
Submitted:
10 December 2023
Posted:
11 December 2023
You are already at the latest version
Abstract
Keywords: Trichoderma sp., biogenic selenium nanoparticles, capping biomolecules, bio-corona, agriculture, plant biostimulants, phytopathogens, physicochemical properties, bioactivity mechanism
Keywords:
Trichoderma sp.
; biogenic selenium nanoparticles
; capping biomolecules
; bio-corona
; agriculture
; plant biostimulants
; phytopathogens
; physicochemical properties
; bioactivity mechanism
1. Introduction
Due to its physical, chemical, and biological characteristics, selenium (Se) is regarded as an element of great potential in a variety of scientific fields, such as biomedical, food, agricultural, and environmental fields [1,2,3]. Selenium is known to have a narrow physiological window [4,5]. Therefore, in areas with selenium deficit in soil, biofortification is considered to produce better results than direct supplementation [3,6]. Beside the biofortification effect, selenium application on plants determines effects similar to plant biostimulants: enhanced nutrient uptake [7], increase resistance to abiotic stress [8,9] and increased crop quality traits [10,11,12].
Scientific research in nano-formulations of micronutrients like Se is gaining more and more ground on account of the undeniable advantages of nanotechnology. Nanoparticles (NPs) are believed to have superior reactive surfaces, due to their high area-to-volume ratio [13,14,15]. Nano-formulations can enhance absorption, sensitivity, stability and resistance [16,17]. Whence, NPs are highly biocompatible and bioavailable [18]. Besides the aforementioned valuable features, selenium nanoparticles (SeNPs) have lower toxicity compared to other forms of Se, due to the zero-valent state of oxidation of nano-selenium [19].
The most common means of obtaining SeNPs have been chemical methods. These approaches are based on reducing selenium using reducing agents. The reactions often take place in the presence of certain emulsifiers or surfactants, which create stable colloidal systems that will finally contain the SeNPs. Physical methods of SeNPs synthesis mainly involve the exposure of selenium precursors to different radiation sources. In this regard, photoablation or different non-ionizing radiations (UV radiation) or ionizing radiation (gamma radiations) are used to target selenium compounds, which will finally lead to SeNPs formation. Apart from being expensive and time consuming, the harsh conditions of the aforementioned techniques can create a toxic processing environment, non-sustainable waste and can interfere in the use of SeNPs in biological systems [20,21,22]. Biological synthesis methods, developed in a green manner, seem to be the light at the end of the tunnel in a resource-consuming scientific outline. These methods are considered safe, eco-friendly, cost-effective, non-toxic and waste minimizing [23,24,25].
Biogenic nanoparticles can be synthetized using microorganisms or plants. NP biosynthesis can occur either intracellularly or extracellularly. Certain biomolecules of the physiological apparatus of microorganisms can carry out reducing and stabilizing functions, vital for SeNPs production [26]. As they provide just the right conditions for this process, microorganisms got their reputation as nano-factories. Their great potential also lies in the possibility of modifying their cellular machinery to facilitate NP synthesis [27].
Fungi are thought to be among the most efficient microorganisms in the matter of biogenic NP synthesis [28]. Besides their capacity to biosynthesize high quantities of NP, the stabilizing enzymes and metabolites, fungi are manageable microorganisms with accelerated growth rate. An interesting and resourceful trait of fungi is their ability to tolerate metals, even in high concentrations [29]. Compared to bacteria, fungi are almost ideal biocatalysts for the biosynthesis of NP since they are known to produce higher levels of bioactive compounds, making them more suitable for large-scale production [30].
Nanoparticles synthesized by fungi have been successfully studied in a wide range of research areas like agroecosystems, plant science, eco-friendly formulations with protective potential for agricultural crops. Abd-Elsalam (2022) predicts that the introduction of large-scale myco-nanoparticles strategies in agriculture will arise in the upcoming years, due to the continuous reports of their newly discovered uses in agri-food fields over the last few years [31].
All things considered, fungal-mediated synthesis of SeNPs is a resourceful research direction. The main principle of this biosynthesis relies on using fungal biomass as a host for the reduction of Se precursors such as selenate (SeVIO4 2) and selenite (SeIVO3 2–) oxyanions to the less toxic Se0. SeNPs synthesis using fungal biomass has been successfully performed using various strains: Gliocladium roseum [32], Aspergillus terreus [33], Alternaria alternata [34], Aspergillus oryzae [35], Penicillium chrysogenum [36], and Trichoderma sp., the latter being detailed below.
The last years have witnessed an unprecedented interest in SeNPs and its applications, especially in the biogenic ones. Several reviews that approach this subject are available [6,24,37,38,39,40,41], including special reviews on the applications of SeNPs in agriculture [39,40,41]. Therefore, the purpose of this work is not an in-depth review of the general aspects of SeNPs, but rather aims to give a critical and focused analysis of the probably most relevant type of SeNPs for agriculture, the so-called trichogenic SeNPs., i.e., SeNPs produced by Trichoderma sp.
Trichoderma sp. is a specific species of interest for the production of SeNPs. Trichogenic SeNPs could find a wide variety of applications, especially in agriculture, as the benefits of Trichoderma sp. uses on plants are well known. Trichoderma colonizes wood, herbaceous plant materials, showing a high level of genetic diversity. The adaptability of Trichoderma sp. to various substrates and its tolerance to toxic compounds (e.g.: from fungicides, herbicides or pollutants) make these fungi excellent soil biocontrol agents [42]. Therefore, using this fungus for SeNP synthesis could potentiate its properties and those of the SeNPs themselves, leading to novel products of commercial and ecological interest [43].
Despite the significant importance of Trichoderma sp. and the SeNPs it can produce, a thorough review focusing on trichogenic SeNPs is still lacking. The current reviews either present various nanoparticles and nanomaterials produced by Trichoderma sp., with a relatively brief mention of trichogenic SeNPs, or present general fungal biosynthesis of SeNPs. Moreover, probably one of the most important aspects for agriculture applications, i.e., the nature of the capping molecules forming the so-called bio-corona of trichogenic SeNPs, has not been enough emphasized. Therefore, we considered necessary to bring to front and into perspective the main aspects of trichogenic biosynthesis of SeNPs.
2. Methods of trichogenic SeNPs biosynthesis
Trichoderma-derived SeNPs are currently in the developmental stages. Several Trichoderma species have been employed in nanotechnology for obtaining NPs of metallic origin: titanium, gold, zinc, silver etc., recently reviewed in [44]. Even more, some strains have substantial potential in the biosynthesis of nanoparticles at industrial level [30,45].
There is limited study on Trichoderma sp. mediated synthesis of SeNPs. The biosynthesis methods are influenced by the fungal strain used, selenium precursor concentration, culture medium where it grows and develops the metabolites important for nanoparticles formation. For production of mycelial biomass involved in biosynthesis, the most common growth media used in studies for Trichoderma strains were Czapek-Dox agar (CDA) [46], Potato dextrose broth (PDB) [23,47,48], AP1 agar supplemented with nutrients [49], and Martin modified broth (MMB) medium [29,50].
The first report describing the biosynthesis methodology based on Trichoderma sp. along with other filamentous fungi (Aspergillus funiculosus, Aspergillus niger, Coriolus versicolor, Rhizopus arrhizus etc.) and yeasts (Saccharomyces cerevisiae, Candida glabrata, etc.) was by Gharieb et al. (1994) [46]. The study was based on understanding the capacity of strains to reduce selenite to Se0. Based on this screening, the authors demonstrated that T. reesei can reduce selenite to elemental selenium. The first hint of the reduction is the red colour change of the growth medium (due to the presence of amorphous elemental selenium).
Later, more strains of Trichoderma were tested together with other methodologies of SeNPs biosynthesis, i.e., in vitro, by using cell lysate, culture filtrate and/or cell walls. Nandini et al. (2017) studied downy mildew control in pearl millet, synthetizing SeNPs by way of T. asperellum, T. harzianum, T. atroviride, T. virens, T. longibrachiatum and T. brevicompactum strains, using three forms of fungal culture (culture filtrate, cell lysate, and crude cell wall) [47]. A similar method was performed on T. atroviride [51]. In another study, eight Trichoderma strains were investigated in order to obtain SeNPs with bioactive metabolite support, with applications against phytopathogens and mycotoxins [48]. Trichoderma sp. WL-Go was subjected to several conditions to identify the optimal ones [50]. T. harzianum appears to be the most studied strain in SeNPs formation, identified as a strain with auspicious biosynthesis potential [48,49,52].
Considering the previous studies, the biogenic synthesis based on Trichoderma sp. has several benefits in terms of efficiency and the generation of diverse metabolites under optimal surroundings [28]. The inoculation conditions serve as key points right from the beginning of the experiment. As a first step Trichoderma sp. needs to be inoculated on a solid growth medium. Potato dextrose agar (PDA) is the classic culture medium in fungal isolation and culture, also valid for Trichoderma sp. [47,48]. The malt extract agar (MEA) is used as well [49]. The development of the mycelial mass in the liquid growth medium takes place in the dark, under static [47,51] or under stirring conditions [23,49]. An essential point is considered to be the growth phase of microorganisms [26].
