
Submitted:
12 December 2023
Posted:
12 December 2023
You are already at the latest version
Abstract
Mulberry (Morus alba L), a significant fruit tree crop, requires magnesium (Mg) for its optimal growth and productivity. Nonetheless, our understanding of the molecular basis underlying magnesium stress tolerance in mulberry plants remains unexplored. In our previous study, we identified several differential candidate genes associated with Mg homeostasis via transcriptome analysis including xyloglucan endotransglucosylase/hydrolase (XTH) gene family. The XTH gene family is crucial for plant cell wall reconstruction and stress responses. These genes have been identified and thoroughly investigated in various plant species. However, there is no research pertaining to XTH genes within the M. alba plant. This research systematically examined the M. alba XTH (MaXTH) gene family at the genomic level using a bioinformatic approach. In total, 22 MaXTH genes were discovered and contained the Glyco_hydro_16 and XET_C conserved domains. The MaXTH’s were categorized into five distinct groups by their phylogenetic relationships. The gene structure possesses four exons and three introns. Furthermore, the MaXTH gene promoter analysis reveals a plethora of cis-regulatory elements; mainly stress responsiveness, growth and development, light responsiveness, and phytohormone responsiveness. GO analysis indicated that MaXTHs encode proteins that exhibit xyloglucan xyloglucosyl transferase and hydrolase activities in addition cell wall biogenesis as well as xyloglucan and carbohydrate metabolic processes. Moreover, a synteny analysis unveiled an evolutionary relationship between the XTH genes in M. alba and those in three other species: A. thaliana, P. trichocarpa, and Zea mays. Expression profiles from RNA-Seq data displayed distinct expression patterns of XTH genes in M. alba leaf tissue during Mg treatments. Real-time quantitative PCR analysis confirmed the expression of the MaXTH genes in Mg stress response. Overall, this research enhances our understanding of the characteristics of MaXTH gene family members and lays the foundation for future functional genomic study in M. alba.
Keywords:
Morus alba
; xyloglucan endotransglucosylase/hydrolase (XTH)
; phylogeny
; magnesium stress
; expression patterns
1. Introduction
Mulberry (Morus spp.) is an essential plant in many Chinese provinces, primarily cultivated for its leaves and fruits. It holds particular significance in sericulture, as mulberry leaves serve as the exclusive food source for domestic silkworms (Bombyx mori L.) [1,2]. Besides its historical role in silkworm rearing, mulberry, especially Morus alba (M. alba), demonstrates potential as a pioneer tree species in marginal environments [3]. Moreover, the leaves of M. alba are of high medicinal value [4] and are believed to possess antioxidant, anti-inflammatory, and anti-allergic properties attributed to various bioactive phytochemicals, including polyphenolic compounds, triterpenoids, and anthocyanins. Although M. alba is of significant economic importance, its growth and development are subject to the influence of nutrient concentrations. High magnesium (Mg) levels or Mg deficiency are among the key factors affecting the growth and development of the plant [5]. Nevertheless, M. alba’s stress response to different Mg concentrations is unclear, especially at the genomic level.
Mg deficiency frequently hinders crop yield in sandy or highly acidic soils, primarily attributable to the high leaching susceptibility of Mg. This occurrence is widely observed and has notable implications for agricultural productivity in such soil conditions [6]. Extensive investigations have scrutinized and unveiled the consequences of Mg-deficiency on plant physiological aspects, including biomass distribution, carbon dioxide (CO2) uptake, protection against photooxidative stress, net CO2 uptake, and biomass distribution [7] resulting in yield reduction and poor fruit quality [8,9]. In response to these challenges, plants have developed intricate regulatory mechanisms, including the involvement of distinct gene families, such as Xyloglucan endotransglucosylase/hydrolases (XTHs) [10]. XTHs genes are classified within the glycoside hydrolase family 16 and are an essential group of enzymes primarily responsible for cleaving and rearranging the xyloglucan backbones within plant cell walls [11,12,13]. Specifically, family members of this gene carry out two distinct biochemical processes that are catalyzed by two specific enzymes: xyloglucan endotransglycosylase (XET) and xyloglucan endohydrolase (XEH) [14]. XET catalyzes the transfer of one xyloglucan molecule to another, resulting in the elongation of xyloglucan, whereas XEH is characterized by hydrolyzing an individual xyloglucan molecule, causing an irreversible reduction in the length of the xyloglucan chain [13].
Several XTHs exhibit both catalytic properties and play an essential role in regulating the extensibility of plant cell walls, root elongation, and plant growth [11,15]. Due to the advancement of sequencing technology and data availability, an expanding repertoire of XTH genes has been discovered and characterized in a broader range of species including Ananas comosus (48) [16], Arachis hypogaea L (58) [17], Glycine max (61) [18] Arabidopsis thaliana [19], lpomoea batatas (36) [20], Oryza sativa (29) [21], Solanum lycopersicum L (37) [11], Nicotiana tabacum (56) [22] Brassica rapa (53) and Brassica oleracea (35) [23]. Nonetheless, the XTH family constituents in mulberry remain undisclosed. Earlier investigations have demonstrated the involvement of XTH genes in numerous crucial processes, particularly the development and growth of plants via the remodeling of plant cell walls. For instance, in Arabidopsis, genes such as AtXTH17, AtXTH18, AtXTH19, and AtXTH20 exhibited specific expression patterns in the root tissues and were significantly involved in the processes of root elongation and the initiation of root hair formation [19,24]. GhXTH1 gene overexpressed in cotton, elongated cotton fibers by 15-20% [25]. In addition, some XTH genes have been reported to play active roles in fruit softening and ripening. PavXTH14 and PavXTH15 expression in cherry fruits resulted in a significant decrease in fruit firmness and altered the constitution of hemicellulose and pectin in the cell wall of the transgenic fruit [26]. Likewise, XTH influenced the softening and ripening of fruits, including tomatoes [27], strawberries [28], kiwi [29], and pears [29]. Several other XTH genes are involved in flower development [29] and leaves [30].
Numerous investigations have also suggested that plant hormones play a role in regulating the activity of XTH genes. For instance, the application of abscisic acid increased the expression of Arabidopsis AtXTH23 [19]. Similarly, the ethylene application induced the expression of banana MA-XETI, which is involved in the ripening and softening of the peel and pulp [30]. Furthermore, under ethylene induction, three CaXTH genes were significantly upregulated in the leaf tissue of hot pepper [31]. Members of the XTH gene family primarily regulate cell wall responses to biotic and abiotic stressors, which affect plant growth. The overexpression of DkXTHI was found to augment the resistance of transgenic Arabidopsis plants to salt, drought-induced stress, and abscisic acid, consequently impacting the development of roots and leaves [32]. Similarly, XTH genes in Chinese cabbage (Brassica rapa L) exhibited an up-regulated expression in response to elevated temperatures [33]. Furthermore, under low temperatures, the DkXTH6 gene in persimmons decreased in expression, while the DkXTH7 gene showed noticeably high transcription levels [34]. A prior proteomic study in maize revealed that XTHs were differently regulated in response to drought stress [35]. Moreover, xyloglucan content was decreased in the Arabidopsis AtXTH31 mutant, which lowered the amount of absorbed Al3+ and increased resistance to aluminum stress [36]. The overexpression of the xyloglucan endotransglucosylase/hydrolase gene in Populus euphratica resulted in increased resistance to cadmium tolerance by limiting cadmium absorption in the root system of transgenic tobacco plants. In addition, the transgenic plants had 56-87% more xyloglucan degrading activity (XDA) than the wild type, which resulted in a 25–27% decrease in the amount of xyloglucan in the root cell walls [37]. Moreover, in Arabidopsis, aluminum tolerance was imparted by the induction of ZmXTH, a gene encoding xyloglucan endotransglucosylase/hydrolase from maize [38]. Similarly, Arabidopsis mutants xth15 and xthI7 exhibited elevated aluminum tolerance in contrast to wild-type plants [36].
These preliminary studies highlight the key role of XTHs in various plants' response to various stresses. However, to the best of our knowledge, there is no functional characterization of Mulberry XTH gene family members. Consequently, there is a necessity for a systemic and comprehensive exploration of the M. alba XTH gene family across the genome. The present investigation conducted an analysis of the XTH gene family within M. alba based on our previous transcriptomic analysis after Mg stress treatment using the available genome data. Subsequently, detailed information, including phylogenetic analysis, gene structure characterization, chromosomal localization, motif analysis, promoter analysis, and syntenic relationships of MaXTH genes, were examined. Furthermore, Real-time quantitative PCR (qRT-PCR) was employed to determine the expression patterns of XTH genes in the leaf tissues of M. alba subjected to various levels of Mg stress. The findings of this study are poised to offer significant insights into the XTH genes in M. alba, contributing to a deeper comprehension and setting the groundwork for the functional analysis of plant XTH genes in mulberry plants.
2. Materials and Methods
2.1. Growth Conditions and Magnesium Treatment for the Mulberry Plant (Morus alba).
Mulberry (), a member of the M. alba (Yu-711) species, was obtained from the National Mulberry GenBank at Jiangsu University of Science and Technology in Zhenjiang, Jiangsu, China. Growth of the mulberry plant materials and Mg treatments followed the methods of our previous the [5,39]. On the 20th day, leaves from all experimental groups were collected from the 3-5 leaf position due to noticeable deficiency symptoms. These sampled leaves were temporarily preserved at a temperature of -80 ℃ for subsequent studies including transcriptome analysis.
2.2. Data Collection and Identification of XTH Gene Family Members in Morus alba
Genetic data in the form of genome sequences (fasta) and annotation files (gff) for three plant species, Arabidopsis thaliana, Populus trichocarpa, and Zea mays, were obtained from the official NCBI website (https://www.ncbi.nlm.nih.gov). We obtained the genome sequence and annotation file for M. alba with the help of Professor Weiguo Zhao of Jiangsu University of Science and Technology provided. To determine the XTH gene in the mulberry plant genome, a HMMER search was executed utilizing a hidden Markov model (HMM) profile comprising binding domains, PF00722 and PF06955, sourced from the Pfam database (http:// pfam. xfam. org/). Sequences with E values <1 was scrutinized, and any short open reading frames (less than 100 in length) were manually sorted out. The filtered sequences with putative PF00722 and PF06955 domains were screened as primary candidate for M. alba XTH genes (MaXTH).
2.3. Gene Structure, Motif Analysis and Sequence Alignment
Each MaXTH gene structure was visualized in TBtools software v1.098769 [40] by utilizing the genome sequence in conjunction with its corresponding annotation file. Employing the online MEME suite (available at https://meme-suite.org/meme/), the conserved motifs within MaXTHs were identified using specific parameters: a search for 10 motifs, with a minimum width of 6 and a maximum width of 55. Subsequently, an analysis of the consensus sequence was conducted, and a web logo was generated through the utilization of the MEME tool. The alignment of multiple MaXTH protein sequences was conducted utilizing the DNAMAN software.
2.4. Physicochemical Properties of the MaXTH Gene Family
The ExPASy online platform (https://web.expasy.org/protparam/) was used to obtain the molecular weight (Mw), isoelectric point (pI), and grand average of hydropathy (GRAVY) data for individual XTH proteins. Additionally, the subcellular localization of MaXTH proteins was predicted using the CELLO online resource (http://cello.life.nctu.edu.tw/) [41].
2.5. Phylogenetic Analysis
The evolutionary relationship of the XTH gene family among different species was assessed following the alignment of XTH protein sequences from M. alba, P. trichocarpa, and A. thaliana in MEGA 7 software using Clustal W [42]. A phylogenetic tree, based on the maximum likelihood method, with 1000 bootstrap replications was generated employing the MEGA 7.0 software. All other phylogeny trees were performed using MEGA 7 software following the same specifications indicated above.
2.6. Analysis of Cis-Regulatory Elements of MaXTH Genes and GO Analysis
The promoter regions located 2000 base pairs prior to the initiation codon (ATG) of MaXTH genes were extracted from the genome of M. alba. Utilizing the PlantCARE online tool (https://bioinformatics.psb.ugent.be/webtools/plantcare/html/) [43], the inherent regulatory elements in these promoter regions were predicted. Subsequently, the outcomes of this predictive analysis were rendered visible through the TBtool software. Gene Ontology (GO) analysis was performed using the web tool Shiny GO (http://bioinformatics.sdstate.edu/go74/).
