Submitted:
11 December 2023
Posted:
12 December 2023
You are already at the latest version
Abstract
Woodland bat assemblages are usually structured in space by distance from the ground, water and obstacles, features often defining chiropteran hunting tactics. Consequently, bat species composition differs strongly among various habitats even within the same forest patch. However, when conducting local bat survey in Wolin National Park (WPN), we revealed unexpected uniformity in qualitative and quantitative structure of bat assemblages, based on mist netting and ultrasound recording. In total, 10 vespertilionid species were detected. In all methods and sampled habitats a single species, Pipistrellus pygmaeus, predominated, while no Barbastella barbastellus, an old forest specialist, was detected, despite the abundance of preferred daily roosts. We also reviewed literature for mist netted bat samples in four different habitats of lowland Polish forests. Samples usually clustered based on habitats and the same habitat classes often clustered very closely, despite representing geographically distant forests. The exception was WPN, where all four habitat classes formed a tightly packed cluster. We suppose that P. pygmaeus might act as a hyperabundant native species, a successful generalist that reduces the contribution of more specialized taxa in the assemblage. It probably benefits from both forest renaturation and anthropogenic cross-boundary subsidy, the latter including both roost availability and prey abundance.
Keywords:
Chiroptera
; Vespertilionidae
; woodlands
; diversity
; Poland
; Wolin National Park
1. Introduction
Forests are one of the most important habitats for insectivorous bats worldwide, providing opportunities for both foraging and roosting [1]. Bats themselves might perform a significant function in suppression of herbivorous insects and, in consequence, driving a top-down trophic cascade in forest ecosystems [2,3]. Thus, insectivorous bats act as agents of biological pest control in silviculture [4,5]. Due to significant interspecific variation in diet among sympatric bat species [6,7], the significance of that function might be affected by taxonomic composition of particular bat assemblages, which, in turn, is affected by a number of factors, acting in various spatial scales.
Chiropteran assemblages are structured mainly by responses to resources, evolutionary and geological history, as well as geographic factors, while the interspecific interactions, like competition, seem to be of lesser importance [8], though not negligible one. In the local scale, however, proportion of particular taxa among foraging or commuting bats in woodland landscape of the temperate zone is shaped primarily by spatial structure of forest habitat [9,10], as bat species partition their niches based on distance from the ground, water and obstacles [11]. These species are adapted to that distances by different wing shapes, flight speed and maneuverability [11], but also echolocation call designs [12]. Strong variation in structure of forest bat assemblages is also caused by tree stand age [13,14], level of natural and human-made disturbance [15], proportion of particular tree taxa [16,17], volume of deadwood [18], elevation [15], size of forest patch [19], location within the patch [20] and land cover adjacent to its borders [14]. As bat faunal size correlates with availability and diversity of roosts [21], while even closely related and morphologically similar species differ in roost selection [22,23], the structure of bat assemblage foraging in woodlands might be expected to result from presence or absence of particular roost types, either within the forest or in its close neighborhood. The majority of methods used to sample bat assemblages (mist netting and ultrasound recording) does not allow to differentiate between various functions of habitat during night-time bat activity (foraging site, drinking site or commuting route [24,25,26]) while different functions might also be expected to explain taxonomic structure of bat samples. The most notable of such differences, hardly linked to spatial structure of forest habitat alone, are those between water bodies and forest roads [20] or between forest rivers and small ponds [27,28]. Irrespectively of casual mechanisms behind the observed variation, strong differences in bat species composition are expected among different habitats even in relatively small geographic scale, e.g. within the same forest patch, mesoregion or protected area.
The aim of that paper is to present bat assemblages of a national park in Central Europe that do not meet the aforementioned expectations, i.e. reveal unusual uniformity in qualitative and quantitative structure among different forest habitats and methods of sampling.
2. Materials and Methods
2.1. Study area
Wolin National Park (WNP) is located in the northwestern part of Poland (Figure 1), on the Baltic Sea Coast, covering a significant portion of the largest Polish island – Wolin, cut from the mainland by Dziwna channel, 90 m wide in the narrowest point and ~4.5 km in the widest. The Park was created in 1960. It covers 10937 ha, including 4648 ha of the forest ecosystems (42,5%) [29]. Most of its woodlands forms a compact block, covering the range of moraine hills with culmination at 115 m a.s.l.. From the north, they are bordered by active cliffs up to 93 m high, made of clay, sand and gravel, falling directly to the Baltic Sea. Lower cliffs, also partially active, fall into the Szczecin Lagoon, bordering park from the south, while in the north-east, sand dunes occur. Four postglacial lakes are located within the park forests or adjacent to them; they are accompanied by three artificial lakes, formed in abandoned chalk pits. The total surface of all that lakes comprises 163 ha. Except those lakes, there are only few small water bodies, all of them are shallow pools on impermeable clays, used by ungulates as drinking holes and wallows. Running waters are extremely scarce, include only four short streams, discharging the lakes, and few, even shorter creeks seeping from the southern cliff and entering the Szczecin Lagoon.
The climate, according to the Köppen classification, is a humid continental one, with a warm summer subtype (Dbf). The mean annual temperature is 9.1 °C, with a mean temperature in January of 0.8 °C and a mean temperature in July of 18.2 °C. The mean annual precipitation is 585 mm, with 30 days of any snow cover on average.
Average age of the tree stands in the WPN is 112 years. Predominant tree species are Scotch pine Pinus sylvestris (54,5% of the stand volume, average age 96 years), beech Fagus sylvatica (26,1%, 131 years) and sessile oak Quercus petraea (11,7%, 106 years). Predominant forest communities are fertile- Galio odorati-Fagetum and acid poor- Luzulo pilosae-Fagetum beech forests, with a significant contribution of acid beech-oak Fago-Quercetum and suboceanic pine- Leucobryo-Pinetum forests. However, only 26% of the communities remain unaltered by previous human interference [30]. Intensive restoration measures were implemented in the last few years, mostly focused on removal of Scotch pine, planted in the past in broadleaved forests. The predominantly moraine, woodland part of the Park is adjacent to the flat, alluvial part, located in regressive delta of Świna channel, covered predominantly by halophytic pastures and reed beds, fed by brackish, hypereutrophic waters of the Szczecin Lagoon.
Only few buildings are present within the studied forest block, mostly belonging to the Park authorities (offices, living quarters, foresters’ lodges and facilities associated with European bison breeding program). However, numerous settlements, including several villages and one city (Międzyzdroje), are located at the park borders. The number of new buildings, ranging from hotels to small cabins, is growing fast around the Park, due to ongoing tourist boom.
2.2. Data collection
Bat survey in Wolin National Park was conducted in 2022-2023, it was originally designed to establish basic inventory of bat fauna, including species composition, distribution, and location of sites of particular importance. Summer part of that survey was based on three methods:
In 10-20th July 2022-2023 bats were captured in mist nets set on forest roads, within forest stands free of understory, along forested shores of lakes and lagoons, over small brooks and streams (either free-flowing or dammed), and small, natural forest pools (always without outflow, used as water holes by ungulates). In total, 17 sites were surveyed (18 nights, in total) (Figure 1). Every night, 3-6 monofilament mist nets (6-12 m each) were set and checked every 15 min. from dusk to dawn. Bats were identified based on external morphological features, both metric and non-metric [31] and, after checking their sex, age and reproductive status, released on spot. Mist netting was conducted solely during rainless nights, with windspeed below 6 m/s. Bats were captured based on license of the Ministry of Climate and Environment DOP-WPN.61.122.2022.MŚP.
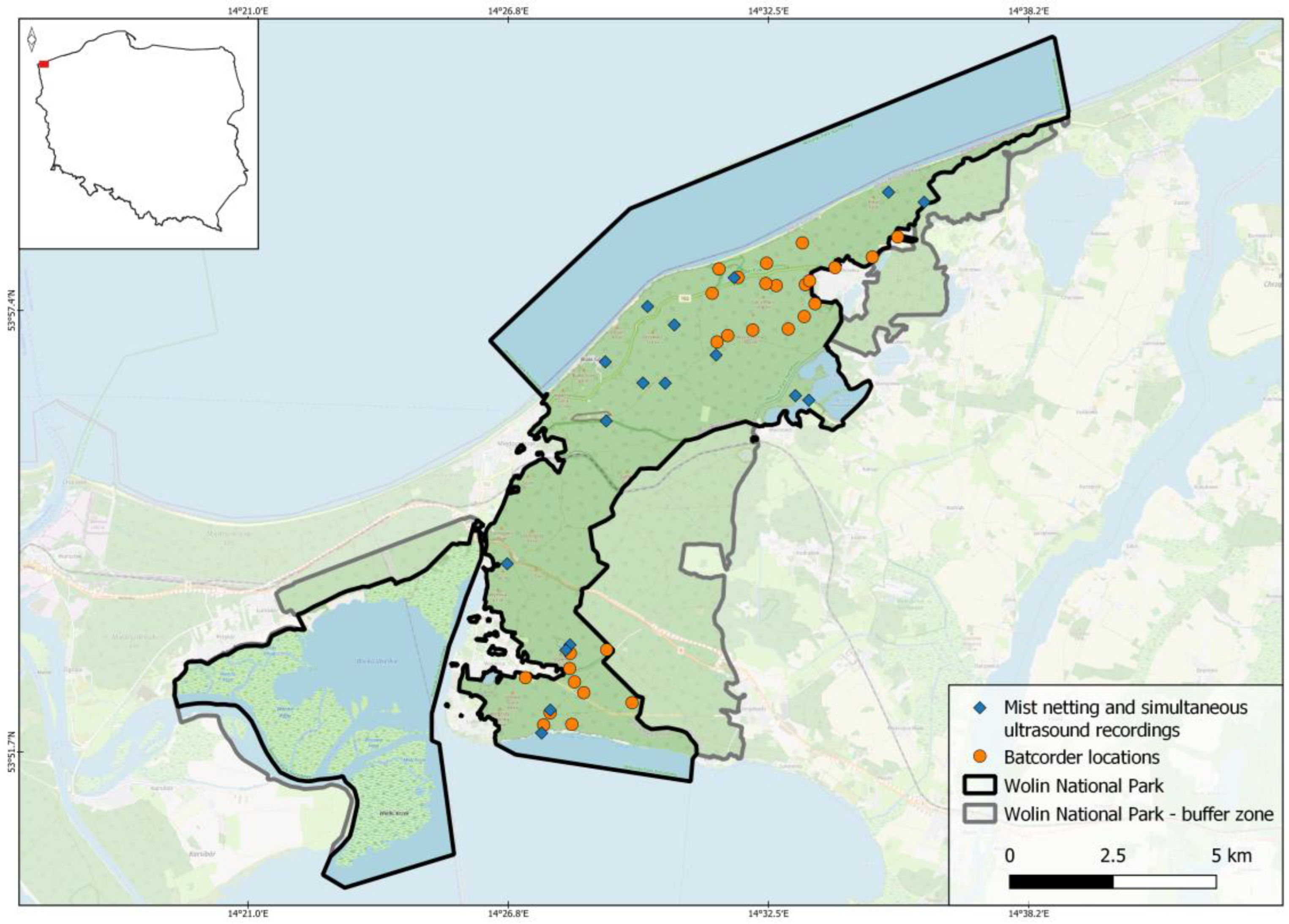
Figure 1.
Location of bat mist netting and ultrasound recording sites in Wolin National Park in 2022-2023.
Figure 1.
Location of bat mist netting and ultrasound recording sites in Wolin National Park in 2022-2023.
During the mist-netting, echolocation calls were recorded automatically, using broadband, full spectrum ultrasound detectors: Pettersson D-1000X (Pettersson Elektronik AB, Sweden), Song Meter Mini Bat and Echo Meter Touch with iPad Air tablet (Wildlife Acoustics, USA) at 16 sites. Detectors were located 100-250 m from the nearest mist net, usually in the canopy gap or at the lake/lagoon shore, at the ground level, with microphone directed to the sky at angle ~45°. Recordings were downloaded to the personal computer and identified automatically, using Wildlife Acoustics Kaleidoscope Pro 5 software that included automatic classifiers for all European bat species, based on maximum-likelihood estimator [32]. We chose Balanced (0) level of sensitivity for the Auto ID function. About 17% of all recordings were identified manually using the BatSound 3.31 program, based on shape of sonograms and oscillograms, frequency of maximum amplitude, call duration, interval length and repetition rate, compared to values of that parameters available in literature [31,33].
From April to October 2022, Batcorder (ecoObs, Germany) ultrasound recorders were set for 1-5 nights (median 2) by members of the WPN staff at 31 sites (88 nights in total) (Figure 1). All recording points where located inside the tree stands. Recorders were hanged on trees at height ~1.5-2.0 m for each night (from dusk to dawn) and removed for the day. Recorded calls were identified automatically, using batIdent 1.5 software [34]. If automatic ID software recognized some files as containing call sequences of western barbastelle Barbastella barbastellus, we checked all files recorded in a particular site manually, using bcAdmin 4.0 software, searching for recordings of that particular species.
