
Submitted:
12 December 2023
Posted:
13 December 2023
You are already at the latest version
Abstract
: Intracellular tripeptide glutathione is an important agent of cell survival in hypoxia. Glutathione covalently binds to SH-groups of hemoglobin cysteine residues, protecting them from irreversible oxidation and changes its affinity to oxygen. Reduced glutathione (GSH) can also form a non-covalent complex with hemoglobin. Previously, we showed that hemoglobin tetramer has four noncovalent binding sites of glutathione GSH molecules inside, two of which are released during hemoglobin transition to deoxy form. In this study, we characterized the conserved cysteine residues and residues of non-covalent glutathione binding sites in the sequences of a number of hypoxia tolerant and hypoxia-sensitive mammals. The alpha subunit of all species considered was found to have no conserved cysteines, whereas the beta subunit contains Cys94 residue, which is conserved across species and whose glutathionylation changes the affinity of hemoglobin for oxygen 5-6-fold. In addition, the key residues of noncovalent glutathione binding sites in both alpha and beta subunits are absolutely conserved in all species considered, suggesting a common mechanism of hemoglobin redox regulation for both hypoxia-sensitive and hypoxia-tolerant mammals. The solvent accessibility of all HbA and HbB residues in oxy- and deoxy- forms was analyzed. The key residue betaCys94 is solvent accessible only in the deoxy form.
Keywords:
hypoxia
; hemoglobin
; glutathione
; redox sensitivity
; hypoxia-tolerant species
; alpha subunit
; beta-subunit
; cysteine residues
1. Introduction
Erythrocytes are hemoglobin-containing cells which can bind O2 with hemes – Fe2+ -containing prosthetic groups in lungs and release it in peripheral tissues [1]. Hemoglobin is a well-studied protein having quaternary structure including two pairs of identical subunits.
Subunits of human hemoglobin (Homo SapienceN) (N- is the notation for the normoxy species) are encoded by genes: HBA1 and HBA2 (α-subunit, P69905), HBB (β-subunit, P69905), HBG1 and HBG2 (γ-subunit), HBD (𝛿-subunit, P02042), HBE1 (ε-subunit, P02100), HBZ (ζ-subunit, P02008), HBQ1 (θ-subunit, P09105). (2022). Expression of these genes is the most significant in the bone marrow, the spleen, placenta and other female reproductive tissues (HBD, HBG1, HBG2 only) or male reproductive tissues (HBZ only). Less expression levels of these genes can be detected in liver, lungs, cerebellum and cerebral cortex tissues [2]. Expression and biosynthesis levels of different hemoglobin subunits depends on the age and probably regulated epigenetically [3].
Hemoglobin is very densely packed in erythrocytes and is surrounded by a thin layer of bound water [4]. High concentration of hemoglobin stabilizes its tetrameric quaternary structure [5]. The binding and releasing of oxygen is cooperative due to the Bohr effect. In different stages of development and in different species, hemoglobin binding affinity for O2 and oxygen release in tissues may differ. Most animal species live and reproduce themselves at a normal level of atmospheric oxygen, about 20%. When it decreases, oxidative stress occurs in tissues and it leads to oxidative modifications of proteins and free-radical damage of cell compartments. However, some animal species are resistant to oxidative stress caused by lack of oxygen in their natural environment. One of the widely known ones is the naked mole rat Heterocephalus glaberHR (HR is the notation for the hypoxia-resistant species), which demonstrates exceptional resistance to hypoxemia (~5% O2) [6,7]. These animals can effectively reproduce themselves even at low oxygen concentrations.
There are several identified mechanisms of hypoxia resistance in mammals [6,8]. For example, increased blood vessel density allows Spalax to tolerate to underground hypoxia [9]. Both humans [10] and animals [11,12] living in hypoxic conditions may have elevated hematocrit, hypoxia-inducible factor - 1alpha (HIF-1α), erythropoietin and hemoglobin levels. In humans living on the Tibetan plateau, hemoglobin concentration is independent of altitude, but it rises with altitude in organisms living on the Andean highland [10]. In the Ethiopian highlands, both hemoglobin concentration and arterial oxygen saturation are independent of altitude [13,14]. This means that hemoglobin concentration and arterial oxygen saturation may be an inherited genetic adaptation. Oxygen affinity and saturation are closely related to the properties of hemoglobin molecules in animal species. It is differences in the sequence and, consequently, the structure of hemoglobin that are responsible for differences in the affinity of oxygen binding in the lungs and the efficiency of its release in body tissues.
In recent studies [15,16], we have shown that hemoglobin tetramers in the oxy-form are depots of glutathione, which is partially released during the transition to the deoxy-form and protects tissues under hypoxia. The formation of a non-covalent complex of hemoglobin with glutathione changes the properties of hemoglobin and leads to slight increase of affinity to oxygen [15,16]. Based on our experimental data and molecular docking data we hypothesised that there are two pairs of glutathione binding sites with different affinity in the central cavity of the oxyhemoglobin tetramer. The high affinity pair of binding sites is located at the interface between α- and β-hemoglobin chains, and the second pair of sites with slightly lower affinity is located between the two β-chains [16]. Under deoxygenation two GSH localised in the “pocket” between two beta-globin chains release [16]. In this study, we characterized the conservation of amino acid residues forming all four GSH binding sites in some hypoxia tolerant and hypoxia-sensitive species.
There is evidence that glutathionylation of the Cys93 of HbB leads to a 5-6 fold increase in oxygen affinity [17]. We have shown that Cys93 of HbB in adult hemoglobin is solvent accessible only in deoxyform, residue Cys112 of HbB is accessible in both oxy and deoxy forms, and Cys104 of HbA is unexcessively inaccessible and its glutathionylation has not been confirmed [15,16]. Oxidative stress can lead to the oxidation of GSH to GSSG and SH groups of proteins to SOH and further to SO2 and SO3 (irreversible oxidation). The reduced form of glutathione interacts with SOH group of protein to form disulfide bond ProteinS-SG and a water molecule or oxidized glutathione interacts with SH group of protein results in protein glutathionylation [18]. Such modifications modulate the function of the protein, affecting its activity. Under normal conditions, glutaredoxin remove glutathionylation from cysteine residues, restoring the original function of the protein. Glutathionylated hemoglobin is the marker of oxidative stress of red blood cells [19]. Here we tested the hypothesis that the number of hemoglobin cysteines available to the solvent should correlate with the severity of oxidative stress conditions to which this species is adapted.