2.7. Chromosomal Localization, Circos, and Synteny Analyses
The length of each chromosome and the chromosomal position of all XTH genes were obtained from the annotated dataset of the M. alba genome. The positional mapping of genes on the chromosomes was visualized through the utilization of TBtools software v1.098769 [40]. Based on the alignment and further examination of the phylogenetic relationship of the MaXTH genes, paralogous genes were discovered [20], and the interconnections of the paralogous genes according to their locations on the chromosomes were exhibited on the circos map constructed in the TBtools. The Multiple Collinearity Scan toolkit (MCScanX), integrated into the TBtool, was applied for the identification of syntenic blocks and specific gene pairs, adhering to its default configurations [44]. Additionally, the Multiple Synteny Plotter program within the TBtool (accessible at https://github.com/CJ-Chen/TBtools) was employed to visually represent the synteny associations among orthologous XTH genes across M. alba, A. thaliana, Zea mays, and Populus trichocarp [45].
2.8. Gene Expression Analysis and qRT-PCR Gene Validation
By using the RNA-seq data obtained from our transcriptome analysis after Mg application (https://ncbi.nlm.nih.gov/subs/sra; accession number: PRJNA951543), this study examined the expression of the XTH genes in M. alba treated with different levels of Mg after 20 days (about 3 weeks) and compared it to the expression of the gene in optimum Mg supply (the control) according to our previous study [5]. To validate the XTH genes identified through high-throughput sequencing, six differentially expressed genes (DEGs) were chosen for real-time quantitative polymerase chain reaction (qRT-PCR) validation, following the established protocol detailed in a prior study [46]. Mulberry leaf samples used for the RNA-seq analysis were also used for total RNA and cDNA synthesis for qRT-PCR validation. Furthermore, the 2−ΔΔCt method [47] was applied to estimate the fold changes in gene expression. The primer sequences and gene names for qRT-PCR validation are enlisted in the supplementary file S1.
3. Results
3.1. Identification and Physiological Features XTH Genes in M. alba
To identify potential candidate genes in response to high levels of magnesium (Mg) or magnesium nutrient starvation in M. alba, RNA-Seq transcriptome analysis for M. alba using the leaves after level of magnesium (Mg) treatments in a previous study (Data under journal consideration). A comprehensive examination of the transcriptome data resulted in the identification of 12 XTH genes. Among them, the candidate MaXTH-1 with the gene ID LOC21405692 had a higher expression level and, therefore, was selected and used as a query gene for the search against the Morus genome using BLASTP and HMMER search. 24 putative XTH proteins were obtained after the search and confirmed in the M. alba genome using their transcript ID. After which, the short and redundant sequences underwent manual scrutiny. Subsequently, the identification of conserved domains was carried out utilizing the Pfam and CDD databases. Finally, the 22 XTH proteins obtained were named MaXTH-1 to MaXTH-22 (Table 1 & Supplementary file S2). Coding sequences (CDS) of the 22 MaXTH proteins ranged from 645 bp (LOC21407360; MaXTH-21) to 1,509 bp (LOC21403517; MaXTH-18), with an average length of 915 bp. The amino acid sequence length of the MaXTH proteins spanned from 214-502, where MaXTH-18 was the longest sequence (501 aa) and MaXTH-21 (214 aa) had the shortest sequence. The molecular weights of MaXTH proteins varied between 24.071 kDa (MaXTH-21) and 56.843 kDa (MaXTH-18), averaging 34.40 kDa. The GRAVYs for all MaXTH genes were negative, ranging from -1003 to -0.099 (Supplementary file S2), suggesting that MaXTHs are likely hydrophilic. The predicted putative localization of the M. alba XTH was prevalent in the extracellular region, while a small number were located in the plasma membrane, mitochondrion, cytoplasmic, vacuole, and nuclear region (Table 1).
3.2. Gene Structure, Conserved Domain, and Motif Analysis
The structural diversity of the 22 MaXTH genes was investigated by determining their exon and intron structures through the alignment of their genomic and CDS sequences using the TBtool software. In addition, a phylogenetic tree was constructed using the complete MaXTH protein sequences, depicted with the distribution of introns and exons (Figure 1). The resulting phylogenetic analysis revealed the categorization of MaXTH genes into five distinct groups: I, II, III, IV and V constituting of 8, 9, 12, 6, and 7 MaXTH genes respectively (Figure 1a). In general, genes that are grouped together share similar structures. For instance, all members of group 1 (MaXTH-1, MaXTH-2, MaXTH-3, MaXTH-4, MaXTH-5, MaXTH-6, and MaXTH-9) contained three exons in their coding region, and two introns (Figure 1b). Members of group II also possessed four exons and three introns except for MaXTH-7 which had three exons. MaXTH group III members constituted four exons, excluding MaXTH-21 and MaXTH-22. Moreover, MaXTH-21 contained no intron. Interestingly, MaXTH-15 had no 5’ or 3’ UTRs.
To investigate the conserved domains of the 22 MaXTH genes, the Pfam and CDD databases were employed. From the results obtained, only four genes, MaXTH-9, MaXTH-18, MaXTH-21, and MaXTH-22, contained a single conserved domain (Glyco_hydro_16). In contrast, the remaining 16 MaXTH genes exhibited the presence of both Glyco_hydro_16 and XET_C conserved domains, as illustrated in Figure 2. MaXTH proteins were further characterized by predicting their potentially conserved motifs employing the MEME online software. A total of ten conserved motifs were identified, each exhibiting different lengths (Figure 3).
The longest motif (motif 1) had 50 amino acids, while the shortest (motif 8) constituted 6 amino acids (Figure 3). The motifs 1, 2, 3, 4, 5, 7 and 10 were present in glycohydro_16, (Figure 4) however not all proteins possessed motif 8. For instance, motifs 8 and 9 were present in the XET C domain. Multiple sequence alignments by DNAMAN software showed that all the MaXTH genes were highly conserved near potential N-linked glycosylation sites (Figure 5) between the amino acid sequences 126-272.
3.3. Evolutionary Relationship of the MaXTH Proteins
To comprehend the evolution and classification of MaXTH proteins, a phylogenetic tree based on the maximum likelihood method was constructed using the 22 MaXTH protein sequences with other XTH sequences from Arabidopsis thaliana (16) and Populus trichocarpa (15). From the results obtained, MaXTH proteins formed 5 different groups with XTH proteins from A. thaliana and P. trichocarpa (Figure 6). Groups I, II, III, IV and V contained 7, 8, 9, 5, and 6 members, respectively. Generally, the phylogenetic analysis depicts that MaXTH genes clustered well with XTH orthologs from P. trichocarpa and A. thaliana. However, the association between MaXTH and P. trichocarpa XTH proteins within group II exhibited a limited correlation, suggesting that the MaXTH proteins present in the branch might be less evolutionarily conserved or more primitive.
3.4. Cis-Acting Regulatory Elements of XTH Genes from Morus alba
To comprehend the transcriptional regulation of MaXTH genes, the 2000 bp promoter sequence of all the MaXTH genes was retrieved from the M. alba genome and analyzed using the PlantCare online database. The data obtained infer that MaXTH promoters possessed several cis-regulatory elements involved in processes such as stress response, hormone regulation, and cell development (Figure 7). Defense and stress-responsive cis-elements were identified for 10 MaXTH genes, including MaXTH-3, MaXTH-5, MaXTH-6, MaXTH-7, MaXTH-12, MaXTH-13, MaXTH-14, MaXTH-15, MaXTH-16 and MaXTH-21. In terms of hormone-responsive elements, 11, 14, 10, and 13 MaXTH genes contained salicylic acid-responsive elements, auxin-responsive elements, Methyl jasmonate (MeJa)-responsive regulatory elements, and gibberellin-responsive elements.
Drought-inducible response elements were detected in 13 MaXTH genes. The promoter sequences of two MaXTH genes constituting MaXTH-8 and MaXTH-18 exhibited cis-regulatory elements associated with wound responsiveness. Light-responsive elements were abundantly detected across all 22 MaXTH genes. Other MaXTH genes were identified to contain cell cycle response elements, endosperm response elements, and meristem expression regulatory elements associated with cell development. These results inferred that MaXTH genes participate in diverse biological processes and exhibit responsiveness to different biotic and abiotic stress factors.
Gene ontology (GO) analysis was conducted to elucidate the roles of MaXTH genes. The proteins encoded by MaXTH gene members exhibited both xyloglucan xyloglucosyl transferase and hydrolase activities (Figure 8A & supplementary file S3). In addition, MaXTH genes were observed to play crucial roles in various biological activities, including the organization of the cell wall, the biogenesis of the cell wall, and processes related to xyloglucan metabolism.
Further, GO analysis revealed that certain members of MaXTH genes were localized in the apoplast and cell wall regions. This observed localization aligns with the predictions made through subcellular localization analysis (Table 1). Notably, MaXTH genes are implicated in cellular glucan metabolic processes and carbohydrate metabolic processes. Additionally, the hierarchical clustering of the functional enrichment pathways depicted in Figure 8B reveals that MaXTH genes collaborate to perform various functions.
3.5. Chromosomal Localization, Circos, and Synteny Analyses
The chromosomal positions of XTH genes in M. alba were located through genome annotation in the TBtools. As depicted in Figure 9, the 22 MaXTH genes were unevenly distributed among the chromosomes across the genome of M. alba (Figure 9). The most significant MaXTH genes were positioned on chromosome one with 6 members. Three MaXTH genes were allotted on chromosomes 7, 9, and 10, and two on chromosome 14. Chromosomes 3, 4, 6, 12, and 13 contained one MaXTH each. Interestingly, no MaXTH genes were found in chromosomes 2, 5, 8 and 11 (Figure 9a). Based on the phylogenetic relationships of the MaXTH protein sequences (Figure 1 a), a total of five MaXTH gene pairs were identified as shown in the circos map (Figure 9b). It was discovered that three (MaXTH-10* and MaXTH-11*, MaXTH-16* and MaXTH-18*, MaXTH-19* and MaXTH-20*) and two pairs (MaXTH-9* and MaXTH-6*, MaXTH-2* and MaXTH-5*) of the MaXTHs gene pairs belonged to inter-chromosomal and intra chromosomal segments (Figure 9b). Furthermore, the gene pairs were associated to the same phylogenetic group. MaXTH-16* and MaXTH-18*, MaXTH-19* and MaXTH-20* belonged to group III, MaXTH-10* and MaXTH-11* were in group IV whereas MaXTH-9* and MaXTH-6*, MaXTH-2* and MaXTH-5* were affiliated to group I.
To delve deeper into the evolutionary correlations and genetic linkage occurrences within the XTH gene family, a systemic map of XTH genes across M. alba and three additional species, encompassing one monocotyledonous (Zea mays) and two dicotyledonous (Populus trichocarpa and Arabidopsis thaliana) plants were constructed using TBtools software (Figure 10).
According to the results of the collinearity analysis, 12 covariate pairs were generally discovered. Among them, 7 pairs were identified in Populus trichocarpa (represented in blue lines), 4 pairs in A. thaliana (represented in green lines), and only one pair was observed in Zea mays (represented in red lines). Furthermore, MaXTH-14 displayed a significant degree of collinearity with three comparable species (two in P. trichocarpa and one each in A. thaliana and Zea mays), inferring that MaXTH genes displayed greater evolutionary differences in P. trichocarpa in contrast to A. thaliana, and Zea mays.
3.6. Expression Profiling of MaXTH Genes under Different Magnesium Treatment and qRT-PCR Validation
Several studies have reported that the XTH gene family play an important role in plant response to abiotic stresses. To validate these accessions, the expression patterns of M. alba XTH genes responding to different treatments of magnesium concentrations were investigated by RNA-seq. RNA from M. alba leaf tissues was sampled on day 20. The XTH genes with expression changes according to the significant differential expression standard (|log2 (Fold change) |≥1 and false discovery rate (FDR) < 0.05) were analysed. The results from our investigation exhibited different expression levels of XTH genes at various concentrations (Figure 11 A-F). At the least concentration of 0 mM/L (T1), 10 MaXTHs were identified, among which six were significantly downregulated. Four XTHs genes including LOC21410403, LOC21405693, and LOC21401284 showed higher expression concentrations (Figure 11A).
