
Submitted:
11 December 2023
Posted:
12 December 2023
You are already at the latest version
Abstract
Accurate and robust methods are needed to monitor crop growth and predict grain yield and quality in breeding programs under variable agrometeorological conditions. Field experiments were conducted during two successive cropping seasons (2021, 2022) at four trial locations (Estonia, Latvia, Lithuania, Norway). The focus was on assessment of grain yield (GY), grain protein content (GPC), and UAV-derived NDVI measured at different plant growth stages. Performance and stability of 16 selected spring wheat genotypes were assessed under two N application rates (75, 150 kg N ha−1) and over contrasted agrometeorological conditions were assessed, and the quantitative relationships between agronomic traits and UAV-derived variables were figured out. None of the traits were subject to a significant (p<0.05) genotype by nitrogen interaction. High-yielding and high-protein genotypes were detected with high WAASB stability, specifically under high and low N rates. This study highlights the significant effect of NDVI analysis on growth stages GS55 and GS75 as key linear predictors, especially in the context of spring wheat GY. However, the effect of these indices depends on the specific growing conditions in the respective locations, thus limiting their universal utility.
Keywords:
genotype
; meteorological parameters
; nitrogen fertilization rate
; multispectral vegetation index
; correlations
1. Introduction
Common wheat (Triticum aestivum L.) is a valuable and extensively cultivated cereal worldwide. Global wheat production has surged from 715 million tons in 2015 to 780 million tons in 2021, with Europe appearing as the second-largest producer of bread wheat [1]. The importance of wheat reaches the Baltic countries (Latvia, Estonia, Lithuania) and the Nordic nations (Denmark, Norway, Sweden, Finland), where wheat production collectively accounts for approximately 7% and 5% of the total European wheat production, respectively [1,2].
The climatic conditions in the Nordic-Baltic region allow for the cultivation of both types of wheat – spring and winter wheat. Notably, most of the harvested area in Baltic countries is dedicated to winter wheat, with only about 20% of the area distributed to spring wheat cultivation [3]. Given that a substantial part of spring wheat production in the Baltic countries and Norway is earmarked for breadmaking, breeding programs in this region place simultaneous emphasis on traits related to productivity and baking quality, including grain yield (GY) and grain protein content (GPC) [4,5,6,7]. Wheat having a moderate-to-high GPC (exceeding 12%) is considered essential for bread making, while wheat with lower GPC levels is typically used for cookies, noodles, or animal feed [8].
GY and GPC are complex quantitative characteristics controlled by multiple genes, influenced by genotype, environment, and genotype - environment interactions. Breeders often rely on grain yield and quality, along with their contributed performance and phenotypic expression, when selecting crop cultivars under mega-environment tests [9]. Abiotic stresses, such as water stress, heat, and nutritional deficiency, can have negative impacts on crop yield and quality [10,11]. Therefore, each genotype should be evaluated for several years in different locations in order to identify high performing and stable genotypes and recommend them for cultivation in target areas.
Wheat, as a cereal crop, has a high demand for nitrogen (N), crucial for plant growth, development, GY formation, and GPC accumulation, essential for baking and processing quality. Wheat growth has an absolute requirement for N, and crop yield and quality rely heavily on substantial N inputs [12].
Crop breeding needs to be intensified and made more efficient, using new phenomics methods to predict GY and GPC, both of which are interrelated parameters [13]. Consequently, remote sensing is gradually gaining recognition as an alternative to traditional, destructive field sampling methods and laboratory testing, serving as a tool for finding the nitrogen (N) status in wheat [14]. Unmanned aerial vehicle (UAV) platforms equipped with multiple sensors, capable of swiftly scanning entire trials, prove to be useful tools for getting non-destructive data on the physiological state of plant canopies [15,16]. Multispectral data have been analysed for UAV-based estimation of wheat grain yield under different water and nitrogen conditions and at various wheat growth stages [17,18].
The Normalized Difference Vegetation Index (NDVI), calculated from multispectral data, stands out as the most popular index for vegetation assessment [19]. This index finds utility in various fields, including agriculture [20]. NDVI is valuable in estimating biomass [21], showing plant stress [22], finding chlorophyll concentration [23], and other characteristics. Numerous studies on wheat have shown the potential of using NDVI readings as a tool to differentiate and find high-yielding wheat genotypes, even in different nitrogen environments [24,25,26].
There is a lack of published results on the agronomic and UAV remote sensing phenotyping of spring wheat genotypes in multi-environment experiments, particularly within the Baltic region’s unique conditions, offering valuable insights for breeding programs. The aim of this study was to identify best performing and stable spring wheat genotypes for grain yield and grain protein content in different Nordic-Baltic environments.
The objectives of this study were:
- assess the variation in GY, GPC and UAV-derived NDVI across different years and nitrogen fertilization rates in the Nordic-Baltic region;
- evaluate the performance and the stability of spring wheat genotypes concerning GY and GPC under varying N fertilization rates and diverse agrometeorological conditions;
- explore the quantitative relationships between GY, GPC, and UAV-derived NDVI variables across multi-environmental trials during different plant growth stages.
Using UAV phenotyping technologies combined with field evaluations, we intended to determine whether superior genotypes can be accurately found with this methodology, and at which stages of plant development assessment should be done.
2. Materials and Methods
2.1. Plant materials
In this experiment, sixteen spring wheat genotypes were used, which include cultivars and improved breeding lines, mostly developed in the last decade in the spring wheat breeding programs of the Baltic States and Norway. These programs include the Agricultural Resources and Economics Institute (AREI, Latvia, LV), the Lithuanian Agricultural and Forestry Research Centre (LAMMC, Lithuania, LT), the Estonian Rural Research and Knowledge Centre (METK, Estonia, EE), as well as the Norwegian University of Life Sciences (NMBU, Norway, NO) and Graminor (Norway). Detailed information on the selected spring wheat genotypes, their origin and descriptions are given in Table S1.
2.2. Field trials
Field experiments were carried out over two consecutive years (2021 and 2022) at four geographically distant locations representing different agro-ecological conditions and soil types in the Nordic and Baltic region: at Centre of Estonian Rural Research and Knowledge (Jogeva, Eastern part of Estonia, 58°76’ N, 26°24’ E), at Stende Research Centre (Dižstende, North-West of Latvia, 57°18’ N, 22°56’ E), at Institute of Agriculture (Dotnuva, in central Lithuania, 55°39’ N, 26°24’ E), and at Vollebekk Research Station (Ås, south-eastern Norway, 59°39’ N, 10°45’ E). The physical and chemical properties of the experimental soils prior to sowing are detailed in Table S2.
In each location spring wheat genotypes were sown in the field trials set up at two contrasting nitrogen (N) fertilization rates applied before sowing: 75 and 150 kg ha-1 N of compound NPK fertilizer, referred to as low N (N75) and high N (N150) rates, respectively. The high N treatment is a typical fertilization rate for spring wheat in Norway and the Baltic states. The low N treatment reflects less intensive management, allowing to evaluate the performance of genotypes at reduced N supply. The experiment was designed in split plots, with the main plot being the N rates (N75, N150) with sub-plot being the wheat genotypes.
The plot size in various locations was 9-15 m2 with seeding rate of 500 kernels m-2 in LV and LT, and 600 kernels m-2 in EE and NO. Trials were sown at the proper time, from 21st of April to 2nd of May depending on local meteorological conditions in each location. Treatment of trials with pesticides included the application of herbicides and fungicides in all locations based on standard procedures. Foliar fungicide 1-2 treatments were applied at each location, but insecticides were used when the control thresholds of insects were exceeded. Following the ripening stage, the trial material was harvested using a combine.
Daily weather data (precipitation amount, mm; °C; average day temperature, °C) were downloaded from services provided by research institutions. The climate normal or long-term average (LTA, 1991–2020) was used to compare two-year data at each location.
2.3. Phenotyping of field traits
Days to maturity (DM, days) were calculated from visually recorded maturity dates when 50% of the plants were in the respective developmental stage. Plant height (PH, cm) was measured manually at the ripening stage in three separate places of the plot and calculated as the average height of a sample of a fertile stem, from soil surface to the top of the spike. Grain yield (GY, g m-2) was recorded from each experimental plot and adjusted to a moisture content of 14%. Grain protein content (GPC, %) was measured using a near-infrared reflectance spectrometer (Perten Inframatic 9200/Perten Instrument AB) in Norway (NO), and near-infrared transmittance whole grain analysers were used in Estonia (EE) (InfratecTM/Foss), Latvia (LV) (InfratecTM NOVA/Foss), and Lithuania (LT) (Infratec 1241/Foss).
2.4. UAV data acquisition and processing
DJI Phantom 4 Multispectral quadcopter (DJI, Inc., Shenzhen, Guangdong, China) was used to acquire image data in LV, LT, and NO. This commercial UAV model is commonly used in agronomy research and monitoring [27]. DJI Phantom 4 RTK (Real Time Kinematics) quadcopter (DJI, Inc., Shenzhen, Guangdong, China) carrying Micasense RedEdge MX multispectral camera (AgEagle Aerial Systems, Seattle, Washington, USA) was used to acquire image data in EE.
Multispectral UAV images were captured at each location from an altitude of 20 meters, resulting in a ground sample distance (GSD) of 1 cm. The images were taken with a minimum of 75% overlap in the front and side directions, employing a 90-degree (NADIR) camera position. Each flight mission generated approximately 5000 TIFF images, with five images acquired for each shot.
Four UAV flight missions were planned in each of eight field trials, corresponding to certain time of plant vegetative and generative development based on Zadoks growth scale [28]: the middle of vegetative wheat plant development (tillering, growth stage 25/GS25), spike emergence or heading (GS55), the middle of grain filling (milk development stage, GS75), and late grain filling (dough development stage, GS85) (Table 1). However, the developmental stages differed slightly between genotypes due to differences in phenological development.
The result was a series of average reflectance values of the field trial plots in each of the five spectral bands (red, green, blue, red-edge, near-infrared). Aerial images were processed with PIX4Dmapper by Pix4D (Lausanne, Switzerland) photogrammetry software for drone mapping. Processed orthophotos were georeferenced with an average RMSE of 3-5 cm. To keep this precision and compatibility between data series taken in different time periods, Ground Control Points (GCP) were set up, measured and kept during the entire vegetation season.
Spectral reflectance values were extracted from final orthophoto mosaics, which were cropped to field boundaries. The zonal statistics tool in QGIS was used to extract mean reflectance values from the orthophoto images, which served as the input for the calculation of NDVI index values. The NDVI was calculated in QGIS based on a combination of visible RED and NIR wavelengths with the following Equation (1):
where RNIR is the reflectance in the near-infrared band, Rred. is the reflectance in the red band.
NDVI = (RNIR-Rred)/(RNIR+Rred),
2.5. Data statistical analysis
The data of 16 genotypes in each of 16 environments (4 locations, 2 years, 2 N rates) was initially investigated using descriptive analysis: maximum and minimum values, variance, standard deviation, and mean. All derived measurements were calculated from the individual plot values.
For split-plot trial design the Linear Mixed-Effects Model ANOVA was applied. In the first model two main factors genotype, N rate and their interaction were fixed, and field block was random; calculations were carried out separately for data of 8 environments (combination of location and year). The partitioning of the percentage of factors was computed from the total sum of the square (TSS, %). In the second model three main factors: genotype, year, N rate and their interactions were fixed, and field block was random; calculations were carried out separately for data of each location. Mean comparisons between genotypes, trial years and N rates were performed using Fisher’s least significant difference (LSD) (p<0.05).
The multivariate stability analysis was performed based on the additive main effects and multiplicative interaction model (AMMI), combining the ANOVA for the genotype and environment main effects with the principal components analysis of genotype x environment interaction [29]. Each year-location combination was considered as an environment; therefore, eight environments were accounted for stability analysis of 16 genotypes under high N and low N rates.
