Submitted:
11 December 2023
Posted:
12 December 2023
You are already at the latest version
Abstract
PPR is a highly contagious viral disease and one of the deadliest affecting wild goats, sheep and small ruminants, however goats are generally more sensitive. The causative agent is the Peste des Petits Ruminants virus (PPRv) which is a single-stranded RNA virus of negative polarity belonging to the Paramyxoviridae family. In February 2020, an active outbreak of PPR was re-ported in a herd of a transhumant farmer in the village of Ngainth Pathé (department of Kounguel, Kaffrine region,Senegal). Of the ten (10) swabs collected from goats, eight (8) came back positive by real-time PCR. The most positive sample was amplified, purified and amplicon sent for sequencing. Phylogenetic analysis showed that the sequence of the PPR virus obtained belonged to lineage IV. These results confirm those found in the latter countries and reinforce the hypothesis of the importance of animal mobility between these neighboring countries. In perspective, following the discovery of this lineage IV in Senegal, a study on its dispersion is underway throughout the national territory. The results that will emerge from this study, asso-ciated with detailed data on animal movements and epidemiological data, will provide appro-priate and effective information to improve PPR surveillance and control strategies with a view to its eradication.
Keywords:
Peste des petits ruminants
; lineage IV
; Molecular characterization
; Senegal
1. Introduction
In Senegal, production of small ruminants is an activity of choice for vulnerable populations such as women and young people because of their short reproductive cycle and low investment costs compared to other livestock such as cattle [1], thus offering huge opportunities for reducing poverty. However, high burden of disease remains a major constraint to small ruminants production. Among these diseases, Peste des Petits Ruminants (PPR) poses the greatest threat to small ruminant livestock. This disease threatens global food security and the rural economy, but its control is complicated, in particular due to significant animal movements with transhumance and commercial movements in the infected regions [2].
PPR is a highly contagious viral disease and one of the deadliest affecting wild goats, sheep and small ruminants; however goats are generally more sensitive. The causative agent is the Peste des Petits Ruminants virus (PPRv) which is a single-stranded RNA virus of negative polarity belonging to the Paramyxoviridae family and the genus of morbilliviruses closely related to the rinderpest virus eradicated successfully worldwide [3]. PPR-related morbidity reaches 100% in extreme situations, with a mortality rate that can reach 90% [4]. Due to the serious impact of the disease on the economy and food security, but also its alarming rate of spread, the World Organization for Animal Health (OMSA) and the United Nations Food and Agriculture (FAO) have chosen PPR as a target for a global eradication program [5]. Based on the molecular characterization of the partial sequence of the N gene, the PPR virus is classified into four genetically different lineages (I, II, III and IV) [6].
Historically, the geographical distribution of these lineages was distinct with the circulation of lineages I, II and III in Africa and that of lineage IV in Asian countries and the Middle East [7]. However, several studies have shown the detection of lineage IV in several African countries over the past decades [8,9,10]. In West Africa, lineage I was last officially detected in 1994 in Senegal [11]. However, since that date, no information is available on the molecular characterization of the PPRv. Until 2010 when the results of the molecular characterization of circulating strains of the PPR virus obtained with the study by Salami H et al between 2010 and 2014 showed that only the lineage II could be detected in Senegal during this period [12]. The study by Tounkara et al in 2019 also confirmed that only lineage II was detected in samples taken in 2013 from Dakar markets [13]. Long time well-known in Asia, lineage IV was subsequently discovered in several African countries, particularly countries bordering Senegal such as Mali [14]. However, until then lineage IV has never been described in Senegal.
In this study, we describe the genetic diversity of the PPRv lineage IV for the first time in Senegal.
2. Materials and Methods
2.1. Outbreak investigation and sample collection
In February 2020, an active outbreak of PPR was reported in a mixed herd of 160 sheep and goats of a transhumant farmer in the village of Ngainth Pathé (department of Kounguel, Kaffrine region). The transhuman farmer had left the village of Tatki Hillo in the department of Podor (northern Senegal) with his herd at the beginning of November 2020. Along the way, he stayed in the Sylvopastoral zone in Téssékré, department of Linguère, region of Louga before reach the locality of Ngainth Pathé in the central zone in the Kaffrine region (see figure below) where the samples were collected. Symptoms observed in infected animals are tearing, coughing, dyspnoea, throwing up, diarrhea and ulcerations in the gingival mucosa. At the start of the infection, 25 of them presented very intense digestive and respiratory syndromes with high mortality in lambs and kids over 80% of the population. All sick animals were sampled by taking an oculo-nasal or rectal swab for animals with diarrhea, for some both types of samples. The swabs were then transported to the laboratory in 2 ml tubes containing 500 µl of viral transport medium and then stored in liquid nitrogen.
