
Submitted:
11 December 2023
Posted:
12 December 2023
You are already at the latest version
Abstract
The fungal order Entomophthorales in the Zoopagomycotina includes many fungal pathogens of arthropods. This review explores five genera in the Erynioideae subfamily, namely Erynia, Furia, Pandora, Strongwellsea, and Zoophthora. This is the largest subfamily in the Entomophthorales including 125 described species. The species diversity, global distribution, and host range of this subfamily are summarized, revealing that relatively few taxa are geographically widespread, and few have broad host ranges, contrasting with a large number of species with single reports, from one location and one host species. The insect orders infected by the greatest numbers of species are the Diptera and Hemiptera. Across the subfamily relatively few species have been cultivated in vitro and those that have require more specialized media than any fungi. Given their potential to attack arthropods and in the fungal evolutionary tree, which species might be adopted for biological control purposes or biotechnological innovations are highlighted. Current challenges in the implementation of these species in biotechnology include the limited ability or difficulty in culturing many in vitro, a correlated paucity in genomic resources, and considerations regarding the host ranges of different species.
Keywords:
insect biocontrol
; cultivability
; genomics
; entomopathogens
; geographic distribution
; host range
1. Introduction
The fungal order Entomophthorales in the Zoopagomycotina includes at least 246 species of arthropod pathogens [1], many of which are well known for the ability to cause epizootics and change the behavior of infected hosts [2]. Their role in biocenoses is extremely important because they can function as regulators of arthropod populations and thus play a role in ecosystem homeostasis. Within the Entomophthorales, the largest family is the Entomophthoraceae, which consists exclusively of arthropod pathogens. This family was divided into two subfamilies in 2005, the Entomophthoroideae and the Erynioideae [3]. The Erynioideae is the larger of these two subfamilies, containing five genera, namely Erynia, Furia, Pandora, Strongwellsea, and Zoophthora. Fungi in these genera use their ballistic conidia to infect a wide range of arthropod species inhabiting agricultural and forest ecosystems, including insect hosts that are recognized as pests of various important crops worldwide [4,5]. In addition to attacking arthropod pests directly damaging crops and forests, directly some of the arthropods hosts of species in this subfamily include vectors of numerous diseases that impact humans, livestock, and crops.
Due to the observed capability of the Entomophthorales to cause massive mortality of insect hosts, questions about their potential use for various biotechnological applications are often raised. However, with the exception of the experimental release of Zoophthora radicans in Australia in the late 1970s [6] no other species of Entomophthorales have been developed thus far for field testing as fungal biological control agents. Species within this group differ from one another in numerous ways that impact their potential development as biopesticides [7]. One of the most important attributes to consider in this regard is the cultivability of these fungal species in vitro, which plays a critical factor in their propagation for potential application as biological control agents. This feature can be expressed to various degrees: from total inability to isolate fungal strains in vitro to routine transfers of the isolates and preservation of their cultures in culture collections, to research on improving in vitro growth toward potential mass production. Additionally, for successful biological control, the host range of the pathogens must be known, and is crucial in both identifying suitable fungi for specific target pests as well as in avoiding potential impacts on non-target arthropods. Furthermore, the natural habitats of these fungi and their geographical distributions are important toward consideration of development for biological control as regions around the world differ in regulation of species to be used for pest control that are native vs. non-native [8]. In addition, knowledge of the habitats where these fungi are naturally active will provide information about their ecological adaptability and long-term survival under diverse conditions.
We have analyzed these vital characteristics for species of the Erynioideae, and we aim to pinpoint which species demonstrate the most potential for future use in biological control. Moreover, we are keen on identifying the specific insect groups for which the application of these species as biological control agents might be most successful. We predict that these findings can be used as bases for screening and selection in future biocontrol research programs with Entomophthorales, paving the way for more effective and sustainable pest management solutions with species in the Erynioideae.
The primary objective of this research was to characterize several critical aspects of the lifestyles of the species within the five genera in the Erynioideae, here referred by the acronym EFPSZ. An overall consideration of these aspects of species in this group has not previously been undertaken. We predict that the assembly of this new information could be important toward potential applications of this group in various aspects of biotechnology, particularly in the development of biological control agents for pest management.
2. Materials and Methods
2.1. Literature analysis
We sought all literature related to the five genera in the Erynioideae through the use of the Web of Science, Scopus, and Google Scholar. We also examined the information associated with the species and strains deposited in the US Department of Agriculture Agricultural Research Service Collection of Entomopathogenic Fungal Cultures (ARSEF), the American Type Culture Collection (ATCC, USA) and CBS-Westerdijk Institute KNAW Fungal Biodiversity Centre, also known as Central Bureau of Fungal Cultures (Netherlands). The traits that were investigated are geographical distribution, host range, type of habitat, and documented ability to grow in vitro.
2.2. Distribution map
A map of the number of recorded species was created using StepMap GmbH software (Berlin, Germany). Countries and regions were colored according to the number of recorded species from light (less than five species) to dark pink (over 50 species described). Green indicates none have been reported. We consider the species: 1) local if the distribution range covers only one continent, 2) cosmopolitan or broadly distributed with records on at least two continents, and 3) ubiquitous if distribution records cover three or more continents.
2.3. Phylogenetic tree
To generate a dataset of EFPSZ taxa, we downloaded 18S and 28S sequences of identified species with accurate nomenclature from GenBank. All sequences were initially aligned, their alignments were manually adjusted, and ambiguous regions were excluded from the alignments using Mesquite 3.04 version [9]. Phylogenetic relationships were determined by the Neighbor Joining (NJ) algorithm, and the tree was visualized in PAUP* 4.0 [10].
3. Results
3.1. Geographic distribution
Species in the EFPSZ group have been recorded from all continents except Antarctica. However, the number of described species differs significantly between countries and regions (Figure 1). Many records are from several Central European countries (especially Switzerland, Poland), and the United Kingdom. Many species have also been found in North America (USA and Canada), other European countries, and China. Only a handful of records have been made in South America and across a considerable area of Asia and Oceania. The continent least documented for these fungi is Africa, where EFPSZ fungi have been reported only from four countries. They are also sparsely reported in countries in South America other than Argentina, Brazil, and Chile and in several countries in the Middle East. However, the map in Figure 1 most likely does not represent the actual species distributions, but rather the situation regarding our knowledge of this fungal group in particular countries where more sampling has occurred. It is obvious that climatic conditions in many countries of Africa or South America might be very favorable for species in the Erynioideae, but little research has yet been undertaken to describe Erynioideae and their host ranges from these regions.
There were clear differences in pattern species distribution among the five genera. Each genus contains both ubiquitous and cosmopolitan species as well as local ones, and they are not grouped in any specific way on the phylogenetic tree (Figure 2). It is very possible that many of the species of these genera that we classify as local have much broader distributions but have not been sampled broadly. We consider the broad distributions of many species as advantageous for future biocontrol agents since this feature might indicate a significant level of adaptability and ability to survive the environmental conditions in different climatic environments due to their ecology, pathogenesis and specialization [11].
Another perspective on geographical distribution can be obtained from analyzing culture collection deposits. The largest insect pathogen collection of the world is the USDA/ARS entomopathogenic fungi collection (ARSEF), which includes almost 15,000 occurrence records [12]. Although ARSEF has deposits from all over the world, most samples are from the USA, and then from Europe. We hypothesize that isolates from the rest of the world are less-represented due to lack of sampling. Despite the scarcity of well recorded data, at least 53 species out of the 125 valid EFPSZ species might be considered as cosmopolitan and at least 25 as ubiquitous (Figure 2, Table 1). Many of these might become ubiquitous as a result of worldwide spread of human agricultural activities, which spreads many crops worldwide along with their pests. One of the best examples of human mediated distribution might be P. gloeospora, found on several continents in mushroom growing farms [13].
3.2. Host specificity
EFPSZ fungi show a range of host-specificity. One third of EFPSZ parasitize two or more insect families. The species Erynia conica, E. rhizospora, E. selpulchralis, P. batallata, P. blunckii, P. echinospora, P. nourii, Z. aphidis, Z. canadensis are pathogenic to representatives of at least two families. An absolute generalist is Z. radicans, which infects insects in seven orders and 21 families (Table 1). However, two thirds of EFPSZ fungal species show some host-specificity and are able to infect only a narrower range of insects, usually attacking members of the same genus or family.
The flies (Order Diptera) are the most frequent hosts for EFPSZ fungi as more than one third of these fungal species were found killing Diptera. Nearly 25 percent of EFPSZ fungi infect insects in the order Hemiptera (30 pathogenic species), and nearly half that number were found infecting Coleoptera (16 pathogens) and Lepidoptera (15 pathogens). Within the Diptera, families most attacked by EFPSZ fungi are Calliphoridae (8 pathogen species), Tipulidae (7), Muscidae, Psychodidae and Sciaridae (6 each), and Chironomidae (5). In the genus Strongwellsea, species specialize exclusively on four dipteran families: Anthomyiidae, Muscidae, Sarcophagidae, and Scatophagidae. Among Hemiptera, the families Aphididae (14), Miridae (7), and Cicadellidae (6) are most infected by EFPSZ fungi. All other insect families have less than 5 pathogenic EFPSZ species infecting them (Figure 3).
3.3. Biological and ecological characteristics of EFPSZ fungi as biocenose components
The majority of species in the Erynioideae primarily infect insects in natural and agricultural environments. These habitats include aquatic biocenoses, forests and natural areas, and agrocenoses. It might be more precise to discuss the distribution of insect hosts, even if fungal infections can lead to infected insects relocating from their typical habitats [85]. Many EFPSZ species infect the imago (adult stage) of hosts. Among holometabolous hosts, species in the genus Strongwellsea only infect adults while some species from the other genera infect larvae. No EFPSZ species have been found attacking insect eggs.
These different host life cycle stages may occur in various ecosystems, so this factor should also be considered. In most cases, infected insects are found and collected on plant parts, partly due to the so-called climbing effect caused by any EFPSZ fungi [166]. This altered behavior may also be attributed to better visibility for researchers compared to the soil surface beneath vegetation. A comprehensive analysis of the distribution of entomophthoralean fungi in European biocenoses, focusing on forests and agrocenoses, was carried out by Bałazy [15]. Bałazy’s analysis emphasizes the importance of insect mobility, particularly because many insect hosts have wings and can migrate to neighboring ecosystems, spreading infection. This mobility is supported by collection of dispersing aphids infected with P. neoaphidis [167] and isolation of P. delphacis from planthoppers caught on a weather ship off the coast of Japan [75].
Most EFPSZ fungi are prevalent in aboveground ecosystems. The cadavers of insects infected with EFPSZ are often found on wild and cultured plants in various ecosystems. The presence of representatives of the genus Zoophthora in particular, is well-documented in numerous agricultural crops, orchards, and different types of forests. Species like P. dipterigena, P. phillonthii, Z. anglica, Z. miridis, Z. opomyzae, Z. petchii, Z. phytonomi, and Z. radicans are commonly observed in annual and perennial crops, meadows, pastures, orchards, and forests. These species are well-adapted to drier habitats. Species of the genera Strongwellsea and Zoophthora appear to be the most adaptable to a wide range of habitats, whether natural or human-created. Furthermore, aphid pathogenic species like P. neoaphidis and P. nouryi, have been found worldwide in many crops and are commonly observed at different temperatures and humidities.
All Entomophthorales require high humidity to release and disperse their conidia [168]. Interestingly, half of the species in the genus Erynia were found in aquatic or notably moist areas, e.g. E. aquatica, E. conica, E. curvispora, E. nematoceris, E. ovispora, E. sepulchralis, and E. variabilis. These species may serve as efficient biological control agents for insects requiring aquatic habitats during specific life stages due to their higher humidity needs compared to other species in this group. Additionally, five species in the genus Pandora, one species in Furia, and one species in Zoophthora were found in moist habitats. However, no Strongwellsea species were recorded in explicitly aquatic or moist environments (Table 1).
Soil is an unusual habitat for predominantly insect pathogenic EFPSZ. Nevertheless, at least one species, Pandora nouryi, infects root aphids (Pemphigus) and follows its hosts to this habitat, becoming a soil dweller [77]. Zoophthora myrmecophaga infects ants that move along their paths on the soil surface. Pandora brahminae, which infects scarabs inhabiting soil surface, also might be considered soil inhabitants.
3.4. Cultivability
Few species in the EFPSZ group have been isolated into pure culture, or even had their cultivability tested. Most species have been described only from insect cadavers and there are no cultures preserved. The largest culture collection of entomopathogenic fungi in the world is the USDA Agricultural Research Service Collection of Entomopathogenic Fungal Cultures (ARSEF, Ithaca, NY, USA), which contains over 14,000 fungal strains, isolated from infected insects or cadavers of insects dying from fungal infections. While most strains belong to the Ascomycota, entomophthoralean fungi are also well represented. This collection preserves 683 total isolates of EFPSZ [12]. These include 28 species known in the genera Erynia, Furia, Pandora, Strongwellsea and Zoophthora, as well as 36 isolates from these genera, which are not yet identified to the species level. Most species are represented by single or just a few isolates. However, there are over a hundred isolates representing species such as P. neoaphidis and Z. radicans, which reflects the easy cultivability of those species.
Few EFPSZ fungi can be cultivated on typical fungal nutritional media such as malt extract or potato dextrose agar or in the corresponding liquid media [168]. In the past, to ensure fungal growth of entomopathogens special media containing animal protein from additives such as liver, extracts of fresh or dried insects, blood serum, or egg yolk were used [21,169], providing the pathogens with specific nutrients absent in the usual laboratory media. Sometimes such rare and exotic media components as fly fat-bodies are used to stimulate spore germination or hyphal growth. Addition of yeast extract, arginine or other aminoacids to the medium substantially improves the growth of entomophthoralean fungi. Nowadays the most commonly used liquid medium for entomophthoralean growth is Grace′s Insect Medium (Sigma-Aldrich, St. Louis, USA), often with additives such as fetal bovine serum (ThermoFisher Scientific, Waltham, USA).
