
Submitted:
11 December 2023
Posted:
12 December 2023
You are already at the latest version
Abstract
Abstract: To explore the interrelationship between groundwater microbial communities and hydrochemical characteristics in Mancheng district, Baoding city, samples from karst groundwater and porous groundwater were collected respectively. The hydrochemical analysis and 16s RNA gene V4-V5 region sequencing method were used to detail the water environmental characteristics and microbial community distribution pattern in the study area. The hydrochemical results reveal significant differences in major ion contents and other hydrochemical indices between karst water and porous water. Microbial community analysis demonstrates that Proteobacteria, Actinobacteria, and Firmicutes are the dominant taxa in both types of water bodies, but their proportions vary significantly in different water bodies. The difference in microbial communities between karst water and porous water reflects the influence of environmental factors such as ion concentration and pH on microbial distribution. Redundancy analysis (RDA) indicates significant associations between microbial community composition and environmental factors in both types of groundwater environments. Studying the distribution pattern of microbial communities and the connection with environmental factors in two types of groundwater provides important basis for interpreting the functional roles of microbes in groundwater ecosystems.
Keywords:
environmental microbiology
; hydrogeology
; hydrochemical characteristics
; microbial ecology
; environmental response characteristics
1. Introduction
Natural spring areas, as the unique zones where groundwater and surface water converge, have abundant water resources and distinct ecological environments. Research into environmental microbial communities and their functions holds significant implications worldwide. In recent years, it has been discovered that microbes in groundwater environments play vital roles in maintaining aquatic ecological stability, protecting water resources, and safeguarding ecosystem health (M R Shields et al.,2016; L Yan et al.,2021; E V Wehncke & N A Mariano, 2020). Studies have shown that groundwater microbes can degrade organic matter, remove heavy metals, and eliminate pollutants such as nitrogen and phosphorus, offering biotechnological methods for protecting and purifying water resources (P Sharma et al.,2021). Additionally, groundwater microbes play key roles in the Earth's biogeochemical cycles, such as those of carbon, nitrogen, and sulfur (C E Wegner et al.,2019; C Rinke et al.,2018). In-depth research into groundwater microbial communities and their functions is of great theoretical significance for understanding global ecosystem functions and elucidating Earth's ecological evolution. The study of groundwater microbes provides new perspectives and methods for ecological protection and the sustainable use of water resources.
The Yimuquan Spring area in Baoding city is located at the southern margin of the North China Plain, in the pre-mountain area, occupying the upstream area of the groundwater dynamics system in central Hebei, and boasts abundant groundwater resources and complex hydrogeological conditions. According to the data from China Water Resources Yearbook (2021), the groundwater resources of Baoding city account for 79.4% of the city's total water resources, playing a crucial role in agricultural production, ecological environment protection, and residential water supply (Y Xiao et al., 2021). However, with the growth of the population and economic development, the water resources of the Yimuquan Spring area in Baoding city are facing increasingly severe pressure, and the scientific management and protection of groundwater resources have become increasingly important. Although groundwater microorganisms play an important role in biogeochemical processes, the relationship between the microbial community structure in the groundwater layer of this area and environmental characteristics has not yet been thoroughly studied (K L Korbel et al., 2022; A Ritter et al., 2021). Therefore, this study aims to conduct a systematic survey of the microbial communities in the groundwater layer in the Yimuquan Spring area in Baoding city, to delve deeply into the distribution and diversity of microbial communities in groundwater and their relationships with environmental factors, and reveal the structure and environmental response characteristics of groundwater microbial communities. In doing so, we hope to understand the ecological functions of microorganisms in groundwater, providing a scientific basis for the protection and rational use of groundwater resources. Ultimately, this work will provide strong support for the sustainable development of this area, the improvement of the ecological environment, and the enhancement of people's livelihood and well-being.
2. Materials and Methods
2.1. Study Area Overview
The study area is primarily located in Mancheng District, Baoding City, Hebei Province, encompassing parts of the upstream and downstream spring areas. It includes a range between 38°56′0"-38°58′30"N and 115°12′30"-115°24′30"E. The climate is a warm temperate semi-humid continental climate, with an average annual temperature of 13℃ and average annual precipitation of 550mm (Y N Pan et al., 2020). The terrain is mainly plain, representing various geological origins such as ancient river floodplains, alluvial fans, and aeolian terraces. The topography is high in the north and low in the south, with altitudes ranging between 50-100 meters (W Chen, 2008). The hydrogeological conditions are complex. The main aquifers include the Quaternary alluvial layer and the Tertiary sedimentary layer. The former is the main aquifer, with a thickness of 20-100 meters. The latter has a thickness of about 50-150 meters and slightly less water content (Z Y Huang et al., 2015).
The study area has abundant overall resources, with the annual available groundwater resources being approximately 936 million cubic meters. Among this, 70% is used for agricultural irrigation, followed by industrial production and domestic water use (Y Xiao et al., 2021). The groundwater quality is safe with low nitrite, nitrate, and heavy metal contents. The concentrations of the former two are far below the values stipulated in the "Groundwater Quality Standards (GB/T 14848-2017)" (D M Han et al., 2021). In addition, the total hardness of groundwater in the study area is relatively high (average 450 mg/L) and exhibits alkaline characteristics (pH values are between 7.5-8.5). The main cations are calcium, magnesium, and sodium, while the main anions are carbonate, bicarbonate, and sulfate. The water chemical type is primarily HCO3·SO4-Ca·Mg (X G Fu et al., 2021).
2.2. Sample Collection and Testing
In January 2022, considering the groundwater flow direction and actual water usage in the study area, we collected nine groundwater samples from the Yimuquan area in Mancheng District, Baoding City. The sampling depth was between 90 and 200 meters (Figure 1). Prior to sampling, we pumped the well for 0.5-2 hours until the water quality was clear and stable. Samples were then taken from the water outlet directly above the submersible pump at the ground surface, without pipe aeration or disinfection. Samples were collected in three parts: Firstly, on-site measurement of water temperature, pH, electrical conductivity (Ec), and oxidation-reduction potential (ORP) was performed using a Hach HQ40D portable multi-parameter water quality analyzer. Secondly, the samples were preserved according to the "Standard Test Methods for Drinking Water" (GB/T 5750-2006), and then the total dissolved solids (TDS) were measured using the dry weighing method. An ICS-600 Ion Chromatograph was used to determine Cl-, SO42-, NO3-, NO2-, and F-. Inductively Coupled Plasma Optical Emission Spectrometry (ICP-OES) was used to determine Na+, Al3+, Fe3+, Mn2+, and TDS. After the tests were completed, the relative measurement error was calculated to be 1.7% based on the standards announced in the "Standard Test Methods for Drinking Water - Quality Control in Water Quality Analysis (GB/T 5750.3-2006)" and DD2014-2015 "Technical Requirements for Quality Control of Sample Analysis in Groundwater Pollution Investigation and Evaluation". Thirdly, 1 L of each water sample was collected and filtered using a 0.22μm water system filter membrane using a suction pump, then stored in a dry ice box. The samples were then transported to Shanghai Personal Biotechnology Co., Ltd. for testing.
DNA extraction was performed using the OMEGA Soil DNA Kit (D5625-01) from Omega Bio-Tek, and the extracted DNA was tested. The DNA concentration was measured using a NanoDrop ND - 1000 spectrophotometer (Thermo Fisher Scientific) at 260 nm and 280 nm, and the quality of the DNA was assessed using 1% agarose gel electrophoresis. The concentration of the DNA solution was adjusted, with the working DNA solution stored at 4°C and the storage solution stored at -20°C. The V4-V5 region of the bacterial 16S rRNA gene was amplified by PCR using the forward primer 515F (5′-GTGCCAGCMGCCGCGGTAA-3′) and the reverse primer 907R (5′-CCGTCAATTCMTTTRAGTTT-3′). A 7-bp barcode specific to the samples was incorporated into the primers for multiplex sequencing. The PCR amplification products were purified using Vazyme VAHTSTM DNA Clean Beads (Vazyme, Nanjing, China) and quantified using the Quant-iT PicoGreen dsDNA Assay Kit (Invitrogen, Carlsbad, CA, USA). After the individual quantification step, the amplified products were pooled in equimolar amounts and sequenced on the Illlumina MiSeq platform using the MiSeq Reagent Kit v3 for 2*250 bp paired-end sequencing.
2.3. Sequence and Data Analysis
Bioinformatic data was processed using QIIME2 software. The DADA2 plugin was used for sequence quality filtering, denoising, merging, and chimera removal. The diversity plugin was used to estimate Alpha diversity data, with samples rarefied to 10,000 sequences per sample. Within the feature classifier plugin, a Bayesian classifier was used to classify ASVs. Subsequently, Alpha diversity indices and Beta diversity indices were calculated and visualized along with clustering results using QIIME2 and R software package (v3.2.0).
Water chemistry data and its correlation with species were compiled and analyzed using Excel 2007, SPSS 25.0, and Canoco5 for data collection, collation, and statistical analysis.
3. Results and Analysis
3.1. Hydrochemical Characteristics
According to the analysis, the groundwater in the study area is weakly alkaline, and there is not much difference in the basic characteristics of the pore water in the upper reaches and the karst water in the middle, with pH ranging from 7.44 to 7.88 and an average value of 7.71 (Table 1). The TDS value is low, with the highest value being only 572.47 mg·L-1 and an average value of 408.99 mg·L-1. In terms of composition, there is not much difference in the cation content of the two, with high levels of Ca2+ and Mg2+, reflecting the lithological conditions of the strata in the area. The content of K+, Na+, and Mg2+ in pore water is slightly lower than that in karst water. As for anionic composition, the SO42- and NO3- levels in karst water are more than twice as high as in pore water, while Cl- and HCO3- are relatively low. The hydrochemical type of all five karst water samples is represented as HCO3-Ca·Mg, while in pore water samples, two types, HCO3-Ca·Mg (BD6 & BD10) and HCO3·Cl-Ca·Mg (BD8 & BD9), are present (Figure 3).
In addition, the redox environment of the two is different, with karst water having a higher amount of dissolved oxygen and slightly weaker oxidizing properties, and pore water having a lower amount of dissolved oxygen and stronger oxidizing properties relative to karst water. This is consistent with the findings of Q C Qin et al., 2020.
3.2. Microbial Community Structure
3.2.1. Diversity Analysis
In this study, 142847-206200 pairs of sequences were obtained, and 117067-168692 groups of high-quality sequences were analyzed and tested. From the rarefaction curves (Figure 4 and Figure 5), it can be seen that with the increase in drawdown depth, the rarefaction curves of Chao1 and Shannon indices gradually become noticeably flat. This suggests that the samples obtained are sufficiently representative in terms of microbial quantity and population richness, and the sequencing depth has met the needs to reflect the diversity and quantity of samples.
From the Alpha index box plot, it can be seen that the Goods_coverage index in the sequenced samples ranges from 0.969 to 0.998, reflecting a high and reliable sequencing depth (Figure 6). In terms of the Chao1 index, the mean of the KW group is 3583.7, which is significantly lower than the mean of 4586.25 in the PW group, reflecting that the samples in the KW group have relatively high richness in combination with the Observed species index. The Shannon and Simpson indices reflect that the samples in the PW group are superior in terms of species diversity. The Faith pd index shows that the samples in the PW group also have advantages in terms of the evolutionary-based diversity and species evenness of the microbiota.