Hypoxia tolerant species have a higher affinity of hemoglobin for oxygen and a higher nitrite reductase activity than sensitive species [20,21]. The neotenic theory of hypoxia tolerance suggests that some hypoxia-tolerant animals use isoforms of hemoglobin that are similar to that of neonatal mammals [7]. This assumption looks logical because fetal and embryonic hemoglobins have greater binding affinity. We decided to see what place in the hemoglobin sequence hierarchy the hemoglobin sequences of hypoxemia-resistant mammals occupy and to test this hypothesis.
2. Results
2.1. Hemoglobins hierarhy
We found references to hypoxia-resistant mammals in the literature and compiled a list of hemoglobins of these species (Supplement Table S1).
Looking at the hierarchical tree of hemoglobin sequences of the selected mammalian species (Figure S1 of the Supplement), it can be seen that the alpha-like sequences, with two exceptions, are well classified. Only the Hemoglobin X sequence A7M7S6 of Mus musculusN (Mouse), which fell into the zeta hemoglobin cluster, and the alpha chain sequence A0A8I6AI13 of Rattus norvegicusN (Rat), which fell into the theta chain cluster, are out of the overall harmonious picture.
At first glance, beta-like sequences do not have such a clear separation of chain types, but this is an artefact of the clustering and tree drawing algorithm (Figure S1 of the Supplement). Two clusters of beta chains could well be placed next to each other, and then two clusters of epsilon chains would be next to each other.
A number of conclusions can be drawn from this figure. For example, the gamma chain of haemoglobin J7LGZ1 of Pantholops hodgsoniiHR is closest to fetal hemoglobin beta P02082 of Capra hircusHR and fetal hemoglobin beta P02081 of Bos taurusHR. The delta chains of Homo sapiensN (P02042), Rhinopithecus roxellanaHR (A0A2K6P3G0) and Theropithecus geladaHR (A0A8D2K6V2) were found to be more similar to the beta hemoglobins of different species.
Considering the hypothesis of neonatal origin of hemoglobins of hypoxia-resistant species, we can conclude that the sequences of both beta chains of aquatic mammals Hydrodamalis gigasHR (A0A481WPM8, A0A481WR13) and the single HbB isoform of Trichechus manatus latirostrisHR (A0A2Y9ECT2) are indeed closer to delta chains of other species. Also, one of the four HbB isoforms A0A8C6RUI9 of Nannospalax galiliHR is more similar to epsilon hemoglobins. Two of the six HbB isoforms of Mus musculusN (P04443, P04444) are much more similar to epsilon hemoglobins, e.g. epsilon hemoglobins A0A452F723 of Capra hircusN and A0A6I9I4T5 of Vicugna pacosHR, then to beta hemoglobins.
Some Uniprot classification of sequences looks wrong. For example, theta hemoglobin P04246 of Sus scrofaN looks like epsilon hemoglobins.
The most dissimilar sequences are the hemoglobin sequences zeta (A0A8C2R2Z3) of Capra hircusN, and alpha (G5BXY4) of Heterocephalus glaberHR (two the lowest branches of the phylogenetic tree, Figure S1 of the Supplement).
2.2. Cysteine residues in HbA and HbB sequences
2.2.1. HbA
Interesting, all alpha hemoglobin isoforms of Bos taurusN (P01966) and Bos mutus grunniensHR (P01968, P01967) contain no cysteine at all (Supplement Table S1).
Most HbA sequences have only one cysteine residue at position 104 like in humans. But all HbA isoforms of Pantholops hodgsoniiHR (F6MFG2 and Q0ZA50), Capra caucasica caucasicaHR (A0A7D5Y015), Capra hircus creticaHR (A0A7D5Y174), Capra nubianaHR (A0A7D5Y1A3) and Capra hircusN (P0CH25, P0CH26) have only one cysteine residue at position 111 not 104 (Supplement Table S1).
Alpha hemoglobin sequences of Ailurus fulgensHR (P18969) and Talpa europaeaN (P01951) contain two cysteines at both positions 104 and 111. Rattus norvegicusN alpha hemoglobin isoform A0A8I6GMB4 contain two cysteines at positions 13 and 104. At very different positions 9 and 25, Heterocephalus glaberHR G5BXY2 sequence contain two cysteines.
Two of four probably functional isoforms of Rattus norvegicusN (A0A1K0FUB4, P01946) contain three cysteines at positions 13, 104 and 111. Both alpha hemoglobin isoforms of Tamias umbrinusHR (T1YT26, T1YVM3) contain three cysteines at 13, 104 and 111 positions.
So, positions 13, 104, 111 are common for the most cysteine residues of HbA, and Heterocephalus glaberHR G5BXY2 sequence contain two cysteines at positions 9 and 25. Of these residues, Cys104 and Cys25 are not solvent accessible (Figure S2 of the Supplement) both in oxy and deoxy forms, the remaining residues are solvent accessible and can theoretically undergo oxidation and glutathionylation. Cysteine 104 of the alpha subunit, in both oxy and deoxy forms, contacts with Gln127 of the beta subunit, so we believe it is important for maintaining the stability of the alpha/beta subunit interface. β Cys13 in mouse HbbD beta globins was more susceptible to disulfide exchange with GSSG than β Cys93 [22] by in vitro studies. Also it is known that β Cys13 is able to bind dimethylarsinous acid in rat blood [23]. The function and role of Heterocephalus glaberHR G5BXY2 hemoglobin cysteine residues has not been studied to date.
2.2.2. HbB
All organisms have at least one cysteine at position 93 in hemoglobin beta chains (Supplement Table S1).
