
Submitted:
12 December 2023
Posted:
13 December 2023
You are already at the latest version
Abstract
As yin and yang, cancer and diabetes are metabolic disorders characterized by deregulated, yet opposing, metabolic alterations. These metabolic shifts reflect a complex network of defective underlying signaling pathways, which ultimately orchestrate both disease pathogenesis and progression. The Warburg effect portrays the chief metabolic phenotype in cancer and is reflected by the dominance of aerobic glycolysis over minimal-to-no viable oxidative phosphorylation in cancer. Conversely, during the pathogenesis of type 2 diabetes (T2D), oxidative phosphorylation is constantly heightened. In advanced insulin resistance and T2D, the metabolic landscape becomes more complex, with exacerbating metabolic alterations affecting oxidative phosphorylation in a tissue-specific manner. Promising nutritional adjuvants, such as alpha-lipoic acid and flavonoids, can reprogram metabolism and exert antitumor and antidiabetic effects. Other pharmacological agents, such as metformin, have therapeutic effects on cancer and T2D. Herein, we discuss the molecular mechanisms and signaling pathways involved in the metabolic shifts in cancer and T2D, and explore the therapeutic strategies under investigation that aim to reverse the metabolic shift in both disease scenarios.
Keywords:
oxidative phosphorylation
; cancer
; mitochondria
; metabolic shift
; type 2 diabetes
; insulin resistance
; therapy
; nutritional adjuvants
; glutaminolysis
1. Introduction
Cellular metabolism involves a series of enzyme-driven biochemical reactions that generate or consume energy. The activity and speed of these reactions fluctuate constantly. These fluctuations are governed by diverse cellular energy requirements, proliferative activities, environmental stressors, and overall functions. Nevertheless, metabolism is now perceived in much broader ways than merely biochemistry; it pervades all facets of biology [1,2].
Under healthy conditions, cells can balance anabolism, catabolism, and waste removal via monitoring and coordinating different metabolic pathways. In various disease states, this intricate balance is lost, resulting in altered metabolism. Usually, genetic reprogramming underlies dysfunctional metabolic switching in cells and tissues. These perturbing shifts in metabolism are different in each disease. Here, we highlight the paradox of the metabolic shift in cancer versus type 2 diabetes (T2D), and its implications in targeted therapy [1,2].
In 1923, Otto Warburg first postulated the Warburg phenomenon, suggesting metabolic rewiring to be one of the hallmarks of cancer, after observing that tumors demonstrate increased glucose uptake. He further hypothesized that cancer cells, due to dysfunctional mitochondria, primarily utilize aerobic glycolysis instead of oxidative phosphorylation (OXPHOS) for rapid energy release, which is required by proliferating cells [3,4]. The Warburg effect involves cytoplasmic anaerobic fermentation of glucose into lactate, despite regular oxygen availability. Aerobic glycolysis ultimately increases cellular anabolism and decreases catabolism. Our knowledge of this phenomenon and its driving forces has been refined and expanded over the past decades. Nevertheless, the two following features remain unaltered with regard to the Warburg effect: increased glucose uptake and lactate production [5,6]. Aerobic glycolysis is markedly heightened in over 70% of cancer types such as lung [7], breast [8], liver [9,10], brain [11], prostate [12], gynecologic [13], and pancreatic cancer [3,14,15]. Similar to solid tumors, hematologic malignancies, such as lymphomas [15,16,17] and leukemias [15,18,19], also demonstrate high aerobic glycolysis and low OXPHOS rates. In certain tumors, the accelerated Warburg effect occurs even in the presence of active or partially active mitochondrial OXPHOS [20,21,22,23]. It is argued that in cancer, minimal activity of mitochondrial OXPHOS is crucial for tumor cell survival [21,24,25,26] and metastasis [27]. As discussed in further sections, accumulating evidence suggests additional reasons why mitochondrial dysfunction plays a role in the preferential glycolytic shift in tumor cells.
In addition to the Warburg effect, cancer cells simultaneously adopt another metabolic pathway called glutaminolysis as part of a metabolic reprogramming strategy to meet their specific energy and biosynthetic demands. With heightened energy demands, cancer cells absorb and utilize more glutamine than normal cells, supplementing glucose as an additional energy source [28]. Glutaminase converts glutamine to glutamate and ammonia [29]. The resultant glutamate enters the tricarboxylic acid (TCA) cycle within mitochondria, supporting energy production and biosynthetic precursor synthesis [30]. Notably, increased glutaminolysis in cancer cell mitochondria induces a metabolic shift from canonical oxidative phosphorylation and ATP production to the synthesis of anabolic intermediates for lipid and amino acid production. Malate, an intermediate in the TCA cycle, is metabolized to pyruvate and lactate, while citrate contributes to lipid metabolism. Both processes generate NADPH molecules, countering specific reactive oxygen species and averting oxidative stress [31]. Furthermore, glutamine serves as a crucial nitrogen source for nucleotide biosynthesis in the cytosol. In purine biosynthesis, two glutamine molecules provide nitrogen atoms for the purine ring formation in inosine monophosphate, a precursor to both adenosine monophosphate and guanosine monophosphate [32]. In pyrimidine biosynthesis, one glutamine molecule contributes the nitrogen atom for the formation of cytidine triphosphate from uridine triphosphate [33]. Collectively, glutamine acts as a signaling molecule, activating essential pathways that promote survival, proliferation, and differentiation. In certain cancers, dysregulation of these pathways may contribute to cancer development and metastasis.
In contrast, the complex metabolic changes underlying T2D are different from those underlying cancer. In established T2D, insulin resistance arises in peripheral tissues, primarily in the skeletal muscle (SKM) [34], and decreases glucose-induced insulin secretion by pancreatic β cells [35,36,37,38]. In healthy SKM, insulin increases the mitochondrial capacity for OXPHOS via an increased expression of mitochondrial OXPHOS-related genes and the posttranslational modification of mitochondrial proteins in the form of phosphorylation [39,40,41,42]. Hyperglycemia induces the release of insulin, which activates mitochondrial respiration [43]. However, chronic hyperglycemia in individuals, due to continuous nutritional overload and decreased physical activity, leads to prolonged hyperactivity of the OXPHOS machinery. This is associated with a consistently excessive release of reactive oxygen species (ROS), leading to oxidative toxicity and insulin resistance in peripheral tissues, eventually resulting in the development of T2D [44,45,46]. However, in established T2D, contradicting findings have been reported on mitochondrial OXPHOS in SKM [41]. Several researchers reported mitochondrial dysfunction and low OXPHOS in SKM [47,48], while others reported normal OXPHOS [49,50,51]. Few studies reported that the liver exhibited normal to even increased mitochondrial OXPHOS [52,53,54]. Some researchers argue that mitochondrial dysfunction contributes to the development of insulin resistance and T2D [47,48,55]. However, the opposite is more often believed to be true, i.e., insulin resistance leads to mitochondrial dysfunction in peripheral tissues [41,56]. In contrast, in T2D, a Warburg-like effect and lactate production also occurs in pancreatic β cells [57,58]. The released lactate could cause insulin resistance by suppressing glycolysis and impairing insulin signaling in SKM [59].
In this review, we explore the metabolic paradigms in cancer and T2D and highlight a few promising nutrigenetic and therapeutic approaches that aim to reverse the metabolic shift and restore a normal balance in both diseases.
2. The Metabolic Shift in Cancer
2.1. Why Do Tumors Adopt Glycolysis over OXPHOS?
Both anaerobic glycolysis and OXPHOS produce cellular energy in the form of adenosine triphosphate (ATP). OXPHOS is much more efficient in generating ATP than glycolysis; it generates approximately 32 ATPs from a single glucose molecule, whereas glycolysis produces only a net of two ATPs (see Figure 1). Briefly, during glycolysis, a single molecule of glucose is converted into two molecules of pyruvate through a series of biochemical reactions that ultimately result in the production of two ATPs, after consuming two ATPs in the glycolytic process. The resultant pyruvate can either enter the Krebs cycle (tricarboxylic acid (TCA) cycle) followed by OXPHOS in the mitochondria in aerobic conditions or it can be converted to lactate in anaerobic conditions. OXPHOS is an oxygen-dependent process that combines the oxidation of nicotinamide adenine dinucleotide (NADH) and flavin adenine dinucleotide (FADH2) with the phosphorylation of ADP to form ATP. In contrast, in anaerobic glycolysis, the conversion of pyruvate to lactate consumes NAD+ to generate NADH. The biochemical landscapes of glycolysis, aerobic glycolysis, and the TCA cycle have been well reviewed by Akram in 2013 [60,61]. Another recent review by Shiva et al. (2020) elegantly describes OXPHOS, which is also known as the electron transport chain (ETC) cycle [62]. The selection of a less efficient metabolic pathway by the cell is attributed to the fast-track generation of ATP by glycolysis, when compared to OXPHOS. Therefore, it was believed that an increase in the rate of aerobic glycolysis would promptly deliver the energy needs of the cell [22,63,64,65,66]. In that sense, an increase in the frequency of aerobic glycolysis translates into an increase in glucose uptake [67] and rapid ATP synthesis. However, this explanation was found to be insufficient to justify the glycolytic surge in tumors even in the presence of functional mitochondria. There are other reasons and theories that emerged, which are discussed in the following sections (see Figure 2). It is worth pointing out that another theory of altered energy metabolism in tumor cells exists and is termed the reverse Warburg effect or tumor symbiosis. This theory highlights a crosstalk between hypoxic tumor cells and normoxic stromal cells, in which the stromal cells take up the lactate generated by the hypoxic tumor cells and use it as fuel to generate ATP via oxidative mechanisms. The ATP generated by normoxic cells then becomes a source of energy for the neighboring hypoxic tumor cells [68]. This lactate shuttle and the interplay between the heterogenic metabolic phenotypes in the core and microenvironment of the tumor assist in the survival of tumor cells [68,69,70,71].
