
Submitted:
11 December 2023
Posted:
12 December 2023
You are already at the latest version
Abstract
Viruses pose a great threat to people’s lives. Enterovirus A71 (EV-A71) infects children and infants all over the world with no FDA-approved treatment to date. Understanding the basic mechanisms of viral processes aids in selecting more efficient drug targets and designing more effective antivirals to thwart this virus. The 5´-untranslated region (5’-UTR) of the viral RNA genome is composed of a cloverleaf structure and an internal ribosome entry site (IRES). Cellular proteins that bind to the cloverleaf structure regulate viral replication, while those that bind to the IRES also known as IRES trans-acting factors (ITAFs) regulate viral translation. In this review, we survey the cellular proteins currently known to bind the 5’-UTR and influence viral translation and replication with emphasis on comparing proteins’ functions and localizations pre- and post-(EV-A71) infection. Comprehensive understanding of how the host cell’s machinery is hijacked and reprogrammed by the virus to facilitate its replication is crucial for developing effective antivirals.
Keywords:
Enterovirus A71 (EV-A71)
; ITAF
; IRES
; 5’-UTR
1. Introduction
Viral infections can cause life threatening illness. Enterovirus A71 (EV-A71), the causative agent of Hand, foot, and mouth disease (HFMD), has resulted in high morbidity and in some cases mortality rates in infants and children under five years old.1 Mild symptoms of HFMD include rashes on the hands, feet, and mouth, fever and painful blister-like sores. In addition, infection with EV-A71 has been known to cause more severe complications such as, brainstem encephalitis, cardiopulmonary failure, acute flaccid paralysis, and death in severe cases.1,2
According to the World Health Organization (WHO), EV-A71 was first isolated in 1969 in California, USA; however, some studies showed that the virus could have emerged in 1963 when a worldwide epidemic occurred in the late twentieth century.3 Outbreaks of EV-A71 have occurred primarily in East Asian countries as well as in Europe and America.4 In China, there were approximately 7.2 million infections and two thousand deaths between 2008 and 2012.5 Waves of infections occurred in the Netherlands, France, and Germany from 2007-2013.5 Most recent incidences of EV-A71 infection include several cases in Spain in 20166 and around 13,000 cases in Gansu Province, China in 2018.7 Also in 2018, the Children’s Hospital of Colorado reported a 2.75 percent increase in confirmed cases as compared to the average from the past five years (2013-2017).8 In the same year, Vietnam reported 53,000 clinical cases of HFMD and 6 deaths.9 Approximately 2,500 Taiwanese individuals were infected with EV-A71 in 2020, who needed emergency or outpatient visits to the hospital.2 Notably, the National Institute of Allergy and Infectious Diseases (NIAD) listed EV-A71 as an emerging infectious disease.10 Due to the severity of EV-A71 infection and its life-threatening complications to children, researchers and scientists have been working on ways to combat this virus. To date, there is no antiviral treatment for EV-A71 infection; however, progress has been made in the development of vaccines. There are three inactivated, whole EV-A71 vaccines available in China. Other types of vaccines are still in the development process.11,12
EV-A71 belongs to the Picornaviridae family, genus Enterovirus, and species Enterovirus A.13 It is a non-enveloped RNA virus with a positive sense, single stranded RNA.14 The viral genome of EV-A71 is around 7,400 nucleotides long and composed of a 5´- untranslated region (5´-UTR), a single open reading frame (ORF) and a 3´-UTR.14 The 5´-UTR of the genome folds into a stable secondary structure that has six stem loops (SLI-VI), as determined by structure prediction software. The SLI is also known as the cloverleaf structure while SLII-VI forms a type I internal ribosome entry site (IRES) (Figure 1b, Figure 2).4,15 The 5´-UTR region is the control hub for EV-A71 genome replication and translation.
Upon infection, EV-A71 causes rapid inhibition of cap-dependent translation machinery of the host cell and utilizes host’s translation machinery for cap-independent translation of viral proteins mediated by its IRES element within the 5´-UTR (Figure 1a).16 Once the viral particle binds to the host cell receptor and the viral genome is released into the cytoplasm, translation results in a single polyprotein encoded by the ORF. This resultant polyprotein is cleaved by cellular and/or viral proteases, mainly 3C protease (3Cpro) to generate a total of 11 viral proteins VP4, VP2, VP3, VP1, 2A, 2B, 2C, 3A, 3B, 3C and 3D. The 3Cpro along with 2Apro also cleaves host cell’s translation initiation factors (eIFs) such as eIF4G, eIF4A, and eIF5B and PABP resulting in the shutdown of host cell’s cap-dependent translation. Inducing stress granule formation by 2Apro and deactivation of eIF2α in response to endoplasmic reticulum (ER) stress induced by EV-A71 infection further contributes to the shutdown of host translation. However, EV-A71 uses a fragment of eIF5B resulting from 3Cpro cleavage as a substitute for eIF2α in order to continue viral RNA translation.16,17 In addition 3Cpro cleaves many other cellular factors to evade cellular responses against viral infections and also to induce cell apoptosis.18–21
For IRES-mediated translation to take place, 40S ribosomal subunit must be recruited on to the IRES elements using eIFs and auxiliary RNA-binding proteins (RBPs) which are known as IRES trans-acting factors (ITAFs). Most ITAFs are nuclear RBPs that are redistributed to the cytoplasm in response to viral infection and cellular stress (Figure 1). The majority of the ITAFs that have been discovered so far are positive regulators of EV-A71 translation. Some of the ITAFs also regulate viral replication via binding to the cloverleaf structure (SLI) of the 5´-UTR. Given the fact that the cellular localization and functional roles of most of these ITAFs have been changed upon EV-A71 infection, it is clear that EV-A71 naturally reprograms the cellular environment to achieve an optimal viral replication. Here, we review up-to-date information on ITAFs of EV-A71 that have been identified so far (Table 1) and how they are repurposed during EV-A71 replication, that will aid in the process of developing more effective antiviral therapies against EV-A71 infection.
2. Cellular IRES trans-acting factors (ITAFs) that regulate EV-A71 translation
2.1. hnRNP A1
Heterogeneous nuclear ribonucleoproteins (hnRNPs) are a family of diverse cellular proteins that play important roles in most aspects of RNA metabolism.22 The quintessential member of this family of proteins is the abundantly expressed hnRNP A1. HnRNP A1 is primarily involved in the regulation of alternative splicing;23 however, it is also known to modulate transcription, translation, stability, and nucleo-cytoplasmic export of mRNAs.24–27 In addition to its regulatory functions in mRNA metabolism, hnRNP A1 is involved in micoRNA (miRNA) processing, telomere maintenance, and modulation of transcription factor activity.24,28,29 HnRNP A1 localizes to the nucleus but has the ability to transport mature mRNAs to the cytoplasm under stress-induced conditions.24,30,31 HnRNP A1 achieves its varied functions in part due to its ubiquity and preference to bind short degenerate sequences, consisting of a minimal 5´-YAG-3´ motif (Figure 3d).32
Upon EV-A71 infection, hnRNP A1 re-localizes to the cytoplasm where it binds specifically to the SLII and SLVI domains of the viral IRES.15,33–36 Individual knockdowns of hnRNP A1 and its homolog hnRNP A2 showed minor effects on IRES activity, viral RNA synthesis, and viral 3C protease (3Cpro). Interestingly, however, simultaneous knockdown of both hnRNP A1 and A2 resulted in a significant attenuation of IRES activity and reduction of viral titers by 4 log10 units, indicating that these two proteins are functionally interchangeable.33 The SLII-hnRNP A1 interaction has been thoroughly characterized biophysically and functionally. HnRNP A1 binds to a phylogenetically conserved bulge loop to change the structure of SLII, which in turn stimulates IRES activity. Mutations or deletions to the bulge loop abrogates hnRNP A1 binding, significantly attenuates IRES translation and inhibits EV-A1 replication by ~5 log10 units.15,34,37 Thus, the recruitment of hnRNP A1 to the EV-A71 5’-UTR is essential for efficient viral replication.
2.2. AUF1 (hnRNP D)
AU-rich element RNA-binding protein 1 (AUF1) also known as hnRNP D is found in 4 isoforms, p37, p40, p42, and p45 that are generated by alternative splicing and numbered based on their molecular weight.38 HnRNP D is primarily responsible for mRNA decay via binding to AU-rich elements (AREs) in order to regulate mRNA turnover.38,39 AREs are destabilizing elements found in most mRNA.40 Additionally, AUF1 is involved in activating telomerase expression, in repressing senescence, and in maintaining normal aging.41 AUF1 is also known to associate with several protein complexes.42–44
Upon EV-A71 cell infection, AUF1 is transported from the nucleus to the cytoplasm and it associates with the EV-A71 5’-UTR.41 AUF1 specifically binds the SLII of the IRES region.41,45 The knockdown of AUF1 showed an increase in IRES activity, production of viral 3Cpro, and viral titers while no effect on viral RNA synthesis or cap-dependent translation.41 Hence, AUF1 is a negative regulator of IRES-dependent translation and viral replication.
Both AUF1 and hnRNP A1 binds to the same bulge region within the SLII. HnRNP A1 binding enhances IRES activity whereas AUF1 binding inhibits IRES activity in a putative mechanism that tunes the levels of IRES-dependent translation to meet the replication needs of EV-A71.45 A competitive binding assay between hnRNP A1 and AUF1 demonstrated that the levels of SLII-bound hnRNP A1 reduced upon increasing levels of AUF1.46 However, the combined knockdowns of hnRNP A1/A2 and AUF1 had no effect on IRES activity shows that IRES has an intrinsic function that can be tuned by these proteins as suggested by Lin et al.46 In addition to EV-A71, AUF1 has been reported to negatively regulate the replication of three related picornaviruses; poliovirus, human rhinovirus, and coxsackievirus in mammalian cells.46,47
Small virus-derived RNAs (at least vsRNA1-4) will be generated in EV-A71 infected cells due to the cleavage of EV-A71 5´-UTR by Dicer. Out of these vsRNAs, vsRNA1 is found to bind the SLII of the IRES to reduce IRES-mediated translation. It has been discovered that vsRNA1 promoted the association of AUF1 with SLII as well as the association of 2 other positive ITAFs, Ago2 and HuR that will be discussed in detail later in the review. It is possible that vsRNA1 control EV-A71 translation and replication via its ability to impact ITAF-IRES interactions at SLII. 16,48
2.3. hnRNP K
HnRNP K is a versatile protein with functional roles in nucleus as well as the cytoplasm.39 It plays diverse roles in mRNA metabolism including regulation of transcription (via binding to CT-rich promoter regions), alternative splicing, mRNA silencing during cell differentiation (via interaction with GSK3β), mRNA stability (during cellular stress conditions) and translation.49–53 HnRNP K interacts with RNA via recognizing CU-rich RNA patches while it can also interact with proteins using its Lysine (K)-rich domains.54
Upon EV-A71 infection, hnRNP K that was localized in the nucleus will be enriched in the cytoplasm.35 Lin et al. showed that hnRNP K binds to the 5’-UTR, specifically at the regions SL I-II and SLIV to regulate EV-A71 replication. The knockdown of hnRNP K in EV-A71 infected cells reduced viral replication and delayed the synthesis of positive and negative RNA strands.35 These results indicate that hnRNP K positively regulates EV-A71 replication. The results by Lin et al.35 don’t show that hnRNP K enhance IRES-mediated translation even though it binds at the SLIV of IRES region. However, interaction of hnRNP K at SLIV may have a stabilization effect on EV-A71 that will promote viral RNA synthesis.
2.4. PCBP1 (hnRNP E1)
Poly(C)-binding protein 1 (PCBP1), also known as hnRNP E1, is a host protein located primarily in the nucleus and can be shuttled between nucleus and cytoplasm. It has three K-homology (KH) domains that mediates RNA binding.55,56 PCBP1 binds to CU-rich regions within the 3’-UTR of mRNA to increase its stability.57 PCBP1 also has roles in modulating alternative splicing, mRNA silencing, transcription and translation.58–61 It is known that PCBP1 interacts with the 5′-UTR of poliovirus (PV) RNA and facilitate the viral RNA replication.56 However, the mechanisms by which PCBP1 facilitate viral replication and translation of many viruses are not known.