At positions 93 and 112, there are cysteines in the sequences of three beta-hemoglobin groups: 1) hominidae and bears Rhinopithecus roxellanaHR (A0A2K6Q8D1), Homo sapiensN (P68871), Theropithecus geladaHR (A0A8D2FZZ9, P02029), Ailurus fulgensHR (P18982), Ursus thibetanusHR (P68012); 2) European hedgehog and mole Erinaceus europaeusN (A0A1S3AE42, A0A1S3WPY1, P02059), Talpa europaeaN (P02061); 3) sealiforms Hydrodamalis gigasHR (A0A481WR13, A0A481WPM8), Dugong dugonHR (A0A481WQB0), Trichechus manatus latirostrisHR (A0A2Y9ECT2).
Two cysteines at positions 93 and 125 appeared in two groups of beta-hemoglobin sequences: five of the eight potential isoforms of Rattus norvegicusN (A0A0G2JTW9, P02091, A0A8I5ZXF1, A0A8L2R1Q6, A0A8L2R6L7) and both two isoforms of Cavia porcellusHR (P02095, A0A7T7JJ63).
Positions 13 and 93 locate cysteines in two of the five potentially functional sequences of Mus musculusN (P02088, P02089) beta hemoglobins.
One of the four sequences of Nannospalax galiliHR HbB (A0A8C6RII4) is the most highly divergent. It contains cysteines at positions 13, 32, 50, and 93.
In the beta chain, Cys93 and Cys32 residues are solvent accessible only in deoxy- form, and Cys112 is not solvent accessible both in oxy and deoxy forms (Figure S3 of the Supplement).
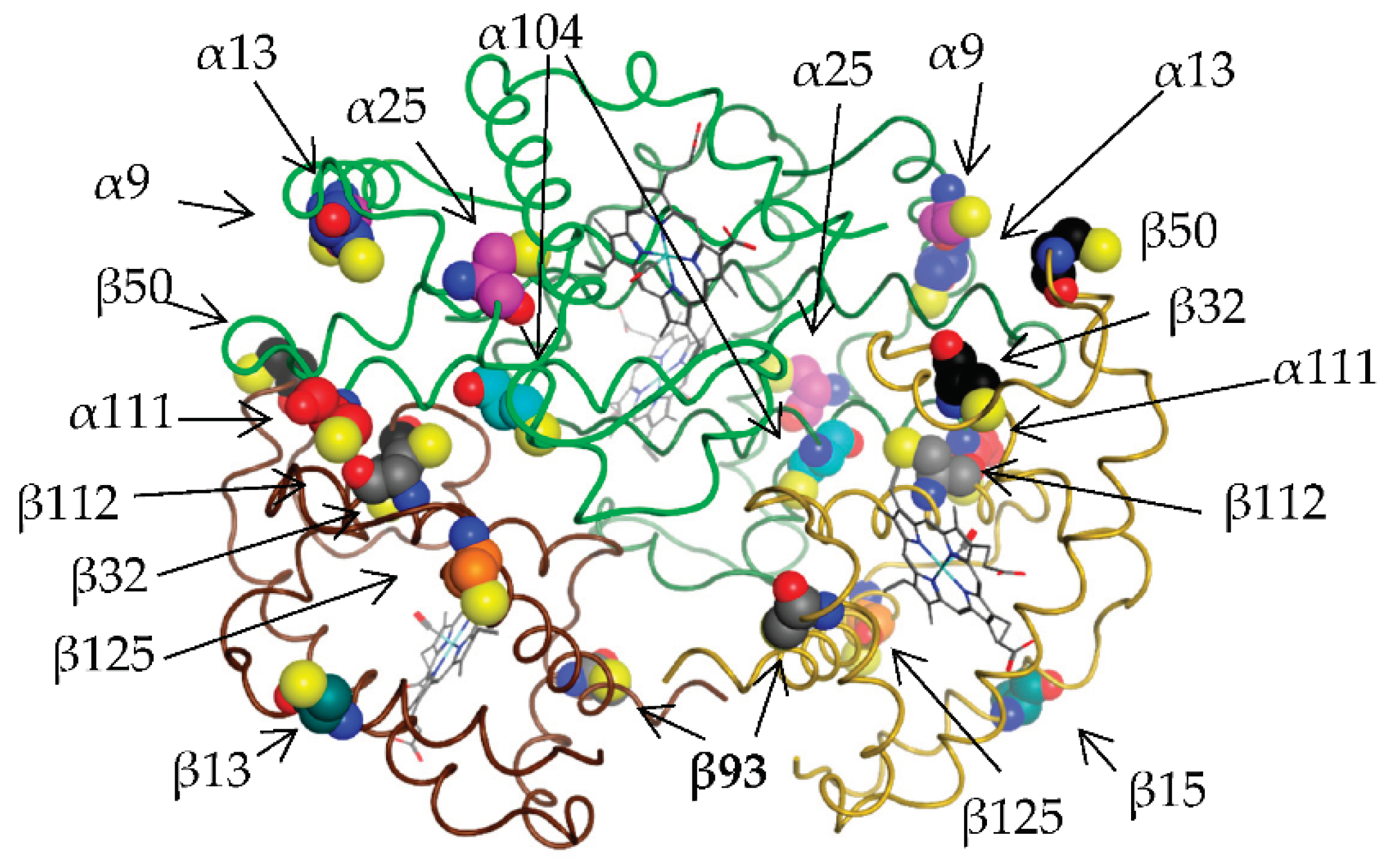
Figure 1.
Spatial arrangement of all possible cysteine residues in the structure of haemoglobin tetramer (on the structure of the oxyform of human hemoglobin 1r1x). Alpha chains are shown with green and dark green line, beta chains with brown and light brown line. The alpha chains show residues 9, 13, 25, 104 and 111. The beta chains show residues 13, 32, 50, 93, 112 and 125. Buried and inaccessible to solvent are residues α104 and β32, the rest are solvent accessible and can be glutathionylated.
Figure 1.