Stability analysis was carried out using WAASB index [30]. The index was calculated as follows (2):
where WAASBi is the weighted average of absolute scores of the i-th genotype; IPCAik is the score of the i-th genotype in the k-th IPCA (interactions principal component axis), and EPk is the amount of the variance explained by k-th IPCA. Genotype with lowest WAASB score deviates least from the mean performance across environments and is therefore considered as most stable.
Pearson correlation analyses were performed between GY, GPC and NDVI values at four plant growth stages in each of 8 environments in high and low N rates separately. We used linear regression models to test the relationships between NDVI values at each plant growth stage (as independent variables) and GY and GPC (as dependent variables). We reported adjusted determination coefficients (adj. R2), root mean squared error (RMSE) and p-values to characterize regression models.
Data results were visualized with MS Excel extension Real Statistics and R 4.2.3 statistical program [31] packages lme4, datasets, agricolae, metan.
3. Results
3.1. Variation of meteorological data
Overall meteorological conditions were markedly different between the trial years of 2021 and 2022. However, in comparison to the 30-year long-term average (LTA) data (1991-2020), there are both similar trends and differences between environments (Figure 1). In 2021, April, May and August experienced lower average daily air temperatures than the LTA in all trial locations, but in June and July, it exceeded the average annual observations by 1.7 to 4.0°C and 1.5 to 4.2°C, respectively (Fig., a). In April, May and June, in all test locations, and in July in EE and LT locations, the amount of precipitation was significantly reduced (Fig., b). In the 2022 season, April and May were generally relatively cool, but in June, the average daily air temperature in all trial locations exceeded the LTA (Figure 1a). In June, the monthly rainfall was reduced in the trials of NO, EE and LV. Heavy rains occurred in LT in June, when 256% of precipitation compared to the LTA was noticed (Figure 1b).
Overall, the growing season of 2022 was considered as more favourable season for wheat growth and productivity than the earlier one due to lower average temperatures and higher rainfall.
3.2. Variation of traits by locations and years
Results of Analysis of variance performed for GY and GPC data showed that in all location the effect (sum of squares) of different years were highly significant (p<0.001) with the highest contribution on both traits’ variability (Table S2). Mean GY values varied significantly by year in each location, with the lowest yielding trial location-year in EE in 2021 (466.8 g m-2), and the highest yielding location-year in LV in 2022 (622.2 g m-2) (Table 2). When comparing the two trial years, the average grain yield was significantly higher in 2022 than in 2021 in three locations (NO, LV, LT), while in LT, it was the opposite.
Mean GPC values also varied substantially from one location-year to another. The mean GPC values among four locations ranged from 11.7 to 15.6% in 2021 and from 10.9 to 14.5% in 2022, with the lowest value in LT in 2022 (10.9%), and the highest value in LV in 2021 (15.6%). Differences were observed between locations on GPC depending on the trial year. In LV, EE, and LT, significantly higher GPC was obtained in 2021, but not in NO, where it was, on contrary, 2.1% higher in 2022.
UAV-derived NDVI obtained at four growth stages showed also significant variation among trial years (Table 3).
Overall, in all four growth stages, mean value differences varied significantly and did not show consistent similar variation compared to data obtained between years. For instance, in GS55, when comparatively the highest mean NDVI values were detected, in LV and LT this trait was significantly higher in 2022, while in EE it was higher in 2021, but in NO in both trial years we obtained similar NDVI values.
3.3. The impact of N rate on phenotypic variation of traits
As shown in Figure 2, the impact of the high N rate was pronounced for GY at all trial locations; significantly higher GY in such conditions were obtained in EE, LV and NO (Figure 2a; Table S4). Opposite results were detected in LT, where in 2021 significantly (p<0.001) higher average GY values were obtained under low N conditions, but in 2022 N treatment did not supply a significant effect. In LV and NO, a higher N regime response to GY variation was detected in 2021 compared to 2022.
High N fertilization rate promoted the formation of significantly (p<0.001) higher GPC at all locations (Figure 2b; Table S4). Significantly higher GPY was harvested under high N fertilization rate in all trial locations, except in 2021 in LT where this trait did not show significant difference (Figure 2c).
The N rate showed a significant effect on the variation of NDVI values measured at the tillering stage (GS25) only in three environments (EE_2021; NO_2021, LT_2022) (Table 4). In all locations, NDVI evaluated during heading (GS55) and milk (GS75) growth stages, and in most cases significantly higher NDVI values were obtained under high N supply compared to those obtained in low N conditions. At all locations the NDVI variation was significantly affected by the N fertilization regime also in the late grain filling stage (GS85), with significantly (except in EE_2022) higher values under high N application.
Analysis of variance confirmed that in all locations-year environments genotype by nitrogen (G x N) interaction was not significant for any of the investigated traits (Table S4).
3.4. Mean performance and stability of wheat genotypes under contrasting environments
The mean performance of GY and GPC, their corresponding WAASB stability indices and trait ranking values under high and low N rates for spring wheat genotypes are presented in Table 5. Genotype effects were significant for investigated traits. Three spring wheat genotypes were detected as top-yielding under both low N (GY 547.4-564.1 g m-2) and high N (GY 592.5-609.5) rates, confirming the fact of the lack of significant G x N interaction effect. Overall, for GY, the ranking order of WAASB values for genotypes differed between the N rates. Genotypes DS-655-7-DH and DS-720-3-DH showed below-average GY stability ranking by WAASB over all investigated environments. In turn, the top-yielding DS-17-16-DH showed the best stability only under low N rate. Genotype 990-2 in the opposite ranked 1st by WAASB under N150, but only ranked 13st under N75. The variety Betong showed comparatively good GY results and stability under both high and low N. The other genotypes, characterized by high stability ranking by WAASB in both N rates showed below-average yield levels. The genotype Zombi had the same stability rank across both N rates.
GPC among different genotypes varied from 11.7 to 14.0% under low N, and from 13.2 to 16.0% under high N. The results showed that genotypes 876 and Runar, with the lowest yield over investigated environments, formed the top-highest (1st; 2nd) GPC in both N rates. However, in both N rates, these genotypes exhibited one of the lowest stability rankings by WAASB among the 16 tested genotypes. The top-yielding genotype DS-17-16-DH formed one of the lowest GPC under both N rates, where ranking of WAASB stability values between both N rates distinctly differed. Voore simultaneously combined above-average GPC (13.1 and 14.7%, under N75 and N150, respectively) and showed a good stability ranking (4th) under high N. Overall, the top-ranking of WAASB values for GPC were detected for genotypes (013-074, 990-2, and Hiie, all grown under high N) with GPC corresponding close to this trait average values.
Genotypes DS-17-16-DH, DS-655-7-DH, DS-720-3-DH, 013-01 and DS-638-5-DH were top-high yielding over all investigated environments, and characterized comparatively by the longest average growing period (DM 106.9-108.6 days) among the 16 tested genotypes (Table 6). All top-yielding genotypes, also obtained the highest NDVI values in GS75 and GS85. Top-yielding genotypes DS-17-16-DH and DS-638-5-DH were characterized with comparatively short PH, but for DS-655-7-DH and DS-720-3-DH this trait was above average value. Wheat genotypes 876 and Runar being in the top of GPC rank, were characterized by the comparatively tallest plants (4th, 1st in rank, respectively), short DM (102.8 and 103.1 days, respectively) and with the lowest NDVI values in all growth stages. Different patterns in terms of NDVI profiles between genotypes were observed. For instance, for DS-655-7-DH comparatively high values of NDVI were detected in all investigated growth stages, but high-protein cultivar Runnar, one of the highest NDVI values showed only in GS25 (Table 6).
3.5. Correlations among NDVI and GY, GPC
As the above-described results obtained in the trial locations, showed a significant effect of year and N rate on the variation of GY and GPC, relationships with NDVI were therefore analysed separately in each of the 16 environments. Correlation analyses indicated generally strong positive relationship between NDVI and GY at N75 rate, which differed between environments and plant development stages (Figure 3a). The strongest and the most consistent correlations were found for LT and NO in 2022, which were statistically significant at all developmental stages (p<0.05), except for LT at GS85. In almost all environments the strongest correlations were found at GS75 stage, all of them, apart from EE in 2022, were significant (p<0.01). Correlations from 2021 were more similar between environments compared to 2022.
At N150 fertilization rate, the overall strength of correlations between NDVI and grain yield (GY) was similar to that at N75. Coefficients were the highest (r>0.75) for NO in 2022 and LT in 2021 (GS55). For most environments and growth stages, the correlation of NDVI with grain yield was positive, except for LV (GS25) where there was a significant (p<0.05) negative correlation (Figure 3b). Overall, at the GS25 stage, correlative relationships between GY and NDVI remained lower compared to the respective results of later growth stages, with the notable exception of EE in 2021, where GS25 had the strongest significant correlation (p<0.01).
For GPC, correlations with NDVI were notably weaker and mostly negative compared to GY or remained non-significant (Figure 4a). At N75, strong negative correlations (r>-0.5) were found for the GS25 stage (in NO in 2022, EE in 2021 and LV 2021). All the significant correlations at N75 between GPC and NDVI remained negative. GS75 was also the stage with the strongest overall correlations, compared to other development stages. However, in most environments at GS25 and GS55 correlations between NDVI and GPC were non-significant.
At N150, the correlation coefficients between GPC and NDVI were mostly negative or non-significant (Figure 4b). A notable exception to this was NO (in 2021 and 2022) where significant negative correlations (r<-0.25) were found at GS55, GS75 and GS85. Similarly, in EE in 2021 and 2022, negative correlation was rather strong (r<-0.5) for GS55 and for GS75 and GS85. In LT in 2022, a positive correlation was significant (p>0.05) at the GS25 – the only significant one for GPC and NDVI at N150.
3.6. Linear relationships among NDVI and GY, GPC
Linear regression analysis showed that most significant relationships between GY and NDVI values were found at the milk growth stage (GS75) and heading growth stage (GS55) (Table 7). Conversely, the dough growth stage (GS85) had only two significant relationships. Specific environments stood out, for instance, NO trials at the GS85 were highly significant (p<0.001) for both years and at both N rates. Lithuania for 2022 showed the highest R2 at GS75 and at N75 (0.82, p<0.001) and very low RMSE value (29.96). Overall, NO trials in 2022 showed the most consistent and significant relationships with GY for both years and N rates.
Differences in significance between two N rates were also present; for GY, trials at N150 showed more significant relationships compared to N75 (23 and 17 respectively, from total of 32 measured relationships in Table 8). This was especially clear at the GS25 and GS85 growth stages. RMSE values for N75 were on average twice smaller than for N150. Additionally, in EE trials, GY at GS75 was non-significant at N75 but highly significant at N150 for both years (adj. R2=0.40 and 0.42). In LT trials, GY was highly significant for both GS55 and GS75 in both years.
GPC showed fewer significant relationships with NDVI values compared to GY. Similarly, as to GY, most environments showed significant relationships at the GS75 and GS85 stages, with N150 having a slightly higher number of those. In contrast to GY, differences in RMSE values for GPC were on average very similar between N75 and N150. From all tested environments, the NO trial in 2022 showed the most consistent and significant relationships with GPC at GS75 and GS85 (adj. R2=0.23-0.55).
4. Discussion
Given that the price of common wheat is determined by GPC in most wheat markets, including the Nordic-Baltic region, this study focused on GY and GPC as the most critical economic traits. This comprehensive multifactorial study of spring wheat genotypes, incorporating remote phenotyping data, marks a pioneering effort under the conditions of the Nordic-Baltic region, supplying practical knowledge for breeders.