Upon EV-A71 infection, PCBP1 primarily localizes in the cytoplasm and co-localizes with the EV-A71 RNA in the ER-derived membrane. PCBP1 specifically binds to the 5’-UTR at SLI and IV using KH1 domain. Knockdown of PCBP1 decreased the level of VP1 viral protein while the overexpression of PCBP1 increased the VP1 production. Further, knockdown of PCBP1 resulted in reduced EV-A71 viral titers while the overexpression of PCBP1 increased the titers. Hence, PCBP1 binds the EV-A71 5´-UTR and positively regulate viral protein expression and virus production.56
2.5. PTB (hnRNP I)
The polypyrimidine tract-binding protein 1 (PTB or PTBP1), also known as hnRNP I, is an RNA binding protein where it interacts with polypyrimidine stretches (CA repeats or CA-rich elements) on RNA mediated by four RNA recognition motifs (RRMs) (Figure 3e).39,62 Even though PTB is mostly known for its role in the regulation of alternative splicing of pre-mRNAs63–65; it is also involved in various other cellular processes including mRNA stabilization, regulation of mRNA translation and miRNA mediated regulation of gene expression.66–68
Upon cell infection, PTB is translocated from the nucleus to the cytoplasm to participate in viral processes. A study by Xi et al. have discovered that PTB specifically binds to the SL VI (stem-loop VI and linker region, nt 564-742) of IRES using RRM1-2. Their results further demonstrate that PTB is a positive ITAF that enhances the EV-A71 IRES-mediated initiation of translation, viral protein expression and virus production.69
2.6. FBP2
The far upstream element binding protein 2 (FBP2 or FUBP2), which is also known as KH-type splicing regulatory protein (KHSRP or KSRP), is a host protein that plays diverse roles including regulation of transcription, pre-mRNA splicing, and mRNA editing.70–72 FBP2 also regulates mRNA degradation via binding to AREs at 3´-ends as well as maturation of miRNA precursors (Figure 3c).73–75 FBP2 can shuttle between the nucleus and the cytoplasm, and its localization can vary in tissue- and context-dependent manner.74
An extensive study by Lin et al. identified FBP2 as a cellular ITAF that binds to the 5’-UTR of EV-A71, in vitro and in vivo.33 A pull-down assay suggests that FBP2 interact with the regions SLI-II (nt 1–167), SLII-III (nt 91–228) and SLVI and spacer region (nt 566–745) (stem-loop VI and spacer region) in EV-A71 5′-UTR via a region that involves at least KH2 and KH4 domains. They further showed that FBP2 is relocalized from the nucleus to the cytoplasm during EV-A71 infection. Knockdown of FBP2 resulted in an increase in viral protein synthesis while FBP2 overexpression resulted in a decrease in viral protein synthesis. Further, FBP2 was shown to negatively regulate IRES-dependent translation to confirm the role of FBP2 as a negative regulator (negative ITAF) of IRES function. The competitive binding assay between PTB and FBP2 suggests that FBP2 acts as a negative ITAF via its ability to bind IRES competitively with positive ITAFs, such as PTB.36
- N- and C-terminus cleaved FBP2
Upon investigating FBP2, truncated products of the protein were detected in EV-A71 infected cells. The nonstructural proteins of EV-A71; 2A, 2B, 2C, 2BC, 3A, 3AB, 3C and 3D, are not responsible for the cleavage FBP2. However, the truncation of FBP2 is a result of viral replication that was found to occur through various virus-induced mechanisms involving caspase activation, proteasomes, and autophagy.76 Five truncated products were detected, two of which, FBP21–503 and FBP2190 –711, bind to the 5’-UTR. Like full length FBP2, FBP2190 –711 negatively regulates viral translation, while FBP21–503 is a positive regulator of IRES-driven translation and viral protein synthesis.68 Hence, EV-A71 infection cleaves FBP2 at the C-terminus to reverse its function from a negative to a positive regulator of viral translation while retaining the ability of FBP2 to bind to the 5’-UTR.76
2.7. FBP1
The far upstream element binding protein 1 (FBP1 or FUBP1), which is highly homologous to FBP2, can bind to RNA or ssDNA.77 FBP1 activates the transcription of a proto-oncogene c-myc (cellular myelocytomatosis oncogene) that mediates cell growth, by binding to the far upstream element (FUSE) upstream of the c-myc promoter.78 It also inhibits the translation of nucleophosmin by binding to the 3’-UTR of its mRNA.79 Additionally, FBP1 plays a role in post-transcriptional regulation of a growth-associated protein 43 (GAP43) in neural development. It promotes GAP43 mRNA degradation through binding to a pyrimidine-rich region at the 3’-UTR of the transcript.77 FBP1 is known to interact with the UTRs Hepatitis C virus (HCV) and Japanese encephalitis virus (JEV) RNA genomes to positively and negatively regulate viral replication, respectively.80,81
A study by Huang et al. identified FBP1 as a positive ITAF for EV-A71 replication.82 Similar to FBP2, FBP1 naturally resides in the nucleus. However, upon EV-A71 infection, FBP1 is redistributed in the cytoplasm, where most of the steps in viral replication takes place.82 In vitro and in vivo studies show that FBP1 binds to the EV-A71 5’-UTR specifically at the linker region downstream of the IRES (nt 686–714). FBP1 contains four KH-type RNA binding domains (KH1-4) flanked by N- and C-terminal domains. However, KH3-4 is enough for FBP1 to bind the EV-A71 5´-UTR. FBP1 has demonstrated that it can positively regulate the IRES-dependent translation of EV71 via binding the 5´-UTR. FBP1 and FBP2 both bind to the linker region downstream of IRES but have opposing roles in IRES-mediated translation by acting as a positive and a negative regulator, respectively. An in vitro competition binding assay between FBP1 and FBP2 revealed that FBP1 outcompeted FBP2 in binding to the IRES linker region. This result suggests that FBP1 may act as a positive ITAF by preventing the negative ITAF, FBP2 binding to the IRES linker region.82
- C-terminus cleaved FBP1
EV-A71 induces the cleavage of FBP1 during the middle stages of infection. The viral protease 2A (2Apro) is responsible for the truncation of FBP1 into its primary cleavage product, FBP11-371. However, unlike in case of FBP2, the FBP1 cleavage is not due to proteasome, lysosome, or caspase activity.83 Similar to intact FBP1, FBP11-371 binds to the linker region of the 5’-UTR but at a different nucleotide sequence (nt 656-674). The non-competitive and simultaneous binding of FBP1 and FBP11-371 additively enhance IRES-mediated translation as well as EV-A71 viral yield.83
2.8. Ago2
The Argonaute 2 (Ago2) protein is implicated in transcriptional and post-transcriptional gene silencing.84 Ago2 is a component of the RNA-induced silencing complex (RISC), where Ago2 binds to a guide RNA, such as a microRNA or a short interfering RNA (Figure 3a).84,85 Silencing occurs through binding of this guide RNA to a complementary strand on the target mRNA.84,85 This will either cause an endonucleolytic cleavage of the mRNA by Ago2 or the inhibition of translation.84–86 The silencing pathway chosen is based on the degree of complementarity between the guide RNA and its target mRNA.84 In addition, Ago 2 was found to upregulate translation via binding to AU-rich elements at the 3'-UTR.87
Ago2 is cellularly localized in the nucleus and the cytoplasm.86 However, Ago2’s localization upon EV-A71 infection has not been reported. Ago2 was identified as an ITAF that specifically binds at the SLII of EV-A71 5´-UTR. The knockdown of Ago2 resulted in reduced IRES activity, demonstrating that Ago2 is a positive ITAF. Further, Ago2 knockdown resulted in a reduction of 3Cpro expression and viral yields.48
2.9. HuR
Human antigen R (HuR), also known as ELAV-like RNA-binding protein 1 (ELAVL1). HuR binds various mRNAs via recognition of AU-rich elements to regulate their stability and translation (Figure 3f).88,89 HuR is predominantly located in the nucleus but shuttles between nucleus and cytoplasm.90 The N-terminal RRM1 and 2 of HuR mediates its interactions with poly-U or AU-rich elements while RRM3 mediates its interactions with the poly-A tail of target mRNAs.90,91 HuR has also shown to positively regulate the expression of Ago2.92
Upon EV-A71 infection, HuR previously localized primarily in the nucleus is translocated into the cytoplasm.93 HuR was identified as an ITAF that specifically binds at the SLII of EV-A71 5´-UTR. The knockdown of HuR resulted in reduced IRES activity, demonstrating that HuR is a positive ITAF. Further, HuR knockdown resulted in a reduction of 3Cpro expression and viral yields.48
- Additive effects of Ago2 and HuR
The simultaneous knockdown of Ago2 and HuR didn’t show an additive effect on the reduction of IRES activity, while the effect was additive on the reduction of 3Cpro expression and viral yields. However, neither independent nor simultaneous knockdown of Ago2 and HuR had an effect on viral RNA replication. These results suggest that Ago2 and HuR are positive regulators of IRES-dependent translation and EV-A71 replication.48
There are many ITAFs that are identified to bind SLII of EV-A71 5´-UTR, including Ago2, HuR, AUF1, hnRNP A1/A2.33,46,48 Competitive binding studies were done to study the dependency of binding of one protein to another. Based on the results from knockdown assays, AUF1, Ago2, or HuR didn’t depend on one another to bind to SLII. However, an increase in vsRNA1 caused an increase in binding of AUF1, Ago2, and HuR to SLII, while the binding of hnRNP A1/A2 were unaffected. Given that AUF1 is a negative regulator and while HuR and Ago2 are positive regulators of IRES-activity, it is speculated that vsRNA1 binding at SLII might alter the binding of AUF1, HuR, and Ago2 at SLII to regulate viral IRES-mediated translation.48
2.10. MOV10
The Moloney Leukemia Virus 10 (MOV10) is a cellular RNA helicase protein found in the cytoplasm.94,95 It colocalizes with Ago2 in the RISC complex to mediate microRNA guided mRNA cleavage.96 MOV10 mediates mRNA degradation of thousands of mRNAs by initially binding to the 3’-UTR and then translocating along the 3’-UTR to unfold the structure and disassemble proteins prior to mRNA degradation.95 C-terminus of MOV10 consists of seven highly conserved helicase motifs that account for its RNA binding activity, while its N-terminal CH-domain mediates protein-protein interactions.97
A study by Wang et al. has demonstrated that MOV10 positively regulates EV-A71 replication.94 The knock down of MOV10 drastically reduced the levels of viral protein as well as positive-strand RNA. MOV10 was found to interact with SLI, the cloverleaf-like structure and the IRES of E-AV71 5′-UTR to facilitate viral RNA replication and IRES-dependent translation, respectively. Further, it was shown that MOV10 brings out the positive regulation of EV-A71 replication through its C-terminus (the site of RNA helicase activity), while the N-terminus possesses a potentially inhibitory effect on viral production by inhibiting viral translation via an unknown mechanism. Upon EV-A71 infection, MOV10 formed distinct perinuclear aggregates and co-localized with processing bodies (P-bodies). In addition to its interaction with Ago2, MOV10 was shown to interact with HuR, another positive ITAF that interacts with the EV-A71 5′-UTR.94
2.11. SIRT1
Silent mating type information regulation 2 homolog 1 (SIRT1) is a member of the sirtuin family. It is a NAD+-dependent deacetylase and involved in a broad range of physiological functions including control of gene expression, metabolism and aging.98 SIRT1 has been implicated in obesity-associated metabolic diseases, cancer, aging, cellular senescence, neurodegeneration, and inflammatory signaling in response to environmental stress.98–100 SIRT1 has been known to positively regulate human immunodeficiency virus 1 (HIV-1) transcription and hepatitis B virus (HBV) replication through the deacetylation of Tat and transcription factor AP-1, respectively.101,102
SIRT1 is localized in the nucleus; however, upon EV-A71 infection, the protein is translocated from nucleus to the cytoplasm.103 According to Han et al., SIRT1 interacts with the SLI (cloverleaf structure) and the SLs II, III and V within IRES of EV-A71 5′-UTR to repress viral RNA transcription and IRES-mediated translation, respectively. At the same time, SIRT1 binds viral 3Dpol protein and results in the repression of viral genome replication.103
2.12. Sam68
The 68 kDa Src-associated protein in mitosis (Sam68), also known as the KH domain containing, RNA binding, signal transduction associated 1 (KHDRBS1) protein that is implicated in many processes including cell cycle and signaling, cell growth, alternative splicing, pre-mRNA splicing and trafficking.104–106 Sam68 belongs to the signal transduction and activation of RNA (STAR) protein family as well as the hnRNP K homology (KH) domain family of RNA-binding proteins.107 It contains one KH domain that interacts with RNA and several proline-rich sequences that facilitate protein-protein interactions with SH3- and WW domain-containing proteins.108 The KH domain of Sam68 recognizes and binds RNA using U(U/A)AA direct repeat motifs.109,110
Sam68 is an ITAF that resides in the nucleus and gets relocated to the cytoplasm upon EV-A71 infection. Similar redistribution of Sam from nucleus to cytoplasm has also been reported in cases of FMDV and poliovirus infections.111,112 Sam68 specifically interacts with the SLIV and V of 5’-UTR of EV-A71 RNA using its hnRNP K homology (KH) domain. Sam68 was found to interact with other ITAFs including Poly(c)-binding protein 2 (PCBP2) and Poly(A)-binding protein (PABP) to facilitate viral replication. Sam68 is a positive regulator of IRES-activity, viral protein expression and viral titer. However, Sam68 doesn’t have any regulatory effect on viral genome replication.113
2.13. FUBP3
The far upstream element-binding protein 3 (FUBP3) is a single-stranded NA-binding protein that recognizes only one strand of the far upstream element (FUSE).71 The structure of FUBP3 consists of four regularly spaced K homology (KH) domains that recognize similar sequences in single-stranded DNA or RNA targets.114 FUBP3 has roles in the regulation of transcription, splicing and translation.115 FUBP3 is known to bind the 3′-UTR of Japanese encephalitis virus (JEV) to regulate RNA replication and promote subsequent viral translation and viral particle production.114
A study by Tsai et al. on the inhibition of EV-A71 replication and internal ribosome entry site (IRES) activity by Kaempferol (a flavonoid) has discovered FUBP3 as a new ITAF that associates with EV71 5′-UTR to enhance the IRES-dependent translation.116 Another study by Huang et al. has reported the interaction of FUBP3 with the 5′-UTR of EV-A71 to regulate its replication in differentiated neuronal cells.117 Upon EV-A71 infection, FBP3 in the nucleus will be relocated into the cytoplasm.117
2.14. GADD34
The growth arrest and DNA damage-inducible protein 34 (GADD34) also known as PPP1R15A is a protein that is upregulated in response to various cell stress inducing stimuli. GADD34 interacts with serine/threonine protein phosphatase 1 (PP1) to dephosphorylate eIF2α, thereby restarting protein synthesis for cells to recover from integrated stress response (ISR).118–120 GADD34 is known to play an important role in regulating the interferon response of virus-induced innate immunity.121,122 While there are many reports regarding the role GADD34 in the inhibition of viral replication121–123, GADD34 is shown to promote the replication of infectious bronchitis virus (IBV).124 GADD34 attenuated HIV-1 replication via inhibition of viral protein expression in a mechanism mediated by 5’-UTR/TAR RNA, probably by modulating TAR RNA structure.125
A study by Li et al. demonstrated that EV-A71 activates GADD34 via viral precursor protein 3CD to promote IRES-mediated viral translation. GADD34 is a short-lived protein and is highly expressed only under the conditions of cellular stress.126 Upon EV-A71 infection, 3CD upregulates GADD34 translation via the upstream open reading frame (uORF) within the 5'-UTR of GADD34. GADD34 is unable to bind directly to EV-A71 5′-UTR. Hence, 3CD binds at the SLI (cloverleaf structure)127 and recruits GADD34 to the 5′-UTR of EVA-A71. Once bound at the 5′-UTR, GADD34 promotes the EV-A71 IRES-activity through its PEST repeats (1, 2 and 3). However, the dephosphorylation of eIF2α by GADD34 was unrelated to these observed effects in the upregulation of EV-A71 replication.128
2.15. DDX3
DEAD-box protein 3 is a DEAD-box RNA helicase that regulates translation and is encoded by the X- and Y-linked paralogs DDX3X and DDX3Y. DDX3X is ubiquitously expressed and essential for viability while DDX3Y is male-specific and shows lower and more variable expression in somatic tissues compared to DDX3X. However, the roles of DDX3X and DDX3Y in translation are functionally redundant.129 DDX3 has a relaxed substrate specificity and is implicated in many cellular processes such as gene expression including transcription, splicing, mRNA export, translation, cell cycle control, regulation apoptosis, and innate immune signaling (Figure 3g).130 It has been known that DDX3 is a prime target for viral manipulation during Hepatitis C virus (HCV), Hepatitis B virus (HBV), Human Immunodeficiency Virus (HIV) and poxvirus infections as viral proteins interact with DDX3 to utilize its function for the process of viral replication.130 It has been shown that DDX3X is specifically required for the translation initiation of transcripts that possess highly stable secondary structures within their 5′-UTR that resist the unwinding activity of eIF4A. DDX3X binds the 5′-UTR via its interactions with eIF4G and PABP, and works in corporation with eIF4A to destabilize the secondary structure and facilitate ribosome entry.131