Due to low probability of correct identification [35], we left calls of representatives of Myotis genus unidentified to the species level, with exception of M. myotis and M. nattereri [31]. If automatic classifiers identified some call sequences as produced by species that have never been recorded in north-western Poland based on hand-examined specimens, we reclassified them to species with the closest call characteristics (Miniopterus schreibersii to Pipistrellus pygmaeus, P. kuhlii and Hypsugo savii to P. nathusii, Nyctalus lasiopterus to N. noctula). Genera Nyctalus, Eptesicus and Vespertilio are also known for strong overlap in call parameters, with the pair N. leisleri – V. murinus being especially difficult to distinguish [33]. Thus, we left only automatic identifications belonging to the species from that group, which presence was confirmed by captures in mist nets, while the remaining ones were reclassified to the broader category Nyctalus/Eptesicus/Vespertilio (NEV).
Location of every mist net or ultrasound recorder was recorded by application Locus Map on Android mobile phone. Classification of each location to the general habitat categories (water bodies: 1 – forest-crossing stream, 2 – lake or lagoon shore, lined by forest, 3 – small forest pool; land: 1 – canopy gap or clearing, 2 – glade, 3 – forest road) was assigned during field work.
2.3. Statistical analysis
Particular bat species differ strongly in sonar range and detectability by ultrasonic microphones, which results in underrepresentation of species producing low-intensity calls [33]. Therefore, we provided two measures of their relative abundance based on data obtained with ultrasound detectors. One is raw number and percentage of echolocation call sequences classified to particular taxa. The second are adjusted numbers and the resulting percentage, obtained by multiplying raw data by detectability coefficients, provided by [33] for open to semi-open environment, as no recording site was located in dense forest understory (Table 1). We decided, however, to provide also raw, unadjusted data, as most studies have not adjusted recording numbers for detectability, while we aimed not only to compare our datasets among habitats and methods, but also to compare the structure of forest bat assemblage in WPN with that revealed in previous papers.
Species composition among habitats and methods was compared with Pearson’s χ2 test. We used capture and acoustic data separately to compute the expected species richness, based on individual abundance rarefaction (species accumulation curve), using PAST ver. 4.07b [36]. This method allowed the in detail comparison of species diversity of bats among habitats. We compared also bat assemblages of WPN mist netted on forest roads, over streams, small bodies of water, as well as at lakes and lagoons with those netted in other forests of lowland part of Poland in years 1990-2023, classified to similar habitats (1 – rivers and streams, 2 – lakes, lagoons and large fish ponds, 3 – small ponds, 4 – forest roads). To perform such comparison, we reviewed all, already published data on bat surveys, where bats were mist netted in summer (Table S1). As morphological features to distinguish P. pygmaeus from P. pipistrellus became known only in early 2000s [37,38], individuals classified as either of them were lumped into one category, similarly as Myotis mystacinus and Myotis alcathoe. We rejected studies, where attribution of sites to habitat classes remained ambiguous. To compare species composition of samples among forest complexes and habitats, we performed cluster analysis in ClustVis, an open-access web tool for visualizing clustering of multivariate data [39], provided by University of Tartu [40]. Samples, characterized by relative abundances of 17 bat species, were clustered with complete linkage method, using two farthest objects from two clusters to be merged, with Euclidean distance as a measure of similarity and ‘tightest cluster first’ as method of tree ordering.
3. Results
3.1. General composition of bat fauna
In total, we captured 455 bats, representing 10 species: greater mouse-eared bat Myotis myotis, Natterer’s bat Myotis nattereri, Daubenton’s bat Myotis daubentonii, serotine Eptesicus serotinus, common pipistrelle Pipistrellus pipistrellus, soprano pipistrelle Pipistrellus pygmaeus, Nathusius’ pipistrelle Pipistrellus nathusii, noctule Nyctalus noctula, lesser noctule Nyctalus leisleri and brown-long eared bat Plecotus auritus. We recorded all the same species with ultrasound detectors, although M. daubentonii could not be reliably distinguished from similar taxa (thus classified to Myotis sp. only). Although batIdent software recognized 12 call sequences of Barbastella barbastellus recorded by Batcorder detectors, we did not confirmed that identification by subsequent manual analysis. Pettersson D-1000X, Mini Bat (MB) and Echo Meter Touch (EMT) detectors recorded 6828 echolocation call sequences, while Batcorders 10 950 such sequences.
3.2. Species composition revealed by different methods
Both in the netted sample and the two datasets obtained by ultrasound recording, P. pygmaeus predominated, the most strongly among netted individuals (60%). It was captured in 14 of 17 sites, being the most frequent species and it was the most numerous netted bat in 13 sites. Its calls were also recorded in the highest number of sites, while in 14 sites they appeared to be the most numerous. Distinguishing features of the netted sample were relatively high percentage of N. noctula and M. daubentonii and frequency of occurrence of M. nattereri almost as high (13 sites) as of P. pygmaeus, although M. nattereri was only an accessory species in the assemblage (5.1%). Batcorders distinguished from the remaining ultrasound detectors by recording unusually low number of N. noctula calls (3.93% of all sequences). On contrary, among recordings obtained with D-1000X, MB and EMT detectors, N. noctula appeared to be subdominant species (29.3%, compared to 36.9% of P. pygmaeus), however, when adjusting results for detectability, its share in the assemblage dropped to just 8.6%. In the adjusted results, Myotis sp. replaced N. noctula in position of the second most abundant taxon (27.5%) (Table 1). Despite strong similarity in overall structure of assemblage among methods, species composition of netted and simultaneously recorded bats differed significantly, when comparing both raw (χ2=356.03, p<0.0001) and adjusted data (χ2=279.33, p<0.0001). However, Batcorders did not differ from the remaining detectors in proportion of recorded taxa (χ2=3336.81, p=1.0).

Table 2.
Abundance (n), dominance (%) and the number of localities (NL) of particular bat species in three datasets used to characterize the summer bat assemblage in the Wolin National Park. NEV – Nyctalus/Eptesicus/Vespertilio, indet. – unidentified. MB – Mini Bat, EMT – Echo Meter Touch.
Table 2.
Abundance (n), dominance (%) and the number of localities (NL) of particular bat species in three datasets used to characterize the summer bat assemblage in the Wolin National Park. NEV – Nyctalus/Eptesicus/Vespertilio, indet. – unidentified. MB – Mini Bat, EMT – Echo Meter Touch.
| Species | mist-netting | D-1000X, MB and EMT detectors | Batcorder detectors | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| raw | adjusted | NL | raw | adjusted | NL | ||||||||
| n | % | NL | n | % | n | % | n | % | n | % | |||
| M. myotis | 1 | 0,2 | 1 | 4 | 0,06 | 5 | 0,09 | 3 | 1 | 0,01 | 1 | 0,01 | 1 |
| M. nattereri | 23 | 5,1 | 13 | 3 | 0,04 | 5 | 0,09 | 2 | 19 | 0,20 | 32 | 0,33 | 5 |
| M. daubentonii | 51 | 11,2 | 8 | - | - | - | - | - | - | - | - | - | - |
| Myotis sp. | - | - | - | 952 | 14,04 | 1590 | 27,46 | 9 | 762 | 8,04 | 1272 | 13,14 | 17 |
| E. serotinus | 5 | 1,1 | 4 | 156 | 2,30 | 98 | 1,70 | 10 | 76 | 0,80 | 48 | 0,49 | 12 |
| P. pipistrellus | 5 | 1,1 | 3 | 554 | 8,17 | 554 | 9,57 | 13 | 664 | 7,01 | 664 | 6,86 | 15 |
| P. pygmaeus | 273 | 60,0 | 14 | 2501 | 36,89 | 2501 | 43,19 | 16 | 5532 | 58,38 | 5532 | 56,99 | 20 |
| P. nathusii | 12 | 2,6 | 9 | 460 | 6,78 | 460 | 7,94 | 15 | 666 | 7,03 | 666 | 6,88 | 18 |
| Pipistrellus sp. | 2 | 0,4 | 1 | 38 | 0,56 | 38 | 0,66 | 6 | 1367 | 14,43 | 1367 | 14,11 | 20 |
| N. noctula | 59 | 13,0 | 5 | 1988 | 29,32 | 497 | 8,58 | 12 | 373 | 3,94 | 93 | 0,96 | 14 |
| N. leisleri | 6 | 1,3 | 1 | 120 | 1,77 | 37 | 0,64 | 8 | 10 | 0,11 | 3 | 0,03 | 4 |
| P. auritus | 18 | 4,0 | 4 | 4 | 0,06 | 5 | 0,09 | 3 | 6 | 0,06 | 7 | 0,08 | 5 |
| total | 455 | 100,0 | 17 | 6780 | 100,00 | 5790 | 100,00 | 16 | 9476 | 100,00 | 9706 | 100,00 | 20 |
| NEV | - | - | - | 18 | - | - | - | - | 325 | - | - | - | - |
| indet. | - | - | - | 30 | - | - | - | - | 1149 | - | - | - | - |
3.3. Structure of bat assemblages in different habitats
P. pygmaeus was a dominant species in all the habitats sampled with mist netting, comprising almost the same percentage of individuals on forest roads, over small standing water bodies and streams (62-65%), while amounting 39% of netted bats at lake and lagoon shores. Habitats differ in the quantitative structure of netted bat samples mostly in the abundance of subdominant and accessory species, most notably N. noctula in forests and at lake shores, M. daubentonii over streams and at lake shores and P. auritus over small forest pools. The only N. leisleri individuals were netted over the small forest pool (Figure 2). Species accumulation curves revealed no differences in diversity among habitats where netting was conducted, with nearly identical trends and almost completely overlapping confidence intervals (Figure 3).
Ultrasound call recording, conducted during mist netting revealed a similar pattern. P. pygmaeus was the first most abundant species over canopy gaps (60%) and glades (49%) but it was the second most abundant species at lake shores (31%), surpassed by N. noctula (36%). Notable subdominant taxa were Myotis sp. at lake shores and N. leisleri over glades (Figure 4). The difference between habitats even diminished when data were adjusted for detectability. P. pygmaeus became the first abundant species in all three habitat classes (37-69%), while Myotis sp. – the second most abundant taxon at lake shores (34%) (Figure 5). Among recorded representatives of the genus Pipistrellus, relatively high abundance of P. nathusii over canopy gaps and clearings is worth noticing, both based on raw (16%, Figure 4) and adjusted data (17%, Figure 5). Despite strong similarity among sampled habitats, species composition among all three habitat classes differed significantly (Table 3). Species accumulation curves revealed negligibly higher diversity of bats flying in canopy gaps/clearings and virtually no differences between glades and lake shores, but confidence intervals overlap strongly in case of all three habitat classes (Figure 6).
3.4. Comparison with the other Polish lowland forests
Review of Polish faunistic literature provided 33 papers, allowing to retrieve data about bat species composition and habitat from 29 lowland forests. Combined with our data from WPN, they provided 55 samples, containing 5866 individual captures in total. The number of captures varied strongly, from 7 to 552 (median 70). More than one habitat class was sampled in 17 forests (Table S1). Samples usually, although not exclusively, clustered based on habitats, with notably high proportion of M. daubentonii and (regionally) N. noctula netted over rivers but high proportion of P. auritus, E. serotinus and (locally) B. barbastellus on forest roads. Lakes and other large water bodies were nested among rivers, revealing very similar structure of bat assemblages, while small ponds represented the most diverse set of data, clustering usually closer with roads than other water bodies. The same habitat classes often clustered very closely, despite representing geographically distant forests (e.g. rivers in the Białowieża Primeval and Kozienicka Forests or roads in Strzeleckie Forests and ‘Meteoryt Morasko’ Nature Reserve, located 250 km and 500 km apart, respectively). The exception was WPN, where all four habitat classes formed a tightly packed cluster, distinguished by unusually high proportion of P. pipistrellus/P. pygmaeus species group, revealed in no other location. In no other forest, all the sampled habitats clustered together, while the two habitat classes from the same forest clustered together only in three cases (Figure 7).
4. Discussion
4.1. General composition of bat fauna
Summer assemblage of bats inhabiting forests of the WPN appeared to consist of 10 species, belonging to five genera and single family Vespertilionidae. These species represent five foraging guilds [8]: open-air aerial foragers (N. noctula, N. leisleri), forest and clearing aerial foragers (E. serotinus, P. pipistrellus, P. pygmaeus, P. nathusii), water-surface foragers (M. daubentonii), foliage gleaners (M. nattereri, P. auritus) and a ground gleaner (M. myotis). The taxonomic and ecological composition of the assemblage is typical for the lowland Central or Northern Europe, similar to that recorded in southern Sweden or Germany [8].
However, not all species, which geographical ranges cover the WPN (i.e. occur in the north-western Poland [41]), nor even all species known to occur within the borders of the Park were recorded during our survey. Two species were netted in the WPN solely during autumn swarming at abandoned post-WW2 bunkers, these were Brandt’s bats Myotis brandtii and western barbastelle B. barbastellus [42]. As both species are regional or facultative migrants, revealing moderate but regular movements between summer and winter areas [43,44], it is quite possible that they do not occur in summer on the island of Wolin at all. Previous acoustic survey in the WPN, based solely on Batcorder detectors, provided some scarce recordings of B. barbastellus, however the authors used exclusively automatic identification and did not verify any IDs manually [45]. Verifying recorded files manually, we provided evidence that batIdent software alone is not a reliable method for detection of B. barbastellus during acoustic surveys due to widespread misidentification. The only published field test, comparing the accuracy of identification for software used in our study (batIdent and Kaleidoscope) unfortunately did not cover B. barbastellus [44].