Most of the studies investigating trichogenic SeNPs used culture filtrate (extracellular content), lysate (intracellular content), and/or cell walls from Trichoderma sp. to perform in vitro bio-assisted SeNPs synthesis. To collect the metabolites for the in vitro bio-assisted synthesis of trichogenic SeNPs, mycelial mat is subjected to several processes such as ground, lysis through sonication, centrifugation. Then, an aqueous solution of Se precursor is added. Sodium selenite (Na2SeO3), a well-known bioactive chemical, has commonly been used as a precursor in trichogenic SeNPs in vitro and in vivo biosynthesis but its concentration varies from study to study [23,46,47,48,49,51,52]. The reaction mixture is usually kept in the dark until the presence of red sediment, because light induces Se reduction as well and could induce formation of other Se forms [23]. Nevertheless, light is a parameter that deserves to be tested with respect to the biological effects of the trichogenic SeNPs. A previous study on SeNPs produced by a culture filtrate of Penicillium crustosum reported enhanced antimicrobial, anticancer, and catalytic activity in the presence of light [29].
The in vivo bio-synthesis, reported in less studies than in vitro one, can result in either intracellular, extracellular or both types of SeNPs, the extracellular ones either by exporting them from the cells or by direct extracellular SeNPs formation [46,49,50]. A brief presentation of the biosynthesis methods in relation to the obtained SeNPs is presented in Table 1.
The pH seems to play an important role, at least with respect to some aspects of trichogenic SeNPs. Diko et al. concluded that the pH is a critical parameter for trichogenic SeNPs synthesis [50]. It is assumed that the alkaline medium prevents the agglomeration of nanoparticles and promotes the stabilization of capping agents from fungi on the NP surfaces, such asproteins and other biomolecules, forming the so-called “bio-corona” [29]. The nature of capping biomolecules has a high impact on the stability of SeNPs with respect to aggregation. An alkaline pH induced an increase in the permeability of fungal cell membrane, which resulted in extracellular SeNPs [26]. The optimum pH for extracellular synthesis of SeNPs from Trichoderma sp. was found to be 8 when performed in vivo [50] and was reported in vitro at pH 8-12 in one study [23]. The in vitro and in vivo biosynthesis of fungal, including trichogenic SeNPs both seem to have advantages and disadvantages. The in vitro biosynthesis has the advantage of an easier manipulation of SeNPs, avoiding the neccessasity of mycelium lysis to recover the SeNPs, lysis which could even result in alterations of their properties. The in vivo biosynthesis could have the advantage of generating specific metabolites and other biomolecules triggered by the interaction with Se. These metabolites might have superior biological properties than the metabolites produced in the absence of Se, properties that would be transfered to SeNPs as well, in the case of these metabolites being part of the capping bio-corona. In-depth studies, comparing the two types of biosynthesis with the same Trichoderma strains, whcih are currently missing, would give more Information in this respect.
3. Mechanism of trichogenic SeNPs biosynthesis
The exact mechanism of SeNPs formation by Trichoderma sp. has not been investigated, but some clues can be obtained from the studies of other fungal species. Most of the studies reported that the synthesis of NPs in fungi might be a defense mechanism for reducing the toxicity of different elements that they encounter. It is already recognized that fungi have great tolerance to metal and non-metal ions [19,37,54]. One of the fungus coping mechanism with this kind of abiotic stress refers to their ability of reducing the metal/non-metal ions to lower the states of oxidation and, implicitly, synthetizing nanoparticles which are usually less toxic (e.g.: reducing Se+4 and Se+6 from selenite (SeO3-2) and, respectively, selenate (SeO4−2) to zero valent selenium – creating SeNPs). This process could be viewed as detoxifying mechanism [54,55,56].
Although accumulation of nanoparticles can cause physiological changes in microorganisms, fungi can tolerate impressive amounts of NP accumulation. Therefore, fungi continue to develop even after the biosynthesis of nanoparticles [37,57]. Synthesis of NP in fungal biomass can occur inside the cytoplasm (absorption), within the periplasm or outside the cell membrane (adsorption). Intracellular synthesis of NP overcomes the disadvantage of obtaining aggregated, clustered, and large-sized nanoparticles. In comparison to intracellular processes, extracellular synthesis offers the advantage of obtaining large quantities of NPs in a relatively pure state, free from other cellular fungal biomolecules or microbial cells, thus making the downstream process easier [58]. Additionally, fungi release extracellular reductive proteins that can be employed in later stages of the process [30,51].
Figure 1.
Possible mechanism of fungal-mediated SeNPs synthesis. NADPH dependent oxidoreductases mediate the reduction of selenium ions from precursors to SeNPs along with quinone derivates which can be electron shuttles in the oxidoreduction process. Capping agents (e.g.: proteins from fungal biomass) can surround the SeNPs, acting as NP stabilizers. This process can occur either intracellularly (after the absorption of the Se ions into the cell) or extracellularly. Some information about membrane transport processes and other metabolites involved in the fungal synthesis of NP are not fully understood yet. The Figure is adapted and based on the Information from [54,59].
Figure 1.
Possible mechanism of fungal-mediated SeNPs synthesis. NADPH dependent oxidoreductases mediate the reduction of selenium ions from precursors to SeNPs along with quinone derivates which can be electron shuttles in the oxidoreduction process. Capping agents (e.g.: proteins from fungal biomass) can surround the SeNPs, acting as NP stabilizers. This process can occur either intracellularly (after the absorption of the Se ions into the cell) or extracellularly. Some information about membrane transport processes and other metabolites involved in the fungal synthesis of NP are not fully understood yet. The Figure is adapted and based on the Information from [54,59].
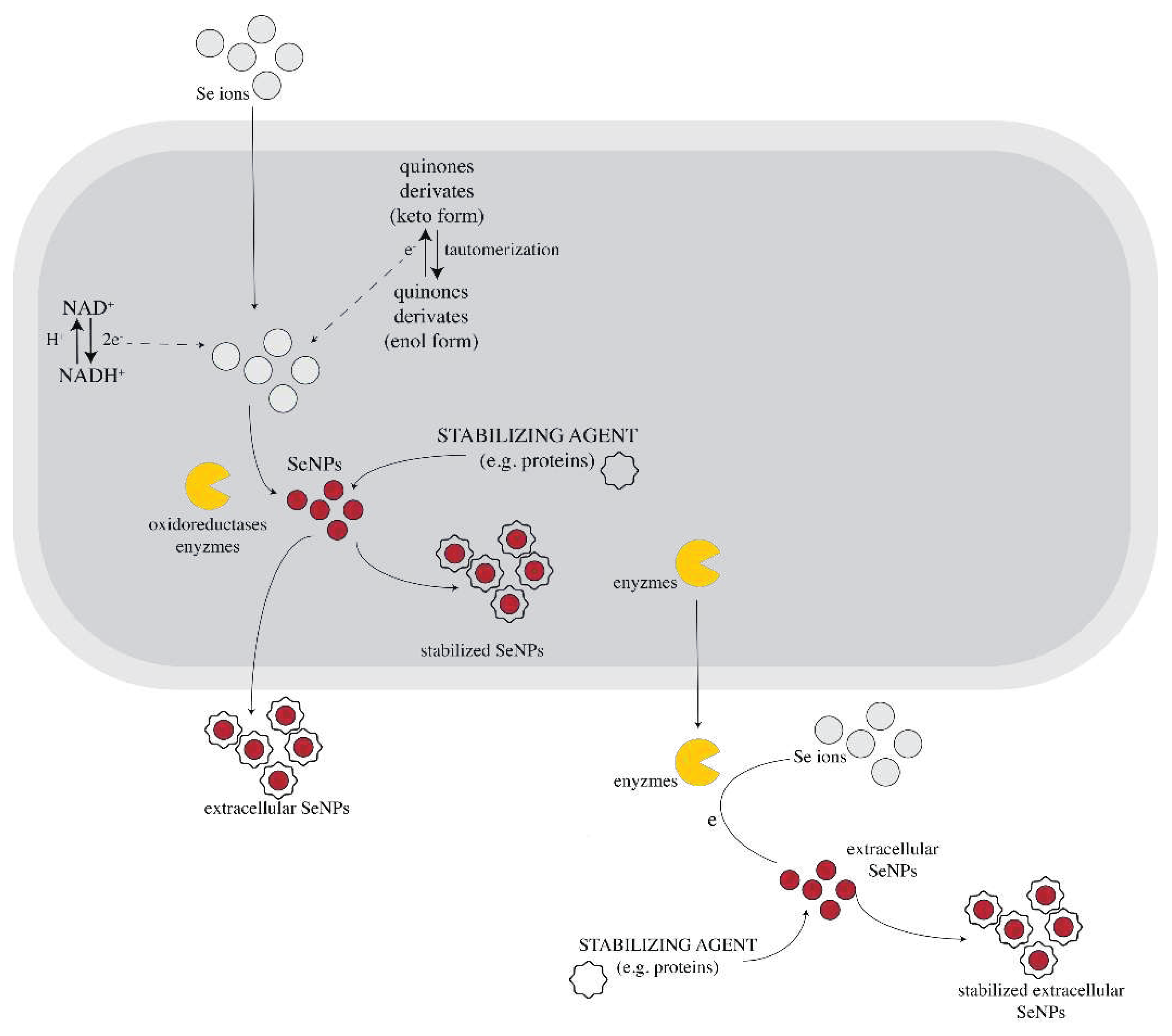
Although exceedingly researched, these superpowers of fungi have not been fully explained yet, the complete mechanism of fungi-mediated NPs synthesis remaining unknown. However, studies confirm that the NADPH – dependent reductases (nitrate (NO3–) and nitrite (NO2−) reductases) from cell wall or plasmatic membrane of the cell play a major role in the fungal NP biosynthesis, including trichogenic synthesis of selenium nanoparticles. These reductases are thought to be catalysts for the reaction between the 2 e- donor – NADPH and inorganic ions [37,54,57,60,61,62]. However, G. Li et al. (2012) showed that NADPH alone is not enough to form these kinds of nanoparticles. Their experiments demonstrate that fungal NP biosynthesis is not possible using just NADPH and NP precursor alone. Moreover, when fungal biomass was added to NADPH and NP precursor, NPs were formed. Therefore, other molecules besides NADPH and oxidoreductases are necessary for nanoparticle synthesis by fungi [62]. For example, in selenite reduction, biomolecules like phenazine-1carboxylic acid and glutathione might be involved in selenite reduction [28]. Some research revealed that there are certain fungal, metabolites that contribute to biogenic NP formation. Compounds like