4.1. Variations in meteorological conditions across trial years and locations resulted in differences in the formation of GY and GPC
The impacts of climatic variability have been seen in the last few decades, where drought stress is a major factor that affects wheat GY and GPC increasing the variability of these traits for Baltic region conditions [7]. Meta-analysis based on wheat data showed that drought significantly decreased GY, and increased GPC [32]. In our study, drought conditions were recorded in 2021 but to a different degree, growing period, and duration across four locations, resulting in substantial GY and GPC variation. In 2021, when higher average daily temperatures and lower precipitation were observed, significantly lower GY was obtained in three out of the four trial locations (EE, LV, and NO). Considering that the complete amount of N fertilizer was applied before sowing, the heavy rainfall experienced in LT in June of 2022 caused leaching of N from upper soil layer, which is likely the main reason why both lower GY and GPC were obtained in this location. Elevated temperatures during grain filling can result in high protein percentages because these conditions tend to reduce starch synthesis more than protein synthesis [33]. In 2021, temperature and moisture conditions were more favorable for formation of higher mean GPC compared to 2022, in EE, LV, and LT, except in NO, where the mean value of the GPC did not exceed 12%, because of comparatively lower mean daily temperature in June and July in this location.
In our study the estimated field traits were significantly affected by N fertilization rates in almost all locations, with some exceptions, which can also be explained by differences in meteorological conditions between locations. It is generally acknowledged that proper increase of nitrogen supply improves plant’s resistance to abiotic stresses, yield and grain quality [34]. Recently several attempts were made to compare plant’s yield penalties due to the drought and heat stresses under low and high nitrogen supply in the field conditions for wheat. Giménez et al., (2021) [35] demonstrated that increased temperature during nights resulted in increased tiller mortality only under high nitrogen supply. Recent study demonstrated that high nitrogen supply mitigated the adverse effect of drought and heat stresses when they were exposed separately. However, when the two stresses were combined, wheat growth and grain yield were affected negatively by high N supply [36]. In 2021, the comparatively heavier drought conditions were in EE and LT, however, the timing of water shortage was different in these locations. In EE, plants experienced drought in June, while the water deficit was mostly in July in LT. In EE, the average yield was significantly higher under high nitrogen supply, albeit it was significantly higher under low nitrogen supply in LT in 2021. The obtained results of yield reduction under high N rate imply that high nitrogen supply under water deficit together with unusually high temperature at anthesis and during grain filling might cause higher plant sensitivity to drought stress. Considering that drought and high temperatures are becoming more frequent due to global warming, the rates of N fertilization might need to be revised to avoid not only pollution of the environment, but also yield penalties.
In this study, UAV-derived NDVI obtained at four growth stages also showed significant variation among trial years, although there was no marked consistency between locations due to the aforementioned differences in meteorological variables. NDVI as a basic measure of plant vitality and vegetation density can be influenced by plant growth stage, vegetation cover percentage, plant health and stress, weather conditions and crop management [37]. In a study by Thapa et al. (2019) [38] in winter wheat, it was detected that as the water stress increased, NDVI values across seasons decreased. Our results showed that NDVI exhibited a significant difference between N treatments consistently in all locations during GS55, GS75, and GS85, with higher values found under high N compared with low N conditions, although in the study of Yousfi et al. (2019) [39], the NDVI index did not show significant differences in contrasting N conditions (N0 and N160).
4.2. Spring wheat genotypes grown under contrasting N rates differed in stability and exhibited different patterns of NDVI profile during the season
Genotypes and their genetic characteristics are also key factors that affect variation of traits in spring wheat. Our results showed that genotype effects were highly significant (p<0.001) for all traits showing the presence of wide genetic variation among the genotypes. It is explainable as the set of 16 genotypes included in this study consisted of both, older and newly registered spring wheat genotypes (Table S1), which also differed significantly in terms of PH and the length of the DM (Table 6). Higher input prices and environmental concerns are now also focused on common wheat breeding programs towards low-input agricultural practices, and, more particularly, low N input management [40]. In our research we were also interested in finding out a significant genotype by nitrogen (G x N) interaction variability for traits of our focus, which would indicate wheat tolerance to N constraints. Analysis of variance showed that in our study, the G x N interaction was not significant for any of the investigated traits (Table S4). Other studies have found both, significant [25] and non-significant [41] effects of this interaction factor on GY variability. Hitz et al. (2017) [25] indicated that to select for high yield potential (or NUE), breeding material must be evaluated under both low and high N conditions. High yield potential only under low N conditions will not be enough to ensure its success, since the ability to respond to high rates of N is also important to achieve wide cultivation of a genotype. The three top-yielding spring wheat genotypes DS-655-7-DH, DS-17-16-DH and DS-720-3-DH (all originated from LT) had been characterized by breeders as high-yielding breeding lines, which was also confirmed in our study, as these genotypes showed the best results under both low and high N rates. Results also showed that grain yield is strongly associated to DM, a longer vegetation period comes with higher GY. In turn, early maturity genotypes Voore, 876 (both from EE) and Runar (NO) provided the lowest yield, but at the same time formed higher GPC under both N rates. The focus on more yield-stable genotypes will prevent N losses in unfavourable years [42]. We found indication that a genotype that is stable with respect to variation in one resource of factors will not always be relatively stable in response to variation in a different resource of factors as also shown by Weiner at al. (2021) [43]. We presented genotype performance and stability under different N rates over eight environments by using WAASB stability index that is based on AMMI model [30]. Only a limited number of published research was found about yield stability investigation under different N rates for wheat [44]. We found that genotypes had different yield stability depending on N rate. DS-17-16-DH was the most stable for GY only under low N, but genotype 990-2 in opposite was the best on WAASB stability under high N. Genotypes Zombi with below-average GY and 013-074 with below-average GPC, showed high WAASB stability ranks across both rates of N.
Analysis of variance showed that comparatively high magnitude of the genotype as a factor effect for NDVI was observed in the milk (GS75) and dough (GS85) development stages except in EE location in 2022 (Table S4). We recognized that all identified high-yielding genotypes, were characterized with the highest NDVI values in GS75 and GS85 (Table 6). During earlier stages of plant development (GS25; GS55), characteristics of those genotypes showed different patterns in terms of NDVI profiles. For instance, for top-yielding DS-655-7-DH comparatively high values of NDVI were detected in all investigated growth stages, while the old, early maturity, high-protein cultivar Runar showed one of the highest NDVI values only in GS25.
4.3. Correlations and linear relationships between GY, GPC, and UAV-derived NDVI highlight key crop growth stages
We used UAV-derived NDVI as the main vegetation index, since it has been widely used in vegetation assessments in general and wheat phenotyping in particular [19]. A study of rapid wheat phenotyping using NDVI in China showed that it had more consistent correlations with grain yield compared to other indices like NDRE and NGRDI [45]. Wheat trials in Australia found that UAV-derived NDVI values were strongly correlated with the hand-held GreenSeeker sensor (R2=0.85) [46], which can be considered as a baseline for such measurements.
Analyzing the relationships between NDVI and grain yield (GY) data in each of the 16 environments, we found the strongest correlations between NDVI and GY at the GS75 (r = 0.74, p<0.01) stage, which which indicates that this period can be used for the exploration of the predictive power of NDVI by linear models. By using the split plot design, Sultana et al. (2014) [47] found that NDVI had the strongest correlations (R2 > 0.9) with GY in booting, grain filling, and maturity stages.
UAV data collected at the heading stage was superior for predicting GPC, while GY was more accurately predicted during the grain filling stage [48]. A study from Idaho, USA [49] reports similar findings with strong linear relationships between GY and NDVI values at the middle phase of plant development (R2=0.78). Yousfi et al. (2016) [50] showed correlation coefficients between GY and NDVI ranging from 0.36 to 0.74.
Another main response variable – grain protein content (GPC) expressed a negative relationship with NDVI, which is typical and has been reported by several studies [51,52]. GPC prediction is less straightforward compared to GY, often requiring machine learning methods and ensembles of predictors [53,54]. Li-Hong et al. (2007) [55] concluded that using single measurement of spectral reflectance was not enough to predict GPC reliably in wheat. A study in China [54] found that in GPC prediction from UAV-derived NDVI using machine learning methods, RMSE did not exceed 0.74. Our study also found a weaker relationship between NDVI and GPC, compared to GY. Research states that the weak relationship between NDVI and GPC makes it a suboptimal index for protein content prediction in wheat [56].
A study using correlation analysis for wheat trials with high-resolution NDVI data for yield found average to strong correlations (r=0.60–0.81) and was able to distinguish between various rates of fertilization [27]. Here we demonstrated that N fertilization rates did not have a significant impact on the strength of correlations between NDVI and GPC, since the highest r values were found at the same locations and growth stages for both N75 and N150.
Differences in correlations between environments were quite pronounced for all yield parameters. Two locations, NO and EE, stood out, especially on significant correlations between NDVI and GY in GS25 and GS85, absent in other environments. Such differences in early and late stages of crop development could be explained by variation in biophysical parameters of environments in trials. Sánchez et al. (2023) [57] mentions that NDVI values are influenced by soil type and moisture as well as local meteorological conditions which are impossible to control in field trials.
Our study proved that UAV-derived NDVI data, combined with field-collected agronomic data, may be successfully employed for multi-environmental trials of wheat phenotyping. One of our main findings is that NDVI values at GS55 and GS75 were the most significant linear predictors for spring wheat GY. Other studies [58,59] reported the highest R2 values for linear relationships between wheat NDVI and GY at these stages. However, it is important to note that uncertainties arise from differences in growing conditions during crop development stages. In our study, GY showed more significant relationships compared to GPC. This is also visible in many studies which use GY as the main response variable see [60].
In our trials, higher N application rate (N150) showed slightly more significant linear relationships compared to N75, although this was not clear from correlation analysis. This effect was expected since N fertilization rate is known to influence NDVI values [24,61], however, the influence on R2 values between environments was quite modest.
Single growing season with a major meteorological influence, for example, significant drought stress, can alter expected NDVI values. Thapa et al. (2019) [38] reports that drought stress at the later stages of wheat development reduced the strength of the relationship of NDVI to GY.
5. Conclusions
Overall, our study reveals the intersection of climate issues and agricultural development in the Nordic and Baltic region. Unusual temperatures and water deficits during critical stages of plant development emphasize significant meteorological differences between years and locations. The study establishes a clear link between these deviations and their effects on nitrogen uptake, causing substantial phenotypic changes and altering trait relationships in different environments. Our findings provide important insights into the development of resilient crops under changing environmental conditions.
In brief, our research identified top-yielding spring wheat genotypes that performed well under both high and low nitrogen rates. Genotypes with early maturity exhibited higher grain protein content. The study revealed variations in grain yield stability among different genotypes depending on nitrogen rates.
The study underscores substantial enhancement in wheat phenotyping research through UAV-derived data, particularly in a multi-environment pilot project. A key finding emphasizes the role of NDVI analysis at growth stages GS55 and GS75, serving as significant linear predictors, particularly for spring wheat grain yield. However, it’s crucial to note that the impact of these indices varies among distinct growing conditions at various locations, limiting their universal applicability. To optimize their effectiveness, a nuanced understanding of the intricate relationships between NDVI values and local growing conditions is essential.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org, Figure S1: title; Table S1: title; Video S1: title. Table S1. Name, origin and description of the spring wheat genotypes selected for study. Table S2. Physical and chemical properties of the experimental soil before sowing at four locations (EE, LV, LT, NO) and two growing year (2021, 2022). Table S3. Results of 3-way ANOVA for GY, GPC and NDVI at different growth stages. Table S4. Results of 2-way ANOVA for GY, GPC and NDVI at different growth stages.