In the 1 mM/L (T2), we detected 14 MaXTHs of which LOC21462237 was upregulated and had the highest expression level (Figure 11B). In contrast, the remaining 3 XTH genes showed were downregulated expression (LOC21390860, LOC21404346 and LOC21405696). M. alba treatment with 2 mM/L (T3) revealed 10 MaXTHs (Figure 11C). For 6 mM/L treatments (T4), it was observed that 3 XTHs (LOC21404346, LOC21404262 and LOC21407360) were low in expression (Figure 11D). In contrast, four XTHs (LOC21410403, LOC21405693, LOC210401284 and LOC21405696) were upregulated (Figure 11D). At the excess Mg of 9 mM/l (T5), all 10 MaXTH genes identified were significantly up-regulated (Figure 11E) with LOC21387254 and LOC21404346 being down-regulated.
Furthermore, six MaXTH genes including MaXTH-17 (LOC21410403), MaXTH-13 (LOC21401284), MaXTH-21 (LOC21407360), MaXTH-1 (LOC21405692), MaXTH-6 (LOC21404263) and MaXTH-10 (LOC21404346) in response to Mg stresses were selected to verify their relative expression level in mulberry leaves through qRT-PCR analysis. The results reveal that the selected genes could be expressed in the mulberry leaves, proving the reliability of the XTH genes identified by the transcriptome data (Figure 12A-E). LOC21410403 (MaXTH-17) exhibited a low expression level at 3 mM/L (CK; optimum concentration for M. alba growth); however, was highly expressed at an elevated concentration of 6 mM/L (Figure 12A). For LOC21401284 (MaXTH 13), the highest expression level was observed at Mg deficiency (0 mM/L) (Figure 12B). LOC21407360 (MaXTH-21) was moderately expressed at 0 mM/L, but was highly expressed in, 3 and 9 mM/L (Figure 12C). The expression of LOC21404263 (MaXTH-6) was highly expressed in 0. 3 and 9 mM/L Mg treatments (Figure 12D). For the LOC21405692 (MaXTH-10), the highest expression level was observed at 3mmol/l and the expression level was reduced at 6 mM/L concentration (Figure 12E). Finally, the expression of LOC21404346 (MaXTH-1) was higher in 0 and 2 mM/L concentrations (Figure 12E). Altogether, our findings confirm that XTH genes family are highly present in M. alba genome and expresses in M. alba leaf tissues in response to Mg imbalances. This outcome strongly suggests that XTH genes family are important in M. alba development and nutritional regulation.
4. Discussion
Mulberry (M. alba) is a plant of considerable economic importance, yet its growth and development are influenced by various abiotic factors such as Mg deficits [5]. Mg serves diverse functions in biological systems [48]. Consequently, gaining insights into how plants respond to both Mg deficiency and excess at the genomic level is essential for effective plant nutrient management. Past research indicates that plants have evolved sophisticated regulatory mechanisms, engaging specific gene families like xyloglucan endotransglucosylase/hydrolases (XTHs) [10] to facilitate their adaptation to Mg stress.
XTHs represent a category of plant enzymes responsible for regulating xyloglucan crosslinking within cell walls, playing a pivotal role in the control of plant growth and development [49,50]. The role of XTH genes is not only limited to cell wall elongation but also plays a part in plant responses to various environmental stresses. The XTH gene family has been identified across diverse plant species, such as A. thaliana [51], wheat [52], grapevine [53], rice [21], peanut [17], barley [54], sweet potato [20] and poplar [55]. Within the scope of this investigation, we present the discovery and characterization of the XTH gene family within the M. alba genome. This includes exploring their phylogenetic relationships, conserved motifs, gene structures, cis-acting regulatory elements, and gene expression patterns in response to Mg starvation, low or high treatments.
Based on the M. alba genome, 22 XTH genes were identified based on our strictest identification workflow and labelled as MaXTH-1 to MaXTH-22. The number of identified XTH genes was notably less compared to various other species, including tobacco (56), wheat (71), Solanum lycopersicum (37), and Glycine max (61) [11,18,22,52]. It is widely acknowledged that the functional attributes of genes are intricately linked to their structural and physicochemical characteristics [20,56]. In this study, the 22 MaXTH protein members displayed significant disparities with respect to protein sequence length, molecular weight, isoelectric point (pI), intron and exon distributions (Table 1). This variation implicates a high diversity among XTH family members in M. alba. Additionally, most MaXTH genes were predicted to be in the extracellular space, while a few were in the plasma membrane, vacuole, mitochondrion, and nuclear region. This is contrary to previous reports for other XTH protein members in other plant species, where the majority of the XTH proteins were in the plasma membrane rather than the extracellular space and other locations [22,53,55]. Further, phylogenetic analysis indicated that MaXTH protein families were clustered into five groups (Figure 1), similarly observed for XTH proteins from sweet cherry [26]. Interestingly, the MaXTH proteins belonging to the same group demonstrated similar gene structures (Figure 1) and conserved sequence expression which is consistent with previously documented literature [11,20], suggesting that XTH members within the same group may exhibit analogous functionalities. Moreover, most of the MaXTH genes demonstrate the presence of two main conserved domains (Glyco_hydro_16 and XET_C domain) (Figure 2). Nevertheless, MaXTH-9, MaXTH-18, MaXTH-21, and MaXTH-22 lacked the XET_C domain. This absence suggests a potential evolutionary divergence, indicating a loss of the XET_C domain during the evolutionary trajectory of XTH proteins in M. alba.
Phylogenetic distribution of XTH proteins from M. alba, A. thaliana, and P. trichocarpa revealed that MaXTH genes could be categorized into five groups (group I-V) (Figure 5). Earlier studies have documented the categorization of XTH gene families into distinct groups in various plant species. In tobacco, for instance, eight family groups were identified [22], while three groups were observed in peanut [17], barley [54], and sweet potato [20]. Poplar, on the other hand, exhibited four distinct groups [55]. The MaXTH genes were observed to cluster better with XTH proteins from P. trichocarpa than A. thaliana, implying a closer evolutionary relationship between XTH proteins in M. alba and those of P. trichocarpa rather than A. thaliana. According to chromosomal localization analysis, it was observed that MaXTHs were heterogeneously distributed on 10 out of the 14 chromosomes of M. alba (Figure 9). Further investigation revealed five gene pairs among the XTH gene families in the M. alba genome. Previous research has indicated that a set of gene functions exhibit high conservation across various plant species [57,58]. Consequently, it is imperative to identify true orthologs in different plant species through the application of synteny analysis. The results obtained from the synteny analysis depicted a significant degree of synteny between the M. alba genome and those of P. trichocarpa and A. thaliana, exhibiting 7 and 4 synthetic blocks of MaXTH between P. trichocarpa and A. thaliana, respectively. In contrast, one synthetic block was identified between Zea mays (Figure 10).
Cis-regulatory elements are essential for regulating gene expression. The comprehension of cis-regulatory elements within the promoter region of genes has the potential to clarify the roles and regulatory mechanisms of specific genes that engage in collaborative interactions with other genes [59,60]. Investigating the cis-regulatory elements of the 22 MaXTH exhibited a quantity of core promoters involved in light responsiveness, hormone responsiveness (Abscisic acid, Salicylic acid, MeJA, Gibberellin), stress responsiveness (temperature, drought, low-defense, anoxic, and stress), growth and development elements (Figure 7). MaXTH promoters contain a variety of elements that respond to environmental and plant hormone stimuli, which might indicate various regulatory or functional mechanisms in response to biotic and abiotic stress factors [20,61]. Besides, there were significant variations in terms of type and quantity and certain elements related to metabolism, and gene expression were unique to specific MaXTH genes. The structural variations of MaXTH proteins could result in modifying protein functions. Several studies have demonstrated that plant XTH proteins have essential roles in plant growth, development, and stress resistance. The presence of numerous cis-elements identified in the promoter region of the M. alba XTH genes suggests that the XTH genes within M. alba possess the capability to adapt to diverse modifications in the plant, particularly responsiveness to light, several hormones, and numerous stress response elements (anaerobic and anoxic specific inducibility).
Analyzing gene expression profiles can advance our understanding of XTHs functions in M. alba growth and development. Analysis of transcriptome data at day 20 after the various magnesium treatments indicated that several XTHs were expressed in response to the treatments (Figure 11A-F). XTH genes, including MaXTH-17(LOC21410403), MaXTH-13(LOC21401284), MaXTH-21(LOC21407360), MaXTH-6 (LOC21404263) and MaXTH-10 (LOC21404346) highly expressed at 0, 1, 2, 6, and 9 mM/L of Mg concentration, respectively (Figure 10A-F) compared to control (3 mM/L, optimum Mg for M. alba growth). Meanwhile, MaXTH-6 (LOC21404263) was downregulated at 2 and 6 mM/L while MaXTH-1 (LOC21405692) exhibited low expression at 6 mM/L. Prior findings indicate that abiotic stressors can induce transcript-level changes in XTH genes. For example, in response to cadmium (Cd) stress, the expression of BnXTH1, BnXTH3, BnXTH6, and BnXTH15 was observed to be upregulated in Boehmeria nivea. Conversely, BnXTH18, BnXTH16, BnXTH17, and BnXTH5 exhibited notable down-regulation under the same Cd stress conditions [62].
Similar contrasting expression patterns of the XTH gene family were identified in Camellia sinensis under fluorine stress where CsXTH7, CsXTH1, CsXTH6, and CsXTH1 were up-regulated, while that of CsXTH3 was down-regulated [63]. Additionally, the expression of PeXTH experienced a notable up-regulation in the roots and leaves of P. euphratica when exposed to Cd stress [37]. Likewise, under Al stress, AtXTH15 and AtXTH14 demonstrated a significant decrease, leading to a reduction in xyloglucan endo transferase (XET) activity and consequently enhancing the aluminum tolerance of A. thaliana [64]. In this study, we observed different expression patterns of MaXTH genes. Changes in the expression of MaXTH genes can affect cell wall flexibility and strength, which are important factors in stress adaptation. The increased expression of certain MaXTH genes might contribute to cell wall remodeling, allowing for better flexibility and adaptation to magnesium stress. Conversely, decreased expression could be associated with a more rigid cell wall structure. These findings indicate the capacity of MaXTH genes to provide defense to the M. alba plant during magnesium starvation, undersupply and excess application. Future works in M. alba should investigate the functional genomic validation of these identified XTH genes and how they regulate Mg nutrition.
Taken together, the results of this research offer novel insights into MaXTH genes under abiotic stress, particularly when subjected to different Mg concentrations. It could be inferred that the MaXTHs might exhibit heightened functionalities related to the cell wall in stressful conditions through interaction with xyloglucan. However, additional molecular and genetic research are required to confirm their roles.
5. Conclusions
In this current investigation, an extensive examination of the M. alba XTH gene family was conducted. The results from the investigation successfully identified and further characterized a total of 22 MaXTH genes. These genes were subsequently categorized into five groups (I to IV) based on their phylogenetic relationships. Gene structure and motif composition were observed to be consistent within each group. A thorough analysis of gene synteny uncovered evidence of evolutionary relationships among XTH genes in M. alba and three other species, A. thaliana, P. trichocarpa, and Zea mays. Gene ontology analysis revealed that MaXTHs are responsible for encoding proteins demonstrating both xyloglucan xyloglucosyl transferase and hydrolase activities. Moreover, MaXTHs actively contribute to the processes of cell wall biogenesis, as well as the metabolic pathways associated with xyloglucan and carbohydrates. Furthermore, specific Cis-acting regulatory elements detected in the promoter region of MaXTH genes suggest their potential involvement in various biological processes such as development, light responsiveness, phytohormone responses, and stress adaptation. Moreover, investigating the expression profiles of MaXTH genes within leaf tissues exposed to different magnesium concentrations revealed diverse patterns of gene expression. Collectively, the findings from this research provide valuable insights into the functions of XTH genes within M. alba and present a better understanding of how mulberry plants respond to various magnesium treatments.
Supplementary Materials
The supplementary data to this article are present as supplementary materials.