Figure 10.
swabs collected from goats, the viral RNA was extracted using the QIAGEN RNeasy mini kit according to the supplier's instructions. The extracted RNAs were then analyzed by Real-time RT-PCR using the qScritpt XLT One step RT-qPCR Tough mix amplification kit from Quantabio with the sets of primers and probes developed by Batten et al [15] in order to estimate the viral load of the samples. All strong positive samples were subjected to conventional RT-PCR using forward primer NP3 (5'-GTCTCGGAAATCGCCTCACAGACT-3') and reverse primer NP4 (3'-CCTCCTCCTGGTCCTCCAGAATCT-5') [16] located at the 3' end of the N gene using the One Step RT-PCR mix kit from Qiagen, making it possible to amplify a segment of 351 base pairs (bp) of the PPRV gene. The PCR was carried out according to the following program: 1 cycle RT (Retro transcription): 30 min at 50°C, 1 cycle of primer hybridization: 15 min at 95°C and 40 cycles of elongation (15sec 95°C 45 sec at 60°C; 45 sec at 72°C). The size of the amplified fragments was visualized after electrophoretic migration of the PCR products in a 1.5% agarose gel with 1X TAE buffer.
Figure 10.
swabs collected from goats, the viral RNA was extracted using the QIAGEN RNeasy mini kit according to the supplier's instructions. The extracted RNAs were then analyzed by Real-time RT-PCR using the qScritpt XLT One step RT-qPCR Tough mix amplification kit from Quantabio with the sets of primers and probes developed by Batten et al [15] in order to estimate the viral load of the samples. All strong positive samples were subjected to conventional RT-PCR using forward primer NP3 (5'-GTCTCGGAAATCGCCTCACAGACT-3') and reverse primer NP4 (3'-CCTCCTCCTGGTCCTCCAGAATCT-5') [16] located at the 3' end of the N gene using the One Step RT-PCR mix kit from Qiagen, making it possible to amplify a segment of 351 base pairs (bp) of the PPRV gene. The PCR was carried out according to the following program: 1 cycle RT (Retro transcription): 30 min at 50°C, 1 cycle of primer hybridization: 15 min at 95°C and 40 cycles of elongation (15sec 95°C 45 sec at 60°C; 45 sec at 72°C). The size of the amplified fragments was visualized after electrophoretic migration of the PCR products in a 1.5% agarose gel with 1X TAE buffer.

2.2. Sequencing and editing
The amplification products were then purified using the PROMEGA SV Gel and PCR clean-up system-50rxn wizard kit. Amplicons thus purified was sent to LGC genomics (Germany) for sequencing using the Sanger method. The analysis of the generated sequences carried out with the Molecular Evolutionary Genetics Analysis 7 (MEGA 7) software [17]. The Bioedit software[18] made it possible to clean the sequences, and to carry out the multiple alignments of the N gene with other PPRV sequences representative of the four genetic lines obtained in Genbank. The two-parameter Kimura model proposed by the MEGA 7 program was used. From this model, phylogenetic trees were constructed in MEGA 7 using the likelihood method (ML) with a repetition of 1000 (bootstraps).
2.3. Nucleotide sequence accession number:
The complete genome sequence of the GTP_SEN-KAFF2021 PPRv strain is available at Genbank under the accession number OR819867.
3. Results
Almost all affected animals expressed very expressive digestive and respiratory syndromes associated with fetid diarrhea, mucopurulent discharge and intense lacrimation then in the final phase a high lethality up to 80% of the affected herd. Goat kids had a mortality rate up to 100% while morbidity also approached 100% for adults who survived.