In the next subsections, we summarize the reasons behind the adoption of glycolysis over OXPHOS in tumor cells.
2.1.1. Reason 1: Mitochondrial Dysfunction
Despite controversies around the presence of functional or partially functional mitochondria in some cancers, we believe these observations are an exception rather than the standard. Discovering signs of mitochondrial activity in some cancer phenotypes does not equate to the presence of normally functional mitochondria. Mitochondrial dysfunction continues to be the leading cause of predominantly occurring glycolysis over OXPHOS in tumor cells. Several mechanisms contribute to the development of mitochondrial dysfunction in tumor cells (see Figure 3); this is fortified by the mutations or transcriptomic dysregulations found in the genes that encode OXPHOS- and glycolysis-related proteins (Table 1 and Table 2).
OXPHOS-related genes are either of mitochondrial DNA (mtDNA) or nuclear DNA (ncDNA) origin. Point mutations are the most common type of mutations in mtDNA reported to date. Even more interesting are the mutations occurring in oncogenes and tumor suppressor genes, which are frequent and characteristic to all cancer types. Many oncogenes and tumor suppressor genes indirectly regulate glycolysis and OXPHOS by regulating the expression of OXPHOS-related or glycolytic proteins (Table 3). Moreover, mitochondrial metabolic dysfunction in cancer can also result from the dysregulation of mitochondrial biogenesis and mitophagy. Regarding cancer, there are other mutations in ncDNA genes that encode TCA cycle-related enzymes, such as succinate dehydrogenase (SDH) [72], fumarate hydratase (FH) [73], and isocitrate dehydrogenase (IDH) [74,75], which have a direct effect on OXPHOS [76] (Table 1). For instance, Bourgeron et al. reported — for the first time —that a mutation in the SDH gene caused mitochondrial ETC deficiency [77]. In 2018, Böttcher et al. discovered that a gain-of-function mutation in IDH leads to enhanced D-2HG production, which triggers the destabilization of the hypoxia-inducible factor-1 alpha (HIF-1α) protein, thus making the cell more dependent on OXPHOS [78].
Likewise, tumorigenic mutations in cardinal oncogenes and tumor suppressor genes significantly contribute to mitochondrial dysfunction in cancer. Although oncogenes and tumor suppressor genes do not directly encode OXPHOS-related or glycolytic proteins, they indirectly regulate the activity of these proteins through cell signaling pathways (Table 2). This strongly abates the rationale claiming the presence of functional mitochondrial OXPHOS in cancer. Common oncogenes and tumor suppressor genes involved in the mitogen-activated protein kinase (MAPK), phosphoinositide 3-kinase (PI3K), and mammalian target of rapamycin (mTOR) pathways have been heavily reported in most cancer types and can affect mitochondrial function (Table 2).
Mitochondrial deregulation is also manifested by an imbalance in the degree of biogenesis of mitochondrial organelles and mitophagy of unhealthy mitochondria. Imbalances in mitochondrial organelle turnover results in an abnormal number of available mitochondrial organelles in the cytoplasm, which have been implicated in cancer progression [79,80]. Autophagy is inhibited by mTOR pathway activity. An active AMP-activated protein kinase (AMPK) pathway inhibits the mTOR pathway, thereby activating autophagy. Autophagy promotes cell survival by recycling cellular organelles to produce energy. Reportedly, there is an association between high autophagic activity and increased cancer resistance to chemotherapy [81,82]. Mitophagy, on the other hand, degrades mitochondria through either the PTEN induced kinase 1 (PINK1)/Parkin (PRKN) or the BCL2 interacting protein 3 (BNIP3)/NIP-3-like protein X (NIX)/FUN14 domain containing 1 (FUNDC1) pathways, or by AMPK activation and consequent phosphorylation of Unc-51 like autophagy activating kinase 1 (ULK1). Interestingly, some studies have shown that the downregulation of PRKN causes a decrease in mitophagy and the accumulation of dysfunctional mitochondria in the cytoplasm. This has been associated with decreased mitochondrial OXPHOS, increased ROS, and increased glycolysis. Therefore, PRKN deficiency contributes towards the Warburg effect in cancer. PRKN deficiency has been observed in several cancer phenotypes in humans, including colorectal cancer [83], glioblastoma [84], melanoma [85], lung cancer [86], and breast cancer [79,87].
Some studies have suggested that the overexpression of uncoupling protein (UCP) promotes aerobic glycolysis, tumor proliferation, and resistance to apoptosis-induced chemotherapy [88,89,90]. UCPs are a family of mitochondrial proteins localized in the mitochondrial membrane that act as anion transporters. UCP2, in particular, is ubiquitously expressed in the body and plays several biological functions and has been shown to play a role in both tumorigenesis and chemoresistance. UCP2 has an antioxidant effect due to its role in transporting protons from the inner mitochondrial membrane to the inner mitochondrial matrix [91]. In 2016, Brandi et al. demonstrated that UCP2 caused the downregulation of OXPHOS-related Complex I (NADH dehydrogenase), Complex IV (cytochrome c oxidase), Complex V (ATPase), and a decrease in mitochondrial oxygen consumption [90].

Table 1.
Common alterations in the genes involved in the TCA cycle and OXPHOS metabolism in cancer.
| Gene | Encoding DNA | Protein | Cycle | Reported dysregulation in cancer | Publications |
|---|---|---|---|---|---|
| Aco2 | Nuclear | Aconitase 2 | TCA (Krebs cycle) |
|
[76,92] |
| IDH1 | Nuclear | Isocitrate Dehydrogenase 1 | TCA | Point mutations | [74,76,93,94,95,96,97] |
| SDH | Nuclear | Succinate Dehydrogenase | TCA and ETC cycle |
|
[72,76,98,99,100,101,102,103,104,105,106] |
| FH | Nuclear | Fumarate Hydratase | TCA |
|
[73,76,107,108,109,110,111] |
Table 2.
Common oncogenes and tumor suppressor genes implicated in mitochondrial dysfunction in cancer.
Table 2.
Common oncogenes and tumor suppressor genes implicated in mitochondrial dysfunction in cancer.
