
Submitted:
08 December 2023
Posted:
11 December 2023
You are already at the latest version
Abstract
Noncoding RNAs (ncRNAs) including circular RNAs (circRNAs) and N6-methyladenosine (m6A) have been shown to play a critical role in the development of various diseases including obesity and metabolic disorder-associated fatty liver disease (MAFLD). Obesity is a chronic disease considered by excessive fat accumulation in the body, which has been increased lately and is the foremost risk factor for the MAFLD. Causes of obesity may involve the interaction of genetic, behavioral, and social factors. The m6A RNA methylation might add a novel inspiration for understanding the development of obesity and MAFLD with post-transcriptional regulation of gene expression. In particular, circRNAs, microRNAs (miRNAs) and m6A might be implicated in the progression of MAFLD. Interestingly, m6A modification could modulate the translation, degradation, and other functions of ncRNAs. miRNAs/circRNAs could also modulate m6A modifications by affecting writers, erasers, and readers. In turn, ncRNAs could modulate the expression of m6A regulators in different ways. However, there is limited evidence on how these ncRNAs and m6A interact to affect the promotion of liver diseases. M6A has been appeared to occur in DNA, RNA and proteins, which may be associated with several biological properties. This study would provide a mechanistic understanding of the m6A modification and ncRNAs in the association on liver diseases, especially for MAFLD. Comprehension of the association between m6A modification and ncRNAs may contribute to the development of treatment tactics for MAFLD.
Keywords:
N6-methyladenosine
; noncoding RNA
; circular RNA
; RNA-binding protein
; APRO family protein
; NAFLD
; MAFLD
; probiotics
; YTHDF2
1. Introduction
Circular RNAs (circRNAs) are a class of single-strand closed RNA molecules [1]. They are generated by the reverse splicing process, which might be involved in the development of many diseases including various liver diseases. Most circRNAs are non-coding RNAs (ncRNAs) while part of cytoplasmic circRNAs with coding capability could be translated into peptides, which may contribute to several important biological and/or physiological processes, since many circRNAs originate form exons and reside in the cytoplasm [2]. Therefore, innovative insight for circRNA-based therapeutic and/or diagnostic strategies has been implied [3]. Fortunately, circRNAs are also observed in eukaryotic cells [4], whose expression pattern might enable them to support the clinical diagnosis as biological markers in many types of disease [5]. In particular, circRNAs are highly stable and easily detected in the circulation, which have been emerging valuable non-coding RNAs representing a promising non-invasive approach for predicting NAFLD [6].
Generally, ncRNAs may be short of the capability in translating into protein/peptide, however, ncRNAs could work as crucial transcripts to regulate the expression of other genes [7]. These ncRNAs may be typically divided into several groups including microRNAs (miRNAs), long ncRNAs (lncRNAs), and circRNAs [8]. In addition, the n6-methyladenosine (m6A) modification in those ncRNAs have been increasingly described to have a dense association with pathological machineries of various diseases. Several crucial regulators derived from m6A modification could affect the profound function of ncRNAs to take part in the initiation and/or progression of related diseases. Remarkably, ncRNAs might mediate the downstream signaling to impact the expression of “readers”, “writers”, and/or “erasers” for the additional modification of m6A functions [9]. The m6A RNA modification may highly regulate hepatic function and development of liver diseases, providing some directions to understand the mechanism of malicious activities in the development of liver diseases [10]. (Figure 1)
The m6A, the most prevalent modification associated with eukaryotic RNAs, could affect several steps of RNA metabolism including translation, splicing, in addition to the decay/stability of RNAs, which may be the epitranscriptomic modification that occurs on the N6 position of adenosine [11]. It is the most abundant modification in eukaryotic RNAs, accounting for nearby 0.4 % of all adenosines in RNA, which may be involved in the functional alteration of RNAs [12].Interestingly, the m6A modification may be closely associated with the initiation and/or development of obesity and non-alcoholic fatty liver disease (NAFLD), which may progress to end-stage liver disease [13]. Obesity is the main risk factor for the NAFLD. The feature of the disease is hepatic steatosis with the accumulation of surplus fat in the liver and metabolic liver dysfunction. Therefore, it has been suggested that NAFLD should be retitled as metabolic disorder-associated fatty liver disease (MAFLD). Here, we use the term MAFLD instead of NAFLD. MAFLD is often supposed to be practically asymptomatic. However, many MAFLD patients complain of exhaustion, which may disturb their quality of life (QOL). Currently, there are no specific pharmacotherapies of MAFLD. Mostly, the treatment may include lifestyle adjustment and medicines for improving fat metabolism and balancing oxidation. Therefore, further research and development of novel therapeutic tactics are seriously required [13]. In addition, obesity is a chronic metabolic disease that is closely related to type 2 diabetes mellitus, cardiovascular diseases, and osteoarthritis [14]. The prevalence of obesity is increasing rapidly, which is realized as a global public health burden [14]. However, the precise mechanism and role of m6A-modified ncRNAs in MAFLD are not well understood. The m6A modification might play a role in the formation of specific and complexed microenvironment in liver [15]. In fact, recent studies have shown that nucleotide methylation may be directly associated with the inflammatory grade, lipid synthesis, and/or oxidative stress, playing a crucial role in the progression of MAFLD [15]. Therefore, m6A modification might be a key player in the pathogenesis of MAFLD, which may provide new mechanistic insight into MAFLD [16]. Recent studies have explored the roles of m6A RNA methylation in the pathogenesis of liver diseases, providing new insights for studying the molecular mechanism of liver diseases [17]. Epigenetic modification occurring in RNA has become the hotspot of the field, offering the potential of m6A as a treatment option for several liver diseases.
2. Association of m6A and ncRNAs in various liver diseases
M6A is a highly prevalent modification in mRNAs and ncRNAs that affects the epigenetic effects, which could play a critical role in cellular physiology and/or pathology processes. The effects of m6A are determined by ‘reader’ (recognition) ‘writer’ (methyltransferase), and ‘eraser’ (demethylase) molecules [18]. (Figure 1) Writers are methyltransferases, including Wilms tumor 1 associated protein (WTAP), methyltransferase-like 3 (METTL3), Vir-like m6A methyltransferase-associated (VIRMA) and methyltransferase-like 14 (METTL14).The AlkB homolog 5 (ALKBH5) and fat mass and obesity-associated protein (FTO) are known as erasers, which are demethylases that could reverse the methylation. The m6A modifications are usually distinguished by m6A-binding reader molecules, including YT521-B homology (YTH) domain-containing-1 and -2 (YTHDC-1 and -2), YTH domain family proteins 1-3 (YTHDF1-3). In addition, heterogeneous nuclear ribonucleoprotein (HNRNP) family including HNRNPC and heterogeneous nuclear ribonucleoprotein A2/B1 (HNRNPA2B1) may have also YTH domain for binding the m6A site [19]. Furthermore, it has been proposed that eukaryotic initiation factor 3 (eIF3) might initiate the translation in a cap-independent manner by binding to the m6A sites in the 5′-UTR of mRNAs, while insulin-like growth factor 2 mRNA-binding protein 1/2/3 (IGF2BP1/2/3) could increase the stability of the target RNAs [19]. The m6A modification could play an indispensable role in the proliferation and/or differentiation of hepatocytes. For example, the m6A content might be decreased in patients with type 2 diabetes, and mRNA expression levels of FTO, METTL3, METTL14, and WTAP seems to be enhanced. Remarkably, the m6A content may be negatively associated with the expression level of METTL3, METTL14, and FTO mRNA. Therefore, high glucose could upregulate the protein expression of FTO in HepG2 cells [20]. In addition, the FTO could decrease the concentration of m6A in RNAs transcript, thereby regulating the expression of related transcripts [21]. It has been demonstrated that m6A modification has an impact on the regulation of carboxylesterase 2 (CES2), a serine esterase responsible for the hydrolysis of endogenous substrates including triglycerides and diacylglycerides, affecting lipid metabolism. [22]. Furthermore, the phosphorylation of p70 ribosomal subunit 6 kinase (S6K) and/or mammalian target of rapamycin (mTOR) might be reduced by the knockout or knockdown of WTAP [23].
Circular RNAs (circRNAs) could also play a part in physiological and/or pathological processes through the similar pathway as a new type of molecule with intriguing functions [24]. In addition, proteins encoded by circRNAs have been confirmed to be related to multiple pathophysiological processes including immunity [25]. The m6A modifications might regulate the metabolism of circRNAs. It has been shown that circRNAs containing m6A residues could be translated into protein by non-cap-dependent structures [26]. The m6A-modified circRNA might be recognized by the YTHDF2 [27], in which the open reading framework (ORF) might be verified for the translational capability of the circRNA. Several effects of the peptide/protein from circRNA translation have been identified [28]. Whereas the FTO demethylase could diminish the rate of circRNAs translation, the METTL3 methyltransferase could boost the translation rate [29].
Remarkably, these proteins may be also significant players in the development of MAFLD. Liver is a vital metabolic and digestive organ in the pathophysiological processes. Recent studies have suggested that m6A RNA modification could regulate hepatic function and development of liver diseases [10,30]. For example, METTL14 could modulate the expression of miR-34a-5p, impairing mitochondrial homeostasis in MAFLD [31]. In addition, many of these epigenetic factors are amenable to dietary or lifestyle interventions. For example, several interventions may include diets low in carbohydrates, free sugars, fructose, and lipids, in addition to healthy eating patterns and probiotics [32]. The nutritional and metabolic status of individuals may interact with epigenetics at different levels [33]. Epigenetic modifications are potentially reversible and could provide new therapeutic avenues for treating these diseases using epigenetic modulators [34]. A number of studies have revealed that m6A modification is closely related to the pathology of MAFLD [35]. Recently, studies have found that the regulation of immune cells by m6A modification may also have a role in MAFLD.