Larger scale production using simpler media has been developed for a few species. A method for producing Z. radicans dry-formulated mycelium has been developed with sporulation equivalent to cadavers. Recent examples of successful production on a large laboratory scale of the fungus P. cacopsyllae are the studies by Muskat et al. ([170,171], in press). This fungus that infects psyllids of the genus Cacopsylla has been fermented, encapsulated, and tested for above ground application.
The most fastidious EFPSZ species can so far only be cultured in vivo. They require the maintenance of an insect colony either in the laboratory or in the natural environment in order to maintain their population and complete their life cycle. The best results are obtained from using the natural hosts of the pathogenic fungus. In vivo production is the most difficult and labor intensive of methods for growing entomopathogenic fungi.
The ability to grow EFPSZ in culture is often connected with the ability of these fungi to infect insects at various stages of their life cycle, or at least stages other than the imago (adult), as seen in species like E. curvispora, Z. bialovienzenzis, Z. lanceolata, Z. phytonomi, and Z. radicans [15,21]. One of the remarkable characteristics of EFPSZ fungi, particularly those with high-host or life-stage specificity is the loss of their vigor and viability after several transfers on laboratory media despite strong initial growth [168,172]. These features of highly host-specific members of the EFPSZ can pose a challenge to mass production in biotechnology.
3.5. Genomics and Biotechnology
In addition to using these species directly for biological control, other potential biotechnology applications based on genes or proteins derived from these fungi are yet to be explored. The early diverging lineages of fungi, also defined as the paraphyletic group not including the Dikarya (e.g. Ascomycota and Basidiomycota), have emerged through the characterization of their genomes and identification of numerous shared genes and traits common to animals that were lost in more derived members of the Dikarya. Such homologs may in the future provide new insights into fundamental biology or even lead to therapeutics for human health. Fungi themselves are also known to produce diverse secondary metabolites/natural products, some of which are used commercially. Genome sequences have revealed evidence of greater biosynthetic capability for such molecules, even among some lineages of early diverging fungi [173]
At present, just a single species has had its genome sequenced from the Erynioideae, i.e. Z. radicans [174], due to the ease of its cultivability. The genome was generated as part of a large collection of species in a study that addressed the ploidy levels of the early fungal lineages, and so features about what this genome contains have not been described yet in detail. This genome sequence carries the information for how Z. radicans completes its lifecycle, including as an entomopathogen, but the mechanism is not immediately obvious. The Z. radicans genome is large, with the assembly at over 650 Mb currently divided over nearly 7,000 scaffolds and estimated to encode over 14,000 genes. The large size is due to the large amount of repetitive DNA within the genome (Figure 4). The entomopathogens in the Hypocreales (Ascomycota) contain numerous gene clusters for the synthesis of secondary metabolites, one of the best known being the cluster for the immunosuppressive cyclosporin from Tolypocladium inflatum [175], as well as other types of toxins (e.g. enterotoxins) with possible roles in altering host behavior [176]. However, such detailed information based on the Z. radicans genome is not yet available.
As pointed out above, one challenge toward generating more genome sequences is being able to obtain sufficient DNA, usually through culturing of isolates in vitro, which has not been possible for many of these species. Difficulties with culturing EFPSZ fungi make sequencing their genomes complicated because of the challenge of isolating high quality and high molecular weight DNA and RNA for sequencing. For many species, extraction of total DNA from the host insect cadaver might be the only option. However, the recent advances in single cell genomics [177], may provide a way in the near future to generate more genomic information about the genetic composition and potential virulence factors of EFPSZ species. Once identified, these genetic components could be utilized in vector-based expression systems for application as biopesticides. There are also a few genomes available for the closely related subfamily – Entomophthoroideae, including Entomophthora muscae, Entomophaga maimaga, Massospora cicadina as well as other species in Entomophthorales Conidiobolus coronatus Neoconidiobolus throboides and Basidiobolus meristosporus [178]. Summarizing all genome features for these Entomophthorales genomes, it can be predicted that the genomes of most EFPSZ fungi are much larger compared to the average ascomycete fungal genome size (40-60 kB) and can reach 600,000–1,000,000 kB in size and consist of many duplicated gene copies and repeated regions [179].
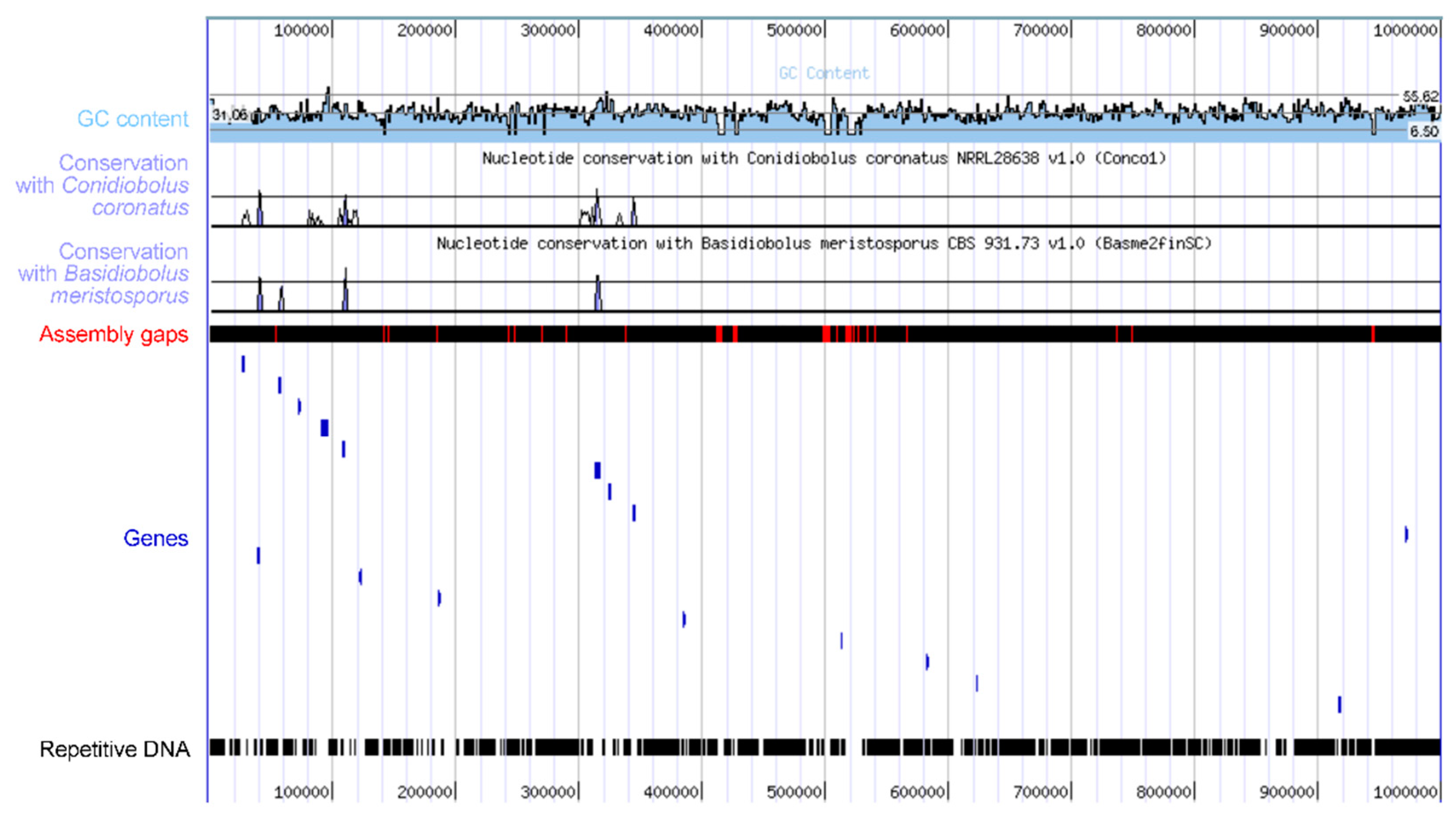
Figure 4.
The Z. radicans genome features large amounts of repetitive DNA between coding regions. This figure is a modified version of the MycoCosm [180] visualization of 1 Mb of sequence on contig 1, showing in blue only 17 genes across the region, separated by repetitive DNA in black. Conservation of DNA sequences with two other sequenced species in the Entomophthorales is low.
Figure 4.
The Z. radicans genome features large amounts of repetitive DNA between coding regions. This figure is a modified version of the MycoCosm [180] visualization of 1 Mb of sequence on contig 1, showing in blue only 17 genes across the region, separated by repetitive DNA in black. Conservation of DNA sequences with two other sequenced species in the Entomophthorales is low.
4. Discussion
The goal of using entomopathogenic fungi in various biotechnological applications, particularly to control populations of agricultural insect pests, has existed for several decades. However, entomophthoralean species have not been successfully developed and applied for biological control, despite numerous attempts. The major challenges in their application as biocontrol agents include the difficulty with cultivation of many species, requirements for specific abiotic conditions in the field, and the potentially low survival rates of these fungi outside of the host. However, EFPSZ fungi possess significant potential, which is still largely unexplored. Recent advances in genome sequencing technologies may allow researchers to access key genetic factors involved in virulence against insect hosts, in even those EFPSZ that cannot be easily cultivated, and biotechnology could then potentially be used to deploy these in various ways for pest control.
The ability of entomophthoralean species to infect insects from different families, or even from different orders, increases the diversity of target insect species for developing efficient biological control measures and selection of suitable pests to control. At the same time, the broad host-ranges of some of these fungi makes use of newly developed biological control remedies riskier as they may also affect other non-target insects, including those that are beneficial for natural ecosystems and humans.
While further research investigating host-range among EFPSZ is needed, the search for potential biocontrol agents within the Erynioideae might use information on current host-ranges and distributions, targeting hosts belonging to insect taxonomic groups that are known to be attacked by EFPSZ (Table 1). Even more important, prediction of possible non-targets of entomopathogenic species should account for insects within those same taxonomic groups in addition to pollinators and other beneficials. To some extent it is an advantage if a species already has been found on several continents and thus might be adapted for use in biological control over a large area. These species with high adaptability to various environments are promising candidates for targeting widespread insect hosts. Each genus of the EFPSZ group has several species that are distributed on at least three continents: in the genus Erynia, E. aquatica, E. conica, E. ovispora, and E. rhizospora. In the genus Furia, F. americana F. gastropachae, F. ithacensis, and F. virescens; and in the genus Pandora, P. blunckii, P. bullata, P. delphacis, P. dipterigena, P. gammae, P. neoaphidis, and P. nouryi. Many ubiquitous species are included in the genus Zoophthora: Z. aphids, Z. geometralis, Z. occidentalis, Z. phalloides, Z. phytonomi and Z. radicans. At least two species of the Strongwellsea, S. castrans and S. magna have been found on two continents. With increasing research on entomophthoralean fungi, we hypothesize that it is likely that a larger number of ubiquitous species will be identified.
Representatives of the genera Erynia, Pandora and Zoophthora are among important infective agents of insects under field conditions. The attempts at application of EFPSZ species on different continents has had success with Z. radicans in Australia using a strain originated from Israel to control the spotted alfalfa aphid, Therioaphis maculata [77].
A lower need for humidity could be considered an advantageous feature for potential biological control preparations using EFPSZ species. Aquatic species like E. aquatica might be successfully used only in wet habitats where they are highly adapted to the moist environment, and this could restrict application compared to the species found in diverse ecosystems. The broad ecological and geographical range of Z. radicans, recorded from numerous agricultural and natural habitats makes this species unique within the EFPSZ.
Cultivability is perhaps the main factor that determines the potential success of any biotechnological application with EFPSZ. If the fungus is hard to cultivate on artificial media, then the only way to apply it as a biological control agent is to keep it in vivo, infecting the insect population either in nature or under lab conditions. However, this is costly, cumbersome, and risky. Development of the biotechnological process and especially scaling it up demands easy culturing without losing virulence. The challenges of isolation into pure culture of the majority of EFPSZ species along with loss of vigor during numerous culture transfers significantly complicates the research on and development of potential biological control.
A problem with the successful pest control with many fungi is the level of susceptibility of the active organisms to external factors, such as fluctuations in temperature, humidity, and rainfall. Climate changes may significantly impact the relationship between fungi, insects and crops, and the interactions among them [181]. Furthermore, additional information needed for the eventual production of EFPSZ as biopesticides would be development of optimal methods for formulation and application.
5. Conclusions
Analysis of the available data on virulence, growth in vivo and in vitro, formulation, and field-testing suggests that one promising candidate for the development of an efficient biological control agents would be the species Z. radicans. This species seems to have potential for control of a range of lepidopteran larvae in many agricultural and forest ecosystems. Another species, P. neoaphidis, has great potential for control of numerous aphid species in cereals and legumes. Of special importance is the worldwide distribution of aphids impacting crops, and thus, the large market that exists. In orchards, P. cacopsyllae has recently proven to possess a potential for control of psyllid pests. A major objective here is that fruits are high value crops, that may favor biological control options. For the moist and aquatic habitats, there are several species infecting dipterans. Erynia aquatica and E. conica may have potential for mosquito control, however little is known about their virulence and growth in vitro. Obviously, the aforementioned factors are not the only factors determining the success or failure of biocontrol development; however, they play essential roles. However, advances in genome sequencing methods may allow researchers to access virulence factors and other genetic factors fro these fungi that could be harnessed for future biotechnological solutions.
