
Submitted:
11 December 2023
Posted:
12 December 2023
You are already at the latest version
Abstract
The viability, productivity and survival of higher plants under the adverse factors influence are largely determined by the functional activity of the antioxidant system. The aim of our work was to investigate changes in formation of high-molecular (superoxide dismutase and peroxidase) and low-molecular (phenolics, including flavanols and proanthocyanidins) antioxidants in callus culture of Camellia sinensis under influence of phenolic precursors (L-phenylalanine – 3 mM, trans-cinnamic acid – 1 mM, naringenin – 0.5 mM). According to the data obtained, precursors effect on tea callus cultures did not lead to significant increasing of superoxide dismutase and peroxidase activity in most cases. However, it led to the increased accumulation of the total phenolics content, as well as flavanols and proanthocyanidins contents. For C. sinensis callus cultures, the most promising regulator of phenolic compounds was L-phenylalanine, in the presence of which its content increased almost twice. Thus, the exogenous effect of various precursors is possible to use for the targeted regulation of the certain phenolics classes accumulation in plant cells.
Keywords:
tea plant
; callus culture
; L-phenylalanine
; trans-cinnamic acid
; naringenin
; superoxide dismutase
; peroxidase
; flavanols
; proanthocyanidins
1. Introduction
Plants in the process of their growth and development are constantly exposed to various environmental factors, which leads to changes in their metabolism and functional activity [1]. This also applies to the antioxidant system, which protects them from the action of reactive oxygen species (ROS) such as superoxide radical (radical anion) О2•-, hydroperoxide radical НО2•, hydroxyl radical НО•, hydrogen peroxide Н2О2, singlet oxygen 1О2 [2,3,4]. These molecules are formed under conditions of normal plant growth during respiration and photosynthesis, however under stressful conditions, the balance between their formation and inactivation is disturbed. The accumulation of these molecules leads to the so-called "oxidative explosion", causing changes in metabolism, initiation of pathological processes, necrotic damage to vegetative and generative organs, and even death of plants [2,4].
Despite all these changes, plants remain viable, which is largely due to the functional activity of their antioxidant system [3]. Its main components are high-molecular and low-molecular antioxidants, which, being present in lower concentrations compared to the oxidized substrate, significantly delay or inhibit its oxidation [4].
High-molecular metabolites of plant tissues are represented by various enzymes, which include superoxide dismutase (SOD), various peroxidases, catalase and other enzymes [3,4]. Among them, SOD scavenging the superoxide radical (О2•–) is considered to be the "primary" protector of cells from the action of ROS [5]. An important role is also assigned to peroxidases, the functional role of which is associated with the neutralization of hydrogen peroxide formed in cells – a very toxic compound for them [2,3].
Low-molecular antioxidants include various compounds of primary and secondary plant metabolism (carotenoids, tocopherols, polyamines, polyphenols, etc.) that can slow down or prevent the oxidation of other compounds [3]. Among low molecular weight antioxidants, polyphenols or phenolic compounds (PCs) – compounds of secondary metabolism synthesized in all plant cells – have attracted great interest [6]. PCs, due to their chemical properties and interaction with ROS, are able to reduce their level, thereby affecting the redox mechanisms in plants and, as a consequence, maintaining their viability [7,8].
Phenolics are extremely diverse in structure and chemical properties [9,10]. They are represented by different classes such as phenylpropanoids, flavonoids and proanthocyanidins [8]. Presently the great success has been achieved in study of their metabolism: the sequence of their formation and the enzymes involved in these reactions have been established (Figure 1). It is well known the role of these metabolites in protecting against UV radiation, various pathogens, mechanical damage and resistance to various external influences is known [6,9]. They are also involved in the formation of plant cell walls and may serve as storage [8]. The issue on the content and composition of the PCs metabolites is of great importance [3,8,9].
In general, the balance of high-molecular and low-molecular antioxidants ensures the viability of plants and their resistance to stress [2,4,6]. At the same time, if the level of high-molecular antioxidants is associated with the "protection" of cells from ROS, then the accumulation of low-molecular metabolites of phenolic nature is also of practical importance, since many of their representatives can be used as pharmacologically valuable drugs for the health of the population [11,12].
One of the promising approaches to the study of plant metabolism is plant cell and tissue cultures grown in vitro [13]. This is due to the fact that they represent a “simpler” biological system compared to a plant, which is grown under strictly controlled conditions and retains the specificity of the metabolism of an intact plant [14]. Using the action of various exogenous factors, it is possible not only to study the mechanisms of its regulation, but also to select conditions for increasing the productivity of cell cultures as potential producers of biologically active plant metabolites for pharmacology [13]. These include the action of biosynthesis precursors of various compounds, including PCs [13,15,16]. However, it is worth noting that the effectiveness of this action depends on the concentration of the compound, the duration of its exposure and the species-specific characteristics of cultures [17,18,19].
The tea plant (Camellia sinensis L.) is one of the unique plants with a specialized metabolism aimed at the formation of PCs [20,21,22]. It is known that the proportion of these secondary metabolites in its tissues can reach 30% by dry weight [23]. C. sinensis callus cultures also retain the ability to form PCs, although at a lower level (no more than 10% by dry weight) [24]. It should also be emphasized that the main components of their phenolic complex are flavans, or flavanols – substances with P-vitamin capillary-strengthening activity. All this indicates that in vitro culture of the tea plant is not only a good model object for studying the regulation of phenolic metabolism, but also a potential producer of biologically active compounds.
Considering all of the above, the purpose of our work was to study the effect of the PCs precursors (L-phenylalanine, trans-cinnamic acid, naringenin) on the antioxidant potential of the C. sinensis callus culture, investigating changes in formation of high-molecular (superoxide dismutase and peroxidase) and low-molecular (phenolics, including flavanols and proanthocyanidins) antioxidants.
2. Results
2.1. Morphological Characteristics of Tea Cells Cultures
Comparison of morphological parameters of in vitro plant cultures exposed to exogenous factors is an important indicator of assessing their physiological state [25]. The tea callus cultures grown in vitro in a liquid nutrient medium were compact, quite dense and had a beige color (Figure 2).
On the main nutrient medium or on the medium with additional L-phenylalanine (PhA), they had a light beige colour. On the medium with naringenin (NG), the calluses were darker in color, and on the medium with trans-cinnamic acid (CA), they were denser and dark beige. All this indicates minor differences in the morphology of tea callus cultures grown on media with various precursors of PCs, relative to the control.
The water content determination in tea callus culture almost did not reveal significant differences between the variants (Table 1). One can only note a statistically significant increase in the water content of the culture under the NG action.
2.2. The Antioxidant System of Tea Cells Cultures
2.2.1. The High-Molecular Antioxidants in Tea Cells Cultures
Determination of the superoxide dismutase (SOD) activity in tea callus cultures revealed statistically significant differences between the control and two experimental (CA and NG) variants (Figure 3a). The highest SOD activity was noted in calli grown on the medium with NG, where it was almost 2.5 times higher the control variant. The CA effect was manifested at a very low level of the SOD activity in tea cells culture. As for the calli exposed to PhA, the enzyme activity was statistically equal to the control values.
Our research also indicated that the peroxidase (POX) activity was the highest in tea cells culture of the control variant (Figure 3b). In the experimental variants, there was a statistically significant trend towards a decrease in POX activity in the series: control-PhA-CA-NG. This decrease in tea cells culture grown on the medium with PhA amounted to 15%, on the medium with CA, to 24%, and on the medium with NG, to 40%.
2.2.2. The Low-Molecular Antioxidants in Tea Cells Cultures
According to the obtained data, when growing tea cells cultures on media with phenolic precursors, the total phenolic content was significantly higher than in the control (Figure 4a). The greatest increase in this parameter was noted on the medium with PhA, when it exceeded the control data twice. In the presence of NG, the content of PCs, increased by 79%, and in tea cells culture treated with CA – by 23%, relative to the control values.
As for the flavanols content, the lowest its accumulation was noted in the tea cells culture of the control (Figure 4b). While in all experimental variants, it was significantly higher. Thus, in the presence of PhA, its content exceeded the control value by 150%. A fairly close effect was observed with the action of NG (an excess by 130%). In the CA variant, this increase was 180%.
Determination of the proanthocyanidins content in tea cells cultures also showed the stimulating effect of all precursors (Figure 4c). The highest its amount was observed in calluses grown on the media with PhA and NG, which almost equally exceeded the values of the control variant (2.3 and 2.2 times, respectively). At the same time, the action of CA increased the content of these compounds only 1.6 times relative to the control variant.
2.3. The Phenylalanine Ammonia-Lyase Activity in Tea Cells Cultures
In experimental variants, the activity of phenylalanine ammonia-lyase (PAL) exceeded that of the control and this effect depended on the precursor added to the nutrient medium (Figure 5). The greatest activity of the enzyme was noted in the callus on the medium with CA, which exceeded the control value by more than 2 times (228%). On the medium with PhA, the enzyme activity in tea callus culture exceeded the control value by 63%, and on the medium with NG – by 25%.
2.4. The Level of Lipid Peroxidation in Tea Cells Cultures
According to our data, the lipid peroxidation (LPO) level was statistically equal in calluses of the control variant and those exposed to PhA and CA treatment (Figure 6). The only exception was the variant with the action of NG, where the level of LPO was twice as high as the control.
3. Discussion
3.1. Morphological Characteristics of Tea Cells Cultures
The water content is an important parameter of the viability of plant cells and tissues. It is worth noting that statistical processing of the data did not reveal a significant effect of treatment with different precursors on this parameter (Table 2).
However, there was a statistically significant increase (2%) in the water content of the culture under the NG action (Table 1). It should be emphasized here that a change in this parameter within 5% does not indicate significant changes in the physiological processes of cells [15]. The described effect of NG is confirmed by the data obtained in the work of Sharma et al. where a similar trend was noted [26]. At the same time, there was shown no change in water content using the example of Carthamus tinctorius seedlings in the presence of the indicated above precursor [27]. It is worth noting that for PhA, a similar to our data effect was noted for the Vitex agnus castus shoot cultures and Cicer arietinum seedlings [28,29]. While the CA effect on the example of Pisum sativum was the opposite and revealed in a decrease in water content [30].