DDX3X shuttles between the nucleus and the cytoplasm and possesses RNA-dependent ATPase/helicase activity.130 Su et al. demonstrated that DDX3X binds the 5′-UTR of EV-A71 and enhances IRES-dependent translation, partly mediated by viral 2Apro and 3Cpro protease activity. DDX3X can bind efficiently to IRES + spacer, SLI-III, SLIV-VI, and SLVI + spacer regions in the EV-A71 5′-UTR, irrespective of EV71 infection. Their results strongly suggest that the truncated eIF4G (cleaved by viral 2Apro) binds specifically to SLV and recruits DDX3X to SLVI or a region nearby to locally unwind the secondary structure of SLVI, thereby facilitating ribosome entry and scanning. They also showed that DDX3X also enhances the IRES activity of coxsackievirus A16, Echovirus 9, EMCV, and HCV implicating DDX3X is general cellular factor for the translation of these highly structured viral IRESs.132
2.16. APOBEC3G
APOBEC3G (apolipoprotein B mRNA-editing enzyme, catalytic polypeptide-like 3G) or A3G is a member of the APOBEC superfamily. A3G is a cytidine deaminase that contains a conserved His-X-Glu and Cys-X-X-Cys Zn2+ coordination motif. A3G is an interferon-inducible cellular protein and plays an important role in defending against viral infections. It has been demonstrated to inhibit the infection of several viruses such as human immunodeficiency virus-1 (HIV-1), T-cell leukemia virus type 1 (HTLV-1), hepatitis B virus (HBV) and hepatitis C virus (HCV).133,134 A3G can inhibit viral replication in a cytidine deaminase activity-dependent (C-terminal domain) manner as well as in a deaminase activity-independent manner where its N-terminal domain’s RNA binding ability that mediates incorporation of A3G into viral particles thereby disrupting reverse transcription or genome encapsidation.135–137
Li et al. demonstrated that A3G inhibits EV-A71 virus replication via competitive binding to the 5′UTR of EV-A71 and inhibiting the 5′UTR activity. A3G binding impaired the interaction between the 5′-UTR and the host protein poly(C)-binding protein 1 (PCBP1), an ITAF which enhances the viral RNA replication and IRES-mediated translation. A3G was shown to bind the SLI and II while PCBP1 is known to bind to SLI and IV of EV-A71 5′-UTR56,138. Hence, A3G binds at SLI and competitively inhibits the binding of PCBP1 at the same location, as suggested by the higher binding affinity of A3G compared to PCBP1 towards the 5′-UTR and the reduction of 5′UTR-PCBP1 interactions with increasing A3G expression levels. However, EV-A71 has developed mechanisms to overcome the suppression by A3G via its degradation through the autophagy–lysosome pathway mediated by viral protein 2Cpro.138
2.17. Staufen1
Staufen1 is one of the two homologs of Staufen, a double-stranded RNA (dsRNA) and tubulin-binding protein.139 Staufen1 contains four dsRNA-binding domains (RBDs) of which dsRBD2-4 are shown to bind dsRNA (Figure 3h).140 Staufen1 is known for its ability to regulate cellular mRNA translation, trafficking, and degradation via Staufen1-mediated RNP formation. Staufen1 enhances the translation efficiency via binding to the 5′-UTR while it can bind to the 3′-UTR to promote mRNA degradation.141,142 A study by Chen et al. has demonstrated that Staufen1 facilitates both translation and replication of the EV-A71 genome. The RBD2-3 of Staufen1 was identified to interact with the 5´-UTR EV-A71 to enhance the IRES activity as well as the translation efficiency. Further, binding of Staufen1 at the 5´-UTR increased the stability of viral RNA.143
Staufen1 has been reported to play roles in the life cycles of other RNA viruses, including Hepatitis C virus (HCV), influenza A virus, and HIV-1. During the infection of HCV, Staufen1 demonstrated roles in the viral replication, translation and trafficking of the HCV genome,144 while it facilitated in the viral particle assembly of HIV-1 and Influenza A viruses.145,146
2.18. hnRNP H and hnRNP F
The two hnRNP proteins, hnRNP F and hnRNP H are closely related and fall under the hnRNP F/H subfamily. They are found to act as activators as well as repressors in regulating alternative splicing, depending on the context of the binding site.147 HnRNP F/H possess three quasi-RNA recognition motifs (qRRMs) that preferably bind to poly(G)-rich sequences in the target exons and/or adjacent introns in order to regulate alternative splicing and 3′-end processing of numerous genes (Figure 3b).148
Both hnRNP H and F are located in the nucleus and are relocated into the cytoplasm upon EV-A71 infection. A study by Tsai et al. on the inhibition of EV-A71 by Kaempferol (a flavonoid) via impairing its replication and internal ribosome entry site (IRES) activity has discovered hnRNP F and hnNNP H as new ITAFs that associates with EV-A71 5′-UTR to enhance the IRES activity. Both hnRNP H and F were discovered initially as kaempferol-induced cellular factors associated with the 5′-UTR of the EV-A71 genome. However, they have carried out individual knockdown of endogenous HNRH1 and HNRPF proteins to show that it resulted in the decreased EV-A71 IRES activity. Unfortunately, the specific binding sites of hnRNP H and F on the EV-A71 5´-UTR have not been mapped.116
2.19. EGR1
Early growth response-1 (EGR1) is a C2H2-type zinc finger protein, and it is a transcription factor that activates many genes essential for growth, proliferation, or differentiation via binding to GC-rich recognition motifs. EGR1 expression is induced in response to various extracellular stimuli including growth factors, hormones, and neurotransmitters. EGR1 then couples extracellular signals to long-term cellular responses by altering gene expression of its target genes.149 EGR1 is found to activate microRNA-141 (miR-141) expression to suppresses eukaryotic initiation factor 4E (eIF4E) production, resulting in the facilitation of EV-A71 replication via shutting off host protein synthesis.150
A study by Song et al. has revealed that the first two zinc fingers of EGR1 bind directly to SLI and SLIV of the EV-A71 5′-UTR to regulate viral replication via enhancing both IRES-mediated translation and RNA replication. Further, this study demonstrated that EGR1 facilitates EV-A71 replication in a manner independent of miR-141 and eIF4E, via its interactions with the 5′-UTR.151
2.20. TIA-1 and TIAR
T-cell intracellular antigen 1 (TIA-1) and TIA-1-related protein (TIAR) have been identified as nucleating components of mammalian stress granules (SGs).152 TIA proteins consist of three RNA-binding domains and a glutamine-rich carboxyl-terminal domain that enable aggregation to insoluble aggregates. TIA proteins have many important roles in regulation of mRNA metabolism especially during environmental stress conditions, including the regulation of splicing, mRNA stability, storage, and translation efficiency. The TIAR/TIA-1 proteins are known to selectively target specific mRNAs via binding to specific AREs located at the 3′ end of the target mRNAs.153 TIA-1 and TIAR has been reported to be translocated from nucleus to cytoplasm to form SG-like granules as a result of poliovirus and enterovirus (CVB3 or EV-A71) infections.154,155
Wang et al. demonstrated that TIA1 and TIAR were recruited into SGs following EV-A71 infection and were localized to the sites of viral replication. Both TIA-1 and TIAR interact only with the SLI (nt 1–105) of EV-A71 5′-UTR and results in positive regulation of viral replication. The silencing of either TIA-1 or TIAR expression significantly reduced not only viral replication but also viral progeny production, via regulation of the level of viral RNA. Even though TIA-1 and TIAR bind to AREs, RNA pull-down assays proved that they didn’t bind to 5′-UTR or 3′-UTR of EV-A71 genome except SLI, making both TIA1 and TIAR distinct from many ITAFs that are shown to interact with multiple loops within 5′-UTR.156
2.21. Additional cellular proteins
In addition to the proteins listed in Table 1, there are many additional cellular proteins that were discovered to associate with EV-A71 5’-UTR while their specific binding sites and roles in EV-A71 viral replication have not yet been determined. These proteins were identified by pull down assays utilizing biotin-labeled 5’-UTR followed by MALDI-TOF MS characterization. Such additional cellular proteins that were discovered by individual studies carried out by Lin et al. and Xi et al. are as follows: TBP-associated factor TAFII 150 (TBP-AFII 150), N-ras upstream protein (Unr), Glycyl-tRNA synthetase (G tRNA synthetase), Keratin, IGF-II mRNA-binding protein 1 (IMP-1), Polypyrimidine tract-binding protein 2 (PTB-2), ErbB3-binding protein (EBP1), Poly(rC)-binding protein 2 (PCBP2), pro alpha (I) collagen, Lrp protein, PTB-associated splicing factor (PSF), Splicing factor, proline- and glutamine-rich (SFPQ), Cationic trypsinogen, CGI-55 protein, Elongation factor 1-gamma (EEF1G), 2-phosphopyruvate-hydratase alpha-enolase, Collagen binding protein 2, Mutant beta-actin, DNA-binding protein.35,36,69
3. Discussion
Upon EV-A71 infection, the host cellular pathways will be repurposed to facilitate viral replication. Some of the main pathways that are perturbed include innate immunity, RNA metabolism, nuclear-cytoplasmic transport, stress granule formation and autophagy. EV-A71 uses two main strategies to achieve the repurposing of cellular machinery. The first strategy is redistribution of cellular proteins from their original locations into the cytoplasm where viral replication takes place. This will perturb their original cellular functions while being utilized solely in different steps of viral replication. The second strategy is the cleavage of key cellular proteins involved in above mentioned cellular pathways, mediated by viral proteases (mainly 3Cpro and 2Apro) or other virus-induced pathways (caspases, proteosome, and autophagy) (Figure 4).
As the viral RNA genome is released into the cytoplasm upon EV-A71 infection, viral proteins are translated in an IRES-dependent manner while the viral genome is replicated via mechanisms mediated by RNA-dependent RNA polymerase (RdRp) 3Dpol. The 5´-UTR of the viral RNA genome, which is composed of the cloverleaf structure (SLI) and the IRES (SLII-VI), functions as a control hub for viral RNA transcription and translation via recruiting viral and cellular proteins (Figure 1). Cellular RNA-binding proteins that are recruited to the IRES and regulate viral translation are called ITAFs. The regulatory effect of these ITAFs on viral IRES-mediated translation can either be positive or negative. Table 1 summarizes the ITAFs associated with the EV-A71 5´-UTR indicating their effect on viral translation and how they are redistributed in the cell upon EV-A71 infection.
It is noteworthy that hnRNP proteins are a key target of EV-A71, possibly due to the fact that hnRNP family proteins are key players in regulating various aspects of mRNA metabolism including alternative splicing, stability, transcription, and translation while each of them are structurally and functionally diverse. The hnRNP proteins are primarily localized in the nucleus and upon EV-A71 infection, they are relocated into the cytoplasm where most of the steps of viral replication take place. Out of the seven hnRNP proteins listed in this review, hnRNP A1, hnRNP E1 (PCBP1), hnRNP I (PTB), hnRNP H, and hnRNP F act as positive ITAFs that bind EV-A71 5´-UTR to enhance IRES-mediated viral translation. The only hnRNP that acts as a negative ITAF is hnRNP D (AUF1). The hnRNP proteins that act as positive ITAFs also play simultaneous roles in other steps of EV-A71 replication such as viral genome replication.
In addition to the hnRNP proteins, FBP1, Ago2, HuR, MOV10, Sam68, FBP3, GADD34, DDX3, HSPA6, Staufen1, and EGR1 act as positive ITAFs while only FBP2, SIRT1, and APOBEC3G act as negative ITAFs. However, EV-A71 has strategies to repurpose even the negative ITAFs into positive ITAFs. One such example is the cleavage of the C-terminus of FBP2 to convert the negative ITAF into a positive ITAF. ITAFs can have different and unique cellular localizations that are related to their cellular function; however, they will be redistributed into the cytoplasm to bind the EV-A71 5´-UTR and enhance the IRES-mediated translation. Figure 1 visually represents the re-localization of cellular proteins before and after EV-A71 infection as well as their binding sites at the 5´-UTR of EV-A71 RNA genome.
Apart from the relocation of cellular ITAFs into the cytoplasm, the levels of mRNA as well as the translation of positive ITAFs can also be enhanced upon EV-A71 infection, as observed in case of GADD34. Further, EV-A71 viral proteins such as 2Apro or 2Cpro play important roles in the process of repurposing cellular proteins to facilitate viral replication, mainly via their protease activity. Their activity is known to facilitate the redistribution of positive ITAFs from nucleus to the cytoplasm via cleaving certain components of nuclear pore complex. Sometimes, their activity can truncate a cellular protein to give it a new feature that is beneficial for the viral replication as observed in case of eIFG4, where truncated version of eIF4G (cleaved by viral 2Apro) binds specifically to SLV of IRES and recruits positive ITAF DDX3X facilitating ribosome entry and scanning. In some cases, even positive ITAFs are cleaved by viral proteases as observed in the cleavage of FBP1 by 2Apro to give rise to an additive enhancement of IRES activity (Figure 4).
It is clear that the mechanism of fine-tuning the IRES-activity to regulate EV-A71 viral replication is a very complex process involving the interplay between the positive and negative cellular ITAFs as well as the viral proteins. Many recent studies have expanded the repertoire of positive and negative ITAFs suggesting new targets for EV-A71 inhibition. Most importantly, studies that identify the interplay between positive and negative ITAFs that bind at the same site, not only help in understanding the means of IRES- mediated translation regulation but also highlights these sites as better candidates of drug targeting. We believe that a comprehensive and generalized understanding of the mechanism of how EV-A71 hijacks and repurposes host cellular proteins for its gain can guide the development of novel and more efficient approaches for drug targeting.
4. Future perspectives
Considering the increased interest in discovering novel small molecule inhibitors of RNA viruses like EV-A71, it is important to implement robust strategies to identify biologically relevant targets for therapeutic intervention. The 5´-UTR of EV-A71 represents a promising target given its central importance in regulating the cellular stages of viral replication cycles. Moreover, the EV-A71 5´-UTR is under high-selective pressure to maintain its overall structure because it associates with a collection of ITAFs to differentially control viral gene expression. Therefore, the next generation of EV-A71 antivirals to be discovered should function by selectively perturbing specific 5´-UTR-ITAF interactions to drive viral replication towards predictable outcomes (Figure 1 and Figure 2).
To date, SLII is the only RNA structure of the EV-A71 5´-UTR where specific binding sites of several of its ITAFs has been biophysically investigated to reveal the competitive interplay between hnRNP A1 and AUF1 for its bulge loop. Further, this region has been successfully targeted by a small molecule antiviral (DMA-135) where its mechanism of action is to allosterically increase the binding affinity of AUF1 to SLII, thus shifting the SLII regulatory axis towards translation repression. These observations suggest that other small molecules with capacity to selectively perturb specific 5´-UTR-ITAF interactions await discovery. To realize this outcome, we believe that it is prudent to better understand the structural-based mechanisms by which ITAFs assemble onto the EV-A71 5´-UTR, characterize the differential cell-type expression patterns of ITAFs upon viral infection, and perform comparative virological and structural studies of related enteroviruses such as EV-D68. Lastly, we must also remember that ITAFs and viral RNA elements are covalently modified so it should be a priority to understand the extent to which such modifications modulate EV-A71 gene expression at a molecular level. The coordination of these studies will accelerate the discovery of the next generation of antiviral agents that target viral RNA complexes and produce a library of chemical biology reagents that can be deployed to selectively modulate conserved host-virus interactions.
Author Contributions
Conceptualization, B.S.T.; RNA 3D structure predictions and preparation of representative protein-RNA complexes, B.H., S.P.; writing-original draft preparation, J.D.-C., C.H., J.K.; writing-review and editing, S.M.A., B.H., S.P., B.S.T.; All authors have read and agreed to the published version of the manuscript.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Nayak, G.; Bhuyan, S. K.; Bhuyan, R.; Sahu, A.; Kar, D.; Kuanar, A. Global Emergence of Enterovirus 71: A Systematic Review. Beni Suef Univ J Basic Appl Sci 2022, 11, 78. [Google Scholar] [CrossRef] [PubMed]
- Hu, Y.-L.; Chen, C.-M.; Wang, E.-T.; Kuo, H.-W.; Shih, W.-L.; Fang, C.-T.; Liu, D.-P.; Chang, L.-Y. The Secular Trend of Enterovirus A71 after the Implementation of Preventive Measures in Taiwan. BMC Public Health 2022, 22, 1483. [Google Scholar] [CrossRef]
- Enterovirus 71. https://www.who.int/teams/health-product-policy-and-standards/standards-and-specifications/vaccine-standardization/enterovirus-71 (accessed 2023-11-08).