3.2.2. Microbial Composition Analysis
At the phylum level, Proteobacteria, Actinobacteria, Bacteroidetes and Firmicutes are the bacterial species with the highest abundance in the groundwater in the study area, with average contents all above 6%, and single abundances accounting for 76.13%~38.53%, 22.45%~3.74%, 22.67%~2.49% of the total community respectively (Figure 7a). In addition, species with an average abundance greater than 3% include Planctomycetes (4.06%), Chloroflexi (4.04%), Omnitrophicaeota (3.95%), and Acidobacteria (3.26%). The groundwater in the study area moves from west to east, resulting in gradient differences in water composition and underground environmental conditions, and the species composition changes accordingly (S Amalfitano A et al., 2014). In this process, the relative abundances of Proteobacteria and Omnitrophicaeota decreased, with decreases of 10.26% and 40.94% respectively. On the other hand, the relative abundances of Actinobacteria, Bacteroidetes, Planctomycetes, Chloroflexi and Acidobacteria increased to varying degrees (20.93%~95.52%). These increases in species abundance may imply a series of diversified ecological niches that have arisen or been enhanced due to the gradual changes in water composition and underground environmental conditions. Bacteroidetes are famous for their ability to decompose polysaccharides, which helps in the degradation of organic matter (D L Kirchman, 2002). Some studies have shown that Planctomycetes can degrade organic substances such as polysaccharides and polyphenols (J Wang et al., 2002). When groundwater flows from mountains to plains, it may change the types and contents of organic matter, thereby benefiting the growth of Planctomycetes. Chloroflexi can also play a key role in the degradation of organic matter and denitrification process (L A Hug et al., 2013). Acidobacteria are involved in various biogeochemical processes such as organic matter degradation and nitrate reduction (R T Jones et al., 2009).
The content of Nitrospirae, Rokubacteria, and Elusimicrobia are relatively uniform, with contents in the KW group of 3.06%, 1.42%, and 1.02% respectively, and in the PW group of 2.63%, 1.47%, and 0.9%. This indicates that they have potential adaptability to environmental changes, or that they are involved in basic biogeochemical processes that are common throughout the groundwater ecosystem. The ecological functions and relative abundance of Rokubacteria and Elusimicrobia in groundwater environments are relatively stable, which may be related to their physiological characteristics and ecological niches (C J Castelle et al., 2015; D P Herlemann et al., 2011). The similar abundance of Nitrospirae in karst water and pore water may imply that the nitrogen cycling processes in these two groundwater environments are relatively similar. Furthermore, the compositional differences between the KW and PW groups are most evident in Actinobacteria, Bacteroidetes, Planctomycetes and Acidobacteria with the above four types of bacteria contributing 9.09% of the positive compositional difference to the PW group. This indicates that they may be better adapted to the environmental conditions of the PW group.
At the genus level, the composition varies greatly among different samples, and there is no obvious dominant genus (Figure 7b). The dominant genus of BD1 is Omnitrophicaeota (13.49%), that of BD3 is Psychrobacter (22.66%) and Glutamicibacter (12.14%), that of BD4 is Gallionella (24.26%) and Sideroxydans (22.47%), and that of BD8 is Ochrobactrum (12.38%), and there is no obvious dominant genus in other samples(Fig.8). Overall, the microbial community structure of the KW group shows a more obvious dominant genus, while the microbial community structure of the PW group is more balanced. This difference may reflect the diversity of environmental conditions and microbial ecological adaptability, suggesting the complexity and uniqueness of microbial ecosystems in each groundwater sample (C A Lozupone and R Knight , 2007; T M Flynn et al., 2013; K Faust and J Raes, 2012).
3.2.3. Analysis of Differences in Microbial Community Composition
Considering the high dispersion at the genus level and no obvious dominant genus in the microbial taxa of the study area's groundwater, PCA analysis was carried out based on the species matrix at the phylum level (C A Lozupone et al., 2007). As can be seen from the figure, PCo1 and PCo2 can represent 30.1% and 19.2% of the variation characteristics, respectively (Figure 9). The distribution of samples in the KW group is generally to the left, mainly concentrated in the area of the PCo1 axis from -0.2 to 0.05. Due to the higher community abundance and diversity in the PW group compared to the KW group, all samples fall within the -0.1 to 0.1 range on the PCo2 axis.
Given that microbial community characteristic data often display a discontinuous distribution or even discrete characteristics, heatmaps and random forest analyses use decision trees to establish non-linear sample classifiers to find marker species, and are widely used in environmental microbial analyses (L Richard, et al., 2017, S Louca et al., 2016). From the heatmap of community composition at the phylum level, it can be seen that Actinobacteria, Bacteroidetes, Firmicutes and Deinococcus-Thermus are more abundant in the PW group, while Omnitrophicaeota and Patescibacteria are more indicative in the KW group. This is related to environmental tolerance as well as dissolved oxygen concentration and organic matter content (J A Vorholt, 2012; C A Nickerson et al., 2004).
Random forest analysis indicates that Actinobacteria, Patescibacteria, Firmicutes, Deinococcus-Thermus, etc., are the intergroup difference marker species calculated by the classifier model (Figure 11). Members of Patescibacteria have a strong ability to survive in anaerobic or microaerobic environments and can use some organic matter as a carbon source and energy (C J Castelle et al., 2017). Firmicutes and Deinococcus-Thermus are better adapted to extreme environments with poor nutritional conditions (S Maloy, 2013; K S Makarova et al., 2001). Combining species biological characteristics, we believe that differences in nutritional conditions and dissolved oxygen content are the main factors in screening for marker species of intergroup differences between the KW and PW groups. This is consistent with the research understanding of D C White et al.,1995 and K Manoj et al., 2016.
3.3. Correlation Analysis of Microbial Community and Environmental Characteristics
In terms of hydrological characteristics, the abundance of microbial species shows a significant correlation with some hydrological characteristic indicators. As can be seen from the correlation heatmap, in the KW environment, various ions in the water body support the abundance of species and to some extent support the development of species evolution and diversity. Among them, the higher ORP representing the level of oxidation is closely associated with the improvement of species diversity (Figure 12). The Chao1 diversity index is significantly positively correlated with ORP (Oxidation-Reduction Potential) and Na+, and significantly negatively correlated with pH. This implies that microbial abundance will be higher in environments with high oxidation-reduction potential and high Na+ concentration, and species abundance will decrease in environments with low pH values. The Shannon diversity index is extremely positively correlated with ORP. This indicates that the diversity of microbial species will increase in environments with high ORP values. The PW environment is different, the overall abundance of species relatively prefers a weakly oxidizing environment, and the diversity and evolution level of species relatively depend on a high dissolved oxygen alkaline environment. The Shannon diversity index is significantly positively correlated with pH and DO (Dissolved Oxygen), and significantly negatively correlated with Ec (Conductivity), Ca2+, Cl-, HCO3-, NO3- and other factors. This implies that in environments with high pH values and high dissolved oxygen concentrations, the diversity of microbial populations will increase, while in environments with high conductivity and high concentrations of ions such as Ca2+, Cl-, HCO3-, NO3-, the diversity of microbial populations will decrease. The Pielou_e uniformity index is significantly positively correlated with pH and DO, and significantly negatively correlated with Ec, Ca2+, Cl-, HCO3-, NO3- and other factors. This means that in environments with high pH values and high dissolved oxygen concentrations, the uniformity of microbial populations will increase, while in environments with high conductivity and high concentrations of ions such as Ca2+, Cl-, HCO3-, NO3-, the uniformity of microbial populations will decrease.
Selecting the main environmental feature indicators, RDA redundancy analysis was performed on the top 20 species in abundance at the phylum level in the samples to intuitively show the relationship between various species and hydrological and hydrochemical characteristic factors (Figure 13 and Figure 14).
Through RDA redundancy analysis, it was found that in the KW group, Proteobacteria only showed a significant positive correlation with NO2-. This may be attributed to some members of Proteobacteria being able to use NO2- for nitrate respiration. Actinobacteria, Cyanobacteria, Deinococcus-Thermus showed positive correlations with HCO3-, Na+, Ca2+, Mg2+, SO42-, with HCO3- reaching a significant level. This represents that the aforementioned microbes require the above ions as key elements for growth and metabolism. Among them, many known species in Actinobacteria can use bicarbonate ions as a carbon source (G M Zaitsev et al., 1998). Omnitrophicaeota, Rokubacteria, Nanoarchaeaeota were negatively correlated with pH and temperature, and positively correlated with ORP, DO, and the main ions in the water body, with pH, ORP, K+, and Cu showing significant correlations. This indicates that these microbes prefer lower pH and temperatures, but require higher oxidation-reduction potential and dissolved oxygen concentrations. Firmicutes, Euryarchaeota, Verrucomicrobia showed negative correlations with Ec, DO, and the main ion content in the water body, and positive correlations with ORP, temperature, F-, silicic acid, and Fe content, with Ec, Mg2+, Cl-, NO3-, F-, silicic acid, and Fe showing significant correlations, indicating that they have adapted to living conditions rich in F-, silicic acid, and Fe. Planctomycetes, Nitrospirae, Thaumarchaeota, Latescibacteria were positively correlated with ORP, silicic acid, and negatively correlated with Ba, Cr and the main ions in the water body, with ORP, Ba, Cr showing significant correlations. This indicates that their survival ability is stronger in environments with high oxidation-reduction potential and silicic acid, but they may be sensitive to the presence of elements such as Ba and Cr.
In the PW group, Proteobacteria, Planctomycetes, Acidobacteria, Nitrospirae were positively correlated with pH, DO, F-, and negatively correlated with Ec, temperature, and other main ions. Among them, pH, HCO3-, Mg2+, Ba, Co, etc., reached extremely significant levels. This implies that these microbes are adapted to high pH, high dissolved oxygen, and F- environments, but are not adapted to high conductivity and temperature, or have difficulty surviving in environments with high main ion concentration. This is consistent with the study by T M Lapara, et al. 2001. These microbes may also use these environmental factors as energy or nutrients through some special biochemical pathways. Bacteroidetes, Chloroflexi, Omnitrophicaeota, Rokubacteria, Euryarchaeota, Deinococcus-Thermus were negatively correlated with pH, DO, F-, and positively correlated with Ec, Ba, Co, Sr, and the main ions in the water body. Among them, Ec, Ca2+, Mg2+, Cl-, SO42-, NO3-, Co, Sr showed extremely significant correlations. This indicates that these microbes may be adapted to low pH, low dissolved oxygen, and low F- environments, and have stronger survival capabilities in high conductivity and ion concentration environments, or can metabolize some substances in high ion environments (Hahn, M. W. 2004). Cyanobacteria, Thaumarchaeota, Latescibacteria were significantly negatively correlated with ORP and significantly positively correlated with NO2-. This may mean that these microbes are more likely to survive in environments with lower oxidation-reduction potential and can use NO2- as an energy source or nutrient (H W Paerl et al., 2011).
As can be seen, with the change in water environmental conditions, the response of species and community composition has changed accordingly, but the response of most species to environmental factors has only changed in degree, not in direction. Individual species such as Omnitrophicaeota, Euryarchaeota, Thaumarchaeota, and Latescibacteria have changed in their response direction to some environmental factors. Among them, Omnitrophicaeota and Euryarchaeota have changed in their response to basic hydrochemical ions. The correlation of Omnitrophicaeota with K+, Na+, SO42- changed from a positive correlation in karst water to a negative correlation (0.767→-0.81*). The correlation of Euryarchaeota with Ec, TDS, Ca2+, Mg2+, Cl-, NO3- changed from a negative correlation in karst water to a positive correlation (-0.82*→0.756). The response changes of Thaumarchaeota and Latescibacteria are consistent, affecting the correlation with ORP (0.803→-0.81*) and NO2- (-0.347→0.883*). These microbes have shown different ecological niche widths in different water body environments, reflecting their outstanding environmental adaptability (G E Hutchinson, 1957). Specifically, the ecological niche widths of Omnitrophicaeota, Thaumarchaeota and Euryarchaeota in pore water significantly exceed those in karst water, implying that the pore water environment may have larger environmental fluctuations, forcing these microbes to expand their resource acquisition strategies (M A Leibold, 1995). On the other hand, the resource selection of these microbes in different water bodies also showed differences, possibly due to the different competitive pressures they face. For example, Omnitrophicaeota mainly uses S5 and S6 resources in karst water, while it mainly uses S4, S8, and S9 resources in pore water, which may be a response to the competitive pressure of other microbial populations in the environment (P Chesson, 2000). In addition, changes in niche width and resource selection may also contribute to the diversity of the ecosystem. Microbes with broader niches are more likely to survive in changing environments, thereby increasing the stability and diversity of the ecosystem (P Chesson, 2000). These findings provide a new perspective for us to understand the adaptability of microbes to environmental changes, and their role in maintaining ecological diversity.