Author Contributions
Weiguo Zhao, Michael Ackah and Blessing Danso: conceived and designed the research. Weiguo Zhao: directed the entire research project and provided funding. Michael Ackah and Blessing Danso: Performed bioinformatics data analysis and manuscripts figures. Xin Jin performed the qRT-PCR experimentation. Derek M. Ayittey, and Frank Kwarteng Amoako: helped in data analyses and manuscript figures preparation. Michael Ackah and Blessing Danso wrote the manuscript draft. Michael Ackah, Blessing Danso, Derek M. Ayittey and Weiguo Zhao: edited the final manuscript. All authors read and approved the final manuscript submission.
Funding
This work was supported by the Key R&D Program of Guangxi (AB23026066), earmarked fund for CARS-18, National Key R&D Program of China (2021YFE0111100), Science and Technology Partnership Program (KY202201002), Ministry of Science and Technology of China, Zhenjiang Science and Technology support project (GJ2021015), the Crop Germplasm Resources Protection Project of the Agriculture Ministry (111721301354052026), and National Infrastructure for Crop Germplasm Resources (NICGR-43).
Institutional Review Board Statement
Not applicable.
Declaration of competing interest
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
References
- Buhroo, Z.I., et al., Trends in development and utilization of sericulture resources for diversification and value addition. International Journal of Entomological Research, 2018. 6(1): p. 27-47. [CrossRef]
- Chauhan, T. and M.K. Tayal, Mulberry sericulture. Industrial entomology, 2017: p. 197-263.
- Zeng, P., et al., Physiological stress responses, mineral element uptake and phytoremediation potential of Morus alba L. in cadmium-contaminated soil. Ecotoxicology and environmental safety, 2020. 189: p. 109973. [CrossRef]
- Zhang, R., et al., Mulberry leaf (Morus alba L.): A review of its potential influences in mechanisms of action on metabolic diseases. Pharmacological Research, 2022. 175: p. 106029. [CrossRef]
- Jin, X., et al., Magnesium Nutrient Application Induces Metabolomics and Physiological Responses in Mulberry (Morus alba) Plants. International Journal of Molecular Sciences, 2023. 24(11): p. 9650. [CrossRef]
- Aitken, R., et al., Response of field-grown maize to applied magnesium in acidic soils in north-eastern Australia. Australian Journal of Agricultural Research, 1999. 50(2): p. 191-198. [CrossRef]
- Ogura, T., et al., Short-Term Magnesium Deficiency Triggers Nutrient Retranslocation in Arabidopsis thaliana. Front Plant Sci, 2020. 11: p. 563. [CrossRef]
- Gerendás, J. and H. Führs, The significance of magnesium for crop quality. Plant and Soil, 2013. 368: p. 101-128. [CrossRef]
- Guo, W., et al., Magnesium deficiency in plants: an urgent problem. Crop J 4: 83–91. 2016. [CrossRef]
- Yang, L.-T., et al., Magnesium deficiency induced global transcriptome change in Citrus sinensis leaves revealed by RNA-Seq. International journal of molecular sciences, 2019. 20(13): p. 3129. [CrossRef]
- Sarmiento-López, L.G., et al., Genome-wide characterization of the xyloglucan endotransglucosylase/hydrolase gene family in Solanum lycopersicum L. and gene expression analysis in response to arbuscular mycorrhizal symbiosis. PeerJ, 2023. 11: p. e15257.
- Pauly, M. and K. Keegstra, Biosynthesis of the plant cell wall matrix polysaccharide xyloglucan. Annual review of plant biology, 2016. 67: p. 235-259. [CrossRef]
- Eklöf, J.M. and H. Brumer, The XTH gene family: an update on enzyme structure, function, and phylogeny in xyloglucan remodeling. Plant physiology, 2010. 153(2): p. 456-466.
- Li, X., et al., Identification and analysis of the xyloglucan endotransferase/hydrolase (XTH) family genes in apple. Scientia Horticulturae, 2023. 315: p. 111990. [CrossRef]
- Stratilová, B., et al., Plant xyloglucan xyloglucosyl transferases and the cell wall structure: subtle but significant. Molecules, 2020. 25(23): p. 5619. [CrossRef]
- Li, Q., et al., Genome-wide identification and characterization of xyloglucan endotransglycosylase/hydrolase in Ananas comosus during Development. Genes, 2019. 10(7): p. 537. [CrossRef]
- Zhu, J., et al., Genome-wide identification of xyloglucan endotransglucosylase/hydrolase gene family members in peanut and their expression profiles during seed germination. PeerJ, 2022. 10: p. e13428. [CrossRef]
- Song, L., et al., Characterization of the XTH gene family: new insight to the roles in soybean flooding tolerance. International Journal of Molecular Sciences, 2018. 19(9): p. 2705. [CrossRef]
- Yokoyama, R. and K. Nishitani, A comprehensive expression analysis of all members of a gene family encoding cell-wall enzymes allowed us to predict cis-regulatory regions involved in cell-wall construction in specific organs of Arabidopsis. Plant and cell physiology, 2001. 42(10): p. 1025-1033. [CrossRef]
- Zhang, J.-Z., et al., Genome-wide identification and expression analysis of the xyloglucan endotransglucosylase/hydrolase gene family in sweet potato [Ipomoea batatas (L.) Lam]. International Journal of Molecular Sciences, 2023. 24(1): p. 775. [CrossRef]
- Yokoyama, R., J.K. Rose, and K. Nishitani, A surprising diversity and abundance of xyloglucan endotransglucosylase/hydrolases in rice. Classification and expression analysis. Plant Physiology, 2004. 134(3): p. 1088-1099. [CrossRef]
- Wang, M., et al., Genome-wide identification and expression profiling analysis of the xyloglucan endotransglucosylase/hydrolase gene family in tobacco (Nicotiana tabacum L.). Genes, 2018. 9(6): p. 273. [CrossRef]
- Wu, D., et al., Genome-wide identification, and phylogenetic and expression profiling analyses, of XTH gene families in Brassica rapa L. and Brassica oleracea L. Bmc Genomics, 2020. 21: p. 1-17. [CrossRef]
- Osato, Y., R. Yokoyama, and K. Nishitani, A principal role for AtXTH18 in Arabidopsis thaliana root growth: a functional analysis using RNAi plants. Journal of plant research, 2006. 119: p. 153-162. [CrossRef]
- Lee, J., et al., Xyloglucan endotransglycosylase/hydrolase genes in cotton and their role in fiber elongation. Planta, 2010. 232: p. 1191-1205. [CrossRef]
- Zhai, Z., et al., Genome-wide identification of the xyloglucan endotransglucosylase/hydrolase (XTH) and polygalacturonase (PG) genes and characterization of their role in fruit softening of sweet cherry. International Journal of Molecular Sciences, 2021. 22(22): p. 12331. [CrossRef]
- Miedes, E. and E.P. Lorences, Xyloglucan endotransglucosylase/hydrolases (XTHs) during tomato fruit growth and ripening. Journal of plant physiology, 2009. 166(5): p. 489-498. [CrossRef]
- Witasari, L.D., et al., Higher expression of the strawberry xyloglucan endotransglucosylase/hydrolase genes Fv XTH 9 and Fv XTH 6 accelerates fruit ripening. The Plant Journal, 2019. 100(6): p. 1237-1253.
- Atkinson, R.G., et al., Analysis of xyloglucan endotransglucosylase/hydrolase (XTH) gene families in kiwifruit and apple. Postharvest Biology and Technology, 2009. 51(2): p. 149-157. [CrossRef]
- Lu, W.J., et al., Cloning and expression analysis of an XET cDNA in the peel and pulp of banana fruit ripening and softening. Acta Botanica Sinica, 2004. 46(3): p. 355-362.
- Cho, S.K., et al., Constitutive expression of abiotic stress-inducible hot pepper CaXTH3, which encodes a xyloglucan endotransglucosylase/hydrolase homolog, improves drought and salt tolerance in transgenic Arabidopsis plants. FEBS letters, 2006. 580(13): p. 3136-3144.
- Han, Y., et al., Overexpression of persimmon DkXTH1 enhanced tolerance to abiotic stress and delayed fruit softening in transgenic plants. Plant cell reports, 2017. 36: p. 583-596. [CrossRef]
- Yang, K.A., et al., Identification of cell wall genes modified by a permissive high temperature in Chinese cabbage. Plant science, 2006. 171(1): p. 175-182. [CrossRef]
- Han, Y., et al., Isolation and characterization of two persimmon xyloglucan endotransglycosylase/hydrolase (XTH) genes that have divergent functions in cell wall modification and fruit postharvest softening. Frontiers in Plant Science, 2016. 7: p. 624. [CrossRef]
- Zhu, J., et al., Cell wall proteome in the maize primary root elongation zone. II. Region-specific changes in water soluble and lightly ionically bound proteins under water deficit. Plant physiology, 2007. 145(4): p. 1533-1548. [CrossRef]
- Zhu, X.F., et al., XTH31, encoding an in vitro XEH/XET-active enzyme, regulates aluminum sensitivity by modulating in vivo XET action, cell wall xyloglucan content, and aluminum binding capacity in Arabidopsis. The Plant Cell, 2012. 24(11): p. 4731-4747. [CrossRef]
- Han, Y., et al., Overexpression of Populus euphratica xyloglucan endotransglucosylase/hydrolase gene confers enhanced cadmium tolerance by the restriction of root cadmium uptake in transgenic tobacco. Environmental and experimental botany, 2014. 100: p. 74-83. [CrossRef]
- Du, H., et al., ZmXTH, a xyloglucan endotransglucosylase/hydrolase gene of maize, conferred aluminum tolerance in Arabidopsis. Journal of Plant Physiology, 2021. 266: p. 153520. [CrossRef]
- Adolf, A., et al., Transcriptome profiling reveals candidate genes associated with cold stress in mulberry. Brazilian Journal of Botany, 2021. 44: p. 125-137. [CrossRef]
- Chen, C., et al., TBtools: An Integrative Toolkit Developed for Interactive Analyses of Big Biological Data. Molecular Plant, 2020. 13(8): p. 1194-1202. [CrossRef]
- Yu, C.S., et al., Prediction of protein subcellular localization. Proteins: Structure, Function, and Bioinformatics, 2006. 64(3): p. 643-651.
- Tamura, K., et al., MEGA6: molecular evolutionary genetics analysis version 6.0. Molecular biology and evolution, 2013. 30(12): p. 2725-2729.
- Lescot, M., et al., PlantCARE, a database of plant cis-acting regulatory elements and a portal to tools for in silico analysis of promoter sequences. Nucleic acids research, 2002. 30(1): p. 325-327. [CrossRef]
- Wang, Y., et al., MCScanX: a toolkit for detection and evolutionary analysis of gene synteny and collinearity. Nucleic acids research, 2012. 40(7): p. e49-e49. [CrossRef]
- Chen, C., et al., TBtools, a toolkit for biologists integrating various biological data handling tools with a user-friendly interface. BioRxiv, 2018. 289660(10.1101): p. 289660.
- Ackah, M., et al., DNA methylation changes and its associated genes in mulberry (Morus alba L.) Yu-711 response to drought stress using MethylRAD sequencing. Plants, 2022. 11(2): p. 190.
- Livak, K.J. and T.D. Schmittgen, Analysis of relative gene expression data using real-time quantitative PCR and the 2− ΔΔCT method. methods, 2001. 25(4): p. 402-408.
- Kwon, M.C., et al., Comparative metabolomics unravel the effect of magnesium oversupply on tomato fruit quality and associated plant metabolism. Metabolites, 2019. 9(10): p. 231. [CrossRef]
- Ishida, K. and R. Yokoyama, Reconsidering the function of the xyloglucan endotransglucosylase/hydrolase family. Journal of Plant Research, 2022. 135(2): p. 145-156. [CrossRef]
- Ma, X., et al., Genome-wide analysis of the XTH gene family and functional analysis of DlXTH23. 5/25 during early longan somatic embryogenesis. Frontiers in Plant Science, 2022. 13: p. 1043464. [CrossRef]
- Maris, A., et al., Differences in enzymic properties of five recombinant xyloglucan endotransglucosylase/hydrolase (XTH) proteins of Arabidopsis thaliana. Journal of Experimental Botany, 2011. 62(1): p. 261-271. [CrossRef]
- Han, J., et al., A Surprising Diversity of Xyloglucan Endotransglucosylase/Hydrolase in Wheat: New in Sight to the Roles in Drought Tolerance. International Journal of Molecular Sciences, 2023. 24(12): p. 9886. [CrossRef]
- Qiao, T., et al., Identification and expression analysis of xyloglucan endotransglucosylase/hydrolase (XTH) family in grapevine (Vitis vinifera L.). PeerJ, 2022. 10: p. e13546.