Author Contributions
Conceptualization, APG and AEH; methodology, APG, AEH, AI and KEB; validation, YN, AEH, APG and AI; investigation, NV, LK, KEB, JE, YN, RGM, VBK and FA; resources, AEH, YN, KEB, AI, RGM and JE; data curation, APG, NV, YN, RGM, LK, AI, KEB and AEH; writing—original draft preparation, APG, AEH, AI and KEB; visualization, APG and AI; supervision, AEH and YN; funding acquisition, AI and APG. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported in part by the U.S. Department of Agriculture, Agricultural Research Service. AI was supported by the Hermon Slade Foundation (HSF22060).
Data Availability Statement
n. a.
Acknowledgments
Authors are thankful to Lucas Beagle, Division 30, and leadership of UES, an Eqlipse company, for the possibility to conduct and publish this study.
Conflicts of Interest
The authors declare no conflict of interest.
References
- N. E. Sacco and A. E. Hajek, “Diversity and Breadth of Host Specificity among Arthropod Pathogens in the Entomophthoromycotina,” Microorganisms, vol. 11, no. 7, p. 1658, 2023. [CrossRef]
- E. Hajek and J. Eilenberg, Natural enemies : an introduction to biological control, Second edition. Cambridge, UK; New York: Cambridge University Press, 2018., 2018. [Online]. Available: http://find.library.duke.edu/catalog/DUKE010116949.
- S. Keller and O. Petrini, “Keys to the identification of the arthropod pathogenic genera of the families Entomophthoraceae and Neozygitaceae (Zygomycetes), with descriptions of three new subfamilies and a new genus,” Sydowia, vol. 57, pp. 23–53, 2005.
- P. A. Shah and J. K. Pell, “Entomopathogenic fungi as biological control agents,” Appl Microbiol Biotechnol, vol. 61, no. 5, pp. 413–423, 2003. [CrossRef]
- J. P. M. Araújo and D. P. Hughes, “Chapter One - Diversity of Entomopathogenic Fungi: Which Groups Conquered the Insect Body?,” in Advances in Genetics, vol. 94, B. Lovett and R. J. St. Leger, Eds., in Genetics and Molecular Biology of Entomopathogenic Fungi, vol. 94. , Academic Press, 2016, pp. 1–39. [CrossRef]
- H. C. Evans, S. L. Elliot, and R. W. Barreto, “Entomopathogenic fungi and their potential for the management of Aedes aegypti (Diptera: Culicidae) in the Americas,” Mem Inst Oswaldo Cruz, vol. 113, no. 3, pp. 206–214, Mar. 2018. [CrossRef]
- B. S. Bamisile, J. A. Siddiqui, K. S. Akutse, L. C. Ramos Aguila, and Y. Xu, “General Limitations to Endophytic Entomopathogenic Fungi Use as Plant Growth Promoters, Pests and Pathogens Biocontrol Agents,” Plants, vol. 10, no. 10, Art. no. 10, 2021. [CrossRef]
- B. I. P. Barratt, “Assessing safety of biological control introductions.,” CABI Reviews, vol. 2011, pp. 1–12, 2012. [CrossRef]
- W. P. Madisson and D. R. Madisson, “Mesquite: a modular system for evolutionary analysis. Version 3.04,” Mesquite project. Accessed: Jun. 23, 2021. [Online]. Available: https://www.mesquiteproject.org/.
- D. L. Swofford, “PAUP*: Phylogenetic Analysis Using Parsimony (and other methods) Version 4.0 beta.” 2001.
- B. Lovett and R. J. St. Leger, “The Insect Pathogens,” Microbiology Spectrum, vol. 5, no. 2, p. 10.1128/microbiolspec.funk-0001–2016, 2017. [CrossRef]
- ARSEF, “ARSEF Catalogues: USDA ARS.” Accessed: Oct. 31, 2023. [Online]. Available: https://www.ars.usda.gov/northeast-area/ithaca-ny/robert-w-holley-center-for-agriculture-health/emerging-pests-and-pathogens-research/docs/mycology/page-2-arsef-catalogues/.
- M. W. Miller and C. B. Keil, “Redescription of Pandora gloeospora (Zygomycetes: Entomophthorales) from Lycoriella mali (Diptera: Sciaridae),” Mycotyaxon, vol. 38, pp. 227–231, 1990.
- J. F. Anderson and S. Z. Rongo, “Entomophthora aquatica sp. n. infecting larvae and pupae of floodwater mosquitoes,” Journal of Invertebrate Pathology, vol. 13, no. 3, pp. 386–393, 1969. [CrossRef]
- S. Bałazy, Flora of Poland. Fungi (Mycota), Entomophthorales, vol. 24. Kraków, Poland: Polish Academy of Sciences, W. Szafer Institute of Botany, 1993.
- M. Gustafsson, “On the species of the genus Entomophthora Fr. in Sweden. 1. Classification and distribution,” Kungliga Landbruks-Högskolans Annaler, vol. 31, pp. 103–121, 1965.
- M. Gustafsson, “On species of the genus Entomophthora Fres. in Sweden. III. Possibility of usage in biological control,” Lantbrukshögsk. Ann., vol. 35, pp. 235–274, 1969.
- S. Keller, “Arthropod-pathogenic Entomophthorales from Switzerland. III. First additions,” Sydowia, vol. 59, no. 1, pp. 75–113, 1991.
- S. Keller, “Arthropod-pathogenic Entomophthorales of Switzerland. II. Erynia, Eryniopsis, Neozygites, Zoophthora and Tarichium,” Sydowia, vol. 43, pp. 39–122, 1991.
- S. Keller and Y. G. C. Dhoi, “Insect pathogenic Entomophthorales from Nepal and India,” Mitteilungen der Schweizerischen Entomologischen Gesellschaft, vol. 80, pp. 211–215, 2007.
- E. Z. Koval, Flora Fungorum Ucrainical. Zygomycotina. Entomophthorales. Kyiv, Ukraine: National Academy of Sciences, 2007.
- D. M. MacLeod and E. Müller-Kögler, “Entomogenous fungi: Entomophthora species with pear-shaped to almost spherical conidia (Entomophthorales: Entomophthoraceae),” Mycologia, vol. 65, no. 4, pp. 823–893, 1973. [CrossRef]
- M. Niell and S. Santamaria, “Additions to the knowledge of Entomopathogenic Entomophthorales (Fungi, Zygomycota) from Spain,” Nova Hedwigia, vol. 73, no. 1, pp. 167–197, 2001. [CrossRef]
- M. Z. Fan and Z. Z. Li, “Two new pathogens of dipteran insects,” Mycotaxon, vol. 50, pp. 307–314, 1994.
- M. Z. Fan and Z. Z. Li, “Erynia chironomis, comb. nov. (Zygomycetes: Entomophthorales),” Mycotaxon, vol. 53, p. 369, 1995.
- Z. Z. Li, Flora fungorum sinicorum. Entomophthorales, vol. 13. Beijing: Science Press, 2000.
- S. Keller, “Arthropod-pathogenic Entomophthorales from Switzerland. III. First additions,” Sydowia, vol. 59, no. 1, pp. 75–113, 2007.
- S. Bałazy, “On some little known epizootics in noxious and beneficial arthropod populations caused by entomophthoralean fungi,” IOBC WPRS Bull., vol. 21, no. 6, pp. 63–68, 2003.
- S. Ben-Ze’ev, Y. Zelig, S. Bitton, and R. G. Kenneth, “The entomophthorales of Israel and their arthropod hosts: Additions 1980–1988,” Phytoparasitica, vol. 16, no. 3, pp. 247–257, 1988. [CrossRef]
- A. A. Evlakhova, Entomopathogenic Fungi. Moscow: Nauka, 1974.
- J. A. Hutchison, “The Genus Entomophthora in the Western Hemisphere,” Transactions of the Kansas Academy of Science (1903-), vol. 66, no. 2, pp. 237–254, 1963. [CrossRef]
- E. Z. Koval, Identification of Entomophilic Fungi of the USSR. Kyiv, Ukraine: Naukova Dumka, 1974.
- D. Leatherdale, “The arthropod hosts of entomogenous fungi in Britain,” Entomophaga, vol. 15, no. 4, pp. 419–435, 1970. [CrossRef]
- M. P. Nadeau, G. B. Dunphy, and J. L. Boisvert, “Entomopathogenic fungi of the order Entomophthorales (Zygomycotina) in adult black fly populations (Diptera: Simuliidae) in Quebec,” Can J Microbiol, vol. 40, no. 8, pp. 682–686, 1994. [CrossRef]
- Z. Z. Li, Z. A. Chen, and Y. W. Xu, “Erynia gigantea, a new pathogen of spittlebug, Aphrophora sp.,” Acta Mycologica Sinica, vol. 9, no. 4, p. 263-265, 1990.
- L.-S. Zha, T.-C. Wen, K. D. Hyde, and J.-C. Kang, “中国虫霉目真菌及其寄主昆虫名录更新,” Mycosystema, vol. 35, no. 6, pp. 666–683, 2016. [CrossRef]
- S. Keller, “New records of Entomophthoraceae (Fungi, Zygomycetes) from aquatic insects,” Mitt. Schweiz. Entomol. Gesell., vol. 78, pp. 333–336, 2005.
- R. A. Humber and I. Ben-Ze‘ev, “Erynia (Zygomycetes: Entomophthorales): Emendation, Synonymy, and Transfers,” Mycotaxon, vol. XIII, no. 3, pp. 506–516, 1981.
- S. Keller and T. Hülsewig, “Amended description and new combination for Entomophthora nebriae Raunkiaer, (1893), a little known entomopathogenic fungus attacking the ground beetle Nebria brevicollis (Fabricius, 1792),” Alpine Entomology, vol. 2, pp. 1–5, 2018. [CrossRef]
- C. Tkaczyk, S. Bałazy, T. Krzyczkowski, and R. Wegensteiner, “Extended studies on the diversity of arthropod-pathogenic fungi in Austria and Poland,” Acta Mycologica, vol. 46, no. 2, Art. no. 2, 2011. [CrossRef]
- C. S. Jia, “Morphology of Pandora dipterigena collected from Lucilia sericata,” Chinese Journal of Vector Biology and Control, vol. 21, no. 6, p. 546-548, 2010.
- R. Thaxter, “The Entomophthoreae of the United States,” Harvard botanical memoirs, vol. 2, pp. 133–201, 1888.
- S. Keller, “Arthropod-pathogenic Entomophthorales from Switzerland. IV. Second addition,” Mitteilungen der Schweizerischen Entomologischen Gesellschaft, vol. 85, pp. 115–130, 2012.
- Y. J. Huang and Z. Z. Li, “Furia fujiana, a new pathogen of pale-lined tiger moth, Spilarctia obliqua,” Acta Mycologica Sinica, vol. 12, no. 1, pp. 1–4, 1993.
- M. J. Filotas, A. E. Hajek, and R. A. Humber, “Prevalence and biology of Furia gastropachae (Zygomycetes: Entomophthorales) in populations of forest tent caterpillar (Lepidoptera: Lasiocampidae),” The Canadian Entomologist, vol. 135, no. 3, pp. 359–378, 2003. [CrossRef]
- D. M. MacLeod and D. Tyrrell, “Entomophthora crustosa n. sp. as a pathogen of the forest tent caterpillar, Malacosoma disstria (Lepidoptera: Lasiocampidae),” The Canadian Entomologist, vol. 111, pp. 1137–1144, 1979.
- R. R. de Moraes, A. E. Loeck, and L. C. Berlamino, “Inimigos Naturais de Rachiplusia Nu (Guenne, 1852) e de Pseudoplusia Includens (Walker, 1857) (Lepidoptera: Noctuidae) em Soja no Rio Grande so Sul. - Portal Embrapa,” Pesq. Agropec. Bras., vol. 26, pp. 57–64, 1991.
- Y. J. Huang, B. N. Zheng, and Z. Z. Li, “Natural and induced epizootics of Erynia ithacensis in mushroom hothouse populations of yellow-legged fungus gnats,” Journal of Invertebrate Pathology, vol. 60, no. 3, p. 254-258, 1992. [CrossRef]
- Ben-Ze‘ev, “Erynia neopyralidarum sp. nov. and Conidiobolus apiculatus, pathogens of pyralid moths components of the misdescribed species, Entomophthora pyralidarum (Zygomycetes: Entomophthorales),” Mycotaxon, vol. 16, no. 1, pp. 273–292, 1982.
- R. M. C. Pena, Y. Kunimi, T. Motobayashi, Y. Endoh, K. Numazawa, and J. Aoki, “Erynia neopyralidarum Ben-Ze’ev (Zygomycetes : Entomophthorales) Isolated from the Mulberry Tiger Moth, Spilosoma imparilis Butler (Lepidoptera : Arctiidae) in Japan,” Applied Entomology and Zoology, vol. 25, no. 4, pp. 503–508, 1990. [CrossRef]
- W. Li, X. F. Wang, Z. H. Li, W. A. Xu, and C. F. Sheng, “Survey of entomophthoralean fungi in Shandong,” Entomological Knowledges, vol. 41, no. 4, pp. 350–353, 2004.
- Z. Z. Li, M. Z. Fan, and B. Huang, “New combinations of entomophthoralean fungi originally in the genus Erynia,” Mycosystema, vol. 17, no. 1, pp. 91–94, 1998.