- Huang, P.-N.; Shih, S.-R. Update on Enterovirus 71 Infection. Curr Opin Virol 2014, 5, 98–104. [Google Scholar] [CrossRef] [PubMed]
- Xu, B.; Wang, J.; Yan, B.; Xu, C.; Yin, Q.; Yang, D. Global Spatiotemporal Transmission Patterns of Human Enterovirus 71 from 1963 to 2019. Virus Evolution 2021, 7, veab071. [Google Scholar] [CrossRef] [PubMed]
- Wörner, N.; Rodrigo-García, R.; Antón, A.; Castellarnau, E.; Delgado, I.; Vazquez, È.; González, S.; Mayol, L.; Méndez, M.; Solé, E.; Rosal, J.; Andrés, C.; Casquero, A.; Lera, E.; Sancosmed, M.; Campins, M.; Pumarola, T.; Rodrigo, C. Enterovirus-A71 Rhombencephalitis Outbreak in Catalonia: Characteristics, Management and Outcome. The Pediatric Infectious Disease Journal 2021, 40, 628. [Google Scholar] [CrossRef]
- Liu, H.; Zhang, Y.; Zhang, H.; Zheng, Y.; Gou, F.; Yang, X.; Cheng, Y.; McClymont, H.; Li, H.; Liu, X.; Hu, W. Prototypes Virus of Hand, Foot and Mouth Disease Infections and Severe Cases in Gansu, China: A Spatial and Temporal Analysis. BMC Infectious Diseases 2022, 22, 408. [Google Scholar] [CrossRef] [PubMed]
- Messacar, K.; Burakoff, A.; Nix, W. A.; Rogers, S.; Oberste, M. S.; Gerber, S. I.; Spence-Davizon, E.; Herlihy, R.; Dominguez, S. R. Notes from the Field: Enterovirus A71 Neurologic Disease in Children - Colorado, 2018. MMWR Morb Mortal Wkly Rep 2018, 67, 1017–1018. [Google Scholar] [CrossRef] [PubMed]
- Nhan, L. N. T.; Hong, N. T. T.; Nhu, L. N. T.; Nguyet, L. A.; Ny, N. T. H.; Thanh, T. T.; Han, D. D. K.; Van, H. M. T.; Thwaites, C. L.; Hien, T. T.; Qui, P. T.; Quang, P. V.; Minh, N. N. Q.; van Doorn, H. R.; Khanh, T. H.; Chau, N. V. V.; Thwaites, G.; Hung, N. T.; Tan, L. V. Severe Enterovirus A71 Associated Hand, Foot and Mouth Disease, Vietnam, 2018: Preliminary Report of an Impending Outbreak. Euro Surveill 2018, 23, 1800590. [Google Scholar] [CrossRef] [PubMed]
- NIAID Emerging Infectious Diseases/Pathogens | NIH: National Institute of Allergy and Infectious Diseases. https://www.niaid.nih.gov/research/emerging-infectious-diseases-pathogens (accessed 2023-11-08).
- Li, M.-L.; Shih, S.-R.; Tolbert, B. S.; Brewer, G. Enterovirus A71 Vaccines. Vaccines (Basel) 2021, 9, 199. [Google Scholar] [CrossRef]
- Yi, E.-J.; Shin, Y.-J.; Kim, J.-H.; Kim, T.-G.; Chang, S.-Y. Enterovirus 71 Infection and Vaccines. Clin Exp Vaccine Res 2017, 6, 4–14. [Google Scholar] [CrossRef]
- Chan, Y.-F.; Sam, I.-C.; AbuBakar, S. Phylogenetic Designation of Enterovirus 71 Genotypes and Subgenotypes Using Complete Genome Sequences. Infect Genet Evol 2010, 10, 404–412. [Google Scholar] [CrossRef] [PubMed]
- Brown, B. A.; Pallansch, M. A. Complete Nucleotide Sequence of Enterovirus 71 Is Distinct from Poliovirus. Virus Res 1995, 39, 195–205. [Google Scholar] [CrossRef] [PubMed]
- Tolbert, M.; Morgan, C. E.; Pollum, M.; Crespo-Hernández, C. E.; Li, M.-L.; Brewer, G.; Tolbert, B. S. HnRNP A1 Alters the Structure of a Conserved Enterovirus IRES Domain to Stimulate Viral Translation. J Mol Biol 2017, 429, 2841–2858. [Google Scholar] [CrossRef] [PubMed]
- Lai, M.-C.; Chen, H.-H.; Xu, P.; Wang, R. Y. L. Translation Control of Enterovirus A71 Gene Expression. J Biomed Sci 2020, 27, 22. [Google Scholar] [CrossRef] [PubMed]
- Sun, D.; Chen, S.; Cheng, A.; Wang, M. Roles of the Picornaviral 3C Proteinase in the Viral Life Cycle and Host Cells. Viruses 2016, 8, 82. [Google Scholar] [CrossRef] [PubMed]
- Weng, K.-F.; Li, M.-L.; Hung, C.-T.; Shih, S.-R. Enterovirus 71 3C Protease Cleaves a Novel Target CstF-64 and Inhibits Cellular Polyadenylation. PLoS Pathog 2009, 5, e1000593. [Google Scholar] [CrossRef] [PubMed]
- Lei, X.; Xiao, X.; Xue, Q.; Jin, Q.; He, B.; Wang, J. Cleavage of Interferon Regulatory Factor 7 by Enterovirus 71 3C Suppresses Cellular Responses. J Virol 2013, 87, 1690–1698. [Google Scholar] [CrossRef] [PubMed]
- Lei, X.; Han, N.; Xiao, X.; Jin, Q.; He, B.; Wang, J. Enterovirus 71 3C Inhibits Cytokine Expression through Cleavage of the TAK1/TAB1/TAB2/TAB3 Complex. J Virol 2014, 88, 9830–9841. [Google Scholar] [CrossRef] [PubMed]
- Li, M.-L.; Lin, J.-Y.; Chen, B.-S.; Weng, K.-F.; Shih, S.-R.; Calderon, J. D.; Tolbert, B. S.; Brewer, G. EV71 3C Protease Induces Apoptosis by Cleavage of hnRNP A1 to Promote Apaf-1 Translation. PLoS One 2019, 14, e0221048. [Google Scholar] [CrossRef]
- Wang, J.; Sun, D.; Wang, M.; Cheng, A.; Zhu, Y.; Mao, S.; Ou, X.; Zhao, X.; Huang, J.; Gao, Q.; Zhang, S.; Yang, Q.; Wu, Y.; Zhu, D.; Jia, R.; Chen, S.; Liu, M. Multiple Functions of Heterogeneous Nuclear Ribonucleoproteins in the Positive Single-Stranded RNA Virus Life Cycle. Front Immunol 2022, 13, 989298. [Google Scholar] [CrossRef]
- Mayeda, A.; Krainer, A. R. Regulation of Alternative Pre-mRNA Splicing by hnRNP A1 and Splicing Factor SF2. Cell 1992, 68, 365–375. [Google Scholar] [CrossRef] [PubMed]
- Jean-Philippe, J.; Paz, S.; Caputi, M. hnRNP A1: The Swiss Army Knife of Gene Expression. Int J Mol Sci 2013, 14, 18999–19024. [Google Scholar] [CrossRef] [PubMed]
- Michael, W. M.; Choi, M.; Dreyfuss, G. A Nuclear Export Signal in hnRNP A1: A Signal-Mediated, Temperature-Dependent Nuclear Protein Export Pathway. Cell 1995, 83, 415–422. [Google Scholar] [CrossRef] [PubMed]
- Henics, T.; Sanfridson, A.; Hamilton, B. J.; Nagy, E.; Rigby, W. F. Enhanced Stability of Interleukin-2 mRNA in MLA 144 Cells. Possible Role of Cytoplasmic AU-Rich Sequence-Binding Proteins. J Biol Chem 1994, 269, 5377–5383. [Google Scholar] [CrossRef] [PubMed]
- Hamilton, B. J.; Burns, C. M.; Nichols, R. C.; Rigby, W. F. Modulation of AUUUA Response Element Binding by Heterogeneous Nuclear Ribonucleoprotein A1 in Human T Lymphocytes. The Roles of Cytoplasmic Location, Transcription, and Phosphorylation. J Biol Chem 1997, 272, 28732–28741. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.-S.; Manche, L.; Xu, R.-M.; Krainer, A. R. hnRNP A1 Associates with Telomere Ends and Stimulates Telomerase Activity. RNA 2006, 12, 1116–1128. [Google Scholar] [CrossRef] [PubMed]
- Guil, S.; Cáceres, J. F. The Multifunctional RNA-Binding Protein hnRNP A1 Is Required for Processing of miR-18a. Nat Struct Mol Biol 2007, 14, 591–596. [Google Scholar] [CrossRef] [PubMed]
- Mili, S.; Shu, H. J.; Zhao, Y.; Piñol-Roma, S. Distinct RNP Complexes of Shuttling hnRNP Proteins with Pre-mRNA and mRNA: Candidate Intermediates in Formation and Export of mRNA. Mol Cell Biol 2001, 21, 7307–7319. [Google Scholar] [CrossRef]
- Piñol-Roma, S.; Dreyfuss, G. Shuttling of Pre-mRNA Binding Proteins between Nucleus and Cytoplasm. Nature 1992, 355, 730–732. [Google Scholar] [CrossRef]
- Jd, L.; Bs, T. Idiosyncrasies of hnRNP A1-RNA Recognition: Can Binding Mode Influence Function. Seminars in cell & developmental biology 2019, 86. [Google Scholar] [CrossRef]
- Lin, J.-Y.; Shih, S.-R.; Pan, M.; Li, C.; Lue, C.-F.; Stollar, V.; Li, M.-L. hnRNP A1 Interacts with the 5’ Untranslated Regions of Enterovirus 71 and Sindbis Virus RNA and Is Required for Viral Replication. J Virol 2009, 83, 6106–6114. [Google Scholar] [CrossRef] [PubMed]
- Levengood, J. D.; Tolbert, M.; Li, M.-L.; Tolbert, B. S. High-Affinity Interaction of hnRNP A1 with Conserved RNA Structural Elements Is Required for Translation and Replication of Enterovirus 71. RNA Biol 2013, 10, 1136–1145. [Google Scholar] [CrossRef] [PubMed]
- Lin, J.-Y.; Li, M.-L.; Huang, P.-N.; Chien, K.-Y.; Horng, J.-T.; Shih, S.-R. Heterogeneous Nuclear Ribonuclear Protein K Interacts with the Enterovirus 71 5’ Untranslated Region and Participates in Virus Replication. J Gen Virol 2008, 89 Pt 10, 2540–2549. [Google Scholar] [CrossRef] [PubMed]
- Lin, J.-Y.; Li, M.-L.; Shih, S.-R. Far Upstream Element Binding Protein 2 Interacts with Enterovirus 71 Internal Ribosomal Entry Site and Negatively Regulates Viral Translation. Nucleic Acids Res 2009, 37, 47–59. [Google Scholar] [CrossRef] [PubMed]
- Davila-Calderon, J.; Li, M.-L.; Penumutchu, S. R.; Haddad, C.; Malcolm, L.; Hargrove, A. E.; Brewer, G.; Tolbert, B. S. Enterovirus Evolution Reveals the Mechanism of an RNA-Targeted Antiviral and Determinants of Viral Replication. bioRxiv February 20, 2023, p 2023.02.20.529064. [CrossRef]
- Fialcowitz, E. J.; Brewer, B. Y.; Keenan, B. P.; Wilson, G. M. A Hairpin-like Structure within an AU-Rich mRNA-Destabilizing Element Regulates Trans-Factor Binding Selectivity and mRNA Decay Kinetics. J Biol Chem 2005, 280, 22406–22417. [Google Scholar] [CrossRef] [PubMed]
- Geuens, T.; Bouhy, D.; Timmerman, V. The hnRNP Family: Insights into Their Role in Health and Disease. Hum Genet 2016, 135, 851–867. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.; Brewer, G. The Regulation of mRNA Stability in Mammalian Cells: 2.0. Gene 2012, 500, 10. [Google Scholar] [CrossRef] [PubMed]
- Pont, A. R.; Sadri, N.; Hsiao, S. J.; Smith, S.; Schneider, R. J. mRNA Decay Factor AUF1 Maintains Normal Aging, Telomere Maintenance, and Suppression of Senescence by Activation of Telomerase Transcription. Mol Cell 2012, 47, 5–15. [Google Scholar] [CrossRef]
- Laroia, G.; Cuesta, R.; Brewer, G.; Schneider, R. J. Control of mRNA Decay by Heat Shock-Ubiquitin-Proteasome Pathway. Science 1999, 284, 499–502. [Google Scholar] [CrossRef]
- Lu, J.-Y.; Bergman, N.; Sadri, N.; Schneider, R. J. Assembly of AUF1 with eIF4G–Poly(A) Binding Protein Complex Suggests a Translation Function in AU-Rich mRNA Decay. RNA 2006, 12, 883–893. [Google Scholar] [CrossRef]
- Sinsimer, K. S.; Gratacós, F. M.; Knapinska, A. M.; Lu, J.; Krause, C. D.; Wierzbowski, A. V.; Maher, L. R.; Scrudato, S.; Rivera, Y. M.; Gupta, S.; Turrin, D. K.; De La Cruz, M. P.; Pestka, S.; Brewer, G. Chaperone Hsp27, a Novel Subunit of AUF1 Protein Complexes, Functions in AU-Rich Element-Mediated mRNA Decay. Mol Cell Biol 2008, 28, 5223–5237. [Google Scholar] [CrossRef] [PubMed]
- Davila-Calderon, J.; Patwardhan, N. N.; Chiu, L.-Y.; Sugarman, A.; Cai, Z.; Penutmutchu, S. R.; Li, M.-L.; Brewer, G.; Hargrove, A. E.; Tolbert, B. S. IRES-Targeting Small Molecule Inhibits Enterovirus 71 Replication via Allosteric Stabilization of a Ternary Complex. Nat Commun 2020, 11, 4775. [Google Scholar] [CrossRef] [PubMed]
- Lin, J.-Y.; Li, M.-L.; Brewer, G. mRNA Decay Factor AUF1 Binds the Internal Ribosomal Entry Site of Enterovirus 71 and Inhibits Virus Replication. PLoS One 2014, 9, e103827. [Google Scholar] [CrossRef] [PubMed]
- Cathcart, A. L.; Rozovics, J. M.; Semler, B. L. Cellular mRNA Decay Protein AUF1 Negatively Regulates Enterovirus and Human Rhinovirus Infections. J Virol 2013, 87, 10423–10434. [Google Scholar] [CrossRef] [PubMed]