| Gene | Class | Genetic alteration | Pathway affected | Effects on OXPHOS (ETC cycle) | The effect of cancer progression | References |
|---|---|---|---|---|---|---|
|
MYC (MYC proto-Oncogene Protein) |
Oncogene | Point mutation, amplification | TGF-β Signaling Pathway | Stimulates mitochondrial biogenesis and function through regulating Transcription Factor A Mitochondrial gene | Self-sufficiency in growth status | [76,112,113,114] |
|
AKT (Alpha Serine/Threonine Kinase) |
Oncogene | Point mutation, amplification, overexpression | AKT pathway |
|
Evade apoptosis | [113,115,116] |
| P53 | Tumor Suppressor gene | Point mutation, Deletion | P53 pathway, Cell Cycle Control: G2/M DNA Damage Checkpoint |
|
Evade apoptosis, insensitivity to anti-growth signals | [113,117,118] |
|
PI3K (Phophatidylinositol-4,5-Bisphosphate 3-Kinase) |
Tumor Suppressor | Point mutation | AKT pathway |
|
Evade apoptosis | [113,119] |
|
PTEN (Phosphatase and Tensin Homolog) |
Tumor Suppressor | Point mutation, deletion | PI3K pathway |
|
Evade apoptosis | [113,120] |
|
MDM2 (Mouse Double Minute 2, Human Homolog Of; P53-Binding Protein) |
Oncogenes | Amplification | Cell Cycle Control: G1/S Checkpoint |
|
Evade apoptosis | [113,121] |
|
BRAF (B-Raf Proto-Oncogene, Serine/Threonine Kinase) |
Oncogenes | Point mutation, amplification, increased expression | MAPK pathway (RAS) | BRAF upregulation inhibits oxidative phosphorylation gene transcription, mitochondrial b, biogenesis, and the expression of PGC1a by targeting the melanocyte lineage factor (MITF). | Self-sufficiency in growth status | [113,122] |
|
KRAS (Kirsten rat sarcoma viral oncogene homolog, GTPase) |
Oncogene | Point mutation | MAPK pathway | KRAS activation of MAPK and PI3K pathways stabilizes and activates hypoxia-inducible factors-1 alpha and -2 alpha (HIF-1α and HIF-2) which facilitates ischemic adaptation. KRAS stimulates aerobic glycolysis by overexpressing Hexokinase, lactate dehydrogenase, and glucose transporters. KRAS induces glutaminolysis by upregualting Glutamate Oxaloacetate Transaminase 1,2. (GOT), leading to aspartate and NADPH generation, and the activation of the NRF2 antioxidant system. Upregulation of RAS leads to increased autophagy and micropinocytosis contributing to the disruption of cellular energy balance and nutrient scavenging. |
Self-sufficiency in growth status | [113,123,124,125,126] |
|
NF-κB (Nuclear Factor Kappa B) |
Oncogene | Amplification, rearrangement, chromosomal translocation in several members of the NF-κB protein family or constitutional activation of NF-κB | NF-κB pathway | NF-κB upregulation and activity cause a decline in mitochondrial respiratory capacity and reduces the expression of key mitochondrial proteins including SDHA, ANT-1, UCP3, and MFN2 and causes increased fission and mitophagy of mitochondrial organelle. It upregulates PGC1α and correlates with high ROS. | Tumor growth | [113,127,128] |
| EGFR (ErbB1 Epidermal Growth Factor Receptor) | Oncogene | Amplification, upregulation | PI3K and MAPK pathway | EGFR modulates mitochondrial function through modification of Cox-II | Self-sufficiency in growth status | [113,129] |
|
IGFR (Insulin-like Growth factor receptor) |
Oncogene | Amplification | AKT, PI3K, and MAPK pathways | Increased IGFR expression alters ATP synthesis and increases mitochondrial function. And decrease mitochondrial ROS production associated with the induction of antioxidant response. | antiapoptotic, cell-survival, and transforming activities |
[113,130] |
|
ErbB2 (HER2, Receptor tyrosine-protein kinase erbB-2 ) |
Oncogene | Amplification | MAPK, PI3K, AKT, and mTOR | ErbB2 overexpression causes downregulation of pro-apoptotic Bcl-2 family protein (Bcl-xS) and increases levels of anti-apoptotic Bcl-xL. This leads to mitochondrial dysfunction and a loss of mitochondrial membrane potential, a 35% decline in ATP levels, and a loss of redox capacity (mitochondrial reductase activity). | Anti-apoptotic and pro-proliferative effects | [113,131] |
|
HIF-1 α (Hypoxia Inducible Factor 1 Subunit Alpha) |
Oncogene | It is stabilized and activated in hypoxic tumor conditions and by inactivating mutations of SDH, FH, IDH as well as due to oncogenic mutation activating other signaling pathways (MAPK, AKT, and mTOR) | HIF-1α induces the expression of pyruvate dehydrogenase kinase 1 (PDK1). PDK1 phosphorylates and inactivates mitochondrial pyruvate dehydrogenase. Enhances the dependence of cells on glycolysis for ATP production instead of OXPHOS. | Metabolism, cell survival, erythropoiesis, angiogenesis | [132,133,134] |
2.1.2. Reason 2: Glycolysis Supports the Proliferative Needs of Cancer Cells
In cancer, tumor cells employ strategies that promote their survival, growth, and invasion. Therefore, it has been theorized that cancer cells use aerobic glycolysis as a trade-off because it supports the biosynthetic anabolic needs pertaining to constant, uncontrolled proliferation [67].
The Warburg effect supplies nucleic acids, proteins, and lipids through certain branching pathways that emanate from glycolysis. For instance, the pentose phosphate pathway (PPP) produces the reducing agent NADPH, which is used in de novo lipid synthesis [135,136,137]. Besides, the diversion of the glycolysis flux towards de novo serine biosynthesis is utilized by the enzyme phosphoglycerate dehydrogenase (PHGDH) [67,138]. Additionally, lactate is produced during the final step of anaerobic glycolysis, along with NAD+. The produced NAD+ acts as a positive feedback mechanism to keep glycolysis active and ensure that the supply chain of building blocks is operating [139]. The produced intracellular lactate is shuttled to the extracellular stroma, giving it its acidic attributes in cancer [140]. An outstanding study by Heiden et al. (2020) revealed that an increased cellular demand for NAD+, which is comparably more than the demand for ATP and the rate of ATP turnover, is what drives the preferential dependence on aerobic glycolysis, rather than OXPHOS, in proliferating cells, such as cancer cells [141]. The NAD+/NADH ratio is critical for several metabolic processes, including nucleotide synthesis, lipid metabolism, amino acid metabolism, and central carbon metabolism [142]. Redox reactions and biosynthetic processes both require NAD+ generation [141]. NAD+ regeneration by the ETC cycle is constrained due to increased mitochondrial membrane potential and decreased ATP synthase activity during OXPHOS in proliferating cancer cells [141]. Owing to the increased NAD+ demand from cancer cells, the cell diverts its metabolic phenotype towards aerobic glycolysis [141].
Undifferentiated stem cells resemble cancer cells in having high proliferative activity, and therefore, they similarly shift their metabolism towards anaerobic glycolysis instead of OXPHOS [143,144]. During stem cell differentiation, cellular metabolism switches back to mitochondrial OXPHOS to generate energy, while the rate of anaerobic glycolysis declines [145]. The dysregulation of the intracellular and extracellular pH of cancer cells that accompanies aerobic glycolysis is another means by which the Warburg effect promotes tumor growth [140]. Dysregulated pH dynamics characterized by extracellular acidic stromal cells and intracellular alkaline cytoplasm are hallmarks of cancer cells and can influence tumor proliferation, metastasis, and metabolic shift [140]. Proliferating cancer cells require an alkaline intracellular pH compared to normal quiescent cells that have an acidic intracellular pH [146,147,148]. The increase in intracellular pH in a cancer cell promotes the glycolytic metabolic shift and confers a proliferative advantage for tumor cells [149,150,151]. For instance, a cytoplasmic alkaline pH is required for growth factors to initiate nucleic acid synthesis [147]. Moreover, an alkaline intracellular pH promotes intracellular protein synthesis and drives other phenotypes of cancer cells [140,152,153].
2.1.3. Reason 3: Activation of HIF-1α by ROS
Another reason for the glycolytic shift in cancer is the accumulation of ROS, which causes the activation of HIF-1α. Usually, in hypoxic conditions, HIF-1α gets activated when the cell senses a low oxygen supply. ROS mimics the hypoxic effect and activates HIF-1α, which then promotes glycolysis by upregulating the expression of several glycolytic enzymes, including hexokinase 2 [154,155], phosphofructokinase [156], phosphoglucomutase 1 [157], enolase [158], pyruvate kinase, pyruvate dehydrogenase (PDH), pyruvate dehydrogenase kinase (PDK) [159], lactate dehydrogenase A (LDHA) [158], monocarboxylate transporter 4 [160,161,162], and glucose transporters, GLUT1 and GLUT3 [163]. Additionally, HIF-1α reduces the OXPHOS capacity by inhibiting mitochondrial biogenesis [164,165], decreasing PDH activity [159], and reducing ETC activity [166].
2.1.4. Reason 4: Dysregulation of the Glycolytic Machinery
Studies have indicated that dysregulation occurs at the level of glycolytic protein expression (Table 3). Subsequent research has elucidated the role of the pyruvate dehydrogenase complex (PDC) in the metabolic switch in tumor cells towards aerobic glycolysis. In 2007, Koukourakis et al. observed a significant decrease or absence of PDC expression and/or an overexpression of PDK in 91% of lung cancer patients tested via immunohistochemistry [7]. Typically, active PDC facilitates the oxidative decarboxylation of pyruvate into acetyl-CoA within the mitochondria. Conversely, PDK phosphorylates and deactivates PDC [15]. When PDC is inactive, pyruvate accumulates in the mitochondria and translocates back to the cytosol, where it is converted to lactate and NADH [15].
In this milieu, NADH was found to play a role in the glycolytic shift by directly or indirectly inhibiting PDC and activating PDK. Recent studies propose that a high concentration of cytosolic NADH, coupled with increased pyruvate, decreased lactate, and an active LDHA enzyme, positively promotes glycolysis in cancer [69,167,168,169].
Certain investigations have also demonstrated that overexpression of the antioxidant UCP2 in cancer cell lines promotes aerobic glycolysis, tumor proliferation, and resistance to apoptosis-induced chemotherapy [88,89,90] (see Figure 2). In 2016, Brandi et al. illustrated that UCP2 upregulates the expression of heterogeneous nuclear ribonucleoprotein A2/B1, which, in turn, regulates the transcription of GLUT1, pyruvate kinase M2 (PKM2), and LDH genes. UCP2 facilitates the metabolic shift in cancer cells towards Warburg's aerobic glycolysis[90].
Table 3.