Spatial arrangement of all possible cysteine residues in the structure of haemoglobin tetramer (on the structure of the oxyform of human hemoglobin 1r1x). Alpha chains are shown with green and dark green line, beta chains with brown and light brown line. The alpha chains show residues 9, 13, 25, 104 and 111. The beta chains show residues 13, 32, 50, 93, 112 and 125. Buried and inaccessible to solvent are residues α104 and β32, the rest are solvent accessible and can be glutathionylated.
2.3. GSH binding pockets conservativity
In our recent work, we found that human oxyhemoglobin can bind four glutathione and identified four GSH binding sites inside the adult oxyhemoglobin tetramer of Homo sapienceN using molecular modeling [16]. Two symmetric sites for oxy- and deoxyhemogemobin include hemoglobin alpha-chain's residues Val1, Leu2, Pro95, Val96, Phe98, Lys99, Ser102, His103, Leu106, Asp126, Lys127, Leu129, Ala130, Ser131, Ser133, Thr134, Thr137, Ser138, Tyr140, among which residues Leu2, Pro95, His103, Leu106, Asp126, Lys127 are absolutely conserved in all organisms, both normoxia and hypoxia-resistant organisms, and residues of beta-chains Val34, Tyr35, Trp37, Arg104, Leu105, Leu106, Asn108, Val109, among which residues Val34, Trp37, Leu105, Leu106 are absolutely conserved, and Tyr35 is replaced by His only in one of four beta-chain isoforms A0A8C6RII4 of Nannospalax galiliHR, and Asn108 is replaced by Asp only in one of three beta-chain isoforms G5BS33 of Heterocephalus glaberHR (Supplement Figures S2, S3).
Two GSH binding sites (in the oxy form only) are located at the exit line from the haemoglobin tetramer cavity between the two beta subunits and include only beta chain residues: Val1, Leu81, Lys82, Glu101, Phe103, Arg104, Gly136, Ala138, Asn139, Ala140, Ala142, His143, His146, of which Val1, Leu81, Phe103, Gly136, Ala138, Ala140, His146 are conserved in all organisms, both normoxia- and hypoxia-resistant, with Lys82 replaced by Asn in both two HbB isoforms A0A481WR13 and A0A481WPM8 of Hydrodamalis gigasHR, and His143 replaced by Ala in all four known HbB isoforms A0A1S3WPY1, P02059, A0A1S3ADW1, A0A1S3AE42 of Erinaceus europaeusN.
3. Discussion
No clear correlation between hypoxia tolerance and the number of cysteine residues in the alpha and beta chains of hemoglobin was found. No single conserved cysteine residue was found in the alpha chain of hemoglobin (Figure S2). At the same time, a conserved cysteine residue betaCys93 was detected in the beta chain, which is present in all studied species, both hypoxia-resistant and normoxic mammals, regardless of how many cysteine residues (from 1 to 6, Supplement Table S1, Figure S3) are present in the isoform. HbB subunit cysteine 93 is known to have several different physiological functions, including protection of iron oxidation by peroxide binding [24], binding of endogenic NO and hydrogen sulfide [25,26], the regulation of dimeric-tetrameric Hb equilibrium[27] and is capable to bind glucose moiety [28]. A perfect review of the role of Cys93 is given by Alayash [25]. Site-directed mutagenesis studies in which βCys93 was replaced by other amino acids (exhibiting different degrees of hydrophobicity) or chemically modified with sulfhydryl reagents unequivocally confirmed the critical role played by this residue in maintaining structural-functional integrity and O2 binding properties of Hb [29]. Other cysteine residues in hemoglobin are not as well studied. In vitro studies showed that β Cys13 in mouse beta hemoglobins was more susceptible to disulfide exchange with GSSG than β Cys93 [22].
A marker of oxidative stress in erythrocytes is glutathionylation of hemoglobin [30], which not only protects protein thiol groups but also changes the affinity of hemoglobin for oxygen. It is important to note that cysteine 93 is the main glutathionylated residue of human hemoglobin, the glutathionylation of which leads to a 5-6-fold increase in the affinity of hemoglobin for oxygen, reducing the cooperativity of binding [17,18,31]. The appearance of glutathionylated hemoglobin with increased affinity to oxygen leads to the fact that such hemoglobin will not only better bind oxygen in the lungs at lower concentrations of oxygen in the air, but also give oxygen at higher concentrations of carbon dioxide and lower concentrations of oxygen in tissues, and, therefore, will be able to deliver oxygen to the farthest, most hypoxic tissues. When oxidative stress increases, the percentage of glutathionylated forms of hemoglobin will increase, improving tissue supply. We believe that this is a mechanism of adaptation of normoxia-resistant species to hypoxia conditions. Hypoxia-resistant species, due to this mechanism, can also expand the range of oxygen and carbon dioxide concentrations acceptable for life. Thus, hemoglobins of both hypoxia-resistant and hypoxia-unstable species are capable of glutathionylation by Cys93, which indicates the existence of a single mechanism of redox regulation of hemoglobin function during the development of oxidative stress in hypoxia-resistant and nonresistant species.
Testing of the neotenic theory hypoxia tolerance [7] has shown that sequences of HbB subunits of some mammalian species, in particular aquatic mammals Hydrodamalis gigasHR and Trichechus manatus latirostrisHR , and some HbB isoforms of Nannospalax galiliHR and Mus musculusN are indeed more similar to fetal and embryonic sequences of other mammals than to adult HbB sequences, but this observation is not widespread. In the other hypoxia tolerant mammalian species considered, the change in hemoglobin affinity for oxygen arose by independent pathways from the adult form of hemoglobin. The adult-type alpha subunit replaces the fetal form at about eight weeks of human fetal age, and the adult version of the beta chain of hemoglobin completely displaces the fetal forms by about 3.5 months of newborn age [3]. This observation is in line with the fact that adult HbA sequences have no similar fetal or embryonic forms of alpha-like subunits among adult HbA sequences of hypoxia tolerant animals. In addition, there are no conserved cysteine residues in HbA, indicating that there is no common mechanism of redox regulation of hemoglobin function by cysteines of the alpha subunit.