penitric acid, chrysogine, chrysogenin, fungisporin, roquefortines or non-enzyme proteins like phytochelatins, methalothenin are thought to play a role in the process. Besides their reduction properties, most of the molecules involved in the fungal-mediated biosynthesis of NPs act also as capping agents that stabilize the NPs [37,49,54,57,61]. Other hypotheses propose that some of these metabolites might be the quinine derivates of anthraquinones and naphthoquinones [49], but this hypothesis requires experimental confirmation. These molecules act as electron shuttles. Quinone shuttles were shown to be necessary for NP production in some cases, as they might act as redox centers and/or electron carriers in the oxidoreduction reactions. There are studies that confirm that the quinones of T. harzianum have great reducing properties [63]. Liang et al. reported for the first time in 2019 the formation of not only elemental Se, but also Se oxide by interaction of sodium selenite and selenate with a T. harzianum strain, which indicates a more complex system than generally assumed [49]. The mechanism generated in vivo could be completely differently than the one in vitro using lysate or filtrate, as most probably Trichoderma produces additional and specific metabolites when in contact with toxic concentrations of Se salts.
Electrostatic interactions might also participate in the fungal NP synthesis. The negatively charged outer membrane surface interacts electrostatically with the inorganic ions, mediating the transportation of ions throughout the cell membrane and promoting complexation of selenium with ligands [54,59]. Clearly, there are more pieces that need to be put together in this metabolic puzzle of fungal mediated NPs synthesis and surely, current research on the topic will continue to provide answers to questions regarding this process.
4. Characterization and manipulation issues of SeNPs
A precise description of SeNP characteristics could be defining for their properties and the correlations with the applications. The shape, size, stability and chemical structure of bio-corona are some of the parameters that were frequently reported in studies of SeNPs. Characterization of these NP is often analyzed in relation with their stabilizing biomolecules, mainly proteins [29]. The main techniques used for NP characterization are Inductively coupled plasma - mass spectrometry (ICP-MS), Inductively coupled plasma - optical emission spectrometry (ICP-OES) and colorimetric methods for quantification, single particle mode ICP-MS (SP-ICP-MS), UV-Vis spectroscopy, Transmission electron microscopy (TEM), Scanning electron microscopy (SEM), X-ray diffraction (XRD), X-ray photoelectron spectroscopy (XPS), Dynamic light scattering (DLS) and zeta potential, Differential centrifugal sedimentation (DCS), Fourier-transform infrared (FTIR) and Raman spectroscopy, SDS-Page electrophoresis for identification of proteins capping the NPs, Mass spectrometry coupled with chromatography for identification of biomolecules from bio-corona,. Another useful technique, which can separate nanoparticles based on its size is Asymmetric flow-field flow fractionation (AF4) coupled with various detectors such as ICP-MS, Multi angle light scattering (MALS), or refractive index detector [64,65], but this technique is less reported in the literature, at least concerning SeNPs. ICP-MS and SP-ICP-MS, along with other techniques are powerful instruments for Se speciation of both anorganic and organic forms of Se [6]. Other methods can be found in recent reviews [64].
One of the most suggestive properties of SeNPs is their color. The post-incubation changes of color in the culture medium to orange or red can be used to confirm the SeNPs formation. The intensity of the color gradually increases with time, as the selenium precursor is being reduced. The color change upon formation of SeNPs is a helpful indicative, as an intense red color can correspond to a high potential of the fungal strain to produce this kind of NPs [28]. The intensity of the color can be documented by UV–Vis spectrophotometric analysis. The UV-Vis spectra of trichogenic SeNPs established that the wavelength at which these kinds of NP absorb light vary greatly, depending on the Trichoderma strain. Previous analysis indicated peaks at 550 nm [47], 259 nm [23], 260 nm [51] or 270 nm [52]. The wavelength showing the maximum absorption can be used for a rough estimation of the size of nanoparticles [66]. Quantification of SeNPs can be obtained by an assay which uses Na2S for dissolution of SeNPs [67].
Transmission electron microscopy (TEM) coupled with energy-dispersive X-ray spectroscopy (EDS) is one of the most popular methods for observing the morphology and composition of biogenic SeNPs, along with Scanning electron microscopy (SEM). These microscopy techniques offer information about the shape, size and intracellular or extracellular distribution of SeNPs. Notwithstanding, some studies point out that an optimum imaging can be achieved especially when the samples that are being subjected to this assay are highly diluted [30]. The surface morphology and size of SeNPs from Trichoderma sp. were observed as hexagonal, spherical or pseudo-spherical and of irregular shapes [23,47,50,51]. The size range of trichogenic nanoparticles observed through TEM is rather broad, depending on the fungal strain. Overall, studies reported measurements from 26.45 nm (T. harzianum) to 312.5 nm (T. virens) [47,48,50,51,52]. Besides TEM analysis, an EDS spectrum can offer information about the degree of purity of the biosynthetized SeNPs [68].
The scattering phenomenon of X-rays by sample electrons at specific diffraction angles is a frequently employed method to determine the crystallographic particularities of biogenic nanoparticles. Depending on the X-ray technique, different morphological particularities can be deduced, like crystal lattice, crystallinity, nanoparticles size distribution and shape. Considering the Bragg’s law, the wide-angle X-ray scattering (WAXS) offers information about the scattering angles induced by sub-nanometric structures (0.1-1 nm), the small-angle X-ray scattering (SAXS) covers the particle sizes from 1-100 nm, whereas the ultra-small-angle X-ray scattering (USAXS) extends the analysis range up to 1000 nm [69]. A complementary technique that is usually discussed together with SAXS and USAXS is the small-angle neutron scattering (SANS), respectively ultra-small-angle neutron scattering (USANS), in which the incoming neutrons interact with the sample nuclei and the neutron scattering is a quantum-mechanical effect [70]. WAXS technique (usually identified as XRD) is usually employed to determine the diffraction patterns of the (bio)synthesized nanoparticles, often in comparison with the crystalline date of the initial reactants to evidence the bio(reduction).
WAXS technique was used to investigate the reduction of selenium with SeNPs formation and the crystallographic structure of SeNPs. A detailed structural analysis can be provided by XRD assay for biogenic SeNPs as well. Several XRD assays confirmed that biogenic SeNPs in general and trichogenic SeNPs in particular appear as amorphous structures [23,47,48,49,68], but there were also reports of crystalline nature [50,51]. The crystalline nature of SeNPs is probably most influenced by the characteristics of the bio-corona, especially its size, but the studies performed previously on trichogenic SeNPs did not investigate this aspect. The XRD studies of trichogenic SeNPs have been performed only in the wide-angle X-ray scattering (WAXS) mode, which gives Information on the crystalline nature of a material.