- L. Villacarlos and N. Wilding, “Four new species of Entomophthorales infecting the leucaena psyllid Heteropsylla cubana in the Philippines,” Mycological Research, vol. 98, no. 2, pp. 153–164, 1994. [CrossRef]
- G. R. Lednev, Pathogens of the insect mycoses. Diagnostics manual. Saint Petersburg: Innovation Center of All-Russian Institute of Plant Protection Ltd., 2003.
- D. C. Steinkraus, A. J. Mueller, and R. A. Humber, “Furia virescens (Thaxter) Humber (Zygomycetes: Entomophthoraceae) Infections in the Armyworm, Pseudaletia unipuncta (Haworth) (Lepidoptera: Noctuidae) in Arkansas with Notes on Other Natural Enemies,” Journal of Entomological Science, vol. 28, no. 4, pp. 376–386, 1993. [CrossRef]
- S. R. Sanchez-Peña, “Entomopathogens from two Chihuahuan desert localities in Mexico,” BioControl, vol. 45, no. 1, pp. 63–78, 2000. [CrossRef]
- S. Keller, T. Hülsewig, and A. B. Jensen, “Fungi attacking springtails (Sminthuridae, Collembola) with a description of Pandora batallata, sp. nov. (Entomophthoraceae),” Sydowia, vol. 75, pp. 37–45, 2022.
- I. Ben-Ze’ev, R. G. Kenneth, and S. Bitton, “The Entomophthorales of Israel and their arthropod hosts,” Phytoparasitica, vol. 9, no. 1, pp. 43–50, 1981. [CrossRef]
- M. Z. Fan, C. Guo, R. G. Liu, D. J. Li, and C. J. Wang, “New record genus Strongwellsea and Pandora blunckii in China,” Journal of Northwest Forestry College, vol. 7, no. 2, pp. 26–30, 1992.
- A. Guzman-Franco, S. Clark, P. Alderson, and J. Pell, “Effect of temperature on the in vitro radial growth of Zoophthora radicans and Pandora blunckii, two co-occurring fungal pathogens of the diamondback moth Plutella xylostella,” BioControl, vol. 53, pp. 501–516, 2007. [CrossRef]
- J. Sun, J. N. Liu, Q. Q. Wu, S. B. Wang, R. S. Cai, and B. Huang, “Investigation of Entomophthoralean resourse in Hefei,” Jorunal of Anhui Agricultural University, vol. 27, no. 3, pp. 240–242, 2000.
- H. Tomiyama and J. Aoki, “Infection of Erynia blunkii (Lak. ex. Zimm.) Rem. et Henn. (Entomophthorales: Entomophthoraceae) in the diamondback moth h, Plutella xylostella L. (Lepidoptera: Yponomeutidae),” Applied Entomology and Zoology, vol. 17, pp. 373–384, 1982. [CrossRef]
- L. Villacarlos, “Updated records of Philippine Entomophthorales,” Philippine Entomologist, vol. 22, no. 1, pp. 22–52, 2008.
- M. Z. Fan, C. Guo, and Z. Z. Li, “New species and a new record of the genus Erynia in China.,” Acta Mycologica Sinica, vol. 10, no. 2, pp. 95–100, 1991.
- D. M. MacLeod, D. Tyrrell, R. S. Soper, and A. J. De Lyzer, “Entomophthora bullata as a pathogen of Sarcophaga aldrichi,” Journal of Invertebrate Pathology, vol. 22, no. 1, pp. 75–79, 1973. [CrossRef]
- T. R. Glare and R. J. Milner, “New Records of Entomophthoran Fungi from Insects in Australia,” Aust. J. Bot., vol. 35, no. 1, pp. 69–77, 1987. [CrossRef]
- S. Keller, “Taxonomic considerations on some species of Erynia (Zygomycetes: Entomophthorales) attacking flies (Diptera),” Sydowia, vol. 45, pp. 252–263, 1993.
- S. Zangeneh and M. Ghazavi, “New Records of Entomophthoralean Fungi from Iran,” Rostaniha, vol. 9, no. 2, pp. 190–203, 2008.
- C. Montalva, K. Collier, C. Luz, and R. A. Humber, “Pandora bullata (Entomophthoromycota: Entomophthorales) affecting calliphorid flies in central Brazil,” Acta Tropica, vol. 158, pp. 177–180, 2016. [CrossRef]
- J. Eilenberg et al., “Pandora cacopsyllae Eilenberg, Keller & Humber (Entomophthorales: Entomophthoraceae), a new species infecting pear psyllid Cacopsylla pyri L. (Hemiptera: Psyllidae),” Journal of Invertebrate Pathology, vol. 200, p. 107954, 2023. [CrossRef]
- R. A. Humber, “Synopsis of a revised classification for the Entomophthorales (Zygomycotina),” Mycotaxon, vol. 34, pp. 441–460, 1989.
- M. K. Miller and J. D. Harper, “Occurrence of Erynia delphacis in the threecornered alfalfa hopper, Spissistilus festinus (Homoptera: Membracidae),” Journal of Invertebrate Pathology, vol. 50, no. 1, pp. 81–83, 1987. [CrossRef]
- Z. Z. Li, M. Z. Fan, and C. F. Qin, “New species and new records of Entomophthorales in China,” Acta Mycologica Sinica, vol. 11, no. 3, pp. 182–187, 1992.
- G. M. I. Lorenz, “Pandora delphacis (Entomophthorales: Entomophthoraceae) an entomopathogenic fungus of the threecornered alfalfa hopper, Spissistilus festinus (Homoptera: Membracidae),” University of Arkansas ProQuest Dissertations Publishing, Ann Arbor, MI, 1995. Accessed: Aug. 25, 2023. [Online]. Available: https://www.proquest.com/openview/74cf7739893ac6f070c578200c04fcfc/1?pq-origsite=gscholar&cbl=18750&diss=y.
- M. Takehiko, S. Hiroki, and S. Mitsuaki, “Isolation of an entomogenous fungus, Erynia delphacis (Entomophthorales : Entomophthoraceae), from migratory planthoppers collected over the Pacific Ocean,” Applied Entomology and Zoology, vol. 33, no. 4, pp. 545–549, 1998. [CrossRef]
- A. Foieri, N. Pedrini, and A. Toledo, “Natural occurrence of the entomopathogenic genus Pandora on spittlebug pests of crops and pastures in Argentina,” Journal of Applied Entomology, vol. 142, no. 3, pp. 363–370, 2017. [CrossRef]
- M. Barta and L. Cagáň, “Aphid-pathogenic Entomophthorales (their taxonomy, biology and ecology),” Biologia, vol. 61, no. 21, pp. S543–S616, 2006. [CrossRef]
- M. Barta and L. Cagáň, “Observations on the Occurrence of Entomophthorales Infecting Aphids (Aphidoidea) in Slovakia,” BioControl, vol. 51, no. 6, pp. 795–808, 2006. [CrossRef]
- I. S. Ben-Ze’ev, “Check-list of fungi pathogenic to insects and mites in Israel, updated through 1992,” Phytoparasitica, vol. 21, no. 3, pp. 213–237, 1993. [CrossRef]
- Y. J. Huang and B. N. Zheng, “A new record species of Erynia from China,” Acta Mycologica Sinica, vol. 9, no. 4, p. 327-328, 1990.
- S. E. Méndez-Sánchez, A. L. Freitas, C. S. de Almeida, G. B. Silva, and L. S. Lima, “Levantamiento preliminar de hongos entomophthorales (Zygomycotina: Zygomycetes), agentes de control natural de insectos al sur de Bahia, Brasil,” Agrotropica, vol. 14, no. 2, Art. no. 2, 2002.
- G. Remaudière and J. P. Latge, “Importancia de los hongos patógenos de insectos (especialmente Aphididae y Cercopidae) en Méjico y perspectivas de uso,” Boletín de sanidad vegetal. Plagas, vol. 11, no. 2, pp. 217–225, 1985.
- A. M. Young, D. Tyrrell, and D. M. MacLeod, “Entomophthora echinospora (Phycomycetes: Entomophthoraceae), a fungus pathogenic on the neotropical cicada, Procollina biolleyi (Homoptera: Cicadidae),” Journal of Invertebrate Pathology, vol. 21, no. 1, pp. 87–90, 1973. [CrossRef]
- P. Baiswar and D. M. Firake, “First record of Pandora formicae on ant, Camponotus angusticollis and Batkoa amrascae on white leaf hopper, Cofana spectra in rice agroecosystem of India,” Journal of Eco-friendly Agriculture, vol. 17, no. 1, Art. no. 1, 2022. [CrossRef]
- J. Małagocka, A. B. Jensen, and J. Eilenberg, “Pandora formicae, a specialist ant pathogenic fungus: New insights into biology and taxonomy,” Journal of Invertebrate Pathology, vol. 143, pp. 108–114, 2017. [CrossRef]
- G. G. Newman and G. R. Carner, “Factors affecting the spore form of Entomophthora gammae,” Journal of Invertebrate Pathology, vol. 26, no. 1, pp. 29–34, 1975. [CrossRef]
- F. Moscardi, B. S. Correra Ferreira, C. B. Hoffmann-Campo, E. B. de Oliveira, and D. G. Boucias, “Ocorrência de entomopatogenos em lepidópteros que atacam a cultura da soja no Paraná,” presented at the Congresso Brasiliero de Entomologia, Londrina, PR: Resumos, 1984, p. 143.
- S. L. DIez and J. C. Gamundi, “Empleo de enfermedades como método de biocontrol de insectos plagas,” Tecnología para el campo. INTA, vol. 10, pp. 19–20, 1985.
- R. A. Samson, H. C. Evans, and J.-P. Latgé, Atlas of Entomopathogenic Fungi. Berlin, Heidelberg: Springer, 1988. [CrossRef]
- J. D. Edelstein and R. E. Lecuona, “Presencia del hongo entomopatógeno Pandora gammae (Weiser) Humber (Zygomycetes: Entomophthorales), en el complejo de ‘orugas medidoras de la soja’ (Lepidoptera: Plusiinae) en Argentina,” RIA. Revista de Investigaciones Agropecuarias, vol. 32, no. 1, pp. 31–38, 2003.
- C. C. López Lastra and A. C. Scorsetti, “Hongos patógenos de insectos en Argentina (Zygomycetes: Entomophthorales,” Revista de Biología Tropical, vol. 54, no. 2, pp. 311–315, 2006. [CrossRef]
- C. C. López Lastra and A. C. Scorsetti, “Revisión de los hongos Entomophthorales (Zygomycota: Zygomycetes) patógenos de insectos de la República Argentina,” Boletín de la Sociedad Argentina de Botánica, vol. 41, no. 1–2, pp. 33–37, 2007.
- Z. Z. Li, B. Huang, and M. Z. Fan, “New species, new record, new combinations and emendation of entomophthoralean fungi pathogenic on dipteran insects,” Mycosystema, vol. 16, no. 2, pp. 91–96, 1997.
- C. S. Jia and B. Hong, “Zoophthora radicans infecting larvae of Cnaphalocrocis medinalis and its epizootic,” Mycosystema, vol. 31, no. 3, pp. 322–330, 2012.
- J. J. Hannam and D. C. Steinkraus, “The natural occurrence of Pandora heteropterae (Zygomycetes: Entomophthorales) infecting Lygus lineolaris (Hemiptera: Miridae),” Journal of Invertebrate Pathology, vol. 103, no. 2, pp. 96–102, 2010. [CrossRef]
- R. J. Milner and J. Bourne, “Influence of temperature and duration of leaf wetness on infection of Acyrthosiphon kondoi with Erynia neoaphidis,” Annals of Applied Biology, vol. 102, no. 1, pp. 19–27, 1983. [CrossRef]
- I. M. Hall, A. D. Lowe, and B. B. Given, “New record of aphid hosts of Entomophthora aphidis and Entomophthora planchoniana in New Zealand,” N. Z. J. Zool., vol. 3, pp. 111–112, 1976.
- R. J. Milner, R. E. Teakle, G. G. Lutton, and F. M. Dare, “Pathogens (Phycomycetes: Entomophthoraceae) of the blue-green aphid Acyrthosiphon kondoi Shinji and other aphids in Australia,” Aust. J. Bot., vol. 28, no. 6, pp. 601–619, 1980. [CrossRef]
- Y. Kobayashi, K. Mogami, and J. Aoki, “Ultrastructural studies on the hyphal growth of Erynia neoaphidis in the green peach aphid, Myzus persicae,” Trans. Mycol. Soc. Jpn., vol. 25, pp. 425–434, 1984.
- W. H. Lu and W. M. Wang, “Investigation and identification of several aphid entomophthoralean fungi in Shandong Province,” Microbiology, vol. 15, p. 155-159, 1988.
- B. Papierok, “On the occurrence of Entomophthorales (Zygomycetes) in Finland. I. Species attacking aphids (Homoptera, Aphididae),” Ann. Entomol. Fenn., vol. 55, pp. 63–69, 1989.
- M.-G. Feng, J. B. Johnson, and L. P. Kish, “Survey of Entomopathogenic Fungi Naturally Infecting Cereal Aphids (Homoptera: Aphididae) of Irrigated Grain Crops in Southwestern Idaho,” Environmental Entomology, vol. 19, no. 5, pp. 1534–1542, 1990. [CrossRef]
- M. G. Feng, R. M. Nowierski, A. L. Scharen, and D. C. Sands, “Entomopathogenic fungi (Zygomycotina: Entomophthorales) infecting cereal aphids (Homoptera: Aphididae) in Montana.,” Pan-Pacific Entomologist, vol. 7, no. 1, pp. 55–64, 1991.