Based on all of the above, it can be concluded that short-term exposure to various precursors of phenolic nature did not cause significant changes in the morphological characteristics of tea callus cultures. Perhaps this effect is due to the regulation of their pro-/antioxidant balance, which is important in maintaining the viability of plant cells [31]. And in this case, an important role is assigned to the functioning of the antioxidant system in plant cells, the main components of which are high-molecular and low-molecular antioxidants [32].
3.2. The Antioxidant System of Tea Cells Cultures
It is known that increased accumulation of ROS causes the development of oxidative stress due to an imbalance between the ROS generation and removal [3]. This leads to a number of pathological processes and plant diseases, necrotic damage to vegetative and generative organs, and even plant death [4]. The main damaging effects of ROS at the cellular level are damage to nucleic acids through the oxidation of deoxyribose, damage to organelles, peptide bond breaks, initiation of lipid peroxidation processes that cause an increase in membrane viscosity and disruption of diffusion processes, accumulation of damaged and self-aggregating proteins. At high levels of ROS accumulation, apoptosis or programmed cell death can occur [3,33].
In order to utilize ROS, plants are able to synthesize various types of antioxidants, enzymatic and non-enzymatic [34]. In this case, the functioning of the organism’s protective systems is of great importance, one of which is the antioxidant system, which includes numerous antioxidants that can slow down or prevent the oxidation of organic substances [3]. The antioxidant system functioning can be regulated by exposure to various exogenous compounds, including phenolic precursors, which is discussed in detail in the sub-chapters below [16,35,36].
3.2.1. The High-Molecular Antioxidants in Tea Callus Cultures
Superoxide dismutase (SOD) is considered to be one of the main high-molecular antioxidants of plant cells and even the "primary" line of their protection against oxidative damage [3]. This enzyme prevents the oxidation of macromolecules by superoxide radicals [5].
According to the statistical processing, the different phenolic precursors treatment influenced the SOD activity (Table 3). Under the CA effect the SOD activity didn’t exceed the control and even was minimal, which to a certain extent correlate with data on Cucumis sativus seedlings, where the absence of changes in the enzyme activity was noted [37]. As for the effects of NG, there are several works confirming our data: a small but statistically significant increase in the activity of this enzyme was shown in Carthamus tinctorius seedlings, while this trend was more pronounced in Phaseolus vulgaris seedlings (almost a two-fold increase relative to the control) [27,36]. However, the PhA treatment did not result any statistically significant changes in relation to control values.
High molecular weight antioxidants also include peroxidases (POX) involved in the detoxification one of the ROS (H2O2) in plant cells [30,38]. According to the obtained data, the POX activity depended on the used precursor (Table 3).
Thus, our data indicate a decrease in POX activity when growing tea callus cultures on a medium with phenolic precursors. A similar trend was noted for Phaseolus vulgaris seedlings, in which, after the action of NG, it decreased by almost three times relative to the control [36]. Using Cucurbita ficifolia seedlings as an example, it was shown that CA treatment did not lead to statistically significant changes in POX activity, as well as PhA treatment, which did not affect this indicator in tomato leaves [37,39]. There are other data as well. Thus, the activity of this enzyme was significantly increased in Fagopyrum esculentum sprouts and in Triticum aestivum leaves under the action of PhA, as well as in tomato, Pisum sativum and Cucumis sativus seedlings under the action of CA [16,30,37,38,40].
There should be also emphasized the tendency of each precursor individually on the both enzymes’ activities. The PhA treatment did not affect the both ones, and the CA action led to the decrease in both cases, while the NG effect revealed in the low POX activity and in the highest SOD activity.
All of the above indicates that the PhA and CA effects did not lead to an increase in the activity of SOD, whereas the presence of NG in the culture medium led to a significant increase in the activity of this enzyme of the antioxidant system. Thus, we can say that the presence of phenolic precursors in the culture medium led to the changes in the activity of such high-molecular antioxidants as SOD and POX. It is worth noting that in the case of the latter enzyme, only a significant decrease in its activity was shown in all experimental variants. As for the activity of such a primary enzyme of the antioxidant system as SOD, the increase in its activity was characteristic only of the NG treatment.
3.2.2. The Low-Molecular Antioxidants in Tea Cells Cultures
The PCs are one of the effective low-molecular antioxidants, which are considered as the "second" line of defense of plant cells from the action of ROS [41,42]. Due to their chemical properties, they interact with active oxygen forms, thereby reducing their toxic effect on cells [9,10]. The statistical processing has shown that the content of different phenolics depended on the treatment with different precursors (Table 4).
Determination of the total phenolic content in tea callus cultures is an important indicator in assessing their ability to accumulate these secondary metabolites [43,44]. On the example of Cicer arietinum leaves and Sequoia sempervirens callus culture, the increase in its content was shown in the presence of PhA, which was similar to our data [35]. While in the cells of the Moringa oleifera plant this trend was significantly less pronounced, herewith in quinoa sprouts there was no statistically significant difference between the experimental and control variants [29,45]. There are data confirming the increase in the PCs accumulation in Carthamus tinctorius and Vigna radiata seedlings in the presence of NG, while in the first case this indicator increased almost two-fold [26,27].
The main components of the phenolic complex, not only of the tea plant, but also of the cultures initiated from it in vitro, are flavanols, which in recent years have also been considered as substances with antioxidant activity [46,47]. According to the data described in the results section, the intake of all phenolic precursors into tea calluses contributed to the activation of the phenolic metabolism, which was accompanied by the accumulation of flavanols characteristic of them. At the same time, the most effective "regulators" of this process were PhA (the initial stages of biogenesis) and NG – an important intermediate of the flavonoid pathway. The example of Triticum aestivum leaves also showed a significant increase in the content of flavanols in the presence of PhA [16].
The tea plant and the callus cultures initiated from it are also characterized by the formation of oligomeric forms of PCs – proanthocyanidins [47,48]. The data obtained by us on the action of PhA are confirmed in the work of Feduraev et al. on the example of wheat leaves [16].
As for the individual effect of each precursor, the upregulating effect of PhA was the most pronounced on all phenolic classes. NG had a similar to PhA effect, which was a little bit less noticeable. While CA treatment had the least expressed effect on these parameters.
All of the above indicates that the cultivation of tea callus culture on a medium with various precursors was accompanied by an increase in the accumulation of low-molecular phenolic antioxidants in them, including flavanols, substances with P-vitamin capillary-strengthening activity. The intensity of this process may depend on the activity of the enzymes involved in their biosynthesis.
3.3. The Phenylalanine Ammonia-Lyase Activity in Tea Cells Cultures
It is known that phenylalanine ammonia-lyase (PAL) is an important enzyme in the initial stages of the PCs biosynthesis [49]. Based on the above presented data about the effect of precursors on the accumulation of phenolic compounds in callus tea cultures, we analyzed its activity in all studied variants (Figure 5). The statistical processing of the data revealed a significant effect of treatment with different precursors on this parameter (Table 5).
In experimental variants, the activity of PAL exceeded that of the control and this effect depended on the precursor added to the nutrient medium. The greatest activity of the enzyme was noted in the callus on the medium with CA, which exceeded the control value by more than 2 times (228%). A different trend was observed in Cichorium intybus calluses when PAL activity decreased significantly relative to control in the presence of CA [50]. On the medium with PhA, the enzyme activity in tea callus culture exceeded the control value by 63%, and on the medium with NG – by 25%. A similar effect of PhA on the PAL activity was observed in the leaves of Ocimum basilicum, Triticum aestivum and in the shoots of Chenopodium quinoa [16,46,51]. It is worth noting that it was significantly more pronounced relative to the control variant, whereas in the case of buckwheat sprouts, this increase was statistically unreliable [40]. And on the example of Glycine max seedlings, a statistically significant decrease in PAL activity in the presence of NG was recorded [49].
It is worth noting that the highest enzyme activity was shown under the CA treatment, although the phenolic compounds content was the lowest one under this precursor effect. Perhaps in this case, further pathways of biosynthesis of polyphenolic compounds, for example lignin and its derivatives, has been activated. This issue is a subject for further research.
Thus, the cultivation of tea callus culture on a medium with phenolic precursors is accompanied by the activation of the key polyphenol biosynthesis enzyme and, as a result, the accumulation of phenolic compounds in them.
3.4. The Level of Lipid Peroxidation in Tea Cells Cultures
Changes in the growing conditions of plant cells, including the administration of additional components into the nutrient medium, can cause changes in the functioning of their antioxidant system [36]. The criterion of this process is the level of lipid peroxidation (LPO), often assessed by the content of malondialdehyde (MDA) [27]. The statistical processing has shown that LPO level depends on the precursor treatment (Table 6). It worth noting that there was a statistically significant change (increasing) in MDA content only under NG action (Figure 6), in other cases its content was equal to the control values.
The data obtained by us are confirmed in a variety of works. Thus, using lentil sprouts and tomato plants as an example, it was shown no statistically significant changes in the LPO level under PhA exposure [39,52]. Also, CA treatment did not lead to significant changes in this parameter in Cucurbita ficifolia seedlings and Tanacetum parthenium leaves [37,53]. However, a different trend was shown on the example of Cucumis sativus seedlings: in this case, the MDA content significantly exceeded the control values [37]. As for the action of NG, which caused the increase in the LPO level, according to our data, different reactions of plant objects to the presence of this precursor are shown in other works. Thus, its effect did not lead to changes in this parameter in the seedlings of Vigna radiata and Phaseolus vulgaris [26,36]. Nevertheless, there are data on the NG treatment of Carthamus tinctorius seedlings, which are similar to the results we obtained [27].
Thus, it can be concluded that the effect of precursors on the LPO level depends both on the nature of this molecule and on the species specificity of the object under study. Moreover, in the case of tea plant cells, the stress effect was shown only under the influence of NG, which was not reflected in morphological parameters, however, a slight increase in water content was noted.