Author Contributions
Conceptualization and methodology, G.B., M.L, M.B., H.K., I.T.; formal analysis, M.B., Z.J., A.L.; Investigation, A.G., A.A, T.M., S.S., M.L., Z.J., A.L., A.I.; resources, M.L.; data curation, A.G., A.A., Z.J.; writing—original draft preparation, M.B., Z.J., Z.R.; writing—review and editing, A.G., A.A., G.B., H.K., M.B., Z.R., Z.J., M.L, A.L., I.T., A.I.; visualization, A.G., Z.J.; project administration, G.B.; funding acquisition, G.B., M.L., M.B., H.K., I.T. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded under the project “NOBALwheat – breeding toolbox for sustainable food system of the NOrdic BALtic region”, which received funding from Iceland, Liechtenstein and Norway through the EEA Grants, with the Research Council of Lithuania (LMTLT) contract No. S-BMT-21-3 (LT08-2-LMT-K-01-032).
Data Availability Statement
Data are available from the authors upon reasonable request.
Acknowledgments
In this section, you can acknowledge any support given which is not covered by the author contribution or funding sections. This may include administrative and technical support, or donations in kind (e.g., materials used for experiments).
Conflicts of Interest
The authors declare no conflict of interest.
References
- FAO 2023. https://www.fao.org/faostat/. (accessed.
- Wirehn, L. Nordic agriculture under climate change: A systematic review of challenges, opportunities and adaptation strategies for crop production. Land Use Policy 2018, 77, 63–74. [Google Scholar] [CrossRef]
- FAO 2021 (accessed.
- Lillemo, M.; Dieseth, J.A. Wheat breeding in Norway. In The World Wheat Book 2, A History of Wheat Breeding; Bonjean, A.P., Angus, W.J., van Ginkel, M., Vol. 1432; 2011; pp 45–75.
- Ruzgas, V.; Liatukas, Z. Wheat breeding in Lithuania. In The World Wheat Book 2, A History of Wheat Breeding; Bonjean, A.P., Angus, W.J., van Ginkel, M., Vol. 1432; 2011; pp 175-192.
- Strazdina, V.; Belicka, I.; Rashal, I.; Grauda, D. History of wheat breeding development in Latvia. In The World Wheat Book 2; Bonjean, A.P., Angus, W.J., van Ginkel, M., Vol. 1432; 2011; pp 213-238.
- Koppel, R.; Ingver, A.; Ardel, P.; Kangor, T.; Kennedy, H. J.; Koppel, M. The variability of yield and baking quality of wheat and suitability for export from Nordic-Baltic conditions. Acta Agric. Scand. Sect. B-Soil Plant Sci. 2020, 70, 628–639. [Google Scholar] [CrossRef]
- Ross, A. S. Genetic and other factors affecting wheat quality. In Achieving Sustainable Cultivation of Wheat, Vol 1: Breeding, Quality Traits, Pests and Diseases, Langridge, P. Ed.; Burleigh Dodds Series in Agricultural Science, Vol. 5; Burleigh Dodds Science Publishing Ltd, 2017; 177-211.
- Cormier, F.; Faure, S.; Dubreuil, P.; Heumez, E.; Beauchêne, K.; Lafarge, S.; Praud, S.; Le Gouis, J. A multi-environmental study of recent breeding progress on nitrogen use efficiency in wheat (Triticum aestivum L.). Theor. Appl. Genet. 2013, 126, 3035–3048. [Google Scholar] [CrossRef]
- Sehgal, A.; Sita, K.; Siddique, K.H.M.; Kumar, R.; Bhogireddy, S.; Varshney, R.K.; HanumanthaRao, B.; Nair, R.M.; Prasad, P.V.V.; Nayyar, H. Drought or/and heat-stress effects on seed filling in food crops: Impacts on functional biochemistry, seed yields, and nutritional quality. Front. Plant Sci. 2018, 9, 1705. [Google Scholar] [CrossRef] [PubMed]
- Kopecká, R.; Kameniarová, M.; Cerny, M.; Brzobohaty, B.; Novák, J. Abiotic Stress in Crop Production. Int. J. Mol. Sci. 2023, 24, 47. [Google Scholar] [CrossRef]
- Hawkesford, M. J. Reducing the reliance on nitrogen fertilizer for wheat production. J. Cereal Sci. 2014, 59, 276–283. [Google Scholar] [CrossRef] [PubMed]
- Marsh, J. I.; Hu, H. F.; Gill, M.; Batley, J.; Edwards, D. Crop breeding for a changing climate: Integrating phenomics and genomics with bioinformatics. Theor. Appl. Genet. 2021, 134, 1677–1690. [Google Scholar] [CrossRef]
- Ma, J. J.; Zheng, B. Y.; He, Y. Applications of a Hyperspectral Imaging System Used to Estimate Wheat Grain Protein: A Review. Front. Plant Sci. 2022, 13, 10. [Google Scholar] [CrossRef]
- Radoglou-Grammatikis, P.; Sarigiannidis, P.; Lagkas, T.; Moscholios, I. A compilation of UAV applications for precision agriculture. Comput. Netw. 2020, 172, 18. [Google Scholar] [CrossRef]
- Xie, C.; Yang, C. A review on plant high-throughput phenotyping traits using UAV-based sensors. Comput. Electron. Agric. 2020, 178, 105731. [Google Scholar] [CrossRef]
- Fei, S. P.; Hassan, M. A.; He, Z. H.; Chen, Z.; Shu, M. Y.; Wang, J. K.; Li, C. C.; Xiao, Y. G. Assessment of Ensemble Learning to Predict Wheat Grain Yield Based on UAV-Multispectral Reflectance. Remote Sens. 2021, 13, 25. [Google Scholar] [CrossRef]
- Prey, L.; Hanemann, A.; Ramgraber, L.; Seidl-Schulz, J.; Noack, P. O. UAV-Based Estimation of Grain Yield for Plant Breeding: Applied Strategies for Optimizing the Use of Sensors, Vegetation Indices, Growth Stages, and Machine Learning Algorithms. Remote Sens. 2022, 14, 17. [Google Scholar] [CrossRef]
- Huang, S.; Tang, L. N.; Hupy, J. P.; Wang, Y.; Shao, G. F. A commentary review on the use of normalized difference vegetation index (NDVI) in the era of popular remote sensing. J. For. Res. 2021, 32, 1–6. [Google Scholar] [CrossRef]
- Bellón, B.; Bégué, A.; Lo Seen, D.; de Almeida, C. A.; Simoes, M. A Remote Sensing Approach for Regional-Scale Mapping of Agricultural Land-Use Systems Based on NDVI Time Series. Remote Sens. 2017, 9, 600. [Google Scholar] [CrossRef]
- Zhu, X. L.; Liu, D. S. Improving forest aboveground biomass estimation using seasonal Landsat NDVI time-series. Isprs J. Photogramm. Remote Sens. 2015, 102, 222–231. [Google Scholar] [CrossRef]
- Beisel, N. S.; Callaham, J. B.; Sng, N. J.; Taylor, D. J.; Paul, A.-L.; Ferl, R. J. Utilization of single-image normalized difference vegetation index (SI-NDVI) for early plant stress detection. Appl. Plant Sci. 2018, 6, e01186. [Google Scholar] [CrossRef] [PubMed]
- Pastor-Guzman, J.; Atkinson, P. M.; Dash, J.; Rioja-Nieto, R. Spatiotemporal Variation in Mangrove Chlorophyll Concentration Using Landsat 8. Remote Sens. 2015, 7, 14530–14558. [Google Scholar] [CrossRef]
- Cabrera-Bosquet, L.; Molero, G.; Stellacci, A. M.; Bort, J.; Nogués, S.; Araus, J. L. NDVI as a Potential Tool for Predicting Biomass, Plant Nitrogen Content and Growth in Wheat Genotypes Subjected to Different Water and Nitrogen Conditions. Cereal Res. Commun. 2011, 39, 147–159. [Google Scholar] [CrossRef]
- Hitz, K.; Clark, A. J.; Van Sanford, D. A. Identifying nitrogen-use efficient soft red winter wheat lines in high and low nitrogen environments. Field Crops Res. 2017, 200, 1–9. [Google Scholar] [CrossRef]
- Naser, M. A.; Khosla, R.; Longchamps, L.; Dahal, S. Using NDVI to Differentiate Wheat Genotypes Productivity Under Dryland and Irrigated Conditions. Remote Sens. 2020, 12, 17. [Google Scholar] [CrossRef]
- Guan, S. L.; Fukami, K.; Matsunaka, H.; Okami, M.; Tanaka, R.; Nakano, H.; Sakai, T.; Nakano, K.; Ohdan, H.; Takahashi, K. Assessing Correlation of High-Resolution NDVI with Fertilizer Application Level and Yield of Rice and Wheat Crops using Small UAVs. Remote Sens. 2019, 11, 19. [Google Scholar] [CrossRef]
- Zadoks, J. C.; Chang, T. T.; Konzak, C. F. A decimal code for the growth stages of cereals. Weed Res. 1974, 14, 415–421. [Google Scholar] [CrossRef]
- Zobel, R.; Wright, M.; Gauch, H. Statistical Analysis of a Yield Trial. Agron. J. 1988, 80, 388–393. [Google Scholar] [CrossRef]
- Olivoto, T.; Lúcio, A. D. C.; da Silva, J. A. G.; Marchioro, V. S.; de Souza, V. Q.; Jost, E. Mean Performance and Stability in Multi- Environment Trials I: Combining Features of AMMI and BLUP Techniques. Agron. J. 2019, 111, 2949–2960. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing. R Foundation for Statistical Computing: Vienna, Austria, 2021. URL https://www.R-project.org/.