- Z. Yu, G. L. Nordin, G. C. Brown, and D. M. Jackson, “Studies on Pandora neoaphidis (Entomophthorales: Entomophthoraceae) Infectious to the Red Morph of Tobacco Aphid (Homoptera: Aphididae),” Environmental Entomology, vol. 24, no. 4, pp. 962–966, 1995. [CrossRef]
- S. S. Tzean, L. S. Hsieh, and W. J. Wu, Atlas of Entomopathogenic Fungi from Taiwan. Taiwan, Republic of China: Council of Agriculture, Executive Yuan, 1997.
- C.-S. Yoon, G.-H. Sung, B.-R. Choi, J.-K. Yoo, and J.-O. Lee, “The Aphid-attacking Fungus Pandora neoaphidis; the First Observation and its Host Range in Korea,” The Korean Journal of Mycology, vol. 26, no. 3, pp. 407–410, 1998.
- J. L. Hatting, R. A. Humber, T. J. Poprawski, and R. M. Miller, “A Survey of Fungal Pathogens of Aphids from South Africa, with Special Reference to Cereal Aphids,” Biological Control, vol. 16, no. 1, pp. 1–12, Sep. 1999. [CrossRef]
- J. L. Hatting, T. J. Poprawski, and R. M. Miller, “Prevalences of fungal pathogens and other natural enemies of cereal aphids (Homoptera:Aphididae) in wheat under dryland and irrigatedconditions in South Africa,” BioControl, vol. 45, no. 2, pp. 179–199, 2000. [CrossRef]
- C. Nielsen, J. Eilenberg, S. Harding, E. Oddsdottir, and G. Halldórsson, “Geographical distribution and host range of entomophthorales infecting the green spruce aphid Elatobium abietinum Walker in Iceland,” J Invertebr Pathol, vol. 78, no. 2, pp. 72–80, 2001. [CrossRef]
- C. Nielsen, C. Sommer, J. Eilenberg, K. S. Hansen, and R. A. Humber, “Characterization of aphid pathogenic species in the genus Pandora by PCR techniques and digital image analysis,” Mycologia, vol. 93, no. 5, pp. 864–874, 2001. [CrossRef]
- E. G. Voronina, G. R. Lednev, and T. J. Mukamolova, “Entomophthoralean fungi,” in Pathogens of Insects: structural and functional aspects, V. V. Glupov., Moscow: Kruglyi god, 2001, pp. 271–351.
- W. Li and C.-F. Sheng, “Occurrence and distribution of entomophthoralean fungi infecting aphids in mainland China,” Biocontrol Science and Technology, vol. 17, no. 4, pp. 433–439, 2007. [CrossRef]
- A. C. Scorsetti, R. A. Humber, J. J. García, and C. C. L. Lastra, “Natural occurrence of entomopathogenic fungi (Zygomycetes: Entomophthorales) of aphid (Hemiptera: Aphididae) pests of horticultural crops in Argentina,” BioControl, vol. 52, no. 5, pp. 641–655, 2007. [CrossRef]
- A. C. Scorsetti, A. Maciá, D. C. Steinkraus, and C. C. López Lastra, “Prevalence of Pandora neoaphidis (Zygomycetes: Entomophthorales) infecting Nasonovia ribisnigri (Hemiptera: Aphididae) on lettuce crops in Argentina,” Biological Control, vol. 52, no. 1, pp. 46–50, 2010. [CrossRef]
- R. Alzugaray et al., “Prospección de agentes de mortalidad natural de áfidos en leguminosas forrajeras en Uruguay,” Agrociencia (Uruguay), vol. 14, no. 1, pp. 27–35, 2010.
- A. Moubasher, M. Abdel-Rahman, A. Abdel-Mallek, and G. Hammam, “Biodiversity of entomopathogenic fungi infecting wheat and cabbage aphids in Assiut, Egypt,” 2010. Accessed: Oct. 29, 2023. [Online]. Available: https://www.semanticscholar.org/paper/Biodiversity-of-entomopathogenic-fungi-infecting-in-Moubasher-Abdel-Rahman/eacd7e417f99819407ecd4adb9c988585db3c1ad.
- I. Ben Fekih, S. Boukhris-Bouhachem, M. Bechir Allagui, A. Bruun Jensen, and J. Eilenberg, “First survey on ecological host range of aphid pathogenic fungi (Phylum Entomophthoromycota) in Tunisia,” Annales de la Société entomologique de France (N.S.), vol. 51, no. 2, pp. 140–144, 2015. [CrossRef]
- I. Ben Fekih et al., “Virulence of Two Entomophthoralean Fungi, Pandora neoaphidis and Entomophthora planchoniana, to Their Conspecific (Sitobion avenae) and Heterospecific (Rhopalosiphum padi) Aphid Hosts,” Insects, vol. 10, no. 2, p. 54, 2019. [CrossRef]
- S. Keller, “Entomophthorales attacking aphids with a description of two new species,” Sydowia, vol. 58, no. 1, pp. 38–74, 2006.
- A. V. Toledo, R. A. Humber, and C. C. L. Lastra, “First and southernmost records of Hirsutella (Ascomycota: Hypocreales) and Pandora (Zygomycota: Entomophthorales) species infecting Dermaptera and Psocodea,” J Invertebr Pathol, vol. 97, no. 2, pp. 193–196, 2008. [CrossRef]
- T. Steenberg, V. Langer, and P. Esbjerg, “Entomopathogenic fungi in predatory beetles (Col.: Carabidae and Staphylinidae) from agricultural fields,” Entomophaga, vol. 40, no. 1, pp. 77–85, 1995. [CrossRef]
- Yu. A. Gres and E. Z. Koval, “A new species Entomorphthora terrestris sp. nova. Affecting the sugar-beet root aphid,” Mikrobiologicheskij Zhurnal, vol. 44, no. 2, pp. 64–69, 1982.
- A. Barta and L. Cagáň, “Pandora uroleuconii sp. nov. (Zygomycetes: Entomophthoraceae), a new pathogen of aphids.,” Mycotaxon, vol. 88, pp. 79–86, 2003.
- J. Eilenberg, V. Michelsen, and R. Humber, “Strongwellsea tigrinae and Strongwellsea acerosa (Entomophthorales: Entomophthoraceae), two new species infecting dipteran hosts from the genus Coenosia (Muscidae),” Journal of Invertebrate Pathology, vol. 175, p. 107444, Jul. 2020. [CrossRef]
- A. Batko and J. Weiser, “On the taxonomic position of the fungus discovered by Strong, Wells, and Apple: Strongwellsea castrans gen. et sp. nov. (Phycomycetes; Entomophthoraceae),” Journal of Invertebrate Pathology, vol. 7, no. 4, pp. 455–463, 1965.
- S. Keller, “Arthropod-pathogenic Entomophthorales of Switzerland. I. Conidiobolus, Entomophaga and Entomophthora,” Sydowia, vol. 40, pp. 122–167, 1987.
- J. Eilenberg, V. Michelsen, A. B. Jensen, and R. A. Humber, “Strongwellsea crypta (Entomophthorales: Entomophthoraceae), a new species infecting Botanophila fugax (Diptera: Anthomyiidae),” Journal of Invertebrate Pathology, vol. 186, p. 107673, 2021. [CrossRef]
- J. Eilenberg, V. Michelsen, A. B. Jensen, and R. A. Humber, “Strongwellsea selandia and Strongwellsea gefion (Entomophthorales: Entomophthoraceae), two new species infecting adult flies from genus Helina (Diptera: Muscidae),” Journal of Invertebrate Pathology, vol. 193, p. 107797, Sep. 2022. [CrossRef]
- R. A. Humber, “The systematics of the genus Strongwellsea (Zygomycetes: Entomophthorales),” Mycologia, vol. 68, no. 5, pp. 1042–1060, 1976.
- J. Eilenberg and V. Michelsen, “Natural host range and prevalence of the genus Strongwellsea (Zygomycota: Entomophthorales) in Denmark,” J Invertebr Pathol, vol. 73, no. 2, pp. 189–198, 1990. [CrossRef]
- J. Eilenberg and A. B. Jensen, “Strong host specialization in fungus genus Strongwellsea (Entomophthorales),” Journal of Invertebrate Pathology, vol. 157, pp. 112–116, 2018. [CrossRef]
- G. Turian, “Entomo-mycoses dans la région de Genéve,” Entomol. Ges., vol. 30, pp. 93–98, 1957.
- Z. Z. Li, “Erynia anhuensis, a new pathogen of aphids,” Acta Mycologica Sinica, vol. 5, no. 1, pp. 1–6, 1986.
- R. X. Gao, G. X. Lin, and X. Guan, “A list of Fujian pathogenic microbes on pest,” Journal of Fujian Agricultural College, vol. 15, no. 4, pp. 300–310, 1986.
- L. T. Villacarlos and B. S. Mejia, “Philippine entomopathogenic fungi I. Occurrence and diversity,” Philippine Agricultural Scientist, vol. 87, no. 3, pp. 249–265, 2004.
- E. G. Voronina and T. J. Mukamolova, Diagnostics of Entomophthoroses of harmful insects. Saint Petersburg: OOO Innovation Center VIZR, 2002.
- Z. Z. Li, Z. A. Chen, C. R. Lu, and H. Z. Hong, “Some entomophthoralean species in Shennongjia,” in Fungi and lichens of Shengnongjia, in Mycological and lichenological expedition to Shengnongjia, Academia Sinica (ed.). , Beijing: World Publishing Corporation, 1989, pp. 79–83.
- D. M. MacLeod, D. Tyrrell, and R. S. Soper, “Entomophthora canadensis n.sp., a fungus pathogenic on the woolly pine needle aphid, Schizolachnus piniradiatae,” Canadian Journal of Botany, vol. 57, no. 23, pp. 2663–2672, 1979. [CrossRef]
- C. Aruta and R. Carvillo, “Identificación de hongos del orden Entomophthorales en Chile. III,” Agro Sur., vol. 17, pp. 10–14, 1989.
- A. E. Hajek et al., “Phylogenetic placement of two species known only from resting spores: Zoophthora independentia sp. nov. and Z. porteri comb nov. (Entomophthorales: Entomophthoraceae),” Journal of Invertebrate Pathology, vol. 140, pp. 68–74, 2016. [CrossRef]
- S. Keller, “Two new species of the genus Zoophthora Batko (Zygomycetes, Entomophthoraceae): Z. lanceolata and Z. crassitunicata,” Sydowia, no. 33, pp. 167–173, 1980.
- C. Nielsen and A. E. Hajek, “Control of Invasive Soybean Aphid, Aphis glycines (Hemiptera: Aphididae), Populations by Existing Natural Enemies in New York State, with Emphasis on Entomopathogenic Fungi,” Environmental Entomology, vol. 34, no. 5, pp. 1036–1047, 2005. [CrossRef]
- K. L. Wu, Z. H. Huang, and C. H. Chen, “A new Zoophthora occidentalis pathogenof Myzus persicae in China,” Fujian Journal of Agricultural Sciences, vol. 29, no. 7, p. 673-677, 2014.
- T. R. Glare, R. J. Milner, and G. A. Chilvers, “The effect of environmental factors on the production, discharge, and germination of primary conidia of Zoophthora phalloides Batko,” Journal of Invertebrate Pathology, vol. 48, no. 3, p. 275, 1986. [CrossRef]
- Uziel and R. G. Kenneth, “Survival of primary conidia and capilliconidia at different humidities in Erynia (subgen. Zoophthora) spp. and in Neozygites fresenii (Zygomycotina: Entomophthorales), with special emphasis on Erynia radicans,” Journal of Invertebrate Pathology, vol. 58, no. 1, pp. 118–126, 1991. [CrossRef]
- R. G. Manfrino, L. A. Castrillo, C. C. L. Lastra, A. V. Toledo, W. Ferrari, and A. B. Jensen, “Morphological and Molecular Characterization of Entomophthorales (Entomophthoromycota: Entomophthoromycotina) from Argentina,” Acta Myc, vol. 55, no. 2, 2020. [CrossRef]
- A. E. Hajek, K. T. Hodge, J. K. Liebherr, W. H. Day, and J. D. Vandenberg, “Use of RAPD analysis to trace the origin of the weevil pathogen Zoophthora phytonomi in North America,” Mycological Research, vol. 100, no. 3, pp. 349–355, 1996. [CrossRef]
- M. Shimazu, “Resting Spore Formation of Entmophthora sphaerosperma FRESENIUS (Entomophthorales : Entmophthoraceae) in the Brown Planthopper, Nilaparvata lugens (STAL)(Hemiptera : Delphacidae),” Appl. entomol. Zool, vol. 14, no. 4, pp. 383–388, 1979. [CrossRef]
- R. J. Milner and R. J. Mahon, “Strain Variation in Zoophthora Radicans, a Pathogen on a Variety of Insect Hosts in Australia,” Australian Journal of Entomology, vol. 24, no. 3, pp. 195–198, 1985. [CrossRef]
- B. Huang, B. N. Zheng, and H. Q. Zhu, “Isolation, identification and biological determination of Erynia radicans,” Practical Forestry Technology, vol. 1998, no. 6, pp. 16–18, 1988.
- S. Galaini-Wraight, S. P. Wraight, R. I. Carruthers, B. P. Magalhães, and D. W. Roberts, “Description of a Zoophthora radicans (Zygomycetes: Entomophthoraceae) epizootic in a population of Empoasca kraemeri (Homoptera: Cicadellidae) on beans in central Brazil,” Journal of Invertebrate Pathology, vol. 58, no. 3, pp. 311–326, 1991. [CrossRef]
- R. Alzugaray, S. Stewart, and M. S. Zebrino, “Epizootia por hongos sobre Epinotia aporema (Wals) (Lepidoptera: Tortricidae) en Uruguay,” Boletín de la Sociedad Zoológica de Uruguay, vol. 7, p. 79, 1992.