The ability of oxyhemoglobin to bind glutathione noncovalently and release it upon deoxygenation [16] also plays an important role in protecting erythrocytes from oxidative stress when passing through tissues with low deoxygenation. Formation of a complex with GSH leads to changes in a number of properties of human hemoglobin, including changes in the heme environment, the environment of tryptophan residues, decreased thermostability of the protein [15], and a slight increase in oxygen affinity [16]. The binding sites of glutathione to oxy- and deoxyhemoglobin were prescanned using molecular docking, based on experimental data indicating that the oxyform binds 4 and the deoxy 2 GSH molecules [16]. Data on thermodynamic parameters of binding indicate that hydrophobic interactions play an important role in the formation of the complex, which correlates well with the modeling data, which testify to the participation of hydrophobic residues in the formation of the complex [16]. In addition, experimental data on a significant change in tryptophan fluorescence confirmed our data [16] that Trp37 (which makes the main contribution to tryptophan fluorescence of hemoglobin) takes part in the formation of the complex [15]. Conserved glutathione binding sites suggest that the mechanism of glutathione release during deoxygenation is also realized in hypoxia-resistant animals. This may play an important role in case of abrupt change of oxygen level in the environment, for example, when a burrowing animal goes to the surface, which leads to oxidative stress in tissues. GSH glutathione is required to protect red blood cells from such tissue stress. It should be noted that it has been shown on human hemoglobin that complexing with GSH increases the affinity of hemoglobin for oxygen by 15% [16]. The release of glutathione from the complex with hemoglobin, thus, reduces the affinity of hemoglobin to oxygen and promotes the release of oxygen into the tissues of the body.
4. Materials and Methods
4.1. Hemoglobin sequences dataset
Some animal species are more resistant to hypoxia than others. This feature allows them to live and reproduce themselves in high-altitude areas, soil and other environments with a low oxygen concentration. Species with known resistance to hypoxia include: Ailurus fulgensHR [32], Bos mutus grunniensHR [33], Canis lupus chancoHR [34], Capra caucasica caucasicaHR, Capra nubianaHR [35], Cavia porcellusHR [36], Chinchilla lanigeraHR [37], Colobus guerezaHR [38], Ctenomys rionegrensisHR [39,40], Heterocephalus glaberHR [6], Lama guanicoeHR [41], Marmota flaviventrisHR [42], Ochotona curzoniaeHR [43], Pantholops hodgsoniiHR [35], Peromyscus maniculatusHR [44], Procavia capensis habessinicaHR [45], Rhinopithecus roxellanaHR [46], Spalacopus cyanusHR [39,40], Spalax ehrenbergiHR [8], Talpa europaeaHR [47], Tamias umbrinusHR [48], Theropithecus geladaHR [49], Ursus thibetanusHR [50], Vicugna pacosHR [51]. We selected all hemoglobin sequences of these species from the Uniprot database (https://www.uniprot.org/). In addition, we took hemoglobin sequences of normoxia species for comparison: Homo sapienceN, Bos TaurusN, Sus scrofaN, Capra hircusN, Mus musculusN, Rattus norvegicusN, Erinaceus europaeusN. A complete list of the hemoglobin sequences reviewed is provided in the Supplement Table S1. In the text normoxic species are indicated by an upper "N" after the species name, hypoxia-resistant species by "HR".
We collected all annotated hemoglobin sequences for all these species (listed in Table S1 of Supplement) from Uniprot database (www.uniprot.org, September 2023).
4.1.1. Alpha hemoglobins
We believe that some of the abovementioned hemoglobin sequences hardly exist in erythrocytes as part of the hemoglobin tetramer in reality, and if they do, they exist only in the form of a monomer and perform another function. For example, the sequences of hemoglobin alpha A0A0A0MP82 and A0A8L2QLW7 of Rattus norvegicus have an insertion after the 94th residue (by numbering of human hemoglobin alpha in alignement) of 43 amino acid residues. Structure modelling showed (Supplement Figure S1) that an insertion at this position of even a small number of amino acid residues disrupts the normal interaction interface with the beta subunit, and an insertion of 43 residues makes complex binding with the beta subunit completely impossible. These sequences were predicted from the rat genome sequence and found as transcripts in the brain [52], but their translation and the presence of a protein product have not been proven. It is also possible that the 43 residue insertion is an intron that should be removed during RNA processing. Such hemoglobin sequences, whose existence in the hemoglobin tetramer seems doubtful, are shown in gray in the Table S1 of the Supplement.
The A0A8I5ZYH2 and A0A8I6AWV0 sequences of Rattus norvegicus contain an extended insertion at the N-terminus of 45 amino acid residues, which, according to structure-by-homology modelling, would not interfere with interaction with the beta chain and tetramer association, but would interfere with tight packing of hemoglobin in the erythrocyte. Also the sequences of alpha-hemoglobin of Sus scrofa A0A4X1UM57, A0A4X1UMN0 and A0A4X1UMN5 have extended insertions of 88 amino acids at the N-terminus. All of these sequences were excluded from further consideration. Multiple alignments and an evolutionary tree were constructed using the remaining alpha chain sequences (Supplement, Figure S2).
4.1.2. Beta hemoglobins
We excluded the sequences Q9TT33 Colobus guereza and E9Q223 Mus musculus, which lack significant sequence fragments from the N- and C-termini respectively, from the sample.
4.1.3. Mu hemoglobins
We excluded the Sus scrofa sequence A0A5G2QP95 due to long insertion of 50 aminoacids after the 31st residue.
4.2. Hemoglobin ASA measurement
Accessible surface area was measured with the program MOE 2019.0102 software.
4.3. Phylogenetic analyses
For analyses of conserved residues and phylogenetic relationships we used UPGMA clustering by similarity using BLOSUM62 matrix by MOE 2019.0102 software.