- Fu, M.-M., C. Liu, and F. Wu, Genome-wide identification, characterization and expression analysis of xyloglucan endotransglucosylase/hydrolase genes family in barley (Hordeum vulgare). Molecules, 2019. 24(10): p. 1935.
- Cheng, Z., et al., Genome-wide identification and expression analysis of the xyloglucan endotransglucosylase/hydrolase gene family in poplar. BMC genomics, 2021. 22: p. 1-13. [CrossRef]
- Yang, Z., R. Zhang, and Z. Zhou, The XTH gene family in Schima superba: Genome-wide identification, expression profiles, and functional interaction network analysis. Frontiers in Plant Science, 2022. 13: p. 911761. [CrossRef]
- Du, H., et al., Genome-wide analysis of the MYB transcription factor superfamily in soybean. BMC plant biology, 2012. 12: p. 1-22. [CrossRef]
- Chen, C., et al., Genome-wide analysis of the WRKY gene family in the cucumber genome and transcriptome-wide identification of WRKY transcription factors that respond to biotic and abiotic stresses. BMC Plant Biology, 2020. 20: p. 1-19. [CrossRef]
- Biłas, R., et al., Cis-regulatory elements used to control gene expression in plants. Plant Cell, Tissue and Organ Culture (PCTOC), 2016. 127: p. 269-287. [CrossRef]
- Han, Y., et al., Genome-wide characterization and expression analysis of bZIP gene family under abiotic stress in Glycyrrhiza uralensis. Frontiers in Genetics, 2021. 12: p. 754237. [CrossRef]
- Xu, P., et al., The brassinosteroid-responsive xyloglucan endotransglucosylase/hydrolase 19 (XTH19) and XTH23 genes are involved in lateral root development under salt stress in Arabidopsis. The Plant Journal, 2020. 104(1): p. 59-75. [CrossRef]
- Ma, Y.-S., et al., Identification of the Xyloglucan Endotransglycosylase/Hydrolase (XTH) Gene Family Members Expressed in Boehmeria nivea in Response to Cadmium Stress. International Journal of Molecular Sciences, 2022. 23(24): p. 16104. [CrossRef]
- Wu, Z., et al., Identification and response analysis of xyloglucan endotransglycosylase/hydrolases (XTH) family to fluoride and aluminum treatment in Camellia sinensis. BMC genomics, 2021. 22(1): p. 1-16. [CrossRef]
- Yang, J.L., et al., Cell wall hemicellulose contributes significantly to aluminum adsorption and root growth in Arabidopsis. Plant physiology, 2011. 155(4): p. 1885-1892. [CrossRef]
Figure 1.
Phylogenetic relationship and gene structure of MaXTH gene family. (A) Phylogenetic tree of the 22 Morus alba XTH gene family (B) gene structure of the MaXTH genes. Pink color; CDS. Green color: UTR. I-V; MaXTH gene family classification.
Figure 1.
Phylogenetic relationship and gene structure of MaXTH gene family. (A) Phylogenetic tree of the 22 Morus alba XTH gene family (B) gene structure of the MaXTH genes. Pink color; CDS. Green color: UTR. I-V; MaXTH gene family classification.

Figure 2.
Domain analysis of the MaXTH gene family.
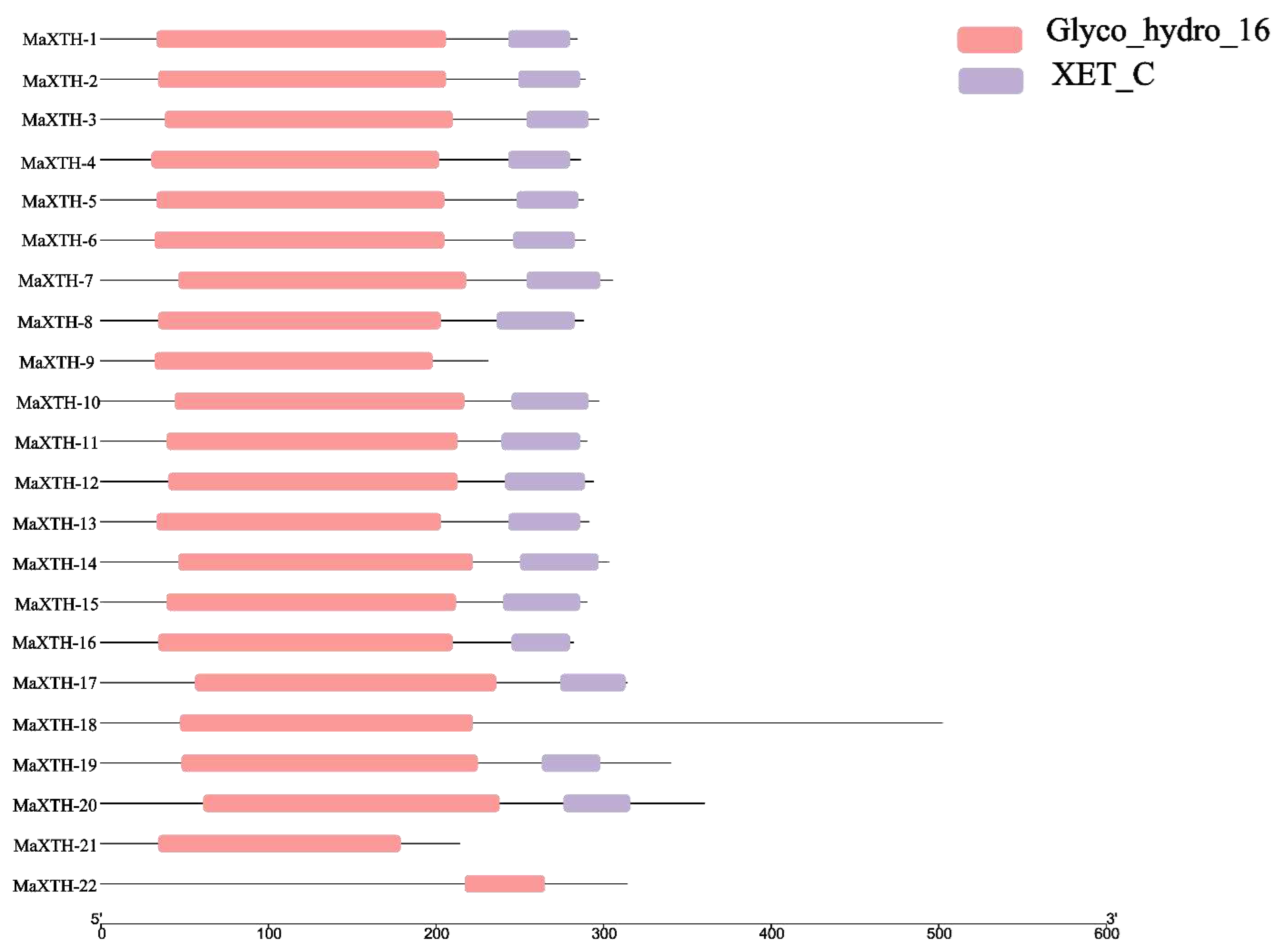
Figure 3.
Motifs sequence logo of MaXTH gene family.
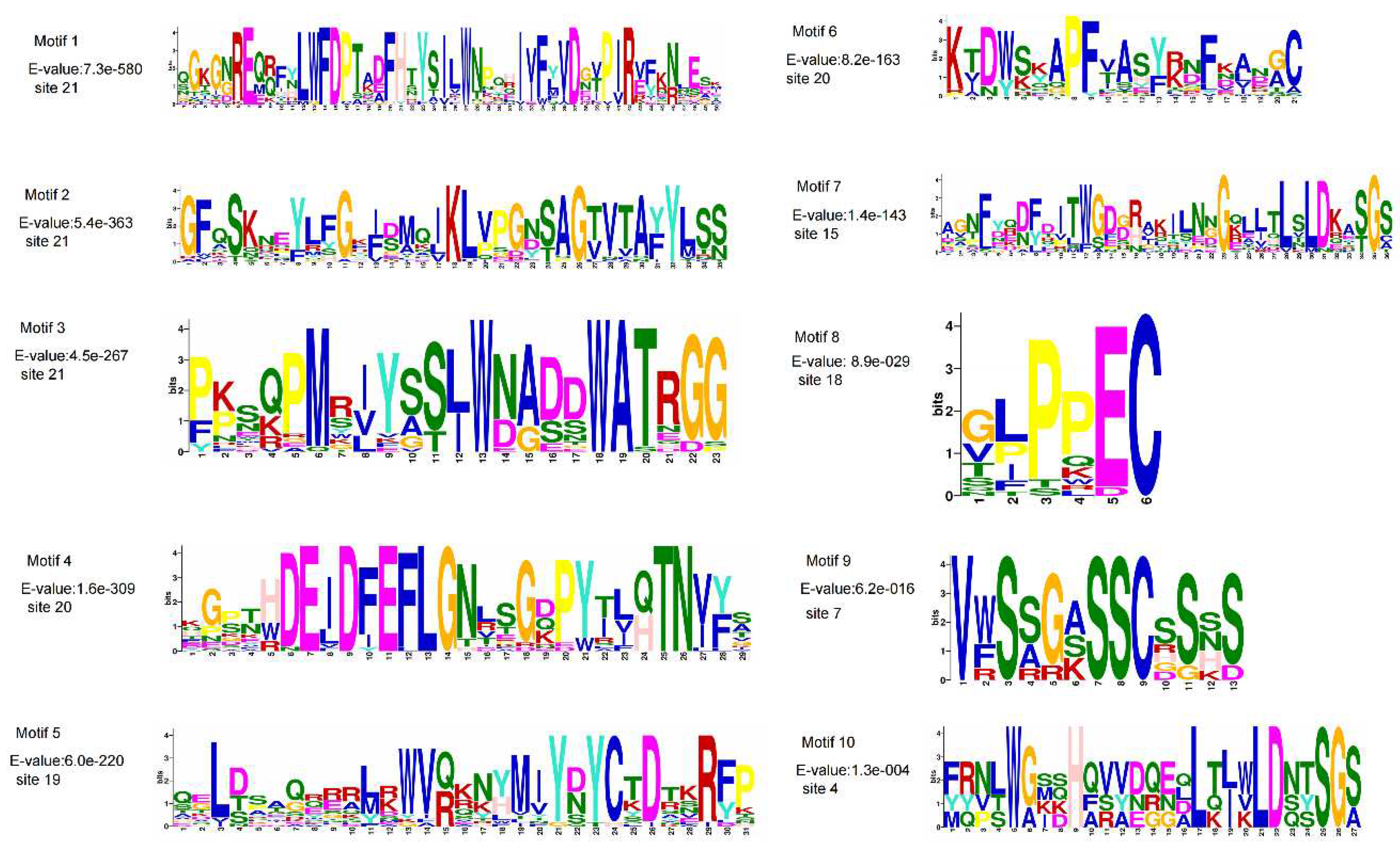
Figure 4.
Conserved Motifs of MaXTH gene family.