3. ncRNAs and RNA-binding proteins involved in mRNA translation
Accumulating evidences have shown that circRNAs as a sponge to absorb miRNA function, involving their connection in the biological process of MAFLD [36,37]. In fact, several circRNAs have recognized to be correlated with miRNAs for mediating the expression of m6A regulators. In addition, circRNAs may have several actions in adjusting cellular function, which might contain to compete the target endogenous RNAs such as miRNAs, known as sponging [38]. For example, it has been shown that Circ_0057558 could act as a miR-206 sponge to inhibit the repression of the ROCK1/AMPK signaling and facilitates lipogenesis, which might contribute to the progression of MAFLD [37]. A single circRNA could attach to one or more miRNAs through its circular sequences. The sponging of miRNA could play a key role within several types of cells in various diseases. Therefore, translational control of RNAs might be a critical factor for the related disease [39].
In the translation of mRNAs, the circularization by binding eIF4G on the 5′-cap of the mRNA and poly(A) binding protein (PABP) on the poly(A) tail might start the mRNA translation. Therefore, obstructing the interaction between eIF4G and PABP might constrain the mRNA translation. In addition, previous studies have shown that PABP-interacting protein 2 (Paip2) could take part in the eIF4G binding for binding to the PABP with a common amino acid sequence of PABP [40]. Then, several miRNAs could be associated to AGO proteins within the complementary 3′ UTR sites. The AGO-miRNA complex is additionally modified by adjacent RNA-binding proteins (RBPs). CircRNAs could also control the gene expression of RNAs by attaching targeted miRNAs in several cells including immune cells and/or cancer stem cells [41]. In cases, certain circRNA could also impede the translation by binding to neighboring PABP [42]. The association between circRNAs and PABP may impact on the combination between PABP and eIF4G, which might specifically influence the translation of several mRNAs [42]. (Figure 2)
Several members of the APRO family have been shown to be implicated in the cytoplasmic mRNA deadenylation and/or RNA turnover [43,44]. Interestingly, the association between APRO family proteins and PABP has been also discovered [45,46]. The N-terminal conserved APRO domain could bind to DNA-binding transcription factors as well as deadenylase subunits of the CCR4/NOT complex [47]. Likewise, some of the APRO family proteins could interact with the cytoplasmic PABP and the poly(A) nuclease complex CCR4/CAF1 [45,46]. APRO proteins, such as Tob1 and Tob2, could concurrently interact with CAF1 protein and PABP, which might trigger the deadenylation of several mRNAs [46]. In addition, Tob1 and Tob2 proteins have an long C-terminal domain with two PAM2 motifs [48]. The antiproliferative effects of Tob1 have been proposed to be involved in the utilization of CAF1/CCR4 complex [49], which may indicate that APRO proteins might exert their function by modulating the metabolism and/or turnover of mRNAs [50]. Another member of APRO family, BTG2, could also interact with CAF1 deadenylase through its APRO domain to regulate cell proliferation [51]. Similarly, the destabilization of mRNAs by BTG1 and BTG2 might contribute for keeping cell quiescence [52]. CircRNAs and miRNAs could inhibit the expression of various mRNAs, which may employ the mRNAs destabilization with APRO family proteins [53]. Therefore, the miRISC complex could interact with PABP, CAF1, and/or CCR4 deadenylases [54]. Importantly, an important component of the miRISC might be the PABP and APRO family proteins, which might be necessary for the miRNA-mediated deadenylation [54]. Since the APRO family proteins have the potential to interact with the CCR4/CAF1 complex, APRO family proteins could be also a significant modulator of miRNAs and circRNAs. Remarkably, the expression of APRO family molecules could be also regulated by certain miRNAs and/or circRNAs [55]. (Figure 2)
Interestingly, it has been reported that overexpression of miR-183-5p in mice could direct to upregulate lipogenic genes for the hepatic triglyceride (TG) accumulation, whereas inhibition of miR-183-5p might improve the hepatic TG accumulation [56]. In this case, Btg1, a member of APRO family, might be a direct target of miR-183-5p [56]. In addition, the BTG2 signaling could play an important role in the development of MAFLD via the regulation of endoplasmic reticulum stress and inflammation. Moreover, NF-κB pathway activation can upregulate BTG2 expression under stress conditions [58]. The correlation between circRNA-HIPK3 and miRNA-29a as well as the correlation between circRNA-0046367 and miRNA-34a expression could influence to the regulation of the Wnt/β catenin pathway, which may provide a new target for treatment of MAFLD [59], in which circRNA sponges may be provided with miRNA-binding sites, thus, overexpression of circRNA could lead to the miRNA suppression and/or upregulation of miRNA target gene expression [60]. In these ways, several studies have reported the role of circRNAs in reducing lipotoxicity and oxidative stress in MAFLD and/or NASH.
4. Attention to YTHDF2, an m6A reader molecule, in the pathogenesis of obesity
It is probable that CCR4/NOT-mediated deadenylation and subsequent 3′-to-5′ exoribonucleolytic degeneration of RNAs can also start the degradation of the m6A mRNAs. However, m6A-containing mRNAs might undergo several different pathways of the RNAs degradation, which may involve the deadenylation via the YT521-B homology (YTH) domain-containing family protein 2 (YTHDF2), an m6A reader protein)-CCR4/NOT (deadenylase) complex as well as via the endoribonucleolytic cleavage by the YTHDF2-ribonuclease (RNase) complex. Consequently, another pathway may be the endoribonucleolytic cleavage by YTHDF2-RNase complex [61]. Suppressed levels of readers/erasers such as YTHDF2 may lead to a hypo-metabolic state in obesity [62]. It has been described that YTHDF2 could regulate the specific mRNA degradation. M6A modification may contribute to metabolic reprogramming in MAFLD, which could play major roles in regulating the innate immune responses [63]. It has been shown that m6A modification is critical in the control of macrophages-directed metabolic programming through the regulation of immune transcripts in MAFLD and obesity [63]. YTHDF2 could bind transcripts carrying m6A in 3’UTR to induce mRNA degradation partially through recruiting the CCR4-NOT deadenylase complex [64]. The binding site of YTHDF2 on m6A might be usually positioned in the 3’-UTR of mRNAs [64,65]. If the m6A is located at the 5’-UTR of mRNAs, YTHDF2 could subsequently facilitate protein translation [66]. Interestingly, it has been suggested that resveratrol may alleviate liver lipid metabolism disorders through the changes of m6A levels in the liver of mice [67]. Moreover, alters in the m6A RNA methylation and lipid metabolism in various tissues may be inter-related mechanisms [68]. Again, the YTHDF2 could modulate the degradation of several mRNAs containing m6A [69]. Remarkably, YTHDF2 could competitively interact with YTHDF3 to accelerate the degradation of specific mRNAs through the Argonaute 2 (AGO2) in a manner independent of m6A [70]. Ago2 is the chief component of the RNA-induced silencing complex. It has been shown that hepatic Ago2 regulates the function of peroxisome proliferator-activated receptor α (PPARα) for oxidative metabolism [71]. YTHDF2 could also control glucose metabolism in the development of various diseases [72]. (Figure 2)
Some m6A-containing circular RNAs could associate with YTHDF2, which may be downregulated by certain RNases [73]. YTHDF2 could directly interact with the SH domain of CNOT1 to recruit the CCR4-NOT complex, thus inducing the deadenylation and/or degradation of m6A-modified circRNAs [74]. The m6A modification and circRNAs could play a key role in the progression of various diseases including MAFLD, which may contribute to some clinical features. For example, m6A methylation and circRNAs could be employed as novel biomarkers for diagnosis and/or therapy prognosis [75]. Several circRNAs could function as sponges of miRNAs, which might play a major role in the pathogenesis of NAFLD by affecting fundamental genes such as PPARs [76]. More studies are required to further explore the relationship between circRNAs and m6A modification in the development of MAFLD.
5. Relationship between YTHDF2 and Diabetes mellitus
Diabetes mellitus (DM) is a systemic metabolic disease, where the body becomes incapable of generating an appropriate quantity of insulin. DM also carries a clinical burden as it is associated with obesity, metabolic syndrome, and MAFLD. There is an enormous economic burden due to healthcare use. Increasing evidence shows that m6A-binding proteins have a vital effect on the development and progression of diabetes [77]. In addition, YTHDF2 and METTL14 may promote hepatic insulin resistance [77].YTHDC1 could also cause glucose intolerance by regulating the splicing of polyadenylation-specific factor 6 (CPSF6) mRNA [78]. As YTHDF2 is an m6A reading protein that could modulate RNA stability, transcription, and translation, m6A modifications of RNAs recognized with the YTHDF2 might be profoundly implicated in DM progression. For example, , activated tumor necrosis factor (TNF) signaling in the TME may promote the YTHDF2 expression, which consequently regulates NF-κB signaling to maintain intra-tumoral Treg survival by accelerating the degradation of m6A-modified transcripts that encode NF-κB-negative regulators [79]. It is well-known that NF-κB signaling is deeply involved in the pathogenesis of DM and/or NAFLD [80]. Therefore, these effects may present new insights into the effect of m6A modification on treating diabetes. Consequently, RNA modifications with N6-methyladenosine m6A have collected broad attention due to their involvement in the onset and progression of DM, obesity and/or MAFLD.