Common alterations reported in glycolysis-related genes in cancer.
| Gene ID | Gene Name | Mutation/deregulation | Function in glycolysis | Publication |
|---|---|---|---|---|
| HK | Hexokinase | Upregulated by p53 in cancer and promotes tumor growth and survival | Phosphorylates glucose when it enters the cells | [76,170,171,172] |
| PFK1 | 6-PhosphoFruktoKinsae-1 | Amplification and/or upregulation, posttranslational modification reported in multiple cancer types. | PFK1 catalyzes the phosphorylation of fructose-6-phosphate (F6P) to fructose-1, 6-bisphosphate (Fru-1,6-P2) using Mg-ATP as a phosphoryl donor. | [76,173,174,175] |
| PK | Pyruvate Kinase | Posttranslational modification or enhanced expression that benefits cancer | PK is involved in the final step of glycolysis, and it mediates the transfer of a phosphate group from phosphoenolpyruvate (PEP) to ADP resulting in pyruvate and ATP. | [76,176,177,178,179] |
| PDK-1 | Pyruvate Dehydrogenase Kinase -1 | Upregulation | PDK is a kinase enzyme that inactivates Pyruvate Dehydrogenase by phosphorylation dephosphorylation at different specific serine residues. PDK decreases the oxidation of pyruvate in mitochondria and increases the conversion of pyruvate to lactate in the cytosol. |
[76,180,181,182] |
2.1.5. Reason 5: AMPK Inhibition in Cancer Leads to a Glycolytic Shift
AMPK is a highly conserved serine/threonine protein complex that acts as a metabolic sensor and a master regulator of cellular metabolic homeostasis [183]. AMPK can be activated by either one of the two cell signals; the first is intracellular Ca2+-dependent, while the second is AMP-dependent (see Figure 4). AMPK is modulated either by phosphorylation or allosteric activation. In response to an increase in the AMP/ATP ratio, liver kinase B1 (LKB1) gets activated, which in turn directly activates AMPK by phosphorylation at Thr172, located in the catalytic subunit of the AMPK protein [184,185,186,187]. The AMP/ATP ratio can be altered during various intracellular states, such as hypoxia, glucose deprivation, calcium concentration, cytokines, and adipokines, and by certain hormones [187]. On the one hand, active AMPK inhibits the biosynthetic pathways in the cell, such as hepatic fatty acid synthesis and protein synthesis. On the other hand, active AMPK activates ATP-generating catabolic pathways, such as fatty acid uptake and oxidation, glycolysis, and mitochondrial biogenesis (see Figure 5). In cancer, AMPK is generally considered as a tumor suppressor [188]. Studies found that the dysregulation of AMPK plays a role in the glycolytic metabolic switch in cancer. Low AMPK expression is further implicated in tumorigenesis by promoting tumor initiation and progression [189]. Inactivation or reduced expression of AMPK in cancer promotes tumor growth and invasiveness [190,191]. This is an expected scenario, considering the activities exerted by AMPK.
A closer look at how AMPK shifts the metabolic phenotype in cancer reveals that AMPK modulates mitochondrial respiration through activating autophagy (including mitophagy) by phosphorylating and activating ULK1; this regulates the localization of an important component of the phagophore called autophagy-related protein 9 (ATG9) [192,193] (see Figure 5). mTOR, on the one hand, can inhibit ULK1/2, thus blocking autophagy [194]. AMPK can also induce mitochondrial biogenesis, which aims to increase the capacity of OXPHOS. Moreover, Faubert et al. (2013) found that AMPK negatively regulates the Warburg effect and suppresses tumor growth in vivo [195]. Faubert et al. documented that knocking down the α catalytic subunit of AMPK accelerates Myc-induced tumorigenesis. Moreover, the inactivation of AMPK caused stabilization of HIF-1α and a glycolytic shift in tumor cells in vitro. Altogether, the inhibition of AMPK in cancer inhibits OXPHOS and activates the Warburg effect in tumor cells (see Figure 5).
More interestingly, in cancer, AMPK acts through the AMPK/tuberous sclerosis complex (TSC)/mTOR signaling axis to regulate the metabolic switch. Inoki and colleagues found that active AMPK phosphorylates and activates TSC2 [196]. Earlier, Inoki had established that both TSC1/2 inhibits the activity of mTOR by inhibiting the phosphorylation of ribosomal protein S6 kinase B1 (S6K) and eukaryotic translation initiation factor 4E-binding protein 1 (4E-BP1), which are downstream targets of mTOR [196,197]. Inoki et al. further reported that TSC1/2 inhibits the phosphorylation of S6K and 4E-BP1 by targeting Ras homolog (Rheb)− the protein that activates the protein kinase activity of mTOR [198]. Inoki et al. showed that TSC2 acts as a GTPase-activating protein and blocks the activity of Rheb and regulates its levels in vitro [198]. In cancer, mTOR is frequently high and has been found to activate aerobic glycolysis via the induction of pyruvate kinase isoenzyme 2 (PKM2) and other glycolytic enzymes [199]. In a recent study by Ling et al. that showed unprecedented results, mTOR was reported to directly inhibit AMPK via the phosphorylation of AMPK α1, S347 and α2, S345 in mammalians, accompanied with reduced phosphorylation of the activation loop T172. A loss of mTOR activity caused AMPK activation independent of the AMP/ATP ratio [200].
Collectively, active AMPK could activate autophagy either directly through the activation of ULK2 or indirectly by activating TSC 2, which further inhibits mTOR.
3. The Metabolic Shift in T2D
The metabolic switch in the case of insulin resistance and T2D occurs at two stages. The first stage occurs during the pathogenesis of insulin resistance and T2D, while the second stage occurs when both insulin resistance and T2D have already manifested clinically. The first stage is characterized by hyperactive OXPHOS and is due to high glucose uptake. In the second stage, distinct metabolic patterns arise in a tissue-specific manner. Conflicting reports on OXPHOS have been described in human and in vitro studies, showing either normal functioning mitochondria and active OXPHOS or dysfunctional mitochondria. However, evidence suggests that if mitochondrial dysfunction does occur, it happens as a result of, rather than being the cause of, insulin resistance and T2D.
3.1. The Metabolic Shift During the Pathogenesis of Insulin Resistance and T2D
Under normal conditions of insulin sensitivity, glucose-stimulated insulin secretion regulates glucose uptake and increases the activity of OXPHOS mitochondrial respiration [43,201]. During the pathogenesis of insulin resistance and towards the emergence of T2D, there is a state of chronic hyperglycemia caused by excessive nutrient supply and/or physical inactivity. Chronic hyperglycemia stimulates the continuous release of insulin from pancreatic β cells [202,203]. Insulin secretion increases glycolysis and pyruvate production [201], and elevates chronic hyperactivity of mitochondrial OXPHOS in response to insulin signaling. This heightened state of OXPHOS results in an augmented state of oxidative stress. In addition to the release of ROS, prolonged hyperglycemia causes glucose toxicity [35,36,37,38,204,205]. Both glucose toxicity and ROS ultimately damage pancreatic β cells and impair their ability to sufficiently secrete insulin [37]. Although ROS production partly results from enhanced mitochondrial respiration under glucose stimulation, the expression of antioxidant genes is unusually low in β cells, leading to ROS accumulation in the cytoplasm, owing to inefficient ROS elimination [206]. Therefore, this is followed by a decrease in insulin secretion and is accompanied by a decrease in the rate of glycolysis [207]. In addition to the impairment of insulin secretion, ROS are major players in developing insulin resistance by rendering cells insensitive to insulin, thus hindering the insulin signaling pathway in peripheral tissues, such as SKM, adipose tissue, and the liver [41,208]. This is because excessive ROS generation activates protein kinase (PK) signaling pathways [209]. Insulin signaling is thus suppressed downstream of the insulin receptor (IR), at the level of IR substrate-1 (IRS-1) and PI3K, which together promote insulin resistance in peripheral tissues [210,211].
3.2. The Metabolic Shift in Established Insulin Resistance and T2D
As mentioned earlier, accumulating evidence reveals that mitochondrial dysfunction is a result, rather than the cause, of insulin resistance in T2D [212,213]. The previous hypothesis, which suggested that insulin resistance is the result of mitochondrial dysfunction, is mostly based on association studies and not on cause-and-effect studies. Several association studies utilized animal models that documented mitochondrial dysfunction in parallel with the presence of insulin resistance. The previously reported presence of mitochondrial dysfunction was tissue-specific, which was mostly found in the SKM of animal models or in patients with T2D or insulin resistance, while active or normal OXPHOS was reported in the liver of patients with T2D or insulin resistance.
Here, we bring forth examples of such association studies. For instance, OXPHOS genes were found to be downregulated in the SKM of patients with T2D [42,214,215] and after high-fat diet consumption [216]. Additionally, both Kelly et al. (2002) [48] and Mogensen et al. (2007) showed impaired mitochondrial respiration in the SKM of patients with T2D, compared to their obese nondiabetic counterparts [217]. Several research groups reported contradictory findings in T2D, whereby the presence of normally functioning mitochondria in SKM was demonstrated [49,50,51].