- R. Alzugaray, M. S. Zerbino, S. Stewart, A. Ribeiro, and J. Eilenberg, “Epizootiologia de hongos Entomophthorales. Uso de Zoophthora radicans (Brefeld) Batko (Zygomicotina: Entomophthorales) para el control de Epinotia aporema (Wals.) (Lepidoptera: Tortricidae) en Uruguay,” Revista de la Sociedad Entomologica Argentina, vol. 58, pp. 307–311, 1999.
- T. Nagahama, H. Sato, M. Shimazu, and J. Sugiyama, “Phylogenetic Divergence of the Entomophthoralean Fungi: Evidence from Nuclear 18S Ribosomal RNA Gene Sequences,” Mycologia, vol. 87, no. 2, pp. 203–209, 1995. [CrossRef]
- L. G. Leite, S. B. Alves, S. P. Wraight, S. Galaini-Wraight, and D. W. Roberts, “Habilidade de infecção de isolados de Zoophthora radicans sobre Empoasca kraemeri,” Sci Agríc., vol. 53, pp. 152–158, 1996.
- C. Lastra and A. Scorsetti, “Revisión de los hongos Entomophthorales (Zygomycota: Zygomycetes) patógenos de insectos de la República Argentina,” Boletín de la Sociedad Argentina de Botánica, vol. 42, no. 1–2, pp. 33–37, 2007.
- A. W. Guzmán-Franco, S. D. Atkins, P. G. Alderson, and J. K. Pell, “Development of species-specific diagnostic primers for Zoophthora radicans and Pandora blunckii; two co-occurring fungal pathogens of the diamondback moth, Plutella xylostella,” Mycol Res, vol. 112, no. Pt 10, pp. 1227–1240, 2008. [CrossRef]
- A. W. Guzmán-Franco, S. J. Clark, P. G. Alderson, and J. K. Pell, “Competition and co-existence of Zoophthora radicans and Pandora blunckii, two co-occurring fungal pathogens of the diamondback moth, Plutella xylostella,” Mycol Res, vol. 113, no. Pt 11, pp. 1312–1321, 2009. [CrossRef]
- L. F. A. Alves, L. G. Leite, and D. G. P. de Oliveira, “Primeiro Registro de Zoophthora radicans (Entomophthorales: Entomophthoraceae) em Adultos da Ampola-da-Erva-Mate, Gyropsylla spegazziniana Lizer & Trelles (Hemiptera: Psyllidae), no Brasil,” Neotrop. entomol., vol. 38, pp. 697–698, 2009. [CrossRef]
- G. M. Mascarin, V. da S. Duarte, M. M. Brandão, and Í. Delalibera, “Natural occurrence of Zoophthora radicans (Entomophthorales: Entomophthoraceae) on Thaumastocoris peregrinus (Heteroptera: Thaumastocoridae), an invasive pest recently found in Brazil,” Journal of Invertebrate Pathology, vol. 110, no. 3, pp. 401–404, 2012. [CrossRef]
- R. G. Manfrino, L. Zumoffen, C. E. Salto, and C. C. López Lastra, “Potential plant–aphid–fungal associations aiding conservation biological control of cereal aphids in Argentina,” International Journal of Pest Management, vol. 59, no. 4, pp. 314–318, 2013. [CrossRef]
- R. G. Manfrino, A. C. Gutiérrez, D. C. Steinkraus, C. E. Salto, and C. C. López Lastra, “Prevalence of entomophthoralean fungi (Entomophthoromycota) of aphids (Hemiptera: Aphididae) on solanaceous crops in Argentina,” Journal of Invertebrate Pathology, vol. 121, pp. 21–23, 2014. [CrossRef]
- R. Manfrino, J. Hatting, R. Humber, C. Salto, and C. Lastra, “Natural occurrence of entomophthoroid fungi (Entomophthoromycota) of aphids (Hemiptera: Aphididae) on cereal crops in Argentina,” Annals of Applied Biology, vol. 164, pp. 1–8, 2014. [CrossRef]
- A. Ribeiro, H. Silva, E. Castiglioni, S. Bartaburu, and J. J. Martínez, “Control natural de Crocidosema (Epinotia) aporema (Walsingham) (Lepidoptera:Tortricidae) por parasitoides y hongos entomopatógenos en Lotus corniculatus y Glycine max,” Agrociencia (Uruguay), vol. 19, no. 2, pp. 36–41, 2015.
- S. Choi et al., “Occurrence of the Entomopathogenic Fungus Zoophthora radicans (Entomophthorales: Entomophthoraceae) in Jeollabuk-do, Korea,” Korean Journal of Applied Entomology, vol. 56, no. 2, pp. 195–201, 2017. [CrossRef]
- A. P. Gryganskyi, B. A. Mullens, M. T. Gajdeczka, S. A. Rehner, R. Vilgalys, and A. E. Hajek, “Hijacked: Co-option of host behavior by entomophthoralean fungi,” PLOS Pathogens, vol. 13, no. 5, p. e1006274, May 2017. [CrossRef]
- M.-G. Feng, C. Chen, S.-W. Shang, S.-H. Ying, Z.-C. Shen, and X.-X. Chen, “Aphid dispersal flight disseminates fungal pathogens and parasitoids as natural control agents of aphids,” Ecological Entomology, vol. 32, no. 1, pp. 97–104, 2007. [CrossRef]
- A. E. Hajek, B. Papierok, and J. Eilenberg, “Methods for study of the Entomophthorales,” in Manual of Techniques in Invertebrate Pathology (Second Edition), L. A. Lacey, Ed., San Diego: Academic Press, 2012, pp. 285–316. [CrossRef]
- E. Müller-Kögler, “Zur Isolierung und Kultur Insektenpathogener Entomophthoraceen,” Entomophaga, vol. 3, pp. 261–275, 1959.
- L. C. Muskat, Y. Kerkhoff, P. Humbert, T. W. Nattkemper, J. Eilenberg, and A. V. Patel, “Image analysis-based quantification of fungal sporulation by automatic conidia counting and gray value correlation,” MethodsX, vol. 8, p. 101218, Jan. 2021. [CrossRef]
- L. C. Muskat, L. M. Görg, P. Humbert, J. Gross, J. Eilenberg, and A. V. Patel, “Encapsulation of the psyllid-pathogenic fungus Pandora sp. nov. inedit. and experimental infection of target insects,” Pest Manag Sci, vol. 78, no. 3, pp. 991–999, 2022. [CrossRef]
- A. E. Hajek, R. A. Humber, and M. H. Griggs, “Decline in virulence of Entomophaga maimaiga (Zygomycetes: Entomophthorales) with repeated in vitro subculture,” Journal of Invertebrate Pathology, vol. 56, no. 1, pp. 91–97, 1990. [CrossRef]
- J. F. Tabima et al., “Phylogenomic analyses of non-Dikarya fungi supports horizontal gene transfer driving diversification of secondary metabolism in the amphibian gastrointestinal symbiont, Basidiobolus,” G3 Genes|Genomes|Genetics, vol. 10, no. 9, pp. 3417–3433, 2020. [CrossRef]
- K. R. Amses et al., “Diploid-dominant life cycles characterize the early evolution of Fungi,” Proceedings of the National Academy of Sciences, vol. 119, no. 36, p. e2116841119, 2022. [CrossRef]
- K. E. Bushley et al., “The Genome of Tolypocladium inflatum: Evolution, Organization, and Expression of the Cyclosporin Biosynthetic Gene Cluster,” PLoS Genet, vol. 9, no. 6, p. e1003496, 2013. [CrossRef]
- C. de Bekker, R. A. Ohm, H. C. Evans, A. Brachmann, and D. P. Hughes, “Ant-infecting Ophiocordyceps genomes reveal a high diversity of potential behavioral manipulation genes and a possible major role for enterotoxins,” Sci Rep, vol. 7, p. 12508, 2017. [CrossRef]
- K. Seto et al., “A combined microscopy and single-cell sequencing approach reveals the ecology, morphology, and phylogeny of uncultured lineages of zoosporic fungi,” mBio, vol. 14, no. 4, pp. e01313-23, 2023. [CrossRef]
- V. Grigoriev et al., “The Genome Portal of the Department of Energy Joint Genome Institute,” Nucleic Acids Research, vol. 40, no. D1, pp. D26–D32, 2012. [CrossRef]
- A. P. Gryganskyi et al., “Sequencing the Genomes of the First Terrestrial Fungal Lineages: What Have We Learned?,” Microorganisms, vol. 11, no. 7, Art. no. 7, 2023. [CrossRef]
- I. V. Grigoriev et al., “MycoCosm portal: gearing up for 1000 fungal genomes,” Nucleic Acids Research, vol. 42, no. D1, pp. D699–D704, 2014. [CrossRef]
- E. Quesada-Moraga, N. González-Mas, M. Yousef-Yousef, I. Garrido-Jurado, and M. Fernández-Bravo, “Key role of environmental competence in successful use of entomopathogenic fungi in microbial pest control,” J Pest Sci, 2023. [CrossRef]
Figure 1.
Number of EFPSZ species recorded for different countries. Green indicates none reported.
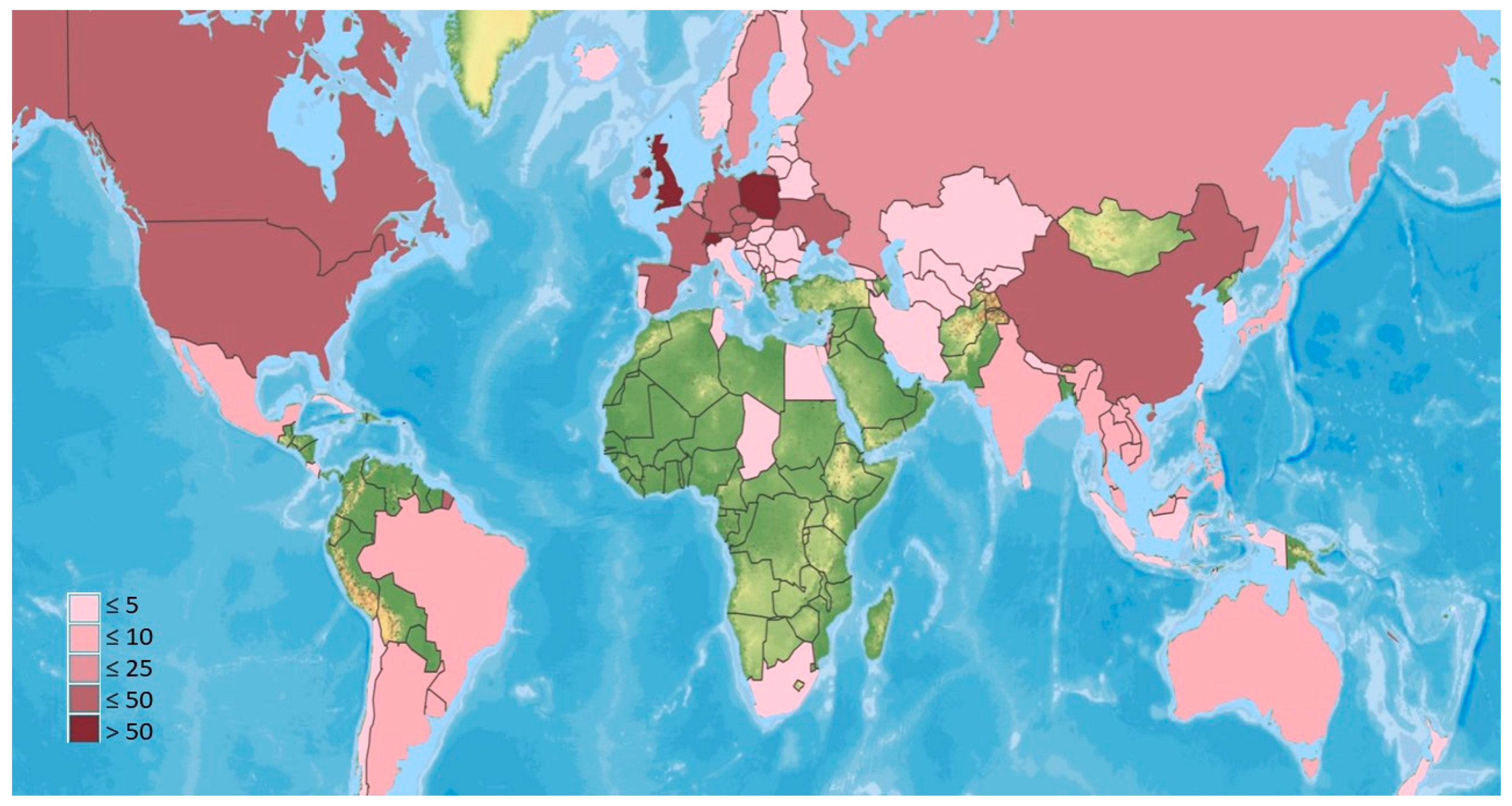
Figure 2.
A phylogenetic tree of species in the genera Erynia, Furia, Pandora, Strongwellsea, and Zoophthora of the subfamily Erynioideae: ~ – aquatic or moist habitats, * – cultivable, generalists in bold, ubiquitous species underlined. Members of the Entomophthoroideae and genus Neoconidiobolus are provided as outgroups.
Figure 2.
A phylogenetic tree of species in the genera Erynia, Furia, Pandora, Strongwellsea, and Zoophthora of the subfamily Erynioideae: ~ – aquatic or moist habitats, * – cultivable, generalists in bold, ubiquitous species underlined. Members of the Entomophthoroideae and genus Neoconidiobolus are provided as outgroups.