It has been reported that miRNA is also a significant bioactive molecule that induces posttranscriptional gene regulation in eukaryotes [81], which could modulate the levels of m6A by targeting the 3'-untranslated region (3'- UTR) of YTHDF2 mRNA in hepatic cells [82]. Interestingly, miR-6125 could inhibit the proliferative ability of colorectal cancer cells by targeting the YTHDF2 mRNA [83], which is also expressed in the cytoplasm of hepatocytes in patients with autoimmune hepatitis [84]. Autoimmune liver diseases include autoimmune hepatitis as well as MAFLD may have an increased risk of DM with displaying glucose disturbances like insulin resistance [85]. In addition, YTHDF2 could increase the degradation of oncogenes or tumor suppressor genes in the m6A-dependent manner [86]. p53 is well known for its tumor suppressor role, which is associated with increased risk for DM, obesity, metabolism, and inflammation. [87,88]. YTHDF2 and m6A modification could regulate the process of autophagy [89], which might be connected to the development of DM and MAFLD [90]. Downregulation of YTHDF2 could enhance the expression of SOCS2, which might negatively regulate the cell proliferation pathway of JAK/STAT signaling [91]. Therefore, YTHDF2 could suppress the phosphorylation of STAT5 for the inhibition of cell growth [92]. Interestingly, miR-223 that has been reported to be abnormally expressed in DM, which might regulate inflammation by targeting different targets [93]. In addition, YTHDF2 could enhance the protein expression of OCT4 through retaining the m6A methylation of the 5'-UTR of OCT4 mRNA [94]. Pancreatic β cell failure is a hallmark of diabetes, in which OCT4 signaling might be involved [95]. Consequently, YTHDF2 could function by regulating the expression of target genes to influence the development of various diseases including DM, obesity and MAFLD. YTHDF2 could also act as a target of miRNA and circRNA to participate in the progression of several diseses. However, the connotation between YTHDF2 and ncRNAs in DM needs further investigation.
6. Future perspectives
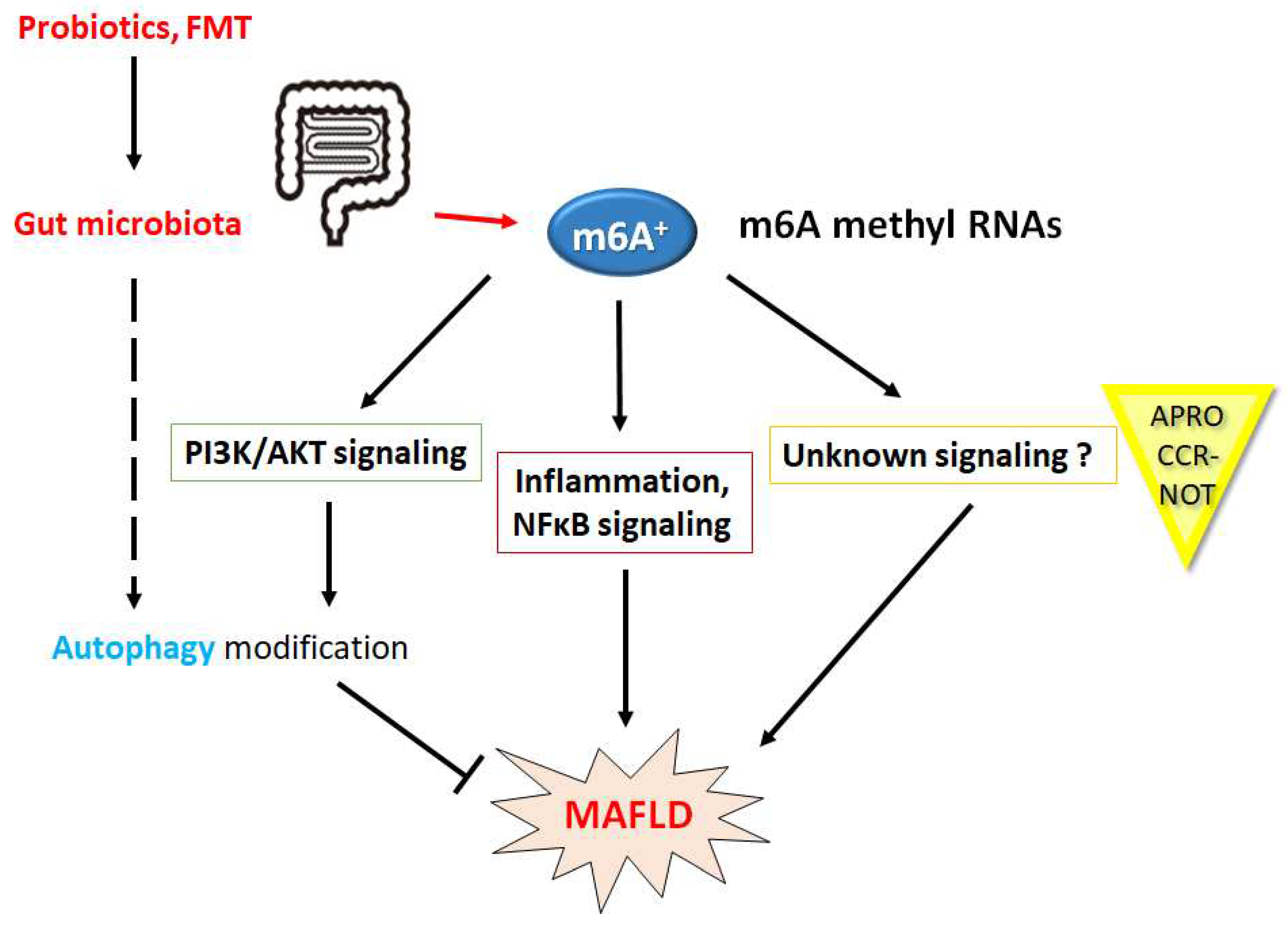
Microbiota collaborations could play critical roles in the development of MAFLD [96]. For example, Fusobacterium nucleatum in gut micribiota may induce a remarkable decline of m6A modifications by downregulation of an m6A methyltransferase METTL3 [97]. In addition, Fusobacterium nucleatum could reduce METTL3 transcription, which might promote the expression of a kinesin family member by reducing its m6A levels and diminishing YTHDF2-dependent mRNA degradation [97]. Non-coding RNAs including circRNAs have been suggested as potential good biomarkers in microbiota-associated diseases [98]. Interestingly, fecal microbiota transplantation (FMT) from Bifidobacterium into mice could significantly suppress the lung metastasis of cancer cells. (Figure 3) Mechanistically, the gut microbiota may impact circRNA expression to regulate levels of resultant miRNAs [99].
The gut microbiota has emerged as a key regulator in the pathogenesis of NAFLD for the development of specific therapies [100]. In presence of microbial dysbiosis, bacteria and their metabolic products may initiate inflammatory pathways which result in hepatocellular inflammation [101]. There might be a mutual relationship between microbiota and host, which could regulate gene expression mostly due to the production of metabolites that are generated after colonic fermentation of bioactive compounds such as short-chain fatty acids (SCFA) and/or polyphenols [102]. This concept might in turn play an important role in the prevention/treatment of various diseases. In fact, the gut microbiota with employing probiotics and/or FMT could modulate the host response to various therapeutics including novel immunotherapies [103]. (Figure 3) Gut microbiota dysbiosis could probably occur due to the use of broad-spectrum antibiotics, which may disturb the microenvironment of gut [104].
Regulators of m6A have not only verified as promising treatment concept but also have been advanced to step into clinical and actual therapies. The application of potent inhibitors or activators to regulate m6A organizers may disturb the development of obesity of DM beneficial for MAFLD patients. Furthermore, the m6A modification and circRNAs could also influence the pathogenesis with affecting the host immune system of patients. Innate immunity is a natural immune defense function during the development with a pivotal role in inflammatory responses [105]. The modification of the m6A gene could affect the role of circRNAs in innate immune regulation. In addition, exogenous circRNAs could precisely trigger an intracellular signaling pathway in immune cells and promote immune activation [106]. Interestingly, non-coding RNAs could exert an impact on the expression of regulator factors of m6A by indirectly modulating downstream targets or directly regulating particular factors of m6A. Modification immune cells would form a valuable feedback loop to mediate the modified beneficial prognosis. Further studies on the interaction of m6A and circRNAs/ncRNAs could uncover the comprehensive mechanism of MAFLD.
7. Conclusions
NcRNAs and m6A modification have been shown to play a critical role in the development and/or progression of various liver diseases including MAFLD with the support of miRNA/circRNA and m6A-reader molecules at the post-transcriptional level. Microbiota collaborations may play critical roles in the disease progression via the modification of m6A levels within various RNAs including ncRNA.
Author Contributions
Conceptualization, MN, NS, YI, and SM; original draft preparation and editing, MN, NS, YI, SY, and SM; visualization, MN, NS, YI and SM; supervision, SM. Each author (MN, NS, YI, SY, and SM) has participated sufficiently in this work of drafting the article and/or revising the article for the important rational content. Then, all authors gave final approval of the version to be submitted. Finally, all authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare that they have no competing financial interests.