Parti-colored bat Vespertilio murinus was recorded more than 100 years ago on the neighboring island of Uznam, about 10 km from the study area, based on collected specimens [46]. Its echolocation calls were identified by batIdent software both during our and earlier Batcorder surveys [45], however relying exclusively on auto ID is even less reliable than in case of western barbastelle. V. murinus is a species widely known from extreme call variation [47], revealing almost complete overlap of all call parameters with four other sympatric species, i.e. N. noctula, N. leisleri, E. serotinus and E. nilssonii [48]. Performance of both batIdent and Kaleidoscope in recognition of V. murinus is generally low (66.7% and 16.7% of correct IDs, respectively; [44]), thus we refrained from including that species in our database. As V. murinus is a long-distance migrant [44], regularly using coastal and insular areas of southern Baltic sea during its spring and autumn movements [49,50,51], it is highly probable that it visits the WPN as well but we cannot provide any reliable evidence that it contributes to the summer bat assemblage of that area. Relatively large amount of capture data (third largest netting sample from any lowland forest in Poland – Table S1), with not even a single V. murinus caught, makes its occurrence in the WPN during summer unlikely. Three other species, Myotis bechsteinii, M. dasycneme and E. nilssonii do also occur in north-western Poland [41], known from sites located 50-100 km south of the WPN [52,53,54]. Their absence within the border of the Park is also notable.
4.2. Species composition revealed by different methods
Both mist nets and ultrasound detectors can yield biased samples of bat assemblages, resulting in different proportions of detected species revealed by either method [55,56], thus both methods are often considered complementary [57]. Mist nets tend to underrepresent species with extremely high flight altitude, high maneuverability and/or high sonar resolution, while ultrasound detectors tend to underrepresent species with low intensity sonar [56]. Some of that features may seem mutually exclusive, e.g. fast-flying species with low maneuverability, like N. noctula, are also hunting at high altitude [11], usually above mist nets, but they can be effectively netted at drinking sites with low clutter [28,58], where they need to decrease their distance from the ground level. Among the highly maneuverable species, water-surface foragers, like M. daubentonii, were considered overrepresented, compared to aerial-hawking pipistrelles [28], probably due to significantly higher flight altitude [59]. Some of that differences, revealed for mist netting at water bodies, hardly refer to mist netting on commuting routes, like forest roads, however. Increased popularity of monofilament (ultra-thin) mist nets, replacing thicker polyester or nylon nets, predominant in earlier studies, may partially reduce bias in capture data and allow capture even those insectivorous bats with highest sonar resolution [60]. In our study, application of monofilament nets allowed to effectively sample even M. nattereri, the species adapted to the smallest target range resolution among the representatives of genus Myotis and, in consequence, to effectively navigate even few centimeters from the obstacles [61].
Bias in acoustic survey is much easier to quantify, as it is linked almost exclusively to call amplitude and resulting sonar range, at least if the species is recognizable based on recordings alone [56]. It explains much higher proportion of P. auritus, emitting low-intensity calls and often relying on passive listening, in our netted sample and much higher proportion of N. noctula, emitting the loudest calls among bats of the WPN, in parallelly collected acoustic samples. That bias might be adjusted for detectability using species-specific coefficients [33], as we did in our study. Applications of that coefficients, although reduced biased towards the loudest species, did not change the overall picture of bat assemblage, with predominance of P. pygmaeus in all three methods. No similar coefficients can be designed for detectability of species with mist netting at present. Still, a significant portion of species cannot be recognized based on echolocation calls, due to strong overlap in call parameters and in some groups (e.g. genus Myotis) that problem cannot be entirely resolved, even with application of discriminant function analysis [62] or artificial neural networks [63]. It applies also to our survey, as our automatic classifiers worked based on similar principals and we had to confirm presence of the most numerous representative of genus Myotis (M. daubentonii) with capture data. Similarly, as mist nets made of mesh of different thickness, also various models of bat detectors may differ strongly in sensitivity and that differences are not even among call frequencies [64,65].
4.2. Uniformization of bat assemblages accros habitats – unique feature of the Wolin National Park?
Available acoustic data on forest bat assemblages in lowland Poland are scarce, restricted to just few woodland areas [13,17,66,67,68], making broadscale comparisons, including cluster analysis, virtually impossible. Thus, we restricted our clustering test solely to capture data, as they are available to numerous locations from all geographical regions, usually collected as a base of local bat surveys (Table S1). Usually, local bat assemblages clustered based on habitat affiliation. Both running and large stagnant waters share predominance of M. daubentonii, a species being a specialist water-surface forager [11]. Thus, only a large surface of open water can support a nursery colony in its vicinity, such function might be played not only by a lake or a group of fishponds but also a river, even small, as on the longer section it may provide large area of water as well [28,69]. Locally, N. noctula and/or N. leisleri, also appear as indicator of forest rivers but it is hardly universal feature of Polish bat assemblages. Valleys of rivers, where noctules predominate, are covered mostly by open meadows [58], contrary to many riverine sites, where only M. daubentonii predominate and river channels are covered by canopy of riparian woodlands [28]. Meanwhile, noctules – hawking prey highly above the ground, and presumably flying down to the water only to drink – require open space, as their fast, unmaneuverable flight preclude them from flying close to obstacles, including vegetation [11].
Divergence of bat assemblages visiting small forest ponds, compared to other water bodies, probably result from their specific function – not foraging but drinking sites. This is why they attract a number of species that do not hunt over water at all, especially gleaners (Plecotus spp., M. myotis, M. bechsteinii, M. nattereri – [11,31]), but in some regions also large aerial hawkers that may hunt over waters only opportunistically (genera Nyctalus and Eptesicus [67]) but primarily visit them to drink. As forest roads act both as commuting routes and foraging sites [24], offering spaces with reduced clutter, they appeared also to be used by members of various foraging guilds. The latter is probably the reason why assemblages of bats using forest roads often cluster together with small forest ponds. Despite different functions, they aggregate a set of various species, revealing not only diverse hunting strategies but also diverse roost preferences – the indicator species of that habitat include obligatory house-dwellers (E. serotinus), old-forest specialists (B. barbastella) and generalists (P. auritus) [31]. However, there are some regions of Poland, where N. noctula can also predominate in samples netted on forest roads; it might be explained by proximity of daily roosts, as that species is also a tree dweller, selecting shelters in sites with reduced clutter [22].
Contrary to the general pattern of clustering similar habitats first, netted samples from the WPN formed a close cluster based on geographic location, a unique position among all analyzed data from lowland Poland, resulting from domination of a single species, P. pygmaeus. Some significant, secondary differences between habitats were revealed and they partially followed the same pattern, as revealed for the remaining Polish forests (higher abundance of P. auritus on small pools, or M. daubentonii over streams and larger waters). These differences were not strong enough to overcome the effect of pipistrelles predominating in every habitat. High abundance and frequency of occurrence of bats from genus Pipistrellus is specific for northern Poland, especially lakelands and Baltic Sea coast, compared to the southern part of the country [41]. That differences are suggested to result from reduced competition with Myotis mystacinus species complex, extremely rare in the north but numerous in the south [70]. It does not explain, however, domination of a single species in all sampled habitats, despite application of various methods and does not explain, why among all Central European pipistrelles, the species occupying the position of dominant is P. pygmaeus.
However, it is impossible to check, if unusual species composition of bats in the WPN is a result of recent changes or it was a feature of that region for a longer time. The only other systematic bat survey of the Park, which quantitative data have ever been published, was restricted to one method (acoustic survey with Batcorders) and one class of habitats (forest roads and interior of tree stands). Yet, it revealed predominance of P. pygmaeus at all sampled sites, but that data were collected only 4-5 years before our study [45].
4.3. Factors behind hyperabundance of Pipistrellus pygmaeus and scarcity of forest specialists
In our knowledge, the only other forest in Central European nemoral zone, where P. pygmaeus was recorded to dominate in all sampled habitats, was Włoszczakowice Forest District in western Poland. P. pygmaeus accounted for 42-48% of echolocation call sequences there, depending on forest type [17]. It might, however, predominate in some extremely depauperated bat assemblages, like in the north-east Scotland [71]. Both areas were sampled using acoustic surveys alone and in no other region, P. pygmaeus predominated among netted bats or in all studied habitats. In Białowieża Primeval Forest, it was the most numerous bat recorded in the strict protection zone of the national park but it was outnumbered by B. barbastellus among recordings from the part of that forest subjected to logging and among bats netted on forest roads [72]. In Gdańsk Pomerania region, also located in northern Poland but further east, P. pygmaeus appeared to be the rarest among three species of pipistrelles, outnumbered not only by P. pipistrellus and P. nathusii but also by N. noctula, E. serotinus and Myotis sp. (69 sites, N=4063 call sequences, [67]).
Pipistrelle bats are considered generalists, representing r selection model on r-K strategy spectrum among chiropteran life histories, due to relatively small body size, short life, high fecundity and dispersal [73]. They can thrive in variable or unpredictable environments and outcompete more specialized via exploitation, which was already suggested as mechanism explaining decline of threatened lesser horseshoe bat Rhinolophus hipposideros, following the expansion of P. pipistrellus in Switzerland, due to complete overlap in spring diet [74]. Thus, may one expect that their anthropogenically-driven expansion may even lead to depauperation of local bat fauna. Their ecological plasticity allows them to predominate in urban and agricultural landscapes, as revealed by acoustic surveys [67,75,76], although, as revealed by our study, they appeared never to predominate among netted bats in forests (Table S1). However, within Pipistrellus pipistrellus species complex, P. pygmaeus is always considered a relative specialist, regarding its habitat and landscape affiliation. It occupies much narrower habitat niche, requiring presence of wetlands, developed riparian zone and broadleaved forests, while its sibling taxon, P. pipistrellus, appears to be eurytopic, widely utilizing an agricultural landscape beyond water [67,77,78,79].
Nonetheless, classification of P. pygmaeus as ‘specialist’ relies solely on habitats used as foraging sites. The remaining aspects of its biology reveal its enormous ecological plasticity and resistance to anthropopressure. The species uses an unusual variety of nursery roosts, from buildings [80], through hollow trees [81,82], spaces under exfoliating bark [82], bat boxes [81] and bridges [83], while its house-dwelling behavior reveals no selection towards building age, material or structure [84]. It also forms nursery colonies much larger than P. pipistrellus [85], furtherly increasing its competitive potential. Moreover, due to flexible foraging strategies, P. pygmaeus is adapted to exploit abundant but ephemeral prey, like small midges, forming large swarms around even-aged, intensively managed plantations of non-native conifers [86]. At least when foraging in riparian and wetland habitats, P. pygmaeus hunts mostly small dipterans, belonging to family Chironomidae, i.e. one of the most abundant group of aquatic insects, often increasing their density with anthropogenic eutrophication of waters [87]. Finally, the species appears not to be affected by exclusion from roosts, easily switching to alternative daily shelters and resume the former activities and their patterns [80].
Therefore, roosting and foraging opportunities in forests of the WPN appear to form a set of conditions able to maintain unusually high abundance of P. pygmaeus to the point that it numerically predominates in all bat assemblages. First, native beech forests of the park, due to consistent strict protection, undergo constant increase in stand age, abundance of dead and dying trees, providing loose bark and crevices, usually preferred by P. pygmaeus [82], but also appearance of canopy gaps. On the other hand, stand conversion conducted in forest communities with anthropogenically altered structure, based on selective cutting of formerly planted Scotch pine, led to increase in abundance of coarse woody debris, appearance of numerous pine snags with vertical splits and loose bark, which were left for natural decay. It also resulted in thinning of the whole stand and appearance of several, small clearings. Both processes – natural and anthropogenic one – can be considered a broad-scale disturbance in forest ecosystems of the WPN, and such disturbances are known to increase the abundance of P. pygmaeus, as it was exemplified by outbreak of bark beetle Ips typographus in Białowieża Primeval Forest [68]. Processes that lead to appearance of numerous snags and deadwood should, however, also favorize old-forest specialists, like B. barbastellus [88], also known to select roosts under loose bark [89] or N. leisleri selecting rot cavities [22]. This is not the case of the WPN, however, where B. barbastellus seem not to occur in summer at all, despite large areas of old beech forests, its optimal habitat [89], while N. leisleri seem to be scarce and local species. The possible explanation for such high dominance indices of P. pygmaeus can be cross-boundary subsidy in both surplus roosts and prey which act in synergy with processes affecting forests within the WPN. First, chaotic urbanization of areas located at the very borders of the Park, as well as unorganized expansion of recreational development in more rural parts of the WPN’s neighborhood may provide abundant new daily shelters. As P. pygmaeus can easily inhabit even new buildings [84], able to enter even very narrow crevices, can benefit rapidly from ongoing urbanization [90]. However, the crucial subsidy that may contribute to the hyperabundance of P. pygmaeus results from location of the WPN at shores of Szczecin Lagoon, subjected to increasing input of nutrients from polluted Odra River, leading to development of hypereutrophic conditions [91]. Such conditions presumably led to exceptional abundance of aquatic nematoceran flies, especially chironomids [87], i.e. predominant prey of P. pygmaeus.