In contrast to the results pertaining the SKM, a study conducted by Takamura et al. (2006) found that genes encoding OXPHOS are upregulated during fasting hyperglycemia in the livers of patients with T2D [52]. In another study conducted on mouse models, hepatic mitochondria showed flexibility in adapting to a high-fat diet and prevented hepatic steatosis through increased OXPHOS activity and ETC uncoupling [218]. Thus, collective data support the rationale that mitochondrial respiration is regulated by different tissue-specific mechanisms, which partly explains why the mitochondrial respiratory reaction to excessive nutrient supply, obesity, physical inactivity, and insulin resistance is not uniform among different organs and tissues and in different reported research contexts. Moreover, other data have demonstrated that mitochondrial inhibition using drugs enhances insulin sensitivity and benefits patients with T2D.
3.3. Mitochondrial Dysfunction is a Result of Insulin Resistance
Recent studies have exploited the cause-and-effect relationship between insulin resistance and mitochondrial dysfunction using pharmacological and transgenic animal model approaches. The results corroborate that mitochondrial dysfunction is a result of insulin resistance and not the opposite. The resultant mitochondrial dysfunction was found to be a protective mechanism to counteract insulin resistance. This was shown in a study by Pospisilik et al. in 2007, who induced OXPHOS deficiency via the genetic ablation of the AIF gene in mouse models. OXPHOS deficiency reduced fat mass, increased insulin sensitivity, and enhanced glucose tolerance. These results established that the inhibition of OXPHOS did not cause insulin resistance in mice, and contrarily, was protective against the development of insulin resistance in obese mice [219].
Nair et al. showed that mitochondrial dysfunction and altered expression of mitochondrial genes are not intrinsic defects in patients with T2D but are rather secondary to abnormal secretion levels of glucose and insulin. In this study, diabetic and nondiabetic individuals exhibited similar mitochondrial content. After low-dose insulin infusion, there was similar ATP production observed in both groups. High-dose insulin, however, showed that ATP production in diabetic patients was lower than that in nondiabetic individuals, along with a reduced expression of peroxisome proliferator-activated receptor G coactivator-1 (PGC-1), citrate synthase, and cytochrome c oxidase [215].
In another study, Roden et al. found that accumulated intramyocellular fatty acyl-CoA causes the downregulation of OXPHOS genes by decreasing the expression of PGC-1. However, they suggested that the vicious cycle of metabolic changes in T2D starts with the increased availability of free fatty acids (FFA), lipid accumulation in myocytes, and impaired lipid oxidation, which may cause mitochondrial dysfunction in the future [220].
An interesting study by Fazakerley et al. found that induced mitochondrial oxidative stress impaired glucose uptake, which was induced by insulin, and decreased the translocation of GLUT4 protein to the cell membrane in adipocytes and myotubes of C57BL/6J mice. However, the induced mitochondrial oxidative stress did not alter the activity of OXPHOS [221]. Interestingly, a review by Lewis et al. (2019) on various experimental designs, which attempted to measure or assess mitochondrial OXPHOS in SKM, has refuted the misconception that mitochondrial OXPHOS is dysfunctional and downregulated in T2D, and proved instead that it is due to the limited oxygen supply to these tissues. They found that there are limitations in the reviewed in vivo and in vitro studies on human mitochondrial SKM function [222]. The authors suggest that the mitochondrial respiratory capacity is intact in T2D when using high-resolution respirometry on isolated mitochondria and that other mitochondrial respiratory inadequacies detected in some in vivo studies are more likely due to changes in mitochondrial fractional volume [222]. These changes could be due to a less active lifestyle or limited oxygen availability in the cytosolic environment [81].
3.4. The Role of ROS in Insulin Resistance and T2D
From a mechanistic standpoint, an elevation in ROS levels can instigate the activation of stress-sensitive serine/threonine kinase signaling pathways, including c-Jun N-terminal kinase (JNK) [223], nuclear factor kappa B (NF-κB) [224,225], p38MAPK [226], and others. These pathways subsequently phosphorylate multiple targets, IRS proteins among them. The heightened serine phosphorylation of IRS diminishes its capacity for tyrosine phosphorylation, potentially hastening the degradation of IRS-1. This provides a plausible explanation for the molecular underpinnings of oxidative stress-induced insulin resistance. Compelling data affirm the crucial role of JNK activation, NF-κB kinase, protein kinase C inhibition, and potentially other stress- and inflammation-activated kinases in the development of oxidative stress-induced insulin resistance. These findings suggest that they could serve as appealing pharmacological targets to enhance insulin sensitivity [211,227].
Al-Mulla and Bitar et al. made a series of interesting discoveries, wherein they explored the role of oxidative stress in insulin resistance and T2D, along with their mechanism of action both in vitro and in vivo. In 2015, they found that oxidative stress and PKA activation were associated with diabetes in Goto-Kakizaki diabetic rat models. They showed that oxidative stress and PKA induced insulin resistance through enhancing cAMP responsive element modulator/Inducible cAMP Early Repressor (CREM/ICER) expression, which reduced IRS-2 expression by inhibiting the transcriptional activity of cAMP response element (CRE) [228]. In another study, they corroborated that T2D instigates a cascade of events that produce ROS (mainly O2) from NADPH oxidase, leading to the oxidation of BH4 and uncoupling of NOS, which ultimately leads to NO inactivation with subsequent peroxynitrite formation. Altogether, an imbalance in the redox state is caused by increased ROS bioavailability and reduced antioxidant capability, which translates into a heightened state of oxidative stress [229,230]. Moreover, the authors demonstrated that the high oxidative stress in T2D is partly attributed to the diminished intracellular stabilization of NRF2 in dermal fibroblasts that were isolated and cultured from Goto-Kakizaki rats. Low NRF2 stabilization caused a decrease in the antioxidant effect of NRF2 in response to glucose-induced oxidative stress in dermal fibroblasts, compared to cells in normoglycemic conditions [230]. Therefore, reduced NRF2 is also associated with higher cellular sensitivity to oxygen free radicals and results in cellular necrosis [230].
In 2012, Bitar and Al-Mulla found that ROS is responsible for the development of insulin-like growth factor 1 (IGF-1) resistance and, consequently, delayed wound healing in a T2D rat model [208]. IGF-1 resistance is another mechanism involved in developing insulin resistance in peripheral tissues. IGF-1 signaling, via the IGF-1 receptor (IGF-1R), uses downstream mediators that are commonly involved in the insulin signaling pathway. Altered IGF-1 function has been implicated in the pathogenesis of insulin resistance and in several other diseases, such as autoimmune diseases, atherothrombosis, osteoporosis, and certain common types of cancer [208]. In their study, IGF-1 activation in the PI3K-AKT-GSK-3ß pathway was attenuated in fibroblasts in vitro that had phenotypic features of diabetes or hypercortisolemia. In contrast, ROS-activated JNK pathway led to the inhibitory phosphorylation of IRS1 at Ser307. Bitar and Al-Mulla showed that ROS, via the activation of JNK-p-IRS1 (Ser307), mediates IGF-1 resistance in T2D [208]. In 2019, Akhter et al. shed light on another mechanism by which oxidative stress is involved in inducing metabolic inflammation in T2D, i.e., through the upregulation of toll-like receptors (2 and 4) , interferon regulatory factors (3 and 5), and other key pro-inflammatory cytokines in peripheral blood mononuclear cells. This mechanism depends on MAPK/NF-κß signaling [231].
3.6. AMPK Inhibition Implicated in Insulin Resistance and T2D
AMPK is regarded as the guardian of mitochondria, a complex master regulator, and a key metabolic sensor. Paradoxically, AMPK is the link that connects the metabolic disturbances in cancer and diabetes, like the concept of Yin and Yang. Although AMPK is inhibited in both diseases, it exerts multi-faceted functions through different signaling pathways. Ultimately, low AMPK activity promotes tumor growth and proliferation and causes insulin resistance in the peripheral tissues of patients with T2D.
As previously discussed, AMPK controls mitochondrial biogenesis, dynamics, and disposal by mitophagy. Therefore, in low cellular ATP states, active AMPK restores ATP homeostasis by increasing mitochondrial ATP production, while low AMPK inhibits autophagy [232]. It was found that T2D is associated with suppressed autophagy and lipid accumulation [233].
AMPK plays an important role in the metabolic shifts associated with insulin resistance and T2D [183]. In animal studies, low AMPK activity contributed to the development of insulin resistance [234,235,236]. Inhibition of AMPK reduced glucose uptake and utilization due to a decline in the phosphorylation of target proteins involved in the trafficking of glucose transporters, GLUT1 and GLUT4. These target proteins are thioredoxin interacting protein, TBC1 domain family member 1 protein, and phospholipase D1. Low AMPK also inhibits FFA β-oxidation in the mitochondria and exacerbates lipid biosynthesis, leading to the accumulation of lipids in cells and tissues. Normally, an active AMPK would enhance the breakdown of lipids by stimulating lipases. The activity of carnitine palmitoyltransferase I (CPT1), which transports FFA into mitochondria, is indirectly stimulated by AMPK. AMPK phosphorylates acetyl-CoA carboxylases 1 and 2, which in turn blocks the production of malonyl-CoA [236]. Malonyl-CoA is a potent inhibitor of the lipid transporter CPT1 [183,236]. Additionally, it was found that active AMPK inhibits hepatic gluconeogenesis by enhancing the expression of the orphan nuclear receptor small heterodimer partner (SHP) gene, which inhibits the transcriptional activity of cAMP responsive element binding protein 1 (CREB). CREB regulates the transcription of hepatic gluconeogenesis genes [237].