Abbreviations
AGO2: Argonaute 2; ALKBH5: AlkB homolog 5; circRNAs: circular RNAs; eIF3: eukaryotic initiation factor 3; FMT: fecal microbiota transplantation; FTO: fat mass and obesity-associated; HNRNPs: heterogeneous nuclear ribonucleoproteins; HNRNPA2B1: heterogeneous nuclear ribonucleoprotein A2/B1; lncRNAs: long ncRNAs; m1A: N1-methyladenosine; m5C: 5-methylcytosine; m6A: N6-methyladenosine; METTL3: methyltransferase like 3; METTL14: methyltransferase like 14; miRNAs: microRNAs; mRNAs: messenger RNAs; mTOR: mammalian target of rapamycin; ncRNAs: non-coding RNAs’ ORF: open reading framework; PABP: poly(A) binding protein; Paip2: PABP-interacting protein 2; PD-1: programmed death 1; PVT1: plasmacytoma variant translocation 1; RBPs: RNA-binding proteins; RNase: ribonuclease; TME: tumor microenvironment; 3′ -UTR: 3′ -untranslated region; WTAP: Wilms’ tumor-associated protein; YTH: YT521-B homology
References
- Liu, H.; Hao, W.; Yang, J.; Zhang, Y.; Wang, X.; Zhang, C. Emerging roles and potential clinical applications of translatable circular RNAs in cancer and other human diseases. Genes Dis. 2023, 10, 1994–2012. [Google Scholar] [CrossRef] [PubMed]
- Das, A.; Gorospe, M.; Panda, A.C. The coding potential of circRNAs. Aging 2018, 10, 2228–2229. [Google Scholar] [CrossRef] [PubMed]
- Meng, J.; Chen, S.; Han, J.-X.; Qian, B.; Wang, X.-R.; Zhong, W.-L.; Qin, Y.; Zhang, H.; Gao, W.-F.; Lei, Y.-Y.; et al. Twist1 Regulates Vimentin through Cul2 Circular RNA to Promote EMT in Hepatocellular Carcinoma. Cancer Res 2018, 78, 4150–4162. [Google Scholar] [CrossRef] [PubMed]
- Memczak, S.; Jens, M.; Elefsinioti, A.; Torti, F.; Krueger, J.; Rybak, A.; Maier, L.; Mackowiak, S.D.; Gregersen, L.H.; Munschauer, M.; et al. Circular RNAs are a large class of animal RNAs with regulatory potency. Nature 2013, 495, 333–338. [Google Scholar] [CrossRef] [PubMed]
- Rybak-Wolf, A.; Stottmeister, C.; Glažar, P.; Jens, M.; Pino, N.; Giusti, S.; Hanan, M.; Behm, M.; Bartok, O.; Ashwal-Fluss, R.; et al. Circular RNAs in the Mammalian Brain Are Highly Abundant, Conserved, and Dynamically Expressed. Mol. Cell 2015, 58, 870–885. [Google Scholar] [CrossRef] [PubMed]
- Zeng, Q.; Liu, C.-H.; Wu, D.; Jiang, W.; Zhang, N.; Tang, H. LncRNA and circRNA in Patients with Non-Alcoholic Fatty Liver Disease: A Systematic Review. Biomolecules 2023, 13, 560. [Google Scholar] [CrossRef]
- Kopp, F.; Mendell, J.T. Functional Classification and Experimental Dissection of Long Noncoding RNAs. Cell 2018, 172, 393–407. [Google Scholar] [CrossRef]
- Zhang, Y.; Mao, Q.; Xia, Q.; Cheng, J.; Huang, Z.; Li, Y.; Chen, P.; Yang, J.; Fan, X.; Liang, Y.; et al. Noncoding RNAs link metabolic reprogramming to immune microenvironment in cancers. J. Hematol. Oncol. 2021, 14, 1–26. [Google Scholar] [CrossRef] [PubMed]
- Feng, H.; Yuan, X.; Wu, S.; Yuan, Y.; Cui, L.; Lin, D.; Peng, X.; Liu, X.; Wang, F. Effects of writers.; erasers and readers within miRNA-related m6A modification in cancers. Cell Prolif. 2023, 56, e13340.
- Xu, K.; Sun, Y.; Sheng, B.; Zheng, Y.; Wu, X.; Xu, K. Role of identified RNA N6-methyladenosine methylation in liver. Anal. Biochem. 2019, 578, 45–50. [Google Scholar] [CrossRef]
- Lee, Y.; Choe, J.; Park, OH.; Kim, YK. Molecular Mechanisms Driving mRNA Degradation by m6A Modification. Trends Genet. 2020, 36, 177–188. [Google Scholar] [CrossRef] [PubMed]
- Fu, Y.; Dominissini, D.; Rechavi, G.; He, C. Gene expression regulation mediated through reversible m6A RNA methylation. Nat. Rev. Genet. 2014, 15, 293–306. [Google Scholar] [CrossRef] [PubMed]
- Kholodenko, IV.; Yarygin, KN. Hepatic Macrophages as Targets for the MSC-Based Cell Therapy in Non-Alcoholic Steatohepatitis. Biomedicines. 2023, 11, 3056. [Google Scholar] [CrossRef] [PubMed]
- Cheng, C.; Yu, F.; Yuan, G.; Jia, J. Update on N6-methyladenosine methylation in obesity-related diseases. Obesity 2023, 32, 240–251. [Google Scholar] [CrossRef] [PubMed]
- Zhang, N.; Tian, X.; Yan, T.; Wang, H.; Zhang, D.; Lin, C.; Liu, Q.; Jiang, S. Insights into the role of nucleotide methylation in metabolic-associated fatty liver disease. Front. Immunol. 2023, 14, 1148722. [Google Scholar] [CrossRef] [PubMed]
- Hu, Y.; Feng, Y.; Zhang, L.; Jia, Y.; Cai, D.; Qian, S.-B.; Du, M.; Zhao, R. GR-mediated FTO transactivation induces lipid accumulation in hepatocytes via demethylation of m6A on lipogenic mRNAs. RNA Biol. 2020, 17, 930–942. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.; Tian, S.; Zheng, X.; Zhang, M.; Zhou, X.; Shang, Y.; Han, Y. N6-methyladenosine RNA methylation in liver diseases: from mechanism to treatment. J. Gastroenterol. 2023, 58, 718–733. [Google Scholar] [CrossRef] [PubMed]
- Meyer, KD.; Jaffrey, SR. Rethinking m6A Readers. ; Writers.; and Erasers. Annu Rev Cell Dev Biol. 2017, 33, 319–42. [Google Scholar] [CrossRef] [PubMed]
- Qin, S.; Zhang, Q.; Xu, Y.; Ma, S.; Wang, T.; Huang, Y.; Ju, S. m6A-modified circRNAs: detections, mechanisms, and prospects in cancers. Mol. Med. 2022, 28, 1–17. [Google Scholar] [CrossRef]
- Yang, Y.; Shen, F.; Huang, W.; Qin, S.; Huang, J.-T.; Sergi, C.; Yuan, B.-F.; Liu, S.-M. Glucose Is Involved in the Dynamic Regulation of m6A in Patients With Type 2 Diabetes. J. Clin. Endocrinol. Metab. 2018, 104, 665–673. [Google Scholar] [CrossRef]
- Li, Z.; Weng, H.; Su, R.; Weng, X.; Zuo, Z.; Li, C.; Huang, H.; Nachtergaele, S.; Dong, L.; Hu, C.; et al. FTO Plays an Oncogenic Role in Acute Myeloid Leukemia as a N 6 -Methyladenosine RNA Demethylase. Cancer Cell 2016, 31, 127–141. [Google Scholar] [CrossRef] [PubMed]
- Takemoto, S.; Nakano, M.; Fukami, T.; Nakajima, M. m6A modification impacts hepatic drug and lipid metabolism properties by regulating carboxylesterase 2. Biochem. Pharmacol. 2021, 193, 114766. [Google Scholar] [CrossRef] [PubMed]
- Bansal, H.; Yihua, Q.; Iyer, S.P.; Ganapathy, S.; Proia, D.; O Penalva, L.; Uren, P.J.; Suresh, U.; Carew, J.S.; Karnad, A.B.; et al. WTAP is a novel oncogenic protein in acute myeloid leukemia. Leukemia 2014, 28, 1171–1174. [Google Scholar] [CrossRef] [PubMed]
- Patop, IL.; Wüst, S.; Kadener, S. Past. ; present.; and future of circRNAs. EMBO J. 2019, 38, e100836. [Google Scholar] [PubMed]
- Lee, C.Q.E.; Kerouanton, B.; Chothani, S.; Zhang, S.; Chen, Y.; Mantri, C.K.; Hock, D.H.; Lim, R.; Nadkarni, R.; Huynh, V.T.V.T.; et al. Coding and non-coding roles of MOCCI (C15ORF48) coordinate to regulate host inflammation and immunity. Nat. Commun. 2021, 12, 1–22. [Google Scholar] [CrossRef] [PubMed]
- Jiang, X.; Liu, B.; Nie, Z.; Duan, L.; Xiong, Q.; Jin, Z.; Yang, C.; Chen, Y. The role of m6A modification in the biological functions and diseases. Signal Transduct. Target. Ther. 2021, 6, 1–16. [Google Scholar] [CrossRef]
- Shao, Y.; Liu, Z.; Song, X.; Sun, R.; Zhou, Y.; Zhang, D.; Sun, H.; Huang, J.; Wu, C.; Gu, W.; Zheng, X.; Jiang, J. ALKBH5/YTHDF2-mediated m6A modification of circAFF2 enhances radiosensitivity of colorectal cancer by inhibiting Cullin neddylation. Clin Transl Med. 2023, 13, e1318. [Google Scholar] [CrossRef] [PubMed]