4. Materials and Methods
4.1. Plant Material and Experimental Conditions
The object of the study, a heterotrophic callus culture, was obtained from the stem of a tea plant young shoots (Camellia sinensis L., Georgian variety). Heller culture medium containing 5 mg/L 2,4-dichlorophenoxyacetic acid, 25 g/L glucose and 7 g/L agar was used for its cultivation [47]. The calluses were grown under the conditions of the growth cabinet at the IPP RAS (25 °C, relative air humidity 70%, darkness 24 h). The duration of the subcultivation was 39 days.
To set up the experiment (Figure 7), callus tissues (the weight of each callus was 220–250 mg) were placed in conical flasks (100 mL). Every flask contained the liquid Heller culture medium (15 mL) of the basic composition (control) or enriched with PhA (3 mM), CA (1 mM) or NG (0.5 mM). The precursors (Serva, Germany) were dissolved in distilled water beforehand, then it was sterilized through membrane filters (MILLEX GV, 0.22µm, Merck, Darmstadt, Germany) and added to the autoclaved liquid Heller culture medium. Precursors’ concentrations were selected in preliminary experiments. The tea callus cultures were grown under conditions in the growth cabinet on a shaker at a frequency of 90 rpm (25 °C, relative air humidity 70%, darkness 24 h) and analyzed on the 7th day. For biochemical studies, callus tissues were frozen with liquid nitrogen and stored at –70 °C.
4.2. Determination of Morphological Characteristics of Tea Cells Cultures
To evaluate the morphological characteristics of tea callus cultures, such indicators as appearance, color and texture were used.
4.3. Determination of Water Content and Dry Weight in Tea Cells Cultures
The tea callus (150 mg FW each) was dried in the thermostat BD-115 (Binder, Tuttlingen, Germany) to a constant weight (70 °C, 48 h). Calculation of water amount and dry weight was carried out using the standard methods [55,56].
4.4. Determination of the Superoxide Dismutase Activity in Tea Cells
The superoxide dismutase (SOD) activity was evaluated by the level of inhibition by superoxide dismutase of the nitroblue tetrazolium (NBT) reduction reaction to formazane by superoxide radicals generating the riboflavin oxidation system [57]. To determine the enzyme activity the frozen material (200 mg FW each) was homogenized in 0.067 M K, Na-phosphate buffer (рН 7,8) at a temperature of +4°C. The homogenate was centrifuged at 7000×g for 20 min (centrifuge Minispin, Göttingen, Germany), the supernatant was used to determine the enzyme. The reaction was carried out by adding to 60 µl supernatant (enzymatic extract) 0.95 ml 0.1 M methionine, 0.32 ml 2 mM NBT and 0.23 ml 0.012 mM riboflavin [58]. The mixture was exposed to fluorescent light for 30 min. The solution absorbance was measured at 560 nm. It was expressed in U/mg protein. One SOD unit was defined as the amount of enzyme required to inhibit 50% of the nitro blue tetrazolium photoreduction in comparison with tubes lacking the plant extract [57,58]. The protein content was determined by the Bradford method [59].
4.5. Determination of the Peroxidases Activity in Tea Cells Culture
To determine the activity of peroxidases (POX) samples of frozen in liquid nitrogen tea callus culture (500 mg FW each) were homogenized in a 0.067 M K,Na-phosphate buffer (pH 7,8) with the addition of insoluble polyvinylpyrrolidone (50-75% by weight of fresh tissue; Serva, Germany). The homogenate was centrifuged at 12000×g for 20 min (centrifuge Minispin, Göttingen, Germany). The obtained supernatant was used to determine the enzyme activity. These manipulations were carried out at a temperature of +4°C. The activity of POX was determined by the change in optical density (wavelength 470 nm) in a reaction mixture of the following composition: 650 µl of 5 mM hydrogen peroxide (Chimmed, Russia), 650 µl of 4 mM guaiacol (Merck, Germany) solution and 400 µl of the initial supernatant [60]. Reaction time was 3 minutes. The POX activity was estimated due to the formation of the reaction product, tetraguaiacol. The enzyme activity was expressed in U/mg protein [61]. The protein content was determined by the Bradford method [59].
4.6. Extraction of Phenolic Compounds from Tea Cells Culture
Samples of frozen in liquid nitrogen tea callus culture (50 mg FW each) were ground in a porcelain mortar, extracted with 96% ethanol [62]. The homogenate was kept at 45 °C (thermostat Gnom, Moscow, Russia) for 30 min; it was centrifuged at 12,000×g for 5 min (centrifuge Minispin, Göttingen, Germany). The sediment was separated and the supernatant was used to determine various phenolic compounds by spectrophotometric methods.
4.7. Determination of Different Phenolic Compounds Classes in Tea Cells Culture
The determination of the total phenolic content quantity was carried out with the Folin–Ciocalteu reagent (Panreac, E.U.) at 725 nm according to our modified method, using a four-point calibration curve of the gallic acid standard (Serva, Germany) [63]. The amount of flavanols was determined with a 1% vanillin (Merck, Germany) in 70% sulfuric acid at 500 nm, using a four-point calibration curve of the epicatechin standard (Serva, Germany) [24]. Proanthocyanidins were determined with butanol reagent (n-butanol:HCl, 95:5 v/v) at 550 nm, using a four-point calibration curve of the cyanidine standard (Sigma, USA) [64].
The total phenolic content was expressed in mg of gallic acid equivalents per g of dry weight (mg GAE·g-1 DW), the content of flavanols in mg of epicatechin equivalents per g of dry weight (mg ECE·g-1 DW), the content of proanthocyanidins in mg of cyanidine equivalents per g of dry weight (mg CE·g-1 DW).
4.8. Determination of the L-Phenylalanine Ammonia-Lyase Activity in Tea Cells Culture
The L-phenylalanine ammonia-lyase (PAL) activity was determined by homogenization of the frozen material (200 mg FW each) in a 0.1 M Na-borate buffer (pH 8.8) containing 0.5 mM EDTA (Reanal, Hungary) and 3 mM dithiothreitol (Reanal, Hungary) with the addition of insoluble polyvinylpyrrolidone (50% by weight of fresh tissue; Serva, Germany) [62]. The homogenate was filtered and centrifuged at 13000×g for 40 min (centrifuge Minispin, Göttingen, Germany) and then the supernatant was used to determine the enzyme activity. These manipulations were carried out at a temperature of +4°C. The PAL activity was determined by the change in optical density (wavelength 290 nm) in a reaction mixture of the following composition: supernatant, 0.1 M Na-borate buffer, 0.02 M L-phenylalanine in ratio 1:1:1 (Serva, Germany). The PAL activity was evaluated by the formation of trans-cinnamic acid from L-phenylalanine for 1 hour. The PAL activity was expressed in U/mg protein [62]. The protein was analyzed according to Bradford [59].
4.9. Determination of the Level of Lipid Peroxidation in Tea Cells Culture
The level of lipid peroxidation was assessed by a reaction with thiobarbituric acid (Dia-m, Russia) and content of its product – malonyl dialdehyde (MDA). Frozen plant material (200 mg FW each) was homogenized in 5 mL of 0.1 M Tris-HCl buffer (pH 7.5; Reanal, Hungary) with 0.35 M NaCl. Then the reaction of the obtained homogenate with 0.5% solution of TBA in a 20% aqueous solution of trichloroacetic acid was carried out. The reaction mixture was kept on a water bath for 30 min, the optical density of the solution was estimated at 532 nm [65]. A molar extinction coefficient equal to 1.56 × 10−5 M–1 cm–1 was used to count the MDA content (µmol MDA g−1 DW) [66].
4.10. Statistical Analysis
All determinations were carried out in three biological and three analytical replicates. The obtained data were statistically processed using Microsoft Excel 2010 14.0 (Redmond, WA, USA) and SigmaPlot 12.2 (Technology Networks, Sudbury, UK) software. The figures show the arithmetic means ± standard deviations (±SDs). Statistical analyses of data were performed using one-way analysis of variance (ANOVA). Mean separation was performed using Normality Test (Shapiro–Wilk) and all Pairwise Multiple Comparison Procedures (Holm-Sidak method). The significant differences at p < 0.05 are denoted by different Latin letters.
5. Conclusions
The antioxidant system plays an important role in preserving the vital activity of plants, as well as their adaptation to changing environmental conditions. Their main components are high-molecular and low-molecular antioxidants, the balance of which determines the presence of active forms of oxygen in the cell and their detoxification. At the same time, high–molecular antioxidants, represented by various enzymes, are considered as the first line of defense, and low-molecular – as the second and possibly, additional ones, the functional activity of which manifests itself at a later stage of exposure to various factors. And this thesis fully corresponds to the data we have obtained for tea cells cultures. After different precursors exposure there was shown a change in enzyme (high-molecular) antioxidants activity. Herewith, the SOD activity varied in two experimental variants: CA treatment contributed to its lowest values, NG treatment – enhanced it. In case of the POX activity, it was always lower under the precursors action than in the control variant. However, the accumulation of low-molecular antioxidants of phenolic nature increased, as well as the activity of the key enzyme of their biosynthesis – PAL. Perhaps it was the accumulation of these metabolites that served as the basis for maintaining the level of LPO in the in most studied samples at the control level.
It is also important to note the fact that using various precursors of phenolic compounds as components of nutrient media for growing plant cell cultures, it is possible, to a certain extent, to regulate the accumulation of substances with antioxidant activity in them, in particular polyphenols. For tea callus cultures, the most promising regulator of their accumulation was PhA, in the presence of which their content increased almost 2 times and manifested itself not only at the level of the total content of these metabolites, but also at the level of flavanols – substances with P-vitamin capillary-strengthening activity.
Based on this, we consider it important and promising to study the regulation of phenolic metabolites under the action of precursors using high-performance liquid chromatography method, as well as molecular genetic approaches. Of great interest is a more detailed study of the balance of high and low molecular weight antioxidants.
