the right border sequence inserted between the marker gene and the gene of interest [11]. Segregation-based strategies are extensively applied for various seed-propagated crops [9,10] but are impractical for vegetatively propagated species, such as potato.
Approaches directly applied to primary transformants, such as the site-specific recombination or systems involving transposon-based and multi auto-transformation vectors are better suited for potato. Various self-excision site-specific recombination systems, including Cre-lox, FLP-FRT, and R/RS, have already been applied to varying degrees for vegetatively propagated horticulture crops, including apricot [12], tomato [13], blood orange [14], strawberries [15], apples [16,17,18] and pears [19]. The Cre-lox and R/RS systems also have also demonstrated effectiveness in generating marker-free transgenic plants of potato. The heat shock inducible Cre-lox systems achieved a 71% excision efficiency of the NPTII antibiotic resistance marker gene in potato cv. ‘Desire’ [20]; while the R/RS recombination tool exhibited lower efficacy, producing only 29% marker-free transgenic plants from regenerated shoots of the same cultivar [21]. The ipt(isopentenyl transferase)-type multi-auto-transformation vector has also been employed to generate marker-free disease-resistant transgenic potato plants [22]. While these approaches have been successful, they do carry certain limitations. These include the need for designing sophisticated constructs, resulting in decreased transformation efficiency; the requirement for additional time for manipulations to re-regenerate marker-free plants after induced excision; the inability to achieve 100% excision efficiency; and unwanted chromosomal rearrangements in the plant genome, as a result of the expression of recombinase systems, leading to genetic and phenotypic changes.
An ideal approach to overcome these limitations involves a marker-free transformation procedure, where a simple expression cassette exclusively encoding the gene of interest is transferred into the plant genome via Agrobacterium or gun-mediated transformation. In practice only a limited number of crops, such as alfalfa [23], peanut [24], tobacco [25] and wheat [26], have been transformed using this approach, with an efficacy ramging from 0.1 to 50%. This straightforward procedure relies on the high regenerative capability of cultivated in vitro tissue to produce plants with a high frequency. Given that potato is among the highly regenerative species, marker-less genetic transformation has been successfully applied to generate of late blight resistant [27,28], virus resistant [29,30] and oxidative stress tolerant transgenic potato lines [31]. The primary challenge associated with marker-less transformation is the necessity for molecular screening of several hundreds of regenerated plant to discover a few transgenic events. Since regenerated plants need to be maintained in vitro, until the results of molecular tests are obtained, transformation without selective genes becomes a time consuming and expensive procedure.
In our current study, we have analyzed the possibility of generation of transgenic potato plants without using selective substances. We employed an expression construct encoding GFP to monitor the transformation process by cultivating internodal explants on both selective and non-selective media. This approach facilitated the analysis of the efficiency of chimeric and transgenic plantlet formation at various time intervals after co-cultivation with Agrobacterium. We also studied the capacity of five potato cultivars to produce transgenic plants without selective pressure and developed a regeneration-specific timeline, aiming to streamline the marker-less genetic transformation by reducing the number of manipulations needed.
2. Materials and Methods
2.1. Plant Material and General Conditions
The plant materials used in this study consisted of various potato germplasms generously provided by the Doka-Gene Technology Ltd, Russia. These included five commercial cultivars: ‘Chicago’, ‘La Strada’, ‘Lion Heart’, ‘Manhattan’ and ‘Pirol’, along with five breeding accessions, specifically #10-9-3, #10-10-10, #12-22-129, #12-36-42 and ‘Indigo’ (formerly #10-17-10). In vitro stock plants of potato were multiplicated in plastic vessels using phytohormone-free Murashige and Skoog (MS) basal medium supplemented with 3% sucrose, solidified with 7 g L−1 agar, and adjusted to a pH 5.8-5.9 before autoclaving. Vessels were maintained in a culture room at 21 ± 2 °C, subjected to a 16 h light / 8 h dark cycle under artificial light (40 μmol m–2 s–1) provided by OSRAM cool white and fluora fluorescent lamps. In all experiments internodal segments of 0.5-1 cm in length, excised from three to four-week-old in vitro plants, were used as explants. Explants were cultured in 100 mm × 20 mm glass Petri dishes containing 25 ml of regeneration medium.
2.2. Potato Regeneration Media
All regeneration media were consisted of MS mineral salts and vitamins, supplemented with 30 g L−1 sucrose and 100 mg L−1 myo-inositol, solidified with 7 g L−1 agar, and adjusted to a pH range of 5.8-5.9. Four combinations of phytogormones were used to induce adventitious regeneration, comprising: zeatin (Zea) 3 mg L−1, indoleacetic acid (IAA) 2 mg L−1, gibberellic acid (GA) 1 mg L−1; zeatin-riboside (Zea-R) 3 mg L−1, IAA 2 mg L−1, GA 1 mg L−1; Zea 3 mg L−1, IAA 0.5 mg L−1; Zea-R 3 mg L−1, IAA 0.5 mg L−1. The preferred combinations of phytohormones were determined through preliminary experiments (data not shown).
2.3. Agrobacterium-mediated Genetic Transformation of Potato With and Without Antibiotic Selection
In these experiments A. tumefaciens strain AGL0 harboring pBIN-mGFP5-ER binary vector was used [32]. T-DNA of this construct consists of the NPTII selectable marker gene under nos promoter and the modified GFP gene driven by CaMV 35S promoter along with a leader sequence for expression localization in the endoplasmic reticulum of the cells. Prior to transformation, colonies of bacteria were cultured overnight in liquid LB medium supplemented with 100 mg L−1 kanamycin at 28 ºC on an orbital shaker. Shortly prior to explants isolation, the agrobacterial suspension was diluted with phytohormon-free MS medium to a final density of OD600 = 0.2. Approximately 50-75 freshly cut intermodal segments were immersed in 15 ml of Agrobacterium suspension and inoculated in a 100 ml glass for 25-30 minutes at room temperature. Following this, the explants were placed onto sterilized paper and blotted dry for 2–3 min under a laminar flow. After removing the excess of bacteria, the explants were transferred to Petri dishes containing the regeneration medium covered with sterilized filter paper, and maintained for 3 days in the dark at 25 ± 1 °C. Post-co-cultivation, groups of 10-12 explants were transferred to 100 mm × 20 mm glass Petri dishes containing 25 ml of fresh regeneration medium supplemented with 500 mg L−1 of the antibiotic cefotaxime. Dishes were kept in the culture room at 21 ± 2 °C under a 16 h light / 8 h dark cycle, with subculturing carried out every 2 weeks. All subsequent media plates contained the same cefotaxime concentration. From the third week after inoculation, plantlets 1.5-3.0 cm in length, regenerated by the explants under-non selective conditions, were cut and transferred to culture vessels (ten plantlets per vessel) containing 100 ml of phytohormone–free MS medium supplemented with 150 mg L−1 cefotaxime. Once the plantlets grew to 5-10 сm in length, shoot tips with one to two expanded leaves were moved into phytohormone–free MS medium containing 100 mg L−1 kanamycin for the indentication of transgenic events through rooting. The step-by-step scheme of non-selective genetic transformation of potato is presented in Figure 1.
Concurrently, the antibiotic-assisted genetic transformation was carried out. After co-cultivation with Agrobacterium in dark for 3 days, a portion of intermodal segments was transferred to the corresponding regeneration medium supplemented with kanamycin (50 mg L−1). Shoot regeneration and transgenic plant selection were then performed as described [32]. To avoid duplication of transgenic lines, only one kanamycin-resistant shoot was collected from each explant.
2.3. GFP Monitoring
Visual screening for transient and stable T-DNA incorporation was conducted using a ZEISS SteREO Discovery.V12 microscope equipped with a PentaFluar S 120 vertical illuminator. In the observations, two commercially available filter sets, namely 38 GFP BP (EX BP 470/40, BS FT 495, EM BP 525/50) and 57 GFP BP (EX BP 470/40, BS FT 495, EM LP 550) from Carl Zeiss MicroImaging GmbH (Germany) were used.
2.4. Generation of Marker-Free Potato Plants for the Silencing of the SteIF4E1 and SteIF4E2 Genes
The agrobacterial strain AGL0 harbouring the pWS-E1-Lhca3 vector designed for the post-transcriptional silencing of the SteIF4E1 and SteIF4E2 genes of potato was used [30]. The expression cassette included the hpRNAi structure, consisted of inverted fragments of SteIF4E1 gene separated by the Strbsc1 intron and controlled by the StLhca3 promoter and terminator. Notably, the T-DNA region of pWS-E1-Lhca3 was not included any selective genes. The agrobacterium-mediated transformation of cv. ‘La Strada’ was carried out as specified in section 2.2. The regeneration media for plantlet development was supplemented with 3 mg L−1 Zea-R, 2 mg L−1 IAA and 1 mg L−1 GA. Plantlets that within the initial 40 days after inoculation were discarded. Two weeks later, all plantlets developed on the explants were cut and transferred to culture vessels (ten plantlets per vessel) containing phytohormone–free MS medium supplemented with 250 mg L−1 cefotaxime. The explants were discarded. Upon the regeneration of plants to a length of 5-8 cm, the two upper leaves of each plant grown in a single vessel were harvested and pooled together for isolation of genomic DNA. The presence of hairpin constructs in the DNA mix was validated using PCR with the primers and conditions described previously [30]. In cases where specific band amplification was observed, each plant from a positive vessel was individually cultivated, sampled for total DNA extraction, and subjected to PCR to identify transgenic events.