- Lin, J.-Y.; Brewer, G.; Li, M.-L. HuR and Ago2 Bind the Internal Ribosome Entry Site of Enterovirus 71 and Promote Virus Translation and Replication. PLoS One 2015, 10, e0140291. [Google Scholar] [CrossRef] [PubMed]
- Stains, J. P.; Lecanda, F.; Towler, D. A.; Civitelli, R. Heterogeneous Nuclear Ribonucleoprotein K Represses Transcription from a Cytosine/Thymidine-Rich Element in the Osteocalcin Promoter. Biochem J 2005, 385 Pt 2, 613–623. [Google Scholar] [CrossRef] [PubMed]
- Cao, W.; Razanau, A.; Feng, D.; Lobo, V. G.; Xie, J. Control of Alternative Splicing by Forskolin through hnRNP K during Neuronal Differentiation. Nucleic Acids Res 2012, 40, 8059–8071. [Google Scholar] [CrossRef] [PubMed]
- Fan, X.; Xiong, H.; Wei, J.; Gao, X.; Feng, Y.; Liu, X.; Zhang, G.; He, Q.-Y.; Xu, J.; Liu, L. Cytoplasmic hnRNPK Interacts with GSK3β and Is Essential for the Osteoclast Differentiation. Sci Rep 2015, 5, 17732. [Google Scholar] [CrossRef] [PubMed]
- Fukuda, T.; Naiki, T.; Saito, M.; Irie, K. hnRNP K Interacts with RNA Binding Motif Protein 42 and Functions in the Maintenance of Cellular ATP Level during Stress Conditions. Genes Cells 2009, 14, 113–128. [Google Scholar] [CrossRef]
- Habelhah, H.; Shah, K.; Huang, L.; Ostareck-Lederer, A.; Burlingame, A. L.; Shokat, K. M.; Hentze, M. W.; Ronai, Z. ERK Phosphorylation Drives Cytoplasmic Accumulation of hnRNP-K and Inhibition of mRNA Translation. Nat Cell Biol 2001, 3, 325–330. [Google Scholar] [CrossRef]
- Bomsztyk, K.; Denisenko, O.; Ostrowski, J. hnRNP K: One Protein Multiple Processes. Bioessays 2004, 26, 629–638. [Google Scholar] [CrossRef]
- Leffers, H.; Dejgaard, K.; Celis, J. E. Characterisation of Two Major Cellular Poly(rC)-Binding Human Proteins, Each Containing Three K-Homologous (KH) Domains. Eur J Biochem 1995, 230, 447–453. [Google Scholar] [CrossRef] [PubMed]
- Luo, Z.; Dong, X.; Li, Y.; Zhang, Q.; Kim, C.; Song, Y.; Kang, L.; Liu, Y.; Wu, K.; Wu, J. PolyC-Binding Protein 1 Interacts with 5′-Untranslated Region of Enterovirus 71 RNA in Membrane-Associated Complex to Facilitate Viral Replication. PLOS ONE 2014, 9, e87491. [Google Scholar] [CrossRef]
- Thyagarajan, A.; Szaro, B. G. Phylogenetically Conserved Binding of Specific K Homology Domain Proteins to the 3’-Untranslated Region of the Vertebrate Middle Neurofilament mRNA. J Biol Chem 2004, 279, 49680–49688. [Google Scholar] [CrossRef] [PubMed]
- Meng, Q.; Rayala, S. K.; Gururaj, A. E.; Talukder, A. H.; O’Malley, B. W.; Kumar, R. Signaling-Dependent and Coordinated Regulation of Transcription, Splicing, and Translation Resides in a Single Coregulator, PCBP1. Proc Natl Acad Sci U S A 2007, 104, 5866–5871. [Google Scholar] [CrossRef]
- Lian, W.-X.; Yin, R.-H.; Kong, X.-Z.; Zhang, T.; Huang, X.-H.; Zheng, W.-W.; Yang, Y.; Zhan, Y.-Q.; Xu, W.-X.; Yu, M.; Ge, C.-H.; Guo, J.-T.; Li, C.-Y.; Yang, X.-M. THAP11, a Novel Binding Protein of PCBP1, Negatively Regulates CD44 Alternative Splicing and Cell Invasion in a Human Hepatoma Cell Line. FEBS Lett 2012, 586, 1431–1438. [Google Scholar] [CrossRef] [PubMed]
- Zhang, T.; Huang, X.-H.; Dong, L.; Hu, D.; Ge, C.; Zhan, Y.-Q.; Xu, W.-X.; Yu, M.; Li, W.; Wang, X.; Tang, L.; Li, C.-Y.; Yang, X.-M. PCBP-1 Regulates Alternative Splicing of the CD44 Gene and Inhibits Invasion in Human Hepatoma Cell Line HepG2 Cells. Mol Cancer 2010, 9, 72. [Google Scholar] [CrossRef]
- Malik, A. K.; Flock, K. E.; Godavarthi, C. L.; Loh, H. H.; Ko, J. L. Molecular Basis Underlying the Poly C Binding Protein 1 as a Regulator of the Proximal Promoter of Mouse Mu-Opioid Receptor Gene. Brain Res 2006, 1112, 33–45. [Google Scholar] [CrossRef] [PubMed]
- Han, S. P.; Tang, Y. H.; Smith, R. Functional Diversity of the hnRNPs: Past, Present and Perspectives. Biochem J 2010, 430, 379–392. [Google Scholar] [CrossRef]
- Patton, J. G.; Mayer, S. A.; Tempst, P.; Nadal-Ginard, B. Characterization and Molecular Cloning of Polypyrimidine Tract-Binding Protein: A Component of a Complex Necessary for Pre-mRNA Splicing. Genes Dev 1991, 5, 1237–1251. [Google Scholar] [CrossRef]
- Hamid, F. M.; Makeyev, E. V. Regulation of mRNA Abundance by Polypyrimidine Tract-Binding Protein-Controlled Alternate 5’ Splice Site Choice. PLoS Genet 2014, 10, e1004771. [Google Scholar] [CrossRef] [PubMed]
- Rahman, M. A.; Masuda, A.; Ohe, K.; Ito, M.; Hutchinson, D. O.; Mayeda, A.; Engel, A. G.; Ohno, K. HnRNP L and hnRNP LL Antagonistically Modulate PTB-Mediated Splicing Suppression of CHRNA1 Pre-mRNA. Sci Rep 2013, 3, 2931. [Google Scholar] [CrossRef] [PubMed]
- Söderberg, M.; Raffalli-Mathieu, F.; Lang, M. A. Identification of a Regulatory Cis-Element within the 3’-Untranslated Region of the Murine Inducible Nitric Oxide Synthase (iNOS) mRNA; Interaction with Heterogeneous Nuclear Ribonucleoproteins I and L and Role in the iNOS Gene Expression. Mol Immunol 2007, 44, 434–442. [Google Scholar] [CrossRef] [PubMed]
- Majumder, M.; Yaman, I.; Gaccioli, F.; Zeenko, V. V.; Wang, C.; Caprara, M. G.; Venema, R. C.; Komar, A. A.; Snider, M. D.; Hatzoglou, M. The hnRNA-Binding Proteins hnRNP L and PTB Are Required for Efficient Translation of the Cat-1 Arginine/Lysine Transporter mRNA during Amino Acid Starvation. Mol Cell Biol 2009, 29, 2899–2912. [Google Scholar] [CrossRef]
- Jafarifar, F.; Yao, P.; Eswarappa, S. M.; Fox, P. L. Repression of VEGFA by CA-Rich Element-Binding microRNAs Is Modulated by hnRNP L. EMBO J 2011, 30, 1324–1334. [Google Scholar] [CrossRef] [PubMed]
- Xi, J.; Ye, F.; Wang, G.; Han, W.; Wei, Z.; Yin, B.; Yuan, J.; Qiang, B.; Peng, X. Polypyrimidine Tract-Binding Protein Regulates Enterovirus 71 Translation Through Interaction with the Internal Ribosomal Entry Site. Virol Sin 2019, 34, 66–77. [Google Scholar] [CrossRef] [PubMed]
- Min, H.; Turck, C. W.; Nikolic, J. M.; Black, D. L. A New Regulatory Protein, KSRP, Mediates Exon Inclusion through an Intronic Splicing Enhancer. Genes Dev 1997, 11, 1023–1036. [Google Scholar] [CrossRef] [PubMed]
- Davis-Smyth, T.; Duncan, R. C.; Zheng, T.; Michelotti, G.; Levens, D. The Far Upstream Element-Binding Proteins Comprise an Ancient Family of Single-Strand DNA-Binding Transactivators *. Journal of Biological Chemistry 1996, 271, 31679–31687. [Google Scholar] [CrossRef] [PubMed]
- Lellek, H.; Kirsten, R.; Diehl, I.; Apostel, F.; Buck, F.; Greeve, J. Purification and Molecular Cloning of a Novel Essential Component of the Apolipoprotein B mRNA Editing Enzyme-Complex. J Biol Chem 2000, 275, 19848–19856. [Google Scholar] [CrossRef]
- Gherzi, R.; Chen, C.-Y.; Trabucchi, M.; Ramos, A.; Briata, P. The Role of KSRP in mRNA Decay and microRNA Precursor Maturation. Wiley Interdiscip Rev RNA 2010, 1, 230–239. [Google Scholar] [CrossRef]
- Trabucchi, M.; Briata, P.; Filipowicz, W.; Rosenfeld, M. G.; Ramos, A.; Gherzi, R. How to Control miRNA Maturation? RNA Biol 2009, 6, 536–540. [Google Scholar] [CrossRef] [PubMed]
- Briata, P.; Chen, C.-Y.; Ramos, A.; Gherzi, R. Functional and Molecular Insights into KSRP Function in mRNA Decay. Biochim Biophys Acta 2013, 1829, 689–694. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.-L.; Kung, Y.-A.; Weng, K.-F.; Lin, J.-Y.; Horng, J.-T.; Shih, S.-R. Enterovirus 71 Infection Cleaves a Negative Regulator for Viral Internal Ribosomal Entry Site-Driven Translation. J Virol 2013, 87, 3828–3838. [Google Scholar] [CrossRef]
- Zhang, J.; Chen, Q. M. Far Upstream Element Binding Protein 1: A Commander of Transcription, Translation and Beyond. Oncogene 2013, 32, 2907–2916. [Google Scholar] [CrossRef]
- Duncan, R.; Bazar, L.; Michelotti, G.; Tomonaga, T.; Krutzsch, H.; Avigan, M.; Levens, D. A Sequence-Specific, Single-Strand Binding Protein Activates the Far Upstream Element of c-Myc and Defines a New DNA-Binding Motif. Genes Dev 1994, 8, 465–480. [Google Scholar] [CrossRef] [PubMed]
- Olanich, M. E.; Moss, B. L.; Piwnica-Worms, D.; Townsend, R. R.; Weber, J. D. Identification of FUSE-Binding Protein 1 as a Regulatory mRNA-Binding Protein That Represses Nucleophosmin Translation. Oncogene 2011, 30, 77–86. [Google Scholar] [CrossRef]
- Zhang, Z.; Harris, D.; Pandey, V. N. The FUSE Binding Protein Is a Cellular Factor Required for Efficient Replication of Hepatitis C Virus. J Virol 2008, 82, 5761–5773. [Google Scholar] [CrossRef]
- Chien, H.-L.; Liao, C.-L.; Lin, Y.-L. FUSE Binding Protein 1 Interacts with Untranslated Regions of Japanese Encephalitis Virus RNA and Negatively Regulates Viral Replication ▿. J Virol 2011, 85, 4698–4706. [Google Scholar] [CrossRef]
- Huang, P.-N.; Lin, J.-Y.; Locker, N.; Kung, Y.-A.; Hung, C.-T.; Lin, J.-Y.; Huang, H.-I.; Li, M.-L.; Shih, S.-R. Far Upstream Element Binding Protein 1 Binds the Internal Ribosomal Entry Site of Enterovirus 71 and Enhances Viral Translation and Viral Growth. Nucleic Acids Res 2011, 39, 9633–9648. [Google Scholar] [CrossRef]
- Hung, C.-T.; Kung, Y.-A.; Li, M.-L.; Brewer, G.; Lee, K.-M.; Liu, S.-T.; Shih, S.-R. Additive Promotion of Viral Internal Ribosome Entry Site-Mediated Translation by Far Upstream Element-Binding Protein 1 and an Enterovirus 71-Induced Cleavage Product. PLoS Pathog 2016, 12, e1005959. [Google Scholar] [CrossRef]
- Höck, J.; Meister, G. The Argonaute Protein Family. Genome Biol 2008, 9, 210. [Google Scholar] [CrossRef] [PubMed]
- Schirle, N. T.; MacRae, I. J. The Crystal Structure of Human Argonaute2. Science 2012, 336, 1037–1040. [Google Scholar] [CrossRef] [PubMed]
- Pantazopoulou, V. I.; Delis, A. D.; Georgiou, S.; Pagakis, S. N.; Filippa, V.; Dragona, E.; Kloukina, I.; Chatzitheodoridis, E.; Trebicka, J.; Velentzas, A. D.; Thiele, M.; Gagos, S.; Thanos, D.; Tseleni-Balafouta, S.; Stravopodis, D. J.; Anastasiadou, E. AGO2 Localizes to Cytokinetic Protrusions in a P38-Dependent Manner and Is Needed for Accurate Cell Division. Commun Biol 2021, 4, 726. [Google Scholar] [CrossRef] [PubMed]
- Vasudevan, S.; Steitz, J. A. AU-Rich-Element-Mediated Upregulation of Translation by FXR1 and Argonaute 2. Cell 2007, 128, 1105–1118. [Google Scholar] [CrossRef] [PubMed]
- Brennan, C. M.; Steitz*, J. A. HuR and mRNA Stability. CMLS, Cell. Mol. Life Sci. 2001, 58, 266–277. [Google Scholar] [CrossRef] [PubMed]
- Lebedeva, S.; Jens, M.; Theil, K.; Schwanhäusser, B.; Selbach, M.; Landthaler, M.; Rajewsky, N. Transcriptome-Wide Analysis of Regulatory Interactions of the RNA-Binding Protein HuR. Molecular Cell 2011, 43, 340–352. [Google Scholar] [CrossRef]
- Fan, X. C.; Steitz, J. A. Overexpression of HuR, a Nuclear-Cytoplasmic Shuttling Protein, Increases the in Vivo Stability of ARE-Containing mRNAs. EMBO J 1998, 17, 3448–3460. [Google Scholar] [CrossRef] [PubMed]