4. Conclusions
Through hydrochemical analysis, it was found that the groundwater in the study area is weakly alkaline. Karst water and pore water show significant differences in terms of major ion composition, dissolved oxygen content, and redox properties, reflecting two distinctly different environments due to the differences in stratum lithology and redox conditions. The groundwater microbial community is mainly composed of Proteobacteria, Actinobacteria, Bacteroidetes, etc., and their relative abundance has significantly changed in the process of groundwater migration from west to east, reflecting the influence of water composition and underground environmental conditions. Different bacterial groups may show different ecological adaptability to changes in specific environmental conditions, implying the complexity and uniqueness of the underground water microbial ecosystem. Through heat map and random forest analysis, we found that Actinobacteria, Bacteroidetes, Firmicutes, etc. are more abundant in the PW group, while Omnitrophicaeota and Cyanobacteria are more indicative in the KW group, which may be related to environmental tolerance, dissolved oxygen concentration, and organic matter content. The existence of these indicator species indicates that the differences in nutritional conditions and dissolved oxygen content are the main factors that form the differences in the microbial community structure in the KW and PW groups in groundwater. Principal Component Analysis (PCA) and Redundancy Analysis (RDA) show that there is a significant correlation between the abundance and diversity of microbial communities and the hydrological features of the environment they inhabit. These environmental features include redox potential, ion concentration, dissolved oxygen concentration, conductivity, and pH. Individual microbial species such as Omnitrophicaeota, Euryarchaeota, Thaumarchaeota, and Latescibacteria show significant adaptive changes in different water environments, which further reflects the important role of microbes in responding to environmental changes and maintaining ecological diversity.
References
- Retter, C. Karwautz, and C. Griebler, "Groundwater Microbial Communities in Times of Climate Change," Current Issues in Molecular Biology, vol. 41, no. 1, pp. 509-538, 2021. [CrossRef]
- A. Lozupone, R. Knight, "Global patterns in bacterial diversity," Proceedings of the National Academy of Sciences of the United States of America, vol. 104, no. 27, 2007.
- A. Nickerson, C. M. Ott, S. J. Mister, B. J. Morrow, L. Burns-Keliher, D. L. Pierson, "Microbial responses to microgravity and other low-shear environments," Microbiology and Molecular Biology Reviews, vol. 68, no. 2, pp. 345-361, 2004.
- E. Wegner, M. Gaspar, P. Geesink, et al., "Biogeochemical Regimes in Shallow Aquifers Reflect the Metabolic Coupling of the Elements Nitrogen, Sulfur, and Carbon," Applied and Environmental Microbiology, vol. 85, no. 9, 2019. [CrossRef]
- C. J. Castelle, C. T. Brown, B. C. Thomas, et al., "Unusual respiratory capacity and nitrogen metabolism in a Parcubacterium (OD1) of the Candidate Phyla Radiation," Scientific Reports, vol. 7, 40101, 2017. [CrossRef]
- C. J. Castelle, K. C. Wrighton, B. C. Thomas, et al., "Genomic expansion of domain archaea highlights roles for organisms from new phyla in anaerobic carbon cycling," Current Biology, vol. 25, no. 6, pp. 690-701, 2015. [CrossRef]
- C. Q. Qin, H. Ding, S. L. Li, et al., "Hydrogeochemical Dynamics and Response of Karst Catchment to Rainstorms in a Critical Zone Observatory (CZO), Southwest China," Frontiers in Water, vol. 2, 577511, 2020. [CrossRef]
- C. Rinke, F. Rubino, L. F. Messer, et al., "A phylogenomic and ecological analysis of the globally abundant Marine Group II archaea (Ca. Poseidoniales ord. nov.)," The ISME Journal, vol. 13, 2018. [CrossRef]
- China Water Resources Bulletin. (2021). Beijing: China Water & Power Press.
- C. White, D. B. Ringelberg, R. J. Palmer, "Quantitative Comparison of the in situ Microbial Communities in Different Biomes," 1995.
- D. L. Kirchman, "The ecology of Cytophaga-Flavobacteria in aquatic environments," FEMS Microbiology Ecology, vol. 39, no. 2, pp. 91-100, 2002.
- D. M. Han, M. J. Currell, H. Guo, "Controls on distributions of sulphate, fluoride, and salinity in aquitard porewater from the North China Plain: Long-term implications for groundwater quality," Journal of Hydrology, 2021.
- D. P. Herlemann, M. Labrenz, K. Jürgens, et al., "Transitions in bacterial communities along the 2000 km salinity gradient of the Baltic Sea," The ISME Journal, vol. 5, no. 10, pp. 1571-1579, 2011.
- V. Wehncke, N. A. Mariano, "Groundwater and Its Role in Maintaining the Ecological Functions of Ecosystems—A Review," [Online]. Available: 10.1007/978-3-030-65443-6_4, 2021.
- E. Hutchinson, "Concluding remarks - cold spring harbor symposia on quantitative biology. Reprinted in 1991: classics in theoretical biology," Bulletin of Mathematical Biology, vol. 53, no. 1507, pp. 193-213, 1957.
- M. Zaitsev, I. V. Tsitko, F. A. Rainey, et al., "New aerobic ammonium-dependent obligately oxalotrophic bacteria: description of Ammoniphilus oxalaticus gen. nov. sp. nov. and Ammoniphilus oxalivorans gen. nov. sp. nov.," International Journal of Systematic Bacteriology, vol. 48, pt. 1, no. 1, pp. 151, 1998. [CrossRef]
- W. Paerl, N. S. Hall, and E. S. Calandrino, "Controlling harmful cyanobacterial blooms in a world experiencing anthropogenic and climatic-induced change," Science of the Total Environment, vol. 409, no. 10, pp. 1739-1745, 2011. [CrossRef]
- A. Vorholt, "Microbial life in the phyllosphere," Nature Reviews Microbiology, vol. 10, no. 12, pp. 828-840, 2012. [CrossRef]
- Wang, C. Jenkins, R. I. Webb, and J. A. Fuerst, "Isolation of Gemmata-like and Isosphaera-like planctomycete bacteria from soil and freshwater," Applied and Environmental Microbiology, vol. 68, no. 1, pp. 417-422, 2002. [CrossRef]
- Faust, J. Raes, "Microbial interactions: from networks to models," Nature Reviews Microbiology, vol. 10, no. 8, pp. 538-550, 2012.
- K. L. Korbel, P. Greenfield, G. C. Hose, "Agricultural practices linked to shifts in groundwater microbial structure and denitrifying bacteria," The Science of the Total Environment, vol. 807, pt. 2, 150870, 2022.
- K. Manoj, N. Shrivastava, P. Teotia, P. Goyal, A. Varma, S. Sharma, N. Tuteja, V. Kumar, "Omics: Tools for Assessing Environmental Microbial Diversity and Composition," pp. 273-283, 2016.
- K. S. Makarova, L. Aravind, Y. I. Wolf, et al., "Genome of the Extremely Radiation-Resistant Bacterium Deinococcus radiodurans Viewed from the Perspective of Comparative Genomics," Microbiology & Molecular Biology Reviews, vol. 65, no. 1, 2001.
- A. Hug, C. J. Castelle, K. C. Wrighton, et al., "Community genomic analyses constrain the distribution of metabolic traits across the Chloroflexi phylum and indicate roles in sediment carbon cycling," Microbiome, vol. 1, no. 1, 22, 2013.
- L. Richard, Marchese, et al., "Comparison of the Predictive Performance and Interpretability of Random Forest and Linear Models on Benchmark Data Sets," Journal of Chemical Information and Modeling, vol. 57, no. 8, pp. 1773-1792, 2017. [CrossRef]
- L. Yan, M. Syrie, K. U. Totsche, R. Lehmann, M. Herrmann, K. Küsel, "Groundwater bacterial communities evolve over time in response to recharge," Water Research, vol. 201, 117290, 2021. [CrossRef]
- A. Leibold, "The Niche Concept Revisited: Mechanistic Models and Community Context," Ecology, vol. 76, no. 5, pp. 1371-1382, 1995.
- M. R. Shields, T. S. Bianchi, Y. Gélinas, et al., "Enhanced terrestrial carbon preservation promoted by reactive iron in deltaic sediments," Geophysical Research Letters, vol. 43, 2016. [CrossRef]
- M. W. Hahn, "Broad diversity of viable bacteria in ‘sterile’ (0.2 μm) filtered water," Research in Microbiology, 2004.
- Chesson, "Mechanisms Of Maintenance Of Species Diversity," 2000, vol. 31, no. 1, pp. 343-366. [CrossRef]
- Sharma, A. K. Pandey, S. H. Kim, et al., "Critical review on microbial community during in-situ bioremediation of heavy metals from industrial wastewater," Environmental Technology & Innovation, vol. 21, 101826, 2021. [CrossRef]
- T. Jones, M. S. Robeson, C. L. Lauber, et al., "A comprehensive survey of soil acidobacterial diversity using pyrosequencing and clone library analyses," The ISME Journal, vol. 3, no. 4, pp. 442-453, 2009.
- Amalfitano, A. Del Bon, A. Zoppini, S. Ghergo, S. Fazi, D. Parrone, P. Casella, F. Stano, and E. Preziosi, "Groundwater geochemistry and microbial community structure in the aquifer transition from volcanic to alluvial areas," Water Research, vol. 65, pp. 384-394, 2014. [CrossRef]
- Louca, S. M. S. Jacques, A. P. F. Pires, J. S. Leal, D. S. Srivastava, L. W. Parfrey, V. F. Farjalla, M. Doebeli, "High taxonomic variability despite stable functional structure across microbial communities," Nature Ecology & Evolution, vol. 1, no. 1, 0015, 2016.
- Maloy, "Brenner's Encyclopedia of Genetics," Brenners Encyclopedia of Genetics, vol. 91, no. 2001, pp. 2-3, 2013.
- M. Flynn, R. A. Sanford, H. Ryu, et al., "Functional microbial diversity explains groundwater chemistry in a pristine aquifer," Bmc Microbiology, vol. 13, no. 1, 146-146, 2013.
- T. M. Lapara, C. H. Nakatsu, L. M. Pantea, "Aerobic Biological Treatment Of a Pharmaceutical Wastewater: Effect Of Temperature On Cod Removal And Bacterial Community Development," Water Research: A Journal of the International Water Association, vol. 35, no. 18, pp. 4417-4425, 2001.
- Chen, "A View on Geomorphologic Zonalization of North China Plain," Geography and Geo-Information Science, 2008.
- G. Fu, Z. Dong, S. Gan, et al., "Groundwater Quality Evaluation for Potable Use and Associated Human Health Risk in Gaobeidian City, North China Plain," Hindawi Limited, 2021.