- Yuan, W.; Zhang, X.; Cong, H. Advances in the protein-encoding functions of circular RNAs associated with cancer (Review). Oncol. Rep. 2023, 50, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Lei, M.; Zheng, G.; Ning, Q.; Zheng, J.; Dong, D. Translation and functional roles of circular RNAs in human cancer. Mol. Cancer 2020, 19, 1–9. [Google Scholar] [CrossRef]
- Wu, Y.; Zeng, Y.; Ren, Y.; Yu, J.; Zhang, Q.; Xiao, X. Insights into RNA N6-methyladenosine in Glucose and Lipid Metabolic Diseases and Their Therapeutic Strategies. Endocrinology 2023, 165. [Google Scholar] [CrossRef]
- Wang, W.; Yan, J.; Han, L.; Zou, Z.-L.; Xu, A.-L. Silencing METTL14 alleviates liver injury in non-alcoholic fatty liver disease by regulating mitochondrial homeostasis. Biomol. Biomed. 2024, 24, 505–519. [Google Scholar] [CrossRef] [PubMed]
- Farías, C.; Cisternas, C.; Gana, J.C.; Alberti, G.; Echeverría, F.; Videla, L.A.; Mercado, L.; Muñoz, Y.; Valenzuela, R. Dietary and Nutritional Interventions in Nonalcoholic Fatty Liver Disease in Pediatrics. Nutrients 2023, 15, 4829. [Google Scholar] [CrossRef] [PubMed]
- Gómez de Cedrón, M.; Moreno Palomares, R.; Ramírez de Molina, A. Metabolo-epigenetic interplay provides targeted nutritional interventions in chronic diseases and ageing. Front Oncol. 2023, 13, 1169168. [Google Scholar] [CrossRef] [PubMed]
- Farsetti, A.; Illi, B.; Gaetano, C. How epigenetics impacts on human diseases. Eur. J. Intern. Med. 2023, 114, 15–22. [Google Scholar] [CrossRef] [PubMed]
- Sun, L.; Chen, X.; Zhu, S.; Wang, J.; Diao, S.; Liu, J.; Xu, J.; Li, X.; Sun, Y.; Huang, C.; et al. Decoding m6A mRNA methylation by reader proteins in liver diseases. Genes Dis. 2024, 11, 711–726. [Google Scholar] [CrossRef]
- Moghadam, S.K.; Bakhshinejad, B.; Khalafizadeh, A.; Hussen, B.M.; Babashah, S. Non-coding RNA-associated competitive endogenous RNA regulatory networks: Novel diagnostic and therapeutic opportunities for hepatocellular carcinoma. J. Cell. Mol. Med. 2021, 26, 287–305. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Tan, QQ.; Tan, XR.; Li, SJ.; Zhang, XX. Circ_0057558 promotes nonalcoholic fatty liver disease by regulating ROCK1/AMPK signaling through targeting miR-206. Cell Death Dis. 2021, 12, 809. [Google Scholar] [CrossRef] [PubMed]
- Huang, A.; Zheng, H.; Wu, Z.; Chen, M.; Huang, Y. Circular RNA-protein interactions: functions, mechanisms, and identification. Theranostics 2020, 10, 3503–3517. [Google Scholar] [CrossRef] [PubMed]
- Larsson, O.; Perlman, D.M.; Fan, D.; Reilly, C.S.; Peterson, M.; Dahlgren, C.; Liang, Z.; Li, S.; Polunovsky, V.A.; Wahlestedt, C.; et al. Apoptosis resistance downstream of eIF4E: posttranscriptional activation of an anti-apoptotic transcript carrying a consensus hairpin structure. Nucleic Acids Res. 2006, 34, 4375–4386. [Google Scholar] [CrossRef]
- Karim, M.M.; Svitkin, Y.V.; Kahvejian, A.; De Crescenzo, G.; Costa-Mattioli, M.; Sonenberg, N. A mechanism of translational repression by competition of Paip2 with eIF4G for poly(A) binding protein (PABP) binding. Proc. Natl. Acad. Sci. 2006, 103, 9494–9499. [Google Scholar] [CrossRef]
- Yang, M.; Qi, M.; Xu, L.; Huang, P.; Wang, X.; Sun, J.; Shi, J.; Hu, Y. Differential host circRNA expression profiles in human lung epithelial cells infected with SARS-CoV-2. Infect Genet Evol. 2021, 93, 104923. [Google Scholar] [CrossRef] [PubMed]
- Ma, W.; Wang, X.; Sun, H.; Xu, B.; Song, R.; Tian, Y.; Zhao, L.; Xu, Y.; Zhao, Y.; Yang, F.; et al. Oxidant stress-sensitive circRNA Mdc1 controls cardiomyocyte chromosome stability and cell cycle re-entry during heart regeneration. Pharmacol. Res. 2022, 184, 106422. [Google Scholar] [CrossRef] [PubMed]
- Ikeda, Y.; Morikawa, S.; Nakashima, M.; Yoshikawa, S.; Taniguchi, K.; Sawamura, H.; Suga, N.; Tsuji, A.; Matsuda, S. CircRNAs and RNA-Binding Proteins Involved in the Pathogenesis of Cancers or Central Nervous System Disorders. Non-Coding RNA 2023, 9, 23. [Google Scholar] [CrossRef] [PubMed]
- Winkler, G.S. The mammalian anti-proliferative BTG/Tob protein family. J. Cell. Physiol. 2009, 222, 66–72. [Google Scholar] [CrossRef] [PubMed]
- Ezzeddine, N.; Chang, T.-C.; Zhu, W.; Yamashita, A.; Chen, C.-Y.A.; Zhong, Z.; Yamashita, Y.; Zheng, D.; Shyu, A.-B. Human TOB, an Antiproliferative Transcription Factor, Is a Poly(A)-Binding Protein-Dependent Positive Regulator of Cytoplasmic mRNA Deadenylation. Mol. Cell. Biol. 2007, 27, 7791–7801. [Google Scholar] [CrossRef] [PubMed]
- Tirone, F. The gene PC3TIS21/BTG2, prototype member of the PC3/BTG/TOB family: Regulator in control of cell growth, differentiation, and DNA repair? J. Cell. Physiol. 2001, 187, 155–165. [Google Scholar] [CrossRef] [PubMed]
- Doidge, R.; Mittal, S.; Aslam, A.; Winkler, G.S. The Anti-Proliferative Activity of BTG/TOB Proteins Is Mediated via the Caf1a (CNOT7) and Caf1b (CNOT8) Deadenylase Subunits of the Ccr4-Not Complex. PLOS ONE 2012, 7, e51331. [Google Scholar] [CrossRef]
- Lim, N.S.; Kozlov, G.; Chang, T.-C.; Groover, O.; Siddiqui, N.; Volpon, L.; De Crescenzo, G.; Shyu, A.-B.; Gehring, K. Comparative Peptide Binding Studies of the PABC Domains from the Ubiquitin-protein Isopeptide Ligase HYD and Poly(A)-binding Protein. J. Biol. Chem. 2006, 281, 14376–14382. [Google Scholar] [CrossRef] [PubMed]
- Horiuchi, M.; Takeuchi, K.; Noda, N.; Muroya, N.; Suzuki, T.; Nakamura, T.; Kawamura-Tsuzuku, J.; Takahasi, K.; Yamamoto, T.; Inagaki, F. Structural basis for the antiproliferative activity of the Tob-hCaf1 complex. J. Biol. Chem. 2009, 284, 13244–13255. [Google Scholar] [CrossRef]
- Ezzeddine, N.; Chen, C.-Y.A.; Shyu, A.-B. Evidence Providing New Insights into TOB-Promoted Deadenylation and Supporting a Link between TOB's Deadenylation-Enhancing and Antiproliferative Activities. Mol. Cell. Biol. 2012, 32, 1089–1098. [Google Scholar] [CrossRef]
- Stupfler, B.; Birck, C.; Séraphin, B.; Mauxion, F. BTG2 bridges PABPC1 RNA-binding domains and CAF1 deadenylase to control cell proliferation. Nat. Commun. 2016, 7, 10811. [Google Scholar] [CrossRef]
- Hwang, S.S.; Lim, J.; Yu, Z.; Kong, P.; Sefik, E.; Xu, H.; Harman, C.C.D.; Kim, L.K.; Lee, G.R.; Li, H.-B.; et al. mRNA destabilization by BTG1 and BTG2 maintains T cell quiescence. Science 2020, 367, 1255–1260. [Google Scholar] [CrossRef] [PubMed]
- Flamand, M.N.; Wu, E.; Vashisht, A.; Jannot, G.; Keiper, B.D.; Simard, M.J.; Wohlschlegel, J.; Duchaine, T.F. Poly(A)-binding proteins are required for microRNA-mediated silencing and to promote target deadenylation in C. elegans. Nucleic Acids Res. 2016, 44, 5924–5935. [Google Scholar] [CrossRef] [PubMed]
- Fabian, M.R.; Mathonnet, G.; Sundermeier, T.; Mathys, H.; Zipprich, J.T.; Svitkin, Y.V.; Rivas, F.; Jinek, M.; Wohlschlegel, J.; Doudna, J.A.; et al. Mammalian miRNA RISC recruits CAF1 and PABP to affect PABP-dependent deadenylation. Mol. Cell 2009, 35, 868–880. [Google Scholar] [CrossRef] [PubMed]