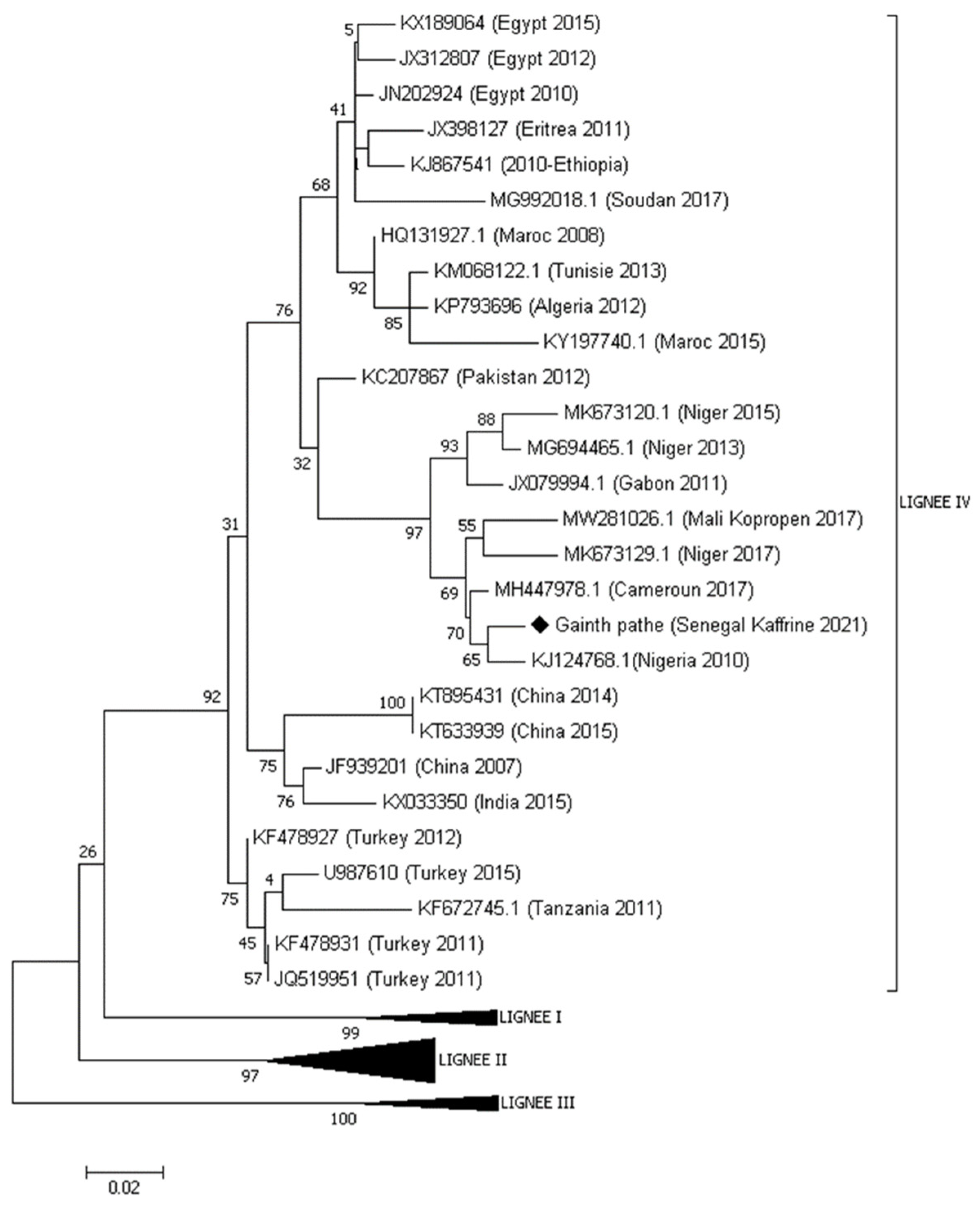
Ten swabs (10) taken were analyzed and eight (8) came back positive by real-time PCR. The most positive sample with a Cycle Threshold (CT) value of 18.38 was amplified with the NP3/NP4 primer pair and then the purified amplicon sent for sequencing. Phylogenetic analysis showed that the sequence of the PPR virus obtained belonged to lineage IV (see figure Phylogenetic tree below).
Figure 2021.
Numbers at nodes are Bootstrap values obtained from 1000 repetitions.