2.5. Molecular Analysis
To analyze integrated T-DNA copies, Southern hybridization was conducted. For the blot analysis, 30 μg of potato genomic DNA was digested overnight at 37°C with 60U HindIII which cleaves the T-DNA of pBIN-mGFP5-ER at a single position (5′ end of the CaMV 35S promoter). Subsequently, the DNA was separated through gel electrophoresis in 0.9% agarose, and then transferred and immobilized onto Hybond N+ membrane (GE Healthcare, Amersham Bioscience, Amersham, UK). Membranes were probed with alkaline phosphatase-labeled probes and detected using CDP-Star detection reagent (Amersham CDP-Star Detection reagent, GE Healthcare) as previuously described [30]. Hybridizing bands were visualized on X-ray film (Retina XBE blue sensitive, Carestream Health INC., NY, United States) at room temperature for 24 h. The DNA probe (711bp) for hybridization was synthesized via PCR using specific primers for the coding region of the GFP gene (forward, 5’-AGTAAAGGAGAAGAACTTTTCACTGGAGTT-3’; reverse, 5’-TTTGTATAGTTCATCCATGCCATGTGT-3’) and labbeled with the alkaline phosphatase using Amersham Gene Images AlkPhos Direct Labelling and Detection System (GE Healthcare, Amersham Bioscience, Amersham, UK).
For qRT-PCR analysis, total RNA was extracted from in vitro grown plants as described [34]. First-strand cDNA was synthesized from 5 μg of each DNase-treated RNA sample using RevertAid Reverse Transcriptase (Thermo Scientific). Gene expression analysis was conducted using Quantitative real-time reverse-transcription PCR (qRT-PCR) with QuantStudio™ 5 Real-Time PCR Cycler (Thermo Fisher Scientific), as previously described [30]. The normalization of the the SteIF4E1 and SteIF4E2 values was performed using the expression levels of the ef1α, a housekeeping potato gene. All primer pairs and analysis conditions were detailed in the previous publication [29]. The reported values represent the mean ± standard deviations (SD) of four biological replicates with three technical replicates.
2.6. Statistical Analysis
The data presented are the mean ± SD. Significant differences were determined via t-test with a threshold of p<0.05. Intra-experimental data were analyzed with Statistica10 software (©StatSoft Inc) using analysis of variance (ANOVA), followed by Tukey's HSD test.
3. Results
3.1. Efficiency of Adventitious Plant Regeneration in Potaoto Cultivars
This experiment describes the regenerative capacity of potato cultivars in producing multiple shoots using zeatin-enriched regeneration media. Each analyzed cultivars was tested on four phytohormonal combinations, with the most favorable combination for adventitious shoot regeneration being determined (data not shown). Summarized results are presented Table 1. All ten cultivars demonstrated the ability to regenerate plantlets from cultured internodal segments. However, the percentage of explants producing shoots varied from 58.75 (‘Indigo’) to 100% (#12-22-129, ‘Pirol’ and #10-9-3). Eight cultivars exhibited an efficiency of plantlet regeneration higher than 93%. Number of regenerated shoots per explant significantly varied depending the genotype. Particularly, ‘Indigo’ (2.4 plantlets per explant) and ‘Manhattan’ (3.5 plantlets per explant) displayed the lowest ability, while ‘Chicago’ and #12-22-129 produced an average of more than 15 shoots per initial explant (Table 1).
The experiment revealed the noticeable difference among cultivars in their timing abilities for generating plantlets. Nodal explants of #12-22-129 and ‘Chicago’ promptly responded to phytohormonal induction, initiating formation of adventitious shoots during the second week following culture initiation. By the end of third week, they notably produced visible plantlets ranging from 0.5 to 2.0 cm in length (Figure 2). Furthermore, the explants of #12-22-129 and ‘Chicago’ were characterized by the emergence of shoots at the both nodal excision sites, ultimately yielding a higher number of shoots compared to other cultivars. In contrast, the explants of ‘Pirol’, ‘Indigo’, ‘Lion Heart’ and #10-9-3 demonstrated a significant delay in plantlet formation, as shoots of a comparable size only emerged at 6th week following culture initiation (Figure 2). Meanwhile, ‘La Strada’, ‘Manhattan’, #10-10-10 and #12-36-42 exhibited an intermediate timing pattern (Figure 2), alongside varying number of regenerated adventitious shoot per explant (Table 1).
Based on the result, five potato cultivars were selected for the non-selective transformation experiment. We used cultivars exhibiting varied timing of plantlets emergence (ranging from early to late), which demonstrated a higher number of regenerating shoots and a higher percentage of explant producing plantlets. These cultivars included #12-22-129 (early type of regeneration), ‘Chicago’ (early/intermediate), ‘La Strada’ (intermediate), #12-36-42 (intermediate/late) and ‘Pirol’ (late).
3.2. Agrobacterium-mediated Genetic Transformation of Potato Without Antibiotic–Assited Selection
In this experiment, nodal explant were transformed with the pBIN-mGFP5-ER vector contained the sequence of green fluorescent protein GFP, allowing for easy monitoring of the generation of transgenic events from single cells to plantlets.
3.2.1. Efficiency of transient expression under selective- and non-selective cultivation
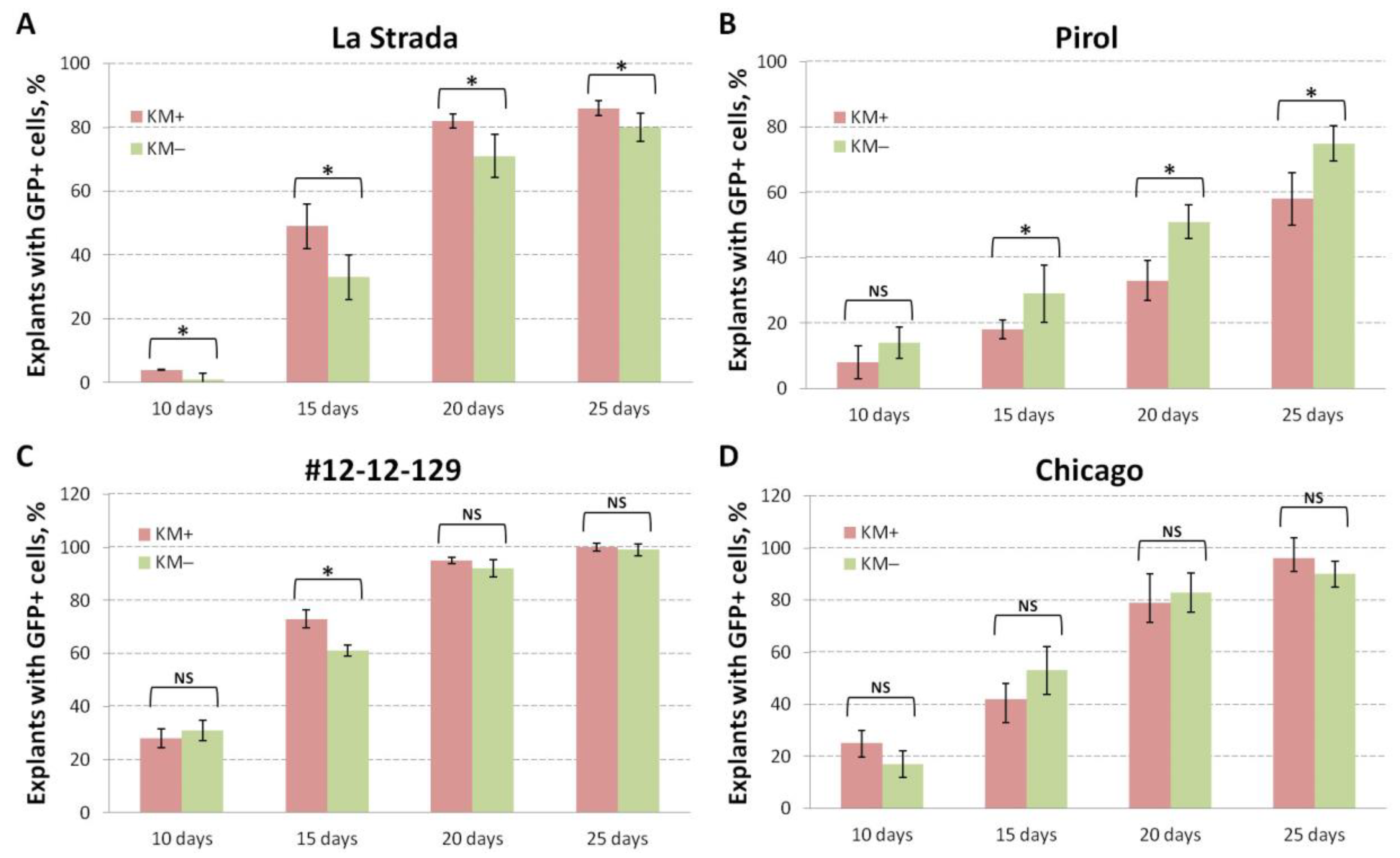
To gain initial insights into the potential of potato cultivars to produce transgenic shoots without selective pressure, transient GFP expression was monitored over four weeks following co-cultivation of nodal segments with agrobactera. The impact of absence or presence of selective pressure was investigated by culturing explants of five genotypes in parallel, both on a medium containing no selective compound and on a medium supplemented with kanamicyn. A significant genotype-dependent effect was observed (Figure 3). Cultivars with earlier regeneration type, such as ‘Chicago’ and #12-22-129 (Figure 3C, 3D), showed earlier transient expression in cells of the explant cuts, particularly during the first two first week of culture, compared to ‘La Strada’ (Figure 3A), ‘Pirol’ (Figure 3B) and #12-36-42. On the whole, nearly all explants (>90%) of ‘Chicago’ and #12-22-129 showcased single cells or cell conglomerates expressing GFP throughout observation poeriod (25th day). The percentage of GFP positive explants in other cultivars was comparatively lower: 84-89% in #12-36-42, 80-86% in ‘Las Strada’ and 58-75% in ‘Pirol’.