Author Contributions
Conceptualization, M.A.A. and N.V.Z.; methodology, M.A.A., T.L.N. and N.V.Z.; formal analysis, M.A.A., T.L.N., V.V.K., E.A.G., P.V.L., and N.V.Z.; investigation, M.A.A., T.L.N., E.A.G., M.Y.Z., V.V.K. and P.V.L.; resources, N.V.Z.; data curation, M.A.A., T.L.N., E.A.G., P.V.L., A.F. and N.V.Z.; writing—original draft preparation, M.A.A., E.A.G. and N.V.Z.; writing—review and editing, M.A.A., E.A.G., A.F. and N.V.Z.; visualization, M.A.A.; supervision, M.A.A., A.F. and N.V.Z.; project administration, M.A.A. and N.V.Z.; funding acquisition, A.F. and N.V.Z. All authors have read and agreed to the published version of the manuscript.
Funding
This research was supported by assignments 122042600086-7 (IPP RAS) of the Ministry of Science and Higher Education of the Russian Federation and by Russian Science Foundation (grant № 23-14-00383).
Data Availability Statement
The data presented in this study are available on request from the corresponding author.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Mareri, L.; Parrotta, L.; Cai, G. Environmental Stress and Plants. Int. J. Mol. Sci., 2022, 23, 5416. [CrossRef]
- Mittler, R., Zandalinas, S. I., Fichman, Y., Van Breusegem, F. Reactive oxygen species signalling in plant stress responses. Nature Reviews Molecular Cell Biology, 2022, 23, 663-679. [CrossRef]
- Hasanuzzaman, M.; Bhuyan, B.; Zulfiqar, F.; Raza, A.; Mohsin, S.; Mahmud, J.; Fujita, M.; Fotopoulos, V. Reactive oxygen species and antioxidant defense in plants under abiotic stress: Revisiting the crucial role of a universal defense regulator. Antioxidants, 2020, 9, 681. [CrossRef]
- Huang, H., Ullah, F., Zhou, D. X., Yi, M., Zhao, Y. Mechanisms of ROS regulation of plant development and stress responses. Frontiers in plant science, 2019, 10, 800. [CrossRef]
- Alscher R.G., Erturk N., Heath L.S. Role of superoxide dismutases (SODs) in controlling oxidative stress in plants. J. Exp. Botany, 2002, 53, 1331–1341. [CrossRef]
- Cheynier, V.; Comte, G.; Davies, K.; Lattanzio, V.; Martens, S. Plant phenolics: Recent advances on their biosynthesis, genetics, and ecophysiology. Plant Physiol. Biochem. 2013, 72, 1–20. [CrossRef]
- Tanase, C.; Coarcă, S.; Muntean, D. A critical review of phenolic compounds extracted from the bark of woody vascular plants and their potential biological activity. Molecules, 2019, 24, 1182. [CrossRef]
- Tijjani, H., Zangoma, M. H., Mohammed, Z. S., Obidola, S. M., Egbuna, C., Abdulai, S. I. Polyphenols: Classifications, biosynthesis and bioactivities. Functional foods and nutraceuticals: Bioactive components, formulations and innovations, 2020, 389-414. [CrossRef]
- Kumar, S., Abedin, M. M., Singh, A. K., Das, S. Role of phenolic compounds in plant-defensive mechanisms. Plant Phenolics in Sustainable Agriculture, 2020, Volume 1, 517-532. [CrossRef]
- Kruk, J., Aboul-Enein, B. H., Duchnik, E., Marchlewicz, M. Antioxidative properties of phenolic compounds and their effect on oxidative stress induced by severe physical exercise. The Journal of Physiological Sciences, 2022, 72, 1-24. [CrossRef]
- Cosme, P., Rodríguez, A.B., Espino, J., Garrido, M. Plant phenolics: Bioavailability as a key determinant of their potential health-promoting applications. Antioxidants, 2020, 9, 1263. [CrossRef]
- Zhang, Y.; Mu, T.; Deng, X.; Guo, R.; Xia, B.; Jiang, L.; Wu, Z.; Liu, M. New Insights of Biological Functions of Natural Polyphenols in Inflammatory Intestinal Diseases. Int. J. Mol. Sci., 2023, 24, 9581. [CrossRef]
- Isah, T.; Umar, S.; Mujib, A.; Sharma, M.; Rajasekharan, P.; Zafar, N.; Frukh, A. Secondary metabolism of pharmaceuticals in the plant in vitro cultures: Strategies, approaches, and limitations to achieving higher yield. Plant Cell Tissue Organ Cult. 2018, 132, 239–265. [CrossRef]
- Chandran, H.; Meena, M.; Barupal, T.; Sharma, K. Plant tissue culture as a perpetual source for production of industrially important bioactive compounds. Biotechnol. Rep. 2020, 26, e00450. [CrossRef]
- Murthy, H.; Lee, J., Paek, Y. Production of secondary metabolites from cell and organ cultures: Strategies and approaches for biomass improvement and metabolite accumulation. Plant Cell Tissue Organ Cult. 2014, 118, 1–16. [CrossRef]
- Feduraev, P.; Skrypnik, L.; Riabova, A.; Pungin, A.; Tokupova, E.; Maslennikov, P.; Chupakhina, G. Phenylalanine and tyrosine as exogenous precursors of wheat (Triticum aestivum L.) secondary metabolism through PAL-associated pathways. Plants, 2020, 9, 476. [CrossRef]
- Musbah, H.; Ibrahim, K.; Ibrahim, K. Effects of feeding tyrosine or phenylalanine on the accumulation of polyphenols in Coleus blumei in vivo and in vitro. J. Biotechnol. 2019, 13, 35–43. [Google Scholar] [CrossRef]
- Muthaiya, M.; Nagella, P.; Thiruvengadam, M.; Mandal, A. Enhancement of the productivity of tea (Camellia sinensis) secondary metabolites in cell suspension cultures using pathway inducers. J. Crop. Sci. Biotechnol. 2013, 16, 143–149. [Google Scholar] [CrossRef]
- Twaij, B.; Jazar, Z.; Hasan, M. The effects of elicitors and precursor on in-vitro cultures of Trifolium resupinatum for sustainable metabolite accumulation and antioxidant activity. Biocatal. Agric. Biotechnol. 2019, 22, 101337. [CrossRef]
- Han, W.-Y.; Huang, J.-G.; Li, X.; Li, Z.-X.; Ahammed, G.; Yan, P.; Stepp, J. Altitudinal effects on the quality of green tea in east China: A climate change perspective. Eur. Food Res. Technol., 2016, 243, 323–330. [CrossRef]
- Zhang, L.; Ho, C.; Zhou, J.; Santos, J.; Armstrong, L.; Granato, D. Chemistry and biological activities of processed Camellia sinensis teas: A comprehensive review. Compr. Rev. Food Sci. Food Saf. 2019, 18, 1474–1495. [Google Scholar] [CrossRef] [PubMed]
- Meng, X.; Li, N.; Zhu, H.; Wang, D.; Yang, C.; Zhang, Y. Plant resources, chemical constituents, and bioactivities of tea plants from the genus Camellia section Thea. J. Agric. Food Chem., 2019, 67, 5318–5349. [CrossRef]
- Liu, Y., Zhao, G., Li, X., Shen, Q., Wu, Q., Zhuang, J., Zhang X., Xia E., Zhang Z., Qian Y., Gao L., Xia, T. Comparative analysis of phenolic compound metabolism among tea plants in the section Thea of the genus Camellia. Food Research International, 2020, 135, 109276. [CrossRef]
- Ossipov, V., Zubova, M., Nechaeva, T., Zagoskina, N., Salminen, J. P. The regulating effect of light on the content of flavan-3-ols and derivatives of hydroxybenzoic acids in the callus culture of the tea plant, Camellia sinensis L. Biochemical Systematics and Ecology, 2022, 101, 104383. [CrossRef]
- Bednarek, P. T., Orłowska, R. Plant tissue culture environment as a switch-key of (epi) genetic changes. Plant Cell, Tissue and Organ Culture (PCTOC), 2020, 140, 245-257. [CrossRef]
- Sharma, P., Gautam, A., Kumar, V., Khosla, R., & Guleria, P. Naringenin reduces Cd-induced toxicity in Vigna radiata (mungbean). Plant Stress, 2021, 1, 100005. [CrossRef]
- Hatamipoor, S., Shabani, L., Farhadian, S. Supportive effect of naringenin on NaCl-induced toxicity in Carthamus tinctorius seedlings. International Journal of Phytoremediation, 2023, 25, 889-899. [CrossRef]
- Skrzypczak-Pietraszek, E.; Piska, K.; Pietraszek, J. Enhanced production of the pharmaceutically important polyphenolic compounds in Vitex agnus castus L. shoot cultures by precursor feeding strategy. Engineering in Life Sciences 2018, 18, 287–297. [Google Scholar] [CrossRef] [PubMed]
- Arora, J., Kanthaliya, B., Joshi, A., Meena, M., Meena, S., Siddiqui, M. H., Alamri, S., Devkota, H. P. Evaluation of Total Isoflavones in Chickpea (Cicer arietinum L.) Sprouts Germinated under Precursors (p-Coumaric Acid and L-Phenylalanine) Supplementation. Plants, 2023, 12, 2823. [CrossRef]
- Kapoor, R. T., Alyemeni, M. N., Ahmad, P. Exogenously applied spermidine confers protection against cinnamic acid-mediated oxidative stress in Pisum sativum. Saudi Journal of Biological Sciences, 2021, 28, 2619-2625. [CrossRef]
- Olszowy, M. What is responsible for antioxidant properties of polyphenolic compounds from plants?. Plant Physiology and Biochemistry, 2019, 144, 135-143. [CrossRef]
- Dumanović, J.; Nepovimova, E.; Natić, M.; Kuča, K.; Jaćević, V. The significance of reactive oxygen species and antioxidant defense system in plants: A concise overview. Frontiers in plant science 2021, 11, 552969. [Google Scholar] [CrossRef] [PubMed]