- Cheng, Q.; Li, Q.; Xu, L.; Jiang, H. Exosomal microRNA-301a-3p promotes the proliferation and invasion of nasopharyngeal carcinoma cells by targeting BTG1 mRNA. Mol. Med. Rep. 2021, 23, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Zhou, X.; Yuan, Y.; Teng, F.; Li, K.; Luo, S.; Zhang, P.; Liu, D.; Zhang, H.; Zhang, J. Obesity-induced upregulation of microRNA-183-5p promotes hepatic triglyceride accumulation by targeting the B-cell translocation gene 1. Life Sci. 2021, 268, 119011. [Google Scholar] [CrossRef]
- Sun, G.; Wang, Y.; Yang, L.; Zhang, Z.; Zhao, Y.; Shen, Z.; Han, X.; Du, X.; Jin, H.; Li, C.; et al. Rebalancing liver-infiltrating CCR3+ and CD206+ monocytes improves diet-induced NAFLD. Cell Rep. 2023, 42, 112753. [Google Scholar] [CrossRef] [PubMed]
- Imran, M.; and Lim, I.K. Regulation of Btg2(/TIS21/PC3) expression via reactive oxygen species-protein kinase C-NF kappa B pathway under stress conditions. Cell Signal. 2013, 25, 2400–2412. [Google Scholar] [CrossRef] [PubMed]
- Abdelgwad, M.; Zakaria, R.; Marzouk, S.; Sabry, D.; Ahmed, R.; Badary, H.A.; Samir, M. The Emerging Role of Circular RNA Homeodomain Interacting Protein Kinase 3 and Circular RNA 0046367 through Wnt/Beta-Catenin Pathway on the Pathogenesis of Nonalcoholic Steatohepatitis in Egyptian Patients. Rep. Biochem. Mol. Biol. 2023, 11, 614–625. [Google Scholar] [CrossRef]
- Fu, L.; Jiang, Z.; Li, T.; Hu, Y.; Guo, J. Circular RNAs in hepatocellular carcinoma: Functions and implications. Cancer Med. 2018, 7, 3101–3109. [Google Scholar] [CrossRef]
- Liu, Y.; Yang, Y.; Lin, Y.; Wei, B.; Hu, X.; Xu, L.; Zhang, W.; Lu, J. N6 -methyladenosine-modified circRNA RERE modulates osteoarthritis by regulating β-catenin ubiquitination and degradation. Cell Prolif. 2023, 56, e13297. [Google Scholar] [CrossRef]
- Rehman, S.; Varma, A.; Gupta, A.; Storey, K.B. The regulation of m6A-related proteins during whole-body freezing of the freeze-tolerant wood frog. Biochem. Cell Biol. 2023, 101, 77–86. [Google Scholar] [CrossRef]
- Qin, Y.; Li, B.; Arumugam, S.; Lu, Q.; Mankash, S.M.; Li, J.; Sun, B.; Li, J.; Flavell, R.A.; Li, H.-B.; et al. m6A mRNA methylation-directed myeloid cell activation controls progression of NAFLD and obesity. Cell Rep. 2021, 37, 109968–109968. [Google Scholar] [CrossRef] [PubMed]
- Du, H.; Zhao, Y.; He, J.; Zhang, Y.; Xi, H.; Liu, M.; Ma, J.; Wu, L. YTHDF2 destabilizes m6A-containing RNA through direct recruitment of the CCR4–NOT deadenylase complex. Nat. Commun. 2016, 7, 12626. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Sun, B.; Jiang, Q.; Wu, R.; Cai, M.; Yao, Y.; Liu, Q.; Shi, H.; Feng, J.; Wang, Y. mRNA m6A plays opposite role in regulating UCP2 and PNPLA2 protein expression in adipocytes. Int. J. Obes. 2018, 42, 1912–1924. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.; Wan, J.; Gao, X.; Zhang, X.; Jaffrey, SR.; Qian, SB. Dynamic m(6)A mRNA methylation directs translational control of heat shock response. Nature. 2015, 526, 591–594. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.; Li, Y.; Yu, J.; Gan, Z.; Wei, W.; Wang, C.; Zhang, L.; Wang, T.; Zhong, X. Resveratrol Attenuates High-Fat Diet Induced Hepatic Lipid Homeostasis Disorder and Decreases m6A RNA Methylation. Front. Pharmacol. 2020, 11. [Google Scholar] [CrossRef] [PubMed]
- Gebeyew, K.; Yang, C.; Mi, H.; Cheng, Y.; Zhang, T.; Hu, F.; Yan, Q.; He, Z.; Tang, S.; Tan, Z. Lipid metabolism and m6A RNA methylation are altered in lambs supplemented rumen-protected methionine and lysine in a low-protein diet. J. Anim. Sci. Biotechnol. 2022, 13, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Boo, SH.; Ha, H.; Lee, Y.; Shin, MK.; Lee, S.; Kim, YK. UPF1 promotes rapid degradation of m6A-containing RNAs. Cell Rep. 2022, 39, 110861. [Google Scholar] [CrossRef]
- Yu, H.; Yang, X.; Tang, J.; Si, S.; Zhou, Z.; Lu, J.; Han, J.; Yuan, B.; Wu, Q.; Lu, Q.; et al. ALKBH5 Inhibited Cell Proliferation and Sensitized Bladder Cancer Cells to Cisplatin by m6A-CK2α-Mediated Glycolysis. Mol. Ther. - Nucleic Acids 2020, 23, 27–41. [Google Scholar] [CrossRef]
- Bhattacharjee, J.; Borra, V.J.; Salem, E.S.B.; Zhang, C.; Murakami, K.; Gill, R.K.; Kim, A.; Kim, J.K.; Salazar-Gonzalez, R.-M.; Warren, M.; et al. Hepatic Ago2 Regulates PPARα for Oxidative Metabolism Linked to Glycemic Control in Obesity and Post Bariatric Surgery. Endocrinology 2021, 162. [Google Scholar] [CrossRef] [PubMed]
- Jin, D.; Guo, J.; Wu, Y.; Yang, L.; Wang, X.; Du, J.; Dai, J.; Chen, W.; Gong, K.; Miao, S.; et al. m6A demethylase ALKBH5 inhibits tumor growth and metastasis by reducing YTHDFs-mediated YAP expression and inhibiting miR-107/LATS2-mediated YAP activity in NSCLC. Mol Cancer. 2020, 19, 40. [Google Scholar] [CrossRef] [PubMed]
- Park, OH.; Ha, H.; Lee, Y.; Boo, SH.; Kwon, DH.; Song, HK.; Kim, YK. Endoribonucleolytic Cleavage of m6A-Containing RNAs by RNase P/MRP Complex. Mol Cell. 2019, 74, 494–507e8. [Google Scholar] [CrossRef] [PubMed]
- Deng, M.; Chen, B.; Liu, Z.; Cai, Y.; Wan, Y.; Zhang, G.; Fan, Y.; Zhang, Y.; Wang, F. YTHDF2 Regulates Maternal Transcriptome Degradation and Embryo Development in Goat. Front. Cell Dev. Biol. 2020, 8, 580367. [Google Scholar] [CrossRef] [PubMed]
- Khalifa, O.; Errafii, K.; Al-Akl, N.S.; Arredouani, A. Noncoding RNAs in Nonalcoholic Fatty Liver Disease: Potential Diagnosis and Prognosis Biomarkers. Dis. Markers 2020, 2020, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Pirola, C.J.; Sookoian, S. Epigenetics factors in nonalcoholic fatty liver disease. Expert Rev. Gastroenterol. Hepatol. 2020, 16, 521–536. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Song, J.; Liu, S.; Zhang, Y.; Qiu, T.; Jiang, L.; Bai, J.; Yao, X.; Wang, N.; Yang, G.; et al. m6A methylation-mediated PGC-1α contributes to ferroptosis via regulating GSTK1 in arsenic-induced hepatic insulin resistance. Sci. Total. Environ. 2023, 905, 167202. [Google Scholar] [CrossRef] [PubMed]
- Yang, K.; Sun, J.; Zhang, Z.; Xiao, M.; Ren, D.; Liu, S.-M. Reduction of mRNA m6A associates with glucose metabolism via YTHDC1 in human and mice. Diabetes Res. Clin. Pr. 2023, 198, 110607. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Dou, X.; Zheng, Z.; Ye, C.; Lu, TX.; Liang, HL.; Wang, L.; Weichselbaum, RR.; He, C. YTHDF2/m6 A/NF-κB axis controls anti-tumor immunity by regulating intratumoral Tregs. EMBO J. 2023, 42, e113126. [Google Scholar] [CrossRef]
- Yang, B.; Lu, L.; Zhou, D.; Fan, W.; Barbier-Torres, L.; Steggerda, J.; Yang, H.; Yang, X. Regulatory network and interplay of hepatokines, stellakines, myokines and adipokines in nonalcoholic fatty liver diseases and nonalcoholic steatohepatitis. Front. Endocrinol. 2022, 13, 1007944. [Google Scholar] [CrossRef]
- Ha, M.; Kim, VN. Regulation of microRNA biogenesis. Nat Rev Mol Cell Biol. 2014, 15, 509–524. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z.; Li, J.; Feng, G.; Gao, S.; Wang, Y.; Zhang, S.; Liu, Y.; Ye, L.; Li, Y.; Zhang, X. MicroRNA-145 Modulates N6-Methyladenosine Levels by Targeting the 3′-Untranslated mRNA Region of the N6-Methyladenosine Binding YTH Domain Family 2 Protein. J. Biol. Chem. 2017, 292, 3614–3623. [Google Scholar] [CrossRef]