SAXS technique uses the diffraction patterns generated by the particle electron density at small diffraction angles, usually at 2θ < 2°, to gather information about the nanoparticle size distribution, nanoparticle shape or nanopores [71]. SAXS has a number of advantages over other analytic techniques used for nanoparticles study, like TEM and DLS, the first one being that it offers an overall view on the bulk size distribution of particle diameters between 1-100 nm, whereas the small grid size of TEM and the reduced concentration of nanoparticles necessary for a good dispersion, together with the natural propensity towards acquiring esthetic images make TEM less relevant for a global picture of the nanoparticle size distribution. SAXS is a non-destructive method that allows the unaltered recovery of the sample after analysis. Compared to DLS, SAXS has a clear advantage in analyzing opaque, colored or semi-translucent colloidal suspensions, and the data reliability for nanoparticles between 1-30 nm is superior for SAXS compared to DLS. Additionally, SAXS allows the morphological determination of size, shape, size distribution, organization and surface structure of the sample electron density differences in their native (bio)synthesized state, liquid, sol-gel or colloidal, and even in situ synthesis progression [71]. Actually, the first SAXS study was performed on "colloidal gold" in 1951, before the term "nanoparticle" being defined [71,72].
The literature of SeNPs characterization using SAXS is scarce, suggesting an available innovative niche at the crossroad of these two directions. In one of the few available studies, spherical red SeNPs with a mean size of 45 nm were obtained by the reduction of Na2SeO3 with L-cysteine and further stabilized with 29 kDa polyvinylpyrrolidone [73]. SAXS analysis performed between 2θ angles 0-1° evidenced a high background noise at 2θ > 0.2°, and the experimental curve was fitted with a Gamma distribution model for particles with 45 nm mean diameter [73]. These SeNPs were used in combination with a boric acid-based fungicide to protect the wood products against the brown-rot Serpula lacrymans. SAXS was used to characterize SeNPs obtained by the reduction of Na2SeO3 with ascorbic acid and stabilized with 200 kDa chitosan as 3% suspension in 2% succinic acid [74]. SAXS performed between the scattering wavevector (q) values from 0-3 nm-1 evidenced a bimodal particle size distribution centered around 2.5 nm and 37 nm in volumetric ratio of 1:2 [74]. These SeNPs were intended for the development of adaptogenic drugs due to their biocompatibility and antioxidant properties. There are no available data of SAXS analysis performed on trichogenic SeNPs, to the best of our knowledge.
XPS assays of trichogenic SeNPs confirmed the presence of elemental Se, by a peak at the binding energy of elemental Se, 55.6 eV [47]. This data confirms the reduction of Se from its precursors and implicitly, the formation of SeNPs [26]. XPS is a powerful technique that can give additional Information on the bio-corona composition and the types and energy of Se interactions, e.g. via 3d orbital electrons, with the molecules in the bio-corona [75,76], but no in-depth analysis is available in the case of trichogenic SeNPs.
Dynamic light scattering (DLS) and SP-ICP-MS tehniques can be used to estimate the size of hydrated SeNPs in solution, including the bio-corona [23]. Whereas TEM gives Information mainly on the size of the selenium core, DLS gives Information on the hydrodynamic radius of the entire structure. SP-ICP-MS can also determine the size of NPs [49]. The zeta potential indicates the net charge of a particle and is a good indicator of NP stability in terms of colloidal dispersion and tendency for aggregation. The closer the value is to zero, the higher is usually the chance that the particles will aggregate due to attractive van der Waals interactions [77]. Values higher than ± 30 mV are considered to be highly stable. But because the zeta potential does not offer Information on the strength of these van der Waals interactions, there are cases of unstable NP suspensions despite high zeta potential and vice versa [78]. The zeta potential is determined by the characteristics of the biomolecules capping the Se core of the NP and forming the bio-corona. The determinations of zeta potential of biogenic SeNPs indicated that these kinds of structures are in general rather stable. To the best of our knowledge, there are only few studies that determined the zeta potential of trichogenic SeNPs [47,50,51,52]. An intriguing aspect is that the zeta potential seems to have significant variations across Trichoderma species and also on the fraction of the culture used, in the case of in vitro bio-assisted synthesis, varying from + 11 mV to as negative as -200 mV [47]. This observation deserves further more in-depth studies, including correlations between the capping biomolecules and zeta potential. Most of the zeta potentials indicating stable colloidal suspensions of SeNPs had negative values, whereas the positive values were in the range indicating unstable systems. Comparison of zeta values of washed and unwashed SeNPs showed that washing and resuspending SeNPs in distilled water can enhance its stability [50,51,78].
Another technique that can give valuable Information on the compounds forming the bio-corona is FTIR. This technique has been applied in almost all studies involving trichogenic SeNPs. Most of the FTIR studies indicated the existence of protein bands, e.g., amide bands I, II, and III in the fingerprint region 1600-1200 cm-1 [23,47,48,50,51,52], although in some cases not properly discussed. A comparison between the FTIR spectra of trichogenic SeNPs obtained with the sterile filtrate and control SeNPs with PDB medium showed spectral differences that indicated a protein-dominant and a saccharide-dominant bio-corona in the trichogenic SeNPs and control SeNPs, respectively [48]. The FTIR spectra of T. harzianum extract and SeNPs indicated as well that proteins from the trichogenic biomass are probably capping the SeNPs [52].
As FTIR data indicate that proteins contribute significantly to the capping and stabilization of trichogenic SeNPs, protein SDS-Page electrophoresis can be a valuable tool that should be used to characterize these proteins. The protein electrophoresis can provide hints on the abundance of capping proteins, its molecular weight, and, in combination with LC-MS/MS, the proteins can be identified. There are almost no studies including electrophoresis analysis of proteins involved in trichogenic SeNP formation. We found one study alone, in which the authors found two protein bands at 15 and 19 kDa proposed to be part of the capping bio-corona [50], but no identification of the bands was performed.
Using liquid chromatography and mass spectrometry (TripleTOF LC-MS), one research group identified 35 various metabolites in the aqueous solution of SeNPs produced by the sterile filtrate of a T. harzianum strain. Among them there were organic acids, amino acids, sugars, and some intermediates from the carbohydrate metabolism, 27 compounds having potent antifungal activity against phytopathogens [48]. This result indicates that the bio-corona can be in fact highly heterogeneous and it partially explains the significant differences in zeta potential of SeNPs among Trichoderma species. It additionally highlights the usefulness of combining various complementary techniques to better understand the properties and role of bio-corona of trichogenic SeNPs, approach which is currently missing.
Table 2 summarizes the Information available from the literature with respect to the composition and the induced physicochemical characteristics of the bio-corona capping trichogenic SeNPs.
Two bottle neck issues in the manipulation of fungal SeNPs is its separation from the fungal biomass in the case of in vivo biosynthesis and especially for intracellularly-formed SeNPs, and respectively, obtaining sterile SeNPs to be biologically tested at laboratory level. The first issue needs disruption of the fungal mycelium which could influence the characteristics of SeNPs. Moreover, the intracellular SeNPs could have different properties compared with the ones released extracellularly in the culture medium during incubation. These aspects are currently still unexplored and should be investigated in the future.
5. Applications of trichogenic SeNPs in plant protection and as plant biostimulants
Improvements in crop yield, management of nutrient levels in soil and among plants, insect control, managing environmental factors have become a priority for the agricultural sector [58]. Nanoparticles could be one of the solutions to satisfy these requirements and it is considered a research direction with great innovative potential. Trichoderma sp. can be used for the biogenic synthesis of NPs with powerful properties needed for the agriculture sector. The use of these species might exhibit its potential to act in synergism with the NP that it produces, opening new leads in creating biotechnologies that might enhance crop quality and resistance against phytopathogens [28,79].
One of the most prevalent bioactivities of trichogenic SeNPs is their antifungal activity [28,38,79,80]. This nano-technology has been proved to suppress sporulation of Sclerospora graminicola, the pathogen that causes downy mildew in pearl millet [47]. Biogenic SeNPs synthetized using T. atroviridae strains have been proved to have great zoosporicidal and antifungal activities for Phytophthora infestans in tomato, which is one of the most devastating pathogens in tomato [51]. The antifungal mechanism could be attributed to the united and synergistic effect of SeNPs and diverse fungal metabolites, including multiple organic acids and its derivates [48]. In addition, SeNPs synthesized by Trichoderma sp. have larvicidal and antifeedant activity, a main component of crop protection [23]. Recently, Helmy et al. showed higher antifungal activity of trichogenic SeNPs against Fusarium oxysporum and higher reduction of Fusarium wilt infection of tomato plant than selenite [81].
Constant mycotoxins contamination is a serious challenge of crop production, food safety animal and human health. Trichoderma sp. derived SeNPs might address these needs as functional biocontrol of mycotoxins in agricultural and food safety procedures [48]. These NPs could represent affordable, environmentally beneficial and non-toxic solutions, outstanding the conventional chemical biocontrol agents and fertilizers [51]. A test conducted by our group on Vigna radiata seeds which germinated in trichogenic SeNPs aqueous solutions validates the non-toxic potential of nanoparticles [82].