We presume that coincidence of spontaneous and active renaturation with provision of surplus anthropogenic roosts and abundance of prey due to contact with waters subjected to eutrophication might create optimal habitat for P. pygmaeus. It might also allow P. pygmaeus to outcompete from tree roosts other bat species that would, otherwise, benefit from renaturation of the Park’s forests but occupy much narrower ecological niches, like B. barbastellus, being a rare example of food specialist among Central European bats [92]. We cannot exclude that success of P. pygmaeus affects even much more common and more generalist tree-dwelling bats that use different hunting tactics (N. noctula), reducing their relative abundance in the quantitative structure of the assemblages. If the affected species are characteristic for particular habitat (e. g. M. daubentonii for water courses and lakes), it increases the similarity of assemblages across habitats and might lead to general depauperation of bat fauna. If the proposed scenario is an accurate model, predominance of P. pygmaeus in all studied forest habitats of the WPN should be treated as an example of hyperabundant native species, i.e. taxon that was not introduced by humans but benefited from their activities, negatively affecting other, less successful native species [93].
5. Conclusions
Our study revealed a unique quantitative structure of forest bat assemblage in central European protected area, characterized by exceptionally high relative abundance of a single generalist species, P. pygmaeus, known for resistance to anthropopressure, flexible foraging strategy and broad spectrum of utilized roosts, from natural to anthropogenic ones. It led to general depauperation of bat fauna, with low proportion of old-forest specialists (absence of B. barbastellus in summer), despite high age of the tree stands and availability of potential roosts. The most notable feature revealed by our survey is high level of similarity in structure of bat assemblages among all sampled habitats, contrary to all the other forests of lowland Poland, covered by our review. We suppose that P. pygmaeus may, in some ecological contexts, act as a hyperabundant native species, at least partially driven by anthropogenic cross-boundary subsidy, to our knowledge the first bat taxon revealing such function. Climate change, eutrophication and urbanization of areas neighboring woodland patches may lead to the similar situations in the future, reducing potential benefits of forest renaturation for diversity of woodland bats.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org. Table S1: Species composition of bats mist netted in four different habitats of 29 forests of lowland Poland obtained from 33 publications, used for cluster analysis (Figure 7); Table S2: Species composition of bats mist netted and recorded in different habitats of the Wolin National Park during survey in 2022-2023 – raw and adjusted data used in Figure 2-6.
Author Contributions
Conceptualization, M.C.; methodology, M.C.; bat mist netting, handling and identification, M.C., Z.W., K.Bo., E.J., J.B., M.J.-J., K.Bi.; ultrasound recording, M.C., Z.W., K.Bo., E.J.; automatic identification of bat call recordings, K.Bo., E.J., J.B., K.Bi.; manual verification of recordings, M.C., Z.W., K.Bo.; preparation of databases, M.C., Z.W., K.Bo.; statistical analysis, M.C.; writing—original draft preparation, M.C.; writing—review and editing, M.C., Z.W., K.Bo., E.J., J.B., M.J.-J., K.Bi.; data visualization, M.C.; map and spatial data, M.J.-J., K.Bi.; preparation of references, M.J.-J.; project administration, M.C., K.Bo. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the Forest Fund of State Forests National Forest Holding (Fundusz Leśny Państwowego Gospodarstwa Leśnego Lasy Państwowe), grant no. EZ.0290.1.22.2022, through the contract no. 97/2022 between the Wolin National Park (recipient of the grant) and the Polish Society for Nature Protection ‘Salamandra’.
Data Availability Statement
All data are provided in the supplementary material.
Acknowledgments
We would like to thank cordially the staff of the Wolin National Park for permission to conduct the study, organization of funding and accommodation, help in selecting sites for mist netting and recording bat calls with Batcorders: Director Wioletta Nawrocka, Vice-Director Alicja Łepek, Konrad Wrzecionkowski, Rafał Mackiewicz, Marek Szwarc, Wanda Janek, Andrzej Jabłonka, Jan Magda, Tomasz Bajor, Łukasz Potocki, Marcin Kądziołka, Artur Szymański and Tomasz Kapral. We also thank all participants of students’ research-training camps in the WPN in 2022-2023, who helped in bat mist netting: Emilia Czabrowska, Ewelina Janikowska, Amelia Rydzyńska, Magda Sitko, Arkadiusz Trzciński, Maja Ura, Maks Wojtkiewicz, Barbara Komosińska, Wiktoria Chudzik, Paulina Kozłowska, Kacper Skokowski, Teresa Kowalewska, Rain Kosatko and Blanka Kwiatkowska.
Conflicts of Interest
Authors declare no conflict of interest.
References
- Law, B.; Park, K.J.; Lacki, M.J. Insectivorous Bats and Silviculture: Balancing Timber Production and Bat Conservation. In Bats in the Anthropocene: Conservation of Bats in a Changing World; Voigt, C., Kingston, T., Eds.; Springer: Cham, Germany, 2016; pp. 105-150. [CrossRef]
- Böhm, S.M.; Wells, K.; Kalko E.K.V. Top-Down Control of Herbivory by Birds and Bats in the Canopy of Temperate Broad-Leaved Oaks (Quercus robur). PLoS ONE 2011, Volume 6(4): e17ila857. [CrossRef]
- Beilke, E.A.; O'Keefe, J.M. Bats Reduce Insect Density and Defoliation in Temperate Forests: An Exclusion Experiment. Ecology 2023, Volume 104(2): e3903. [CrossRef]
- Charbonnier, Y.; Barbaro, L.; Theillout, A.; Jactel, H. Numerical and Functional Responses of Forest Bats to a Major Insect Pest in Pine Plantations. PLoS ONE 2014, Volume 9(10): e109488. [CrossRef]
- Ancillotto, L.; Rummo, R.; Agostinetto, G.; Tommasi, N.; Garonna A.P.; Benedetta, F.; Bernardo, U.; Galimberti, A.; Russo, D. Bats as suppressors of agroforestry pests in beech forests. Forest Ecology and Management 2022, Volume 522, 120467. [CrossRef]
- Vaughan, N. The diets of British bats (Chiroptera). Mammal Review 1997, Volume 27, pp. 77–94. [CrossRef]
- Vesterinen, E.; Puisto, A.; Blomberg, A.; Lilley, T. Table for five, please: Dietary partitioning in boreal bats. Ecology and Evolution 2018, Volume 8(3). [CrossRef]
- Findley, J. Bats. A community perspective. Cambridge Studies in Ecology; Cambridge University Press: New York, 1995; 167 pp.
- Jung, K.; Kaiser, S.; Böhm, S.; Nieschulze, J.; Kalko, E.K.V. Moving in three dimensions: effects of structural complexity on occurrence and activity of insectivorous bats in managed forest stands. Journal of Applied Ecology 2012, Volume 49, pp. 523–531. [CrossRef]
- Allegrini, C.; Korine, C.; Krasnov, B.R. Insectivorous Bats in Eastern Mediterranean Planted Pine Forests—Effects of Forest Structure on Foraging Activity, Diversity, and Implications for Management Practices. Forests 2022, Volume 13, no. 9: 1411. [CrossRef]
- Baagøe, H. J. The Scandinavian bat fauna: adaptive wing morphology, and free flight in the field. In Recent advances in the study of bats; Fenton M.B, Racey, P.A., Rayner J.M.V; Cambridge University Press: New York, 1987; pp. 57-74.
- Neuweiler, G. Foraging Ecology and Audition in Echolocating Bats. TREE 1989, Volume 6, pp. 160-166. [CrossRef]
- Węgiel, A.; Grzywiński, W.; Ciechanowski, M. The foraging activity of bats in managed pine forests of different ages. European Journal of Forest Research 2019, Volume 138, pp. 383–396. [CrossRef]
- Vlaschenko, A.; Kravchenko, K.; Yatsiuk, Y.; Hukov, V.; Kramer-Schadt, S.; Radchuk, V. Bat Assemblages Are Shaped by Land Cover Types and Forest Age: A Case Study from Eastern Ukraine. Forests 2022, Volume 13, no. 10: 1732. [CrossRef]
- Piksa, K.; Brzuskowski, T.; Zwijacz-Kozica, T. Distribution, Dominance Structure, Species Richness, and Diversity of Bats in Disturbed and Undisturbed Temperate Mountain Forests. Forests 2022, Volume 13, no. 1: 56. [CrossRef]
- Kaňuch, P.; Danko, Š.; Celuch, M.; Krištín, A.; Pjenčák, P.; Matis, Š.; Šmídt, J. Relating bat species presence to habitat features in natural forests of Slovakia (Central Europe), Mammalian Biology 2008, Volume 73, Issue 2, pp. 147-155. [CrossRef]
- Węgiel, A.; Grzywiński, W.; Jaros, R.; Łacka, A.; Węgiel, J. Comparison of the Foraging Activity of Bats in Coniferous, Mixed, and Deciduous Managed Forests. Forests 2023, Volume 14, no. 3: 481. [CrossRef]
- Tillon, L.; Bouget, C.; Paillet, Y.; Aulagnier, S. How does deadwood structure temperate forest bat assemblages? European Journal of Forest Research 2016, Volume 135, pp. 433–449. [CrossRef]
- Lesinski, G.; Kowalski, M.; Wojtowicz, B.; Gulatowska, J.; Lisowska, A. Bats on forest islands of different size in an agricultural landscape. Folia Zoologica 2007, Volume 56(2), pp. 153–161.
- Ceľuch, M.; Kropil, R. Bats in a Carpathian beech-oak forest (Central Europe): Habitat use, foraging assemblages and activity patterns. Folia Zoologica 2008, Volume 57(4), pp. 358–372.
- Humphrey, S.R. Nursery Roosts and Community Diversity of Nearctic Bats. Journal of Mammalogy 1975, Volume 56, Issue 2, pp. 321–346. [CrossRef]
- Ruczyński, I.; Bogdanowicz, W. Roost Cavity Selection by Nyctalus noctula and N. leisleri (Vespertilionidae, Chiroptera) in Białowieża Primeval Forest, Eastern Poland. Journal of Mammalogy 2005, Volume 86, Issue 5, pp. 921–930. [CrossRef]
- Lučan, R.K.; Andreas, M.; Benda, P.; Bartonička, T.; Březinová, T.; Hoffmannová, A.; Hulová, Š.; Hulva, P.; Neckářová, J.; Reiter, A.; Svačina, T.; Šálek, M.; Horáček, I. Alcathoe Bat (Myotis alcathoe) in the Czech Republic: Distributional Status, Roosting and Feeding Ecology. Acta Chiropterologica 2009, Volume 11(1), pp. 61-69. [CrossRef]
- Lesiński, G.; Olszewski, A.; Popczyk, B. Forest roads used by commuting and foraging bats in edge and interior zones. Polish Journal of Ecology 2011, Volume 59, pp. 611-616.
- Seibold, S.; Buchner, J.; Bässler, C.; Müller, J. Ponds in acidic mountains are more important for bats in providing drinking water than insect prey. Journal of Zoology 2013, Volume 290, pp. 302-308. [CrossRef]
- Russo, D.; Ancillotto, L.; Cistrone, L.; Libralato, N.; Domer, A.; Cohen, S.; Korine, C. (2019), Effects of artificial illumination on drinking bats: a field test in forest and desert habitats. Animal Conservation 2019, Volume 22, pp. 124-133. [CrossRef]
- Kowalski, M.; Krasnodębski, I.; Sachanowicz, K.; Dróżdż, R.; Wojtowicz, B. Skład gatunkowy, wybiórczość kryjówek i miejsc żerowania nietoperzy w Puszczy Kozienickiej. Kulon 1996, Volume 1 (1-2), pp. 25-41.
- Ciechanowski M. Community structure and activity of bats (Chiroptera) over different water bodies. Mammalian Biology 2002, Volume 67, pp. 276-285. [CrossRef]
- Woliński Park Narodowy 2022. O parku. Ogólnie – położenie, powierzchnia, historia. Available online: https://wolinpn.pl/o-parku/ogolnie-polozenie-powierzchnia-historia/ (accessed on 18 November 2023).
- Woliński Park Narodowy 2022. Ekosystemy leśne Wolińskiego Parku Narodowego. Statystyka. Available online: https://wolinpn.pl/przyroda-parku/statystyka/ (accessed on 18 November 2023).
- Dietz, C.; Kiefer, A. Bats of Britain and Europe; Bloomsbury Natural History: London, 2016; 398 pp.
- Coleman, L.S.; Ford W.M.; Dobony, C.A.; Britzke, E.R. A comparison of passive and active acoustic sampling for a bat community impacted by white-nose syndrome. Journal of Fish and Wildlife Management 2014, Volume 5, pp. 217-226. [CrossRef]
- Barataud, M. Acoustic Ecology of European Bats. Species Identification, Study of Their Habitats and Foraging Behaviour; Biotope – Muséum national d’Historie naturelle: Paris, 2014; 352 pp.