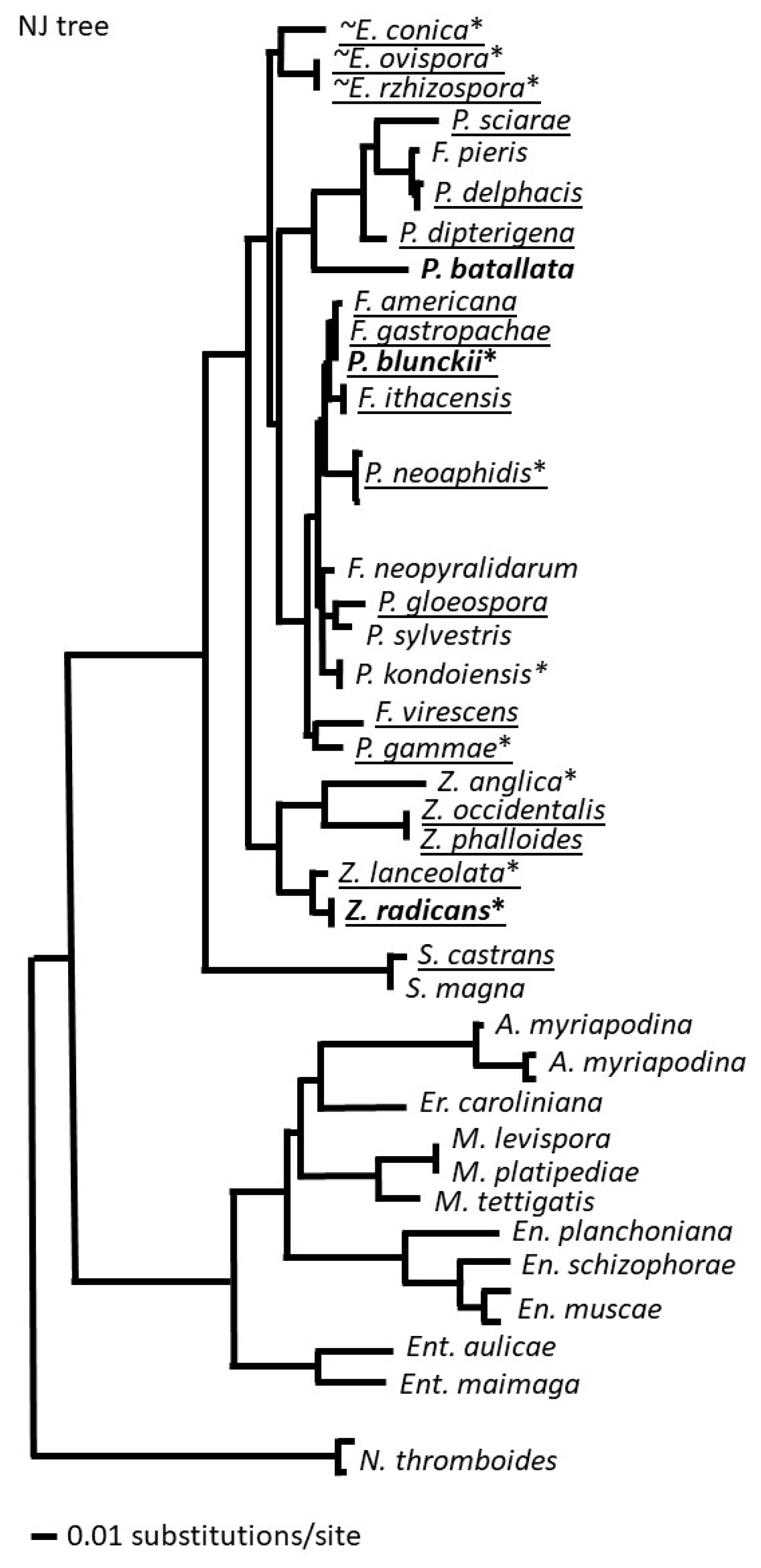
Figure 3.
How many EFPSZ species infect different insect orders. Smaller fractions from left to right: Dermaptera, Trichoptera, Plecoptera, Psocodea, Endognatha, Mesostigmata, Orthoptera, Scatophagidae.
Figure 3.
How many EFPSZ species infect different insect orders. Smaller fractions from left to right: Dermaptera, Trichoptera, Plecoptera, Psocodea, Endognatha, Mesostigmata, Orthoptera, Scatophagidae.
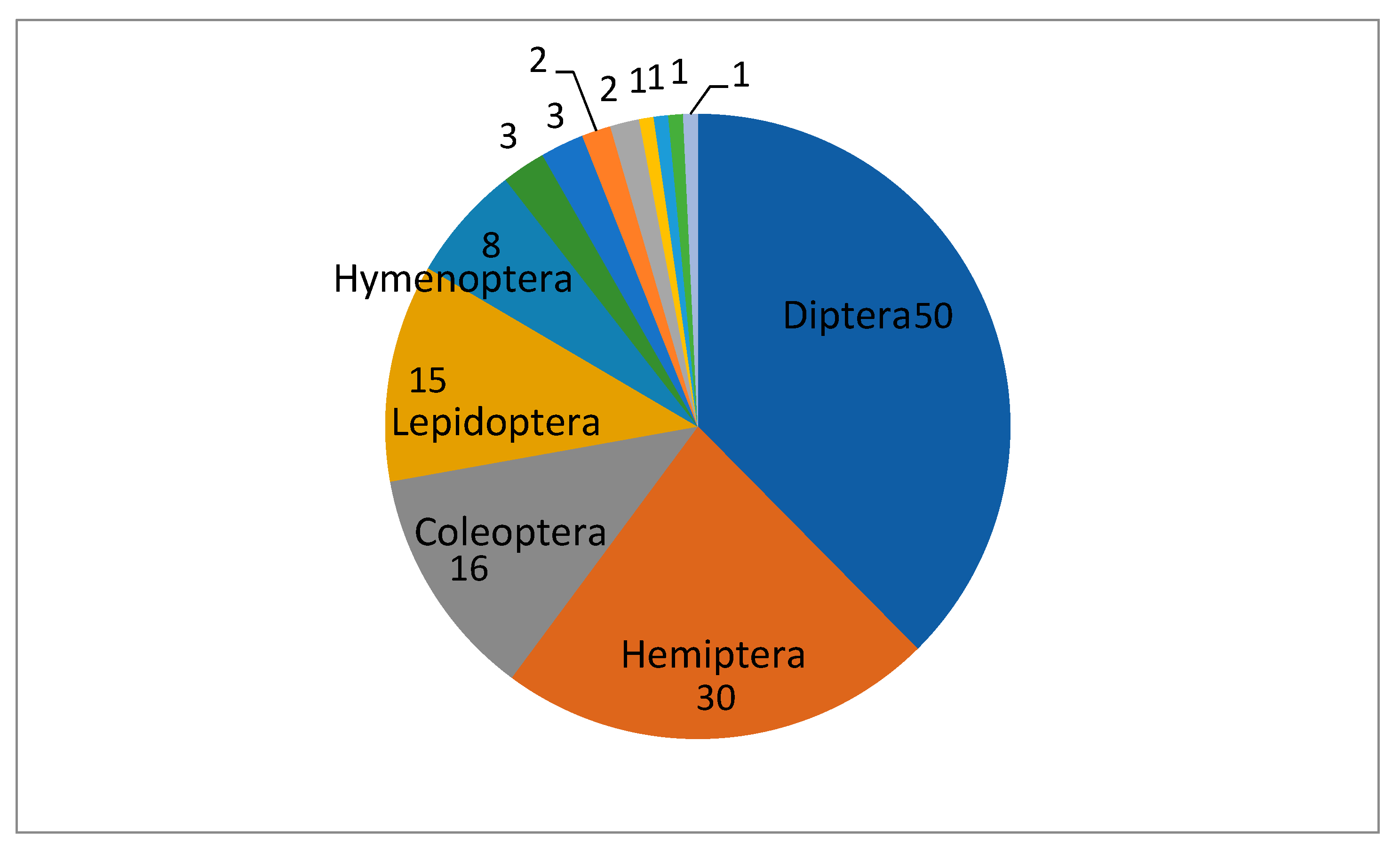

Table 1.
Geographic distributions and insect hosts of EFPSZ fungi.
| Species | Host Order (all Insecta, except as noted) | Host family | Location in details | Reference (based on literature search and ARSEF collection records) |
|---|---|---|---|---|
| ~Erynia aquatica* (2) | &Diptera | Culicidae | Europe: Poland, RF, Spain, Sweden, Switzerland, Ukraine; Nepal; USA | [14,15,16,17,18,19,20,21,22,23] |
| ~E. chironomi | Diptera | Chironomidae | China; Sweden, all of Europe | [15,17,24,25,26] |
| E. cicadellis | Hemiptera | Cicadellidae | Switzerland | [27] |
| ~E. conica* (6) | Diptera, Trichoptera | Chaoboridae, Chironomidae, Culicidae,Psychodidae, Simuliidae, Tipulidae | Asia incl. Israel; Australia; Europe: Poland, RF, Spain, Switzerland, UK, Ukraine; USA | [15,18,19,21,23,28,29,30,31] |
| ~E. curvispora* (3) | &Diptera | Chironomidae, Culicidae, Psychodidae, Simuliidae | China; Europe: Belarus, Estonia, Poland, RF, Switzerland, Ukraine; Israel; NA | [15,18,19,21,26,29,32,33,34] |
| E. delpiniana | Diptera | Muscidae | Italy | [15] |
| E. fluvialis | Diptera | Nematocera | Switzerland | [27] |
| E. gigantea | Hemiptera | Aphrophoridae | China | [35,36] |
| ~E. gracilis | Diptera | Minute gnats | Switzerland; eastern USA | [15,37] |
| ~E. henrici | Diptera | Culicidae | France; Israel | [38] |
| E. jaczewskii | Coleoptera | Carabidae | Ukraine | [21] |
| E. nebriae | Coleoptera | Carabidae | Denmark, Germany | [39] |
| ~E. ovispora* (2) | Diptera | Calliphoridae, Lonchaeidae, Muscidae, Psychodidae, Sarcophagidae, Syrphidae, Tipulidae | Asia: Israel, China, RF; Europe: Austria, Poland, RF, Sakartvelo, Sweden, Switzerland, Ukraine; NA | [15,16,17,18,19,21,29,30,36,40] |
| ~E. plecopteri | Plecoptera | Nemouridae | Europe: Spain, Switzerland, UK; NZ | [15,23,27] |
| ~E. rhizospora (8) | Diptera, Trichoptera | Hydropsychidae, Phryganeidae | China; Europe: Spain, Sweden, Switzerland, UK; USA | [15,18,19,31,41,42] |
| ~E. sepulchralis (1) | Diptera, Trichoptera | Anthomyzidae, Rhagionidae, Syrphidae, Tipulidae | Europe: Poland, Ukraine; USA | [15,21,42] |
| ~E. thurgoviensis | Diptera | Psychodidae | Switzerland | [43] |
| E. tumefacta | Diptera | Muscidae | Switzerland | [27] |
| ~E. variabilis | Diptera | Psychodidae | Europe: Poland, Ukraine, Spain, Sweden, Switzerland; NA | [15,18,19,21,23,31,42] |
| Furia americana (5) | Diptera | Calliphoridae, Muscidae, Sarcophagidae | Brazil; Europe: Italy, Switzerland, UK; USA | [15,18,19,26,27,31,33,42] |
| F. creatonoti | Lepidoptera | Arctiidae, Erebidae | China, Sri Lanka, Taiwan | [15,22,26] |
| F. ellisiana | Dermaptera | Forficulidae | Europe: Poland, Switzerland, UK | [15] |
| F. fujiana | Lepidoptera | Erebidae | China | [26,44] |
| F. fumimontana | Diptera | suborder Brachycera | NA; Poland | [3,15] |
| F. gastropachae (13) | Lepidoptera | Lasiocampidae, Noctuidae | Brazil; China; Europe: Ukraine Spain; NA: Canada, USA | [15,21,23,26,45,46,47] |
| F. ithacensis (4) | Diptera | Empididae, Rhagionidae, Sciaridae | China; Europe: Poland, Spain; USA | [15,23,26,48] |
| ~F. montana | Diptera | Chironomidae | UK; USA | [15,18,19,31,33,42] |
| F. neopyralidarum (1) | Lepidoptera | Erebidae, Piralidae, Tortricidae | Israel, Japan | [15,49,50] |
| F. pieris (1) | Lepidoptera | Pieridae, Zygaeindae | China; NA incl. USA | [15,49,50] |
| F. shandongensis | Dermaptera | Forficulidae | China | [26,51] |
| F. triangularis | Hemiptera | Psyllidae | China, Philippines | [26,52,53] |
| F. virescens (4) | Lepidoptera | Noctuidae | Asia: China, Turkmenistan; Europe: Czech, Finland, Germany, Poland, RF, Spain, Switzerland, UK, Ukraine; NA | [15,18,19,23,30,31,32,33,42,54,55] |
| F. vomitoriae | Diptera | Calliphoridae, Stratiomyiidae, Syrphidae | Europe: Austria, Czech, Poland, RF; Mexico | [15,40,56] |
| F. zabri | Coleoptera | Carabidae | Europe: Czech, Ukraine; Uzbekistan | [15,21] |
| Pandora aleurodis | Hemiptera | Aleyrodidae | Romania | [15] |
| P. batallata | Entognatha, Symphypleona | Sminthuridae | Germany | [57] |
| ~P. bibionis | Diptera | Bibionidae, Sciaridae | China; Switzerland | [26,43,51] |
| P. blunckii (34) | Hymenoptera, Lepidoptera | Plutellidae, Tenthredinidae, Tortricidae | Asia: Israel, China, Japan, Philippines; Australia; Europe (not reported in Poland); Mexico | [15,18,19,26,41,58,59,60,61,62,63] |
| P. borea | Diptera | Calliphoridae, Muscidae, Sarcophagidae | China | [26,64] |
| P. brahminae | Coleoptera | Scarabaeidae | Bharat, China | [15,26] |
| P. bullata | Diptera | Calliphoridae, Sarcophagidae | Brazil; Australia; Europe: Spain, Switzerland, UK; Iran; NA: USA, Canada; SА | [15,18,19,23,27,65,66,67,68,69] |
| P. cacopsyllae* | Hemiptera | Psyllidae | Denmark | [70] |
| P. calliphorae | Diptera | Anthomyiidae | China; France | [26,71] |
| ~P. chironomid | Diptera | Chironomidae | China | [24] |
| P. cicadellis | Homoptera | Cicadellidae | China | [26,52] |
| P. dacnusae | Hymenoptera | Braconidae | Poland | [15] |
| P. delphacis (64) | Hemiptera | Delphacidae | Asia: Bharat, China, East Asia, Indonesia, Japan, Philippines; SA: Brazil, Argentina; Switzerland; USA | [15,26,43,62,63,72,73,74,75,76] |
| ~P. dipterigena (8) | Diptera | Calliphoridae, Muscidae, Mycetophilidae,Psychodidae,Rhagionidae, Sciaridae, Syrphidae, Tachinidae, Tipulidae | Asia: Bharat, China, Indonesia, Iran, Israel; Brazil; Europe: Austria, Poland, RF, Sakartvelo, Spain, Sweden, Switzerland, UK, Ukraine; NA: Mexico, USA | [15,16,17,21,23,26,30,31,33,40,41,42,68,77,78,79,80,81,82] |
| P. echinospora | Diptera, Hemiptera | Aphididae, Formicidae, Lauxaniidae | Asia: China, Israel; Europe: Austria, Poland, Sakartvelo, Spain, Sweden, Switzerland, UK, Ukraine; NA: Costa Rica, USA | [15,16,21,23,26,27,29,33,40,42,58,67,83] |
| P. formicae | Hymenoptera | Formicidae | Bharat; Europe incl. Denmark | [15,84,85] |
| P. gammaeI* | &Lepidoptera | Erebidae, Noctuidae | Asia: China, Israel, Turkmenistan; Australia; Europe: Poland, RF, Slovakia, Switzerland, Ukraine; NA incl. Mexico; SA: Argentina, Brazil | [15,18,19,26,30,32,56,66,79,86,87,88,89,90,91,92] |
| ~P. gloeospora | Diptera | Mycetophilidae, Psychodidae, Sciaridae | China; Europe: France, Ukraine; USA | [13,15,21,22,26,93] |
| P. guangdongensis | Hemiptera | Miridae | China | [94] |
| P. heteropterae (1) | Hemiptera | Miridae | NA incl. USA; Poland | [15,95] |
| P. kondoiensis* (5) | Hemiptera | Aphididae | Australia; China | [15,26,64,96] |
| P. lipae | Coleoptera | Cantharidae | Denmark, France, Poland, Switzerland | [15,19] |
| ~P. longissimi | Diptera | Limoniidae | Switzerland | [27] |
| P. minutispora | Hemiptera | Miridae | Czechia, Switzerland | [15,18] |
| P. muscivora | Diptera | Calliphoridae, Drosophilidae, Muscidae, Tachinidae | Canada; Europe: Poland, UK, Ukraine | [15,21,33] |
| P. myrmecophaga | Hymenoptera | Formicidae | Brazil; Europe: Czechia, Germany, Poland, Sweden, Switzerland, Ukraine, former Yugoslavia; Philippines | [15,18,19,21,63] |