- Kabbage, M., Kessens, R., Bartholomay, L. C., Williams, B. The life and death of a plant cell. Annual Review of Plant Biology, 2017, 68, 375-404. [CrossRef]
- Kolupaev, Y.E.; Kolupaev, Y.E.; Horielova, E.I.; Yastreb, T.O.; Ryabchun, N.I. State of antioxidant system in triticale seedlings at cold hardening of varieties of different frost resistance. Cereal Research Communications 2020, 48, 165–171. [Google Scholar] [CrossRef]
- El-Hawary, S.S.; Abd El-Kader, E.M.; Rabeh, M.A.; Abdel Jaleel, G.A.; Arafat, M.A.; Schirmeiste, T.; Abdelmohsen, U.R. Eliciting callus culture for production of hepatoprotective flavonoids and phenolics from Sequoia sempervirens (D. Don Endl). Nat. Prod. Res. 2020, 34, 3125–3129. [CrossRef]
- Ozfidan-Konakci, C.; Yildiztugay, E.; Alp, F.N.; Kucukoduk, M.; Turkan, I. Naringenin induces tolerance to salt/osmotic stress through the regulation of nitrogen metabolism, cellular redox and ROS scavenging capacity in bean plants. Plant Physiology and Biochemistry 2020, 157, 264–275. [Google Scholar] [CrossRef] [PubMed]
- Ding, J. U., Sun, Y., Xiao, C. L., Shi, K., Zhou, Y. H., & Yu, J. Q. Physiological basis of different allelopathic reactions of cucumber and figleaf gourd plants to cinnamic acid. Journal of Experimental Botany, 2007, 58, 3765-3773. [CrossRef]
- Yadav, V., Singh, N. B., Singh, H., Singh, A., Hussain, I. Putrescine affects tomato growth and response of antioxidant defense system due to exposure to cinnamic acid. International Journal of Vegetable Science, 2019, 25, 259-277. [CrossRef]
- Almas, H. I., -un-Nisa, Z., Anwar, S., Kausar, A., Farhat, F., Munawar, M., Khalizadieh, R. Exogenous application of methionine and phenylalanine confers salinity tolerance in tomato by concerted regulation of metabolites and antioxidants. Journal of Soil Science and Plant Nutrition, 2021, 21, 3051-3064. [CrossRef]
- Świeca, M. Elicitation and treatment with precursors of phenolics synthesis improve low-molecular antioxidants and antioxidant capacity of buckwheat sprouts. Acta Scientiarum Polonorum Technologia Alimentaria, 2016, 15, 17-28. [CrossRef]
- Aryal, S.; Baniya, K.; Danekhu, K.; Kunwar, P.; Gurung, R.; Koirala, N. Total phenolic content, flavonoid content and antioxidant potential of wild vegetables from Western Nepal. Plants, 2019, 8, 96. [CrossRef]
- Kiokias, S.; Oreopoulou, A. Review of the health protective effects of phenolic acids against a range of severe pathologic conditions (including coronavirus-based infections). Molecules 2021, 26, 5405. [CrossRef]
- Nechaeva, T.; Nikolaeva, T.; Zagoskina, N. Salicylic and hydroxybenzoic acids affect the accumulation of phenolic compounds in tea-plant cultures in vitro. Biol. Bull. Russ. Acad. Sci. 2020, 47, 374–380. [CrossRef]
- Ricco, M.; Bari, M.; Catalano, A.; López, P.; Dobrecky, C.; Teves, S.; Posadaz, A.; Becher, M.; Ricco, R.; Wagner, M.; et al. Dynamics of polyphenol biosynthesis by calli cultures, suspension cultures and wild specimens of the medicinal plant Ligaria cuneifolia (Ruiz & Pav.) Tiegh. (Loranthaceae). Analysis of their biological activity. Plants 2021, 10, 1713. [CrossRef]
- Atteya, A. K., El-Serafy, R. S., El-Zabalawy, K. M., Elhakem, A., Genaidy, E. A. Exogenously supplemented proline and phenylalanine improve growth, productivity, and oil composition of salted moringa by up-regulating osmoprotectants and stimulating antioxidant machinery. Plants, 2022, 11, 1553. [CrossRef]
- Meng, X.H.; Liu, C.; Fan, R.; Zhu, L.F.; Yang, S.X.; Zhu, H.T.; Zhang, Y.J. Antioxidative flavan-3-ol dimers from the leaves of Camellia fangchengensis. J. Agric. Food Chem. 2018, 66, 247–254. [CrossRef]
- Zubova, M.; Nechaeva, T.; Kartashov, A.; Zagoskina, N. Regulation of the phenolic compounds accumulation in the tea-plant callus culture with a separate and combined effect of light and cadmium ions. Biol. Bull. Russ. Acad. Sci. 2020, 47, 593–604. [CrossRef]
- Yu, X.; Xiao, J.; Chen, S.; Yu, Y.; Ma, J.; Lin, Y.; Li, R.; Lin, J.; Fu, J.; Zhou, Q.; et al. Metabolite signatures of diverse Camellia sinensis tea populations. Nat. Commun. 2020, 11, 5586. [CrossRef]
- Bido, G. D. S., Ferrarese, M. D. L. L., Marchiosi, R., Ferrarese-Filho, O. Naringenin inhibits the growth and stimulates the lignification of soybean root. Brazilian Archives of Biology and Technology, 2010, 53, 533-542. [CrossRef]
- Shilpa, K., & Lakshmi, B. S. Influence of exogenous cinnamic acid on the production of chlorogenic acid in Cichorium intybus L cell culture. South African Journal of Botany, 2019, 125, 527-532. [CrossRef]
- Świeca, M. Hydrogen peroxide treatment and the phenylpropanoid pathway precursors feeding improve phenolics and antioxidant capacity of quinoa sprouts via an induction of L-tyrosine and L-phenylalanine ammonia-lyases activities. Journal of Chemistry, 2016, 2016. [CrossRef]
- Koca, N., Karaman, Ş. The effects of plant growth regulators and L-phenylalanine on phenolic compounds of sweet basil. Food chemistry, 2015, 166, 515-521. [CrossRef]
- Świeca, M.; Sęczyk, Ł.; Gawlik-Dziki, U. Elicitation and precursor feeding as tools for the improvement of the phenolic content and antioxidant activity of lentil sprouts. Food Chemistry 2014, 161, 288–295. [Google Scholar] [CrossRef] [PubMed]
- Hojati, M.; Modarres-Sanavy, S.A.M.; Enferadi, S.T.; Majdi, M.; Ghanati, F.; Farzadfar, S. Differential deployment of parthenolide and phenylpropanoids in feverfew plants subjected to divalent heavy metals and trans-cinnamic acid. Plant Soil 2016, 399, 41–59. [CrossRef]
- Pardossi, A., Vernieri, P., Tognoni, F. Involvement of abscisic acid in regulating water status in Phaseolus vulgaris L. during chilling. Plant Physiology, 1992, 100, 1243-1250. [CrossRef]
- Tomilova, S.V.; Kochkin, D.V.; Tyurina, T.M.; Glagoleva, E.S.; Labunskaya, E.A.; Galishev, B.A.; Nosov, A.M. Growth and biosynthetic profiles of callus and suspension cell cultures of two rare foxglove species, Digitalis grandiflora Mill. and D. ciliata Trautv. Plant Cell Tissue Organ Cult. (PCTOC) 2022, 149, 213–224. [Google Scholar] [CrossRef]
- Beauchamp, C.; Fridovich, I. Superoxide dismutase: improved assays and an assay applicable to acrylamide gels. Analytical Biochemistry, 1971, 44, 276-287. [CrossRef]
- Cavalcanti, F. R., Oliveira, J. T. A., Martins-Miranda, A. S., Viégas, R. A., Silveira, J. A. G. Superoxide dismutase, catalase and peroxidase activities do not confer protection against oxidative damage in salt-stressed cowpea leaves. New Phytologist, 2004 163, 563-571. [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein–dye binding. Anal. Biochem., 1976, 72, 248-254. [CrossRef]
- Henry, E. W., Jordan III, W. The enzymic response of pea (Pisum sativum) stem sections to applied indoleacetic acid, gibberellic acid and ethrel: catalase, peroxidase and polyphenol oxidase. Zeitschrift für Pflanzenphysiologie, 1977, 84, 321-327. [CrossRef]
- Galibina, N. A., Ershova, M. A., Ignatenko, R. V., Nikerova, K. M., Sofronova, I. N., Borodina, M. N. Cytogenetic and Biochemical Characteristics of Callus Pinus sylvestris L. Russian J. Plant Physiol., 2023, 70, 10. [CrossRef]
- Olenichenko, N. A., Zagoskina, N. V., Astakhova, N. V., Trunova, T. I., & Kuznetsov, Y. V. Primary and secondary metabolism of winter wheat under cold hardening and treatment with antioxidants. Applied Biochemistry and Microbiology, 2008, 44, 535-540. [CrossRef]
- Nikolaeva, T.N.; Lapshin, P. V.; Zagoskina, N. V. Method for determining the total content of phenolic compounds in plant extracts with Folin–Denis reagent and Folin–Ciocalteu reagent: Modification and comparison. Russian J. Bioorganic Chemistry. 2022, 48, 1519-1525. [CrossRef]
- Ossipova, S.; Ossipov, V.; Haukioja, E.; Loponen, J.; Pihlaja, K. Proanthocyanidins of mountain birch leaves: Quantification and properties. Phytochem. Anal. 2001, 12, 128–133. [CrossRef]
- Senthilkumar, M.; Amaresan, N.; Sankaranarayanan, A. Estimation of malondialdehyde (MDA) by thiobarbituric acid (TBA) assay. In Plant-Microbe Interactions; Springer Protocols Handbooks: New York, NY, USA, 2021; pp. 103–105. [CrossRef]
- Hodges, D.M.; DeLong, J.M.; Forney, C.F.; Prange, R.K. Improving the thiobarbituric acid-reactive-substances assay for estimating lipid peroxidation in plant tissues containing anthocyanin and other interfering compounds. Planta, 1999, 207, 604–611. [CrossRef]
Figure 1.