- N. Pan, Q. Li, Y. Liu, et al., "Hydroclimate variations in the Northern China Plain and their possible socio-cultural influences," [Online]. Available: 10.1007/978-3-658-03097-1_8, [Accessed: 22-Jul-2023].
- Xiao, Q. Hao, et al., "Accessible Phreatic Groundwater Resources in the Central Shijiazhuang of North China Plain: Perspective From the Hydrogeochemical Constraints," Frontiers in Environmental Science, vol. 9, 747097, 2021. [CrossRef]
- Y. Huang, Y. Pan, H. Gong, et al., "Subregional-scale groundwater depletion detected by GRACE for both shallow and deep aquifers in North China Plain," Geophysical Research Letters, vol. 42, no. 6, pp. 1791-1799, 2015.
Figure 1.
Geographical location and distribution of sampling points.
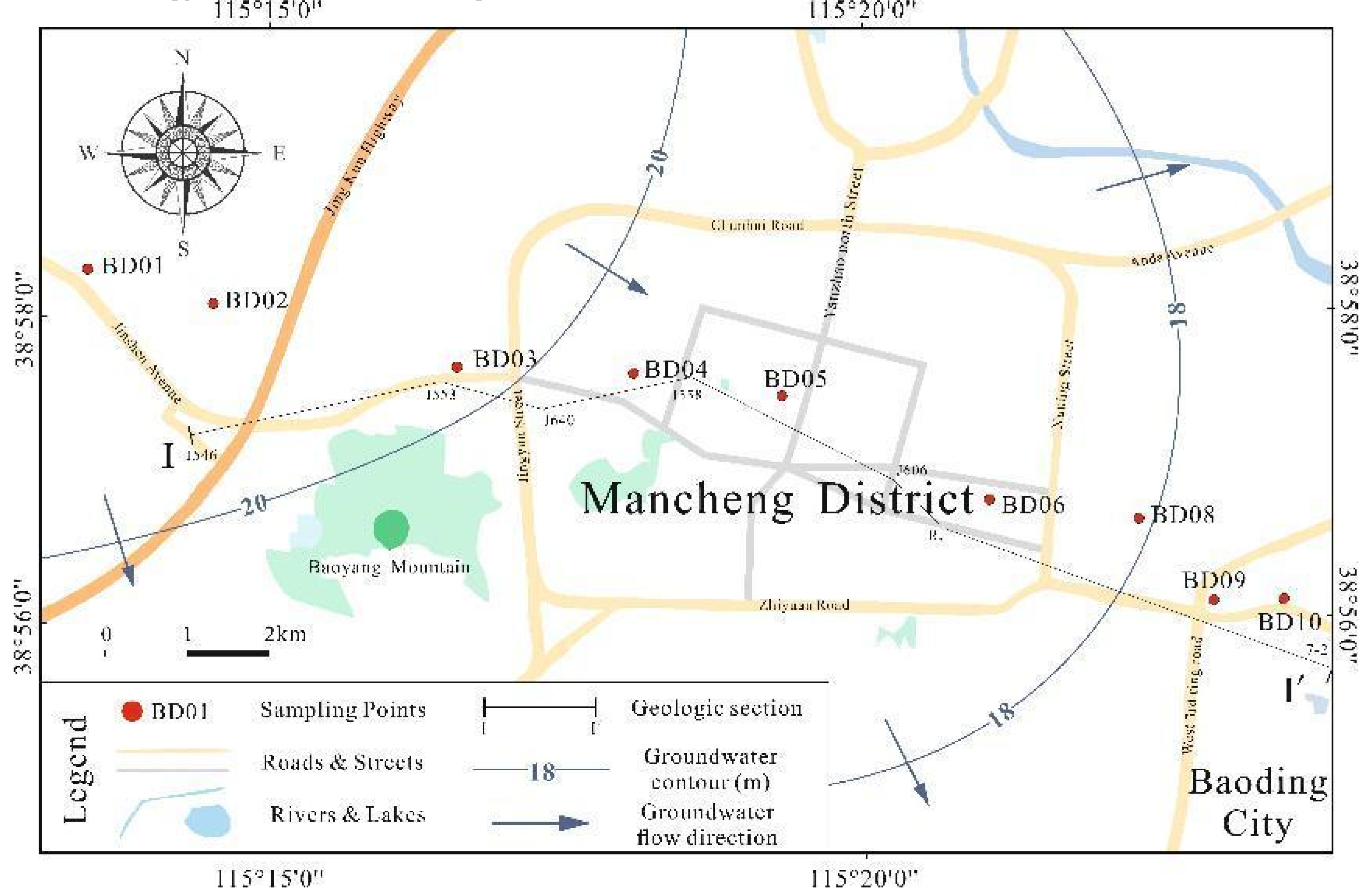
Figure 2.
Geological profile and hydrological characteristics of the study area.
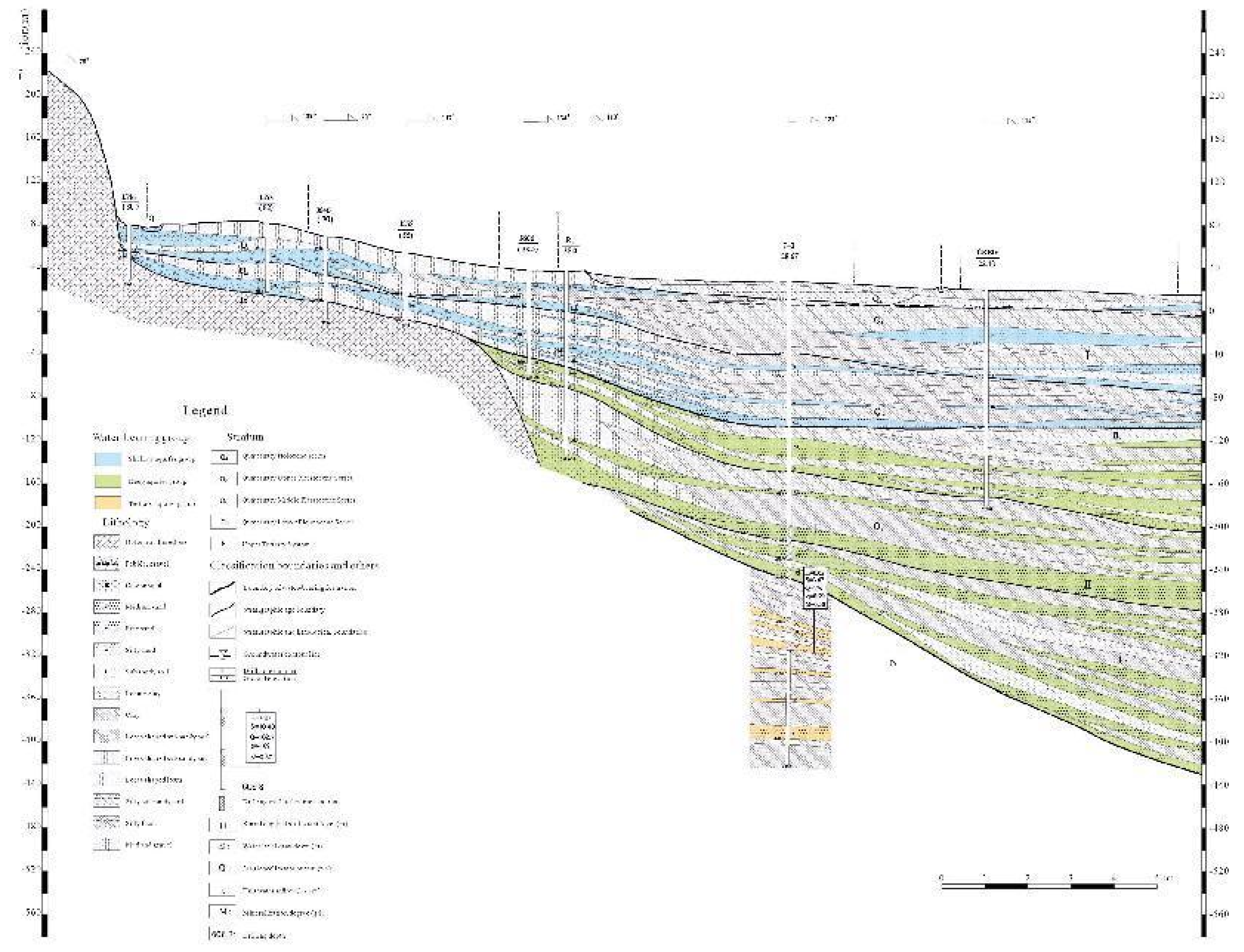
Figure 3.
Piper diagram of groundwater in Yimuquan area.
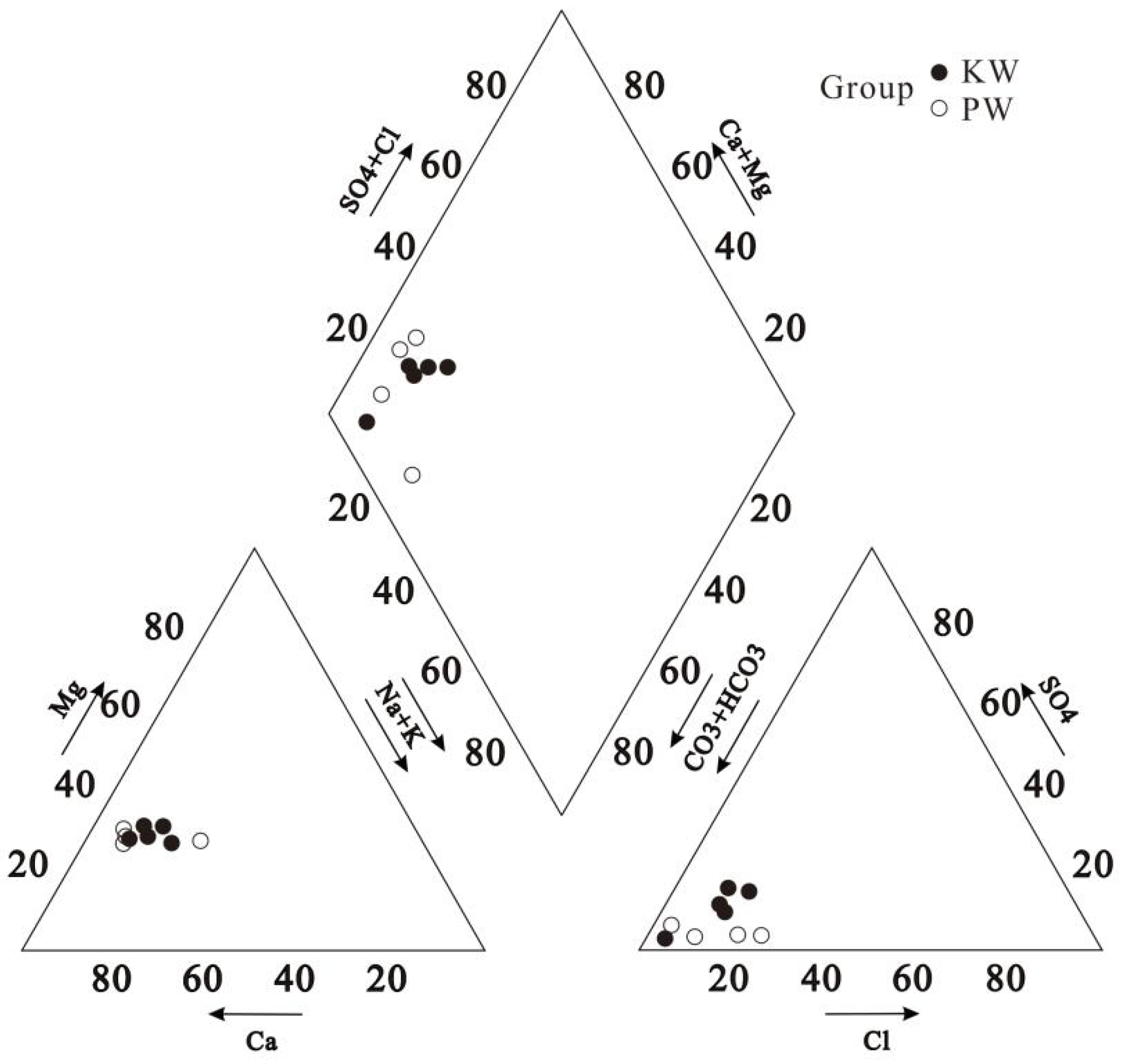
Figure 4.