5. Conclusions
Our data suggest that there is a common mechanism of redox regulation of hemoglobin function during oxidative stress in hypoxia-resistant and hypoxia-sensitive mammalian species. This mechanism includes deposition by oxyhemoglobin of reduced glutathione molecules, which are partially released during deoxygenation, as well as glutathionylation of betaCys93 under conditions of oxidative stress to produce hemoglobin with higher affinity to oxygen. This mechanism allows the mammalian organism to adapt to unfavorable conditions and increase the range of oxygen and carbon dioxide concentrations suitable for life.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org. Table S1: List of hemoglobins of hypoxia tolerant and sensitive species. Figure S1: Phylogenetic reconstruction of hemoglobin protein sequences by similarity using BLOSUM62 matrix UPGMA clustering; Figure S2: Accessible surface area of amino acid residues of the alpha-subunit of human hemoglobin in the oxy-form (based on the oxy 1r1x(red) and deoxy 1a3n(blue) structures); Figure S3: Accessible surface area of amino acid residues of the beta-subunit of human hemoglobin in the oxy-form (based on the oxy 1r1x(red) and deoxy 1a3n(blue) structures); Figure S4: Sequence alignment of hemoglobin alpha-subunit both hypoxia-tolerant and hypoxia-sensitive species; Figure S5: Sequence alignment of hemoglobin beta-subunit both hypoxia-tolerant and hypoxia-sensitive species.
Author Contributions
Conceptualization, IYP and AAA; methodology, YLO; software, SYS; validation, SYS and IYP; formal analysis, AAA; investigation, YLO; resources, SYS; data curation, AAA; writing—original draft preparation, AAA, SYS, IYP; writing—review and editing, AAA, IYP; visualization, AAA; supervision, IYP; project administration, IYP; funding acquisition, IYP. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by Russian Science Foundation grant #19-14-00374.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Data is contained within the article or supplementary material.
Conflicts of Interest
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
References
- Ciaccio, C.; Coletta, A.; Coletta, M. Role of Hemoglobin Structural-Functional Relationships in Oxygen Transport. Mol. Aspects Med. 2022, 84, 101022. [Google Scholar] [CrossRef]
- Karlsson, M.; Zhang, C.; Méar, L.; Zhong, W.; Digre, A.; Katona, B.; Sjöstedt, E.; Butler, L.; Odeberg, J.; Dusart, P. A Single–Cell Type Transcriptomics Map of Human Tissues. Sci. Adv. 2021, 7, eabh2169. [Google Scholar] [CrossRef]
- Wood, W.G. Haemoglobin Synthesis during Human Fetal Development. Br. Med. Bull. 1976, 32, 282–287. [Google Scholar] [CrossRef] [PubMed]
- Latypova, L.; Puzenko, A.; Poluektov, Y.; Anashkina, A.; Petrushanko, I.; Bogdanova, A.; Feldman, Y. Hydration of Methemoglobin Studied by in Silico Modeling and Dielectric Spectroscopy. J. Chem. Phys. 2021, 155, 015101. [Google Scholar] [CrossRef] [PubMed]
- Reeder, B.J.; Wilson, M.T. Hemoglobin and Myoglobin Associated Oxidative Stress: From Molecular Mechanisms to Disease States. Curr. Med. Chem. 2005, 12, 2741–2751. [Google Scholar] [CrossRef] [PubMed]
- Pamenter, M.E. Adaptations to a Hypoxic Lifestyle in Naked Mole-Rats. J. Exp. Biol. 2022, 225, jeb196725. [Google Scholar] [CrossRef] [PubMed]
- Johansen, K.; Lykkeboe, G.; Weber, R.E.; Maloiy, G.M.O. Blood Respiratory Properties in the Naked Mole Rat Heterocephalus Glaber, a Mammal of Low Body Temperature. Respir. Physiol. 1976, 28, 303–314. [Google Scholar] [CrossRef] [PubMed]
- Li, F.; Qiao, Z.; Duan, Q.; Nevo, E. Adaptation of Mammals to Hypoxia. Anim. Models Exp. Med. 2021, 4, 311–318. [Google Scholar] [CrossRef]
- Shams, I.; Avivi, A.; Nevo, E. Oxygen and Carbon Dioxide Fluctuations in Burrows of Subterranean Blind Mole Rats Indicate Tolerance to Hypoxic–Hypercapnic Stresses. Comp. Biochem. Physiol. A. Mol. Integr. Physiol. 2005, 142, 376–382. [Google Scholar] [CrossRef] [PubMed]
- Beall, C.M. Andean, Tibetan, and Ethiopian Patterns of Adaptation to High-Altitude Hypoxia. Integr. Comp. Biol. 2006, 46, 18–24. [Google Scholar] [CrossRef]
- Arieli, R.; Heth, G.; Nevo, E.; Hoch, D. Hematocrit and Hemoglobin Concentration in Four Chromosomal Species and Some Isolated Populations of Actively Speciating Subterranean Mole Rats in Israel. Experientia 1986, 42, 441–443. [Google Scholar] [CrossRef] [PubMed]
- Shams, I.; Avivi, A.; Nevo, E.; Ivanitskaya, E. Assignment of Erythropoietin (EPO) to Blind Subterranean Mole Rat Chromosome 1q by in Situ Hybridization. Cytogenet. Genome Res. 2005, 108, 362. [Google Scholar] [CrossRef] [PubMed]