| P. neoaphidis* (TYPE, 173) | Hemiptera | Aphididae | Worldwide, less frequent in tropics: Africa: Egypt, Tunisia, South Africa; Asia: Bharat, China, Iran, Israel, Japan, Korea, Nepal, Philippines, Taiwan; Australia; Europe: Austria, Bosnia & Hercegovina, Denmark, Finland, France, Iceland, Latvia, Poland, Portugal, RF, Serbia, Slovakia, Spain, Switzerland, UK, Ukraine; NA: Canada, Mexico, USA; NZ; SA: Argentina, Brazil, Chile, Uruguay | [15,18,19,20,21,26,29,30,40,51,54,56,58,61,63,68,77,78,82,96,97,98,99,100,101,102,103,104,105,106,107,108,109,110,111,112,113,114,115,116,117,118] |
| P. nouryi* (5) | Hemiptera & Psocodea | Aphididae, Pseudocaeciliidae | Argentina; Asia: China, Israel; Australia; Europe: Central, Northern & Western, incl. Slovakia; NA | [15,26,51,58,78,100,109,110,112,119,120] |
| P. phalangicida | Mesostigmata & Opiliones (Arachnida) | Parasitidae | Poland, Sweden, UK | [15,33] |
| P. philonthi | Coleoptera | Staphylinidae | Denmark, Poland, Switzerland | [15,27,121] |
| P. phyllobii | Coleoptera | Curculionidae | Poland | [15] |
| P. polonae-majoris | Hemiptera | Cicadellidae, Jassidae | Poland, Ukraine | [15,21] |
| P. psocopterae | Psocodea | Prionoglarididae | France | [15,18,19] |
| ~P. sciarae | Diptera | Sciaridae | Europe: Austria, Denmark, Switzerland, Ukraine; NZ; USA | [15,21,27,31,40] |
| P. shaanxiensis | Diptera | Calliphoridae | China | [24,26] |
| P. sylvestris sp. nova | Lepidoptera | Erebidae | USA | Hajek & Gryganskyi, in press |
| P. terrestris | Hemiptera | Aphididae | Ukraine | [119,122] |
| P. uroleuconii | Hemiptera | Aphididae | Slovakia | [123] |
| Strongwellsea acerosa | Diptera | Muscidae | Europe | [124] |
| S. castrans (2) | Diptera | Anthomyiidae | China; Europe: Czechia, Denmark, Switzerland, UK; USA | [26,59,125,126] |
| S. crypta | Diptera | Anthomyiidae | Denmark | [127] |
| S. gefion | Diptera | Muscidae | Europe | [128] |
| S. magna | Diptera | Muscidae | China; USA | [36,129] |
| S. pratensis | Diptera | Muscidae | Switzerland | [27] |
| S. selandia | Diptera | Muscidae | Europe | [128] |
| S. tigrinae | Diptera | Muscidae | Europe | [124] |
| Strongwellsea sp. nov. | Diptera | Calliphoridae | Europe | [130] |
| Strongwellsea sp. nov. | Diptera | Sarcophagidae | Europe | JE, unpublished |
| Strongwellsea sp. nov. | Diptera | Scatophagidae | Europe | [131] |
| Strongwellsea sp. nov. | Diptera | Anthomyiidae | Europe | JE et al., unpublished |
| Zoophthora anglica* (5) | Coleoptera | Elateridae | Denmark, France, Poland, Romania, Switzerland, UK, Ukraine | [15,21,132] |
| Z. anhuiensis | Hemiptera | Aphididae | China | [15,26,61,133] |
| Z. aphidis* (1) | Hemiptera, Lepidoptera | Aphididae, Cicadellidae, Delphacidae, Erebidae | Asia: China, Philippines, Taiwan; Europe: Armenia, Belarus, Estonia, Lithuania, Moldova, RF, Sweden, UK, Ukraine; NA: Canada, Puerto Rico, USA | [17,21,30,31,33,105,111,134,135,136] |
| Z. aphrophorae | Hemiptera | Aphrophoridae, Cicadellidae, Miridae, Psyllidae | UK | [33] |
| Z. arginis | Hymenoptera | Argidae | Germany, Poland | [15,28] |
| Z. athaliae | Hymenoptera | Tenthredinidae | China; Switzerland | [15,26,27,64] |
| Z. autumnalis | Diptera | Dryomyzidae | Poland | [15] |
| Z. bialovienzensis* | &Lepidoptera | Geometridae, Pyralidae | Poland, Ukraine | [15,21] |
| Z. brevispora | Lepidoptera | Geometridae | Poland | [15] |
| Z. canadensis | Hemiptera, Lepidoptera | Aphididae, Geometridae | China; NA | [137,138] |
| Z. crassispora | Lepidoptera | Tortricidae | Poland | [15] |
| Z. crassitunicata | Coleoptera | Cantharidae | Austria, Switzerland | [15,27,40] |
| Z. elateridiphaga | Coleoptera | Elateridae | Switzerland | [19] |
| Z. erinacea | Hemiptera | Aphididae | Israel; Slovakia | [15,58,78,119] |
| Z. falcata | Hymenoptera | Formicidae | Poland | [15] |
| Z. forficulae | Dermaptera | Forficulidae | Europe: Poland, Switzerland, UK; NA | [15,31,33,43] |
| Z. geometralis | &Lepidoptera | Geometridae, Yponomeutidae | Europe: Austria, Sweden, Ukraine; NA | [15,16,21,40,42] |
| Z. giardia | Orthoptera | Tettigoniidae | France, Germany, Poland | [15] |
| Z. humberi | Diptera | Tipulidae | Chile | [15,139] |
| Z. ichneumonis* | &Hymenoptera | Ichneumonidae | Poland, Switzerland, Ukraine | [15,21,27] |
| Z. independentia | Diptera | Tipulidae | USA | [140] |
| Z. lanceolata* (3) | Diptera | Drosophilidae, Empididae | Europe: France, Poland, Spain, Switzerland, Ukraine; Israel | [15,21,23,29,141] |
| Z. larvivore | Coleoptera | Cantharidae | Poland | [15] |
| Z. miridis | Hemiptera | Miridae | Poland, Spain, Switzerland | [15,23,141] |
| ~Z. nematoceris | Diptera | Bibionidae, Sciaridae | Poland, Spain, Switzerland | [15,23,141] |
| Z. obtuse | Diptera | Brachycerous or Calyptrate | Poland, Switzerland | [15,43] |
| Z. occidentalis (7) | Hemiptera | Aphididae | Asia incl. China; Europe: Poland, Slovakia, Spain, Switzerland, UK; NA; SA incl. Chile | [15,23,27,31,33,77,78,102,103,139,142,143] |
| Z. opomyzae | Diptera | Opomyzidae | Austria, Germany, Poland | [15,28,40] |
| Z. orientalis | Hemiptera | Aphididae | Israel | [29] |
| Z. pentatomis | Hemiptera | Pentatomidae | China | [26,51,52,137] |
| Z. petchii | Hemiptera | Aphrophoridae, Cercopidae, Cicadellidae, Delphacidae | Asia: China, Israel; Europe: Austria, Switzerland | [15,27,28,29,73] |
| Z. phalloides (2) | Hemiptera | Aphididae | Argentina; Asia: Israel Korea; Europe: Germany, Poland, Slovakia, Switzerland, UK; Australia NZ; NA incl. Mexico | [15,18,19,28,29,77,78,82,98,106,144,145,146] |
| Z. phytonomi* | &Coleoptera | Curculionidae | Asia: Israel, Uzbekistan; Australia; Europe: Poland, Romania, Ukraine; USA | [15,21,29,147] |
| Z. porteri | Diptera | Tipulidae | Ukraine; USA | [21,140] |
| Z. psyllae | Hemiptera | Psyllidae, Triozidae | Poland, Spain, Switzerland | [15,23,27] |
| Z. radicans* (305) | Diptera, Coleoptera, Hemiptera, Homoptera, Hymenoptera, &Lepidoptera, Plecoptera | Agromyzidae, Aphididae, Aphrophoridae, Aleurodidae, Argidae, Chironomidae, Chrysomelidae, Cicadellidae, Crambidae, Delphacidae, Geometridae, Miridae, Muscidae, Nemouridae, Pentatomidae, Pieridae, Plutellidae, Psyllidae, Triozidae, Thaumastocoridae, Tortricidae | Africa: South Africa, Tchad; Asia: China, Indonesia, Israel, Japan, Korea, Kyrgyzstan, Malaysia, Philippines; Australia, NZ; Europe: Belarus, Denmark, Estonia, France, Moldova, Poland, RF, Sakartvelo, Serbia, Slovakia, Sweden, Switzerland, UK, Ukraine, former Yugoslavia; NA: Canada, Cuba, Mexico, Puerto Rico, USA; SA: Argentina, Brazil, Uruguay | [15,16,18,19,21,26,29,30,31,33,51,56,60,61,63,77,78,82,91,92,94,112,113,115,121,137,146,148,149,150,151,152,153,154,155,156,157,158,159,160,161,162,163,164,165] |
| Z. rhagonycharum | Coleoptera | Cantharidae | Europe: Denmark, Poland, Switzerland; NA | AEH, person. observations, [15,27] |
| Z. suturalis | Coleoptera | Chrysomelidae | France, UK | [15] |
| Z. tachypori | Coleoptera | Staphylinidae | Poland | [15] |
| Z. viridis | Hemiptera | Miridae | Western Europe incl. Germany, Switzerland | [15,19] |
| Total 123 species; in ARSEF – 683 specimens of 28 species | Total 14 orders | Total 76 families | Total 57 countries, in ARSEF specimens from 29 countries. |
Continents and countries placed alphabetically, ~ - aquatic or moist habitats, * - cultivable, & - infects other or more stages than adult, in parentheses number of specimens in ARSEF culture collection, NA – North America, RF – Russian federation, SA – South America, UK – United Kingdom.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