The biostimulant effects of biogenic SeNPs have been revealed by previous biotests. Zogra et al. (2021) highlighted that in addition to beneficial microorganisms, the use of biogenic SeNPs is known as an environmentally friendly approach, to enhance crop production by alleviating biotic and abiotic stresses (alleviating drought, salinity, heavy metal, heat stresses, and bacterial and fungal diseases in plants) [83]. Joshi et al. demonstrated enhanced resistance of tomato to late blight disease, by priming mechanisms [53]. The application of biogenic SeNPs in plants can improve crop value. Moreover, treatment of crops with biogenic SeNPs could lead to the production of edible foods fortified with Se, which might be of great importance in managing deficiencies of this micronutrient [38]. Table 3 summarizes the previously reported biological effects of trichogenic SeNPs with impact in agriculture and food industry.
For a proper optimization of the trichogenic SeNPs bioactivity and use in agriculture the mechanism behind the biological effects needs to be understood in-depth, such as the priming mechanisms mentioned above, as well as the properties that correlate with its bioactivity. The first step is to establish these correlations, in this sense only a handful of studies providing some Information with respect to trichogenic SeNPs. Nandini et al. found an inversely proportional correlation, but with a modest correlation coefficient, between the growth and proliferation inhibition of S. graminicola and the size of the SeNPs produced by various fractions of Trichoderma spp. Cultures [47]. The reason for this type of correlation is not straightforward, as the sizes of SeNPs were in general larger than 50 nm, reaching microparticle regime (> 100 nm). Translocation of SeNPs of this size into the cells would be rather difficult, although the smallest nanoparticles present are probably translocated. Other explanations might contribute more, such as a different rate of selenium disproportionation or differences in the biomolecules forming the bio-corona. It would have been interesting to perform a correlation analysis with the zeta potential as well, which varied largely in their study, depending on the Trichoderma strain and culture fraction used, we mentioned above.
In another study, the antifeedant activity of trichogenic SeNPs against Spodoptera litura was directly proportional to the nanoparticle concentration [23]. Other correlations between trichogenic SeNPs activity and their properties are not available according to our survey. Questions remain open on if and how the capping bio-corona variation, in relation also with the SeNP crystalliny, affects the bioactivity, considering the differences between Trichoderma species observed. There is no systematic study available to assign particular capping composition to a specific effect. Moreover, few investigations studied to what extend the effects observed are due to the capping biomolecules and to what extend to released soluble Se species or small trichogenic SeNPs that could translocate into the cells, and if there is a synergism between these contributions. The work of Hu et al. showed that the trichogenic SeNPs significantly enhanced the antifungal activity against phytopathogens compared with standard SeNPs [48]. The enhanced activity was despite a lower absorption of selenium from trichogenic SeNPs compared with standard SeNPs, as determined by ICP-MS, which indicates that it was mainly due to metabolites such as organic acids. An analysis of speciation would have helped to estimate absorbed elemental Se from other Se species. Moreover, we believe that the constituents of the NP could influence each other. For example, the capping bio-corona might influence the Se disproportionation reactions and the disproportionation could influence the bio-corona stability. For example, the protein corona was proposed to reduce the oxidation of trichogenic SeNPs [81]. Therefore, the stability of SeNPs should be considered not only from the colloidal point of view, but also the oxidation state of Se and bio-corona integrity. Bacterial proteins were shown to influence the size of SeNPs [84] and this is probably available for fungal proteins as well.
In addition, for agricultural applications in open-fields, one has to consider the effects of soil microbiota and composition on SeNPs. Microorganisms, including fungi, can induce not only reduction, but also oxidation and methylation of selenium species, which could influence the Se disproportionation and the relative abundance of various states of Se [49,85].
6. Conclusions
Selenium nanoparticles (SeNPs) have been intensely studied over the last decade due to their extraordinary properties and their various biological activities. The present review confirms that the biosynthesis of selenium nanoparticles involving Trichoderma sp. reveal a promising potential, in an eco-friendly manner. The SeNPs obtained through trichogenic biosynthesis are non-toxic and have enhanced biological activity compared with chemically synthesized SeNPs. The current study described as well several documented hypotheses on the fungal-mediated NP biosynthesis, that could explain the synthesis mechanism of trichogenic SeNPs. Biogenic nanoparticles, including the trichogenic SeNPs, can be characterized by a wide range of analysis methods, depending on the aim of their final use. Many pieces are still missing in the puzzle and more complementary physicochemical and biological investigations should be performed, as well as correlation analysis for optimizing the biological activity. Applications of trichogenic selenium nanoparticles target mainly the agri-food sector, aiming to improve crop quality.
Author Contributions
Conceptualization, F.O. and D.C.-A.; writing—original draft preparation, L.T.C, V.B., and Ș.-O.D.; writing—review and editing, I.C.F., D.C.-A., and F.O..; visualization, D.C.-A. and F.O.; supervision, I.C.F. and F.O.; project administration, D.C.-A.; funding acquisition, D.C.-A. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by Ministry of Research, Innovation and Digitization, CNCS/CCCDI—UEFISCDI, project number 107PCE/2021, within PNCDI III.
Data Availability Statement
Not applicable
Conflicts of Interest
The authors declare no conflict of interest
References
- Nayak, V.; Singh, K.R.; Singh, A.K.; Singh, R.P. Potentialities of selenium nanoparticles in biomedical science. New Journal of Chemistry 2021, 45, 2849–2878. [Google Scholar] [CrossRef]
- Xiao, D.; Li, T.; Huang, X.; Zhu, K.; Li, Z.; Dong, Y.; Wang, L.; Huang, J. Advances in the Study of Selenium-Enriched Probiotics: From the Inorganic Se into Se Nanoparticles. Molecular Nutrition & Food Research 2023, 2300432.
- Schiavon, M.; Nardi, S.; Dalla Vecchia, F.; Ertani, A. Selenium biofortification in the 21 st century: status and challenges for healthy human nutrition. Plant and Soil 2020, 453, 245–270. [Google Scholar] [CrossRef]
- Banerjee, M.; Chakravarty, D.; Kalwani, P.; Ballal, A. Voyage of selenium from environment to life: Beneficial or toxic? Journal of Biochemical and Molecular Toxicology 2022, 36, e23195. [Google Scholar] [CrossRef]
- Reich, H.J.; Hondal, R.J. Why nature chose selenium. ACS chemical biology 2016, 11, 821–841. [Google Scholar] [CrossRef]
- Constantinescu-Aruxandei, D.; Frîncu, R.M.; Capră, L.; Oancea, F. Selenium Analysis and Speciation in Dietary Supplements Based on Next-Generation Selenium Ingredients. Nutrients 2018, 10, 1466. [Google Scholar] [CrossRef]
- Poblaciones, M.J.; Broadley, M.R. Foliar selenium biofortification of broccolini: effects on plant growth and mineral accumulation. Journal of Horticultural Science & Biotechnology 2022, 97, 730–738. [Google Scholar] [CrossRef]
- Liu, H.D.; Xiao, C.M.; Qiu, T.C.; Deng, J.; Cheng, H.; Cong, X.; Cheng, S.Y.; Rao, S.; Zhang, Y. Selenium Regulates Antioxidant, Photosynthesis, and Cell Permeability in Plants under Various Abiotic Stresses: A Review. Plants-Basel 2023, 12. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, R.; Waraich, E.A.; Nawaz, F.; Ashraf, M.Y.; Khalid, M. Selenium (Se) improves drought tolerance in crop plants–a myth or fact? Journal of the Science of Food and Agriculture 2016, 96, 372–380. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.; Wang, J.H.; Wu, H.H.; Yuan, Q.H.; Wang, J.H.; Cui, J.; Lin, A.J. Effects of selenium fertilizer application and tomato varieties on tomato fruit quality: A meta-analysis. Scientia Horticulturae 2022, 304. [Google Scholar] [CrossRef]