- Wan, C.; Dang, P.; Gao, L.; Wang, J.; Tao, J.; Qin, X.; Feng, B.; Gao, J. How Does the Environment Affect Wheat Yield and Protein Content Response to Drought? A Meta-Analysis. Front. Plant Sci. 2022, 13, 896985. [Google Scholar] [CrossRef] [PubMed]
- Asseng, S.; Martre, P.; Maiorano, A.; Roetter, R. P.; O’Leary, G. J.; Fitzgerald, G. J.; Girousse, C.; Motzo, R.; Giunta, F.; Babar, M. A.; et al. Climate change impact and adaptation for wheat protein. Glob. Chang. Biol. 2019, 25, 155–173. [Google Scholar] [CrossRef] [PubMed]
- Abid, M.; Tian, Z.; Ata-Ul-Karim, S. T.; Cui, Y.; Liu, Y.; Zahoor, R.; Jiang, D.; Dai, T. Nitrogen Nutrition Improves the Potential of Wheat (Triticum aestivum L.) to Alleviate the Effects of Drought Stress during Vegetative Growth Periods. Front. Plant Sci. 2016, 7, 981. [Google Scholar] [CrossRef]
- Giménez, V. D.; Miralles, D. J.; García, G. A.; Serrago, R. A. Can crop management reduce the negative effects of warm nights on wheat yield? Field Crops Res. 2021, 261, 108010. [Google Scholar] [CrossRef]
- Ru, C.; Hu, X. T.; Chen, D. Y.; Wang, W. E.; Zhen, J. B.; Song, T. Y. Individual and combined effects of heat and drought and subsequent recovery on winter wheat (Triticum aestivum L.) photosynthesis, nitrogen metabolism, cell osmoregulation, and yield formation. Plant Physiol. Biochem. 2023, 196, 222–235. [Google Scholar] [CrossRef]
- Forkel, M.; Carvalhais, N.; Verbesselt, J.; Mahecha, M. D.; Neigh, C. S. R.; Reichstein, M. Trend Change Detection in NDVI Time Series: Effects of Inter-Annual Variability and Methodology. Remote Sens. 2013, 5, 2113–2144. [Google Scholar] [CrossRef]
- Thapa, S.; Rudd, J. C.; Xue, Q. W.; Bhandari, M.; Reddy, S. K.; Jessup, K. E.; Liu, S. Y.; Devkota, R. N.; Baker, J.; Baker, S. Use of NDVI for characterizing winter wheat response to water stress in a semi-arid environment. J. Crop Improv. 2019, 33, 633–648. [Google Scholar] [CrossRef]
- Yousfi, S.; Gracia-Romero, A.; Kellas, N.; Kaddour, M.; Chadouli, A.; Karrou, M. .; Araus, J. L.; Serret, M. D. Combined Use of Low-Cost Remote Sensing Techniques and δ13C to Assess Bread Wheat Grain Yield under Different Water and Nitrogen Conditions. Agronomy 2019, 9, 285. [Google Scholar] [CrossRef]
- Laperche, A.; Brancourt-Hulmel, M.; Heumez, E.; Gardet, O.; Hanocq, E.; Devienne-Barret, F.; Le Gouis, J. Using genotype x nitrogen interaction variables to evaluate the QTL involved in wheat tolerance to nitrogen constraints. Theor. Appl. Genet. 2007, 115, 399–415. [Google Scholar] [CrossRef]
- Mroz, T.; Dieseth, J. A.; Lillemo, M. Historical grain yield genetic gains in Norwegian spring wheat under contrasting fertilization regimes. Crop Sci. 2022, 62, 997–1010. [Google Scholar] [CrossRef]
- Zörb, C.; Ludewig, U.; Hawkesford, M. J. Perspective on Wheat Yield and Quality with Reduced Nitrogen Supply. Trends Plant Sci. 2018, 23, 1029–1037. [Google Scholar] [CrossRef]
- Weiner, J.; Du, Y. L.; Zhao, Y. M.; Li, F. M. Allometry and Yield Stability of Cereals. Front. Plant Sci. 2021, 12, 5. [Google Scholar] [CrossRef]
- Ali, M. Stability Analysis of Bread Wheat Genotypes under Different Nitrogen Fertilizer Levels. J. Plant Prod. 2017, 8, 261–275. [Google Scholar] [CrossRef]
- Hassan, M. A.; Yang, M. J.; Fu, L. P.; Rasheed, A.; Zheng, B. Y.; Xia, X. C.; Xiao, Y. G.; He, Z. H. Accuracy assessment of plant height using an unmanned aerial vehicle for quantitative genomic analysis in bread wheat. Plant Methods 2019, 15, 12. [Google Scholar] [CrossRef]
- Duan, T.; Chapman, S. C.; Guo, Y.; Zheng, B. Dynamic monitoring of NDVI in wheat agronomy and breeding trials using an unmanned aerial vehicle. Field Crops Res. 2017, 210, 71–80. [Google Scholar] [CrossRef]
- Sultana, S. R.; Ali, A.; Ahmad, A.; Mubeen, M.; Zia-Ul-Haq, M.; Ahmad, S.; Ercisli, S.; Jaafar, H. Z. E. Normalized Difference Vegetation Index as a Tool for Wheat Yield Estimation: A Case Study from Faisalabad, Pakistan. Sci. World J. 2014, 2014, 725326. [Google Scholar] [CrossRef]
- Sandhu, K. S.; Mihalyov, P. D.; Lewien, M. J.; Pumphrey, M. O.; Carter, A. H. Combining Genomic and Phenomic Information for Predicting Grain Protein Content and Grain Yield in Spring Wheat. Front. Plant Sci. 2021, 12, 14. [Google Scholar] [CrossRef] [PubMed]
- Walsh, O. S.; Marshall, J. M.; Nambi, E.; Jackson, C. A.; Ansah, E. O.; Lamichhane, R.; McClintick-Chess, J.; Bautista, F. Wheat Yield and Protein Estimation with Handheld and Unmanned Aerial Vehicle-Mounted Sensors. Agronomy 2023, 13, 14. [Google Scholar] [CrossRef]
- Yousfi, S.; Kellas, N.; Saidi, L.; Benlakehal, Z.; Chaou, L.; Siad, D.; Herda, F.; Karrou, M.; Vergara, O.; Gracia, A.; et al. Comparative performance of remote sensing methods in assessing wheat performance under Mediterranean conditions. Agric. Water Manag. 2016, 164, 137–147. [Google Scholar] [CrossRef]
- Feng, M.-c.; Xiao, L.-j.; Zhang, M.-j.; Yang, W.-d.; Ding, G.-w. Integrating Remote Sensing and GIS for Prediction of Winter Wheat (Triticum aestivum) Protein Contents in Linfen (Shanxi), China. PLoS ONE 2014, 9, e80989. [Google Scholar] [CrossRef] [PubMed]
- Stoy, P. C.; Khan, A. M.; Wipf, A.; Silverman, N.; Powell, S. L. The spatial variability of NDVI within a wheat field: Information content and implications for yield and grain protein monitoring. PLoS ONE 2022, 17, e0265243. [Google Scholar] [CrossRef] [PubMed]
- Barmeier, G.; Hofer, K.; Schmidhalter, U. Mid-season prediction of grain yield and protein content of spring barley cultivars using high-throughput spectral sensing. Eur. J. Agron. 2017, 90, 108–116. [Google Scholar] [CrossRef]
- Fu, Z. P.; Yu, S. S.; Zhang, J. Y.; Xi, H.; Gao, Y.; Lu, R. H.; Zheng, H. B.; Zhu, Y.; Cao, W. X.; Liu, X. J. Combining UAV multispectral imagery and ecological factors to estimate leaf nitrogen and grain protein content of wheat. Eur. J. Agron. 2022, 132, 15. [Google Scholar] [CrossRef]
- Xue, L. H.; Cao, W. X.; Yang, L. Z. Predicting grain yield and protein content in winter wheat at different N supply levels using canopy reflectance spectra. Pedosphere 2007, 17, 646–653. [Google Scholar] [CrossRef]
- Wang, K.; Huggins, D. R.; Tao, H. Y. Rapid mapping of winter wheat yield, protein, and nitrogen uptake using remote and proximal sensing. Int. J. Appl. Earth Obs. Geoinf. 2019, 82, 10. [Google Scholar] [CrossRef]
- Sánchez, N.; Plaza, J.; Criado, M.; Pérez-Sánchez, R.; Gómez-Sánchez, M. A.; Morales-Corts, M. R.; Palacios, C. The Second Derivative of the NDVI Time Series as an Estimator of Fresh Biomass: A Case Study of Eight Forage Associations Monitored via UAS. Drones 2023, 7, 19. [Google Scholar] [CrossRef]
- Marti, J.; Bort, J.; Slafer, G.A.; Araus, J.L. Can wheat,yield be assessed by early measurements of Normalized Difference Vegetation Index? Ann. Appl. Biol. 2007, 150, 253–257. [Google Scholar] [CrossRef]
- Chandel, N. S.; Tiwari, P. S.; Singh, K. P.; Jat, D.; Gaikwad, B. B.; Tripathi, H.; Golhani, K. Yield prediction in wheat (Triticum aestivum L.) using spectral reflectance indices. Curr. Sci. 2019, 116, 272–278. [Google Scholar] [CrossRef]
- Noureldin, N. A.; Saudy, H. S.; Ashmawy, F.; Saed, H. M. Grain yield response index of bread wheat cultivars as influenced by nitrogen levels. Ann. Agric. Sci. 2013, 58, 147–152. [Google Scholar] [CrossRef]
- Walsh, O. S.; Torrion, J. A.; Liang, X.; Shafian, S.; Yang, R.; Belmont, K. M.; McClintick-Chess, J. R. Grain yield, quality, and spectral characteristics of wheat grown under varied nitrogen and irrigation. Agrosystems Geosci. Environ. 2020, 3, e20104. [Google Scholar] [CrossRef]
Figure 1.
Meteorological parameters at the field trial locations (NO – Norway, EE - Estonia, LV – Latvia, LT - Lithuania) and seasons (2021, 2022) compared to 30-year long-term average (LTA) data (1991-2020): (a) - deviation from the mean daily temperature from LTA; (b) - monthly rainfall as a percentage of the LTA.
Figure 1.
Meteorological parameters at the field trial locations (NO – Norway, EE - Estonia, LV – Latvia, LT - Lithuania) and seasons (2021, 2022) compared to 30-year long-term average (LTA) data (1991-2020): (a) - deviation from the mean daily temperature from LTA; (b) - monthly rainfall as a percentage of the LTA.
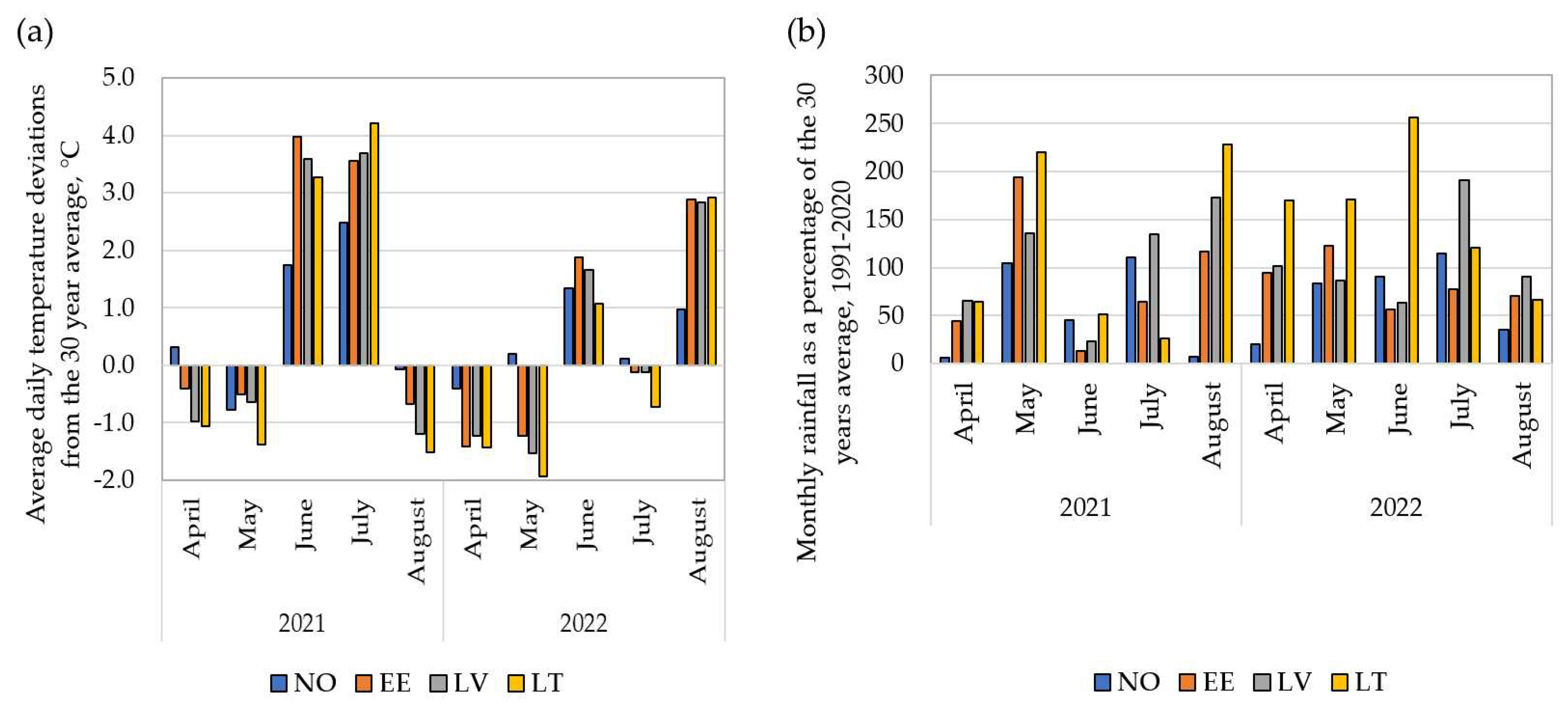
Figure 2.