The main stages of phenolic compounds biosynthesis. PAL—L-phenylalanine ammonia-lyase; C4H—cinnamic acid 4-hydroxylase; 4CL—4-coumarate-CoA-ligase; CHS—chalcone synthase; CHI—chalcone isomerase.
Figure 1.
The main stages of phenolic compounds biosynthesis. PAL—L-phenylalanine ammonia-lyase; C4H—cinnamic acid 4-hydroxylase; 4CL—4-coumarate-CoA-ligase; CHS—chalcone synthase; CHI—chalcone isomerase.
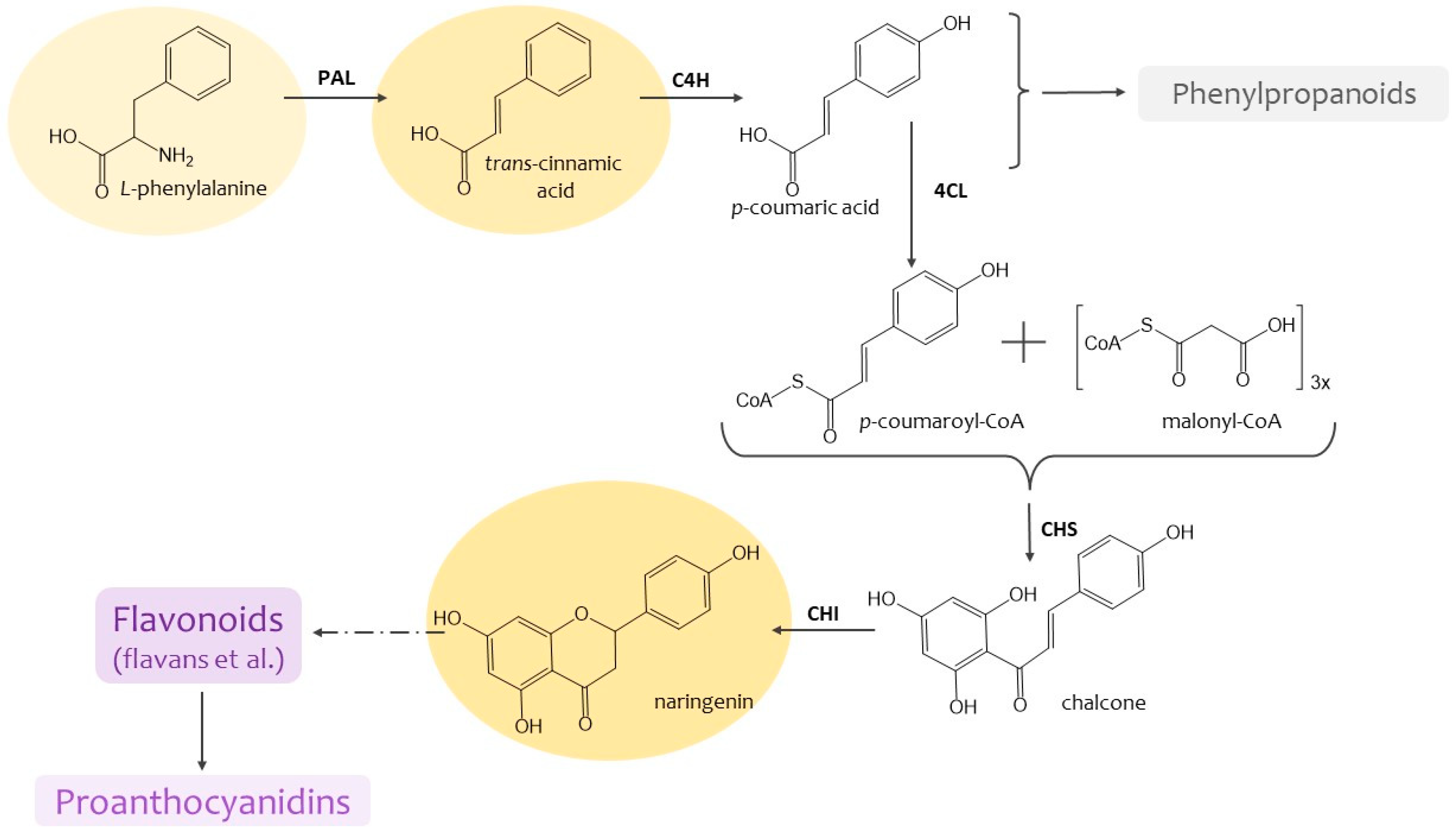
Figure 2.
The tea cells cultures grown for 7 days on the liquid Heller culture medium of the basic composition (Control) or enriched with L-phenylalanine (PhA; 3 mM), trans-cinnamic acid (CA; 1 mM) or naringenin (NG; 0.5 mM).
Figure 2.
The tea cells cultures grown for 7 days on the liquid Heller culture medium of the basic composition (Control) or enriched with L-phenylalanine (PhA; 3 mM), trans-cinnamic acid (CA; 1 mM) or naringenin (NG; 0.5 mM).

Figure 3.
The activity of level of superoxide dismutase – SOD (a), and peroxidase – POX (b), in tea cells cultures grown for 7 days on the liquid Heller culture medium of the basic composition (Control) or enriched with L-phenylalanine (PhA; 3 mM), trans-cinnamic acid (CA; 1 mM) or naringenin (NG; 0.5 mM). Results are expressed as means ± SDs, n = 3. The significant differences at p < 0.05 are denoted by different Latin letters.
Figure 3.
The activity of level of superoxide dismutase – SOD (a), and peroxidase – POX (b), in tea cells cultures grown for 7 days on the liquid Heller culture medium of the basic composition (Control) or enriched with L-phenylalanine (PhA; 3 mM), trans-cinnamic acid (CA; 1 mM) or naringenin (NG; 0.5 mM). Results are expressed as means ± SDs, n = 3. The significant differences at p < 0.05 are denoted by different Latin letters.
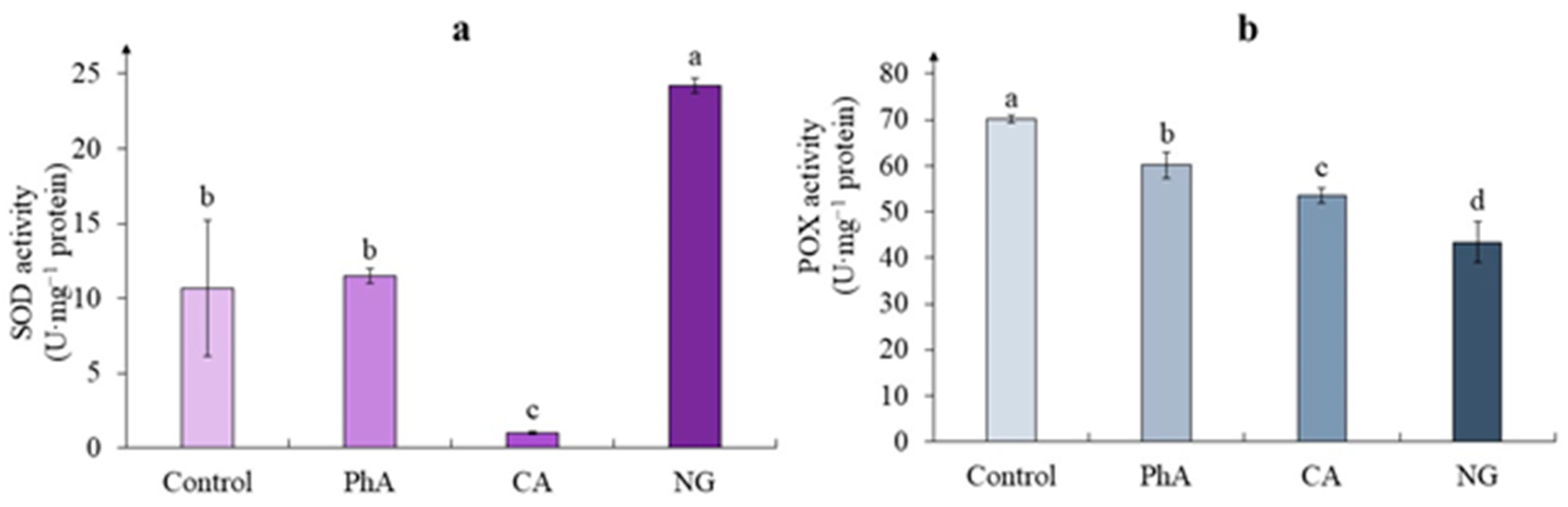
Figure 4.
The total phenolic content – TPC (a), the flavanols content – FLC (b), the proanthocyanidins content – PAC (c) in tea cells cultures grown for 7 days on the liquid Heller culture medium of the basic composition (Control) or enriched with L-phenylalanine (PhA; 3 mM), trans-cinnamic acid (CA; 1 mM) or naringenin (NG; 0.5 mM). Results are expressed as means ± SDs, n = 3. The significant differences at p < 0.05 are denoted by different Latin letters.
Figure 4.
The total phenolic content – TPC (a), the flavanols content – FLC (b), the proanthocyanidins content – PAC (c) in tea cells cultures grown for 7 days on the liquid Heller culture medium of the basic composition (Control) or enriched with L-phenylalanine (PhA; 3 mM), trans-cinnamic acid (CA; 1 mM) or naringenin (NG; 0.5 mM). Results are expressed as means ± SDs, n = 3. The significant differences at p < 0.05 are denoted by different Latin letters.