- Dima, S.-O.; Neamțu, C.; Desliu-Avram, M.; Ghiurea, M.; Capra, L.; Radu, E.; Stoica, R.; Faraon, V.-A.; Zamfiropol-Cristea, V.; Constantinescu-Aruxandei, D. Plant Biostimulant Effects of Baker’s Yeast Vinasse and Selenium on Tomatoes through Foliar Fertilization. Agronomy 2020, 10, 133. [Google Scholar] [CrossRef]
- Dima, Ș.-O.; Constantinescu-Aruxandei, D.; Tritean, N.; Ghiurea, M.; Capră, L.; Nicolae, C.-A.; Faraon, V.; Neamțu, C.; Oancea, F. Spectroscopic Analyses Highlight Plant Biostimulant Effects of Baker’s Yeast Vinasse and Selenium on Cabbage through Foliar Fertilization. Plants 2023, 12, 3016. [Google Scholar] [PubMed]
- Kumar, A.; Prasad, K.S. Role of nano-selenium in health and environment. Journal of Biotechnology 2021, 325, 152–163. [Google Scholar] [CrossRef] [PubMed]
- Devi, M.S.; Srinivasan, S.; Muthuvel, A. Selenium nanomaterial is a promising nanotechnology for biomedical and environmental remediation: A detailed review. Biocatalysis and Agricultural Biotechnology 2023, 102766. [Google Scholar] [CrossRef]
- Manjunatha, C.; Rao, P.P.; Bhardwaj, P.; Raju, H.; Ranganath, D. New insight into the synthesis, morphological architectures and biomedical applications of elemental selenium nanostructures. Biomedical Materials 2021, 16. [Google Scholar] [CrossRef]
- Hadrup, N.; Ravn-Haren, G. Absorption, distribution, metabolism and excretion (ADME) of oral selenium from organic and inorganic sources: A review. Journal of Trace Elements in Medicine and Biology 2021, 67. [Google Scholar] [CrossRef] [PubMed]
- Gu, X.; Gao, C.Q. New horizons for selenium in animal nutrition and functional foods. Animal Nutrition 2022, 11, 80–86. [Google Scholar] [CrossRef] [PubMed]
- Ferrari, L.; Cattaneo, D.; Abbate, R.; Manoni, M.; Ottoboni, M.; Luciano, A.; von Holst, C.; Pinotti, L. Advances in selenium supplementation: From selenium-enriched yeast to potential selenium-enriched insects, and selenium nanoparticles. Animal Nutrition 2023, 14, 193–203. [Google Scholar] [CrossRef]
- Parvin, S.; Khan, S.; Alam, P.; Khan, T.H.; Khataibeh, M.; Khan, M.A.; Samad, A.; Baker, A.; Mansoor, S. A Review on Potentialities of Selenium Nanoparticles and Its Application Using Air Borne Fungus. Applied Ecology and Environmental Sciences 2021, 9, 607–612. [Google Scholar] [CrossRef]
- Bisht, N.; Phalswal, P.; Khanna, P.K. Selenium nanoparticles: A review on synthesis and biomedical applications. Materials Advances 2022, 3, 1415–1431. [Google Scholar] [CrossRef]
- Wadhwani, S.A.; Shedbalkar, U.U.; Singh, R.; Chopade, B.A. Biogenic selenium nanoparticles: current status and future prospects. Applied microbiology and biotechnology 2016, 100, 2555–2566. [Google Scholar] [CrossRef]
- Vijayakumar, S.; Chen, J.; Divya, M.; Durán-Lara, E.F.; Prasannakumar, M.; Vaseeharan, B. A Review on Biogenic Synthesis of Selenium Nanoparticles and Its Biological Applications. Journal of Inorganic and Organometallic Polymers and Materials 2022, 1–16. [Google Scholar] [CrossRef]
- Arunthirumeni, M.; Veerammal, V.; Shivakumar, M.S. Biocontrol efficacy of mycosynthesized selenium nanoparticle using Trichoderma sp. on insect pest Spodoptera litura. Journal of Cluster Science 2022, 33, 1645–1653. [Google Scholar] [CrossRef]
- Reddy, B.; Bandi, R. Synthesis of selenium nanoparticles by using microorganisms and agri-based products. In Agri-Waste and Microbes for Production of Sustainable Nanomaterials; Elsevier: 2022; pp. 655-683.
- Mates, I.; Antoniac, I.; Laslo, V.; Vicas, S.; Brocks, M.; Fritea, L.; Milea, C.; Mohan, A.; Cavalu, S. Selenium nanoparticles: Production, characterization and possible applications in biomedicine and food science. Sci. Bull. B Chem. Mater. Sci. UPB 2019, 81, 205–216. [Google Scholar]
- Zhang, H.; Zhou, H.; Bai, J.; Li, Y.; Yang, J.; Ma, Q.; Qu, Y. Biosynthesis of selenium nanoparticles mediated by fungus Mariannaea sp. HJ and their characterization. Colloids and Surfaces A: Physicochemical and Engineering Aspects 2019, 571, 9–16. [Google Scholar] [CrossRef]
- Ghosh, S.; Ahmad, R.; Zeyaullah, M.; Khare, S.K. Microbial nano-factories: synthesis and biomedical applications. Frontiers in Chemistry 2021, 9, 626834. [Google Scholar] [CrossRef]
- Alghuthaymi, M.A.; Abd-Elsalam, K.A.; AboDalam, H.M.; Ahmed, F.K.; Ravichandran, M.; Kalia, A.; Rai, M. Trichoderma: An Eco-Friendly Source of Nanomaterials for Sustainable Agroecosystems. Journal of Fungi 2022, 8, 367. [Google Scholar] [CrossRef]
- Fouda, A.; Hassan, S.E.-D.; Eid, A.M.; Abdel-Rahman, M.A.; Hamza, M.F. Light enhanced the antimicrobial, anticancer, and catalytic activities of selenium nanoparticles fabricated by endophytic fungal strain, Penicillium crustosum EP-1. Scientific Reports 2022, 12, 1–16. [Google Scholar] [CrossRef]
- Zielonka, A.; Klimek-Ochab, M. Fungal synthesis of size-defined nanoparticles. Advances in Natural Sciences: Nanoscience and Nanotechnology 2017, 8, 043001. [Google Scholar] [CrossRef]
- Abd-Elsalam, K.A. Fungal Cell Factories for Sustainable Nanomaterials Productions and Agricultural Applications; Elsevier: 2022.
- Srivastava, N.; Mukhopadhyay, M. Biosynthesis and structural characterization of selenium nanoparticles using Gliocladium roseum. Journal of Cluster Science 2015, 26, 1473–1482. [Google Scholar] [CrossRef]
- Zare, B.; Babaie, S.; Setayesh, N.; Shahverdi, A.R. Isolation and characterization of a fungus for extracellular synthesis of small selenium nanoparticles. Nanomedicine journal 2013, 1, 13–19. [Google Scholar]
- Sarkar, J.; Dey, P.; Saha, S.; Acharya, K. Mycosynthesis of selenium nanoparticles. Micro & nano letters 2011, 6, 599–602. [Google Scholar]
- Mosallam, F.M.; El-Sayyad, G.S.; Fathy, R.M.; El-Batal, A.I. Biomolecules-mediated synthesis of selenium nanoparticles using Aspergillus oryzae fermented Lupin extract and gamma radiation for hindering the growth of some multidrug-resistant bacteria and pathogenic fungi. Microbial pathogenesis 2018, 122, 108–116. [Google Scholar] [CrossRef]
- Amin, M.A.; Ismail, M.A.; Badawy, A.A.; Awad, M.A.; Hamza, M.F.; Awad, M.F.; Fouda, A. The Potency of fungal-fabricated selenium nanoparticles to improve the growth performance of Helianthus annuus L. and control of cutworm Agrotis ipsilon. Catalysts 2021, 11, 1551. [Google Scholar] [CrossRef]
- Sarkar, J.; Mridha, D.; Davoodbasha, M.A.; Banerjee, J.; Chanda, S.; Ray, K.; Roychowdhury, T.; Acharya, K.; Sarkar, J. A State-of-the-Art Systemic Review on Selenium Nanoparticles: Mechanisms and Factors Influencing Biogenesis and Its Potential Applications. Biological Trace Element Research 2023, 1–37. [Google Scholar] [CrossRef] [PubMed]
- Garza-García, J.J.; Hernández-Díaz, J.A.; Zamudio-Ojeda, A.; León-Morales, J.M.; Guerrero-Guzmán, A.; Sánchez-Chiprés, D.R.; López-Velázquez, J.C.; García-Morales, S. The role of selenium nanoparticles in agriculture and food technology. Biological Trace Element Research 2021, 1–21. [Google Scholar] [CrossRef] [PubMed]
- Hashem, A.H.; Abdelaziz, A.M.; Attia, M.S.; Salem, S.S. Selenium and Nano-Selenium-Mediated Biotic Stress Tolerance in Plants. In Selenium and Nano-Selenium in Environmental Stress Management and Crop Quality Improvement; Hossain, M.A., Ahammed, G.J., Kolbert, Z., El-Ramady, H., Islam, T., Schiavon, M., Eds.; Springer International Publishing: Cham, 2022; pp. 209–226. [Google Scholar]
- Song, J.; Yu, S.; Yang, R.; Xiao, J.; Liu, J. Opportunities for the use of selenium nanoparticles in agriculture. NanoImpact 2023, 31, 100478. [Google Scholar] [CrossRef] [PubMed]
- Samynathan, R.; Venkidasamy, B.; Ramya, K.; Muthuramalingam, P.; Shin, H.; Kumari, P.S.; Thangavel, S.; Sivanesan, I. A Recent Update on the Impact of Nano-Selenium on Plant Growth, Metabolism, and Stress Tolerance. Plants 2023, 12, 853. [Google Scholar] [CrossRef] [PubMed]
- Tyśkiewicz, R.; Nowak, A.; Ozimek, E.; Jaroszuk-Ściseł, J. Trichoderma: The Current Status of Its Application in Agriculture for the Biocontrol of Fungal Phytopathogens and Stimulation of Plant Growth. International Journal of Molecular Sciences 2022, 23, 2329. [Google Scholar] [CrossRef] [PubMed]
- Ogunleye, G.E.; Adebayo-Tayo, B.C.; Oyinlola, K.A. Biological evaluation of extracellular mycosynthesized silver nanoparticles by Trichoderma asperellum. BioMetals 2022, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Castro-Longoria, E. Biosynthesis of Metal-Based NanoparticlesNanoparticles by TrichodermaTrichodermaand Its Potential Applications. In Advances in Trichoderma Biology for Agricultural Applications, Amaresan, N., Sankaranarayanan, A., Dwivedi, M.K., Druzhinina, I.S., Eds.; Springer International Publishing: Cham, 2022; pp. 433-463.