- Uren, P. J.; Burns, S. C.; Ruan, J.; Singh, K. K.; Smith, A. D.; Penalva, L. O. F. Genomic Analyses of the RNA-Binding Protein Hu Antigen R (HuR) Identify a Complex Network of Target Genes and Novel Characteristics of Its Binding Sites. J Biol Chem 2011, 286, 37063–37066. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Zhang, Y.; Zhang, J.; Ma, J.; Xu, X.; Wang, Y.; Zhou, Z.; Jiang, D.; Shen, S.; Ding, Y.; Zhou, Y.; Zhuang, R. Silencing of HuR Inhibits Osteosarcoma Cell Epithelial-Mesenchymal Transition via AGO2 in Association With Long Non-Coding RNA XIST. Frontiers in Oncology 2021, 11. [Google Scholar] [CrossRef]
- Zhu, Y.; Wang, B.; Huang, H.; Zhao, Z. Enterovirus 71 Induces Anti-Viral Stress Granule-like Structures in RD Cells. Biochem Biophys Res Commun 2016, 476, 212–217. [Google Scholar] [CrossRef]
- Wang, H.; Chang, L.; Wang, X.; Su, A.; Feng, C.; Fu, Y.; Chen, D.; Zheng, N.; Wu, Z. MOV10 Interacts with Enterovirus 71 Genomic 5’UTR and Modulates Viral Replication. Biochem Biophys Res Commun 2016, 479, 571–577. [Google Scholar] [CrossRef] [PubMed]
- Gregersen, L. H.; Schueler, M.; Munschauer, M.; Mastrobuoni, G.; Chen, W.; Kempa, S.; Dieterich, C.; Landthaler, M. MOV10 Is a 5’ to 3’ RNA Helicase Contributing to UPF1 mRNA Target Degradation by Translocation along 3’ UTRs. Mol Cell 2014, 54, 573–585. [Google Scholar] [CrossRef] [PubMed]
- Meister, G.; Landthaler, M.; Peters, L.; Chen, P. Y.; Urlaub, H.; Lührmann, R.; Tuschl, T. Identification of Novel Argonaute-Associated Proteins. Curr Biol 2005, 15, 2149–2155. [Google Scholar] [CrossRef] [PubMed]
- Abudu, A.; Wang, X.; Dang, Y.; Zhou, T.; Xiang, S.-H.; Zheng, Y.-H. Identification of Molecular Determinants from Moloney Leukemia Virus 10 Homolog (MOV10) Protein for Virion Packaging and Anti-HIV-1 Activity*. Journal of Biological Chemistry 2012, 287, 1220–1228. [Google Scholar] [CrossRef] [PubMed]
- Rahman, S.; Islam, R. Mammalian Sirt1: Insights on Its Biological Functions. Cell Commun Signal 2011, 9, 11. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, H.; Schoonjans, K.; Auwerx, J. Sirtuin Functions in Health and Disease. Molecular Endocrinology 2007, 21, 1745–1755. [Google Scholar] [CrossRef] [PubMed]
- Kwon, H.-S.; Ott, M. The Ups and Downs of SIRT1. Trends Biochem Sci 2008, 33, 517–525. [Google Scholar] [CrossRef] [PubMed]
- Pagans, S.; Pedal, A.; North, B. J.; Kaehlcke, K.; Marshall, B. L.; Dorr, A.; Hetzer-Egger, C.; Henklein, P.; Frye, R.; McBurney, M. W.; Hruby, H.; Jung, M.; Verdin, E.; Ott, M. SIRT1 Regulates HIV Transcription via Tat Deacetylation. PLoS Biol 2005, 3, e41. [Google Scholar] [CrossRef] [PubMed]
- Ren, J.-H.; Tao, Y.; Zhang, Z.-Z.; Chen, W.-X.; Cai, X.-F.; Chen, K.; Ko, B. C. B.; Song, C.-L.; Ran, L.-K.; Li, W.-Y.; Huang, A.-L.; Chen, J. Sirtuin 1 Regulates Hepatitis B Virus Transcription and Replication by Targeting Transcription Factor AP-1. J Virol 2014, 88, 2442–2451. [Google Scholar] [CrossRef]
- Han, Y.; Wang, L.; Cui, J.; Song, Y.; Luo, Z.; Chen, J.; Xiong, Y.; Zhang, Q.; Liu, F.; Ho, W.; Liu, Y.; Wu, K.; Wu, J. SIRT1 Inhibits EV71 Genome Replication and RNA Translation by Interfering with the Viral Polymerase and 5′UTR RNA. J Cell Sci 2016, 129, 4534–4547. [Google Scholar] [CrossRef]
- Matter, N.; Herrlich, P.; König, H. Signal-Dependent Regulation of Splicing via Phosphorylation of Sam68. Nature 2002, 420, 691–695. [Google Scholar] [CrossRef]
- Paronetto, M. P.; Cappellari, M.; Busà, R.; Pedrotti, S.; Vitali, R.; Comstock, C.; Hyslop, T.; Knudsen, K. E.; Sette, C. Alternative Splicing of the Cyclin D1 Proto-Oncogene Is Regulated by the RNA-Binding Protein Sam68. Cancer Res 2010, 70, 229–239. [Google Scholar] [CrossRef] [PubMed]
- Sánchez-Jiménez, F.; Sánchez-Margalet, V. Role of Sam68 in Post-Transcriptional Gene Regulation. Int J Mol Sci 2013, 14, 23402–23419. [Google Scholar] [CrossRef]
- Fumagalli, S.; Totty, N. F.; Hsuan, J. J.; Courtneidge, S. A. A Target for Src in Mitosis. Nature 1994, 368, 871–874. [Google Scholar] [CrossRef] [PubMed]
- Macias, M. J.; Wiesner, S.; Sudol, M. WW and SH3 Domains, Two Different Scaffolds to Recognize Proline-Rich Ligands. FEBS Lett 2002, 513, 30–37. [Google Scholar] [CrossRef] [PubMed]
- Galarneau, A.; Richard, S. The STAR RNA Binding Proteins GLD-1, QKI, SAM68 and SLM-2 Bind Bipartite RNA Motifs. BMC Molecular Biology 2009, 10, 47. [Google Scholar] [CrossRef] [PubMed]
- Lin, Q.; Taylor, S. J.; Shalloway, D. Specificity and Determinants of Sam68 RNA Binding: IMPLICATIONS FOR THE BIOLOGICAL FUNCTION OF K HOMOLOGY DOMAINS *. Journal of Biological Chemistry 1997, 272, 27274–27280. [Google Scholar] [CrossRef] [PubMed]
- Lawrence, P.; Schafer, E. A.; Rieder, E. The Nuclear Protein Sam68 Is Cleaved by the FMDV 3C Protease Redistributing Sam68 to the Cytoplasm during FMDV Infection of Host Cells. Virology 2012, 425, 40–52. [Google Scholar] [CrossRef] [PubMed]
- McBride, A. E.; Schlegel, A.; Kirkegaard, K. Human Protein Sam68 Relocalization and Interaction with Poliovirus RNA Polymerase in Infected Cells. Proc Natl Acad Sci U S A 1996, 93, 2296–2301. [Google Scholar] [CrossRef]
- Zhang, H.; Song, L.; Cong, H.; Tien, P. Nuclear Protein Sam68 Interacts with the Enterovirus 71 Internal Ribosome Entry Site and Positively Regulates Viral Protein Translation. J Virol 2015, 89, 10031–10043. [Google Scholar] [CrossRef]
- Xu, P.; Tong, W.; Chen, Y.-M. FUSE Binding Protein FUBP3 Is a Potent Regulator in Japanese Encephalitis Virus Infection. Virology Journal 2021, 18, 224. [Google Scholar] [CrossRef] [PubMed]
- Sharma, M.; Anandram, S.; Ross, C.; Srivastava, S. FUBP3 Regulates Chronic Myeloid Leukaemia Progression through PRC2 Complex Regulated PAK1-ERK Signalling. Journal of Cellular and Molecular Medicine 2023, 27, 15–29. [Google Scholar] [CrossRef] [PubMed]
- Tsai, F.-J.; Lin, C.-W.; Lai, C.-C.; Lan, Y.-C.; Lai, C.-H.; Hung, C.-H.; Hsueh, K.-C.; Lin, T.-H.; Chang, H. C.; Wan, L.; Sheu, J. J.-C.; Lin, Y.-J. Kaempferol Inhibits Enterovirus 71 Replication and Internal Ribosome Entry Site (IRES) Activity through FUBP and HNRP Proteins. Food Chemistry 2011, 128, 312–322. [Google Scholar] [CrossRef] [PubMed]
- Huang, H.-I.; Chang, Y.-Y.; Lin, J.-Y.; Kuo, R.-L.; Liu, H.-P.; Shih, S.-R.; Wu, C.-C. Interactome Analysis of the EV71 5′ Untranslated Region in Differentiated Neuronal Cells SH-SY5Y and Regulatory Role of FBP3 in Viral Replication. PROTEOMICS 2016, 16, 2351–2362. [Google Scholar] [CrossRef] [PubMed]
- Brush, M. H.; Weiser, D. C.; Shenolikar, S. Growth Arrest and DNA Damage-Inducible Protein GADD34 Targets Protein Phosphatase 1α to the Endoplasmic Reticulum and Promotes Dephosphorylation of the α Subunit of Eukaryotic Translation Initiation Factor 2. Mol Cell Biol 2003, 23, 1292–1303. [Google Scholar] [CrossRef]
- Zhu, T.; Jiang, X.; Xin, H.; Zheng, X.; Xue, X.; Chen, J.-L.; Qi, B. GADD34-Mediated Dephosphorylation of eIF2α Facilitates Pseudorabies Virus Replication by Maintaining de Novo Protein Synthesis. Veterinary Research 2021, 52, 148. [Google Scholar] [CrossRef] [PubMed]
- Pakos-Zebrucka, K.; Koryga, I.; Mnich, K.; Ljujic, M.; Samali, A.; Gorman, A. M. The Integrated Stress Response. EMBO Rep 2016, 17, 1374–1395. [Google Scholar] [CrossRef] [PubMed]
- Dalet, A.; Argüello, R. J.; Combes, A.; Spinelli, L.; Jaeger, S.; Fallet, M.; Vu Manh, T.-P.; Mendes, A.; Perego, J.; Reverendo, M.; Camosseto, V.; Dalod, M.; Weil, T.; Santos, M. A.; Gatti, E.; Pierre, P. Protein Synthesis Inhibition and GADD34 Control IFN-β Heterogeneous Expression in Response to dsRNA. EMBO J 2017, 36, 761–782. [Google Scholar] [CrossRef]
- Clavarino, G.; Cláudio, N.; Couderc, T.; Dalet, A.; Judith, D.; Camosseto, V.; Schmidt, E. K.; Wenger, T.; Lecuit, M.; Gatti, E.; Pierre, P. Induction of GADD34 Is Necessary for dsRNA-Dependent Interferon-β Production and Participates in the Control of Chikungunya Virus Infection. PLoS Pathog 2012, 8, e1002708. [Google Scholar] [CrossRef]
- Minami, K.; Tambe, Y.; Watanabe, R.; Isono, T.; Haneda, M.; Isobe, K.-I.; Kobayashi, T.; Hino, O.; Okabe, H.; Chano, T.; Inoue, H. Suppression of Viral Replication by Stress-Inducible GADD34 Protein via the Mammalian Serine/Threonine Protein Kinase mTOR Pathway. J Virol 2007, 81, 11106–11115. [Google Scholar] [CrossRef]
- Wang, X.; Liao, Y.; Yap, P. L.; Png, K. J.; Tam, J. P.; Liu, D. X. Inhibition of Protein Kinase R Activation and Upregulation of GADD34 Expression Play a Synergistic Role in Facilitating Coronavirus Replication by Maintaining De Novo Protein Synthesis in Virus-Infected Cells. J Virol 2009, 83, 12462–12472. [Google Scholar] [CrossRef]
- Ishaq, M.; Marshall, H.; Natarajan, V. GADD34 Attenuates HIV-1 Replication by Viral 5’-UTR TAR RNA-Mediated Translational Inhibition. Virology 2020, 540, 119–131. [Google Scholar] [CrossRef] [PubMed]
- Brush, M. H.; Shenolikar, S. Control of Cellular GADD34 Levels by the 26S Proteasome. Mol Cell Biol 2008, 28, 6989–7000. [Google Scholar] [CrossRef] [PubMed]
- Harris, K. S.; Xiang, W.; Alexander, L.; Lane, W. S.; Paul, A. V.; Wimmer, E. Interaction of Poliovirus Polypeptide 3CDpro with the 5’ and 3’ Termini of the Poliovirus Genome. Identification of Viral and Cellular Cofactors Needed for Efficient Binding. J Biol Chem 1994, 269, 27004–27014. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Li, W.; Zhang, S.; Qiu, M.; Li, Z.; Lin, Y.; Tan, J.; Qiao, W. Enterovirus 71 Activates GADD34 via Precursor 3CD to Promote IRES-Mediated Viral Translation. Microbiol Spectr 10 (1), e01388-21. [CrossRef]
- Venkataramanan, S.; Gadek, M.; Calviello, L.; Wilkins, K.; Floor, S. N. DDX3X and DDX3Y Are Redundant in Protein Synthesis. RNA 2021, 27, 1577–1588. [Google Scholar] [CrossRef] [PubMed]
- Schröder, M. Human DEAD-Box Protein 3 Has Multiple Functions in Gene Regulation and Cell Cycle Control and Is a Prime Target for Viral Manipulation. Biochem Pharmacol 2010, 79, 297–306. [Google Scholar] [CrossRef] [PubMed]
- Soto-Rifo, R.; Rubilar, P. S.; Limousin, T.; de Breyne, S.; Décimo, D.; Ohlmann, T. DEAD-Box Protein DDX3 Associates with eIF4F to Promote Translation of Selected mRNAs. EMBO J 2012, 31, 3745–3756. [Google Scholar] [CrossRef] [PubMed]
- Su, Y.-S.; Tsai, A.-H.; Ho, Y.-F.; Huang, S.-Y.; Liu, Y.-C.; Hwang, L.-H. Stimulation of the Internal Ribosome Entry Site (IRES)-Dependent Translation of Enterovirus 71 by DDX3X RNA Helicase and Viral 2A and 3C Proteases. Frontiers in Microbiology 2018, 9. [Google Scholar] [CrossRef] [PubMed]
- Gao, J.; Choudhry, H.; Cao, W. Apolipoprotein B mRNA Editing Enzyme Catalytic Polypeptide-like Family Genes Activation and Regulation during Tumorigenesis. Cancer Sci 2018, 109, 2375–2382. [Google Scholar] [CrossRef]