Box-plot showing the phenotypic distribution of (a) grain yield (GY), (b) grain protein content (GPC) for 16 spring wheat genotypes grown under 16 environments. The black horizontal line in each boxplot is the median, the lower and upper box edges are the first and third quartiles, respectively, and the whiskers are the data minimum and maximum. The black circle in each plot is the mean for that class. Outliers are shown as open circles; EE–Estonia, LV–Latvia, LT–Lithuania, NO–Norway; 2021 and 2022 – year of trials; N75 – N rate with 75 kg N ha-1; N150 - N rate with 150 kg N ha-1; ab – significant differences (p<0.05) between the mean values of two N rates within each location-year are shown by different superscript letters.
Figure 2.
Box-plot showing the phenotypic distribution of (a) grain yield (GY), (b) grain protein content (GPC) for 16 spring wheat genotypes grown under 16 environments. The black horizontal line in each boxplot is the median, the lower and upper box edges are the first and third quartiles, respectively, and the whiskers are the data minimum and maximum. The black circle in each plot is the mean for that class. Outliers are shown as open circles; EE–Estonia, LV–Latvia, LT–Lithuania, NO–Norway; 2021 and 2022 – year of trials; N75 – N rate with 75 kg N ha-1; N150 - N rate with 150 kg N ha-1; ab – significant differences (p<0.05) between the mean values of two N rates within each location-year are shown by different superscript letters.
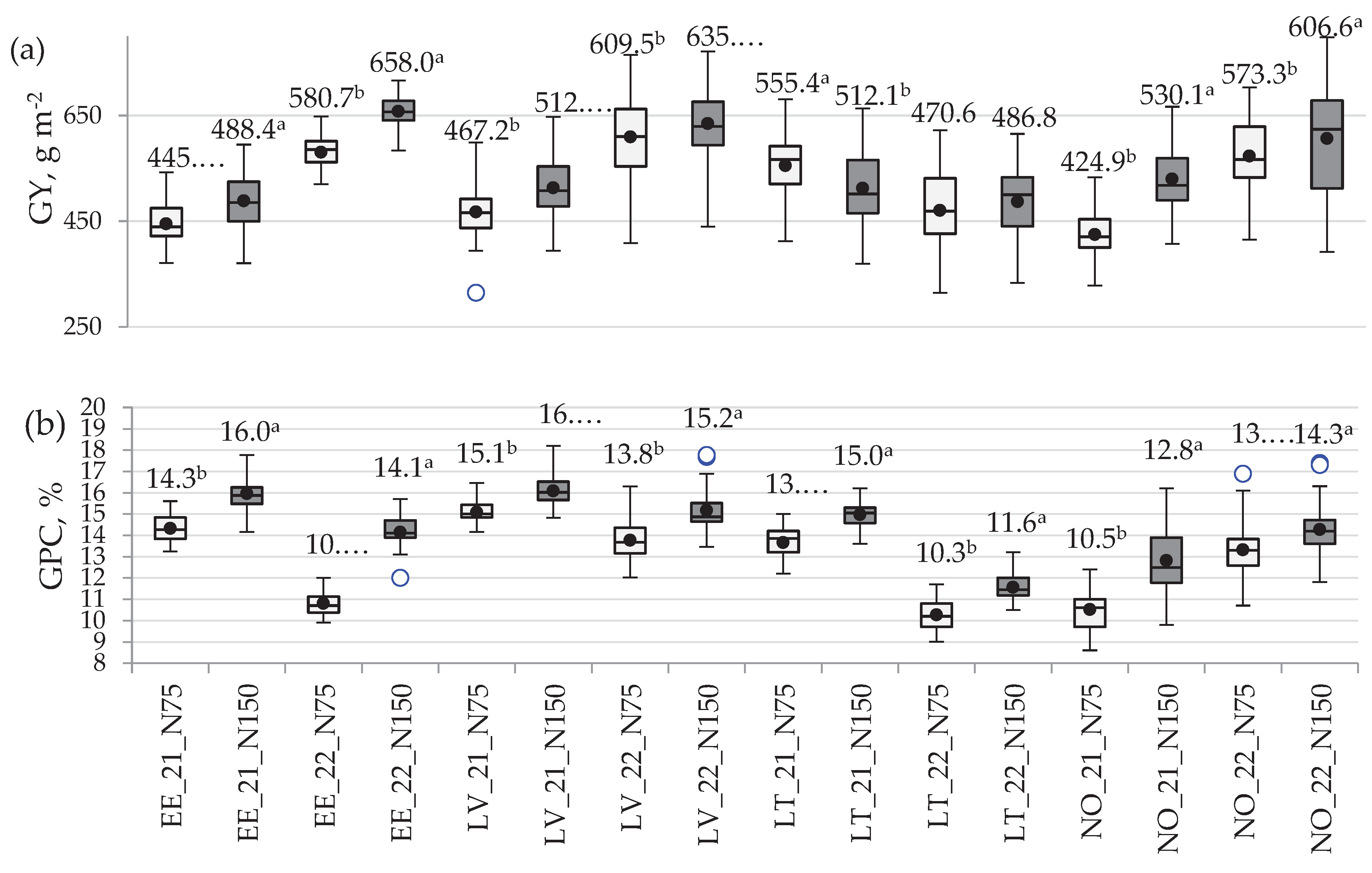
Figure 3.
Correlation coefficients (r) among NDVI and grain yield across 16 spring wheat genotypes grown under two N rates: (a) 75 kg N ha-1, (b) N150 - 150 kg N ha-1 in 8 environments; EE - Estonia, LV-Latvia, LT – Lithuania, NO – Norway; NDVI - Normalized Difference Vegetation Index; GS25 – tillering growth stage; GS55 – heading growth stage; GS75 – milk growth stage; GS85 – dough growth stage; ***p<0.001; **p<0.01; * p<0.05.
Figure 3.
Correlation coefficients (r) among NDVI and grain yield across 16 spring wheat genotypes grown under two N rates: (a) 75 kg N ha-1, (b) N150 - 150 kg N ha-1 in 8 environments; EE - Estonia, LV-Latvia, LT – Lithuania, NO – Norway; NDVI - Normalized Difference Vegetation Index; GS25 – tillering growth stage; GS55 – heading growth stage; GS75 – milk growth stage; GS85 – dough growth stage; ***p<0.001; **p<0.01; * p<0.05.
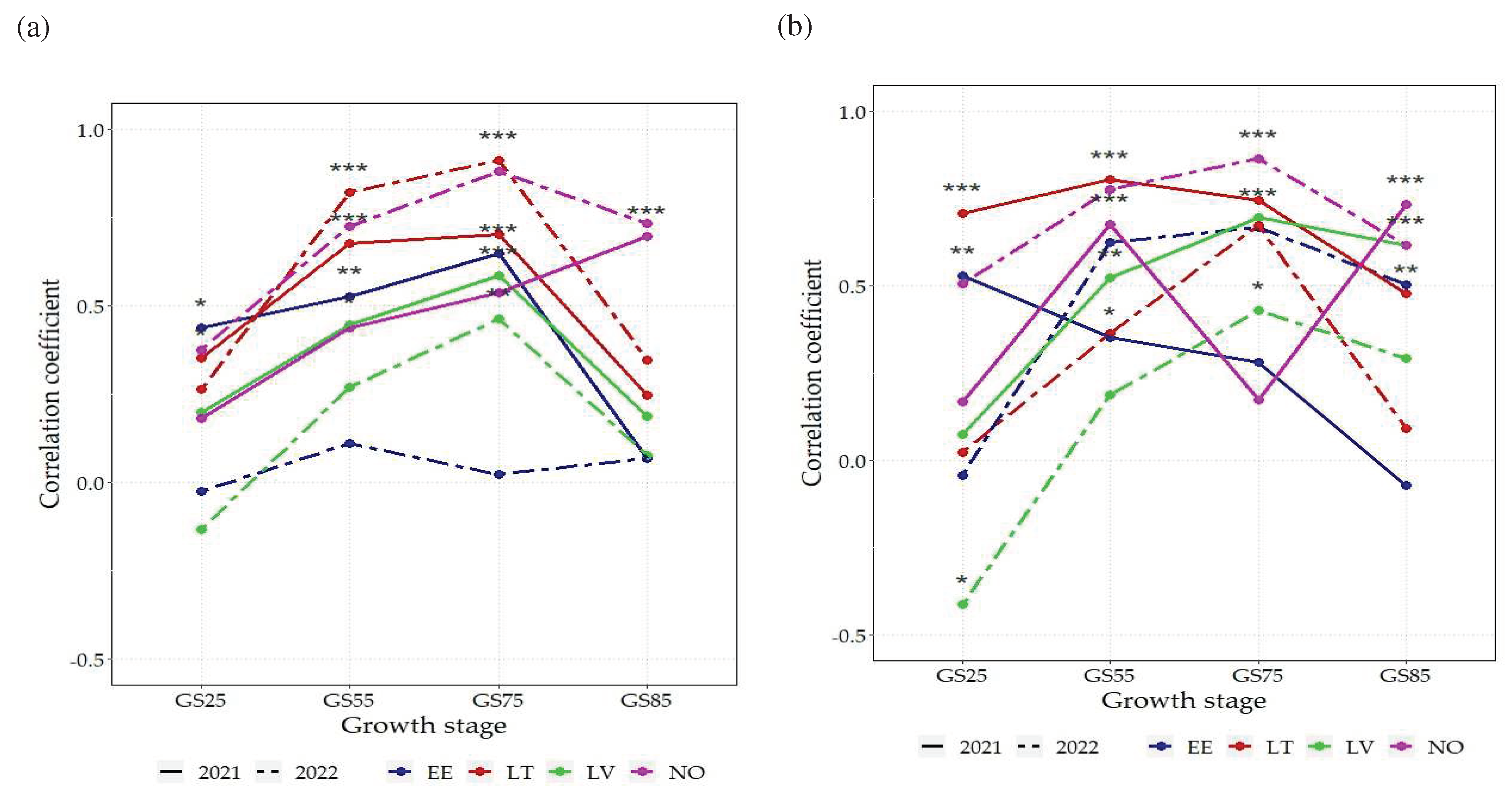
Figure 4.
Correlation coefficients (r) among NDVI and grain protein content across 16 spring wheat genotypes grown under two N rates: (a) 75 kg N ha-1, (b) N150 - 150 kg N ha-1 in 8 environments. EE - Estonia, LV-Latvia, LT – Lithuania, NO – Norway; NDVI - Normalized Difference Vegetation Index; GS25 – tillering growth stage; GS55 – heading growth stage; GS75 – milk growth stage; GS85 – dough growth stage; ***p<0.001; **p<0.01; * p<0.05.
Figure 4.
Correlation coefficients (r) among NDVI and grain protein content across 16 spring wheat genotypes grown under two N rates: (a) 75 kg N ha-1, (b) N150 - 150 kg N ha-1 in 8 environments. EE - Estonia, LV-Latvia, LT – Lithuania, NO – Norway; NDVI - Normalized Difference Vegetation Index; GS25 – tillering growth stage; GS55 – heading growth stage; GS75 – milk growth stage; GS85 – dough growth stage; ***p<0.001; **p<0.01; * p<0.05.