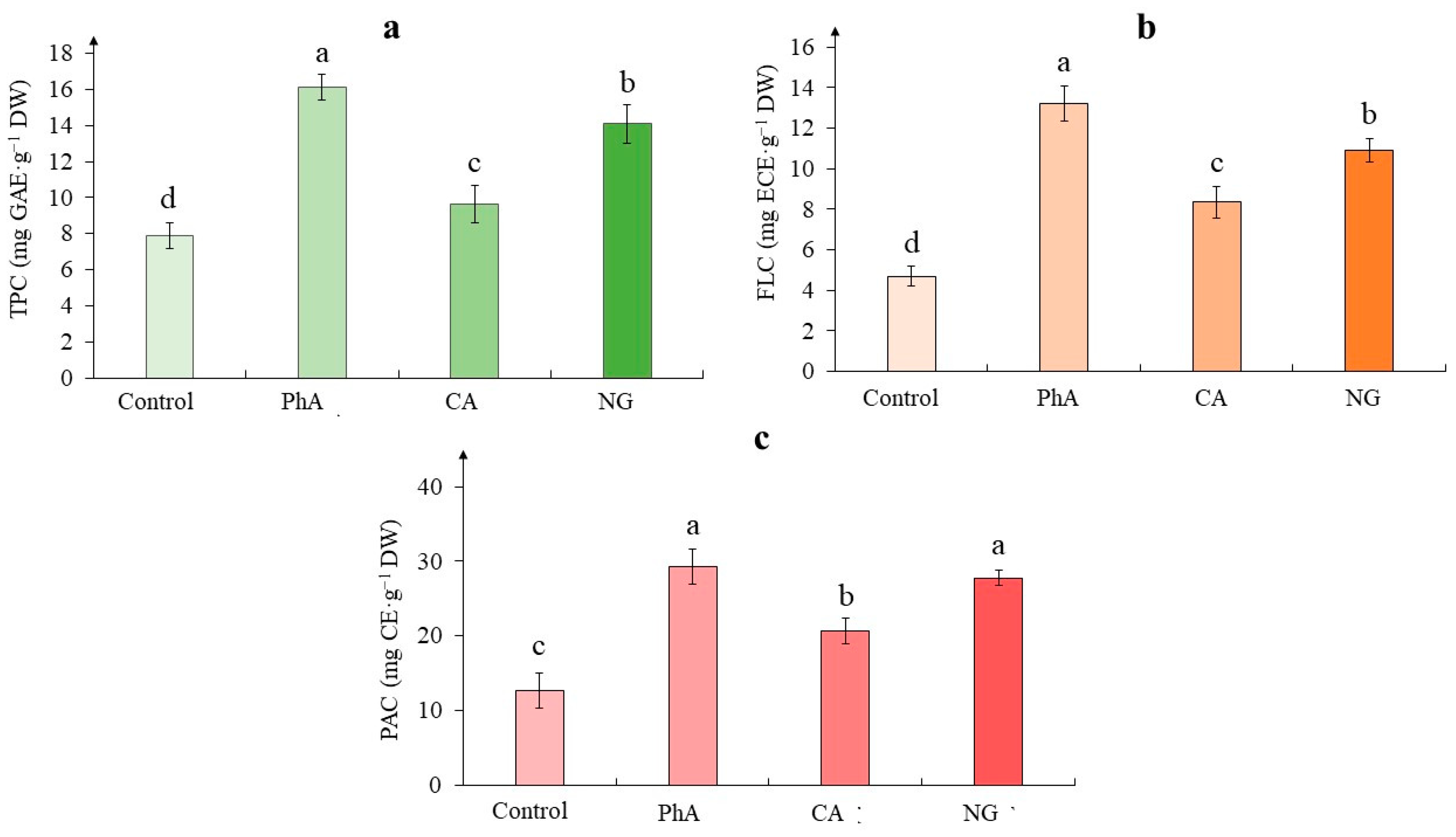
Figure 5.
The level of phenylalanine ammonia-lyase (PAL) activity in tea callus cultures grown for 7 days on the liquid Heller culture medium of the basic composition (Control) or enriched with L-phenylalanine (PhA; 3 mM), trans-cinnamic acid (CA; 1 mM) or naringenin (NG; 0.5 mM). Results are expressed as means ± SDs, n = 3. The significant differences at p < 0.05 are denoted by different Latin letters.
Figure 5.
The level of phenylalanine ammonia-lyase (PAL) activity in tea callus cultures grown for 7 days on the liquid Heller culture medium of the basic composition (Control) or enriched with L-phenylalanine (PhA; 3 mM), trans-cinnamic acid (CA; 1 mM) or naringenin (NG; 0.5 mM). Results are expressed as means ± SDs, n = 3. The significant differences at p < 0.05 are denoted by different Latin letters.
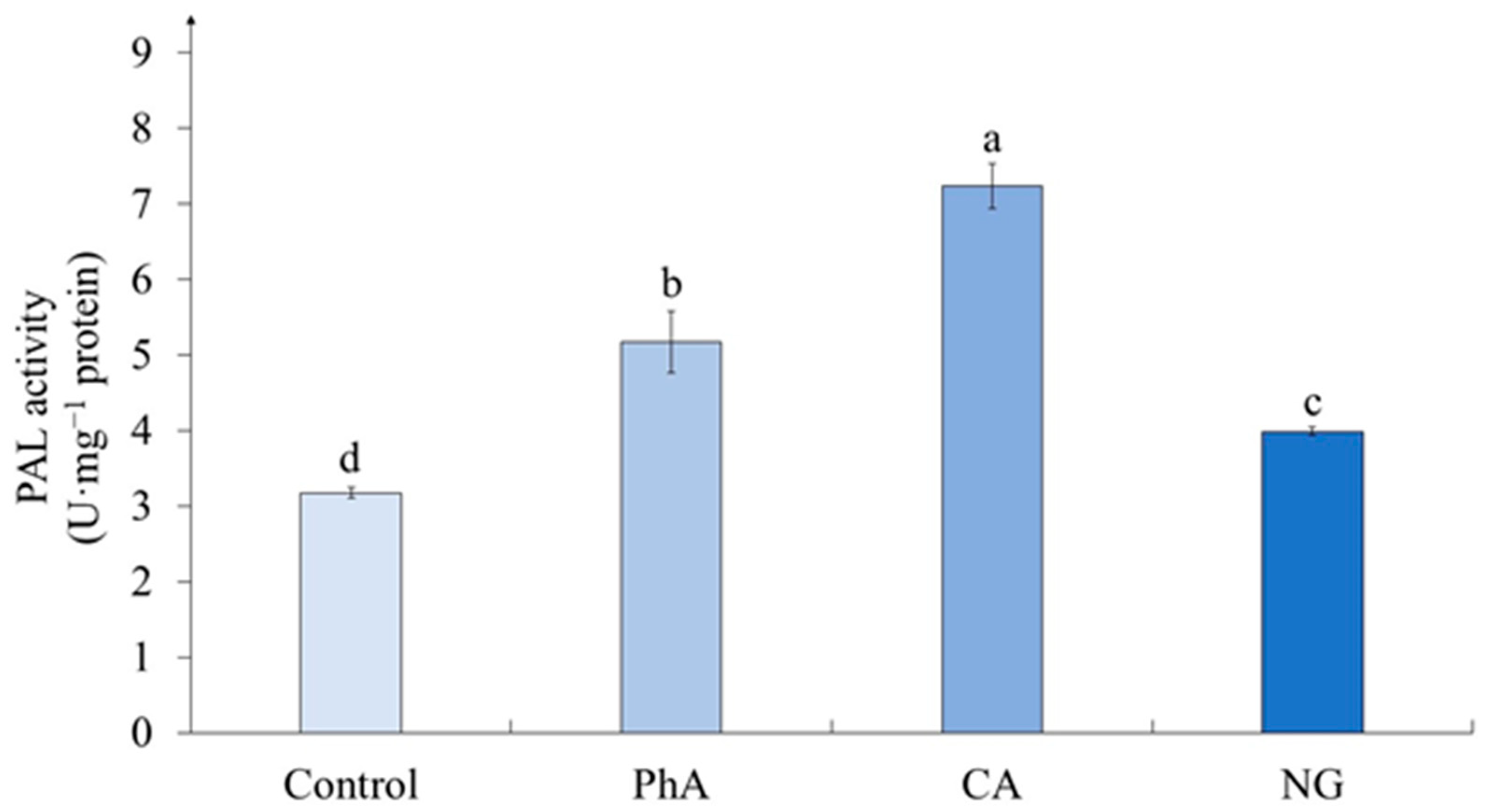
Figure 6.
The level of lipid peroxidation (LPO) in tea cells cultures grown for 7 days on the liquid Heller culture medium of the basic composition (Control) or enriched with L-phenylalanine (PhA; 3 mM), trans-cinnamic acid (CA; 1 mM) or naringenin (NG; 0.5 mM). Results are expressed as means ± SDs, n = 3. The significant differences at p < 0.05 are denoted by different Latin letters.
Figure 6.
The level of lipid peroxidation (LPO) in tea cells cultures grown for 7 days on the liquid Heller culture medium of the basic composition (Control) or enriched with L-phenylalanine (PhA; 3 mM), trans-cinnamic acid (CA; 1 mM) or naringenin (NG; 0.5 mM). Results are expressed as means ± SDs, n = 3. The significant differences at p < 0.05 are denoted by different Latin letters.