- Wang, H.; Zhong, M.; Li, Y.; Li, K.; Wu, S.; Guo, T.; Cen, S.; Jiang, J.; Li, Z.; Li, Y. APOBEC3G Is a Restriction Factor of EV71 and Mediator of IMB-Z Antiviral Activity. Antiviral Research 2019, 165, 23–33. [Google Scholar] [CrossRef]
- Fehrholz, M.; Kendl, S.; Prifert, C.; Weissbrich, B.; Lemon, K.; Rennick, L.; Duprex, P. W.; Rima, B. K.; Koning, F. A.; Holmes, R. K.; Malim, M. H.; Schneider-Schaulies, J. The Innate Antiviral Factor APOBEC3G Targets Replication of Measles, Mumps and Respiratory Syncytial Viruses. Journal of General Virology 2012, 93, 565–576. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, D. H.; Hu, J. Reverse Transcriptase- and RNA Packaging Signal-Dependent Incorporation of APOBEC3G into Hepatitis B Virus Nucleocapsids. Journal of Virology 2008, 82, 6852–6861. [Google Scholar] [CrossRef] [PubMed]
- Okada, A.; Iwatani, Y. APOBEC3G-Mediated G-to-A Hypermutation of the HIV-1 Genome: The Missing Link in Antiviral Molecular Mechanisms. Front Microbiol 2016, 7, 2027. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Ning, S.; Su, X.; Liu, X.; Wang, H.; Liu, Y.; Zheng, W.; Zheng, B.; Yu, X.-F.; Zhang, W. Enterovirus 71 Antagonizes the Inhibition of the Host Intrinsic Antiviral Factor A3G. Nucleic Acids Research 2018, 46, 11514–11527. [Google Scholar] [CrossRef] [PubMed]
- Wickham, L.; Duchaîne, T.; Luo, M.; Nabi, I. R.; DesGroseillers, L. Mammalian Staufen Is a Double-Stranded-RNA- and Tubulin-Binding Protein Which Localizes to the Rough Endoplasmic Reticulum. Mol Cell Biol 1999, 19, 2220–2230. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y. K.; Furic, L.; Desgroseillers, L.; Maquat, L. E. Mammalian Staufen1 Recruits Upf1 to Specific mRNA 3’UTRs so as to Elicit mRNA Decay. Cell 2005, 120, 195–208. [Google Scholar] [CrossRef] [PubMed]
- Abdullah, S. W.; Wu, J.; Wang, X.; Guo, H.; Sun, S. Advances and Breakthroughs in IRES-Directed Translation and Replication of Picornaviruses. mBio 2023, 14, e0035823. [Google Scholar] [CrossRef] [PubMed]
- Furic, L.; Maher-Laporte, M.; DesGroseillers, L. A Genome-Wide Approach Identifies Distinct but Overlapping Subsets of Cellular mRNAs Associated with Staufen1- and Staufen2-Containing Ribonucleoprotein Complexes. RNA 2008, 14, 324–335. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.-M.; Ou, B.-T.; Chen, C.-Y.; Chan, H.-H.; Chen, C.-J.; Wang, R. Y. Staufen1 Protein Participates Positively in the Viral RNA Replication of Enterovirus 71. Viruses 2019, 11, 142. [Google Scholar] [CrossRef]
- Blackham, S. L.; McGarvey, M. J. A Host Cell RNA-Binding Protein, Staufen1, Has a Role in Hepatitis C Virus Replication before Virus Assembly. J Gen Virol 2013, 94 Pt 11, 2429–2436. [Google Scholar] [CrossRef]
- Abrahamyan, L. G.; Chatel-Chaix, L.; Ajamian, L.; Milev, M. P.; Monette, A.; Clément, J.-F.; Song, R.; Lehmann, M.; DesGroseillers, L.; Laughrea, M.; Boccaccio, G.; Mouland, A. J. Novel Staufen1 Ribonucleoproteins Prevent Formation of Stress Granules but Favour Encapsidation of HIV-1 Genomic RNA. J Cell Sci 2010, 123 Pt 3, 369–383. [Google Scholar] [CrossRef] [PubMed]
- de Lucas, S.; Peredo, J.; Marión, R. M.; Sánchez, C.; Ortín, J. Human Staufen1 Protein Interacts with Influenza Virus Ribonucleoproteins and Is Required for Efficient Virus Multiplication. J Virol 2010, 84, 7603–7612. [Google Scholar] [CrossRef] [PubMed]
- Mauger, D. M.; Lin, C.; Garcia-Blanco, M. A. hnRNP H and hnRNP F Complex with Fox2 To Silence Fibroblast Growth Factor Receptor 2 Exon IIIc. Molecular and Cellular Biology 2008, 28, 5403–5419. [Google Scholar] [CrossRef] [PubMed]
- Xu, C.; Xie, N.; Su, Y.; Sun, Z.; Liang, Y.; Zhang, N.; Liu, D.; Jia, S.; Xing, X.; Han, L.; Li, G.; Tong, T.; Chen, J. HnRNP F/H Associate with hTERC and Telomerase Holoenzyme to Modulate Telomerase Function and Promote Cell Proliferation. Cell Death Differ 2020, 27, 1998–2013. [Google Scholar] [CrossRef] [PubMed]
- Thiel, G.; Cibelli, G. Regulation of Life and Death by the Zinc Finger Transcription Factor Egr-1. Journal of Cellular Physiology 2002, 193, 287–292. [Google Scholar] [CrossRef] [PubMed]
- Ho, B.-C.; Yu, S.-L.; Chen, J. J. W.; Chang, S.-Y.; Yan, B.-S.; Hong, Q.-S.; Singh, S.; Kao, C.-L.; Chen, H.-Y.; Su, K.-Y.; Li, K.-C.; Cheng, C.-L.; Cheng, H.-W.; Lee, J.-Y.; Lee, C.-N.; Yang, P.-C. Enterovirus-Induced miR-141 Contributes to Shutoff of Host Protein Translation by Targeting the Translation Initiation Factor eIF4E. Cell Host & Microbe 2011, 9, 58–69. [Google Scholar] [CrossRef]
- Song, Y.; Cheng, X.; Yang, X.; Zhao, R.; Wang, P.; Han, Y.; Luo, Z.; Cao, Y.; Zhu, C.; Xiong, Y.; Liu, Y.; Wu, K.; Wu, J. Early Growth Response-1 Facilitates Enterovirus 71 Replication by Direct Binding to the Viral Genome RNA. The International Journal of Biochemistry & Cell Biology 2015, 62, 36–46. [Google Scholar] [CrossRef]
- Kedersha, N. L.; Gupta, M.; Li, W.; Miller, I.; Anderson, P. RNA-Binding Proteins TIA-1 and TIAR Link the Phosphorylation of eIF-2 Alpha to the Assembly of Mammalian Stress Granules. J Cell Biol 1999, 147, 1431–1442. [Google Scholar] [CrossRef]
- Gottschald, O. R.; Malec, V.; Krasteva, G.; Hasan, D.; Kamlah, F.; Herold, S.; Rose, F.; Seeger, W.; Hänze, J. TIAR and TIA-1 mRNA-Binding Proteins Co-Aggregate under Conditions of Rapid Oxygen Decline and Extreme Hypoxia and Suppress the HIF-1α Pathway. J Mol Cell Biol 2010, 2, 345–356. [Google Scholar] [CrossRef]
- White, J. P.; Lloyd, R. E. Poliovirus Unlinks TIA1 Aggregation and mRNA Stress Granule Formation. Journal of Virology 2011, 85, 12442–12454. [Google Scholar] [CrossRef]
- Wu, S.; Wang, Y.; Lin, L.; Si, X.; Wang, T.; Zhong, X.; Tong, L.; Luan, Y.; Chen, Y.; Li, X.; Zhang, F.; Zhao, W.; Zhong, Z. Protease 2A Induces Stress Granule Formation during Coxsackievirus B3 and Enterovirus 71 Infections. Virol J 2014, 11, 192. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Wang, H.; Li, Y.; Jin, Y.; Chu, Y.; Su, A.; Wu, Z. TIA-1 and TIAR Interact with 5′-UTR of Enterovirus 71 Genome and Facilitate Viral Replication. Biochemical and Biophysical Research Communications 2015, 466, 254–259. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Reprogramming of the cellular environment upon EV-A71 infection. Following viral infection, several host RNA binding proteins, collectively referred to as ITAFs, change their sub-cellular localization. (a) These cellular ITAFs are primarily localized in the nucleus or cytoplasm while some of them can be shuttled between nucleus and cytoplasm based on their cellular functions. Regardless, upon EV-A71 infection, all ITAFs will be relocalized into the cytoplasm to regulate IRES-mediated translation. (b) The 5´-UTR of EV-A71 genome consists of the cloverleaf structure (SLI) and the IRES (SLII-VI). Cellular ITAFs bind to specific region(s) within IRES to either positively or negatively regulate IRES-mediated translation. Positive and negative ITAFs are indicated in green and red, respectively. The ITAFs, of which the effect on translation is not discovered yet, are indicated in grey. Some ITAFs can also bind the SLI region to regulate viral genome transcription.
Figure 1.
Reprogramming of the cellular environment upon EV-A71 infection. Following viral infection, several host RNA binding proteins, collectively referred to as ITAFs, change their sub-cellular localization. (a) These cellular ITAFs are primarily localized in the nucleus or cytoplasm while some of them can be shuttled between nucleus and cytoplasm based on their cellular functions. Regardless, upon EV-A71 infection, all ITAFs will be relocalized into the cytoplasm to regulate IRES-mediated translation. (b) The 5´-UTR of EV-A71 genome consists of the cloverleaf structure (SLI) and the IRES (SLII-VI). Cellular ITAFs bind to specific region(s) within IRES to either positively or negatively regulate IRES-mediated translation. Positive and negative ITAFs are indicated in green and red, respectively. The ITAFs, of which the effect on translation is not discovered yet, are indicated in grey. Some ITAFs can also bind the SLI region to regulate viral genome transcription.
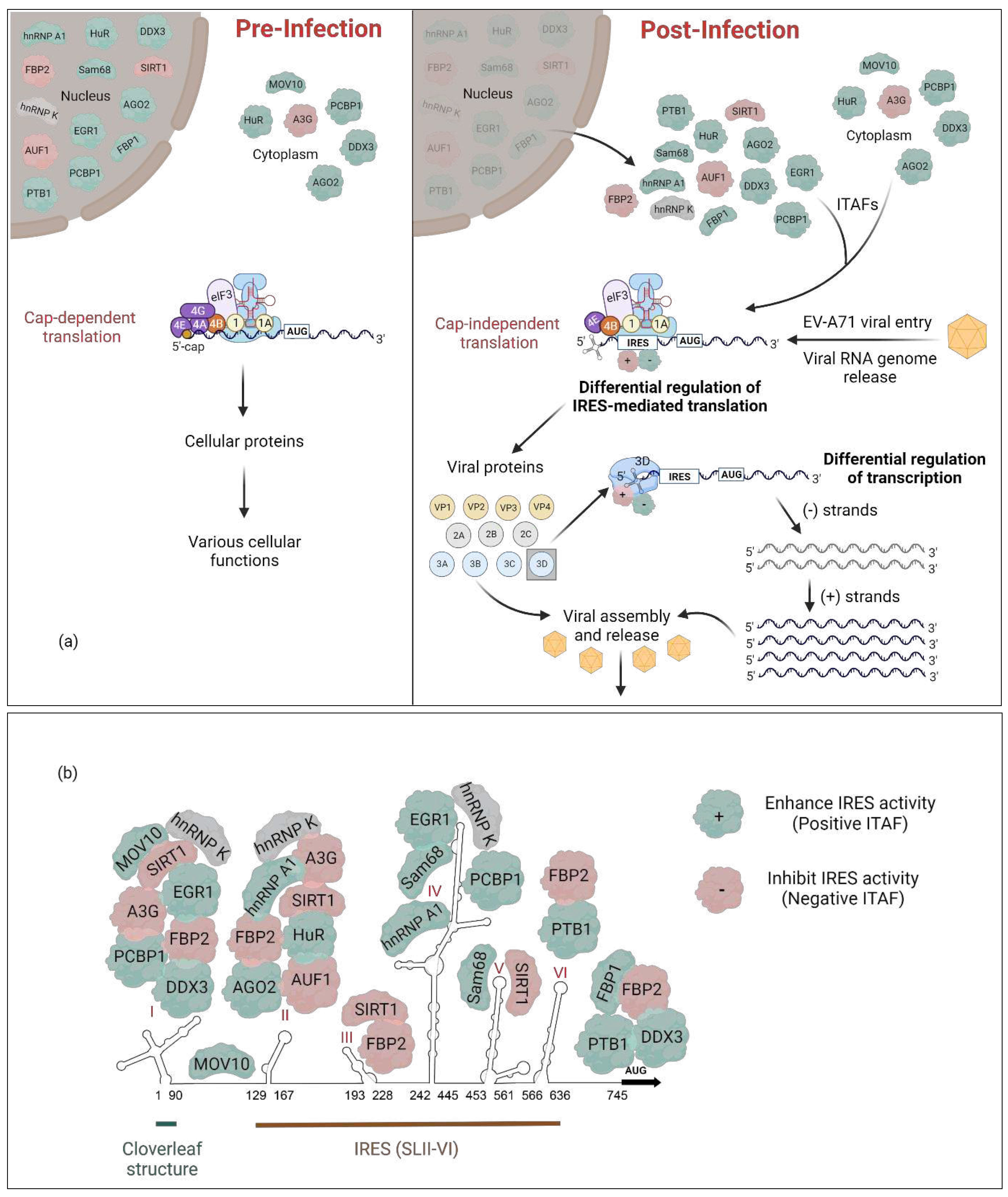
Figure 2.
The EV-A71 5´-UTR contains phylogenetically conserved stem-loops (SLs) that adopt well-defined 3D structures. Depicted is a predicted secondary structure of the 5´-UTR of EV-A71, and 3D models of each individual stem-loop. Nucleotides are color coded where Adenine, Uracil, Guanine, and Cytosine are colored in red, green, blue, and yellow, respectively. Colored RNA regions are predicted to be single stranded and most likely to interact with listed ITAFs. 3D structures of SL I and II are published PDB structures while SLs III-VI were produced using the FAFFAR module of the ROSETTA software suit. The lowest energy structure for each stem-loop been selected from a pool of 10,000 structures. The structures of SLI (8DP3) and SLII (5V17) were obtained from the PDB.
Figure 2.
The EV-A71 5´-UTR contains phylogenetically conserved stem-loops (SLs) that adopt well-defined 3D structures. Depicted is a predicted secondary structure of the 5´-UTR of EV-A71, and 3D models of each individual stem-loop. Nucleotides are color coded where Adenine, Uracil, Guanine, and Cytosine are colored in red, green, blue, and yellow, respectively. Colored RNA regions are predicted to be single stranded and most likely to interact with listed ITAFs. 3D structures of SL I and II are published PDB structures while SLs III-VI were produced using the FAFFAR module of the ROSETTA software suit. The lowest energy structure for each stem-loop been selected from a pool of 10,000 structures. The structures of SLI (8DP3) and SLII (5V17) were obtained from the PDB.
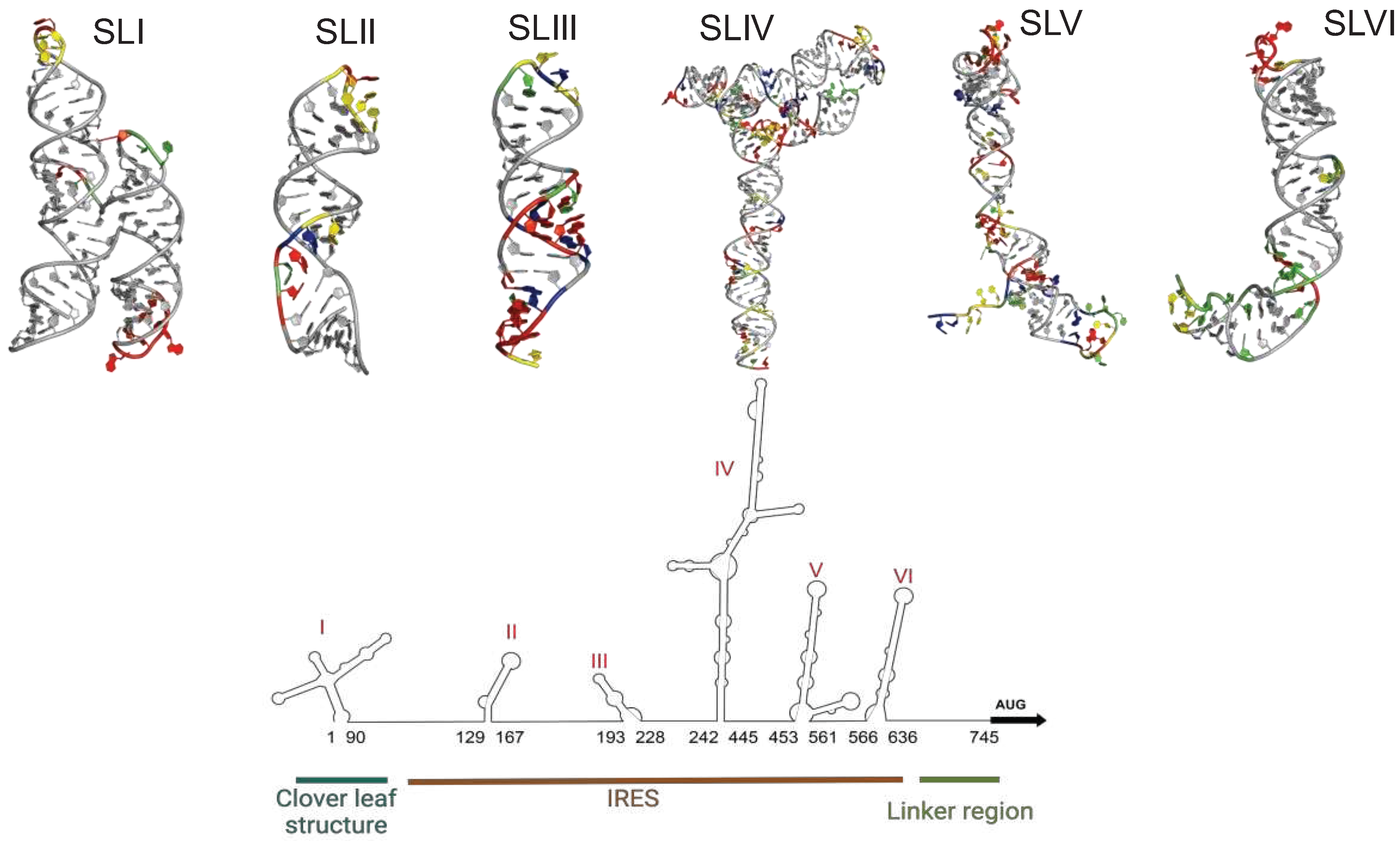
Figure 3.
Cellular ITAFs bind with specificity to short and degenerate sequence motifs. Depicted are published PDB structures of cellular ITAFs bound to RNA. These structures have been experimentally determined by both X-ray and nuclear magnetic resonance (NMR) spectroscopy. RNA has been color coded where Adenine, Uracil, Guanine, and Cytosine are colored in red, green, blue, and yellow, respectively. ITAFs that bind to double stranded RNA have the RNA depicted as a cartoon structure. The identity and PDB ids of the depicted ITAFs are as follows: (a) Argonaute 2 (5KI6), (b) hnRNP F (2KFY), (c) Far-Upstream Binding Protein 2 (4B8T), (d) hnRNP A1 (4YOE), (e) Polypyrimidine tract-binding protein 1 (2N3O), (f) Human Antigen R (6G2K), (g) DEAD-Box Protein 3 (6O5F), (h) Staufen homolog 1 (6HTU).