Taken together, the inhibition of AMPK causes a metabolic shift in T2D through several mechanisms that decrease glucose utilization, inhibit FFA β-oxidation, cause accumulation of lipids in tissues, and activate hepatic gluconeogenesis. Generally, these changes are known to be implicated in developing insulin resistance (see Figure 6).
4. Metabolic Therapeutic Approaches in Cancer and T2D
Like Yin and Yang, cancer and T2D are metabolic disorders characterized by opposing metabolic switches and divergent underlying signaling pathways, yet they intertwine towards the master regulator AMPK. In cancer, the Warburg glycolytic shift promotes malignant transformation, tumor progression, invasiveness, and resistance to chemotherapy and/or radiotherapy [119,238,239]. Nonetheless, during the pathogenesis of T2D, there is hyperactivity and dominance of mitochondrial OXPHOS. Therapeutic and/or nutritional targeting of either of the two metabolic shifts is a promising approach to correct the metabolic imbalance and restore homeostasis [20,239,240,241].
Although tumors are predominately glycolytic, they vary in their phenotypic features associated with proliferation, invasion, metastasis, and resistance to therapy. The characteristics of the metabolic phenotype for each cancer determine its rate of proliferation and resistance to chemotherapy [151]. Thus, cancer therapy needs to be customized to target the underlying causative metabolic dysfunction. Therapeutic attempts to target cellular metabolism in cancer aim at the inhibition of Warburg glycolysis and/or the activation of OXPHOS to confer antiproliferative activity. In addition, metabolic inhibition has shown the ability to sensitize chemoresistant tumor cells to treatment. Furthermore, based on previous research, there have been suggestions to reestablish the metabolic imbalance in cancer by targeting the tumor microenvironment symbiotic crosstalk.
Certain pharmacological agents and nutrients have been able to correct and reverse metabolic imbalances in cancer and are gaining validation through in vitro and in vivo analysis, and in clinical trials.
Whereas in T2D, targeted metabolic inhibition using nutritional and/or pharmacological compounds could prevent insulin resistance and improve insulin sensitivity in prediabetic and diabetic animal models, and in diabetic patients. These therapies aim to inhibit mitochondrial OXPHOS activity [41,219]. The use of certain nutrients and dietary supplements as metabolic treatments or adjuvants in T2D is gaining attention owing to the encouraging results obtained in the past decade. In the next subsections, we summarize the pharmacological based approaches to target mitochondrial metabolism in cancer and T2D.
4.1. Pharmacological-Based Approaches Targeting Mitochondrial Metabolism in Cancer
4.1.1. BACH1 Depletion Activates OXPHOS and Sensitizes Tumor Cells to Metformin
Among the genes related to ROS homeostasis, BTB domain and CNC homolog 1 (BACH1) is a heme-binding transcription factor that represses the oxidative stress response and is a negative regulator of ROS-induced cellular senescence directed by p53 [242,243]. BACH1 is upregulated in breast and other types of cancer; it is proposed to be a marker of poor prognosis and high metastatic rate in breast cancer. For instance, triple-negative breast cancer (TNBC) cells reprogram their metabolism by increasing BACH1 expression to direct their metabolism away from the TCA cycle, which could be a protective mechanism that enhances their proliferative potential. On the one hand, it prevents the accumulation of ROS by shutting down mitochondrial metabolism. Thus, BACH1 may provide a mechanism by which tumor cells evade oxidative stress-induced senescence.
In 2019, Rosner et al. [244] showed that the combined therapeutic use of metformin with BACH1 inhibitor (hemin) could reverse chemoresistance in TNBC cells. BACH1 targets mitochondrial metabolism by repressing key ETC genes (UQCRC1 and ATP5D, both negatively correlated with BACH1 in TNBC), which are predominantly involved in the OXPHOS pathway. Metformin is known to mainly inhibit mitochondrial ETC Complex-I, along with other metabolic targets. Metformin was able to inhibit the growth of tumor cells and decrease tumor cell viability in BACH1-depleted TNBC cells. However, control cells that expressed BACH1 did not respond to metformin treatment, and the TNBC cells continued to grow and proliferate. Downregulating BACH1 in tumors using hemin, both in vitro and in vivo, resulted in an increased expression of mitochondrial inner membrane genes involved in ETC, and promoted mitochondrial respiration. TNBC cells that were depleted of BACH1 exhibited higher oxygen consumption, lower lactate production, higher glucose utilization in the TCA cycle, increased ATP generation, higher TCA cycle intermediates production, and decreased glycolysis-related intermediates [244]. Rosner attempted to reprogram the metabolic pathway in TNBC tumors resistant to ETC inhibition therapy because of high BACH1 expression. Inhibiting BACH1 expression sensitized tumor cells to metformin, both in vitro and in vivo. For further details regarding interventional clinical trials investigating the impact of metformin on various types of cancer, we have compiled a table retrieved from clinicaltrials.gov on November 23, 2023. This table encompasses both completed and ongoing studies that have reached Phase 2 or Phase 3. It is important to note that trials that were withdrawn, suspended, or terminated have been excluded (Refer to Supplementary Table S1).
Cellular senescence is mainly mediated by the tumor suppressor p53, which serves as a barrier to malignant transformation [245]. The upregulation of BACH1 in TNBC cells has been suggested to prevent oxidative stress-induced senescence. This rationale is supported by the findings of Dohi et al. who demonstrated that BACH1 forms a complex with p53, histone deacetylase 1, and nuclear co-repressor. The formation of this complex prevents p53 from inducing an effective oxidative stress response by promoting histone deacetylation [243]. Furthermore, Wiel et al. showed that stabilizing BACH1 using antioxidants in a p53-/- background in lung cancer models increased metastasis, glucose uptake, glycolysis rate, and lactate secretion in mouse and human lung cancer cells. Hence, in scenarios marked by lower oxidative stress, BACH1 promotes glycolysis-dependent lung cancer metastasis independently of p53 [246]. Multiple microRNAs (miRs) were found to target the post-transcriptional regulation of BACH1 and reduce cancer progression; such as miR-142-3p, which can target BACH1 in breast cancer cells, leading to reduced cellular proliferation, invasion, and migration [247]. The induction of miR-330 also inhibits the proliferation of colorectal cancer cells by suppressing BACH1 gene expression [248].
In addition to these studies, BACH1 was also found to be linked to an age-dependent decline in adaptive homeostasis. Its levels were elevated in various tissues, including the heart, liver, and lungs, in aging mice [245]. Furthermore, BACH1 expression was higher in human bronchial epithelial cells obtained from older adults compared to those from young adult donors [249]. Thus, BACH1 attenuates adaptive redox homeostasis in both aging mice and older individuals. Taken together, these studies show that BACH1 is a potential metabolism-targeting therapy for cancer. This suggests that the inhibition of BACH1 can modulate the metabolic profile in resistant cancers such that the OXPHOS pathway is restored, glycolysis is reduced or omitted, cancer growth is halted, and sensitizes cancer cells to therapy.
4.1.2. Dichloroacetate and EGFR-Inhibitors Reverse the Warburg Effect in Cancer
Sun et al. demonstrated that the generic drug dichloroacetate (DCA) can reverse the glycolytic phenotype in metastatic breast cancer cells, both in vitro and in vivo, and can inhibit tumor growth and metastasis [250]. DCA works by inhibiting PDK activity, wherein PDK inactivates PDH via phosphorylation. PDH controls the conversion of pyruvate to acetyl Co-A, which in turn enters the TCA cycle and generates ATP via the action of OXPHOS. Thus, treatment with DCA stops the inhibition of PDH, increases the flux of pyruvate into the mitochondria, and promotes mitochondrial OXPHOS over glycolysis [250].
In 2015, De Rosa et al. demonstrated that the use of EGFR-inhibitors, including erlotinib or WZ4002 in human non–small cell lung cancer cell lines (H1975, HCC827, and H1993) and PHA-665,752 in the H1993 cell line, succeeded in the reversal of the Warburg effect and reactivation of OXPHOS in these cell lines [251]. This effect was mediated through the upregulation of ETC mitochondrial complexes, in addition to reduced expression levels of key glycolysis enzymes, such as hexokinase II and p-PKM2 Tyr105. Concomitantly, decreased lactate secretion and increased intracellular ATP levels were observed in response to EGFR inhibition [251]. In conclusion, these results revealed that the effective inhibition of EGFR signaling can reverse the Warburg effect in cancer cell lines and restore OXPHOS.