- Beall, C.M. Two Routes to Functional Adaptation: Tibetan and Andean High-Altitude Natives. Proc. Natl. Acad. Sci. U. S. A. 2007, 104, 8655–8660. [Google Scholar] [CrossRef] [PubMed]
- Beall, C.M.; Strohl, K.P.; Blangero, J.; Williams-Blangero, S.; Almasy, L.A.; Decker, M.J.; Worthman, C.M.; Goldstein, M.C.; Vargas, E.; Villena, M.; et al. Ventilation and Hypoxic Ventilatory Response of Tibetan and Aymara High Altitude Natives. Am. J. Phys. Anthropol. 1997, 104, 427–447. [Google Scholar] [CrossRef]
- Kuleshova, I.D.; Zaripov, P.I.; Poluektov, Y.M.; Anashkina, A.A.; Kaluzhny, D.N.; Parshina, E.Y.; Maksimov, G.V.; Mitkevich, V.A.; Makarov, A.A.; Petrushanko, I.Y. Changes in Hemoglobin Properties in Complex with Glutathione and after Glutathionylation. Int. J. Mol. Sci. 2023, 24, 13557. [Google Scholar] [CrossRef] [PubMed]
- Fenk, S.; Melnikova, E.V.; Anashkina, A.A.; Poluektov, Y.M.; Zaripov, P.I.; Mitkevich, V.A.; Tkachev, Y.V.; Kaestner, L.; Minetti, G.; Mairbäurl, H.; et al. Hemoglobin Is an Oxygen-Dependent Glutathione Buffer Adapting the Intracellular Reduced Glutathione Levels to Oxygen Availability. Redox Biol. 2022, 58, 102535. [Google Scholar] [CrossRef]
- Craescu, C.T.; Poyart, C.; Schaeffer, C.; Garel, M.C.; Kister, J.; Beuzard, Y. Covalent Binding of Glutathione to Hemoglobin. II. Functional Consequences and Structural Changes Reflected in NMR Spectra. J. Biol. Chem. 1986, 261, 14710–14716. [Google Scholar] [CrossRef]
- Mieyal, J.J.; Gallogly, M.M.; Qanungo, S.; Sabens, E.A.; Shelton, M.D. Molecular Mechanisms and Clinical Implications of Reversible Protein S-Glutathionylation. Antioxid. Redox Signal. 2008, 10, 1941–1988. [Google Scholar] [CrossRef]
- D, G.; I, D.-D.; A, M.; D, B.; A, S.; R, R. Membrane Skeletal Protein S-Glutathionylation in Human Red Blood Cells as Index of Oxidative Stress. Chem. Res. Toxicol. 2019, 32. [Google Scholar] [CrossRef]
- Jensen, F.B. Red Blood Cell pH, the Bohr Effect, and Other Oxygenation-Linked Phenomena in Blood O2 and CO2 Transport. Acta Physiol. Scand. 2004, 182, 215–227. [Google Scholar] [CrossRef]
- Jensen, F.B.; Kolind, R.A.H.; Jensen, N.S.; Montesanti, G.; Wang, T. Interspecific Variation and Plasticity in Hemoglobin Nitrite Reductase Activity and Its Correlation with Oxygen Affinity in Vertebrates. Comp. Biochem. Physiol. A. Mol. Integr. Physiol. 2017, 206, 47–53. [Google Scholar] [CrossRef] [PubMed]
- Hempe, J.M.; Ory-Ascani, J.; Hsia, D. Genetic Variation in Mouse Beta Globin Cysteine Content Modifies Glutathione Metabolism: Implications for the Use of Mouse Models. Exp. Biol. Med. 2007, 232, 437–444. [Google Scholar] [CrossRef]
- Lu, M.; Wang, H.; Li, X.-F.; Arnold, L.L.; Cohen, S.M.; Le, X.C. Binding of Dimethylarsinous Acid to Cys-13α of Rat Hemoglobin Is Responsible for the Retention of Arsenic in Rat Blood. Chem. Res. Toxicol. 2007, 20, 27–37. [Google Scholar] [CrossRef]
- Balagopalakrishna, C.; Abugo, O.O.; Horsky, J.; Manoharan, P.T.; Nagababu, E.; Rifkind, J.M. Superoxide Produced in the Heme Pocket of the Beta-Chain of Hemoglobin Reacts with the Beta-93 Cysteine to Produce a Thiyl Radical. Biochemistry 1998, 37, 13194–13202. [Google Scholar] [CrossRef]
- Alayash, A.I. βCysteine 93 in Human Hemoglobin: A Gateway to Oxidative Stability in Health and Disease. Lab. Invest. 2021, 101, 4–11. [Google Scholar] [CrossRef]
- Kosmachevskaya, O.V.; Nasybullina, E.I.; Shumaev, K.B.; Novikova, N.N.; Topunov, A.F. Effect of Iron–Nitric Oxide Complexes on the Reactivity of Hemoglobin Cysteines. Appl. Biochem. Microbiol. 2020, 56, 512–520. [Google Scholar] [CrossRef]
- O’Neill, J.S.; Reddy, A.B. Circadian Clocks in Human Red Blood Cells. Nature 2011, 469, 498–503. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Bhatt, V.S.; Sun, G.; Wang, P.G.; Palmer, A.F. Site-Selective Glycosylation of Hemoglobin on Cys Β93. Bioconjug. Chem. 2008, 19, 2221–2230. [Google Scholar] [CrossRef]
- Cheng, Y.; Shen, T.-J.; Simplaceanu, V.; Ho, C. Ligand Binding Properties and Structural Studies of Recombinant and Chemically Modified Hemoglobins Altered at Beta 93 Cysteine. Biochemistry 2002, 41, 11901–11913. [Google Scholar] [CrossRef]
- T, N. Protein Glutathionylation and Oxidative Stress. J. Chromatogr. B Analyt. Technol. Biomed. Life. Sci. 2007, 855. [Google Scholar] [CrossRef]
- Mitra, K.; Ubarretxena-Belandia, I.; Taguchi, T.; Warren, G.; Engelman, D.M. Modulation of the Bilayer Thickness of Exocytic Pathway Membranes by Membrane Proteins Rather than Cholesterol. Proc. Natl. Acad. Sci. 2004, 101, 4083–4088. [Google Scholar] [CrossRef]
- Hu, Y.; Thapa, A.; Fan, H.; Ma, T.; Wu, Q.; Ma, S.; Zhang, D.; Wang, B.; Li, M.; Yan, L.; et al. Genomic Evidence for Two Phylogenetic Species and Long-Term Population Bottlenecks in Red Pandas. Sci. Adv. 2020, 6, eaax5751. [Google Scholar] [CrossRef]