Figure 3.
Cellular ITAFs bind with specificity to short and degenerate sequence motifs. Depicted are published PDB structures of cellular ITAFs bound to RNA. These structures have been experimentally determined by both X-ray and nuclear magnetic resonance (NMR) spectroscopy. RNA has been color coded where Adenine, Uracil, Guanine, and Cytosine are colored in red, green, blue, and yellow, respectively. ITAFs that bind to double stranded RNA have the RNA depicted as a cartoon structure. The identity and PDB ids of the depicted ITAFs are as follows: (a) Argonaute 2 (5KI6), (b) hnRNP F (2KFY), (c) Far-Upstream Binding Protein 2 (4B8T), (d) hnRNP A1 (4YOE), (e) Polypyrimidine tract-binding protein 1 (2N3O), (f) Human Antigen R (6G2K), (g) DEAD-Box Protein 3 (6O5F), (h) Staufen homolog 1 (6HTU).
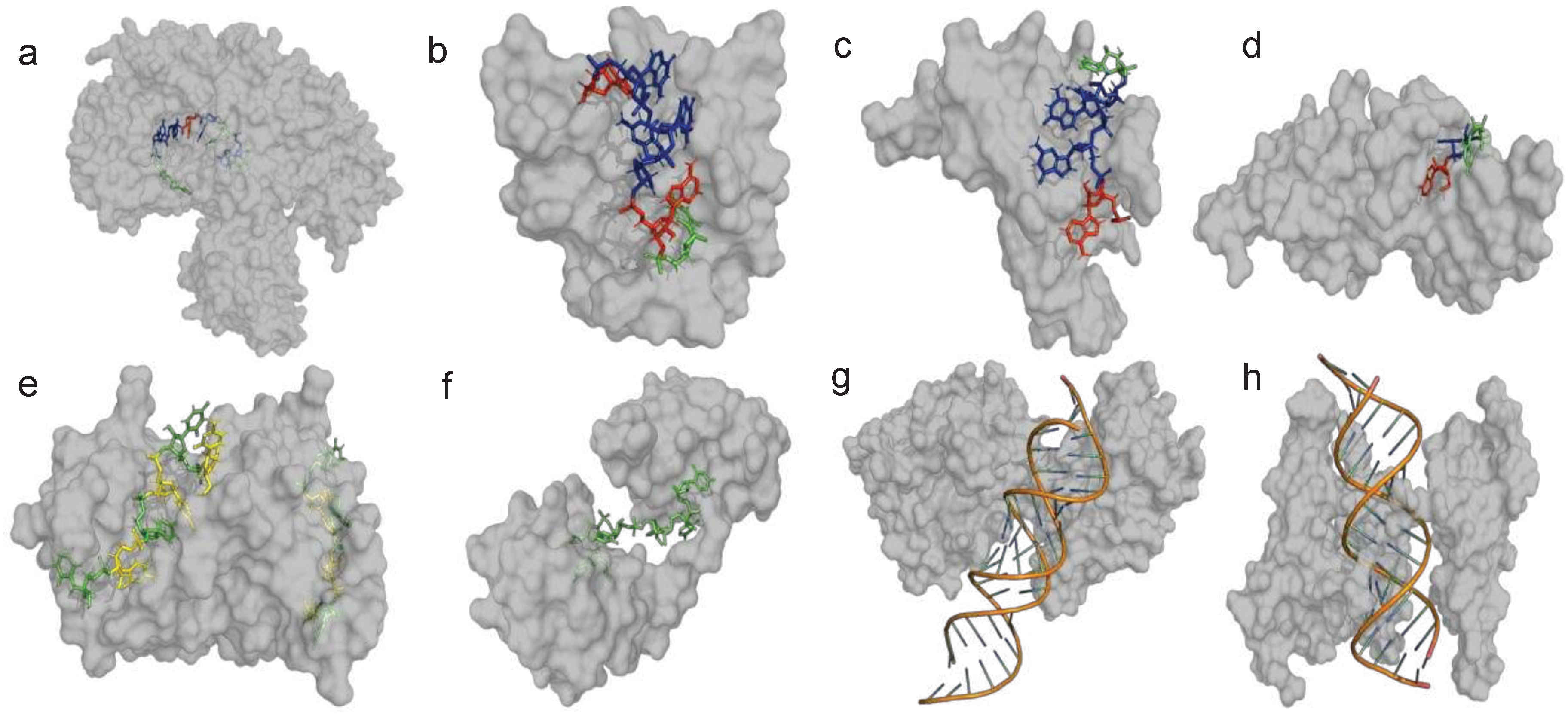
Figure 4.
EV-A71 infection perturbs normal RNA metabolism, which leads to a shift in the cellular homeostasis. Shown in steps 1-7 are some of the main cellular mechanisms that occur in the host cell. The mechanisms that are illustrated as faded are the ones that are affected by EV-A71 infection and are inhibited specifically due to the relocalization of cellular proteins to be used as ITAFs in EV-A71 IRES-mediated (cap-independent) translation. A few of the ITAFs; hnRNPA1, FBP1, FBP2, GADD34, and HuR are used to represent the whole repertoire of ITAFs, to help explain different viral mechanisms adapted by EV-A71 to facilitate its IRES-mediated translation. Positive ITAFs are colored in green while negative ITAFs are colored in red. These ITAFs are shown at the cellular functions that they regulate prior to EV-A71 infection.
Figure 4.
EV-A71 infection perturbs normal RNA metabolism, which leads to a shift in the cellular homeostasis. Shown in steps 1-7 are some of the main cellular mechanisms that occur in the host cell. The mechanisms that are illustrated as faded are the ones that are affected by EV-A71 infection and are inhibited specifically due to the relocalization of cellular proteins to be used as ITAFs in EV-A71 IRES-mediated (cap-independent) translation. A few of the ITAFs; hnRNPA1, FBP1, FBP2, GADD34, and HuR are used to represent the whole repertoire of ITAFs, to help explain different viral mechanisms adapted by EV-A71 to facilitate its IRES-mediated translation. Positive ITAFs are colored in green while negative ITAFs are colored in red. These ITAFs are shown at the cellular functions that they regulate prior to EV-A71 infection.
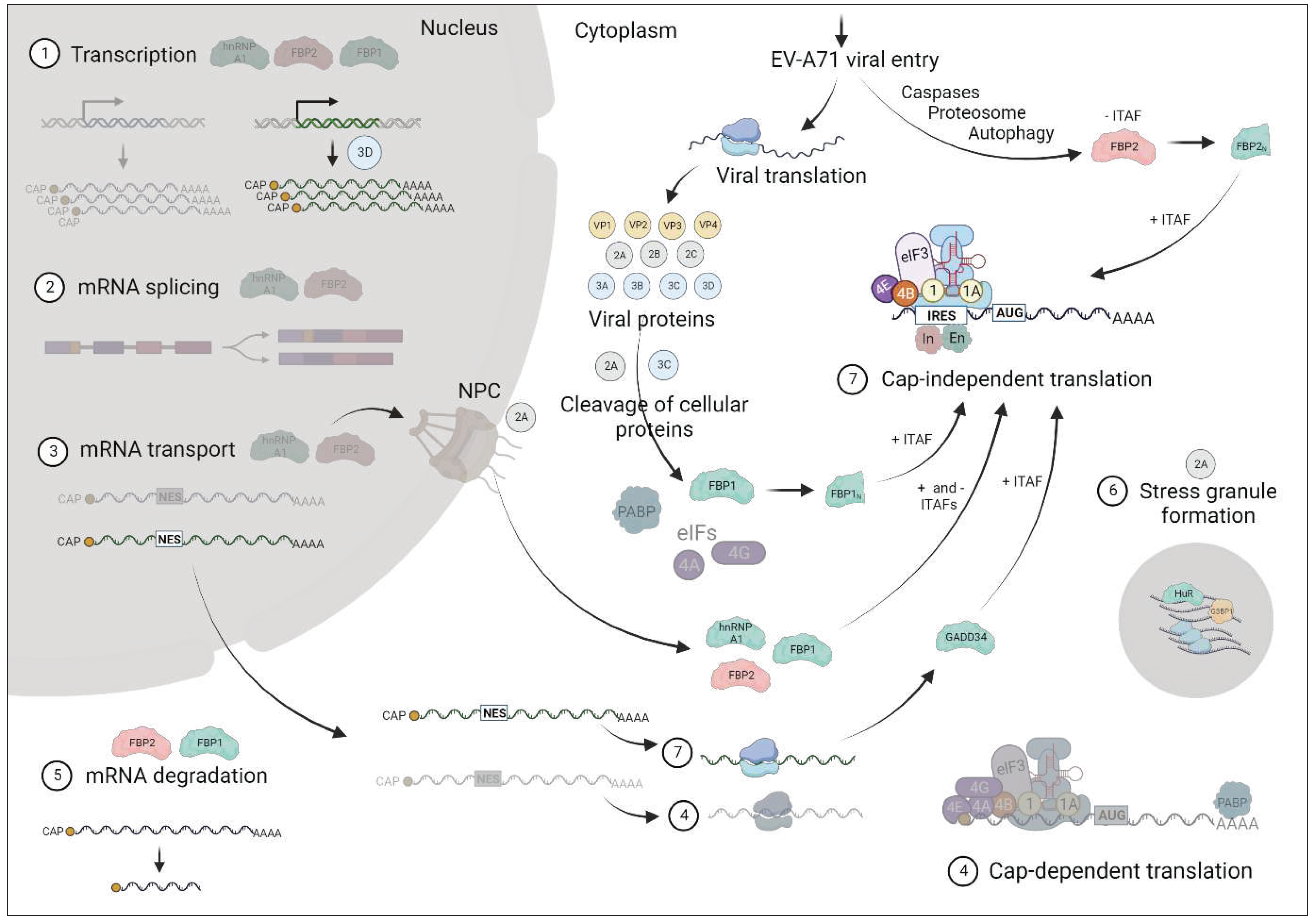

Table 1.
Cellular proteins (ITAFs) that are repurposed upon EV-A71 infection. The ITAFs, their cellular distributions pre- and post-infection, their regulatory site(s) within the IRES (if known), and their effect on IRES-mediated translation are indicated. The PDB IDs of the proteins (bound to RNA) shown in Figure 3 are indicated in Bold.
Table 1.
Cellular proteins (ITAFs) that are repurposed upon EV-A71 infection. The ITAFs, their cellular distributions pre- and post-infection, their regulatory site(s) within the IRES (if known), and their effect on IRES-mediated translation are indicated. The PDB IDs of the proteins (bound to RNA) shown in Figure 3 are indicated in Bold.
| ITAF | Other Names | Cellular distribution | Regulatory Site | IRES activity | Reference(s) | PDB ID(s) | |
|---|---|---|---|---|---|---|---|
| Pre-infection | Post-infection | ||||||
| Heterogeneous nuclear ribonucleoprotein A1 | hnRNP A1 | Nucleus | Cytoplasm | SLII and IV | Enhancement | [34] |
4YOE: 1.92 Å 5MPL: NMR 5MPG: NMR |
| AU-rich element RNA-binding protein 1 | AUF1, hnRNP D |
Nucleus | Cytoplasm | SLII | Inhibition | [45], [46] | 1WTB: NMR 1X0F: NMR |
| Heterogeneous nuclear ribonucleoprotein K | hnRNP K | Nucleus | Cytoplasm | SLI, II, and IV | - | [35] | 1J5K: NMR 1ZZI: 1.80Å 1ZZJ: 2.30 Å 7CRE: 3.00 Å |
| Poly(c)-binding protein 1 | PCBP1, hnRNP E1 |
Nucleus, Cytoplasm |
Cytoplasm | SLI and IV | Enhancement | [56] | 1ZTG: 3 Å 3VKE: 1.77 Å |
| Polypyrimidine tract-binding protein 1 | PTB, PTB1 PTBP1 hnRNP I |
Nucleus | Cytoplasm | SLVI + linker (564 - 742 nt) | Enhancement | [69] |
2N3O: NMR 2AD9: NMR 2ADB: NMR 2ADC: NMR |
| Far upstream binding protein 2 | FBP2, FUBP2 KSRP, KHSRP |
Nucleus | Cytoplasm | SLI-SLII (1-167) SLII-SLIII (91-228) SLVI + linker (566-745) | Inhibition | [36] | 4B8T: NMR |
| C-terminus cleaved FBP2 | FBP21–503 | - | - | 5’-UTR | Enhancement | [76] | |
| N-terminus cleaved FBP2 | FBP2190 –711 | - | - | 5’-UTR | Inhibition | [76] | |
| Far upstream binding protein 1 | FBP1 FUBP1 |
Nucleus | Cytoplasm | Linker (686-714 nt) | Enhancement | [82] | 1J4W: NMR |
| Cleaved FBP1 | FBP11-371 | - | - | Linker (656-674 nt) | Enhancement | [83] | |
| Argonaute 2 | Ago2 | Nucleus, Cytoplasm (P-bodies) |
- | SLII | Enhancement | [48] | 5KI6: 2.15 Å |
| Human Antigen R | HuR, ELAVL11 |
Nucleus, shuttles to Cytoplasm2 |
Cytoplasm3 | SLII | Enhancement | [48] | 4ED5: 2.00 Å 6G2K: 2.00 Å 6GC5: 1.90 Å 6GD2: 1.90 Å |
|
Moloney leukemia virus 10 (C-terminus domain) |
MOV10 | Cytoplasm | Cytoplasm (P-bodies & aggregates perinuclear) |
SLI and IRES (Excluding the linker region) |
Enhancement | [98] | |
| Silent mating type information regulation 2 homolog 1 | SIRT1 | Nucleus | Cytoplasm | SLI, II, III and V |
Inhibition | [107] | |
| 68-kDa Src-associated protein in mitosis | Sam68, KHDRBS1 |
Nucleus | Cytoplasm | SLIV and V | Enhancement | [117] | |
| Far upstream element-binding protein 3 | FUBP3 | Nucleus | Cytoplasm | 5’-UTR | Enhancement | [120,121] | |
| growth arrest and DNA damage-inducible protein 34 | GADD34, PPP1R15A |
ER membrane Mitochondial membrane |
- | 5’-UTR | Enhancement | [131,132] | |
| DEAD-box protein 3 | DDX3 | Nucleus Cytoplasm |
- | Full 5’-UTR | Enhancement | [136] | 6O5F: 2.5 Å |
| apolipoprotein B mRNA-editing enzyme, catalytic polypeptide-like 3G | APOBEC3G, A3G |
Cytoplasm (mainly) Nucleus P-bodies |
Cytoplasm Virions |
SLI and II | Inhibition | [56], [142] | 5ZVA: 2.30 Å 5ZVB: 2.00 Å 6BUX: 1.86 Å 7UXD: 1.50 Å |
| Staufen homolog 1 | Staufen1 | Rough ER Cytoplasm |
- | 5’-UTR | Enhancement | [147] | 6HTU: 2.89 Å |
| Heterogeneous nuclear ribonucleoprotein H | HNRNP H | Nucleus | Cytoplasm | 5’-UTR | Enhancement | [120] | |
| Heterogeneous nuclear ribonucleoprotein F | HNRNP F | Nucleus | Cytoplasm | 5’-UTR | Enhancement | [120] |
2KFY: NMR 2KG0: NMR 2KG1: NMR |
| Early growth response-1 | EGR1 | Nucleus | Cytoplasm | SLI and IV | Enhancement | [155] | 4R2A: 1.59 Å |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