4.1.3. Metformin Activates AMPK to Induce Apoptosis in Cancer
Targeting AMPK in cancer cells to either sensitize tumor cells to chemotherapy, cause cell cycle arrest, or induce apoptosis are promising therapeutic approaches. For instance, the activation of AMPK inhibited cervical cancer cell proliferation through AKT/FOXO3a/FOXM1 signaling cascade by counteracting the function of Forkhead box M1 (FOXM1) [190]. Previously, several pharmacological AMPK activators, such as metformin, the AMP-mimetic 5-aminoimidazole-4-carboxamide (AICAR), and the ATPase inhibitor A23187 were able to suppress cervical cancer cell growth by activating AMPK [190].
In 2008, Keith et al. were able to induce cell cycle arrest in the MDA-MB-231 breast cancer cell line by treating the cells with metformin only in the presence of cyclin-dependent kinase inhibitors (p27kip and/or p21cip1). Metformin was able to activate the AMPK pathway and downregulate cyclin D1 [252]. Mills et al. further demonstrated that the LKB1-AMPK pathway regulates p27kip1 phosphorylation; they were able to induce apoptosis in cell lines after AMPK activation in the absence of p27kip1. Downstream of AMPK, p27Kip1 is phosphorylated at Thr198, which stabilizes p27 leading to autophagy and cell-cycle progression. When p27 was knocked down in the cancer cell line, LKB1-AMPK activation induced apoptosis [253].
4.1.4. Targeting PI3K/AKT pathway in cancer
The dysregulation of the PI3K/AKT pathway is a common feature in many cancers [254]. Evidence indicates that inhibiting PI3K/AKT pathway hinders tumor progression [254]. However, the use of PI3K-AKT-mTOR inhibitors in treating various cancer types has been observed to induce hyperglycemia in patients [255]. A study by Khan et al., investigated the clinical data of 341 cancer patients from 12 phase I clinical trials treated with PI3K , AKT or mTOR inhibitors as well as dual inhibitors. There was evident hyperglycemia in 87.4% of these patients. However, a grade three hyperglycemia was only seen in 6.7% of these patients. Hence, hyperglycemia was mostly manageable in those patients. Thus caution is necessary when treating cancer patients who are also diabetics with PI3K-AKT-MTOR inhibitors [255]. This study may seem paradoxical, as the inhibition of PI3K-AKT-mTOR pathway, which generally leads to inhibition of cell proliferation, is expected to activate the AMPK pathway. AMPK activation would exhibit beneficial effect in diabetes and lower glucose levels. However, this is not the case with PI3K-AKT-mTOR inhibitors alone. Nevertheless, the combination of Metformin and PI3K-AKT-mTOR inhibitors in vitro enhances apoptosis of ovarian cancer cells [256] and induces drug sensitivity in pancreatic cancer cells [257].
PI3K produces Phosphatidylinositol (3,4,5)-trisphosphate (PIP3), which, in turn, activates phospholipase D (PLD) [258]. PLD catalyzes the hydrolysis of the membrane phospholipid, phosphatidylcholine, to generate choline and metabolically active phosphatidic acid (PA) [259]. PA is a signaling lipid involved in processes such as cell proliferation and vesicular trafficking. PLD can influence mTOR activity by generating PA [260], which directly activates mTOR complex 1 (mTORC1) under certain conditions [261]. PA stimulates mTORC1 function and suppresses the activation of mTORC2 as part of a mTORC1/2 feedback loop [262]. PI3K inhibitors decrease PLD activation after insulin receptor stimulation [263], and the mutation of the PIP3 binding site on PLD prevents PLD activation and membrane recruitment [264]. A study by Toschi et al. (2009) demonstrates that by inhibiting PLD activity, mTORC2 could be targeted therapeutically with rapamycin [265]. Thus, the combination of rapamycin, metformin, and PI3K/PLD inhibitors can have a favorable therapeutic outcome in cancer therapy.
PIP3 generation mediates downstream signaling events that inhibit glycogen synthase kinase-3β (GSK-3β) [266,267]. GSK-3β, in turn, hinders NRF2 by directing it towards ubiquitination and subsequent degradation [268]. NRF2 plays a pivotal role in combating oxidative stress and regulating redox homeostasis, thereby safeguarding cells against carcinogenesis (reviewed in [269]). However, studies over the last decade reveal a "dark side" of NRF2 [270], where its constitutive stabilization leads to increased glutaminolysis [271], cancer progression [272], metastasis [273], and chemoresistance [274,275]. Indeed, NRF2 redirects glucose and glutamine into anabolic pathways during metabolic reprogramming [271,276]. Consequently, strategies such as inhibiting PI3K and NRF2 or activating GSK-3β, along with NRF2 repressor Kelch-like ECH-associated protein 1 (KEAP1) [277], hold promising therapeutic potential against cancer.
4.2. Pharmacological-Based Approaches Targeting Mitochondrial Metabolism in T2D
4.2.1. Apoptosis-Inducing Factor Ablation in Diabetic Mice Inhibited OXPHOS
A study by Penninger‘s team in 2007 showed that global or tissue-specific gene ablation (liver and muscle) of apoptosis-inducing factor (AIF) in mice caused a deficiency in OXPHOS, which was accompanied by improved glucose tolerance, increased insulin sensitivity, and reduced fat mass [219]. AIF has been known to cause progressive OXPHOS dysfunction in mice [278,279]. Mutation analysis performed in several model organisms found that AIF was an essential regulatory gene for maintaining fully active and functional mitochondrial ETC [280,281]. Therefore, AIF deletion caused a progressive loss of ETC activity and function [278,280]. In the study by Penninger’s team, impaired OXPHOS prevented weight gain, insulin resistance, and T2D, which is contrary to other studies reporting that OXPHOS deficiency is associated with insulin resistance and T2D [219].
4.2.2. Metformin as a Metabolic Inhibitor in T2D
For decades, metformin has shown great success in the treatment of T2D. Metformin can stimulate glucose uptake and glycolysis in patients with T2D [201]. Glycolysis plays two major roles in glucose homeostasis. The first role is through inhibiting hepatic gluconeogenesis, thereby decreasing the amount of glucose released into the blood [201,282] and the second role is through enhancing insulin secretion by pancreatic β cells [201,207,283]. In this context, metformin works by augmenting glycolysis, which leads to a decrease in liver gluconeogenesis. Metformin exerts its effects by suppressing mitochondrial OXPHOS via inhibiting Complex I (NADH dehydrogenase) of the ETC [284,285]. Inhibiting Complex I increases the AMP/ATP ratio, which further activates AMPK [183]. The idea behind metabolic inhibition is that any injury caused to the mitochondrial metabolic machinery leads to the activation of AMPK to compensate for the mitochondrial dysfunction. Metformin was also found to exert its function by upregulating UCP2 in adipocytes in mouse models, thus playing a protective role against oxidative damage [286]. Therefore, AMPK activation is an effective therapeutic strategy for enhancing insulin sensitivity in T2D [41].
4.2.3. Targeting PI3K/AKT pathway in T2D
Su, et al. (2018) in a comprehensive review explain effects of PI3K-AKT signaling on obesity and T2D. The review summarizes the findings of many studies done in-vitro and in-vivo on diabetic cells and mouse models in which the activity of the PI3K-AKT was targeted [287].
Su, et al. argue that under normal physiologic conditions, PI3K-AKT pathway actively regulates body functions including metabolism and proliferation. PI3K-AKT pathway regulates glucose metabolism through FOXO1 and GSK-3. PI3K-AKT also regulates lipid metabolism through mTORC1 and SREBP. Active AKT inhibits FOXO1 which reduces glucose levels [288,289]. Similarly, active AKT inhibits mTOR complex 1 which consequently reduces lipid and protein production [290]. GSK3 is inhibited also by AKT which leads to glycogen synthesis, thus reducing glucose levels [291]. Lipid metabolism is regulated by AKT activity through sterol regulatory element-binding proteins (SREBP). SREBP increases fatty acid and cholesterol accumulation [287,290,292].
However, when there is chronic excessive energy intake as in obesity, PI3K-AKT signaling becomes suppressed. A state in which re-activating PI3K-AKT would lessen obesity and insulin resistance. Nevertheless, it is in the established disease states of cancer and/or obesity is where there is dysregulation and/or overexpression of PI3K-AKT. At which point, therapeutic inhibition of PI3K-AKT becomes an effective anti-obesity and anti-cancer treatment approach [287].
Thus, Su et al, details the mechanism by which of PI3K-AKT pathway acts in an organ-specific manner [287]. That further explains why targeting PI3K whether by inhibition or activation would be favorable depending on the context [287]. For instance, one study showed that pharmacological inhibition of PI3K-AKT activity reduced adiposity and metabolic syndrome in obese mice and rhesus monkeys [293]. They used two small molecules with selective inhibitory action on PI3K (CNIO-PI3Ki and GDC-0941) as pharmacological inhibitors [293]. In contrast, over expression of FAM3A in the liver activates PI3K p110α-AKT signaling in the liver and decreases hepatic gluconeogenesis and lipogenesis [294].