Chao1 exponential sparse curve.
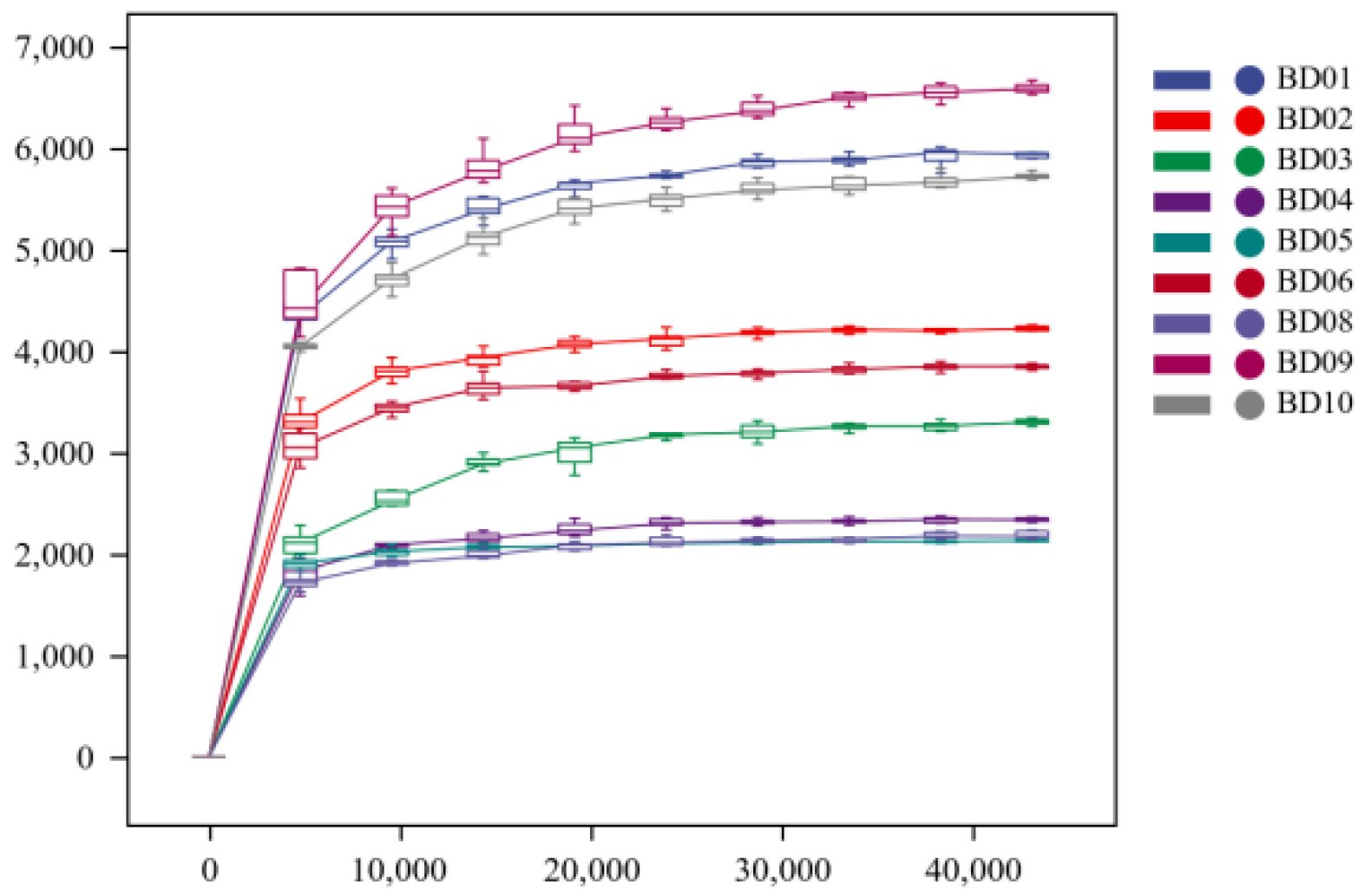
Figure 5.
Shannon exponential sparse curve.
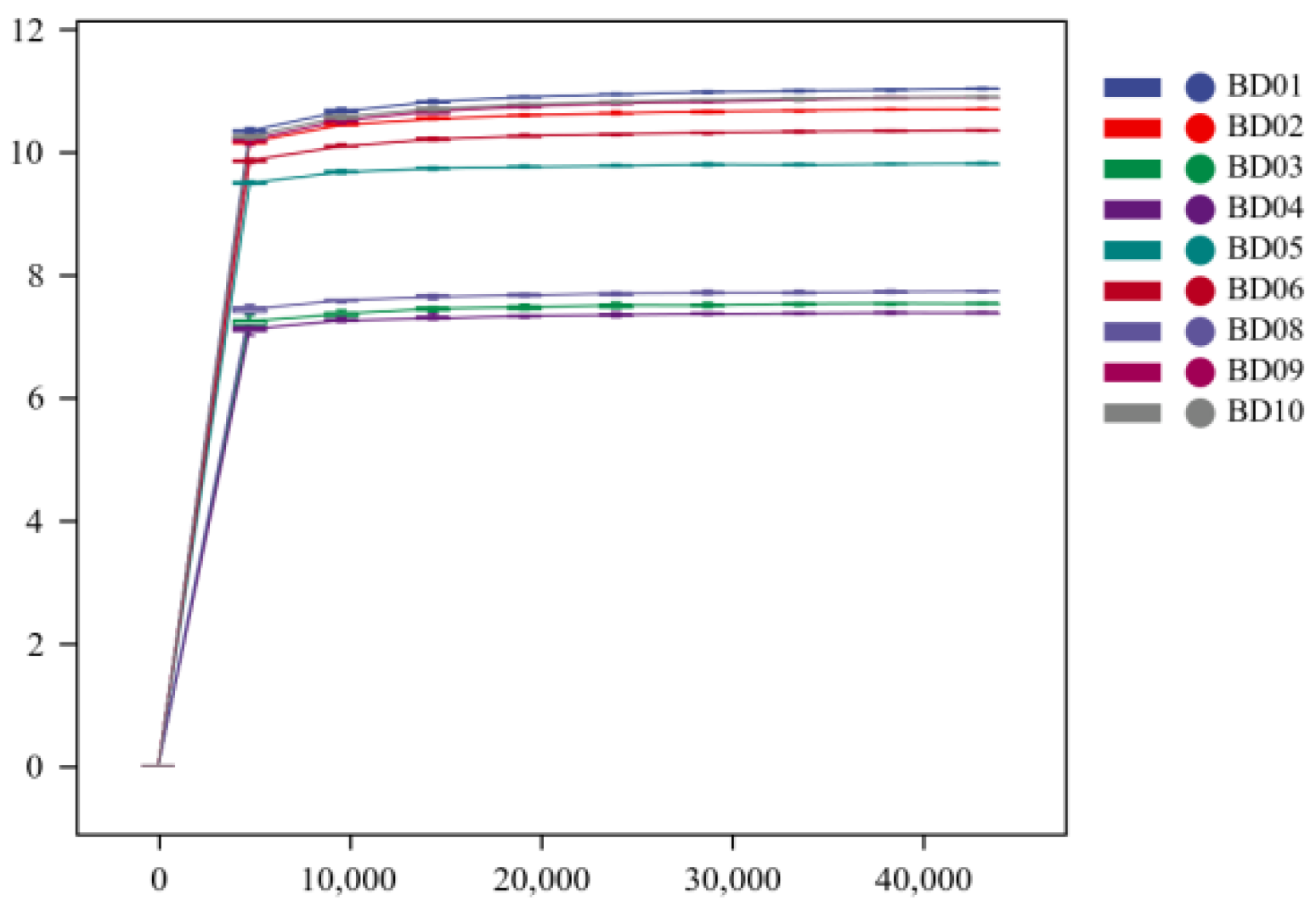
Figure 6.
Box chart of Alpha index of groundwater microbial community.
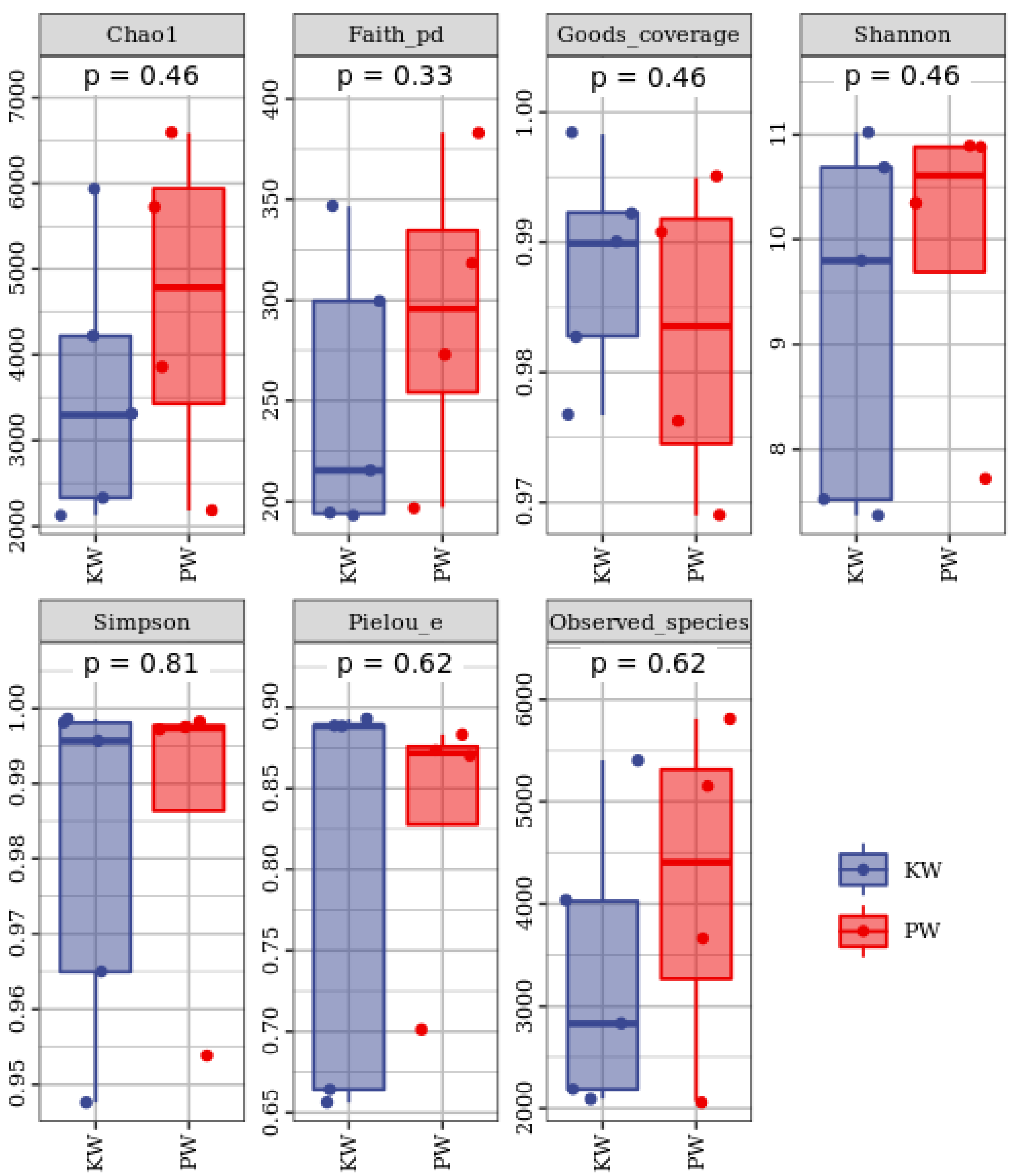
Figure 7.