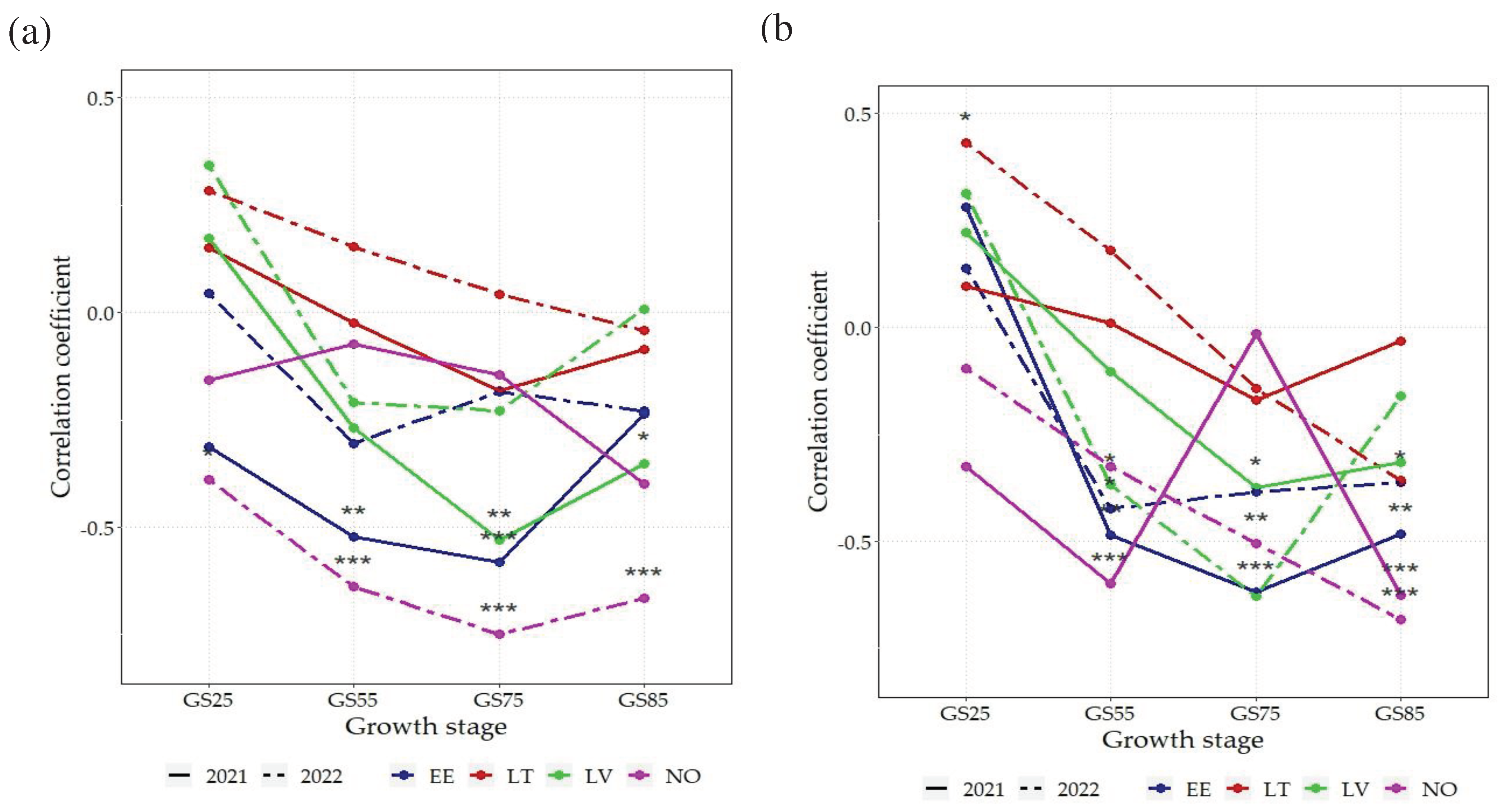

Table 1.
UAV flight mission dates for multi-location trials.
| Stage of plant development | LV | EE | LT | NO | |||||
|---|---|---|---|---|---|---|---|---|---|
| 2021 | 2022 | 2021 | 2022 | 2021 | 2022 | 2021 | 2022 | ||
| Middle of vegetative plant development | GS25 | 05.29 | 05.25 | 05.31 | 06.06 | 05.28 | 05.23 | 05.28 | 05.31 |
| Spike emergence or heading | GS55 | 06.21 | 06.30 | 06.28 | 06.30 | 06.24 | 06.27 | 06.23 | 06.29 |
| Middle of grain filling | GS75 | 07.13 | 07.22 | 07.12 | 07.21 | 07.09 | 07.19 | 07.13 | 07.21 |
| Late grain filling | GS85 | 07.27 | 08.12 | 07.26 | 08.09 | 07.23 | 08.08 | 07.26 | 08.09 |
*GS – approximate growth stage based on Zadoks growth scale [28]; LV-Latvia, EE-Estonia, LT – Lithuania; NO-Norway.
Table 2.
Mean values by years in the trial locations averaged over N fertilization rates for grain yield (GY) and grain protein content (GPC).
Table 2.
Mean values by years in the trial locations averaged over N fertilization rates for grain yield (GY) and grain protein content (GPC).
| Year | Location | |||
|---|---|---|---|---|
| NO | EE | LV | LT | |
| GY, g m-2 | ||||
| 2021 | 477.5±79.1b | 466.8±51.9b | 490.0±57.2b | 533.7±70.7a |
| 2022 | 589.9±94.6a | 619.3±50.5a | 622.2±75.7a | 478.7±73.3b |
| GPC, % | ||||
| 2021 | 11.7±1.7b | 15.1±1.1a | 15.6±0.8a | 14.3±0.9a |
| 2022 | 13.8±1.4a | 12.5±1.8b | 14.5±1.2b | 10.9±0.9b |
*NO – Norway, EE - Estonia, LV– Latvia, LT – Lithuania; the values with different superscript letters in a row are significantly different between years (p<0.05).
Table 3.
Mean values by years in the trial locations averaged over N fertilization rates for UAV-derived NDVI at four wheat plant growth stages.
Table 3.
Mean values by years in the trial locations averaged over N fertilization rates for UAV-derived NDVI at four wheat plant growth stages.
| Year | Location | |||
|---|---|---|---|---|
| NO | EE | LV | LT | |
| GS25 | ||||
| 2021 | 0.536±0.05a | 0.637±0.05a | 0.479±0.03a | 0.544±0.08b |
| 2022 | 0.466±0.10b | 0.515±0.09b | 0.408±0.04b | 0.574±0.06a |
| GS55 | ||||
| 2021 | 0.889±0.03 | 0.903±0.01a | 0.864±0.02b | 0.870±0.03b |
| 2022 | 0.882±0.03 | 0.894±0.02b | 0.905±0.01a | 0.885±0.02a |
| GS75 | ||||
| 2021 | 0.821±0.04b | 0.791±0.03 | 0.763±0.05b | 0.755±0.06a |
| 2022 | 0.837±0.04a | 0.782±0.07 | 0.803±0.04a | 0.729±0.06b |
| GS85 | ||||
| 2021 | 0.478±0.09b | 0.445±0.06 | 0.349±0.04b | 0.350±0.04a |
| 2022 | 0.577±0.10a | 0.425±0.08 | 0.366±0.05a | 0.269±0.03b |
*NO – Norway, EE – Estonia, LV – Latvia, LT – Lithuania; the values with different superscript letters in a row are significantly different between years (p<0.05); GS25 –tillering growth stage; GS55 – heading growth stage; GS75 – milk growth stage; GS85 – dough growth stage.
Table 4.
Mean values of UAV-derived NDVI obtained at four growth stages from eight environments (combination of locations x years) under low and high N application (N75, N150).
Table 4.
Mean values of UAV-derived NDVI obtained at four growth stages from eight environments (combination of locations x years) under low and high N application (N75, N150).
| Environment | GS25 | GS55 | GS75 | GS85 | ||||
|---|---|---|---|---|---|---|---|---|
| N75 | N150 | N75 | N150 | N75 | N150 | N75 | N150 | |
| EE_2021 | 0.613b | 0.660a | 0.898b | 0.908a | 0.784 | 0.797 | 0.430b | 0.461a |
| EE_2022 | 0.506 | 0.521 | 0.890 | 0.897 | 0.780 | 0.783 | 0.414 | 0.436 |
| LV_2021 | 0.477 | 0.475 | 0.857b | 0.871a | 0.748b | 0.778a | 0.335b | 0.363a |
| LV_2022 | 0.403 | 0.412 | 0.901b | 0.910a | 0.799 | 0.807 | 0.351b | 0.382a |
| LT_2021 | 0.546 | 0.542 | 0.868 | 0.872 | 0.753 | 0.757 | 0.323b | 0.346a |
| LT_2022 | 0.602a | 0.546b | 0.878b | 0.891a | 0.702b | 0.755a | 0.260b | 0.277a |
| NO_2021 | 0.520b | 0.552a | 0.866b | 0.912a | 0.791b | 0.850a | 0.421b | 0.535a |
| NO_2022 | 0.467 | 0.466 | 0.885 | 0.880 | 0.837 | 0.836 | 0.547b | 0.606a |
*EE - Estonia, LV-Latvia, LT – Lithuania, NO – Norway; NDVI - Normalized Difference Vegetation Index; GS25 –tillering growth stage; GS55 – heading growth stage; GS75 – milk growth stage; GS85 – dough growth stage. NDVI mean values followed by the different letter within columns of each pair of two N rates (N75 - 75 kg N ha-1, N150 - 150 kg N ha-1) are significantly different (p<0.05).
Table 5.
Grain yield (GY), grain protein content (GPC) and corresponding Weighted Average of Absolute Scores (WAASB) stability index estimates and their ranking for genotypes in the trial averaged over 8 environments (combination of location and year) under low N and high N rates.
Table 5.
Grain yield (GY), grain protein content (GPC) and corresponding Weighted Average of Absolute Scores (WAASB) stability index estimates and their ranking for genotypes in the trial averaged over 8 environments (combination of location and year) under low N and high N rates.
| Genotype | GY | WAASB_GY | GPC | WAASB_GPC | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| N75 | N150 | N75 | N150 | N75 | N150 | N75 | N150 | |||||||||
| g m-2 | R | g m-2 | R | value | R | value | R | % | R | % | R | % | R | % | R | |
| DS-655-7-DH | 564.1 | 1 | 592.5 | 3 | 2.7 | 11 | 3.54 | 12 | 12.7 | 7 | 14.3 | 7 | 0.39 | 12 | 0.33 | 10 |
| DS-17-16-DH | 558.1 | 2 | 609.5 | 1 | 1.13 | 1 | 2.63 | 9 | 12.0 | 15 | 13.6 | 15 | 0.20 | 2 | 0.35 | 12 |
| DS-720-3-DH | 547.4 | 3 | 607.7 | 2 | 4.23 | 16 | 2.83 | 10 | 13.0 | 5 | 14.5 | 4 | 0.22 | 6 | 0.20 | 5 |
| DS-638-5-DH | 539.9 | 4 | 575.6 | 6 | 3.28 | 12 | 4.18 | 16 | 11.7 | 16 | 13.2 | 16 | 0.39 | 13 | 0.59 | 15 |
| Betong | 538.6 | 5 | 570.9 | 7 | 1.73 | 4 | 2.02 | 6 | 12.7 | 9 | 14.2 | 11 | 0.22 | 5 | 0.28 | 7 |
| 013-01 | 538.4 | 6 | 581.7 | 4 | 4.22 | 15 | 3.79 | 15 | 12.3 | 14 | 13.7 | 14 | 0.42 | 15 | 0.61 | 16 |
| 013-032 | 528.7 | 7 | 580.6 | 5 | 2.34 | 10 | 1.43 | 4 | 12.5 | 12 | 14.2 | 12 | 0.42 | 14 | 0.34 | 11 |
| Robijs | 523.6 | 8 | 554.8 | 9 | 3.7 | 14 | 3.68 | 14 | 12.4 | 13 | 14.1 | 13 | 0.30 | 8 | 0.33 | 9 |
| 013-074 | 521.4 | 9 | 564.4 | 8 | 1.98 | 7 | 2.30 | 7 | 12.5 | 11 | 14.5 | 6 | 0.17 | 1 | 0.17 | 3 |
| 990-2 | 507.4 | 10 | 528.6 | 13 | 3.48 | 13 | 1.04 | 1 | 12.7 | 8 | 14.3 | 8 | 0.31 | 10 | 0.13 | 1 |
| Hiie | 505.9 | 11 | 511.5 | 15 | 1.20 | 2 | 3.20 | 11 | 12.6 | 10 | 14.2 | 10 | 0.21 | 3 | 0.14 | 2 |
| Zombi | 496.6 | 12 | 533.1 | 11 | 1.49 | 3 | 1.14 | 3 | 13.1 | 3 | 14.5 | 5 | 0.30 | 9 | 0.28 | 8 |
| Caress | 493.9 | 13 | 548.4 | 10 | 1.85 | 6 | 1.86 | 5 | 12.7 | 6 | 14.3 | 9 | 0.21 | 4 | 0.22 | 6 |
| Voore | 489.3 | 14 | 532.1 | 12 | 2.13 | 9 | 1.06 | 2 | 13.1 | 4 | 14.7 | 3 | 0.24 | 7 | 0.19 | 4 |
| 876 | 477.9 | 15 | 516.4 | 14 | 1.77 | 5 | 2.43 | 8 | 14.0 | 1 | 16.0 | 1 | 0.52 | 16 | 0.48 | 13 |
| Runar | 422.3 | 16 | 454.5 | 16 | 2.09 | 8 | 3.59 | 13 | 13.5 | 2 | 15.6 | 2 | 0.32 | 11 | 0.52 | 14 |
| Mean | 515.8a | × | 553.9b | × | 2.46 | × | 2.54 | × | 12.7b | × | 14.4a | × | 0.3 | × | 0.32 | × |
| LSD | 29.03 | × | 34.2 | × | × | × | × | × | 0.26 | × | 0.38 | × | × | × | × | × |
*R – ranking order; Mean values of GY and GPC followed by the different letter within columns of two N rates (N75; N150) are significantly different (p<0.05); N75 – 75 kg N ha-1; N150 – 150 kg N ha-1; LSD - least significant difference (α=0.05).