4. Discussion
This is the first time that this lineage IV is reported in Senegal. This new situation in the epidemiology of PPR in Senegal was predictable considering the epidemiological context of PPR prevailing in neighboring countries and animal mobility. The phylogenetic tree shows that the Ngainth Pathé sequence of Kaffrine fit into a cluster with 69% node support comprising sequences identified in 2010 in Nigeria [10] and in 2017 in Mali, Niger and Cameroon [14,19,20]. These results confirm the conclusions drawn from the study by Tounkara et al. in 2021 on the risk of introduction of lineage 4 in Senegal due to animal mobility between Senegal and Mali[14]. Indeed, Mali is the largest exporter of animals during Muslim holidays to cross-border countries. In the same dynamic, the similarity of sequences from Nigeria and Niger had already been described by Souley et al [19] suggesting a cross-border movement of PPR between neighboring countries and beyond to Cameroon. This explains this grouping of these sequences belonging to these countries in the same cluster. Phylogenetic analysis identified the presence of multiple PPRv genetic clades that can be considered as part of different transmission networks evolving in parallel in West Africa. These results highlight the importance of animal mobility in the transmission of PPR in the region. With these results, it would be ideal to combine genetic data with mobility networks to identify key sites for virus entry and spread in given areas. Such information could strengthen our ability to develop locally adapted control and surveillance strategies, using, among other risk factors, information on animal mobility.
5. Conclusions
We report the first detection of lineage IV of the PPRv in an outbreak in Ngainth Pathé village (Kaffrine region) in Senegal in February 2021. Phylogenetic analysis showed that the sequence found during this outbreak is genetically linked with sequences found in Mali, Niger, Nigeria and Cameroon. These results confirm those found in the latter countries and reinforce the hypothesis of the importance of animal mobility between these neighboring countries.
In perspective, following the discovery of this lineage IV in Senegal, a study on its dispersion is underway throughout the national territory. The results that will emerge from this study, associated with detailed data on animal movements and epidemiological data, will provide appropriate and effective information to improve PPR surveillance and control strategies with a view to its eradication.
Author Contributions
“Conceptualization, A.B.; methodology, A.B.; M.N.;G.L.D. software, A.B.; validation, M.D. and M.M.L.; formal analysis, A.B.; writing—original draft preparation, A.B. and G.L.D; writing—review and editing, A.B.;M.D.; supervision, M.M.L.; funding acquisition, M.D., M.M.L, All authors have read and agreed to the published version of the manuscript.
Funding
This work was funded as part of the Epidemiology and Control of Peste des Petits Ruminants (EcoPPR) with funding from the EC-IFAD (grand code 2000002577) and CGIAR Research Program on Livestock and continued under the CGIAR Initiative Sustainable Animal Productivity for Livelihoods, Nutrition and Gender (SAPLING). CGIAR research is supported by contributions from the CGIAR Trust Fund. CGIAR is a global research partnership for a food-secure future dedicated to transforming food, land, and water systems in a climate crisis. The funders had no role in study design, data collection and analysis, the decision to publish, or the preparation of the manuscript.
Institutional Review Board Statement
The study was conducted in accordance with the Declaration of Helsinki and approved by the ILRI Institutional Research Ethics Committee (IREC), reference: ILRI-IREC2020-16.
Acknowledgments
The authors are grateful for the administrative and technical support they received. The authors would like to thank Sakhewar Diouf head of veterinary station of Ngainth Pathé.
Conflicts of Interest
The authors declare that they have no known conflicts of interest or personal relationships that could have appeared to influence the work reported in this paper.
References
- Bureau d’Analyse Macro-Economique(BAME), ISRA : Nouvelle Initiative Sectorielle pour le Développement de l’Elevage, situation et perspective du sous-secteur de l’élevage,2004.
- Fakri, F., Embarki, T., Parida, S., et al. Re-emergence of Peste des Petits Ruminants virus in 2015 in Morocco: Molecular characterization and experimental infection in Alpine goats. Veterinary Microbiology, 2016, vol. 197, p. 137-141.
- Pastoret, P. P., Yamanouchi, K., Mueller-Doblies, U., Rweyemamu, M. M., Horzinek, M., & Barrett, T. Rinderpest—an old and worldwide story: history to c. 1902. In Rinderpest and peste des petits ruminants, 2006, pp. 86-VI. Academic Press.
- Kumar, N.; Maherchandani, S.; Kashyap, S.K.; Singh, S.V.; Sharma, S.; Chaubey, K.K.; Ly, H. Peste Des Petits Ruminants Virus Infection of Small Ruminants: A Comprehensive Review. Viruses 2014, 6, 2287–2327. [Google Scholar] [CrossRef] [PubMed]
- OIE&FAO. Global control and eradication of PPR. http://www.oie.int/eng/PPR2015/doc/PPR-Advocacy-EN.pdf. Accessed on 30 Dec 2019.