- Brabant, R.; Laurent, Y.; Dolap, U.; Degraer, S.; Poerink, B.J. Comparing the results of four widely used automated bat identification software programs to identify nine bat species in coastal Western Europe. Belgian Journal of Zoology 2018, Volume 148(2), pp. 119–128. [CrossRef]
- Parsons, K.N.; Jones, G. Acoustic identification of twelve species of echolocating bat by discriminant function analysis and artificial neural networks. Journal of Experimental Biology 2000, Volume 203(Pt 17), pp. 2641-2656. [CrossRef]
- Hammer, R.; Harper, D.A.T.; RYAN P.D. PAST: Paleontological statistics software package for education and data analysis. Palaeontologia Electronica 2001, Volume 4, pp. 1-9.
- Häussler, U.; Nagel, A.; Braun, M.; Arnold, A. External characters discriminating sibling species of European pipistrelles, Pipistrellus pipistrellus (Schreber, 1774) and P. pygmaeus (Leach, 1825). Myotis 1999, Volume 37, pp. 27-40.
- Helversen, O.V.; Holderied, M. 2003. Zur Unterscheidung von Zwergfledermaus (Pipistrellus pipistrellus) und Mückenfledermaus (Pipistrellus mediterraneus/pygmaeus) im Feld. Nyctalus (N.F.) 2003, Volume 8(5), pp. 420-426.
- Metsalu, T.; Vilo, J. ClustVis: a web tool for visualizing clustering of multivariate data using Principal Component Analysis and heatmap. Nucleic Acids Research 2015, Volume 43(W1), pp. 566-570. [CrossRef]
- ClustVis. Available online: https://biit.cs.ut.ee/clustvis/ (accessed on 23.10.2023).
- Sachanowicz, K.; Ciechanowski, M.; Piksa, K. 2006. Distribution patterns, species richness and status of bats in Poland. Vespertilio 2006, Volume 9-10, pp. 151-173.
- Bidziński, K.; Ciechanowski, M.; Jankowska-Jarek, M.; Wikar, Z. (Department of Vertebrate Ecology and Zoology, University of Gdańsk, Poland). Unpublished data, 2023.
- Gaisler, J.; Hanák, V.; Hanzal, V.; Jarský, V. Results of bat banding in the Czech and Slovak Republics, 1948-2000. Vespertilio 2003, Volume 7, pp. 3-61.
- Hutterer, R.; Ivanova, T.; Meyer-Cords, C.H.; Rodrigues, L. Bat migration in Europe. A review of banding data and literature. Federal Agency for Nature Conservation in Germany 2005, Volume 28.
- Zyska, W.; Dylawerski, M.; Mackiewicz, R.; Skórkowski, R.; Szwarc, M.; Walczak, M.; Zyska, M.; Zyska, W. Występowanie nietoperzy w siedliskach leśnych Wolińskiego Parku Narodowego w latach 2018-2019 w tle wyników ocen prowadzonych w ostatnich 30 latach. Przegląd Przyrodniczy 2020, Volume 31, pp. 64-81.
- Ruprecht, A.L. Bats (Chiroptera). In: Atlas of Polish mammals; Pucek, Z., Raczyński, J.; PWN: Warsaw, Poland, 1983; pp. 62–82, maps 27–67.
- Schaub, A.; Schnitzler, H.U. Echolocation behavior of the bat Vespertilio murinus reveals the border between the habitat types “edge” and “open space”. Behavioral Ecology and Sociobiology 2007, Volume 61, pp. 513–523. [CrossRef]
- Zingg, P.E. (1990). Akustische Artidentifikation von Fledermäusen (Mammalia: Chiroptera) in der Schweiz. Revue Suisse de Zoologie 1990, Volume 97, pp. 263-294. [CrossRef]
- Baagøe, H. J. 2001. Danish bats (Mammalia: Chiroptera): Atlas and analysis of distribution, occurrence and abundance. Steenstrupia; Zoological Museum, University of Copenhagen, Denmark, 2001, 26, pp. 1–117.
- Ciechanowski, M.; Jakusz-Gostomska, A.; Żmihorski, M. Empty in summer, crowded during migration? Structure of assemblage, distribution pattern and habitat use by bats (Chiroptera: Vespertilionidae) in a narrow, marine peninsula. Mammal Research 2016, Volume 61(1), pp. 45-55. [CrossRef]
- Keišs, O.; Spalis, D.; Pētersons, G. Funnel trap as a method for capture migrating bats in Pape, Latvia. Environmental and Experimental Biology 2021, Volume 19(1), 7–10. [CrossRef]
- Ciechanowski, M.; Szkudlarek, R. Pierwsze stwierdzenie mroczka pozłocistego Eptesicus nilssonii (Keyserling & Blasius, 1839) na Pomorzu. Nietoperze 2003, Volume 4, pp. 105–107.
- Ciechanowski, M.; Sachanowicz, K.; Kokurewicz, T. Rare or underestimated? – The distribution and abundance of the pond bat (Myotis dasycneme) in Poland. Lutra 2007, Volume 50, pp. 107-134.
- Ciechanowski, M. 2021. Myotis bechsteinii (Kuhl, 1817). Atlas ssaków Polski. Instytut Ochrony Przyrody PAN, Kraków, Poland. Available online: https://www.iop.krakow.pl/Ssaki/gatunek/160 (accessed on 6th November 2023).
- Murray, K.; Britzke, E.; Hadley, B.; Robbins, L. Surveying bat communities: A comparison between mist nets and the Anabat II bat detector system. Acta Chiropterologica 1999, Volume 1, pp. 105-112.
- Flaquer, C.; Torre, I.; Arrizabalaga, A. Comparison of Sampling Methods for Inventory of Bat Communities, Journal of Mammalogy 2007, Volume 88, Issue 2, pp. 526–533. [CrossRef]
- MacSwiney G., M.C.; Clarke, F.M.; Racey, P.A. What you see is not what you get: the role of ultrasonic detectors in increasing inventory completeness in Neotropical bat assemblages. Journal of Applied Ecology 2008, Volume 45, pp. 1364-1371. [CrossRef]
- Rachwald, A.; Boratyński, P.; Nowakowski, W.K. 2001. Species composition and night-time activity of bats flying over rivers in Białowieża Primeval Forest (Eastern Poland). Acta Theriologica 2001, Volume 46(3), pp. 235-242. [CrossRef]
- Ciechanowski, M.; Zając, T.; Biłas, A.; Dunajski, R. Spatiotemporal variation in activity of bat species differing in hunting tactics: effects of weather, moonlight, food abundance, and structural clutter. Canadian Journal of Zoology 2007, Volume 85(12), pp. 1249-1263. [CrossRef]
- Chaves-Ramírez, S.; Castillo-Salazar, C.; Sánchez-Chavarría, M.; Solís-Hernández, H.; Chaverri, G. Comparing the efficiency of monofilament and regular nets for capturing bats. Royal Society Open Science 2021, Volume 8:211404. [CrossRef]
- Siemers, B.; Schnitzler, HU. Echolocation signals reflect niche differentiation in five sympatric congeneric bat species. Nature 2004, Volume 429, pp. 657–661. [CrossRef]
- Vaughan, N.; Jones, G.; Harris, S. Identification of British bat species by multivariate analysis of echolocation call parameters. Bioacoustics 1997a, Volume 7(3), pp. 189 - 207. [CrossRef]
- Walters, C.L.; Freeman, R.; Collen, A.; Dietz, C.; Brock Fenton, M.; Jones, G.; Obrist, M.K.; Puechmaille, S.J.; Sattler T.; Siemers, B.M.; Parsons, S.; Jones, K.E. A continental-scale tool for acoustic identification of European bats. Journal of Applied Ecology 2012, Volume 49, Issue 5, pp. 1064-1074. [CrossRef]
- Fenton, M.B. Choosing the 'correct' bat detector. Acta Chiropterologica 2000, Volume 2, pp. 215-224.
- Adams, A.M.; Jantzen, M.K.; Hamilton, R.M; Fenton, M.B. Do you hear what I hear? Implications of detector selection for acoustic monitoring of bats. Methods in Ecology and Evolution 2012, Volume 3, pp. 992-998. [CrossRef]
- Rachwald, A.; Wodecka, K.; Malzahn, E; Kluziński, L. Bat activity in coniferous forest areas and the impact of air pollution. Mammalia 2004, Volume 68, no. 4, pp. 445-453. [CrossRef]
- Ciechanowski, M. Habitat preferences of bats in anthropogenically altered, mosaic landscapes of northern Poland. European Journal of Wildlife Research 2015, 61, pp. 415–428. [CrossRef]
- Rachwald, A.; Boratyński, J.S.; Krawczyk, J.; Szurlej, M.; Nowakowski, W.K. Natural and anthropogenic factors influencing the bat community in commercial tree stands in a temperate lowland forest of natural origin (Białowieża Forest). Forest Ecology and Management 2021, Volume 479, 118544. [CrossRef]
- Dietz, M.; Encarnação, J.A.; Kalko, E.K.V. Small scale distribution patterns of female and male Daubenton's bats (Myotis daubentonii), Acta Chiropterologica 2006, Volume 8(2), pp. 403-415. [CrossRef]
- Kurek, K.; Tołkacz, K.; Mysłajek, R. Low abundance of the whiskered bat Myotis mystacinus (Kuhl, 1817) in Poland — consequence of competition with pipistrelle bats? Applied Ecology and Environmental Research 2017, Volume 15, pp. 241–248. [CrossRef]
- Rachwald, A.; Bradford, T.; Borowski, Z.; Racey, PA. Habitat Preferences of Soprano Pipistrelle Pipistrellus pygmaeus (Leach, 1825) and Common Pipistrelle Pipistrellus pipistrellus (Schreber, 1774) in Two Different Woodlands in North East Scotland. Zoological Studies 2016, Volume 55:e22. [CrossRef]
- Rachwald, A. Występowanie mopka zachodniego Barbastella barbastellus (Schreber, 1744) w Puszczy Białowieskiej na tle innych gatunków nietoperz. In Inwentaryzacja wybranych elementów przyrodniczych i kulturowych Puszczy Białowieskiej; Matuszkiewicz, J.M, Tabor, J., Eds.; Instytut Badawczy Leśnictwa, Sękocin Stary, Poland, 2022; pp. 713–737. [CrossRef]
- Gaisler J. 1989. The r-K selection model and life history strategies in bats. In European bat research; Hanak V. Horácek I. Gaisler J., Eds.; Charles University Press, Prague, Czechoslovakia, 1989; pp. 117–124.
- Arlettaz, R.; Godat, S.; Meyer, H. Competition for food by expanding pipistrelle bat populations (Pipistrellus pipistrellus) might contribute to the decline of lesser horseshoe bats (Rhinolophus hipposideros). Biological Conservation 2000, Volume 93, Issue 1, pp. 55-60. [CrossRef]
- Mickleburgh, S. Distribution and status of bats in the London area. In European bat research; Hanak, V., Horácek, I., Gaisler, J., Eds.; Charles University Press, Prague, Czechoslovakia, 2989, pp. 327–329.
- Glendell, M.; Vaughan, N. Foraging activity of bats in historic landscape parks in relation to habitat composition and park management. Animal Conservation 2002, Volume 5, pp. 309-316. [CrossRef]
- Vaughan, N.; Jones, G.; Harris, S. Habitat Use by Bats (Chiroptera) Assessed by Means of a Broad-Band Acoustic Method. Journal of Applied Ecology 1997b, Volume 34(3), pp. 716–730. [CrossRef]
- Russ, J.M.; Montgomery, W.I. Habitat associations of bats in Northern Ireland: implications for conservation. Biological Conservation 2002, Volume 108, Issue 1, pp. 49-58. [CrossRef]
- Sattler, T.; Bontadina, F.; Hirzel, A.H.; Arlettaz, R. Ecological niche modelling of two cryptic bat species calls for a reassessment of their conservation status. Journal of Applied Ecology 2007, Volume 44, pp. 1188-1199. [CrossRef]
- Stone, E.L.; Zeale, M.R.K.; Newson, S.E.; Browne, W.J.; Harris, S.; Jones, G. (2015). Managing conflict between bats and humans: The response of soprano pipistrelles (Pipistrellus pygmaeus) to exclusion from roosts in houses. PLoS ONE 2015, Volume 10(8), [e0131825]. [CrossRef]
- Michaelsen, T.C.; Jensen, K.H.; Högstedt, G. Roost Site Selection in Pregnant and Lactating Soprano Pipistrelles (Pipistrellus pygmaeus Leach, 1825) at the Species Northern Extreme: The Importance of Warm and Safe Roosts. Acta Chiropterologica 2014, Volume 16(2), pp. 349-357. [CrossRef]
- Dietz, M.; Brombacher, M.; Erasmy, M.; Fenchuk, V.; Simon, O. Bat Community and Roost Site Selection of Tree-Dwelling Bats in a Well-Preserved European Lowland Forest. Acta Chiropterologica 2018, Volume 20(1), pp. 117-127. [CrossRef]
- Bashta, A.-T. Bridge structures and habitats of bats (Chiroptera): species and spatial diversity. Theriologia Ukrainica 2022, Volume 24, pp. 86-103.