- Khabat, V.; Mansoori, G.A.; Karimi, S. Biosynthesis of silver nanoparticles by fungus Trichoderma Reesei. Insciences J 2011, 1, 65–79. [Google Scholar]
- Gharieb, M.; Wilkinson, S.; Gadd, G. Reduction of selenium oxyanions by unicellular, polymorphic and filamentous fungi: cellular location of reduced selenium and implications for tolerance. Journal of Industrial Microbiology 1995, 14, 300–311. [Google Scholar] [CrossRef]
- Nandini, B.; Hariprasad, P.; Prakash, H.S.; Shetty, H.S.; Geetha, N. Trichogenic-selenium nanoparticles enhance disease suppressive ability of Trichoderma against downy mildew disease caused by Sclerospora graminicola in pearl millet. Scientific Reports 2017, 7, 2612. [Google Scholar] [CrossRef] [PubMed]
- Hu, D.; Yu, S.; Yu, D.; Liu, N.; Tang, Y.; Fan, Y.; Wang, C.; Wu, A. Biogenic Trichoderma harzianum-derived selenium nanoparticles with control functionalities originating from diverse recognition metabolites against phytopathogens and mycotoxins. Food Control 2019, 106, 106748. [Google Scholar] [CrossRef]
- Liang, X.; Perez, M.A.M.-J.; Nwoko, K.C.; Egbers, P.; Feldmann, J.; Csetenyi, L.; Gadd, G.M. Fungal formation of selenium and tellurium nanoparticles. Applied microbiology and biotechnology 2019, 103, 7241–7259. [Google Scholar] [CrossRef] [PubMed]
- Diko, C.S.; Zhang, H.; Lian, S.; Fan, S.; Li, Z.; Qu, Y. Optimal synthesis conditions and characterization of selenium nanoparticles in Trichoderma sp. WL-Go culture broth. Materials Chemistry and Physics 2020, 246, 122583. [Google Scholar] [CrossRef]
- Joshi, S.M.; De Britto, S.; Jogaiah, S.; Ito, S.-i. Mycogenic selenium nanoparticles as potential new generation broad spectrum antifungal molecules. Biomolecules 2019, 9, 419. [Google Scholar] [CrossRef]
- Saravanakumar, K.; Sathiyaseelan, A.; Zhang, X.; Park, S.; Wang, M.-H. Purinoceptor targeted cytotoxicity of adenosine triphosphate-conjugated biogenic selenium nanoparticles in human colon cancer cells. Pharmaceuticals 2022, 15, 582. [Google Scholar] [CrossRef] [PubMed]
- Joshi, S.M.; De Britto, S.; Jogaiah, S. Myco-engineered selenium nanoparticles elicit resistance against tomato late blight disease by regulating differential expression of cellular, biochemical and defense responsive genes. Journal of Biotechnology 2021, 325, 196–206. [Google Scholar] [CrossRef] [PubMed]
- Choi, Y.; Lee, S.Y. Biosynthesis of inorganic nanomaterials using microbial cells and bacteriophages. Nature Reviews Chemistry 2020, 4, 638–656. [Google Scholar] [CrossRef]
- Shoeibi, S.; Mozdziak, P.; Golkar-Narenji, A. Biogenesis of Selenium Nanoparticles Using Green Chemistry. Topics in Current Chemistry 2017, 375, 88. [Google Scholar] [CrossRef]
- Zambonino, M.C.; Quizhpe, E.M.; Jaramillo, F.E.; Rahman, A.; Santiago Vispo, N.; Jeffryes, C.; Dahoumane, S.A. Green synthesis of selenium and tellurium nanoparticles: current trends, biological properties and biomedical applications. International Journal of Molecular Sciences 2021, 22, 989. [Google Scholar] [CrossRef]
- Tsivileva, O.; Pozdnyakov, A.; Ivanova, A. Polymer nanocomposites of selenium biofabricated using fungi. Molecules 2021, 26, 3657. [Google Scholar] [CrossRef] [PubMed]
- Annamalai, J.; Ganesan, S.; Murugan, K.; Janjaroen, D. Recent breakthroughs set by fungal enzymes in the biosynthesis of nanoparticles. In Fungal Cell Factories for Sustainable Nanomaterials Productions and Agricultural Applications; Elsevier: 2023; pp. 131-162.
- Bahrulolum, H.; Nooraei, S.; Javanshir, N.; Tarrahimofrad, H.; Mirbagheri, V.S.; Easton, A.J.; Ahmadian, G. Green synthesis of metal nanoparticles using microorganisms and their application in the agrifood sector. Journal of Nanobiotechnology 2021, 19, 1–26. [Google Scholar] [CrossRef]
- Dhillon, G.S.; Brar, S.K.; Kaur, S.; Verma, M. Green approach for nanoparticle biosynthesis by fungi: current trends and applications. Critical reviews in biotechnology 2012, 32, 49–73. [Google Scholar] [CrossRef]
- Khandel, P.; Shahi, S.K. Mycogenic nanoparticles and their bio-prospective applications: current status and future challenges. Journal of Nanostructure in Chemistry 2018, 8, 369–391. [Google Scholar] [CrossRef]
- Li, G.; He, D.; Qian, Y.; Guan, B.; Gao, S.; Cui, Y.; Yokoyama, K.; Wang, L. Fungus-mediated green synthesis of silver nanoparticles using Aspergillus terreus. Int J Mol Sci 2012, 13, 466–476. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.-Y.; Lo, C.-T.; Chen, C.; Liu, M.-Y.; Chen, J.-H.; Peng, K.-C. Efficient isolation of anthraquinone-derivatives from Trichoderma harzianum ETS 323. Journal of Biochemical and Biophysical Methods 2007, 70, 391–395. [Google Scholar] [CrossRef] [PubMed]
- Fuentes-Cervantes, A.; Ruiz Allica, J.; Calderón Celis, F.; Costa-Fernández, J.M.; Ruiz Encinar, J. The Potential of ICP-MS as a Complementary Tool in Nanoparticle–Protein Corona Analysis. Nanomaterials 2023, 13, 1132. [Google Scholar] [PubMed]
- Nwoko, K.C.; Liang, X.; Perez, M.A.M.J.; Krupp, E.; Gadd, G.M.; Feldmann, J. Characterisation of selenium and tellurium nanoparticles produced by Aureobasidium pullulans using a multi-method approach. Journal of Chromatography A 2021, 1642, 462022. [Google Scholar] [CrossRef]
- Nikam, P.B.; Salunkhe, J.D.; Minkina, T.; Rajput, V.D.; Kim, B.S.; Patil, S.V. A review on green synthesis and recent applications of red nano Selenium. Results in Chemistry 2022, 100581. [Google Scholar] [CrossRef]
- Biswas, K.C.; Barton Ll Fau - Tsui, W.L.; Tsui Wl Fau - Shuman, K.; Shuman K Fau - Gillespie, J.; Gillespie J Fau - Eze, C.S.; Eze, C.S. A novel method for the measurement of elemental selenium produced by bacterial reduction of selenite.
- El-Sayyad, G.S.; El-Bastawisy, H.S.; Gobara, M.; El-Batal, A.I. Gentamicin-assisted mycogenic selenium nanoparticles synthesized under gamma irradiation for robust reluctance of resistant urinary tract infection-causing pathogens. Biological trace element research 2020, 195, 323–342. [Google Scholar] [CrossRef]
- Hamley, I.W. Small-angle scattering: theory, instrumentation, data and applications John Wiley & Sons, Inc.,: 111 River Street, Hoboken, NJ 07030, USA, 2022; p. 278.