The presence of the antibiotic in the medium noticeably effected the transient expression in two cultivars. The percentage of ‘La Strada’ explants exhibiting GFP expression was higher in the presence of kanamycin throughout the observation period, especially during at the first twoe weeks following inoculation (Figure 3A). In contrast, the cultivation of nodal explants of ‘Pirol’ without antibiotic proved beneficial, as a higher portion of explants displayed GFP expression (Figure 3B). However, there was no clear effect of selective pressure in other three cultivars, as an equivalent number of explants exhibited cell fluorescence during throughout the observation period, or the difference was not statistically proved (Figure 3C,D).
Figure 3.
The percentage of explants displaying the GFP expression in cells of potato cultivars, including ‘La Strada’ (a), ‘Pirol’ (b), # 12-12-129 (c), and ‘Chicago’ (d) under selective- (Km+) and non-selective (KM-) cultivation. “*” indicate statistically significant differences calculated according t-test ( p ≤ 0.05), ns - non-significant.
Figure 3.
The percentage of explants displaying the GFP expression in cells of potato cultivars, including ‘La Strada’ (a), ‘Pirol’ (b), # 12-12-129 (c), and ‘Chicago’ (d) under selective- (Km+) and non-selective (KM-) cultivation. “*” indicate statistically significant differences calculated according t-test ( p ≤ 0.05), ns - non-significant.
3.2.2. Efficiency of regeneration of GFP positive transgenic plants under non-selective and selective cultivation
In the absence of antibiotic in regeneration medium, the emergence of non-transgenic, transgenic and chimerical buds was observed on the cut ends of explants during the initial subcultures (Figure 4A,B). These buds easily progressed into shoots, leading to the development of both chimeric and transgenic individuals among the predominant non-transgenic plantlets, as confirmed by GFP observation. Chimeric plantlets exhibited sectored GFP expressing tissues (Figure 4C), while completely transgenic plants demonstrated clear green fluorescence throughout the entire plantlet, inclusive of stem, petioles and leaves (Figure 4D,E). Typically, only a single GFP expressing plantlet was selected from a single explant during observation, albeit occasionally two independent GFP positive plantlets were observed simultaneously among the cluster of non-transgenic shoots (Figure 4F).
Each chimeric and fully transgenic plantlet was regularly separated from the cultivated explants and independently proliferated on a kanamycin-free medium. At this stage, it became challenging to monitor the in vitro plant growth with GFP observation due to technical limitations (maintaining aseptic conditions became difficult with increased sizes) and physical issues (GFP fluorescence diminished in mature tissue and was obscured by chlorophyll fluorescence and tissues autoflorescence). As the T-DNA of the pBIN-mGFP5-ER vector also contains the NPTII gene, after several rounds of sub-culturing without selective pressure, all the putative transgenic plants were subsequently tested by rooting in medium supplemented with a lethal dose of kanamicyn (100 mg L−1). The presence of the selective antibiotic prevented the growth and rooting of non-transgenic plants (Figure S2). The rooting was used with the aim to clarify the transgenic status of plantles, after subculturing and proliferation without addition of selective compound. The results of two independent experiments, each involved 50-75 explants for cultivar, are summarized in Table 2.
In our experiment, we employed a 'blind' approach, where GFP positive plantlets (both chimeric and non-chimeric) were not specifically tracked in relation to the mother explant. For this reason, the frequency of transformation was defined to the total number of detached plantlets at the end of experiment. The efficiency of transformation varied among the analysed cultivars, ranging from 0.9% (‘Chiacago’) to 2.6% (#12-36-42). Upon rooting analysis, we transformed into non-transgenic plants during subsequent antibiotic-free sub-cultivations. Specifically, from half (Pirol, #12-22-129 and #12-36-42) to third (‘Chicago’ and ‘La Strada’) of the plantlets, which initially exhibited GFP expression in tissues at the point of detachment, subsequently became non-transgenic (Table 2).
It's noteworthy that, the number of explants that initially produced GFP positive transgenic buds exceeded the resulting number of transgenic shoots (Table 2). Considering the high portion of chimeric escapes, it's plausible that some of the generated stable transformants are duplicates, potentially stemming from the same transgenic cells aggregate of explant at various stages of experiments. To address this, we carried out a molecular characterization of several transgenic events through Southern blot analysis. For instance, the analysis of 13 out of 15 transgenic plants of ‘Chicago’ using a GFP probe revealed the disparate pattern of T-DNA insertions (Figure 5). Similarly, the analysis of insertion patterns in transgenic plants of ‘Pirol’, which we initially suspected to be duplicates due to their origin from the same explants, confirmed their independent origin (Figure 5).
In a parallel experiment, we also performed kanamicin-assisted genetic transformation of the same cultivars to compare the efficiency of transgenic plants production. Monitoring the transformation process revealed the significant suppression of plant regeneration in cultured explants, alongside the emphasized formation of aggregates of transgenic cells and callus, which subsequently produced single and multiple shoots with distinct GFP expression (Figure S3). When measuring the efficiency of genetic transformation in marker-assisted selection, defined as the percentage of initially inoculated explants that produced transgenic shoots, our analysis indicated markedly higher transformation abilities in potato cultivars under selective pressure compared to the non-selective approach (Table 2). Contrary to the non-selective approach, we observed a substantial genotypic effect. While two cultivars, #12-22-129 and ‘Chicago’, showed 100% and 98% efficiency of genetic transformation, respectively, ‘Pirol’ and ‘La Strada’ yielded transgenic events at half the efficiency, specifically 44% and 47%, respectively.
3.2.3. Impact of the regeneration type on the discovering of GFP positive plants under non-selective pressure
In the present study, we found that the genotype-specific timing of plantlet regeneration significantly influenced the discovering of the GFP positive plants. In Figure 5 we have combined the number of detached plantles with the occurrence of explants formed GFP+ buds, along with the number of detached chimeric/non-chimeric plantlets throughout the entire regeneration stage. The results for two contrasting genotypes are presented. Considering the regeneration experiments, #12-22-129, characterized as a 'fast' regenerating cultivar, produced shoots much earlier than the ‘late’ regenerating cultivar ’Pirol’. By the 50th day post agrobacterial inoculation, almost 100 plantlets were detached from Pirol’ explants, while with #12-22-129, over half of all regenerated explants (nearly 1000 shoots) had already been collected. observed in #12-22-129 explants within the second week of cultivation, whereas in 'Pirol', they emerged 10-15 days later. After 45 days of cultivation, no new explants with GFP positive buds were found on #12-22-129 explants, whereas in 'Pirol', the number of explants with such buds continued to appear until the 60th day of the experiment (Figure 5).
The dynamics of detaching GFP expressing plantlets revealed ‘empty’ periods of culture, when most picked plantlets were non-transgenic (Figure 6). Consequently, in the ‘early’ cultivar #12-22-129, the majority of GFP+ shoots (approximately 75%) were discovered between 30 and 60 days of cultivation. In contrast, about three-quarters of putative transgenic plantlets of the ‘late’ cultivar ‘Pirol’ were generated by cultivated explants between 50 and 80 days of the experiment (Figure 6). Analyzing cultivars with an ‘intermediate’ type of regeneration also highlighted that earlier and the later periods of plants regeneration were ineffective in collecting GFP expressing plantlets. For instance, in ‘Chicago’, a cultivar characterized by an early/intermediate type of regeneration, the majority of putative transgenic events (over 80%) were collected between 30 and 60 days of culture. In ‘La Strada’ and #12-36-42, the formation of GFP positive plantlets was more extensive and consistent between 40 to 80 days, although half of putative transgenic plants were detached between 60 and 80 days of culture
3.3. Marker-Free Transformation of Potato cv. ‘La Stada’ for Silencing of eIF4E1 and eIF4E2 genes
In this experiment, prompted by a partner company's request, we endeavored to apply the experience of non-selective genetic transformation for producing transgenic plants of 'La Strada' using the vector pWS-E1-Lhca3 [30], which devoid any marker gene sequence in T-DNA. As selective substances couldn't be utilized for screening, the molecular assessment of hundreds of regenerated plantlets became essential to detect a few stable transgenic events. Considering that ‘La Strada’ is characterized as a cultivar of ‘intermediate’ regeneration type, we made the decision to discard all plantlets produced by explants until the 40th day post co-cultivation to reduce the number of items subjected to analysis. Subsequently, we allowed the novel plantlets to grow for the next two weeks, then collected all regenerants at once and transferred them to multiplication medium for elongation. No additional plantlets were collected thereafter to avoid the possibility of duplicating transgenic events. In total, 368 plantlets were taken from 51 inoculated explants. The plantlets were cultivated in groups of 10 plants in plastic containers until they grew sufficiently for upper leaves to be collected for DNA extraction. To minimize molecular handling, two leaves from each plantlet grown in a single culture vessel were collected under aseptic conditions and mixed into a single sample for DNA extraction. The PCR analysis of 38 samples, using a pair of primers corresponding to the chimeric union of the left arm and the rbsc1 intron of the hairpin construct of pWS-E1-Lhca3, led to the detecting of a specific band in two DNA mixes (Figure 7).
Following this, the second round of PCR analysis involved new DNA samples extracted from leaves of twenty plants initially cultured in two ‘positive’ vessels, ultimately identifying two transgenic events (Figure 7). Since the discovered plants were collected from independent explants (with all collected regenerants tracked relative to their mother explants), the efficiency of genetic transformation in this experiment was determined to be 3.9% (equating to two hairpin positive events per 51 explants). When defining the efficiency of marker-free transformation as the percentage of PCR positive plants to all collected plants, the transformation rate appeared to be 0.5%.