- Jenkins, E.V.; Laine, T.; Morgan, S.E.; Cole, K.R.; Speakman, J.R. Roost selection in the pipistrelle bat, Pipistrellus pipistrellus (Chiroptera: Vespertilionidae), in northeast Scotland. Animal Behaviour 1998, Volume 56(4), pp. 909-917. [CrossRef] [PubMed]
- Barlow, K.E.; Jones, G. Roosts, echolocation calls and wing morphology of two phonic types of Pipistrellus pipistrellus. Zeitschrift für Säugetierkunde 1999, Volume 64, pp. 257–268.
- Kirkpatrick, L.; Graham, J.; McGregor, S.; Munro, L.; Scoarize, M. Flexible foraging strategies in Pipistrellus pygmaeus in response to abundant but ephemeral prey. PLOS ONE 2018, Volume 13(10): e0204511. [CrossRef]
- Carr, J.F.; Hiltunen, J.K. CHANGES IN THE BOTTOM FAUNA OF WESTERN LAKE ERIE FROM 1930 TO 1961. Limnology and Oceanography 2003, Volume 10, pp. 551-569. [CrossRef]
- Rachwald, A.; Ciesielski, M.; Szurlej, M.; Żmihorski, M. Following the damage: Increasing western barbastelle bat activity in bark beetle infested stands in Białowieża Primeval forest. Forest Ecology and Management 2022, Volume 503, 119803. [CrossRef]
- Russo, D.; Cistrone, L.; Jones, G.; Mazzoleni, S. Roost selection by barbastelle bats (Barbastella barbastellus, Chiroptera: Vespertilionidae) in beech woodlands of central Italy: consequences for conservation. Biological Conservation 2004, Volume 117, Issue 1, pp. 73-81. [CrossRef]
- Cel’uch, M.; Danko. S.; Kaňuch P. On urbanisation of Nyctalus noctula and Pipistrellus pygmaeus in Slovakia. Vespertilio 2006, 9-10, pp. 219-221.
- Friedland, R.; Schernewski, G.; Gräwe ,U.; Greipsland, I.; Palazzo, D.; Pastuszak, M. Managing Eutrophication in the Szczecin (Oder) Lagoon-Development, Present State and Future Perspectives. Frontiers in Marine Science 2019, Volume 5. [CrossRef]
- Andreas, M.; Reiter, A.; Benda, P. Prey Selection and Seasonal Diet Changes in the Western Barbastelle Bat (Barbastella barbastellus). Acta Chiropterologica 2012, Volume 14(1), pp. 81-92. [CrossRef]
- Moore, J.H.; Gibson, L.; Amir, Z.; Chanthorn, W.; Ahmad, A.H.; Jansen, P.A.; Mendes, C.P.; Onuma, M.; Peres, C.A.; Luskin, M.S. The rise of hyperabundant native generalists threatens both humans and nature. Biological Reviews 2023, Volume 98, Issue 5, pp. 1829-1844. [CrossRef]
- Szyp, E. Nietoperze Brodnickiego Parku Krajobrazowego. MSc Thesis.. Department of Vertebrate Zoology, Institute of Biology, Nicolaus Copernicus University, Toruń, Poland, 1996.
- Ciechanowski, M.; Koziróg, L.; Dariusz, J.; Przesmycka, A.; Świątkowska, A.; Kisicka, I.; Kasprzyk, K. Bat fauna of the Iława Lakeland Landscape Park (northern Poland. Myotis 2002, Volume 40, pp. 33-45.
- Bugajna B. Wstępne badania nad nietoperzami (Chiroptera) rezerwatu „Meteoryt Morasko”. Rocznik Naukowy Polskiego Towarzystwa Ochrony Przyrody Salamandra 1996, Volume 1, pp. 217-218.
- Ciechanowski, M.; Czablewska, A.; Mączyńska, M.; Narczyński, T.; Przesmycka, A.; Zapart, A.; Jarzembowski, T.; Rachwald, A. Nietoperze (Chiroptera) Parku Krajobrazowego „Mierzeja Wiślana”. Nietoperze 2008, Volume 9(2), pp. 203-224.
- Piskorski, M.; Urban, P. Nietoperze Południoworoztoczańskiego Parku Krajobrazowego. Nietoperze 2003, Volume 4(1), pp. 21-25.
- Sachanowicz, K.; Krasnodębski, I. Skład gatunkowy i antropogeniczne kryjówki nietoperzy w Lasach Łukowskich. Nietoperze 2003, Volume 4(1), pp. 27-38.
- Ciechanowski, M. Chiropterofauna Puszczy Darżlubskiej. Nietoperze 2003, Volume 4(1), pp. 45 – 59.
- Ciechanowski, M. Struktura przestrzenna zespołu i dynamika aktywności nietoperzy (Chiroptera) w krajobrazie leśno-rolniczym północnej Polski. PhD Thesis, Department of Vertebrate Ecology and Zoology, University of Gdańsk, Poland, 2005.
- Ignaczak M. Nietoperze rezerwatu „Bukowa Góra”. Nietoperze 2003, Volume 4(1), pp. 101 – 102.
- Ciechanowski, M.; Duriasz, J. Nietoperze (Chiroptera) Parku Krajobrazowego Wzgórz Dylewskich Nietoperze 2005, Volume 6(1-2), pp. 25-36.
- Ciechanowski, M.; Anikowska, U.; Nalewaja, A.; Przesmycka, A.; Biała, A. Nietoperze (Chiroptera) Parku Krajobrazowego „Dolina Słupi”. Nietoperze 2006, Volume 7(1 – 2), pp. 19 – 37.
- Lesiński, G.; Gulatowska, J.; Kowalski, M.; Fuszara, E.; Fuszara, M.; Wojtowicz, B. Nietoperze Wysoczyzny Płońskiej. Nietoperze 2006, Volume 7(1 – 2), pp. 39 – 55.
- Piskorski, M. Fauna nietoperzy Parku Krajobrazowego Lasy Janowskie. Nietoperze 2007, Volume 8(1 – 2), pp. 3 – 11.
- Piskorski, M.; Gwardjan, M.; Kowalski, M.; Wojtowicz, B.; Urban, M.; Bochen, R. Fauna nietoperzy Parku Krajobrazowego Lasy Strzeleckie. Nietoperze 2009, Volume 10(1 – 2), pp. 15 – 22.
- Łochyński, M.; Grzywiński, W. Nietoperze Parku Krajobrazowego Puszcza Zielonka. Nietoperze 2009, Volume 10(1 – 2), pp. 23 – 35.
- Piskorski M. Fauna nietoperzy Lasów Sobiborskich. Nietoperze 2008, Volume 9(1), pp. 3 – 17.
- Kmiecik, A.; Kmiecik, P.; Grzywiński, W. 2010. Chiropterofauna Wigierskiego Parku Narodowego. Nietoperze 2010, Volume 11(1 – 2), pp. 11 – 29.
- Postawa, T.; Gas, A. Fauna nietoperzy Wigierskiego Parku Narodowego (północno-wschodnia Polska). Studia Chiropterologica 2003, Volume 3–4, pp. 31–42.
- Sachanowicz, K.; Marzec, M.; Ciechanowski, M.; Rachwald, A.; Nietoperze Puszczy Rominckiej. Nietoperze 2001, Volume 2(1), pp. 109-115.
- Kowalski, M.; Ostrach-Kowalska, A.; Krasnodębski, I.; Sachanowicz, K.; Ignaczak, M.; Rusin, A. Nietoperze Parków Krajobrazowych: Górznieńsko-Lidzbarskiego i Welskiego. Nietoperze 2001, Volume 2, pp. 117-124.
- Ignaczak, M.; Radzicki, G.; Domański, J. Nietoperze Parku Krajobrazowego Międzyrzecza Warty i Widawki. Nietoperze 2001, Volume 2, pp. 125-134.
- Wikar, Z.; Ciechanowski, M. Ssaki rezerwatu przyrody „Studnica” i jego otoczenia. Chrońmy Przyrodę Ojczystą 2019, Volume 75(5), pp. 374 – 387.
- Ciechanowski, M.; Przesmycka, A.; Sachanowicz, K. 2006. Nietoperze (Chiroptera) Wdzydzkiego Parku Krajobrazowego. Parki Narodowe i Rezerwaty Przyrody 2006, Volume 25(4), pp. 85 – 100.
- Apoznański, G.; Kokurewicz, T.; Błesznowska, J.; Kwasiborska, E.; Marszałek, T.; Górska, M. Use of Coniferous Plantations by Bats in Western Poland During Summer Months”. Baltic Forestry 2020, Volume 26, pp. 1–8. [CrossRef]
- Ciechanowski, M. Ssaki (Mammalia). In Przyroda rezerwatów Kurze Grzędy i Staniszewskie Błoto na Pojezierzu Kaszubskim; Herbich, J., Ciechanowski, M., Eds.; Fundacja Rozwoju Uniwersytetu Gdańskiego, Gdańsk, Poland, 2009; pp. 256-264.
- Jarzembowski, T.; Ostrach-Kowalska, A.; Rymarzak, G. Chiropterofauna Kaszubskiego Parku Krajobrazowego. Przegląd przyrodniczy 1997, Volume 8(3), pp. 123-127.
- Ciechanowski, M. Ssaki (Mammalia). In Przyroda projektowanego rezerwatu "Dolina Mirachowskiej Strugi" na Pojezierzu Kaszubskim; Ciechanowski, M., Fałtynowicz, W., Zieliński S.; Eds.; Acta Botanica Cassubica; Gdańsk, Poland, 2004; Volume 4, pp. 110-121.
- Wojtaszyn, G. Nietoperze Pojezierza Wałeckiego. Przegląd przyrodniczy 2002, Volume 13(1-2), pp. 199-211.
- Lesiński, G.; Gryz, J.; Rachwald, A.; Krauze-Gryz, D. Bat assemblages in fragmented forest complexes near Rogów (central Poland). Forest Research Papers 2019, Volume 79, pp. 253-260. [CrossRef]
- Górecki, M.T. Nietoperze Chiroptera okolic Skierniewic. Przegląd przyrodniczy 1998, Volume IX(3), pp. 101 – 108.
- Lesiński, G.; Hejduk, J.; Gajęcka, K.; Górecki, M.; Janus, K.; Zieleniak, A. Nietoperze Bolimowskiego Parku Krajobrazowego i terenów otaczających. Parki Narodowe i Rezerwaty Przyrody 2018, Volume 37, pp. 65-80.
Figure 2.
Structure of bat assemblages in four habitats within forests of the Wolin National Park, as revealed by mist netting. Sample size (the number of captures) in brackets.
Figure 2.
Structure of bat assemblages in four habitats within forests of the Wolin National Park, as revealed by mist netting. Sample size (the number of captures) in brackets.
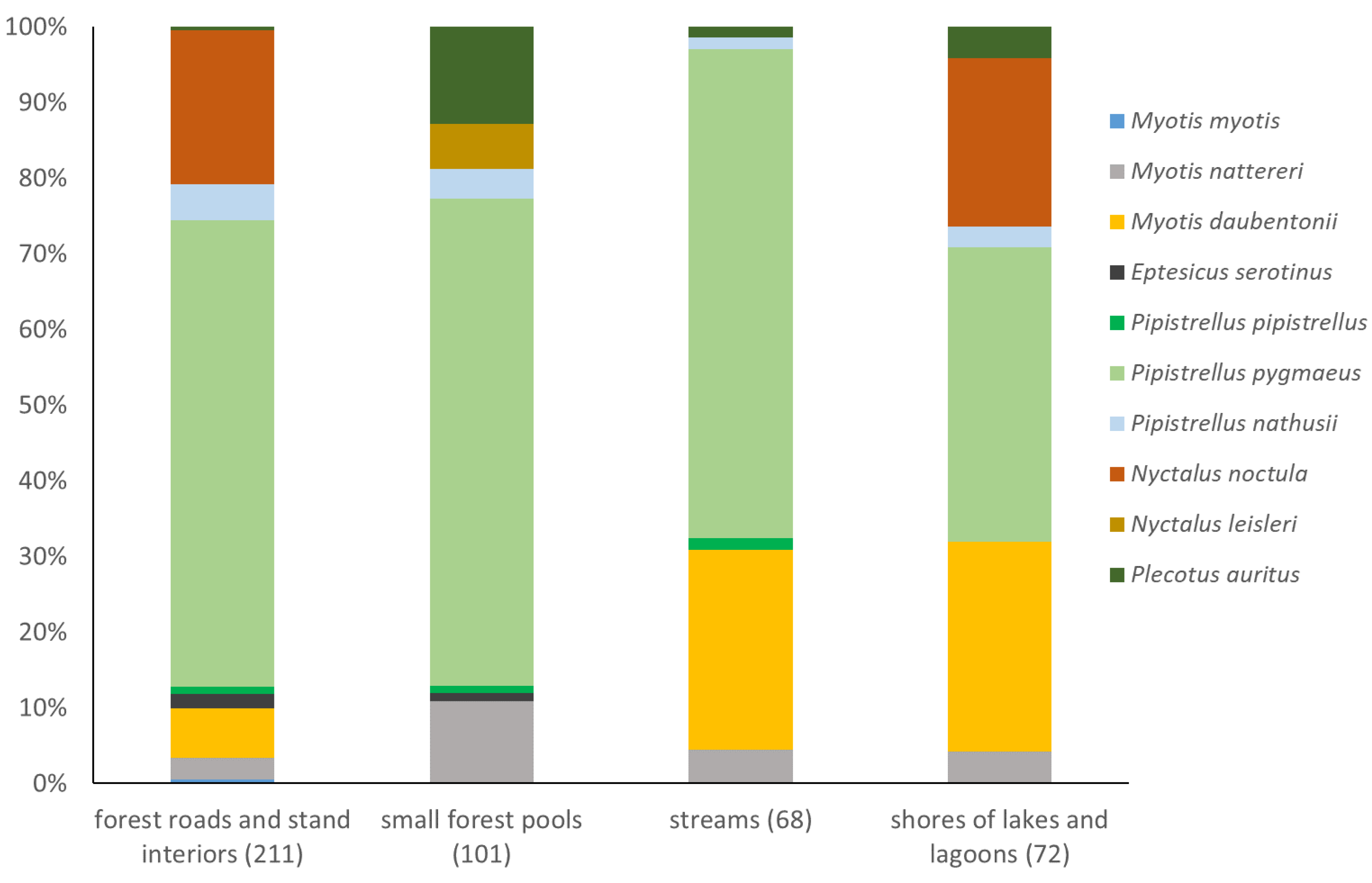
Figure 3.