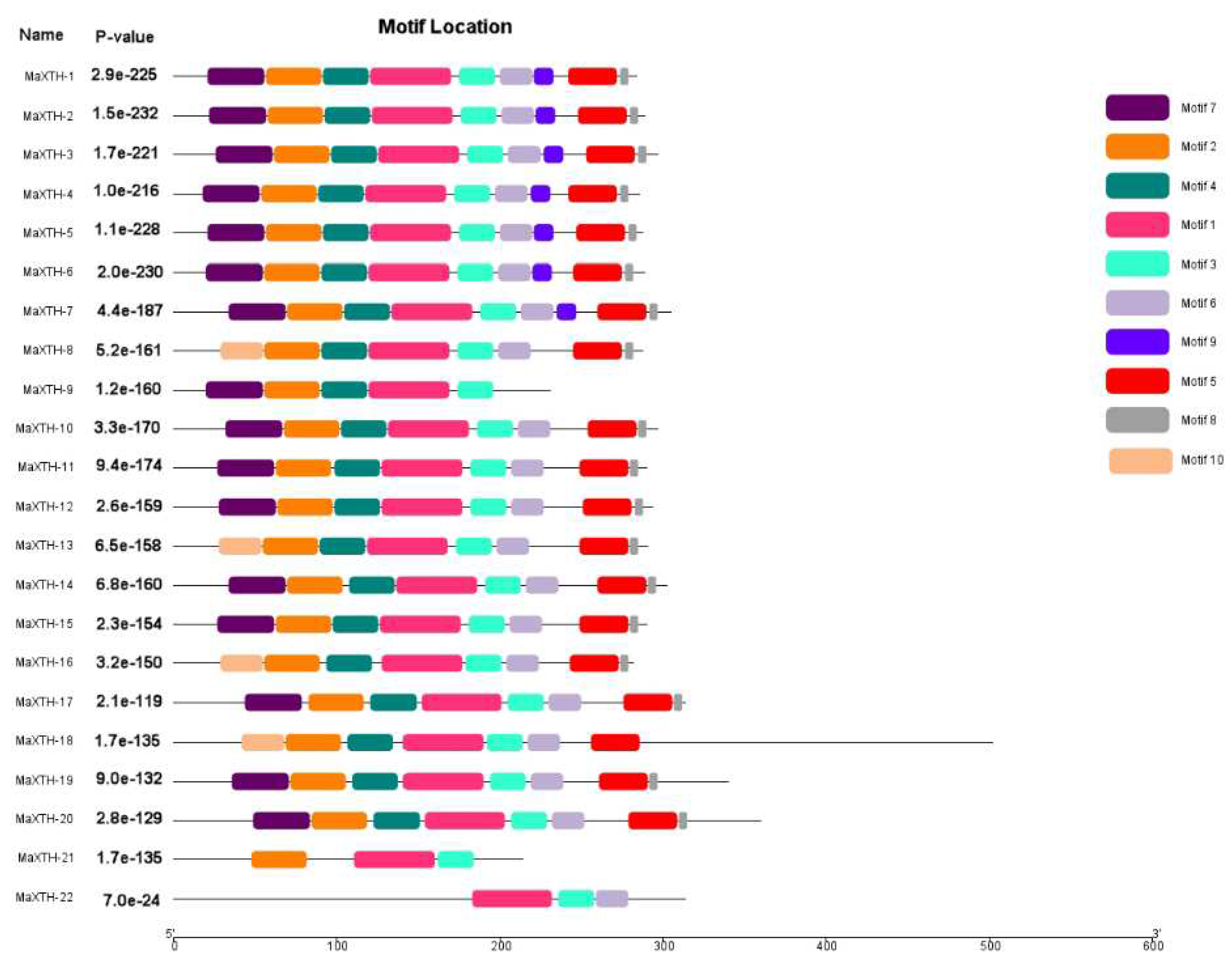
Figure 5.
Multiple sequence analysis of the 22 MaXTH family genes. Asterisks highlight the N-linked glycosylation sites.
Figure 5.
Multiple sequence analysis of the 22 MaXTH family genes. Asterisks highlight the N-linked glycosylation sites.

Figure 6.
Maximum likelihood Phylogenetic relations of XTH protein family in Morus alba (red circles), Populus trichocarpa (blue triangle), and Arabidopsis thaliana (green square).
Figure 6.
Maximum likelihood Phylogenetic relations of XTH protein family in Morus alba (red circles), Populus trichocarpa (blue triangle), and Arabidopsis thaliana (green square).
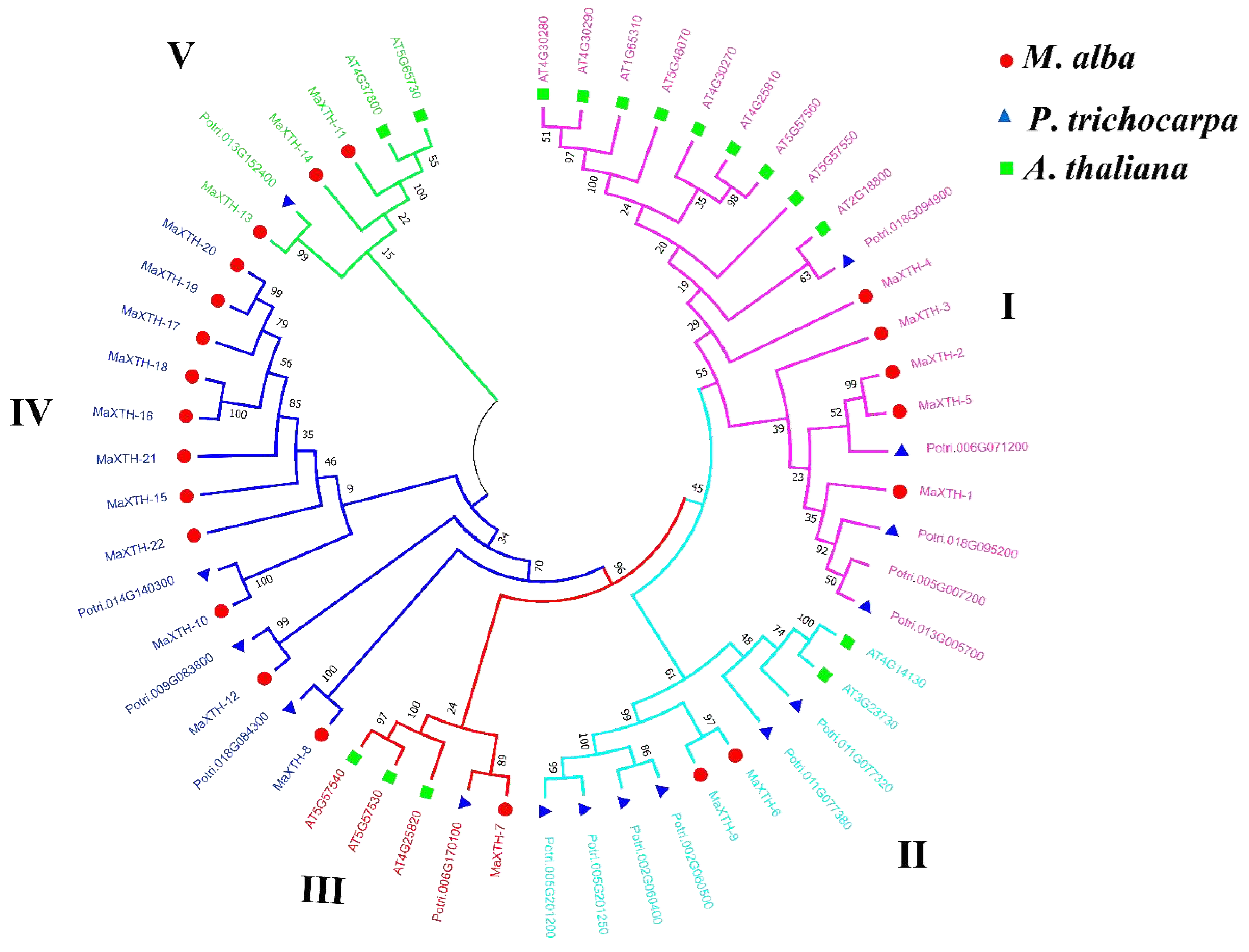
Figure 7.
Cis-elements predicted within the 2 kb sequences upstream of the M. alba XTH gene promoters. (A) Phylogenetic tree of M. alba XTH genes, clustered using MEGA7.0 (B) Distribution of cis-acting elements, with distinct colors in each box denoting different promoters.
Figure 7.
Cis-elements predicted within the 2 kb sequences upstream of the M. alba XTH gene promoters. (A) Phylogenetic tree of M. alba XTH genes, clustered using MEGA7.0 (B) Distribution of cis-acting elements, with distinct colors in each box denoting different promoters.
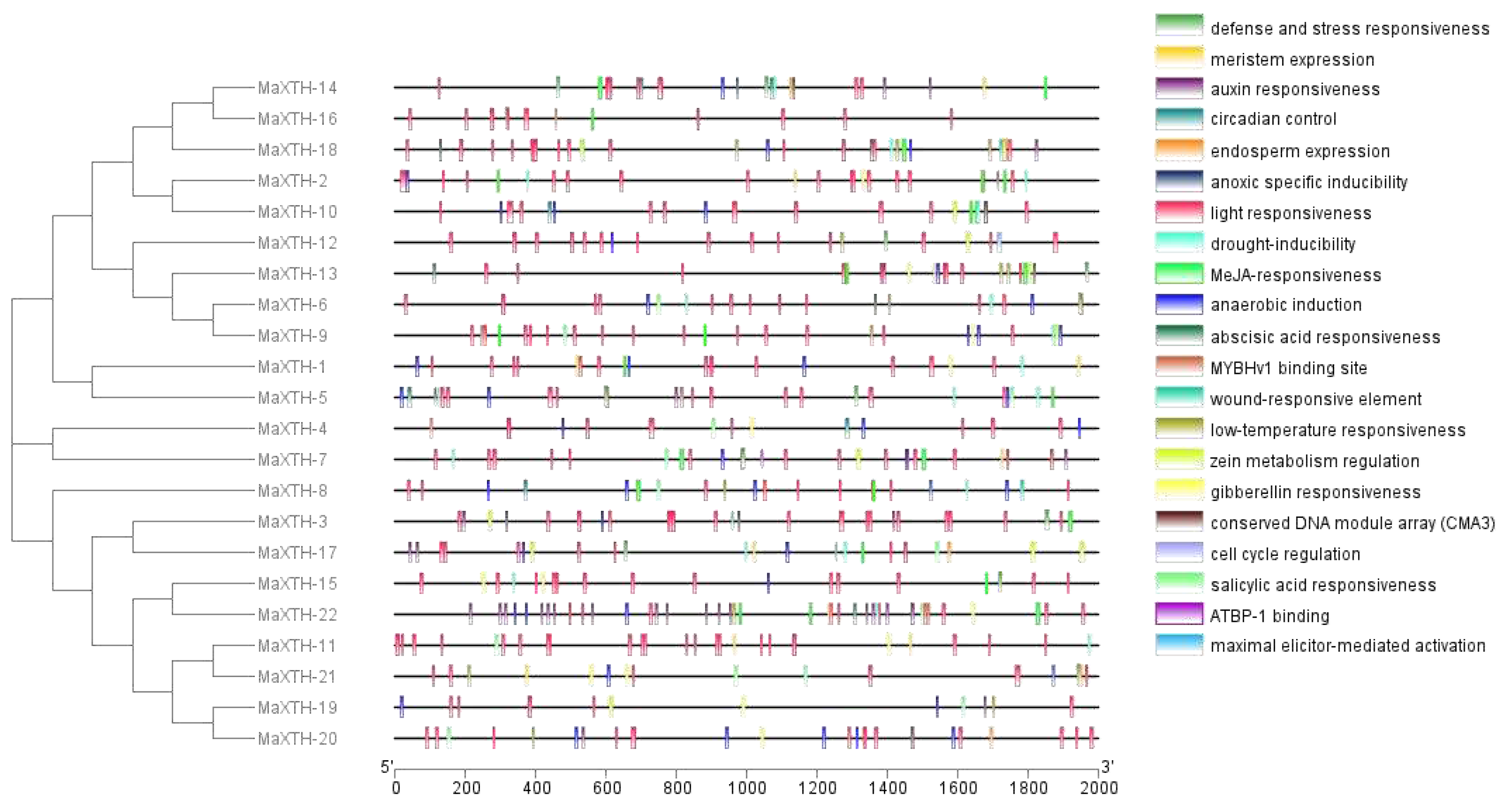
Figure 8.
Gene ontology (GO) annotation and functional clustering. (A) GO analysis of the 22 XTH genes from Morus alba and (B) A hierarchical clustering tree summarizing the correlation among significant pathways listed in the enrichment tab. Pathways with many shared genes are clustered together. Bigger dots indicate more significant p-values.