Following the multiplication of two transgenic plants and after several rounds of sub-culturing, their total RNA was extracted to confirm the activity of the introduced expression cassette. The introduced hairpin construct was specifically engineered to to downregulate the endogenous expression of two homological genes encoding the eukaryotic translation initiation factors eIF4E1 and eIF4E2 of potato. To assess the impact, the level of expression of the two targeted genes was determined by qRT-PCR. The analysis yielded compelling results that discovered plants are not chimeric, as a significant decrease in the amount of RNA transcript specific for SteIF4E1 and SteIF4E2 compared to non-transgenic 'La Strada' (Figure 8). On average, the endogenous level of SteIF4E1 gene expression in non-transgenic plants was over 70 times higher than that in the samples of two transgenic lines. The successful introduction and expression of the marker-free hairpin construct also led to a remarkable 7-9 fold decrease in transcript levels of the endogenous SteIF4E2 gene in both transgenic lines (Figure 8).
4. Discussion
The successful genetic transformation of potato hinges on efficient plant regeneration protocols and optimized methodologies for recognizing transgenic tissues. An increasing number of scientific reports on genetically engineered potato published since the late 1980s, alongside the worldwide commercial releases of GM varieties, indicate that potato is a relevant crop for improvement through biotechnology [2,3,4,5]. Given the increased likelihood of obtaining transgenic events when more cells are capable of differentiating into shoots, we initially screened ten potato cultivars for their regeneration potential to select the most suitable ones for antibiotic-free transformations. Typically, a two-stage regeneration protocol is used in potato, where the stage of shoots regeneration follows the stage of callus induction [4]. Cytokinins play a paramount role in initiating morphogenesis in potato. Although BAP (6-benzylaminopurine), kinetin and TDZ (thidiazuron) are reported to stimulate regeneration in potato, zeatin currently is known as the most popular cytokinin for generating transgenic potato plants. Most transformation protocols involve supplementing the regeneration medium with 1-5 mg L−1 of zeatin along with low level of auxin.
In the present study various zeatin-containing variant of the regeneration medium combined with different auxins were used, including twin variants where zeatin was replaced with zeatin-ribozide, which has been reported to exhibit superior regeneration frequency and even enable one-step transformation of potato [34]. The tested potato cultivars responded positively to various plant phytohormone combinations, with two variants proving to be more effective in practice. The combination of 3 mg L−1 Zea and 0.5 mg IAA L−1 was found better for five cultivars, while the combination of the same level of zeatin along with a significantly higher concentration of IAA (2 mg L−1) and the additional incorporation of gibberellic acid (1 mg L−1) to support the enhanced outgrowth of morphogenic buds was essential for successful regeneration in ‘Chicago’, ‘Lion Heart’, and #10-9-3. Moreover, while the replacement by the more expensive zeatin-ribozide generally had no significant effect in listed above cultivars, it proved more successful in two other cultivars ('Indigo' and 'La Strada'). Nonetheless zeatin-ribozide was more successful in two cultivars (‘Indigo’, ‘La Strada’). This observation aligns with the early notion that application of zeatin-ribozide serves as viable alternative for specific cultivars to achieve higher efficiency of regeneration, an important prerequisite for successful genetic transformation [34].
The experiments revealed the pivotal importance of the timeline of plantlets regeneration from internodal explants. We observed substantial disparities between genotypes in the speed of adventitious shoot formation. Certain cultivars, such as ‘Chicago’ and #12-22-129, initiated the development of plantlets much earlier than others. In contrast, the cultivars characterized by a late regeneration, such as Pirol’, #10-9-3, ‘Indigo’ and ‘Lion Heart’ started the development of plantlets when the cultivars characterized by an early regeneration had already produced a maximum number of shoots. This genotype-specific property can be taken into account when potato varieties with different types of regeneration periods undergo marker-free transformation. Our results suggest that ‘early regenerative’ cultivars produced the majority of transgenic plants (approximately 75%) between 30 and 60 days post inoculation with Agrobacterium, whereas the ‘late’ cultivars preferably generated transgenic events during the 50-80 days of cultivation. Similarly, cultivars with an intermediate regeneration timeline predominantly generate transgenic plants between 40 and 70 days after co-cultivation with Agrobacterium. These findings could be leveraged to potentially reduce the number of analysed regenerated plantlets, given that numerous manipulations such as DNA extraction, PCR analysis, and prolonged in vitro cultivation of each regenerated plant are necessary to detect transgenics. Our data are consistent with those reported by Jo et al. 2014 [28], who, under non-selective conditions, also observed differences between three potato genotypes in the number of PCR positive plants over the number of tested regenerated shoots. In their study, shoots were only collected at three-week intervals, and only a portion of the generated shoots were analyzed. However, the timeline-dependant effect was clearly observed, as a higher portion of PCR+ plants of ‘Potae9’ and ‘Bintje’ was found between 31-50 and 31-70 days, respectively, while explants of ‘Atlantic’ produced a significant portion of transgenic plants during the 50-90 days of culture [28].
The results we have obtained here align with previous studies, suggesting that antibiotic-free transformation is less dependent on genotype compared to antibiotic-assisted protocols [26,28]. In our findings, the efficiency of marker-assisted transformation displayed substantial genotype dependency and varied significantly from 44 to 100% (Table 2). Conversely, it's challenging to argue for a prominent influence of genotype under non-selective conditions, since the transformation efficiency varied within a narrow range of 0.9 to 2.7%. A similar genotype independent trend was observed in marker-less transformation of three potato cultivars with varying genetic background, where the efficiency varied between 0.6 and 2.4% [28]. Notably, an average marker-free transformation efficiency of 2.2% was reported for cv. ‘Superior’ in another study [31], in which leaf explants were used instead of intermodal segments for generation of plantlets. Moreover, in report of Vetten et al. [27], the highly transformable cultivar 'Carnico' exhibited slightly higher efficacy, achieving 4.5% in marker-less transformation, while the cultivars with lower marker-assisted transformation abilities demonstrated a lower percentage of PCR-positive shoots recovery (0.6 – 2.4%). In the present study, the lowest antibiotic-free transformation efficiency was detected in ‘Chicago’ (0.9%), despite the variety demonstrating an extremely high efficiency in kanamycin-assisted selection (100%). When considering transformation efficiency, it's important to note that marker-free selection is calculated relative to the total number of plants analyzed, whereas antibiotic-assisted transformation is defined as the proportion of explants producing transgenic plants. Due to this distinction, the efficiency of the same varieties when using the two protocols is not directly comparable. Furthermore, the ability to generate more (or fewer) plantlets from the same number of cultured cells may explain why a specific cultivar exhibits reduced marker-free transformation efficiency compared to others.
While marker-less transformation could be perceived as a simpler approach, low transformation percentages are commonly reported [23,24,25,26,27,28,29,30], because it is assumed that without the use of selective agents, the proportion of Agrobacterium-treated cells that can become transgenic is limited. Surprisingly, in our study, the number of explants displayed GFP expression until 30 days after agrobacterial inoculation was either equal to or even higher without the selective pressure in four potato cultivars. Notably, only cv. ‘La Strada’ demonstrated better transient expression when kanamycin-assisted selection was used; however, the efficiency of stable genetic transformation was only 47%, lower than in the other genotypes. These findings suggest that a high level of transient expression does not necessarily lead to high efficiency of transformation in potato. Nevertheless, the presence of explants with high transient expression, regardless of selective pressure, makes the generation of fully transgenic plants in the absence of marker-assisted selection a realistic prospect.
In our experiments, the detection of putative transgenic plants during the stage of kanamycin-free regeneration facilitated by a relatively straightforward screening process based on GFP fluorescence. Given the presence of the NPTII gene in the transferred expression cassette, transgenic events were identified after the finishing of experiments through rooting in the presence of kanamycin.
Following the established timeline of transgenic plants emergence, we endeavored to adopt a 'blind' detection approach for ‘La Strada’ when a minimal gene expression cassette was used for producing transgenic plants. The results, based on the PCR and qp-PCR analysis, underscored the reproducibility of the suggested timeline-based transformation approach, as independent marker-free transgenic plants were successfully produced according to the PCR and qp-PCR analyses. Despite a significant reduction in the number of inoculated explants and the one-time collection of plants regenerated between 40 and 55 days, we were able to generate transgenic plants with a frequency of 0.5% (per all analyzed plants) or 1.9% (per inoculated explants). Notably, a similar marker-less transformation efficiency of 0.5% was previously achieved when employing the timeline-based approach to generate intragenic PPV-resistant plants of the cv. 'Pirol' using the same minimal gene expression cassette [30]. Considering that the transgenic plants of 'La Strada' produced in the present study demonstrated a significantly higher RNAi-mediated silencing of target genes than the resistant intragenic line of 'Pirol', we hope to further co confirm the viral resistance in both new lines during a greenhouse test.
While the ability to generate transgenic elite potato cultivars without using selectable marker genes may not be as effective as marker-assisted selection, we believe it will significantly facilitate the potential commercial release of new biotech potato varieties produced via cisgenic and intragenic strategies of genetic transformation. This novel advancement offers promising prospects for the development and deployment of biotech potato, underlining the ongoing advancements in genetic engineering and crop improvement.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org, Figure S1: The percentage of explants displaying the GFP expression in cells of potato cv. #12-36-42 under selective- (kanamycin +) and non-selective (kanamycin -) cultivation. Figure S2: Identification of transgenic events by in vitro rooting of putative transgenic plants (cv. #12-36-42) on the medium, supplemented with a lethal dose of kanamicyn (100 mg L−1). Figure S3: Regeneration of transgenic potato plants under kanamycin assisted selection. Figure S4: Genotype-dependant timeline for formation of GFP expressed buds and plantlets of ‘Chicago’ (early/intermediate type of regeneration), ‘La Srada’ (intermediate type of regeneration) and #12-36-42 (intermediate/late type of regeneration) under non selective conditions in comparison with the all regenerated shoots.