The phylum and genus composition of the groundwater microbial community.
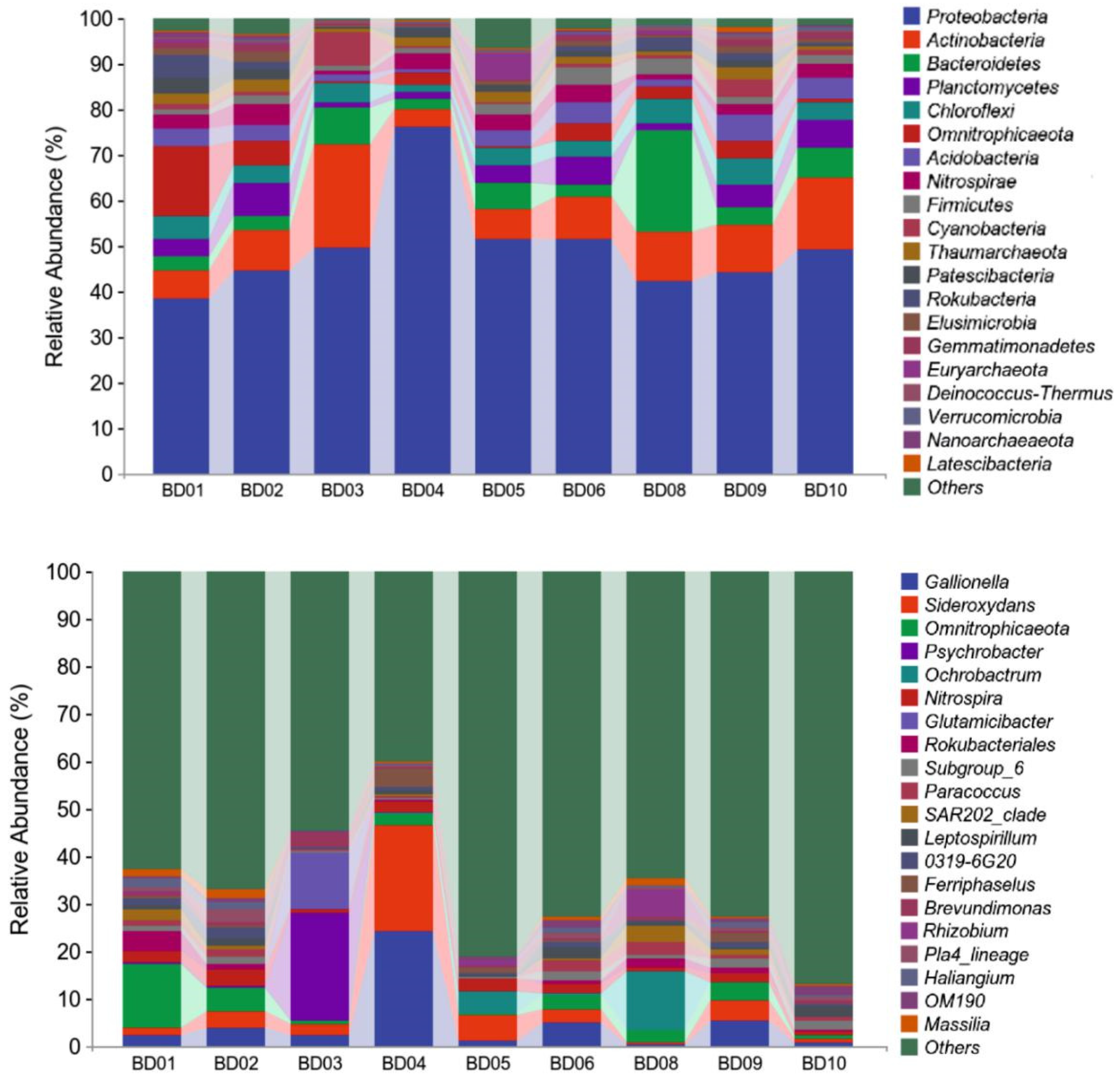
Figure 8.
Composition diagram of common bacterial genera in Yimuquan spring area.
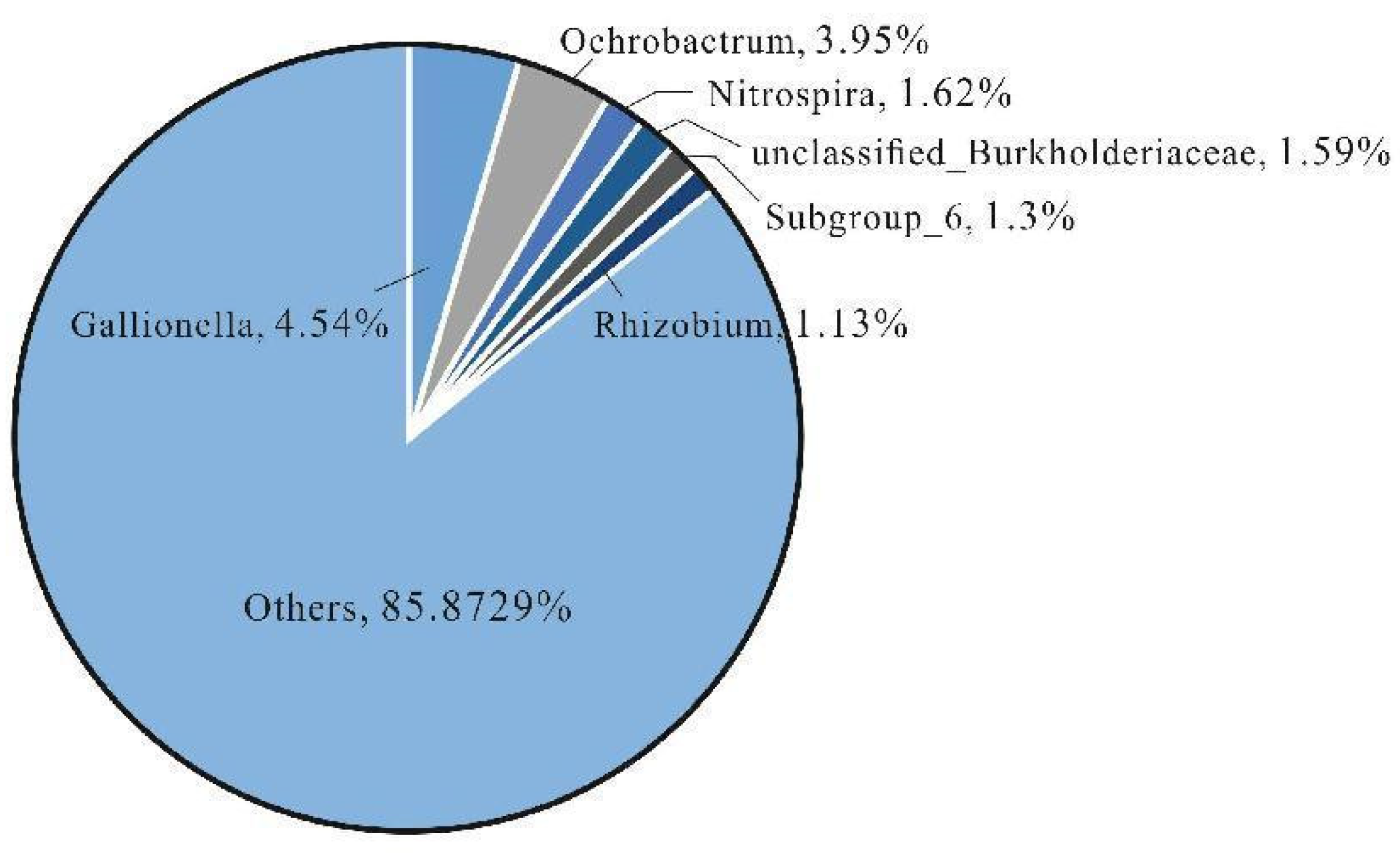
Figure 9.
Principal coordinate analysis of samples in Yimuquan area.
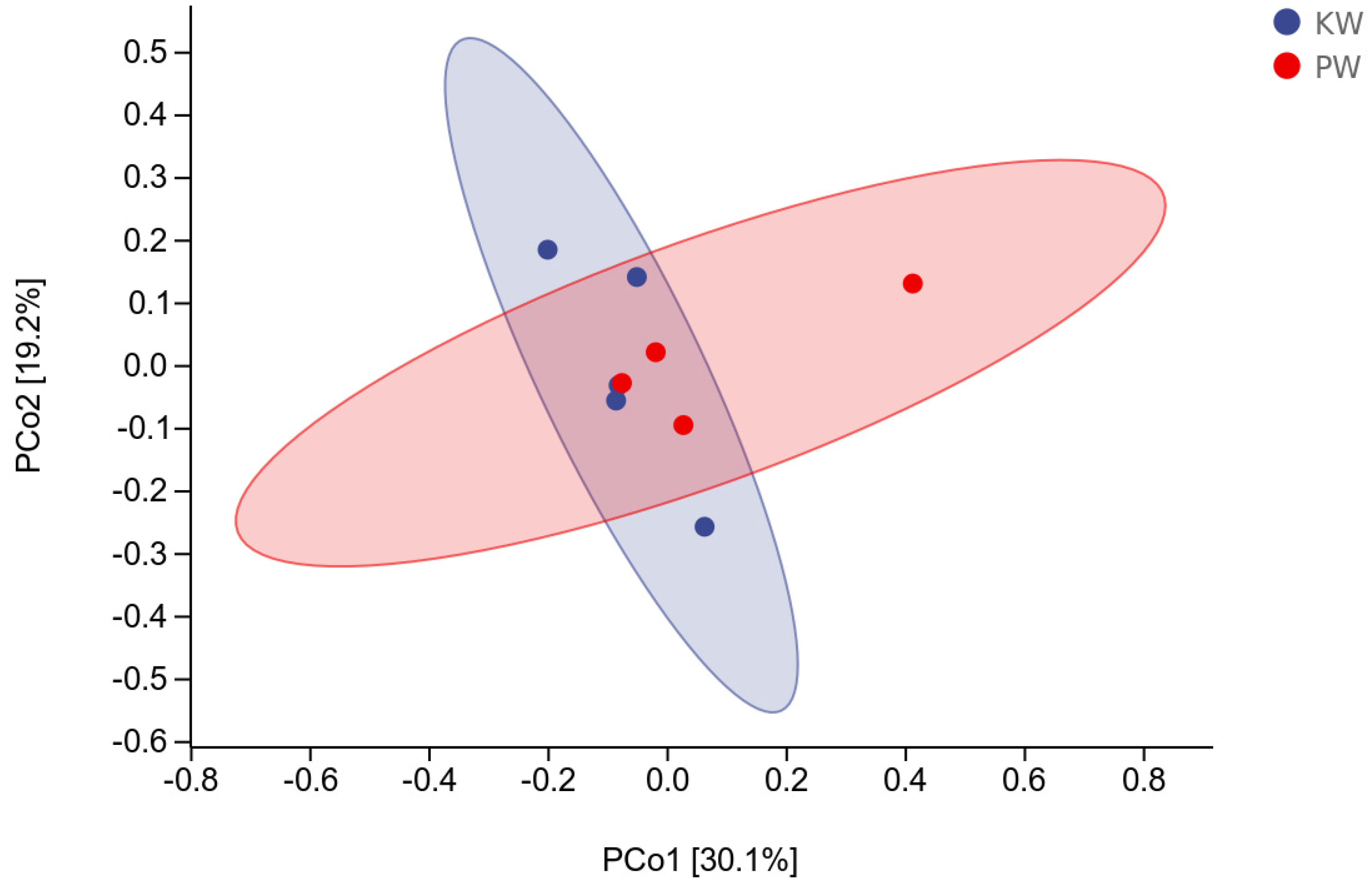
Figure 10.