Rarefaction curves for cumulative numbers of species plotted against numbers of bats mist netted in four habitats within forests of the Wolin National Park.
Figure 3.
Rarefaction curves for cumulative numbers of species plotted against numbers of bats mist netted in four habitats within forests of the Wolin National Park.
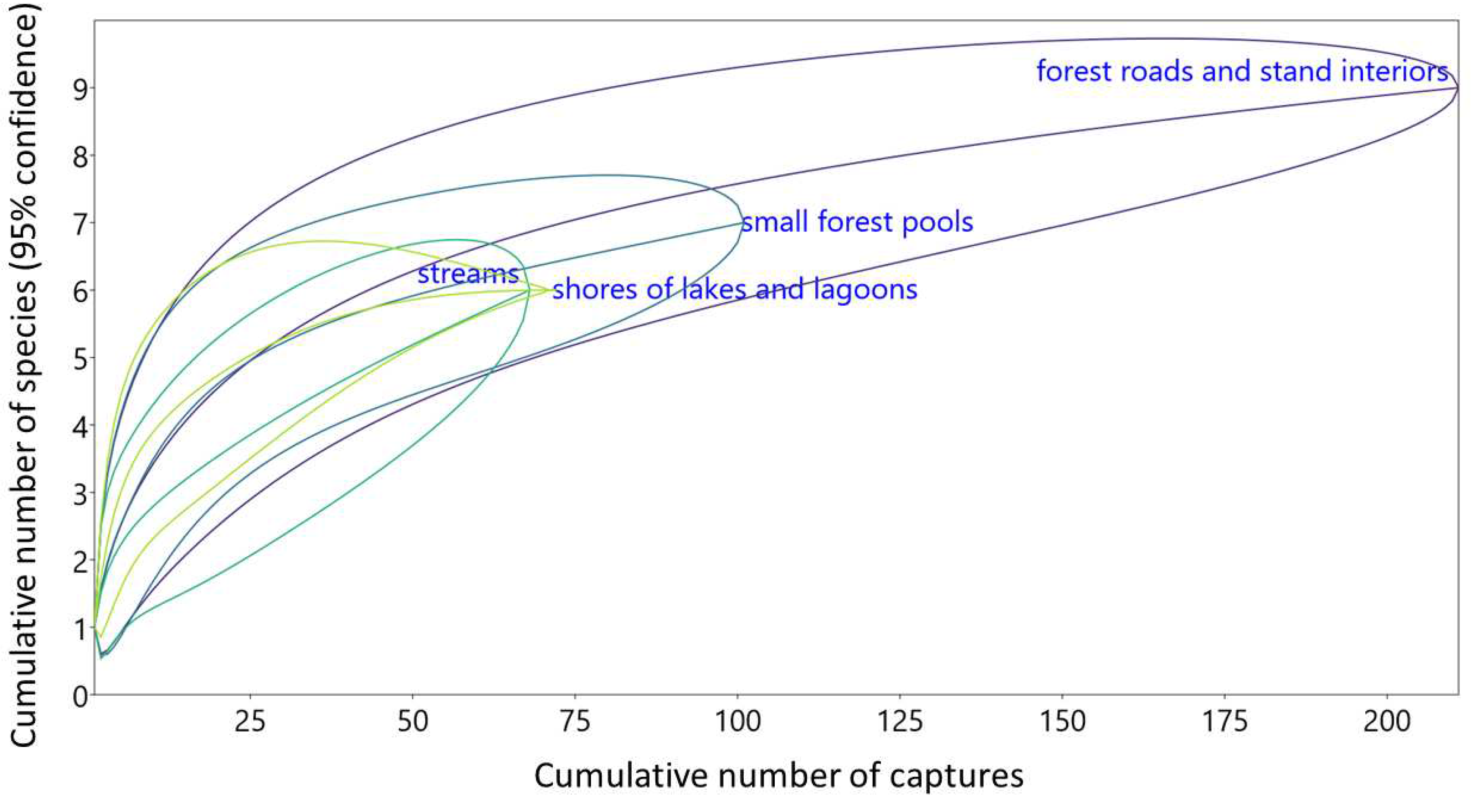
Figure 4.
Structure of bat assemblages in three habitats within forests of Wolin National Park, as revealed by ultrasound recordings (D-1000X, Mini Bat and Echo Meter Touch devices, exclusively). Sample size (the raw number of echolocation call sequences) in brackets.
Figure 4.
Structure of bat assemblages in three habitats within forests of Wolin National Park, as revealed by ultrasound recordings (D-1000X, Mini Bat and Echo Meter Touch devices, exclusively). Sample size (the raw number of echolocation call sequences) in brackets.
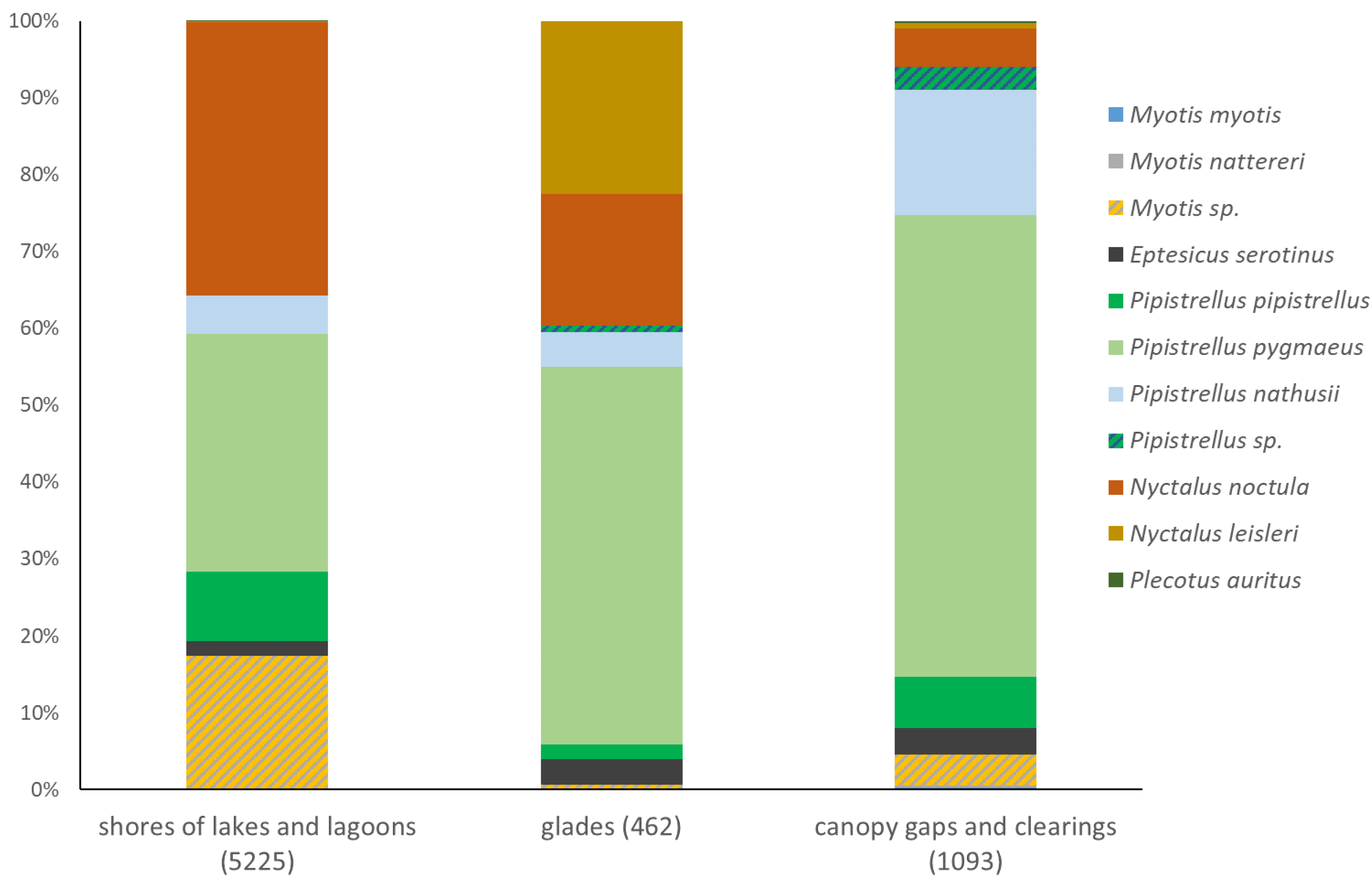
Figure 5.
Structure of bat assemblages in three habitats within forests of the Wolin National Park, as revealed by ultrasound recordings (D-1000X, Mini Bat and Echo Meter Touch devices, exclusively), with proportion of each species adjusted for detectability by multiplying the numbers of call sequences by coefficient from Table 1.
Figure 5.
Structure of bat assemblages in three habitats within forests of the Wolin National Park, as revealed by ultrasound recordings (D-1000X, Mini Bat and Echo Meter Touch devices, exclusively), with proportion of each species adjusted for detectability by multiplying the numbers of call sequences by coefficient from Table 1.
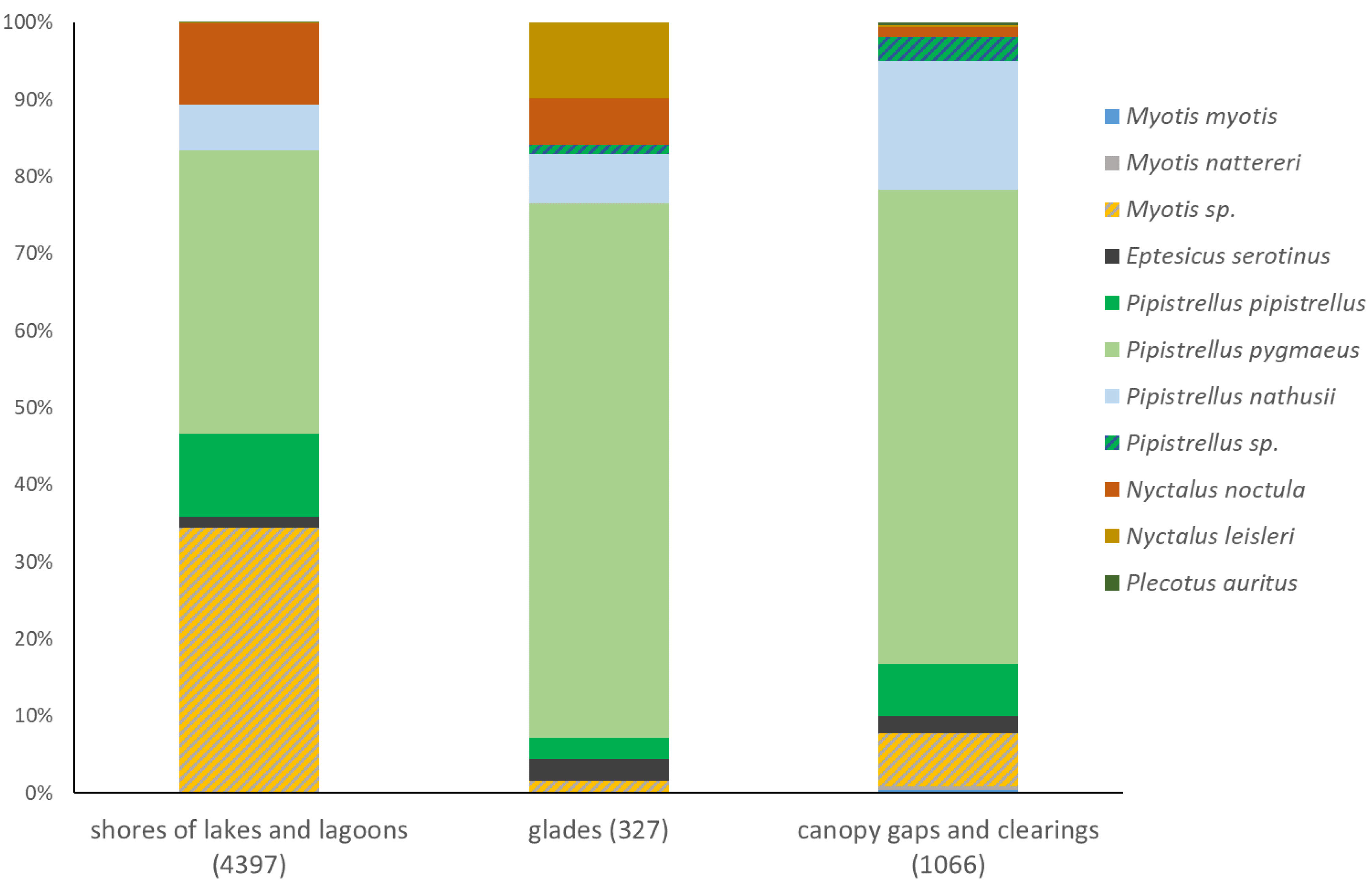
Figure 6.