Author Contributions
D.M. and S.D. conceived the research, D.M, A.O., A.K., T.S., V.T. and A.S.P carried out the experiments, S.D supervised, validated, and acquired financing; D.M. wrote the manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the Russian Science Foundation, grant No 22-14-00118.
Data Availability Statement
The data presented here are available upon request from the corresponding authors.
Acknowledgments
The authors would like to thank the staff of the large-scale research facilities “FITOTRON” (registration number 2-2.9) of the Branch of Shemyakin-Ovchinnikov Institute of Bioorganic Chemistry for the growing of the donor and regenerated plants.
Conflicts of Interest
The authors declare no conflict of interest.
References
- An, G.; Watson, B.D.; Chiang, C.C. Transformation of Tobacco, Tomato, Potato, and Arabidopsis thaliana Using a Binary Ti Vector System. Plant Physiol. 1986, 81, 301–305. [Google Scholar] [CrossRef]
- Si, H.; Zhang, N.; Tang, X.; Yang, J.; Wen, Y.; Wang, L.; Zhou, X. Transgenic Research in Tuber and Root Crops. In Genetic Engineering of Horticultural Crops; Elsevier, 2018; pp. 225–248. ISBN 9780128104392. [Google Scholar]
- Del Mar Martínez-Prada, M.; Curtin, S.J.; Gutiérrez-González, J.J. Potato improvement through genetic engineering. GM Crops Food 2021, 12, 479–496. [Google Scholar] [CrossRef] [PubMed]
- Nahirñak, V.; Almasia, N.I.; González, M.N.; Massa, G.A.; Décima Oneto, C.A.; Feingold, S.E.; Hopp, H.E.; Vazquez Rovere, C. State of the Art of Genetic Engineering in Potato: From the First Report to Its Future Potential. Front. Plant Sci. 2021, 12, 768233. [Google Scholar] [CrossRef] [PubMed]
- Hameed, A.; Zaidi, S.S.-E.-A.; Shakir, S.; Mansoor, S. Applications of New Breeding Technologies for Potato Improvement. Front. Plant Sci. 2018, 9, 925. [Google Scholar] [CrossRef]
- De Steur, H.; Van Loo, E.J.; Maes, J.; Gheysen, G.; Verbeke, W. Farmers’ Willingness to Adopt Late Blight-Resistant Genetically Modified Potatoes. Agronomy 2019, 9, 280. [Google Scholar] [CrossRef]
- Tuteja, N.; Verma, S.; Sahoo, R.K.; Raveendar, S.; Reddy, I.N.B.L. Recent advances in development of marker-free transgenic plants: regulation and biosafety concern. J. Biosci. 2012, 37, 167–197. [Google Scholar] [CrossRef] [PubMed]
- Breyer, D.; Kopertekh, L.; Reheul, D. Alternatives to Antibiotic Resistance Marker Genes for In Vitro Selection of Genetically Modified Plants – Scientific Developments, Current Use, Operational Access and Biosafety Considerations. Critical Reviews in Plant Sciences 2014, 33, 286–330. [Google Scholar] [CrossRef]
- Singh, R.; Kaur, N.; Praba, U.P.; Kaur, G.; Tanin, M.J.; Kumar, P.; Neelam, K.; Sandhu, J.S.; Vikal, Y. A Prospective Review on Selectable Marker-Free Genome Engineered Rice: Past, Present and Future Scientific Realm. Front. Genet. 2022, 13, 882836. [Google Scholar] [CrossRef]
- Sharma, U.; Kaur, A.; Sandhu, J.S. Production of Marker-Free Transgenic Plants. In Agricultural Biotechnology: Latest Research and Trends; Kumar Srivastava, D., Kumar Thakur, A., Kumar, P., Eds.; Springer Nature Singapore: Singapore, 2021; pp. 223–251. ISBN 978-981-16-2338-7. [Google Scholar]
- Matheka, J.M.; Anami, S.; Gethi, J.; Omer, R.A.; Alakonya, A.; Machuka, J.; Runo, S. A new double right border binary vector for producing marker-free transgenic plants. BMC Res. Notes 2013, 6, 448. [Google Scholar] [CrossRef]
- Petri, C.; López-Noguera, S.; Wang, H.; García-Almodóvar, C.; Alburquerque, N.; Burgos, L. A chemical-inducible Cre-LoxP system allows for elimination of selection marker genes in transgenic apricot. Plant Cell Tiss Organ Cult 2012, 110, 337–346. [Google Scholar] [CrossRef]
- Zhang, Y.; Li, H.; Ouyang, B.; Lu, Y.; Ye, Z. Chemical-induced autoexcision of selectable markers in elite tomato plants transformed with a gene conferring resistance to lepidopteran insects. Biotechnol. Lett. 2006, 28, 1247–1253. [Google Scholar] [CrossRef] [PubMed]
- Peng, A.; Xu, L.; He, Y.; Lei, T.; Yao, L.; Chen, S.; Zou, X. Efficient production of marker-free transgenic ‘Tarocco’ blood orange (Citrus sinensis Osbeck) with enhanced resistance to citrus canker using a Cre/loxP site-recombination system. Plant Cell Tiss Organ Cult 2015, 123, 1–13. [Google Scholar] [CrossRef]
- Schaart, J.G.; Krens, F.A.; Pelgrom, K.T.B.; Mendes, O.; Rouwendal, G.J.A. Effective production of marker-free transgenic strawberry plants using inducible site-specific recombination and a bifunctional selectable marker gene. Plant Biotechnol. J. 2004, 2, 233–240. [Google Scholar] [CrossRef] [PubMed]
- Vanblaere, T.; Szankowski, I.; Schaart, J.; Schouten, H.; Flachowsky, H.; Broggini, G.A.L.; Gessler, C. The development of a cisgenic apple plant. J. Biotechnol. 2011, 154, 304–311. [Google Scholar] [CrossRef] [PubMed]
- Timerbaev, V.; Mitiouchkina, T.; Pushin, A.; Dolgov, S. Production of Marker-Free Apple Plants Expressing the Supersweet Protein Gene Driven by Plant Promoter. Front. Plant Sci. 2019, 10, 388. [Google Scholar] [CrossRef] [PubMed]
- Pompili, V.; Dalla Costa, L.; Piazza, S.; Pindo, M.; Malnoy, M. Reduced fire blight susceptibility in apple cultivars using a high-efficiency CRISPR/Cas9-FLP/FRT-based gene editing system. Plant Biotechnol. J. 2020, 18, 845–858. [Google Scholar] [CrossRef]
- Righetti, L.; Djennane, S.; Berthelot, P.; Cournol, R.; Wilmot, N.; Loridon, K.; Vergne, E.; Chevreau, E. Elimination of the nptII marker gene in transgenic apple and pear with a chemically inducible R/Rs recombinase. Plant Cell Tiss Organ Cult 2014, 117, 335–348. [Google Scholar] [CrossRef]