Figure 8.
Gene ontology (GO) annotation and functional clustering. (A) GO analysis of the 22 XTH genes from Morus alba and (B) A hierarchical clustering tree summarizing the correlation among significant pathways listed in the enrichment tab. Pathways with many shared genes are clustered together. Bigger dots indicate more significant p-values.
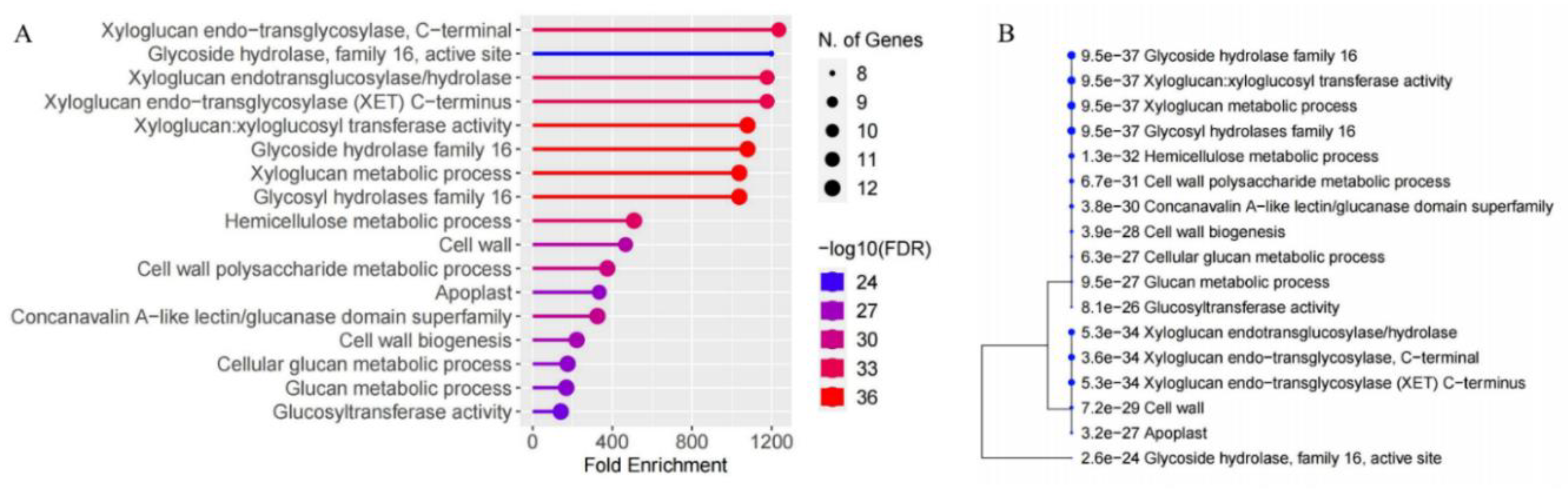
Figure 9.
(A) Chromosomal localization pattern of MaXTH genes. The scale bar on the left represents the length of the chromosomes. (B) The collinearity analysis of MaXTH genes represented via circos map, exhibiting the synteny relationships among the genes. Gene pairs are represented by pink, cyan, and blue lines, while distinct colored labels outside the chromosomes denote gene names belonging to the same phylogenetic family.
Figure 9.
(A) Chromosomal localization pattern of MaXTH genes. The scale bar on the left represents the length of the chromosomes. (B) The collinearity analysis of MaXTH genes represented via circos map, exhibiting the synteny relationships among the genes. Gene pairs are represented by pink, cyan, and blue lines, while distinct colored labels outside the chromosomes denote gene names belonging to the same phylogenetic family.
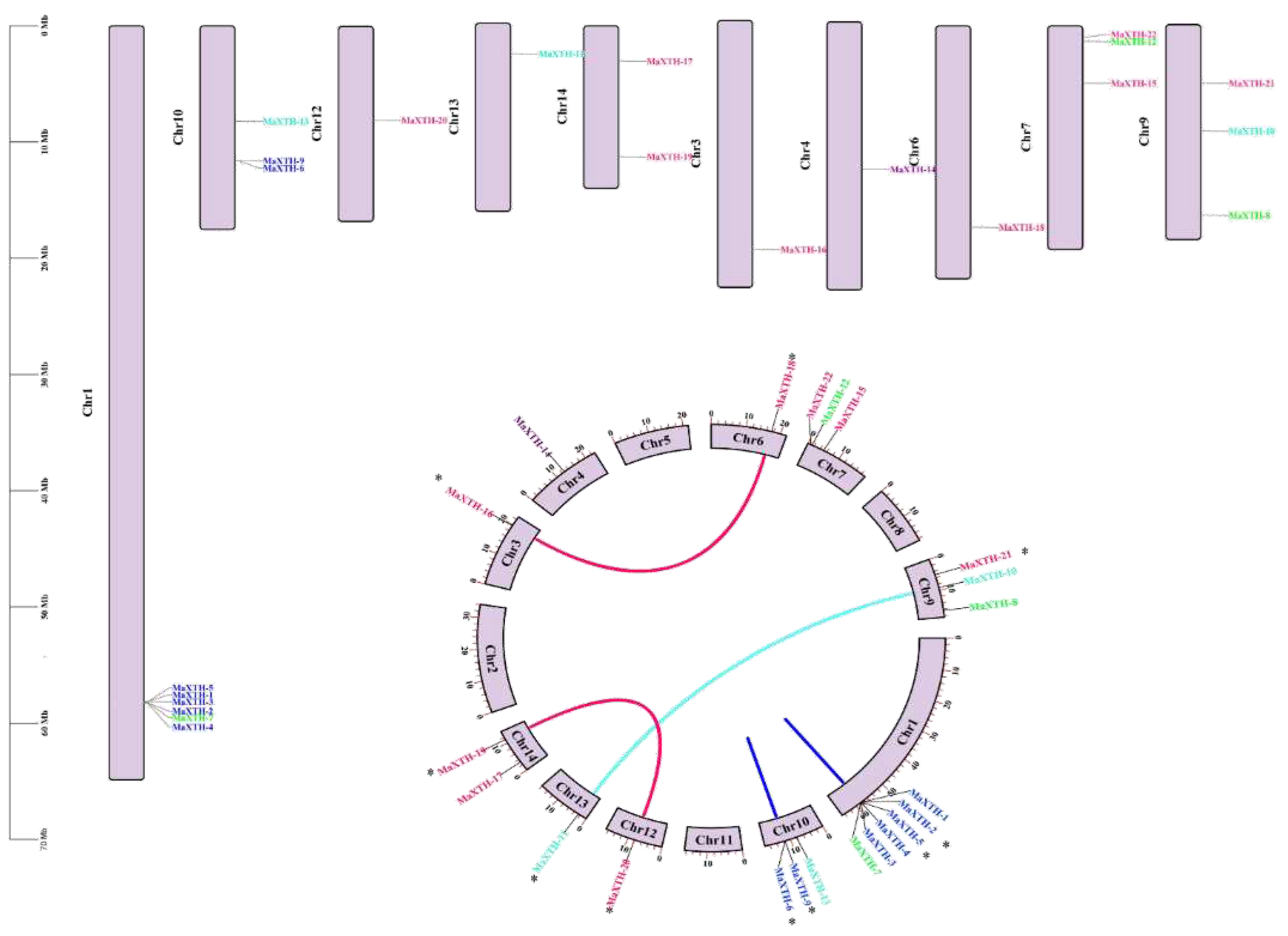
Figure 10.
Collinearity analysis of XTH genes from M. alba and other plant species: Populus trichocarpa, Arabidopsis thaliana and Zea mays. The presented data delineates XTH syntenic gene pairs through distinct colored lines: blue lines signify pairs between Morus alba and Populus trichocarpa, green lines denote pairs between M. alba and Arabidopsis thaliana, and a red line signifies pairs between M. alba and Zea mays. Additionally, grey lines elucidate the presence of orthologous genes of Morus alba shared with three other plant species.
Figure 10.
Collinearity analysis of XTH genes from M. alba and other plant species: Populus trichocarpa, Arabidopsis thaliana and Zea mays. The presented data delineates XTH syntenic gene pairs through distinct colored lines: blue lines signify pairs between Morus alba and Populus trichocarpa, green lines denote pairs between M. alba and Arabidopsis thaliana, and a red line signifies pairs between M. alba and Zea mays. Additionally, grey lines elucidate the presence of orthologous genes of Morus alba shared with three other plant species.
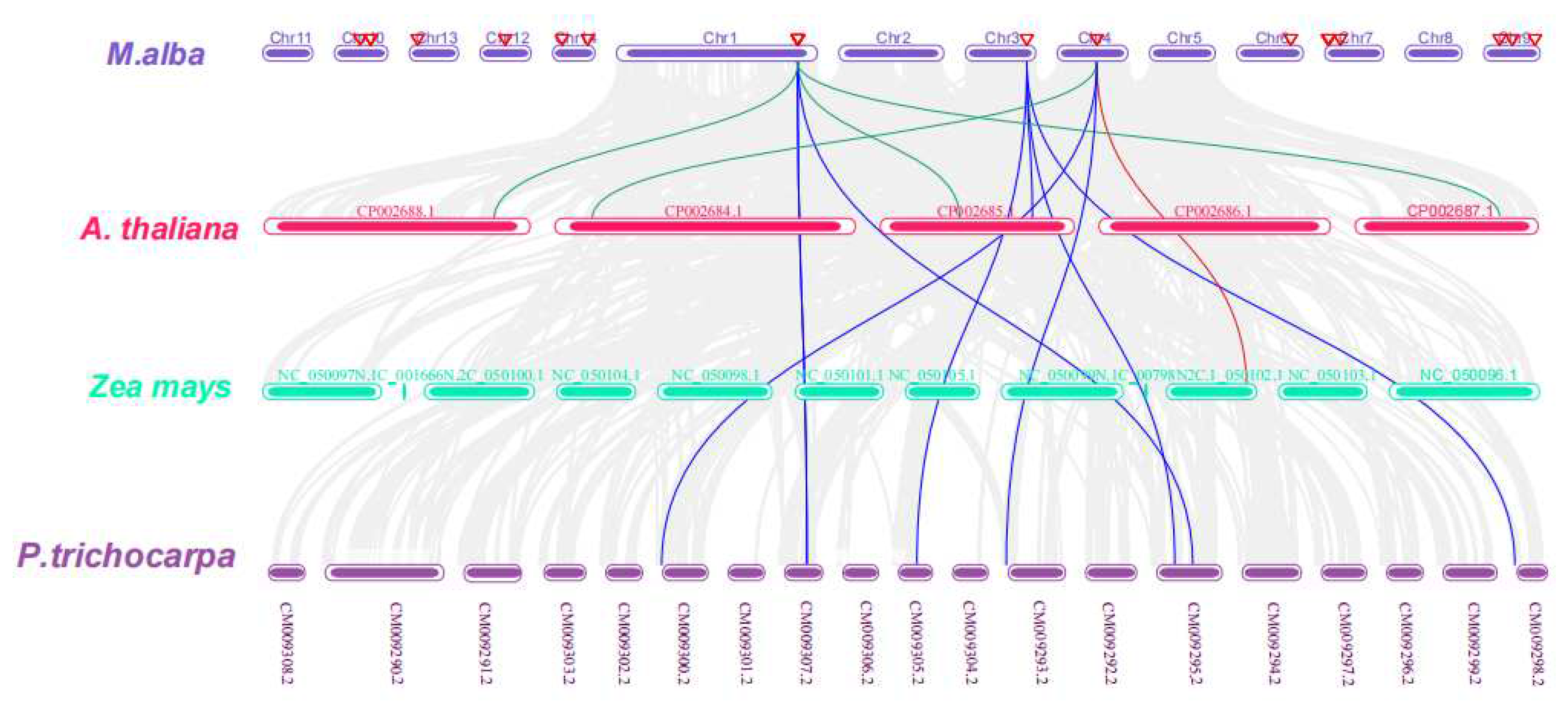
Figure 11.
Heatmap of the relative gene expression pattern of the XTH gene family based on gene relative expression in Morus alba under different magnesium treatments. (A) T1; 0 mmol/l, (B) T2; 1 mmol/l, (C) T3; 2 mmol/l, (D) T4; 6mmol/l and (E) T5; 9mmol/l. CK represents the optimum concentration of magnesium for M. alba growth (3mmol/l). From red to green show the concentration level of the gene expression.