Table 6.
Mean values of plant height (PH), days to heading (DH) and NDVI at four growth stages and their ranking for genotypes averaged over 16 environments (combination of locations x years x N rates).
Table 6.
Mean values of plant height (PH), days to heading (DH) and NDVI at four growth stages and their ranking for genotypes averaged over 16 environments (combination of locations x years x N rates).
| Genotype* | PH, cm | R | DM, days | R | NDVI | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| GS25 | R | GS55 | R | GS75 | R | GS85 | R | |||||
| DS-17-16-DH | 86.3 | 12 | 106.9 | 5 | 0.512 | 11 | 0.886 | 8 | 0.809 | 5 | 0.427 | 5 |
| DS-655-7-DH | 91.4 | 5 | 107.8 | 4 | 0.533 | 3 | 0.896 | 4 | 0.823 | 3 | 0.433 | 4 |
| DS-720-3-DH | 90.1 | 7 | 108.6 | 2 | 0.526 | 6 | 0.897 | 3 | 0.822 | 4 | 0.449 | 3 |
| 013-01 | 88.5 | 8 | 108.4 | 3 | 0.493 | 15 | 0.904 | 2 | 0.837 | 1 | 0.491 | 2 |
| DS-638-5-DH | 85.5 | 14 | 111.0 | 1 | 0.504 | 13 | 0.910 | 1 | 0.833 | 2 | 0.511 | 1 |
| Betong | 86.1 | 13 | 104.9 | 10 | 0.518 | 8 | 0.877 | 12 | 0.770 | 11 | 0.379 | 10 |
| 013-032 | 86.4 | 11 | 106.6 | 6 | 0.505 | 12 | 0.891 | 6 | 0.805 | 6 | 0.405 | 8 |
| 013-074 | 88.2 | 9 | 104.8 | 11 | 0.523 | 7 | 0.882 | 10 | 0.767 | 12 | 0.371 | 12 |
| Robijs | 94.0 | 3 | 106.1 | 8 | 0.570 | 1 | 0.894 | 5 | 0.795 | 7 | 0.410 | 6 |
| Caress | 80.4 | 15 | 105.4 | 9 | 0.499 | 14 | 0.883 | 9 | 0.780 | 9 | 0.382 | 9 |
| 990-2 | 91.1 | 6 | 106.2 | 7 | 0.472 | 16 | 0.876 | 14 | 0.780 | 8 | 0.407 | 7 |
| Zombi | 76.9 | 16 | 104.7 | 12 | 0.526 | 5 | 0.889 | 7 | 0.775 | 10 | 0.369 | 13 |
| Voore | 87.5 | 10 | 104.1 | 14 | 0.515 | 10 | 0.876 | 13 | 0.754 | 14 | 0.376 | 11 |
| Hiie | 95.5 | 2 | 104.5 | 13 | 0.532 | 4 | 0.879 | 11 | 0.758 | 13 | 0.360 | 14 |
| 876 | 93.0 | 4 | 102.8 | 16 | 0.516 | 9 | 0.871 | 15 | 0.738 | 15 | 0.357 | 16 |
| Runar | 97.7 | 1 | 103.1 | 15 | 0.568 | 2 | 0.870 | 16 | 0.716 | 16 | 0.360 | 15 |
| LSD | 1.81 | × | 0.48 | × | 0.031 | × | 0.001 | × | 0.019 | × | 0.020 | × |
| Mean | 88.7 | × | 106.0 | × | 0.519 | × | 0.886 | × | 0.785 | × | 0.405 | × |
*Genotypes are ranked based on GY over 16 environments (combination of year x location x N rates); PH – plant height; DM – days to maturity; R – ranking order; N75 – 75 kg N ha-1; N150 – 150 kg N ha-1; GS25 – tillering growth stage; GS55 – heading growth stage; GS75 – milk growth stage; GS85 – dough growth stages. LSD - least significant difference (α=0.05).
Table 7.
Values of RMSE, adjusted R2 and corresponding significance of linear regression analyses in multi-environment trials for the relationships between grain yield (GY) and grain protein content (GPC) as dependent variables and NDVI as the independent variable.
Table 7.
Values of RMSE, adjusted R2 and corresponding significance of linear regression analyses in multi-environment trials for the relationships between grain yield (GY) and grain protein content (GPC) as dependent variables and NDVI as the independent variable.
| Environment* | Grain yield (GY) | Grain protein content (GPC) | ||||||
|---|---|---|---|---|---|---|---|---|
| RMSE | adj. R2 | RMSE | adj. R2 | |||||
| N75 | N150 | N75 | N150 | N75 | N150 | N75 | N150 | |
| GS25 | ||||||||
| EE_2021 | 35.09 | 46.49 | 0.26 ** | 0.16 * | 0.65 | 0.72 | -0.03 | 0.01 |
| EE_2022 | 32.00 | 31.62 | -0.03 | -0.03 | 0.56 | 0.71 | -0.03 | -0.01 |
| LV_2021 | 52.16 | 50.52 | 0.01 | -0.03 | 0.51 | 0.73 | 0.00 | 0.02 |
| LV_2022 | 79.26 | 61.47 | -0.01 | 0.14 * | 0.93 | 0.93 | 0.09 | 0.07 |
| LT_2021 | 57.96 | 50.18 | 0.10 * | 0.49 *** | 0.69 | 0.7 | -0.01 | -0.02 |
| LT_2022 | 70.02 | 71.97 | 0.04 | -0.03 | 0.63 | 0.58 | 0.05 | 0.16 * |
| NO_2021 | 46.24 | 66.82 | 0.00 | 0.00 | 0.93 | 1.48 | -0.01 | 0.08 |
| NO_2022 | 70.96 | 91.32 | 0.11 * | 0.23 ** | 1.23 | 1.18 | 0.12 * | -0.02 |
| GS55 | ||||||||
| EE_2021 | 38.68 | 43.98 | 0.1 * | 0.25 ** | 0.57 | 0.65 | 0.21 ** | 0.18 ** |
| EE_2022 | 31.81 | 24.68 | -0.02 | 0.37 *** | 0.53 | 0.65 | 0.06 | 0.15 * |
| LV_2021 | 47.66 | 43.2 | 0.17 * | 0.25 ** | 0.5 | 0.74 | 0.04 | -0.02 |
| LV_2022 | 76.99 | 66.32 | 0.04 | 0.00 | 0.97 | 0.91 | 0.01 | 0.11 * |
| LT_2021 | 45.60 | 42.24 | 0.44 *** | 0.64 *** | 0.7 | 0.64 | -0.03 | 0.13 * |
| LT_2022 | 41.58 | 67.02 | 0.66 *** | 0.10 * | 0.65 | 0.63 | -0.01 | 0 ns |
| NO_2021 | 42.29 | 49.90 | 0.16 * | 0.44 *** | 0.94 | 1.25 | -0.03 | 0.34 *** |
| NO_2022 | 52.59 | 66.73 | 0.51 *** | 0.59 *** | 1.02 | 1.12 | 0.39 *** | 0.08 |
| GS75 | ||||||||
| EE_2021 | 39.68 | 39.40 | 0.05 | 0.4 *** | 0.53 | 0.54 | 0.32 *** | 0.44 *** |
| EE_2022 | 32.00 | 23.56 | -0.03 | 0.43 *** | 0.55 | 0.66 | 0.00 | 0.12 * |
| LV_2021 | 43.17 | 36.41 | 0.32 *** | 0.47 *** | 0.44 | 0.69 | 0.26 ** | 0.11 * |
| LV_2022 | 70.83 | 61.00 | 0.19 ** | 0.16 * | 0.96 | 0.76 | 0.02 | 0.37 *** |
| LT_2021 | 44.18 | 47.45 | 0.47 *** | 0.54 *** | 0.69 | 0.65 | 0.00 | 0.12 * |
| LT_2022 | 29.96 | 53.13 | 0.82 *** | 0.44 *** | 0.66 | 0.63 | -0.03 | -0.01 |
| NO_2021 | 39.62 | 66.75 | 0.27 ** | 0.00 | 0.93 | 1.56 | -0.01 | -0.03 |
| NO_2022 | 36.08 | 53.46 | 0.77 *** | 0.74 *** | 0.88 | 1.02 | 0.55 *** | 0.23 ** |
| GS85 | ||||||||
| EE_2021 | 41.24 | 51.58 | -0.03 | -0.03 | 0.6 | 0.72 | 0.11 * | 0.01 |
| EE_2022 | 31.93 | 27.36 | -0.03 | 0.23 ** | 0.54 | 0.67 | 0.02 | 0.1 * |
| LV_2021 | 52.28 | 39.91 | 0.00 | 0.36 *** | 0.49 | 0.71 | 0.09 * | 0.07 |
| LV_2022 | 79.74 | 64.50 | -0.03 | 0.06 | 0.99 | 0.96 | -0.03 | -0.01 |
| LT_2021 | 60.01 | 62.53 | 0.03 | 0.2 ** | 0.7 | 0.67 | -0.03 | 0.07 |
| LT_2022 | 68.10 | 71.68 | 0.09 | -0.02 | 0.66 | 0.6 | -0.03 | 0.1 * |
| NO_2021 | 33.68 | 46.16 | 0.47 *** | 0.52 *** | 0.87 | 1.22 | 0.13 * | 0.37 *** |
| NO_2022 | 52.13 | 83.35 | 0.52 *** | 0.36 *** | 0.99 | 0.86 | 0.42 *** | 0.45 *** |
*Environments: 4 locations - Estonia (EE), Latvia (LV), Lithuania (LT), Norway (NO); 2 years - 2021 and 2022; 2 N rates - 75 kg N ha-1, N150 - 150 kg N ha-1) across four crop growth stages: GS25 – tillering growth stage; GS55 – heading growth stage; GS75 – milk growth stage; GS85 – dough growth stages. RMSE - root means square error; * p<0.05; **p<0.01; ***p<0.001.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