- Kwiatek, O., Minet, C., Grillet, C., Hurard, C., Carlson, E., Karimov, B.,Diallo, A. Peste des Petits Ruminants (PPR) outbreak in Tajikistan. Journal of Comparative Pathology, 2007, 136, 111–119. [CrossRef]
- Banyard, A. C., Parida, S., Batten, C., Oura, C., Kwiatek, O., & Libeau, G. Global distribution of peste des petits ruminants virus and prospects for improved diagnosis and control. Journal of General Virology, 2010, ,91, 2885–2897. [CrossRef]
- Kwiatek, O., Ali, Y. H., Saeed, I. K., Khalafalla, A. I., Mohamed, O. I.,Obeida, A. A., Libeau, G. Asian lineage of peste des petits ruminants virus, Africa. Emerging Infectious Diseases, 2011, 17, 1223–123. [CrossRef]
- Dundon, W., Diallo, A., & Cattoli, G. Peste des petits ruminants in Africa: A review of currently available molecular epidemiological data, 2020. Archives of Virology, 2020, 165, 2147–2163.
- Woma, T. Y., Adombi, C. M., Yu, D., Qasim, A. M., Sabi, A. A., Maurice, N. A., Olaiya, O. D., Loitsch, A., Bailey, D., Shamaki, D., Dundon, W.G., & Quan, M. Co-circulation of Peste des Petits Ruminants Virus Asian lineage IV with Lineage II in Nigeria. Transboundary and Emerging Diseases,2016, 63, 235–242.
- Tounkara, K., Bataille, A., Adombi, C. M., Maikano, I., Djibo, G., Settypalli, T. B. K., Loitsch, A., Diallo, A., & Libeau, G. First genetic characterization of Peste des Petits Ruminants (PPR) from Niger: On the advancing front of the Asian virus lineage. Transboundary and Emerging Diseases,2018, 65, 1145–1151.
- Salami, H., Croville, G., Kwiatek, O., Mariette, J., Klopp, C., Valiere, S., Libeau, G. Complete genome sequence of a field strain of peste des petits ruminants virus isolated during 2010-2014 epidemics in Senegal. Genome announcements, 2014, vol. 2, no 5, p. 10.1128/genomea. 00772-14. [CrossRef]
- Tounkara, K., Kwiatek, O., Niang, M., Abou Kounta Sidibe, C., Sery, A., Dakouo, M., ... & Bataille, A. Corrigendum: Genetic Evidence for Transboundary Circulation of Peste Des Petits Ruminants Across West Africa. Frontiers in Veterinary Science, 2019, vol.6, p. 380.
- Tounkara, K, kwiatek, O, Sidibe, C.A. K, et al. Persistence of the historical lineage I of West Africa against the ongoing spread of the Asian lineage of peste des petits ruminants virus. Transboundary and Emerging Diseases, 2021, vol. 68, no 6, p. 3107-3113. [CrossRef]
- Batten, Carrie A., Banyard, Ashley C., King, Donald P., et al. A real time RT-PCR assay for the specific detection of Peste des petits ruminants virus. Journal of virological methods, 2011, vol. 171, no 2, p. 401-404.
- Couacy-hymann, E., Roger, F., Hurard, C., Guillou, J., Libeau, G., AND Diallo, A. Rapid and sensitive detection of peste des petits ruminants virus by a polymerase chain reaction assay. Journal of Virological Methods,2002, 100, 1, 17–25.
- Tamura K, Stecher G, Peterson D, Filipski A. and Kumar S: 1993-2023. MEGA7: Molecular Evolutionary Genetics Analysis Version 7.0.26 Molecular Biology and Evolution. 30: 2725-2729.
- HALL, T.A. BioEdit: a user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucleic acids symposium series. 1999. p. 95-98.
- Souley, M. M., Ibrahim, A. S., Sidikou, D., Dundon, W. D., Cattoli, G., Abdou, A., Soumana, F., & Yaou, B. Molecular epidemiology of peste des petits ruminants in Niger: An update. Transboundary and Emerging Diseases,2019, 67, 1388–1392.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