Rarefaction curves for cumulative numbers of species plotted against numbers of bat call sequences recorded in three habitats within forests of the Wolin National Park (D-1000X, Mini Bat and Echo Meter Touch ultrasound recorders, exclusively).
Figure 6.
Rarefaction curves for cumulative numbers of species plotted against numbers of bat call sequences recorded in three habitats within forests of the Wolin National Park (D-1000X, Mini Bat and Echo Meter Touch ultrasound recorders, exclusively).
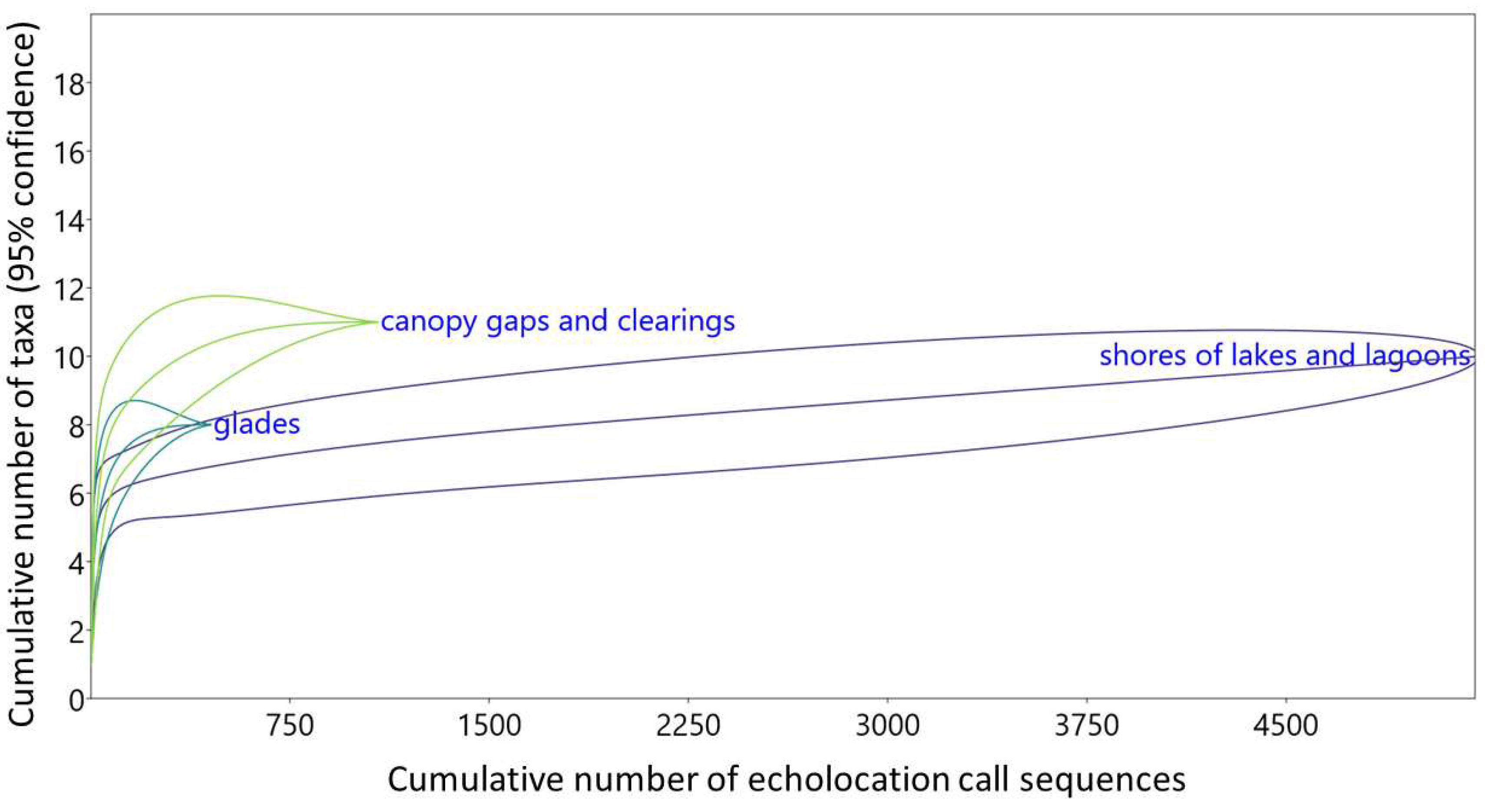
Figure 7.
Comparison of species composition of bats mist netted in lowland Polish forests, based on the literature analysis (Table S1), subjected to cluster analysis (clustering method – complete linkage, Euclidean distance, tightest cluster first). Habitat: RS – rivers and streams, L – lakes, large fishponds and lagoons, SP – small ponds, FR – forest roads and tree stand interiors. Species: Mm – Myotis myotis, Mbc – M. bechsteinii, Mn – M. nattereri, Mms/Malc – M. mystacinus/alcathoe, Mbr – M. brandtii, Md – M. daubentonii, Mds – M. dasycneme, Vm – Vespertilio murinus, En – Eptesicus nilssonii, Es – E. serotinus, Ppip/Ppyg – Pipistrellus pipistrellus/pygmaeus, Pn – P. nathusii, Nn – Nyctalus noctula, Nl – N. leisleri, Pa – Plecotus auritus, Pas – P. austriacus, Bb – Barbastella barbastellus. Site names: F – Forest, Fs – Forests, NP – National Park, LP – Landscape Park, NR – Nature Reserve. Sample sizes (numbers of captures) in brackets.
Figure 7.
Comparison of species composition of bats mist netted in lowland Polish forests, based on the literature analysis (Table S1), subjected to cluster analysis (clustering method – complete linkage, Euclidean distance, tightest cluster first). Habitat: RS – rivers and streams, L – lakes, large fishponds and lagoons, SP – small ponds, FR – forest roads and tree stand interiors. Species: Mm – Myotis myotis, Mbc – M. bechsteinii, Mn – M. nattereri, Mms/Malc – M. mystacinus/alcathoe, Mbr – M. brandtii, Md – M. daubentonii, Mds – M. dasycneme, Vm – Vespertilio murinus, En – Eptesicus nilssonii, Es – E. serotinus, Ppip/Ppyg – Pipistrellus pipistrellus/pygmaeus, Pn – P. nathusii, Nn – Nyctalus noctula, Nl – N. leisleri, Pa – Plecotus auritus, Pas – P. austriacus, Bb – Barbastella barbastellus. Site names: F – Forest, Fs – Forests, NP – National Park, LP – Landscape Park, NR – Nature Reserve. Sample sizes (numbers of captures) in brackets.
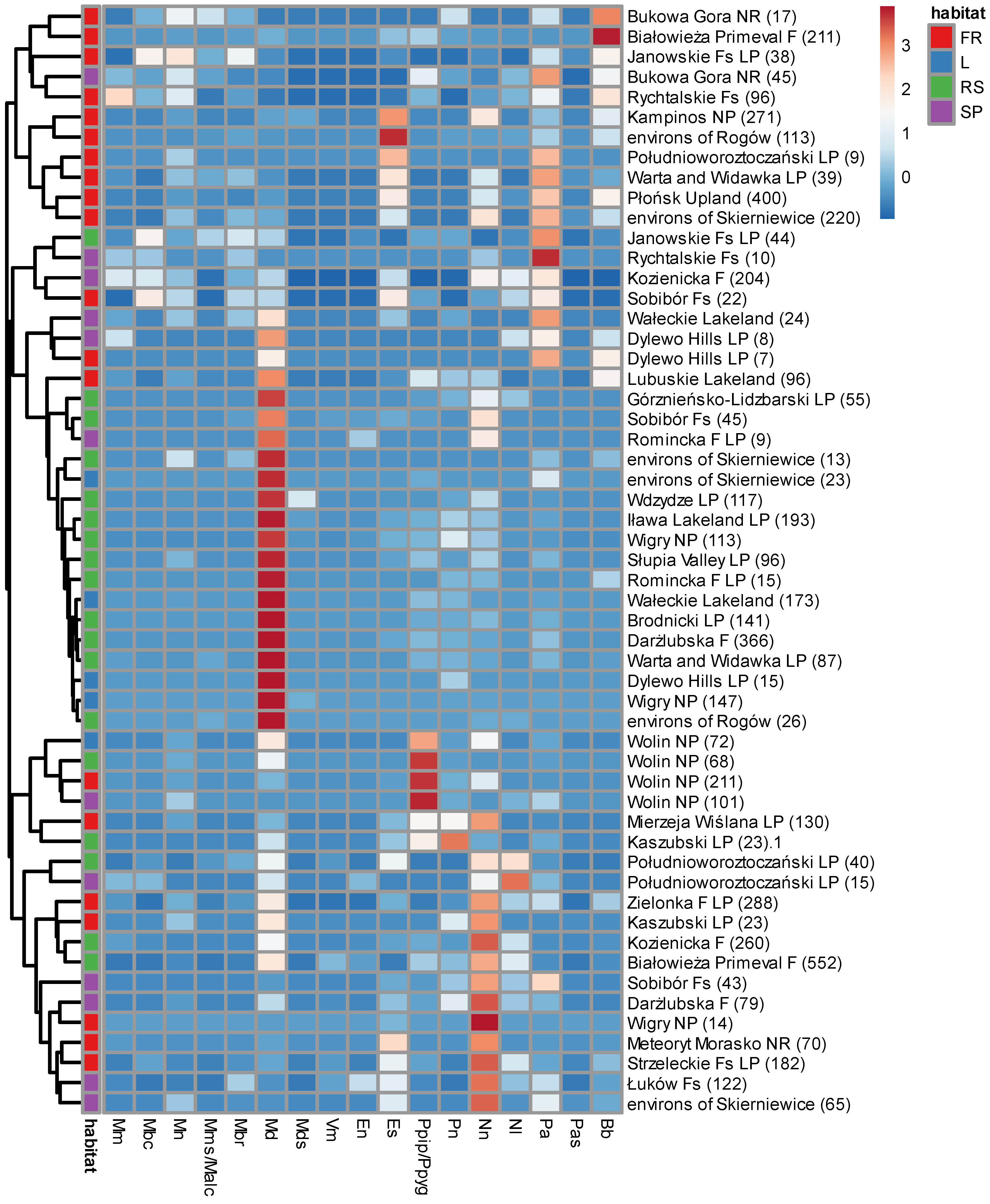
Table 1.
Coefficients used to adjust the numbers of bat passes (call sequences) belonging to particular species in open to semi-open environment [33].
Table 1.
Coefficients used to adjust the numbers of bat passes (call sequences) belonging to particular species in open to semi-open environment [33].
| Taxon | Detectability coefficient |
|---|---|
| Myotis myotis | 1.25 |
| Small Myotis1 | 1.67 |
| Eptesicus serotinus | 0.63 |
| Pipistrellus spp. | 1.00 |
| Nyctalus noctula | 0.25 |
| Nyctalus leisleri | 0.31 |
| Plecotus auritus | 1.25 |
1M. nattereri or any representative of M. daubentonii/mystacinus/brandtii, classified as Myotis sp.
Table 3.
Comparison of bat species composition (proportion of particular taxa among recorded echolocation call sequences) among three habitats, as revealed by ultrasound detectors set simultaneously with mist netting (D-1000X, Mini Bat and Echo Meter Touch devices, exclusively). Lakes – shores of lakes and lagoons, gaps – canopy gaps and clearings.
Table 3.
Comparison of bat species composition (proportion of particular taxa among recorded echolocation call sequences) among three habitats, as revealed by ultrasound detectors set simultaneously with mist netting (D-1000X, Mini Bat and Echo Meter Touch devices, exclusively). Lakes – shores of lakes and lagoons, gaps – canopy gaps and clearings.
| Data | Habitat pair | Χ2 | p |
|---|---|---|---|
| Raw | Lakes - glades | 1224.619 | <0.0001 |
| Lakes - gaps | 799.968 | <0.0001 | |
| Glades - gaps | 336.433 | <0.0001 | |
| Adjusted for detectability | Lakes - glades | 621.029 | <0.0001 |
| Lakes - gaps | 574.69 | <0.0001 | |
| Glades - gaps | 159.295 | <0.0001 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