- Orbegozo, J.; Solorzano, D.; Cuellar, W.J.; Bartolini, I.; Roman, M.L.; Ghislain, M.; Kreuze, J. Marker-free PLRV resistant potato mediated by Cre-loxP excision and RNAi. Transgenic Res. 2016, 25, 813–828. [Google Scholar] [CrossRef]
- Kondrák, M.; van der Meer, I.M.; Bánfalvi, Z. Generation of marker- and backbone-free transgenic potatoes by site-specific recombination and a bi-functional marker gene in a non-regular one-border agrobacterium transformation vector. Transgenic Res. 2006, 15, 729–737. [Google Scholar] [CrossRef]
- Khan, R.S.; Ntui, V.O.; Chin, D.P.; Nakamura, I.; Mii, M. Production of marker-free disease-resistant potato using isopentenyl transferase gene as a positive selection marker. Plant Cell Rep. 2011, 30, 587–597. [Google Scholar] [CrossRef]
- Ferradini, N.; Nicolia, A.; Capomaccio, S.; Veronesi, F.; Rosellini, D. Assessment of simple marker-free genetic transformation techniques in alfalfa. Plant Cell Rep. 2011, 30, 1991–2000. [Google Scholar] [CrossRef] [PubMed]
- Bhatnagar, M.; Prasad, K.; Bhatnagar-Mathur, P.; Narasu, M.L.; Waliyar, F.; Sharma, K.K. An efficient method for the production of marker-free transgenic plants of peanut (Arachis hypogaea L.). Plant Cell Rep. 2010, 29, 495–502. [Google Scholar] [CrossRef] [PubMed]
- Li, B.; Xie, C.; Qiu, H. Production of selectable marker-free transgenic tobacco plants using a non-selection approach: chimerism or escape, transgene inheritance, and efficiency. Plant Cell Rep. 2009, 28, 373–386. [Google Scholar] [CrossRef] [PubMed]
- Richardson, T.; Thistleton, J.; Higgins, T.J.; Howitt, C.; Ayliffe, M. Efficient Agrobacterium transformation of elite wheat germplasm without selection. Plant Cell Tiss Organ Cult 2014, 119, 647–659. [Google Scholar] [CrossRef]
- Vetten, N. de; Wolters, A.-M.; Raemakers, K.; van der Meer, I.; ter Stege, R.; Heeres, E.; Heeres, P.; Visser, R. A transformation method for obtaining marker-free plants of a cross-pollinating and vegetatively propagated crop. Nat. Biotechnol. 2003, 21, 439–442. [Google Scholar] [CrossRef] [PubMed]
- Jo, K.-R.; Kim, C.-J.; Kim, S.-J.; Kim, T.-Y.; Bergervoet, M.; Jongsma, M.A.; Visser, R.G.F.; Jacobsen, E.; Vossen, J.H. Development of late blight resistant potatoes by cisgene stacking. BMC Biotechnol. 2014, 14, 50. [Google Scholar] [CrossRef] [PubMed]
- Bai, Y.; Guo, Z.; Wang, X.; Bai, D.; Zhang, W. Generation of double-virus-resistant marker-free transgenic potato plants. Progress in Natural Science 2009, 19, 543–548. [Google Scholar] [CrossRef]
- Miroshnichenko, D.; Timerbaev, V.; Okuneva, A.; Klementyeva, A.; Sidorova, T.; Pushin, A.; Dolgov, S. Enhancement of resistance to PVY in intragenic marker-free potato plants by RNAi-mediated silencing of eIF4E translation initiation factors. Plant Cell Tiss Organ Cult 2020, 140, 691–705. [Google Scholar] [CrossRef]
- Ahmad, R.; Kim, Y.-H.; Kim, M.D.; Phung, M.-N.; Chung, W.-I.; Lee, H.-S.; Kwak, S.-S.; Kwon, S.-Y. Development of selection marker-free transgenic potato plants with enhanced tolerance to oxidative stress. J. Plant Biol. 2008, 51, 401–407. [Google Scholar] [CrossRef]
- Haseloff, J.; Siemering, K.R. The uses of green fluorescent protein in plants. Methods Biochem. Anal. 2006, 47, 259–284. [Google Scholar]
- Miroshnichenko, D.; Firsov, A.; Timerbaev, V.; Kozlov, O.; Klementyeva, A.; Shaloiko, L.; Dolgov, S. Evaluation of Plant-Derived Promoters for Constitutive and Tissue-Specific Gene Expression in Potato. Plants (Basel) 2020, 9. [Google Scholar] [CrossRef] [PubMed]
- Meisel, L.; Fonseca, B.; González, S.; Baeza-Yates, R.; Cambiazo, V.; Campos, R.; Gonźalez, M.; Orellana, A.; Retamales, J.; Silva, H. A rapid and efficient method for purifying high quality total RNA from peaches (Prunus persica) for functional genomics analyses. Biol. Res. 2005, 38, 83–88. [Google Scholar] [CrossRef] [PubMed]
- Trujillo, C.; Rodríguez-Arango, E.; Jaramillo, S.; Hoyos, R.; Orduz, S.; Arango, R. One-step transformation of two Andean potato cultivars (Solanum tuberosum L. subsp. andigena). Plant Cell Rep. 2001, 20, 637–641. [Google Scholar] [CrossRef]
Figure 1.
Scheme for plant regeneration of transgenic potato under non selective conditions.
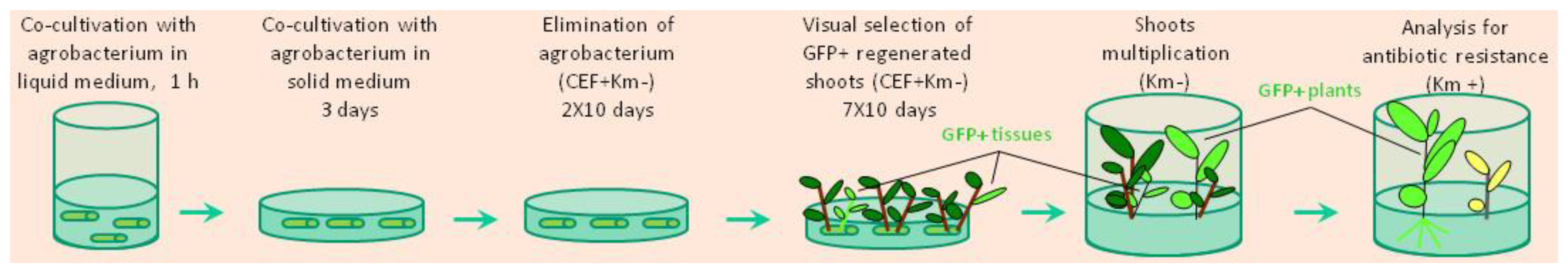
Figure 2.
Plantlets regeneration from in vitro cultured internodal segments of various potato cultivars.
Figure 2.
Plantlets regeneration from in vitro cultured internodal segments of various potato cultivars.

Figure 4.
Regeneration of transgenic potato plants under non selective conditions. (a) and (b), Formation of GFP expressing buds on the cut edge of explants; (c) Development of chimeric plantlets with the GFP positive sector on expanding leaf; (d) and (e), Regeneration of single transgenic plantlets of #12-22-129 and ‘Chicago’ with a visible green fluorescence throughout all tissues; (F) Two independent transgenic shoots generated by a intermodal explant of ‘Pirol’ among the non-transgenic plantlets.
Figure 4.
Regeneration of transgenic potato plants under non selective conditions. (a) and (b), Formation of GFP expressing buds on the cut edge of explants; (c) Development of chimeric plantlets with the GFP positive sector on expanding leaf; (d) and (e), Regeneration of single transgenic plantlets of #12-22-129 and ‘Chicago’ with a visible green fluorescence throughout all tissues; (F) Two independent transgenic shoots generated by a intermodal explant of ‘Pirol’ among the non-transgenic plantlets.
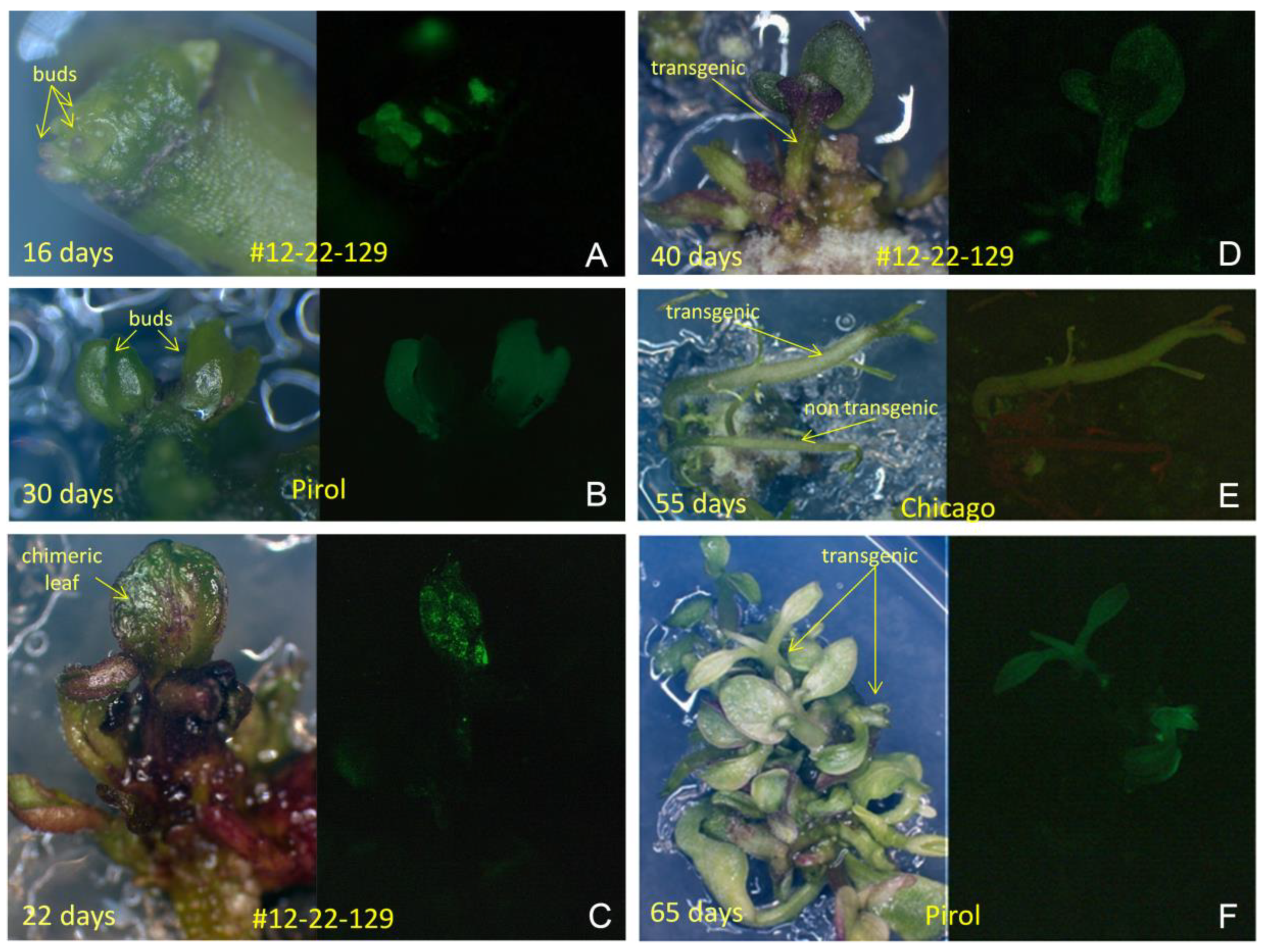
Figure 5.
Example of the Southern blot analysis for GFP insertions in selected potato lines of ‘Pirol’ and ‘Chicago’ transformed with the binary vector pBIN-mGFP5-ER.
Figure 5.