- Li, H.; Zhang, N.; Jiao, X.; Wang, C.; Sun, W.; He, Y.; Ren, G.; Huang, S.; Li, M.; Chang, Y.; et al. Downregulation of microRNA-6125 promotes colorectal cancer growth through YTHDF2-dependent recognition of N6-methyladenosine-modified GSK3β. Clin Transl Med. 2021, 11, e602. [Google Scholar] [CrossRef] [PubMed]
- Saeki, C.; Matsuzaki, J.; Kuroda, M.; Fujita, K.; Ichikawa, M.; Takizawa, S.; Takano, K.; Oikawa, T.; Nakanuma, Y.; Saruta, M.; et al. Identification of circulating microRNAs as potential biomarkers for hepatic necroinflammation in patients with autoimmune hepatitis. BMJ Open Gastroenterol. 2022, 9, e000879. [Google Scholar] [CrossRef] [PubMed]
- Jensen, A.-S.H.; Ytting, H.; Winther-Sørensen, M.; Burisch, J.; Bergquist, A.; Gluud, L.L.; Albrechtsen, N.J.W. Autoimmune liver diseases and diabetes. Eur. J. Gastroenterol. Hepatol. 2023, 35, 938–947. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Zhao, X.; Wang, W.; Shi, H.; Pan, Q.; Lu, Z.; Perez, S.P.; Suganthan, R.; He, C.; Bjørås, M.; et al. Ythdf2-mediated m6A mRNA clearance modulates neural development in mice. Genome Biol. 2018, 19, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Burgdorf, K.S.; Grarup, N.; Justesen, J.M.; Harder, M.N.; Witte, D.R.; Jørgensen, T.; Sandbæk, A.; Lauritzen, T.; Madsbad, S.; Hansen, T.; et al. Studies of the Association of Arg72Pro of Tumor Suppressor Protein p53 with Type 2 Diabetes in a Combined Analysis of 55,521 Europeans. PLOS ONE 2011, 6, e15813. [Google Scholar] [CrossRef] [PubMed]
- Kung, C.-P.; Leu, J.I.-J.; Basu, S.; Khaku, S.; Anokye-Danso, F.; Liu, Q.; George, D.L.; Ahima, R.S.; Murphy, M.E. The P72R Polymorphism of p53 Predisposes to Obesity and Metabolic Dysfunction. Cell Rep. 2016, 14, 2413–2425. [Google Scholar] [CrossRef]
- Paramasivam, A.; Priyadharsini, J.V. The emerging role of m6A modification in autophagy regulation and its implications in human disease. Epigenomics 2022, 14, 565–568. [Google Scholar] [CrossRef]
- Zhuang, S.; Zhou, X.; Yang, X.; Chang, D.; Chen, T.; Sun, Y.; Wang, C.; Zhang, C.; Jiang, J.; Chen, Y.; et al. Dendrobium mixture ameliorates hepatic injury induced by insulin resistance in vitro and in vivo through the downregulation of AGE/RAGE/Akt signaling pathway. Heliyon 2023, 9, e22007. [Google Scholar] [CrossRef]
- Chen, M.; Wei, L.; Law, C.; Tsang, F.H.; Shen, J.; Cheng, C.L.; Tsang, L.; Ho, D.W.; Chiu, D.K.; Lee, J.M.; et al. RNA N6-methyladenosine methyltransferase-like 3 promotes liver cancer progression through YTHDF2-dependent posttranscriptional silencing of SOCS2. J. Hepatol. 2017, 67, 2254–2270. [Google Scholar] [CrossRef]
- de Bock, C.E.; Demeyer, S.; Degryse, S.; Verbeke, D.; Sweron, B.; Gielen, O.; Vandepoel, R.; Vicente, C.; Bempt, M.V.; Dagklis, A.; et al. HOXA9 Cooperates with Activated JAK/STAT Signaling to Drive Leukemia Development. Cancer Discov. 2018, 8, 616–631. [Google Scholar] [CrossRef] [PubMed]
- Aziz, F. The emerging role of miR-223 as novel potential diagnostic and therapeutic target for inflammatory disorders. Cell. Immunol. 2016, 303, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Huang, S.; Zhuang, H.; Ruan, S.; Zhou, Z.; Huang, K.; Ji, F.; Ma, Z.; Hou, B.; He, X. YTHDF2 promotes the liver cancer stem cell phenotype and cancer metastasis by regulating OCT4 expression via m6A RNA methylation. Oncogene 2020, 39, 4507–4518. [Google Scholar] [CrossRef]
- Liu, F.; Cai, Z.; Yang, Y.; Plasko, G.; Zhao, P.; Wu, X.; Tang, C.; Li, D.; Li, T.; Hu, S.; et al. The adipocyte-enriched secretory protein tetranectin exacerbates type 2 diabetes by inhibiting insulin secretion from β cells. Sci Adv. 2022, 8, eabq1799. [Google Scholar] [CrossRef] [PubMed]
- Morishita, A.; Oura, K.; Tadokoro, T.; Fujita, K.; Tani, J.; Kobara, H.; Ono, M.; Himoto, T.; Masaki, T. MicroRNAs and Nonalcoholic Steatohepatitis: A Review. Int. J. Mol. Sci. 2023, 24, 14482. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Zhang, L.; Li, M.; Zhang, Y.; Sun, M.; Wang, L.; Lin, J.; Cui, Y.; Chen, Q.; Jin, C.; et al. Fusobacterium nucleatum reduces METTL3-mediated m6A modification and contributes to colorectal cancer metastasis. Nat. Commun. 2022, 13, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Fardi, F.; Khasraghi, L.B.; Shahbakhti, N.; Naseriyan, A.S.; Najafi, S.; Sanaaee, S.; Alipourfard, I.; Zamany, M.; Karamipour, S.; Jahani, M.; et al. An interplay between non-coding RNAs and gut microbiota in human health. Diabetes Res. Clin. Pr. 2023, 201, 110739. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Z.; Huang, J.; Li, X.; Xing, J.; Chen, Q.; Liu, R.; Hua, F.; Qiu, Z.; Song, Y.; Bai, C.; et al. Gut microbiota regulate tumor metastasis via circRNA/miRNA networks. Gut Microbes 2020, 12. [Google Scholar] [CrossRef]
- Marra, F.; Svegliati-Baroni, G. Lipotoxicity and the gut-liver axis in NASH pathogenesis. J. Hepatol. 2018, 68, 280–295. [Google Scholar] [CrossRef]
- Makri, E.; Goulas, A.; Polyzos, S.A. Epidemiology, Pathogenesis, Diagnosis and Emerging Treatment of Nonalcoholic Fatty Liver Disease. Arch. Med Res. 2020, 52, 25–37. [Google Scholar] [CrossRef] [PubMed]
- Gadecka, A.; Bielak-Zmijewska, A. Slowing Down Ageing: The Role of Nutrients and Microbiota in Modulation of the Epigenome. Nutrients 2019, 11, 1251. [Google Scholar] [CrossRef] [PubMed]
- Pitt, J.M.; Vétizou, M.; Daillère, R.; Roberti, M.P.; Yamazaki, T.; Routy, B.; Lepage, P.; Boneca, I.G.; Chamaillard, M.; Kroemer, G.; et al. Resistance Mechanisms to Immune-Checkpoint Blockade in Cancer: Tumor-Intrinsic and -Extrinsic Factors. Immunity 2016, 44, 1255–1269. [Google Scholar] [CrossRef]
- Blaser, M.J. Antibiotic use and its consequences for the normal microbiome. Science 2016, 352, 544–545. [Google Scholar] [CrossRef] [PubMed]
- Vesely, M.D.; Kershaw, M.H.; Schreiber, R.D.; Smyth, M.J. Natural Innate and Adaptive Immunity to Cancer. Annu. Rev. Immunol. 2011, 29, 235–271. [Google Scholar] [CrossRef]
- Chen, Y.G.; Kim, M.V.; Chen, X.; Batista, P.J.; Aoyama, S.; Wilusz, J.E.; Iwasaki, A.; Chang, H.Y. Sensing Self and Foreign Circular RNAs by Intron Identity. Mol. Cell 2017, 67, 228–238. [Google Scholar] [CrossRef]
Figure 1.
Schematic diagram for the association of noncoding RNAs and m6A methyl RNAs to biological function and liver diseases. In the beginning, the m6A modification may be controlled by methyltransferases “writers” and demethylases “erasers”. The m6A-RNAs binding proteins including YTHDF2 are called “readers”, which have been suggested in several RNAs activities including translation as well as sponging. Consequently, m6A methylation in noncoding RNAs could be an important tool for diagnostics and/or therapeutics in several diseases including MAFLD.
Figure 1.
Schematic diagram for the association of noncoding RNAs and m6A methyl RNAs to biological function and liver diseases. In the beginning, the m6A modification may be controlled by methyltransferases “writers” and demethylases “erasers”. The m6A-RNAs binding proteins including YTHDF2 are called “readers”, which have been suggested in several RNAs activities including translation as well as sponging. Consequently, m6A methylation in noncoding RNAs could be an important tool for diagnostics and/or therapeutics in several diseases including MAFLD.