Figure 11.
Heatmap of the relative gene expression pattern of the XTH gene family based on gene relative expression in Morus alba under different magnesium treatments. (A) T1; 0 mmol/l, (B) T2; 1 mmol/l, (C) T3; 2 mmol/l, (D) T4; 6mmol/l and (E) T5; 9mmol/l. CK represents the optimum concentration of magnesium for M. alba growth (3mmol/l). From red to green show the concentration level of the gene expression.
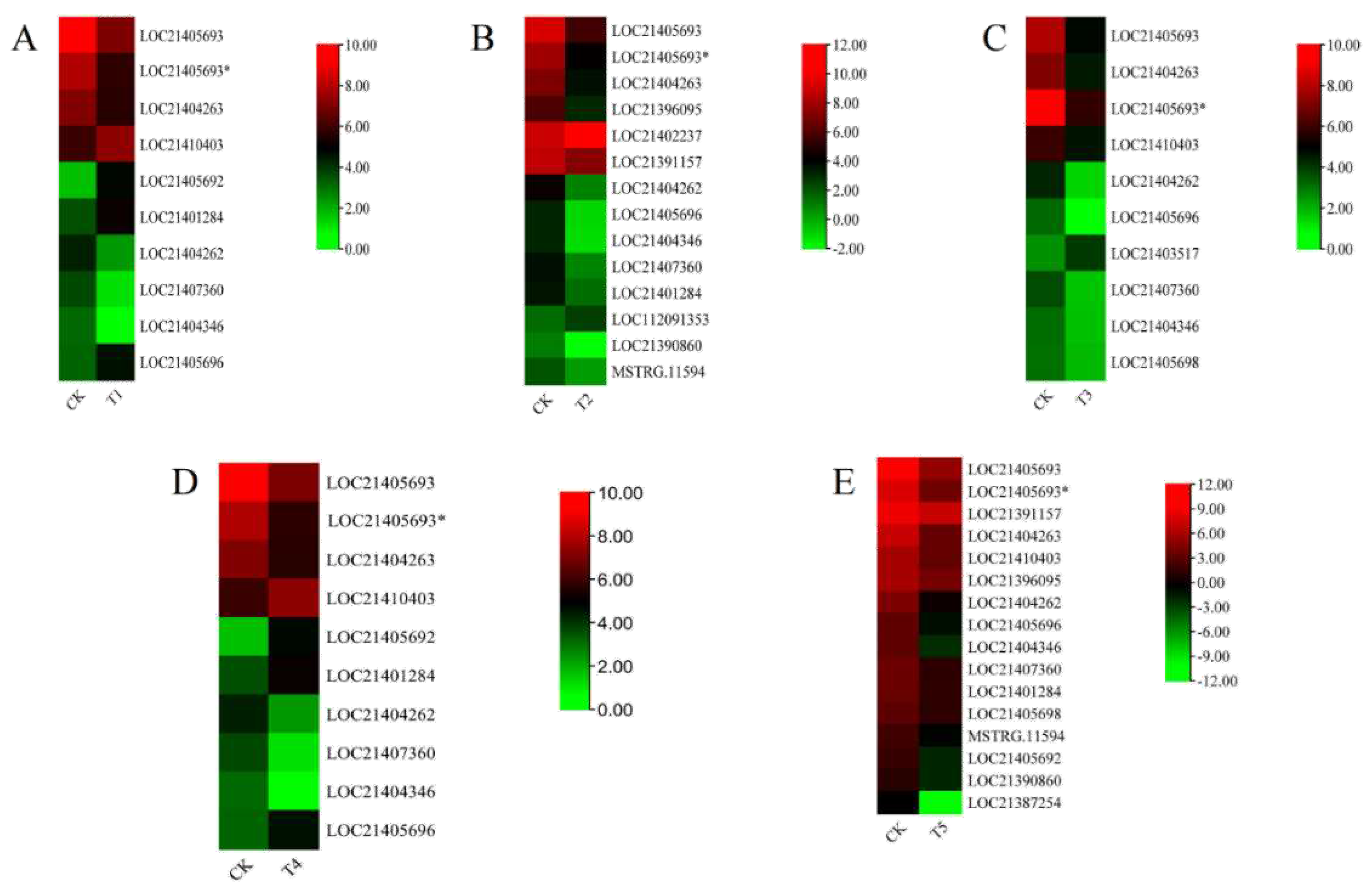
Figure 12.
The verification of relative expression levels of six MaXTH genes by RT-qPCR under different magnesium treatments. (A) LOC21410403 gene, (B) LOC21401284 gene, (C) LOC21407360 gene, (D) LOC21404263 gene, (E) LOC21405692 gene and (F) LOC21404346 gene. Bars are means of three replicates.
Figure 12.
The verification of relative expression levels of six MaXTH genes by RT-qPCR under different magnesium treatments. (A) LOC21410403 gene, (B) LOC21401284 gene, (C) LOC21407360 gene, (D) LOC21404263 gene, (E) LOC21405692 gene and (F) LOC21404346 gene. Bars are means of three replicates.
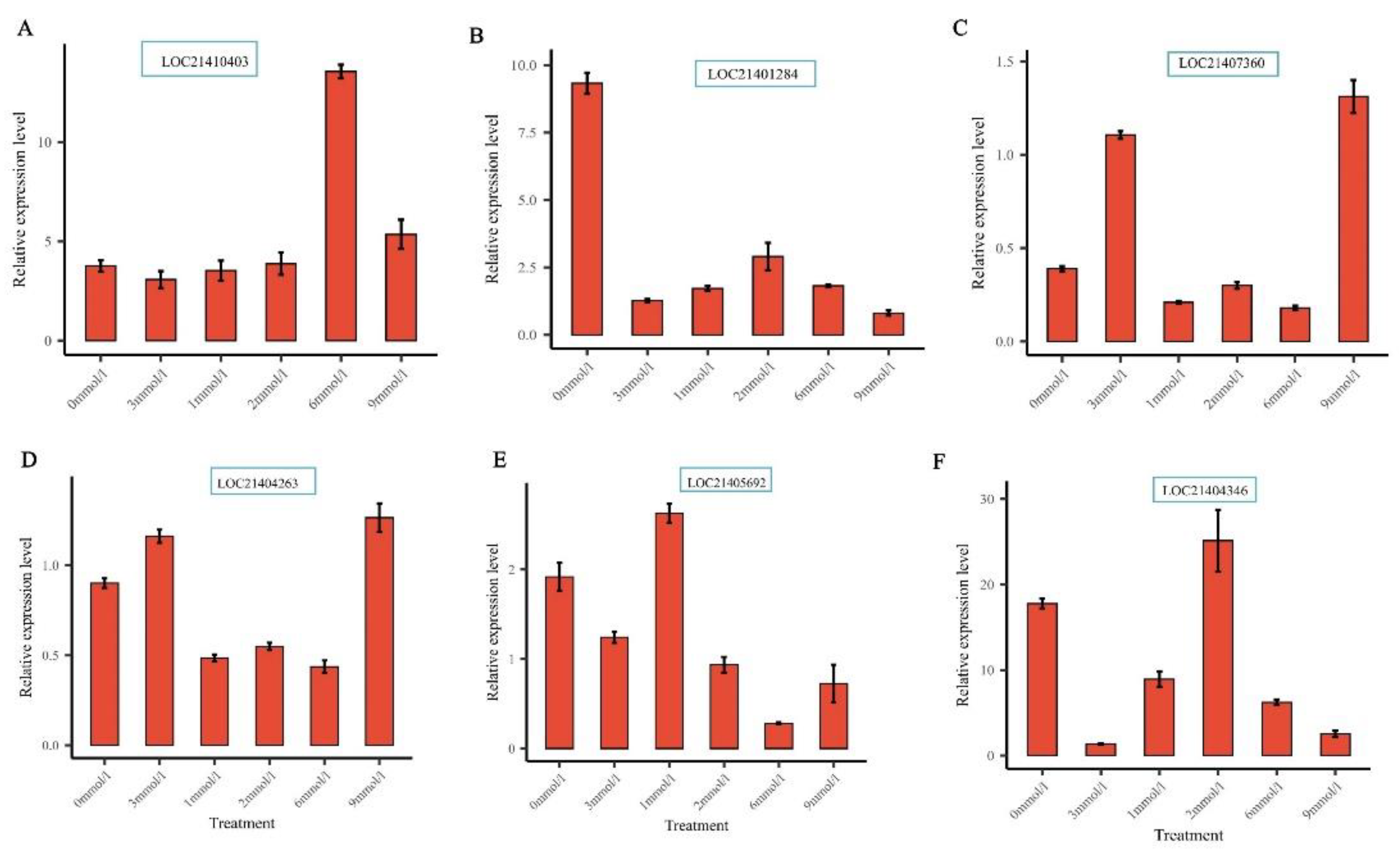

Table 1.
Physiological characteristics of XTH gene family in Morus alba.
| Gene ID | Gene Name | Chromosome | CDS (bp) | Protein Length (aa) | Exons | pI | Protein Molecular Weight (kDa) |
Sublocalization |
| LOC21405692 | MaXTH-1 | 1 | 855 | 284 | 3 | 8.15 | 32.220 | Extracellular |
| LOC21405693 | MaXTH-2 | 1 | 870 | 289 | 3 | 6.31 | 32.211 | Extracellular |
| LOC21405698 | MaXTH-3 | 1 | 894 | 297 | 3 | 5.95 | 33.323 | Extracellular |
| LOC21405697 | MaXTH-4 | 1 | 861 | 286 | 3 | 4.96 | 32.655 | Extracellular |
| LOC21405696 | MaXTH-5 | 1 | 867 | 288 | 3 | 6.21 | 32.501 | Extracellular |
| LOC21404263 | MaXTH-6 | 10 | 870 | 289 | 3 | 8.94 | 32.771 | Extracellular |
| LOC21405699 | MaXTH-7 | 1 | 918 | 305 | 3 | 6.6 | 35.306 | Extracellular |
| LOC21387185 | MaXTH-8 | 9 | 867 | 288 | 4 | 8.96 | 32.893 | Extracellular |
| LOC21404262 | MaXTH-9 | 10 | 813 | 231 | 4 | 5.28 | 25.884 | Extracellular |
| LOC21404346 | MaXTH-10 | 13 | 894 | 297 | 5 | 8.87 | 34.529 | Extracellular |
| LOC21391157 | MaXTH-11 | 9 | 873 | 290 | 4 | 6.24 | 33.270 | Extracellular |
| LOC21387254 | MaXTH-12 | 7 | 885 | 294 | 4 | 8.56 | 34.306 | Extracellular |
| LOC21401284 | MaXTH-13 | 10 | 885 | 291 | 4 | 5.71 | 33.172 | Extracellular |
| LOC21396095 | MaXTH-14 | 4 | 912 | 303 | 4 | 4.72 | 35.292 | Extracellular |
| LOC21390452 | MaXTH-15 | 7 | 873 | 290 | 4 | 5.09 | 33.165 | Extracellular |
| LOC21405370 | MaXTH-16 | 3 | 849 | 282 | 4 | 9.34 | 32.603 | Extracellular, Mitochondrial |
| LOC21410403 | MaXTH-17 | 14 | 945 | 314 | 4 | 7.67 | 35.265 | Extracellular, Vacuole |
| LOC21403517 | MaXTH-18 | 6 | 1,509 | 502 | 4 | 9.74 | 56.843 | Plasma membrane |
| LOC21402237 | MaXTH-19 | 14 | 1,023 | 340 | 4 | 6.27 | 38.748 | Extracellular |
| LOC21391267 | MaXTH-20 | 12 | 1,083 | 360 | 4 | 8.73 | 41.304 | Extracellular |
| LOC21407360 | MaXTH-21 | 9 | 645 | 214 | 1 | 5.52 | 24.071 | Cytoplasmic, Extracellular |
| LOC21390860 | MaXTH-22 | 7 | 945 | 314 | 3 | 5.68 | 35.599 | Cytoplasmic, Nuclear |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