Example of the Southern blot analysis for GFP insertions in selected potato lines of ‘Pirol’ and ‘Chicago’ transformed with the binary vector pBIN-mGFP5-ER.
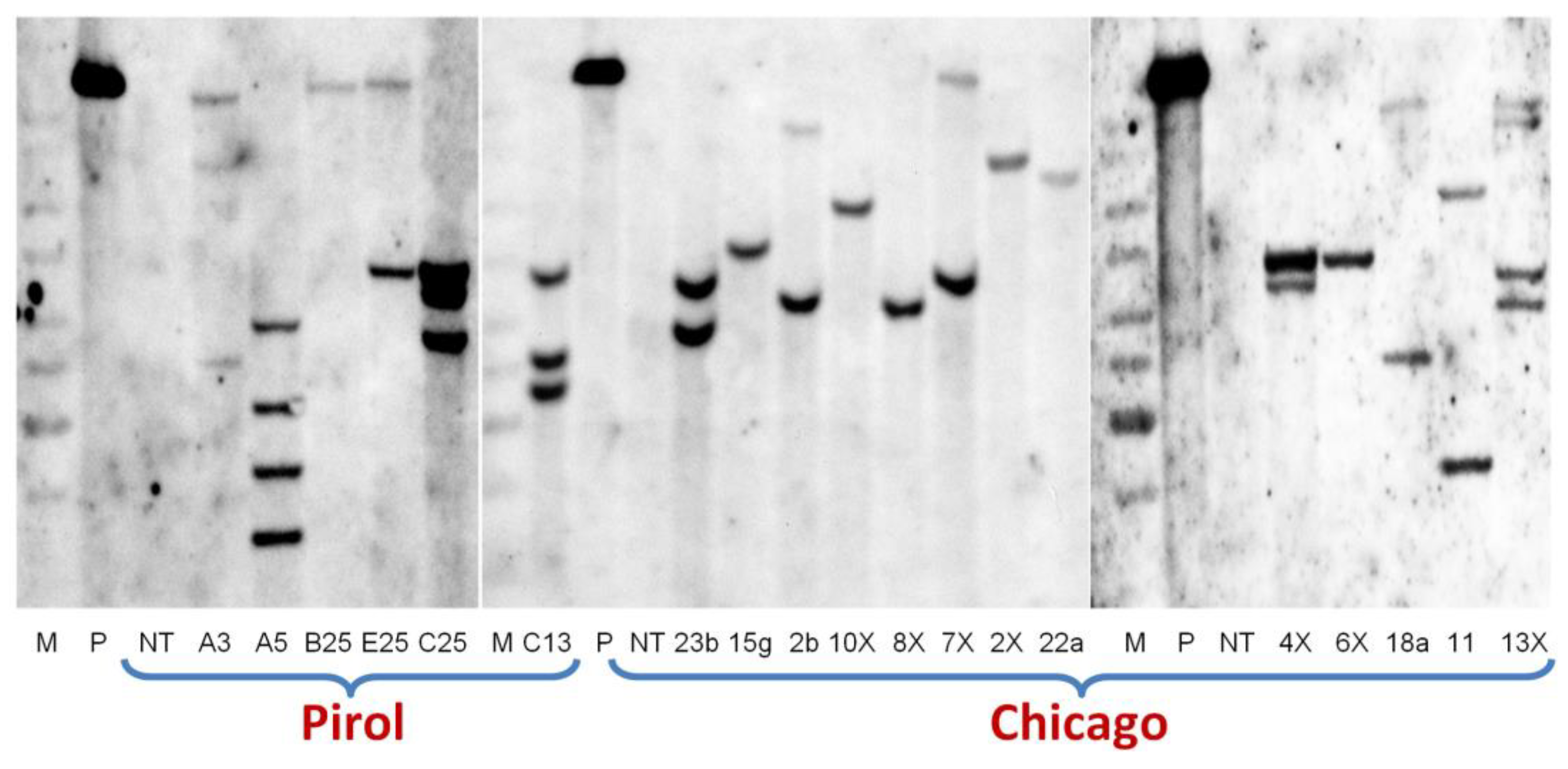
Figure 6.
Genotype-dependant timeline for formation of GFP expressed buds and plantlets of #12-22-129 (‘early’ type of regeneration) and ‘Pirol’ (‘late’ type of regeneration) under non selective conditions in comparison with the all regenerated shoots.
Figure 6.
Genotype-dependant timeline for formation of GFP expressed buds and plantlets of #12-22-129 (‘early’ type of regeneration) and ‘Pirol’ (‘late’ type of regeneration) under non selective conditions in comparison with the all regenerated shoots.
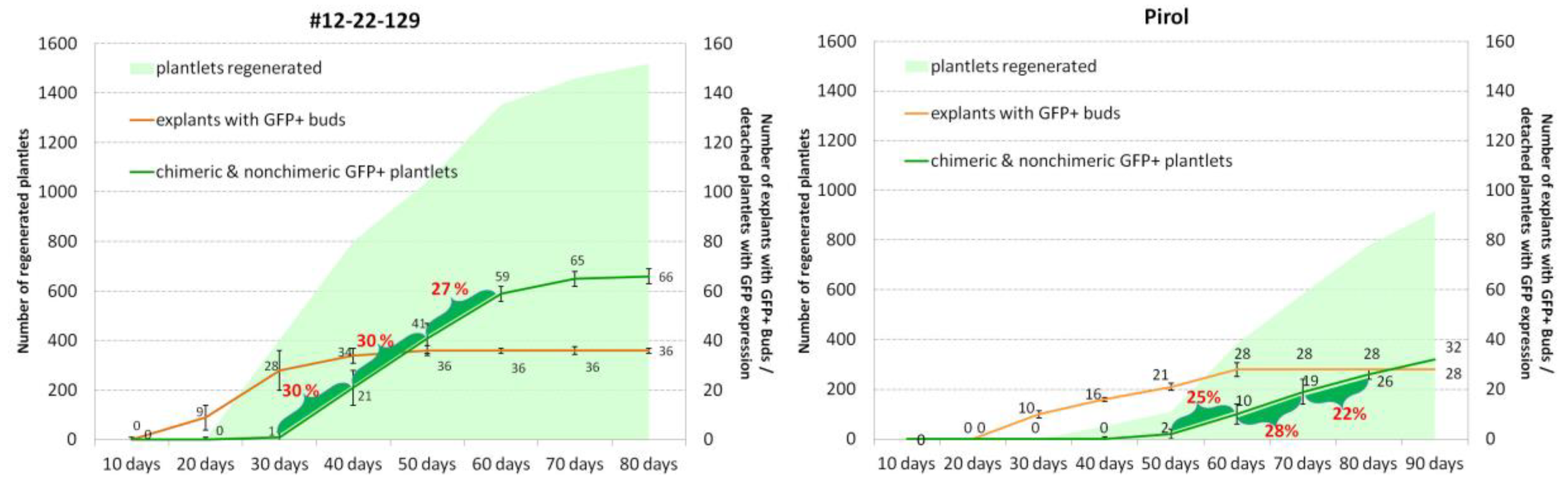
Figure 7.
Generation of marker-free transgenic potato plants of cv. ‘La Stada’ by a markerless vector pWS-E1-Lhca3 encoding the hairpin construct for the RNAi-mediated silencing of the SteIF4E1 and SteIF4E2 genes.
Figure 7.
Generation of marker-free transgenic potato plants of cv. ‘La Stada’ by a markerless vector pWS-E1-Lhca3 encoding the hairpin construct for the RNAi-mediated silencing of the SteIF4E1 and SteIF4E2 genes.
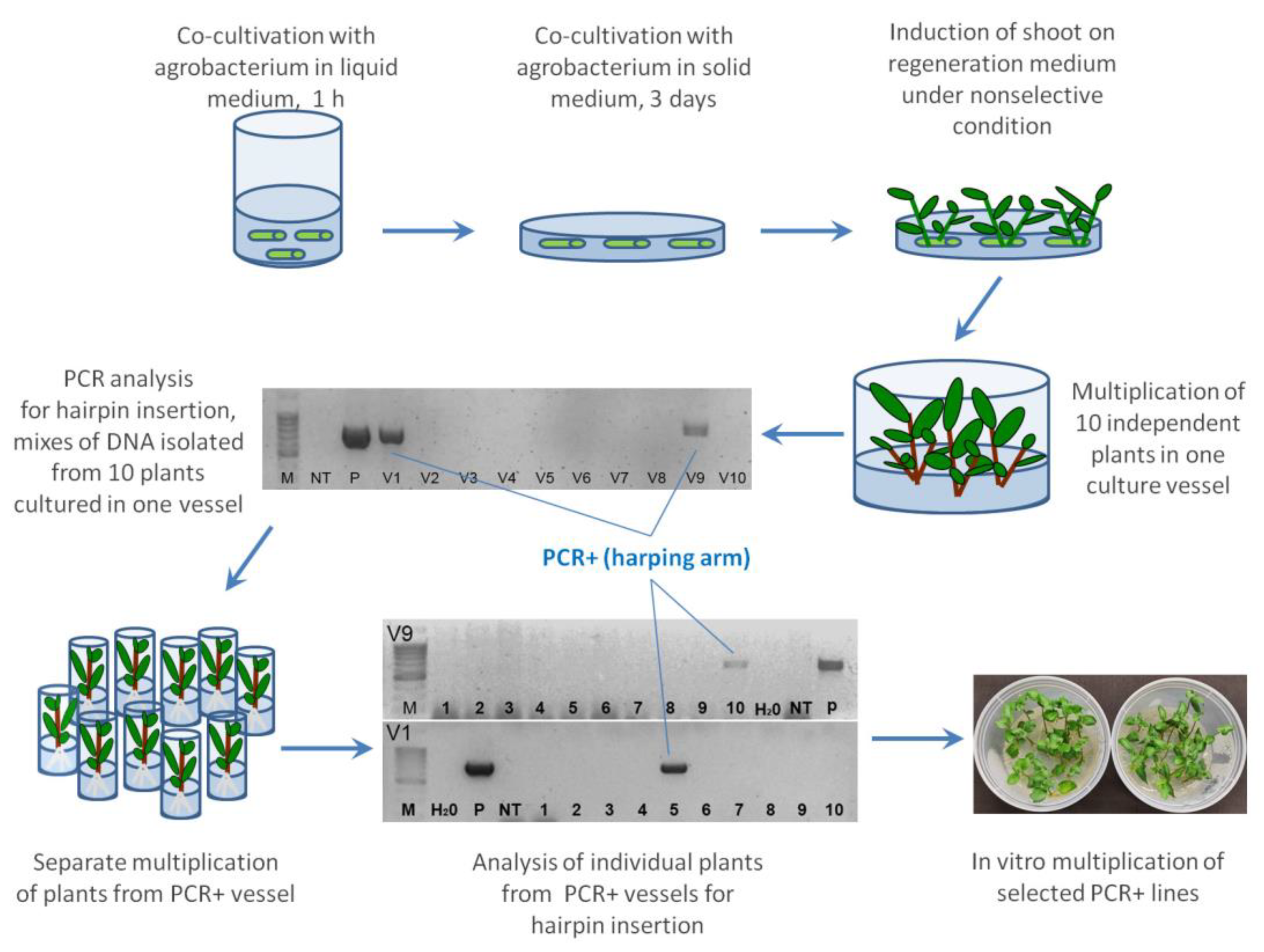
Figure 8.