Heat map of samples on phylum level.
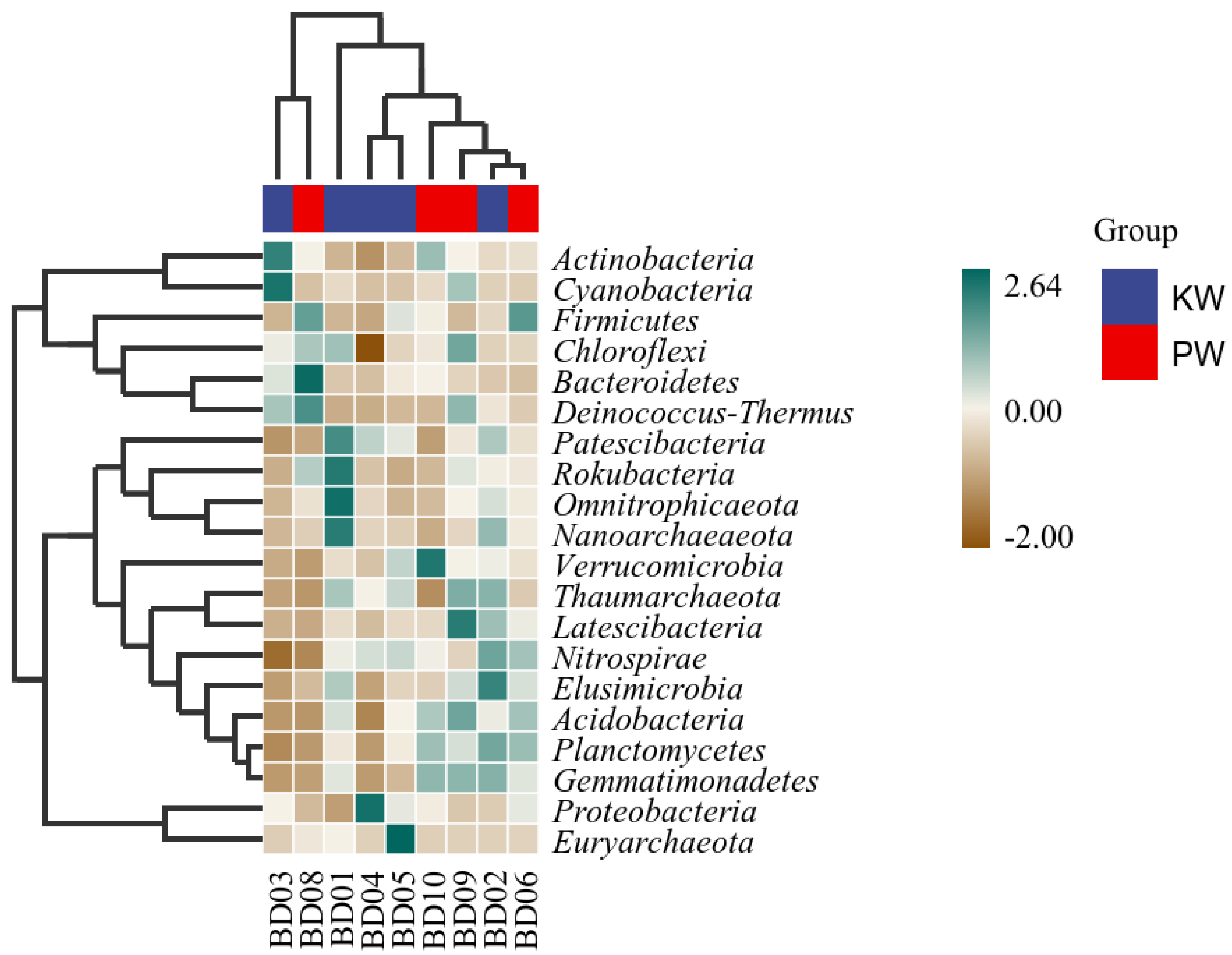
Figure 11.
Random forest analysis on phylum level.
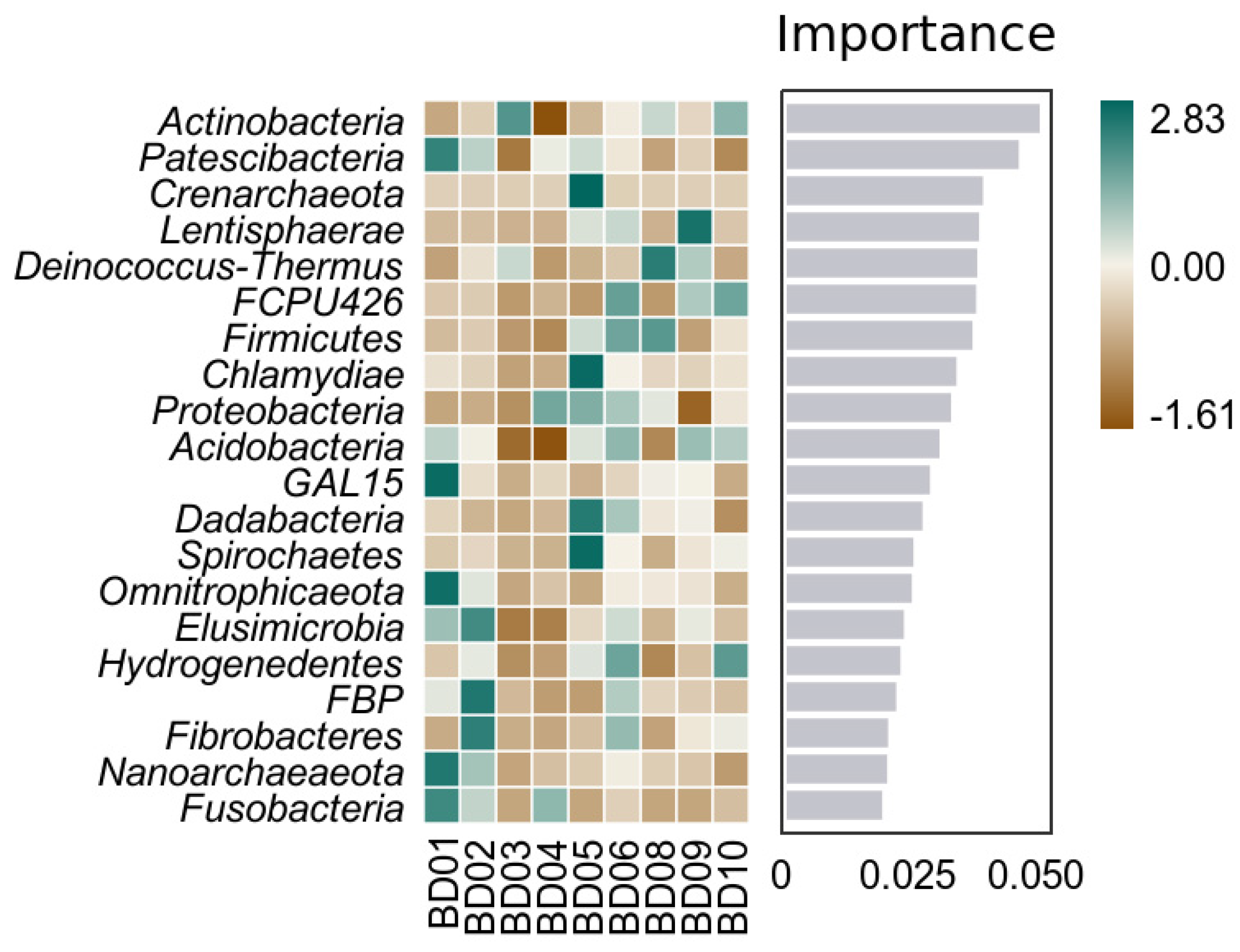
Figure 12.
Heat map of correlation between environmental factors and alpha index.
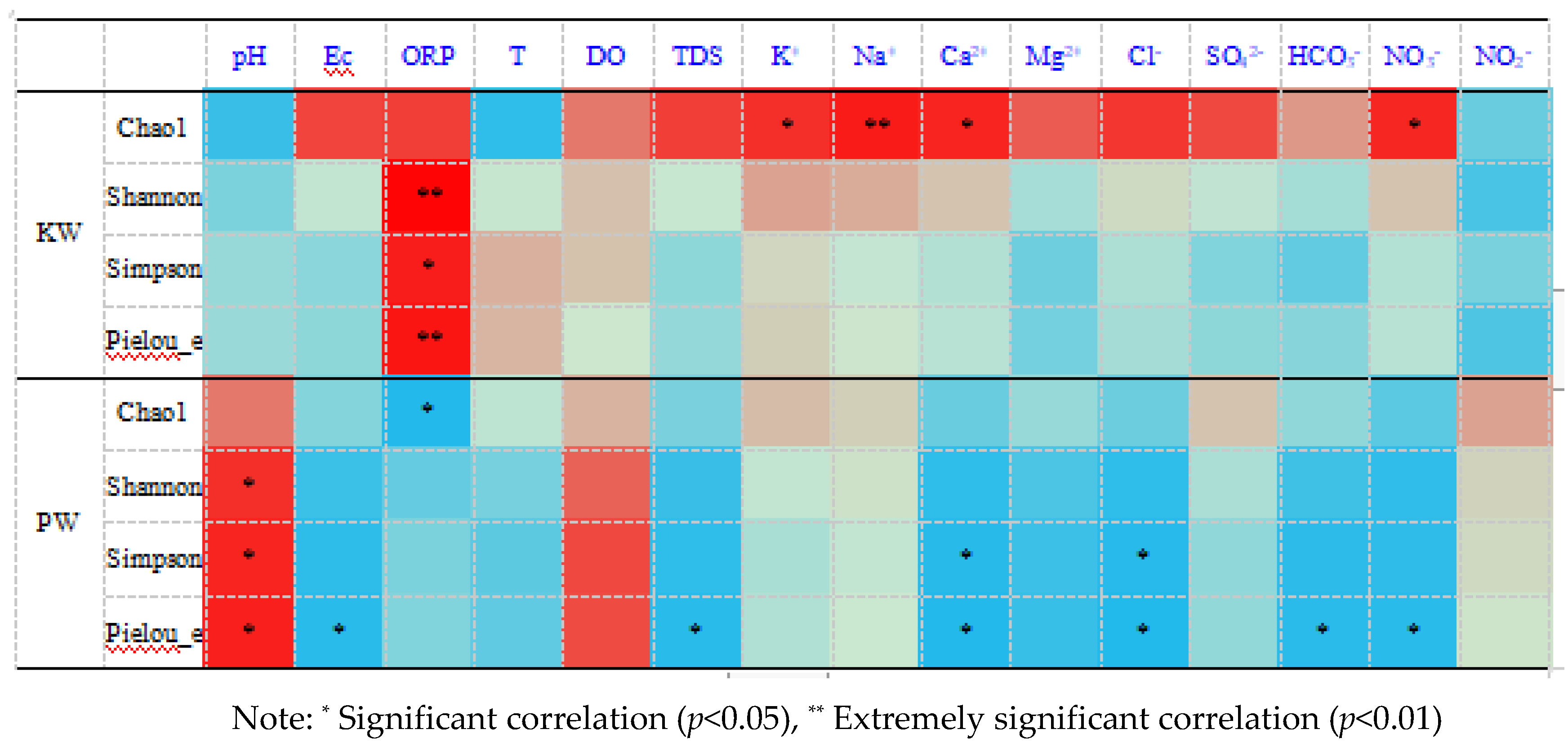
Figure 13.