- Gommes, C.J.; Jaksch, S.; Frielinghaus, H. Small-angle scattering for beginners. Journal of Applied Crystallography 2021, 54, 1832–1843. [Google Scholar] [CrossRef]
- Li, T.; Senesi, A.J.; Lee, B. Small Angle X-ray Scattering for Nanoparticle Research. Chemical Reviews 2016, 116, 11128–11180. [Google Scholar] [CrossRef] [PubMed]
- Turkevich, J.; Hubbell, H.H. Low Angle X-Ray Diffraction of Colloidal Gold and Carbon Black1a. Journal of the American Chemical Society 1951, 73, 1–7. [Google Scholar] [CrossRef]
- Gablech, E.; Fohlerova, Z.; Svec, K.; Zales, F.; Benada, O.; Kofronova, O.; Pekarkova, J.; Caha, O.; Gablech, I.; Gabriel, J.; et al. Selenium nanoparticles with boron salt-based compound act synergistically against the brown-rot Serpula lacrymans. International Biodeterioration & Biodegradation 2022, 169. [Google Scholar] [CrossRef]
- Apryatina, K.V.; Murach, E.I.; Amarantov, S.V.; Erlykina, E.I.; Veselov, V.S.; Smirnova, L.A. Synthesis of a Bioactive Composition of Chitosan-Selenium Nanoparticles. Applied Biochemistry and Microbiology 2022, 58, 126–131. [Google Scholar] [CrossRef] [PubMed]
- Borgatta, J.R.; Lochbaum, C.A.; Elmer, W.H.; White, J.C.; Pedersen, J.A.; Hamers, R.J. Biomolecular corona formation on CuO nanoparticles in plant xylem fluid. Environmental Science: Nano 2021, 8, 1067–1080. [Google Scholar] [CrossRef]
- Chen, W.; Li, X.; Cheng, H.; Zhan, X.; Xia, W. Synthesis, characterization, and anticancer activity of protamine sulfate stabilized selenium nanoparticles. Food Research International 2023, 164, 112435. [Google Scholar] [CrossRef] [PubMed]
- Dahman, Y. Chapter 5 - Nanoparticles**By Yaser Dahman, Hoda Javaheri, Jiafu Chen, and Basel Al-Chikh Sulaiman. In Nanotechnology and Functional Materials for Engineers, Dahman, Y., Ed.; Elsevier: 2017; pp. 93-119.
- Bhattacharjee, S. DLS and zeta potential–what they are and what they are not? Journal of controlled release 2016, 235, 337–351. [Google Scholar] [CrossRef] [PubMed]
- Ramírez-Valdespino, C.A.; Orrantia-Borunda, E. Trichoderma and nanotechnology in sustainable agriculture: a review. Frontiers in Fungal Biology 2021, 61. [Google Scholar] [CrossRef]
- Shahbaz, M.; Akram, A.; Raja, N.I.; Mukhtar, T.; Mashwani, Z.U.; Mehak, A.; Fatima, N.; Sarwar, S.; Haq, E.U.; Yousaf, T. Green synthesis and characterization of selenium nanoparticles and its application in plant disease management: a review. Pakistan Journal of Phytopathology 2022, 34, 189–102. [Google Scholar] [CrossRef]
- Helmy, E.; Salah, R.; El-Shazly, M.M.; Alqhtani, A.; Pokoo-Aikins, A.; Yosri, M. Investigation of the Impact of Mycogenic Titanium and Selenium Nanoparticles on Fusarium Wilt Infection of Tomato Plant. Journal of Pure and Applied Microbiology 2023, 17. [Google Scholar] [CrossRef]
- Bărbieru, O.-G.; Dimitriu, L.; Călin, M.; Răut, I.; Constantinescu-Aruxandei, D.; Oancea, F. Plant Biostimulants Based on Selenium Nanoparticles Biosynthesized by Trichoderma Strains. Proceedings 2019, 29, 95. [Google Scholar]
- Zohra, E.; Ikram, M.; Omar, A.A.; Hussain, M.; Satti, S.H.; Raja, N.I.; Ehsan, M. Potential applications of biogenic selenium nanoparticles in alleviating biotic and abiotic stresses in plants: A comprehensive insight on the mechanistic approach and future perspectives. Green Processing and Synthesis 2021, 10, 456–475. [Google Scholar] [CrossRef]
- Dobias, J.; Suvorova, E.I.; Bernier-Latmani, R. Role of proteins in controlling selenium nanoparticle size. Nanotechnology 2011, 22, 195605. [Google Scholar] [CrossRef] [PubMed]
- Nancharaiah, Y.V.; Lens, P.N.L. Ecology and Biotechnology of Selenium-Respiring Bacteria. Microbiology and Molecular Biology Reviews 2015, 79, 61–80. [Google Scholar] [CrossRef]

Table 1.
Parameters of the biosynthesis of trichogenic selenium nanoparticles.
| Trichoderma strain | Se precursor | Substrate concentration (mM) | Types of synthesis | Size (nm) | Ref. |
|---|---|---|---|---|---|
| Trichoderma sp. | Na2SeO3 | 20 mM | in vitro (intracellular)* | 40-100 | [23] |
| T. reesei | Na2SeO3 | 1, 5, 10 mM | in vivo (intracellular, extracellular) |
- | [46] |
|
T. brevicompactum T. asperellum, T. atroviride, T. harzianum, T. virens, T. longibrachiatum |
Na2SeO3 | 25 mM | in vitro (intracellular, extracellular, cell walls) |
49- 312 | [47] |
|
T. harzianum (three strains), T. koningii, T. longibranchiatum, T. atroviride, T. asperellum, T. virens |
Na2SeO3 | 5 mM | in vitro (intracellular, extracellular) |
50-60 | [48] |
| T. harzianum | Na2SeO3 | 1 mM | in vivo (intracellular, extracellular) |
[49] | |
| Trichoderma sp. WL-Go | SeO2 | 2 mM | in vivo (intracellular, extracellular) |
147 | [50] |
| T. atroviride (Tri_AtJSB2) sp. | Na2SeO3 | 25 mM | in vitro (intracellular, extracellular, cell walls) |
60- 123 | [51] |
| T. harzianum | Na2SeO3 | 25 mM | in vitro (intracellular) | 26 | [52] |
| T. atroviride (Tri_AtJSB2) sp. | Na2SeO3 | 25 mM | in vitro (extracellular) | 60-123 (Z av. 98.5, ref. 32) | [53] |
*in vitro intracellular – mycelium lysate, in vitro extracellular – culture filtrate, Z av – Z average.
Table 2.
Composition and physicochemical characteristics of bio-corona capping trichogenic SeNPs.
| Trichoderma strain | Bio-corona composition | Zeta potential (mV) | Crystallinity | Methods | Ref. |
|---|---|---|---|---|---|
| Trichoderma sp. | apparently, proteins | - | crystalline | FTIR, XRD | [23] |
|
T. brevicompactum T. asperellum, T. atroviride, T. harzianum, T. virens, T. longibrachiatum |
proteins | −200 to + 11.8 | amorphous/ nano-crystalline | FTIR, XRD | [47] |
|
T. harzianum (three strains), T. koningii, T. longibranchiatum, T. atroviride, T. asperellum, T. virens |
Proteins, organic acids, amino acids, sugars, intermediates from the carbohydrate metabolism | - | amorphous | FTIR, LC-MS/MS, XRD |
[48] |
| T. harzianum | - | - | amorphous + Se oxide |
XRD | [49] |
| Trichoderma sp. WL-Go | proteins, sugars | -24.6 | crystalline | FTIR, zeta potential, XRD | [50] |
| T. atroviride (Tri_AtJSB2) sp. | proteins | −49.3 to −43.7 | crystalline | FTIR, zeta potential, XRD | [51] |
| T. harzianum | proteins, hydroxyl molecules |
−37.8 ± 0.36 | crystalline | FTIR, zeta potential, XRD | [52] |
Table 3.
Biological effects of trichogenic SeNPs with applications in agriculture and food industry.
Table 3.
Biological effects of trichogenic SeNPs with applications in agriculture and food industry.
| Trichoderma strain | Phytopathogen / pest | Plant | Effect | Ref. |
|---|---|---|---|---|
| Trichoderma sp. | Spodoptera litura | castor leaves | larvicidal and antifeedant activity | [23] |
|
T. brevicompactum, T. asperellum, T. atroviride, T. harzianum, T. virens T. longibrachiatum |
Sclerospora graminicola | pearl millet [Pennisetum glaucum (L.) R. Br.] | Sporulation and downy mildew inhibition | [47] |
|
T. harzianum (three strains), T. koningii, T. longibranchiatum, T. atroviride, T. asperellum, T. virens |
Fusarium verticillioide, Alternaria alternata, F. graminearum | - | Antifungal, functional biocontrol of mycotoxins | [48] |
| T. atroviride (Tri_AtJSB2) sp. | Phytophthora infestans, Pyricularia grisea, Colletotrichum capsica, Alternaria solani | one-month-old chili (Capsicum annuum L.) cultivar ArkaKhyati and tomato leaves | zoosporicidal and antifungal activities | [51] |
| T. atroviride (Tri_AtJSB2) sp. | Phytophthora infestans | tomato | priming effect, enhanced resistance to late blight disease | [53] |
| T. harzianum | F. oxysporum, Aspergillus niger, Aspergillus flavus, Aspergillus fumigatus, Penicillium marnefeii, Candida albicans, Candida lipolytica, Salmonella typhimurium, Bacillus subtilis, Pseudomonas aeruginosa, Methicillin Resistant Staphylococcus aureus (MRSA), Staphylococcus aureus, Escherichia coli, Enterococcus faecalis | tomato | antimicrobial activity, reduction of Fusarium wilt infection | [81] |
| Trichoderma spp. | Fusarium sp. | Vigna radiata | Reduced toxicity, antifungal, biostimulant | [82] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