Relative expression of SteIF4E1 and SteIF4E2 genes determined by RT-qPCR in transgenic plants of ‘La Strada’ carrying the hairpin construct from pWS-E1-Lhca3 vector. The presented values are means of three replicates and the bars are standard deviations. Means having the same letter in the column has no significant differences according to Duncan’s multiple range test (p < 0.001).
Figure 8.
Relative expression of SteIF4E1 and SteIF4E2 genes determined by RT-qPCR in transgenic plants of ‘La Strada’ carrying the hairpin construct from pWS-E1-Lhca3 vector. The presented values are means of three replicates and the bars are standard deviations. Means having the same letter in the column has no significant differences according to Duncan’s multiple range test (p < 0.001).
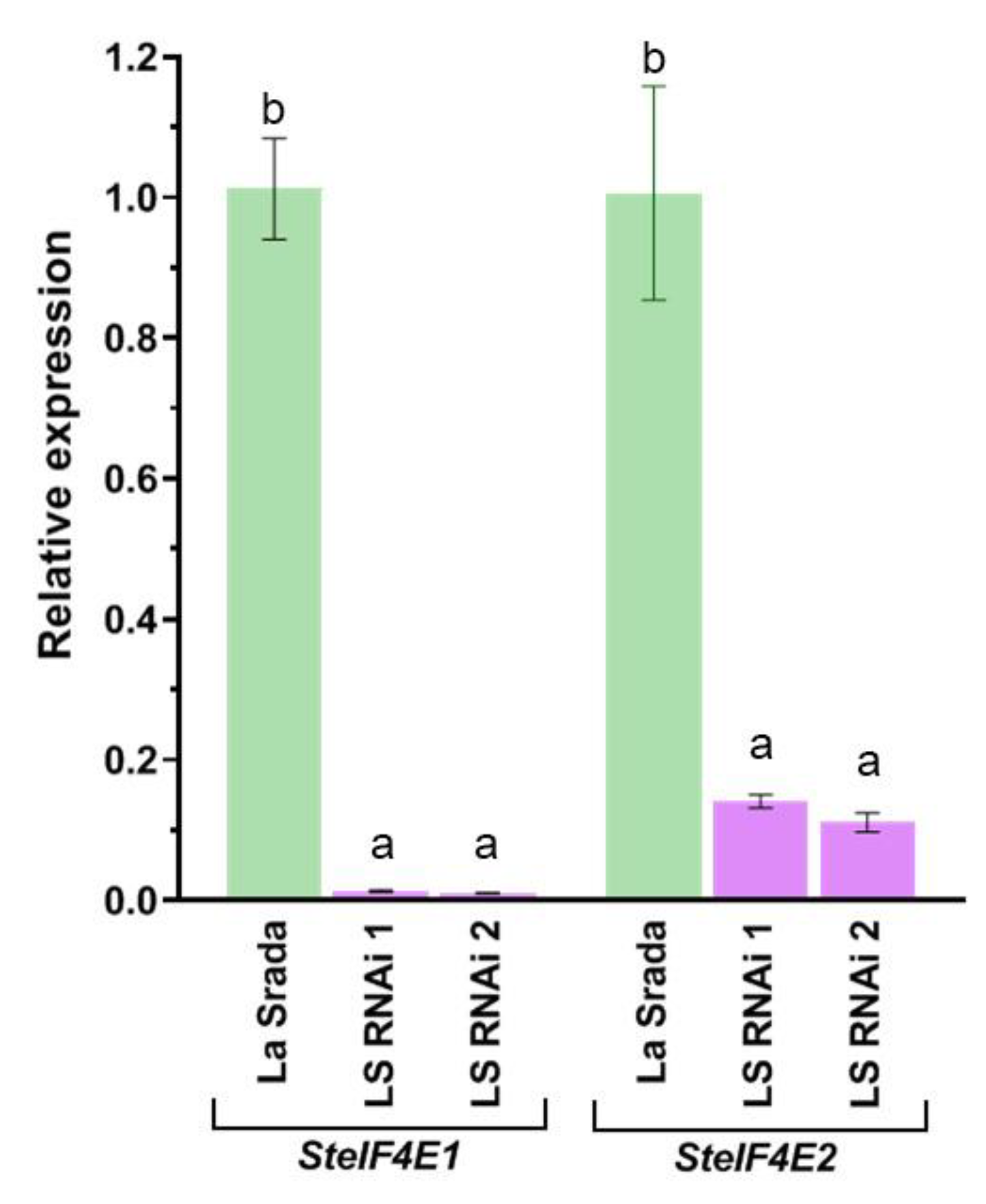

Table 1.
The efficiency of plant regeneration from internodal segments of potato cultivars.
| Cultivar | Combination of phytohormones* | Regeneration Type |
Efficiency of Plantlet Regeneration (%) |
Number of Plantlets per Explant |
|---|---|---|---|---|
| Chicago | 3 Zea + 2 IAA+ 1 GA | early/medium | 98.3 ± 3.3 | 15.2 ± 3.9 |
| Indigo | 3 Zea-R + 0.5 IAA | late | 75.4 ± 2.8 | 2.4 ± 0.3 |
| Lion Heart | 3 Zea + 2 IAA+ 1 GA | late | 93.1 ± 6.3 | 6.3 ± 0.4 |
| La Strada | 3 Zea-R + 2 IAA+ 1 GA | medium | 97.8 ± 3.8 | 12.3 ± 2.9 |
| Manhattan | 3 Zea + 0.5 IAA | medium | 76.2 ± 10.6 | 3.5 ± 1.9 |
| Pirol | 3 Zea + 0.5 IAA | late | 100 ± 0.0 | 8.2 ± 2.0 |
| #10-9-3 | 3 Zea + 2 IAA+ 1 GA | late | 100 ± 0.0 | 6.6 ± 1.8 |
| #10-10-10 | 3 Zea + 0.5 IAA | medium | 93.6 ± 6.3 | 4.8 ± 0.6 |
| #12-22-129 | 3 Zea + 0.5 IAA | early | 100 ± 0.0 | 15.0 ± 1.6 |
| #12-36-42 | 3 Zea + 0.5 IAA | medium/late | 95.0 ± 4.4 | 6.4 ± 2.1 |
* All regeneration media were consisted of MS mineral salts and vitamins, supplemented with 30 g L−1 sucrose, 100 mg L−1 myo-inositol, and solidified with 7 g L−1 agar. Various conbinations of phytogormones were used to induce adventitious regeneration, incuding: 3 mg L−1 of zeatin (Zea) or zeatin-riboside (Zea-R), 2 or 0.5 mg L−1 of indoleacetic acid (IAA), 1 mg L−1 of gibberellic acid (GA).
Table 2.
The efficiency of antibiotic-free genetic transformation of five potato cultivars.
| Cultivar | Regeneration Type | Number of Explants with GFP+ buds* |
Number of Shoots regenerated (90 days)* |
Number of GFP+ shoots detached from explants* |
Number of stable transgenic events (GFP+)* |
Percentage of GFP+ stable events per 100 regenerated shoots (%) |
Transformation efficiency with antibiotic- assisted selection**(%) |
|---|---|---|---|---|---|---|---|
| #12-22-129 | early | 36 | 1519 | 66 | 32 | 2.1 ± 0.4 | 100 |
| Chicago | early/intermediate | 16 | 1554 | 22 | 15 | 0.9 ± 0.1 | 98 |
| La Strada | intermediate | 30 | 1434 | 40 | 29 | 2.0 ± 0.5 | 47 |
| #12-36-42 | intermediate/late | 39 | 964 | 50 | 25 | 2.6 ± 0.4 | 68 |
| Pirol | late | 29 | 916 | 33 | 16 | 1.8 ± 0.7 | 44 |
* Average number per 100 inoculated explants, ** kanamycin-assisted transformation frequency defined as the percentage of initially inoculated explants produced GFP positive events.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