Analysis of relationships between samples, influencing factors and genera.
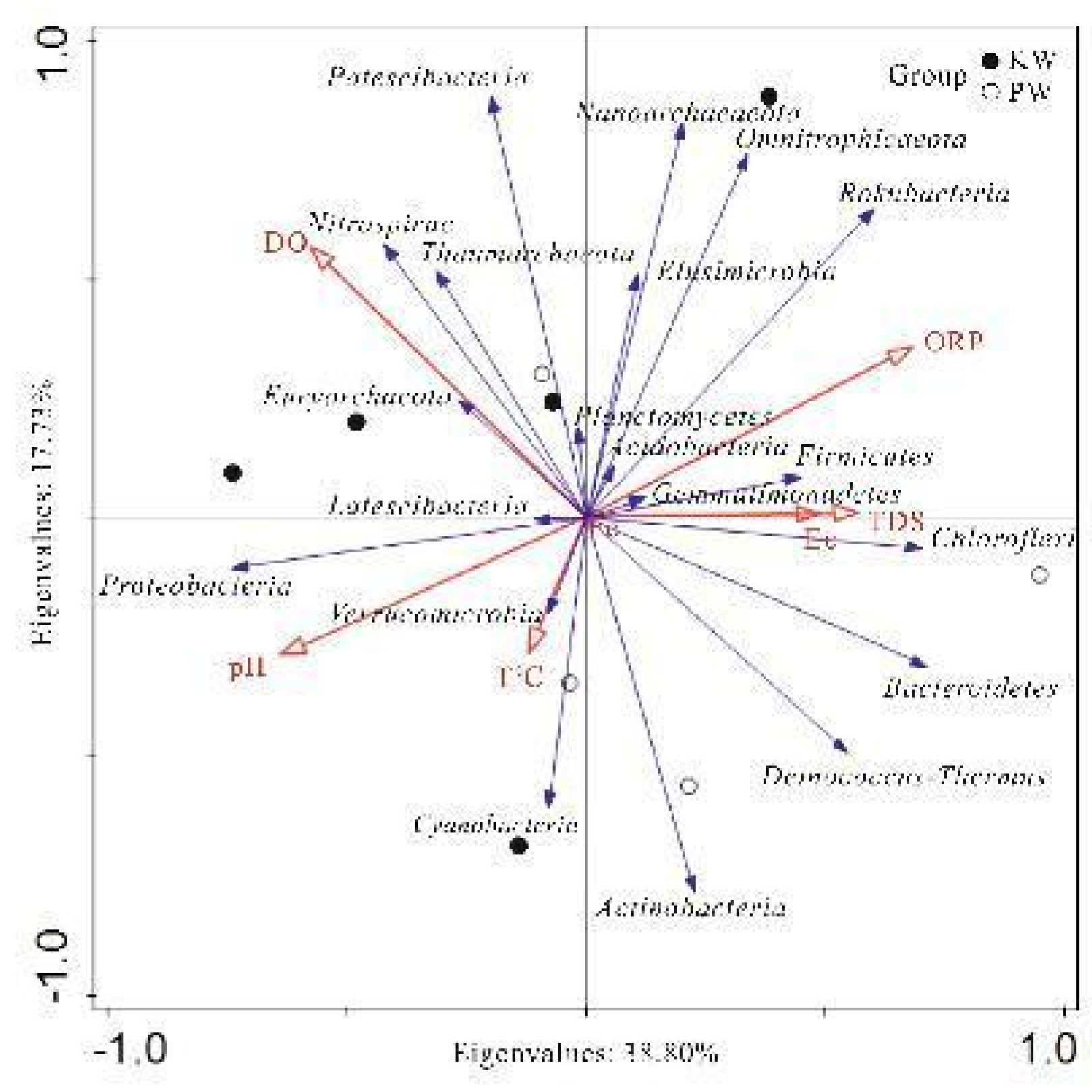
Figure 14.
Analysis of relationships between samples,influencing factors and genera.
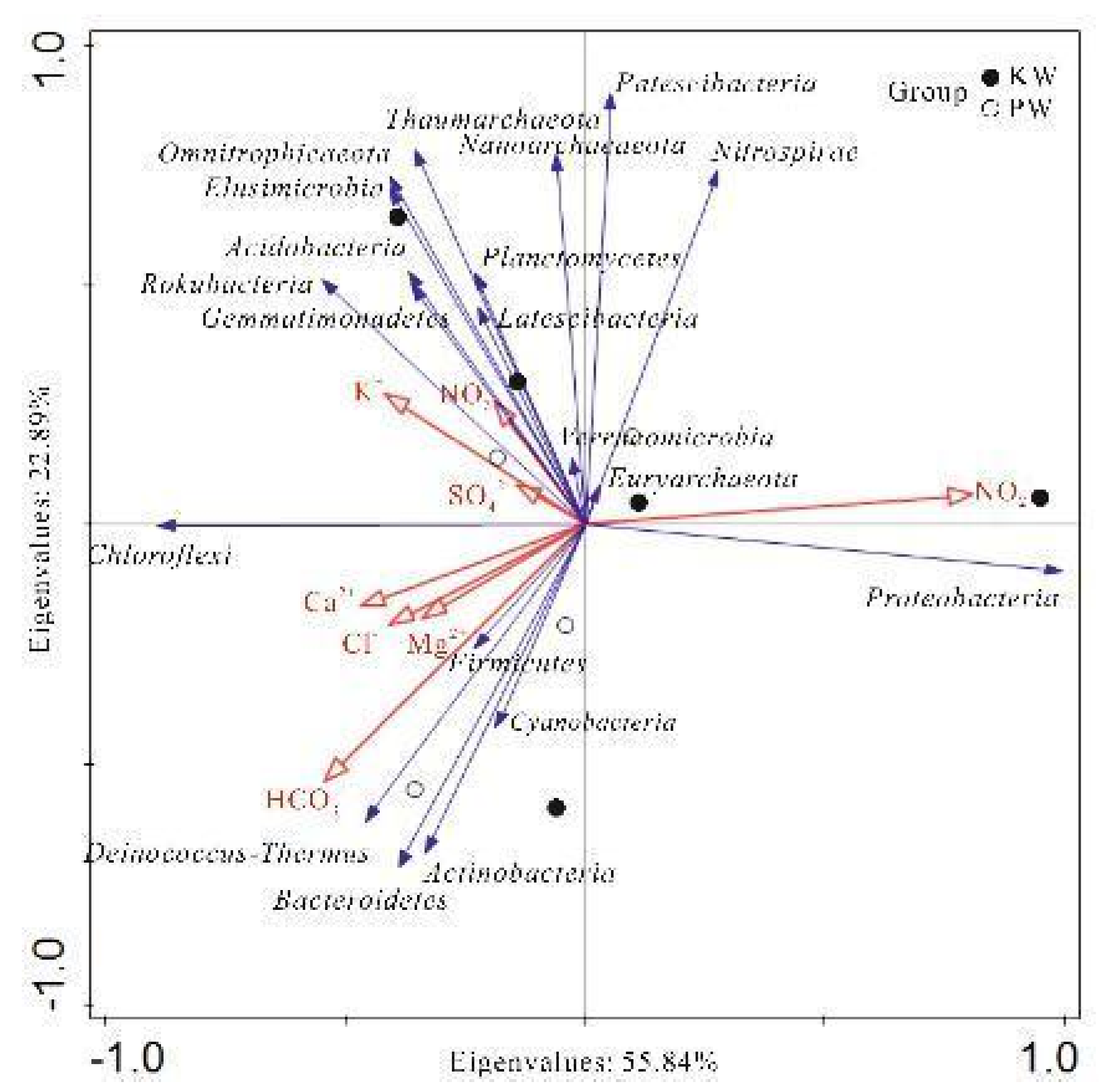

Table 1.
Hydrochemical characteristics and major ion contents of groundwater in the Yimuquan area.
| pH | Ec (μs/cm) |
ORP (mv) |
DO (mg/L) |
TDS (mg/L) |
K+ (mg/L) |
Na+ (mg/L) |
Ca2+ (mg/L) |
Mg2+ (mg/L) |
Cl- (mg/L) |
SO42- (mg/L) |
HCO3- (mg/L) |
NO3- (mg/L) |
NO2- (mg/L) |
F- (mg/L) |
H2SiO3 (mg/L) |
||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Karst Water (n=5) |
Avg | 7.72 | 863.8 | 132.36 | 8.976 | 405.23 | 1.62 | 21.33 | 87.86 | 45.13 | 46.99 | 46.21 | 316.30 | 68.16 | 0.01 | 0.17 | 13.63 |
| Max | 7.83 | 1032 | 163.8 | 9.9 | 487.21 | 3.44 | 35.75 | 106.92 | 54.79 | 74.87 | 71.07 | 344.68 | 109.75 | 0.04 | 0.30 | 15.56 | |
| Min | 7.49 | 550 | 96.2 | 7.46 | 277.03 | 0.84 | 8.58 | 64.10 | 28.40 | 11.73 | 8.96 | 300.23 | 8.64 | 0.00 | 0.11 | 12.35 | |
| Pore Water (n=4) |
Avg | 7.70 | 812.75 | 166.1 | 6.92 | 413.69 | 1.22 | 17.24 | 93.64 | 43.75 | 70.67 | 18.36 | 362.19 | 26.42 | 0.01 | 0.17 | 18.29 |
| Max | 7.88 | 1129 | 190.3 | 9.09 | 572.47 | 1.56 | 29.26 | 135.75 | 56.96 | 139.40 | 22.59 | 418.98 | 41.04 | 0.02 | 0.25 | 23.96 | |
| Min | 7.44 | 581 | 128.8 | 5.18 | 275.33 | 0.86 | 9.95 | 56.36 | 32.68 | 12.66 | 11.93 | 310.58 | 10.65 | 0.00 | 0.14 | 14.85 |
Note: ORP: Oxidation-Reduction Potential; DO: dissolved oxygen.; TDS: total dissolved solids
Table 2.
Changes in phylum niche in karst water and pore water.
| phylum | Karst water | Pore Water | ||
| Levins niche | Types of commonly used resources | Levins niche | Types of commonly used resources | |
| Omnitrophicaeota | 11.1427 | S5=11.32%,S6=10.62% | 8.6607 | S4=11.65%,S8=13.92%,S9=17.23% |
| Thaumarchaeota | 4.1453 | S1=34.59%,S3=17.67%,S5=25.14% | 6.0497 | S9=32.35% |
| Euryarchaeota | 3.3207 | S3=26.77%,S4=34.46%,S5=32.99% | 9.355 | S4=11.69%,S6=12.36%,S8=12.34% |
| Latescibacteria | 5.1979 | S2=32.32%,S4=18.00%,S8=14.49% | 3.1762 | S3=21.15%,S10=50.18% |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