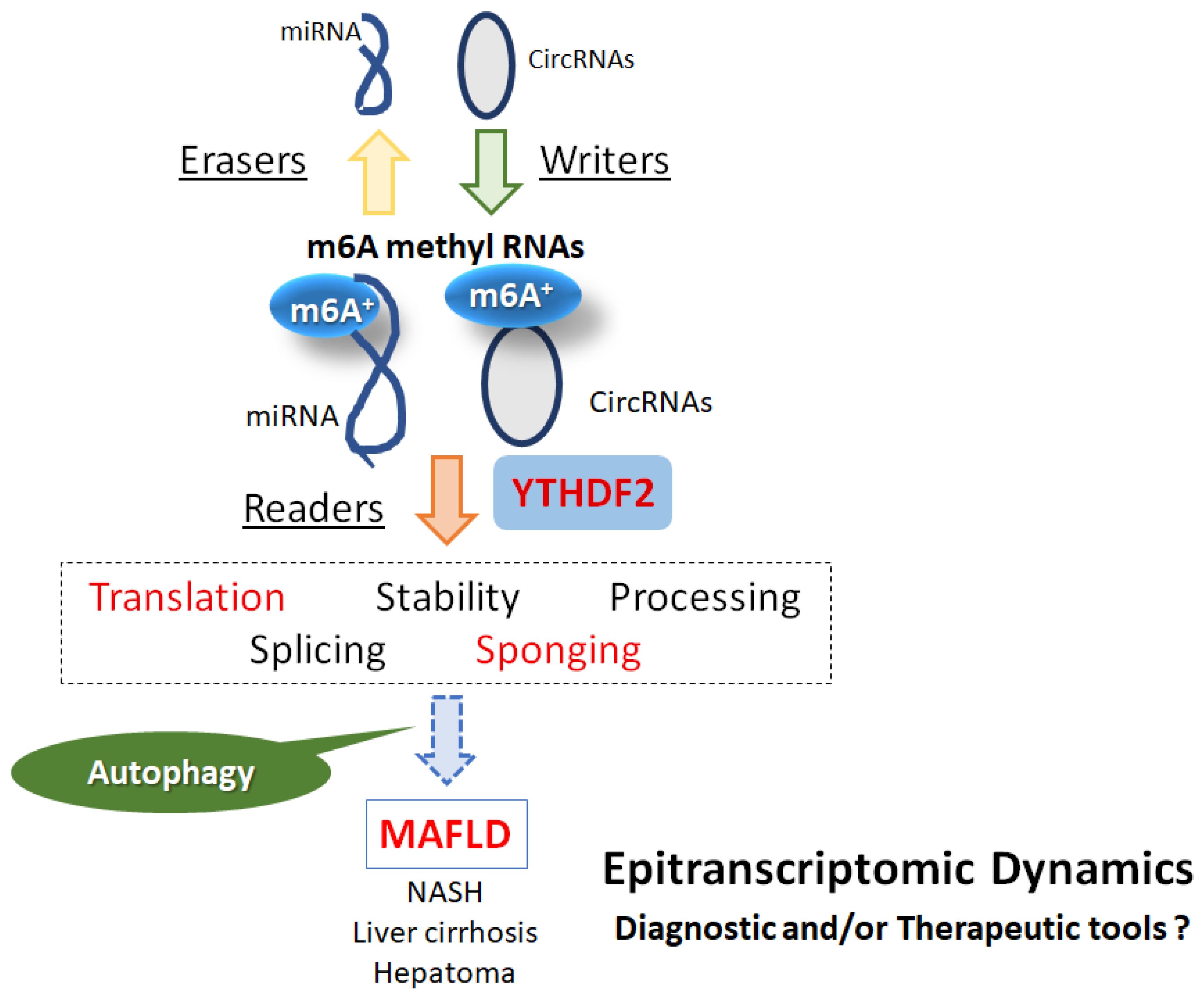
Figure 2.
Image of miRNAs, circRNAs and m6A-mediated inhibition of mRNA translation. RNA-induced silencing complex (miRISC) with certain miRNA could assist the deadenylation and/or mRNA degradation process by CAF1/CCR4/NOT1 molecules with PABP and APRO protein complex, in which GW182 and AGO2 might further assemble the miRISK complex. Subsequently, the miRNAs, circRNAs and m6A with YTHDF2 might play a dynamic role in the regulation of post-transcriptional gene expression via the regulation of mRNA translation and degradation of mRNAs. In attendance, APRO family molecules may interact with PABP to recruit the CAF1/CCR4/NOT1 complex. Note that some critical pathways have been misplaced for clarity. Abbreviations: ORF, open reading frame; “?” means for author speculation.
Figure 2.
Image of miRNAs, circRNAs and m6A-mediated inhibition of mRNA translation. RNA-induced silencing complex (miRISC) with certain miRNA could assist the deadenylation and/or mRNA degradation process by CAF1/CCR4/NOT1 molecules with PABP and APRO protein complex, in which GW182 and AGO2 might further assemble the miRISK complex. Subsequently, the miRNAs, circRNAs and m6A with YTHDF2 might play a dynamic role in the regulation of post-transcriptional gene expression via the regulation of mRNA translation and degradation of mRNAs. In attendance, APRO family molecules may interact with PABP to recruit the CAF1/CCR4/NOT1 complex. Note that some critical pathways have been misplaced for clarity. Abbreviations: ORF, open reading frame; “?” means for author speculation.
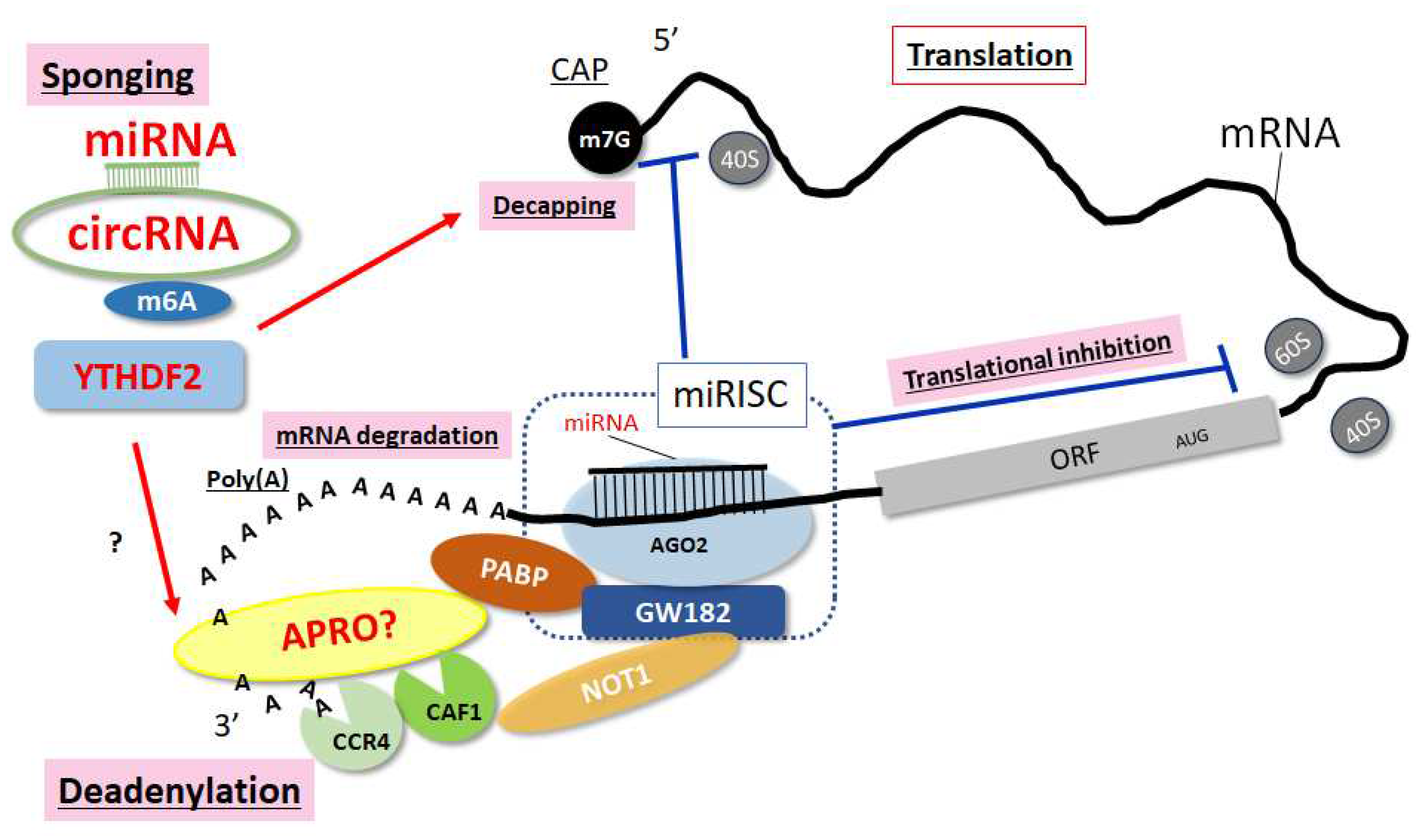
Figure 3.
Schematic demonstration of the potential strategies against the pathogenesis of various diseases including MAFLD. Some kinds of probiotics and/or fecal microbiota transplantation (FMT) might assist the alteration of gut microbial population for the modification of m6A levels and/or ncRNAs, which might be advantageous for the treatment of several diseases including MAFLD. Note that some of important activities such as autophagy initiation, inflammatory reaction, and ROS production have been misplaced for clarity.
Figure 3.
Schematic demonstration of the potential strategies against the pathogenesis of various diseases including MAFLD. Some kinds of probiotics and/or fecal microbiota transplantation (FMT) might assist the alteration of gut microbial population for the modification of m6A levels and/or ncRNAs, which might be advantageous for the treatment of several diseases including MAFLD. Note that some of important activities such as autophagy initiation, inflammatory reaction, and ROS production have been misplaced for clarity.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