- Qiu, Q.; Zhang, G.; Ma, T.; Qian, W.; Wang, J.; Ye, Z.; Cao, C.; Hu, Q.; Kim, J.; Larkin, D.M.; et al. The Yak Genome and Adaptation to Life at High Altitude. Nat. Genet. 2012, 44, 946–949. [Google Scholar] [CrossRef]
- Werhahn, G.; Senn, H.; Ghazali, M.; Karmacharya, D.; Sherchan, A.M.; Joshi, J.; Kusi, N.; López-Bao, J.V.; Rosen, T.; Kachel, S.; et al. The Unique Genetic Adaptation of the Himalayan Wolf to High-Altitudes and Consequences for Conservation. Glob. Ecol. Conserv. 2018, 16, e00455. [Google Scholar] [CrossRef]
- Signore, A.V.; Storz, J.F. Biochemical Pedomorphosis and Genetic Assimilation in the Hypoxia Adaptation of Tibetan Antelope. Sci. Adv. 2020, 6, eabb5447. [Google Scholar] [CrossRef] [PubMed]
- Pairet, B.; Jaenicke, E. Structure of the Altitude Adapted Hemoglobin of Guinea Pig in the R2-State. PloS One 2010, 5, e12389. [Google Scholar] [CrossRef] [PubMed]
- Hopkins, S.R.; Powell, F.L. Common Themes of Adaptation to Hypoxia: Insights from Comparative Physiology. Hypoxia Genes Bedside 2001, 153–167. [Google Scholar]
- Yu, L.; Wang, X.; Ting, N.; Zhang, Y. Mitogenomic Analysis of Chinese Snub-Nosed Monkeys: Evidence of Positive Selection in NADH Dehydrogenase Genes in High-Altitude Adaptation. Mitochondrion 2011, 11, 497–503. [Google Scholar] [CrossRef] [PubMed]
- Tomasco, I.H.; Boullosa, N.; Hoffmann, F.G.; Lessa, E.P. Molecular Adaptive Convergence in the α-Globin Gene in Subterranean Octodontid Rodents. Gene 2017, 628, 275–280. [Google Scholar] [CrossRef] [PubMed]
- Pejo, M.; Tomasco, I.H. Adaptive Evolution of β-Globin Gene in Subterranean in South America Octodontid Rodents. Gene 2021, 772, 145352. [Google Scholar] [CrossRef] [PubMed]
- Storz, J.F. Hemoglobin Function and Physiological Adaptation to Hypoxia in High-Altitude Mammals. J. Mammal. 2007, 88, 24–31. [Google Scholar] [CrossRef]
- Storz, J.F. Hemoglobin–Oxygen Affinity in High-Altitude Vertebrates: Is There Evidence for an Adaptive Trend? J. Exp. Biol. 2016, 219, 3190–3203. [Google Scholar] [CrossRef] [PubMed]
- Luo, Y.; Gao, W.; Gao, Y.; Tang, S.; Huang, Q.; Tan, X.; Chen, J.; Huang, T. Mitochondrial Genome Analysis of Ochotona Curzoniae and Implication of Cytochrome c Oxidase in Hypoxic Adaptation. Mitochondrion 2008, 8, 352–357. [Google Scholar] [CrossRef] [PubMed]
- Storz, J.F.; Cheviron, Z.A.; McClelland, G.B.; Scott, G.R. Evolution of Physiological Performance Capacities and Environmental Adaptation: Insights from High-Elevation Deer Mice (Peromyscus Maniculatus). J. Mammal. 2019, 100, 910–922. [Google Scholar] [CrossRef] [PubMed]
- Signore, A.V.; Paijmans, J.L.; Hofreiter, M.; Fago, A.; Weber, R.E.; Springer, M.S.; Campbell, K.L. Emergence of a Chimeric Globin Pseudogene and Increased Hemoglobin Oxygen Affinity Underlie the Evolution of Aquatic Specializations in Sirenia. Mol. Biol. Evol. 2019, 36, 1134–1147. [Google Scholar] [CrossRef] [PubMed]
- Yu, L.; Wang, G.-D.; Ruan, J.; Chen, Y.-B.; Yang, C.-P.; Cao, X.; Wu, H.; Liu, Y.-H.; Du, Z.-L.; Wang, X.-P. Genomic Analysis of Snub-Nosed Monkeys (Rhinopithecus) Identifies Genes and Processes Related to High-Altitude Adaptation. Nat. Genet. 2016, 48, 947–952. [Google Scholar] [CrossRef] [PubMed]
- Jelkmann, W.; Oberthür, W.; Kleinschmidt, T.; Braunitzer, G. Adaptation of Hemoglobin Function to Subterranean Life in the Mole, Talpa Europaea. Respir. Physiol. 1981, 46, 7–16. [Google Scholar] [CrossRef]
- Revsbech, I.G.; Tufts, D.M.; Projecto-Garcia, J.; Moriyama, H.; Weber, R.E.; Storz, J.F.; Fago, A. Hemoglobin Function and Allosteric Regulation in Semi-Fossorial Rodents (Family Sciuridae) with Different Altitudinal Ranges. J. Exp. Biol. 2013, 216, 4264–4271. [Google Scholar] [CrossRef]
- Chiou, K.L.; Janiak, M.C.; Schneider-Crease, I.A.; Sen, S.; Ayele, F.; Chuma, I.S.; Knauf, S.; Lemma, A.; Signore, A.V.; D’Ippolito, A.M. Genomic Signatures of High-Altitude Adaptation and Chromosomal Polymorphism in Geladas. Nat. Ecol. Evol. 2022, 6, 630–643. [Google Scholar] [CrossRef]
- Zhu, C.; Xu, W.; Li, J.; Liu, C.; Hu, M.; Yuan, Y.; Yuan, K.; Zhang, Y.; Song, X.; Han, J.; et al. Draft Genome Assembly for the Tibetan Black Bear (Ursus Thibetanus Thibetanus). Front. Genet. 2020, 11. [Google Scholar] [CrossRef]
- Storz, J.F.; Moriyama, H. Mechanisms of Hemoglobin Adaptation to High Altitude Hypoxia. High Alt. Med. Biol. 2008, 9, 148–157. [Google Scholar] [CrossRef]
- Richter, F.; Meurers, B.H.; Zhu, C.; Medvedeva, V.P.; Chesselet, M.-F. Neurons Express Hemoglobin Alpha- and Beta-Chains in Rat and Human Brains. J. Comp. Neurol. 2009, 515, 538–547. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