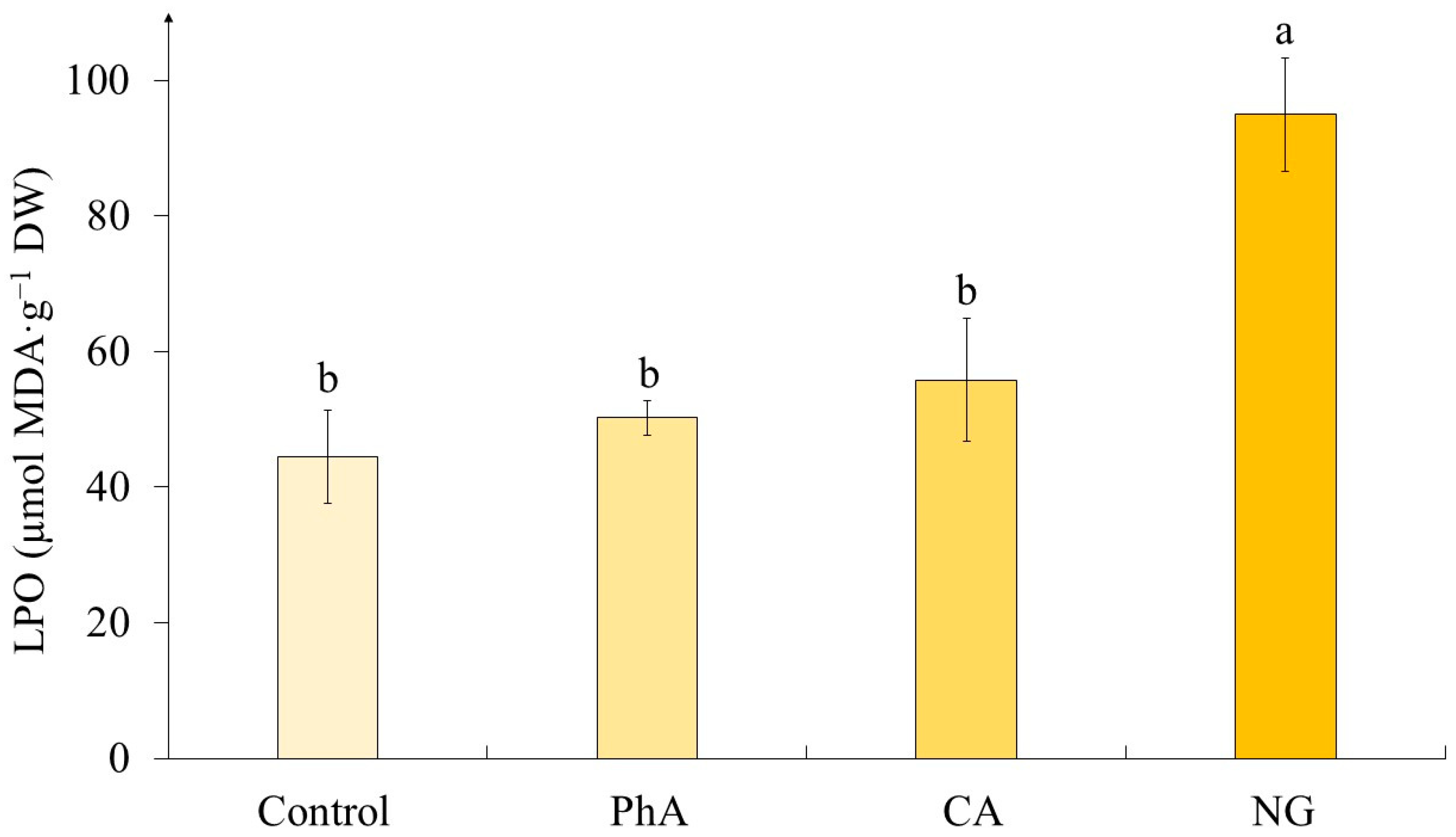
Figure 7.
Experimental design.
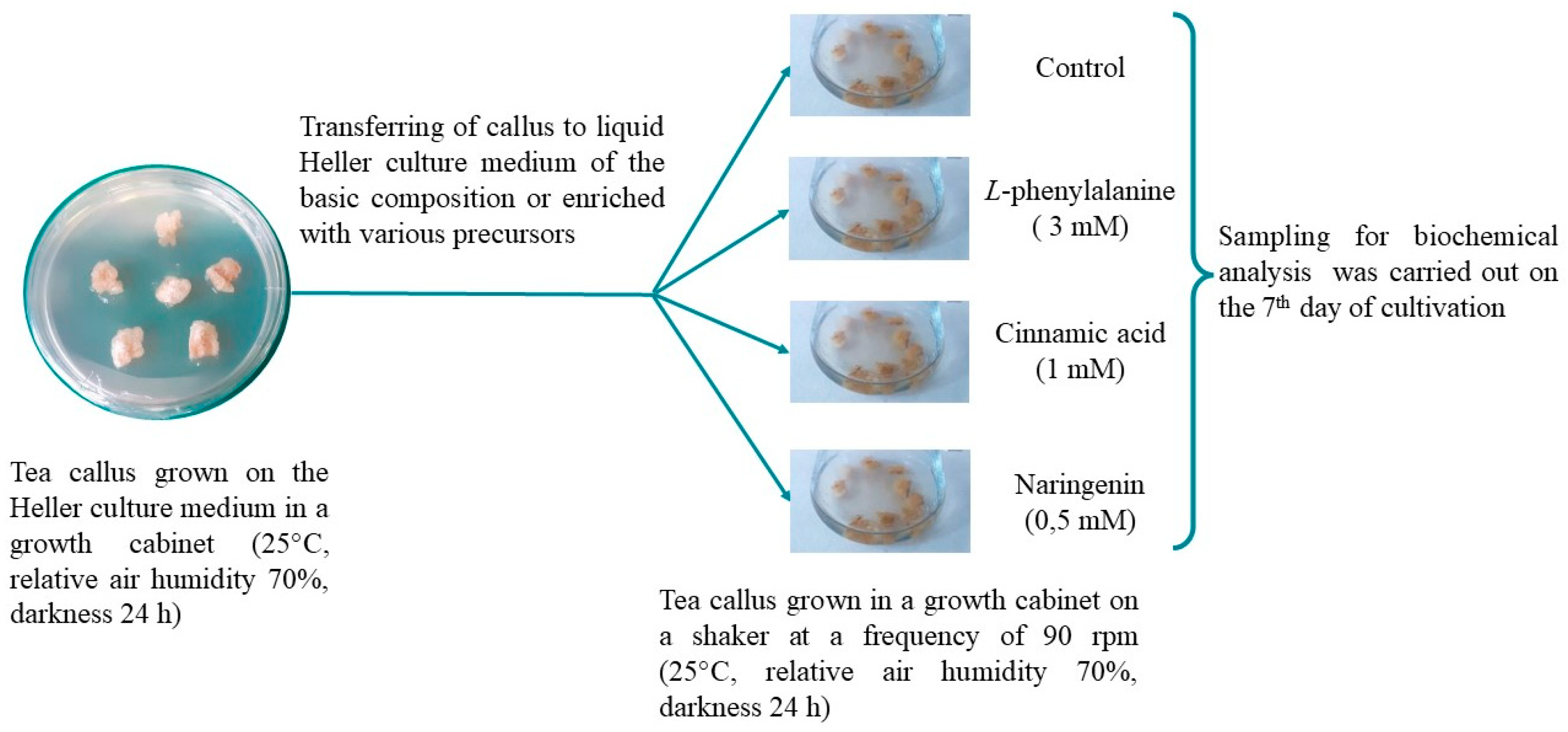

Table 1.
Water content in tea cells cultures grown for 7 days on Heller’s liquid culture medium of the basic composition (Control) or enriched with L-phenylalanine (PhA, 3 mM), trans-cinnamic acid (CA, 1 mM) or naringenin (NG, 0.1 mM).
Table 1.
Water content in tea cells cultures grown for 7 days on Heller’s liquid culture medium of the basic composition (Control) or enriched with L-phenylalanine (PhA, 3 mM), trans-cinnamic acid (CA, 1 mM) or naringenin (NG, 0.1 mM).
| Variants | Water content, % |
|---|---|
| Control | 92.13 ± 0.02 b |
| PhA | 92.05 ± 0.25 b |
| CA | 92.42 ± 0.94 b |
| NG | 94.01 ± 0.23 a |
Results are expressed as means ± standard deviations, n = 3. The significant differences at p<0.05 are indicated by different Latin letters.
Table 2.
One-way ANOVA showing the effect of the precursor’s treatments on water content in Camellia sinensis callus cultures.
Table 2.
One-way ANOVA showing the effect of the precursor’s treatments on water content in Camellia sinensis callus cultures.
| Sum of Squares | df | Mean Square | F | P |
| 7.822 | 3 | 2.607 | 3.492 | 0.129 |
Table 3.
One-way ANOVA showing the effect of the precursor’s treatments on the activity of superoxide dismutase (SOD) and guaiacol-dependent peroxidases (POX) in Camellia sinensis cells cultures.
Table 3.
One-way ANOVA showing the effect of the precursor’s treatments on the activity of superoxide dismutase (SOD) and guaiacol-dependent peroxidases (POX) in Camellia sinensis cells cultures.
| Determination | Sum of Squares | df | Mean Square | F | P |
|---|---|---|---|---|---|
| SOD activity | 42575.27 | 3 | 14191.76 | 17.212 | 0.005 |
| POX activity | 65.28 | 3 | 21.76 | 16.485 | <0,001 |
Table 4.
One-way ANOVA showing the effect of the precursor’s treatments on total phenolic compounds, flavanols, proanthocyanidins content in Camellia sinensis callus cultures.
Table 4.
One-way ANOVA showing the effect of the precursor’s treatments on total phenolic compounds, flavanols, proanthocyanidins content in Camellia sinensis callus cultures.
| Determination | Sum of Squares | df | Mean Square | F | P |
|---|---|---|---|---|---|
| Total phenolic content | 245.598 | 3 | 81.866 | 17.096 | <0,001 |
| Flavanols content | 132.252 | 3 | 44.084 | 8.811 | 0.001 |
| Proanthocyanidins content | 791.121 | 3 | 263.707 | 15.22 | <0,001 |
Table 5.
One-way ANOVA showing the effect of the precursor’s treatments on the activity of L-phenylalanine-ammonia lyase in Camellia sinensis callus cultures.
Table 5.
One-way ANOVA showing the effect of the precursor’s treatments on the activity of L-phenylalanine-ammonia lyase in Camellia sinensis callus cultures.
| Sum of Squares | df | Mean Square | F | P |
| 2218.885 | 3 | 739.628 | 47.576 | <0,001 |
Table 6.
One-way ANOVA showing the effect of the precursor’s treatments on the lipid peroxidation level in Camellia sinensis callus cultures.
Table 6.
One-way ANOVA showing the effect of the precursor’s treatments on the lipid peroxidation level in Camellia sinensis callus cultures.
| Sum of Squares | df | Mean Square | F | P |
| 4702.636 | 3 | 1567.545 | 10.176 | 0.004 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