5. Nutritional Therapeutic Approaches in Cancer and T2D
Two interesting dietary compounds that can be taken orally are alpha-lipoic acid (ALA) and flavonoids. They are natural compounds with minimal-to-no side effects. These nutritional supplements have shown promising results in vitro and in vivo, as well as in combination therapies in patients with diabetes and cancer [295,296,297,298].
5.1. Nutritional- and Dietary-Based Approaches Targeting Mitochondrial Metabolism in Cancer
Reportedly, nutrients, dietary supplements, and dietary habits are beneficial in the treatment and prevention of cancer and diabetes. Although their underpinning mechanisms of action are still being investigated, studies have shown that nutrients and/or dietary approaches are effective metabolic modulators in metabolic-based diseases. Herein, we discuss the mechanisms by which ALA, flavonoids, Omega-3 polyunsaturated fatty acids (ω-3 PUFA), and artemisinin can modulate the metabolic imbalance in cancer and/or diabetes and reverse the disease state.
5.1.1. Alpha-lipoic Acid as a Metabolic Modulator in Cancer
ALA is a naturally occurring dithiol compound that is produced physiologically in the body from octanoic acid in the mitochondria. It can also be found in a variety of food and dietary supplements. ALA, through its various metabolic regulatory effects, can inhibit the proliferation, migration, and invasion of tumor cells and can induce apoptosis [299]. ALA has been well known for acting as a metal chelator and a ROS scavenger [299,300,301]. More importantly, ALA acts as a cofactor for several enzyme complexes, such as PDC, and promotes mitochondrial respiration [299]. Earlier studies reported that treatment with ALA decreased serum levels of pyruvate and lactate in both lean and obese individuals with T2D [302]. Later, ALA was reported to increase PDC activity in rat hepatocytes and in the mitochondria of hepatocytes in diabetic rat models [303,304]. In 2004, Patel et al. reported that ALA could minimize or block the inhibitory phosphorylation of the E1 subunit of the PDC complex via pyruvate kinase, thereby increasing the activity of PDC [303].
ALA is well known for its antioxidant action that increases glutathione peroxidase activity, which in turn reduces oxidative stress [300,305,306]. These antioxidant effects were seen in advanced-stage cancer patients who were administered ALA treatment for 10 consecutive days [306,307]. Nevertheless, ALA also plays the role of a prooxidant by increasing the production of free oxygen radicals in the mitochondria of colon cancer cell lines, but not in nontransformed cells [305]. This prooxidant effect is a result of ALA stimulating mitochondrial OXPHOS and inducing a cytotoxic effect on cancer cells both in in vitro and in vivo models [299,308]. In 2005, Wenzel et al. discovered that ALA induced mitochondrial OXPHOS in a colon cancer cell line (HT-29) and stimulated apoptosis [305]. ALA-induced apoptosis also occurs selectively in colon cancer cells, but not in the nontransformed cells [305]. Moreover, ALA-induced apoptosis ensued predominantly via the intrinsic mitochondrial apoptotic pathway and was independent of p53 [309]. Wenzel et al. further reported that ALA along with its reduced form, i.e., dihydrolipoic acid (DHLA), were able to trigger apoptosis in cancer cells by increasing the production of mitochondrial ROS, following an increased influx of lactate and pyruvate into the mitochondria. This effect was also associated with the downregulation of antiapoptotic protein BCL-Xl [305]. While studying two ovarian cancer cell lines (cisplatin-resistant and cisplatin-sensitive), Kafar et al. showed that ALA treatment induced apoptosis via downregulating the gene expression of antiapoptotic genes, MCL-1 and BCL-Xl, and by upregulating the expression of Bim, a pro-apoptotic gene [310]; another study showed similar results [300]. In line with these findings, KIM et al. reported that ALA treatment caused apoptosis in a dose-dependent manner in an in vitro setting in the MDA-MB-231 breast cancer cell line [300]. ALA promoted apoptosis by increasing the mRNA and protein expression of BAX and by decreasing the mRNA and protein expression of BCL2 [300]. In other cellular contexts, ALA prevented apoptosis [311,312]. Interestingly, ALA successfully reversed the Warburg effect and inhibited glycolysis through the inhibition of PDK [296,313].
ALA has also been shown to modulate mitochondrial metabolism through the activation of AMPK signaling. Shen et al. (2007) revealed that ALA activated AMPK with an increased phosphorylation of AMPK at Thr172 in C2CL2 myotubes. Shen and colleagues found that ALA acted by enhancing Ca+2/calmodulin-dependent protein kinase kinase (CAMKK), and not through AMP-LKB1 signaling [314]. ALA activation of AMPK and the subsequent inhibition of mTOR-S6 signaling suppressed thyroid cancer cell proliferation in vivo in several thyroid cancer cell lines, including BCPAP, HTH-83, CAL-62, and FTC-133 [301]. Additionally, ALA decreased the migration and invasion of cancer cells in thyroid cancer cell lines by inhibiting transforming growth factor β (TGFβ) production and signaling cascade [301]. ALA further induced cell cycle arrest through the upregulation of cyclin-dependent kinase inhibitors, p27kip1 and p21cip1 [315].
Altogether, studies point towards a pleotropic effect of ALA on cancer cells depending on the type of cell and tumor.
5.1.2. Flavonoids as a Metabolic Modulator in Cancer
Similar to ALA, flavonoids are a family of natural polyphenolic compounds found in fruits and vegetables that have promising anticancer effects [297,298,316]. They appear to modulate mitochondrial metabolism in cancer and reverse Warburg glycolysis [317,318,319]. In 2017, Wei et al. reported that Didymin, a natural flavonoid, inhibited the proliferation of the liver cancer cell line HepG2 by decreasing cyclin B1, cyclin D1, and cyclin CDK4 [317]. Didymin also induced apoptosis in HepG2 cells by altering the BCL-2/BAX ratio and by stimulating caspase-mediated apoptosis. Moreover, Wei et al. showed that Didymin could downregulate the ERK/MAPK and PI3K/Akt pathways by upregulating the Raf kinase inhibitory protein (RKIP). This study confirmed earlier observations made by Singhal et al. (2012), which demonstrated, both in vivo and in vitro, that Didymin induced G2/M arrest and apoptosis in neuroblastoma cells and upregulated RKIP [320]. Zhao et al. recently reported that brosimone I, another flavonoid, induced apoptosis and cell cycle arrest through ROS-mediated endoplasmic reticulum stress and AMPK pathway activation in the human colon cancer cell line HCT116. The activation of AMPK depended on an increase in Ca+2 ions and the activation of the CaMKKβ-AMPK pathway, but not on AMP [318]. Reportedly, other members of the flavonoid family have reversed Warburg glycolysis and promoted OXPHOS in in vitro and in vivo preclinical cancer studies [321]. Table 4 summarizes a few of these findings pertaining to a group of flavonoids.
Although preclinical studies have positioned dietary flavonoids as potential candidates for treating and/or preventing cancer [316,321,345], these supplements have not yet shown substantial efficacy in clinical trials. In 2020, Bisol et al. published a systematic review of clinical trials where flavonoids were studied as potential therapeutic agents in cancer [346]. They identified 22 Phase II and one Phase III clinical trials that administered flavonoids as either monotherapy or in combination with other chemotherapeutic agents. Twelve of these clinical trials enrolled patients with solid tumors, while the other 11 trials included patients with hematopoietic or lymphoid malignancies [346]. Overall, low rates of complete or partial response to flavonoids treatment were reported in clinical trials. Additionally, positive outcomes were mostly associated with hematopoietic or lymphoid tumors, compared to solid tumors [346]. These clinical trials had various limitations, including small sample sizes, variations in the administered doses, design of randomized trials, and tumor subtypes of patients [346]. In addition to this, the limited bioavailability and varied absorption of administered flavonoids further limit its efficacy [347]. Studies are being performed to improve the bioavailability of flavonoids by enhancing the metabolic stability and absorption of the administered flavonoids [347,348].
Therefore, a greater number of well-designed clinical trials are required to test the efficacy of flavonoids for cancer treatment.
5.2. Nutritional- and Dietary-Based Approaches Targeting Mitochondrial Metabolism in T2D
Accumulating evidence shows that flavonoids and ALA have beneficial effects as adjuvant and dietary supplements for the treatment of patients with T2D.
5.2.1. ALA as a Metabolic Modulator in T2D
ALA functions as an antioxidant and an anti-inflammatory agent and is reportedly beneficial in treating patients with T2D. It exerts its antioxidant effects by quenching ROS; chelating metallic ions; and reducing the oxidized forms of glutathione, vitamin C, and vitamin E. Moreover, it boosts the antioxidant machinery by enhancing the NRF-2-mediated antioxidant gene expression. It also acts by activating AMPK in the SKM and inhibiting NFκB [295]. Moreover, ALA activates hepatic AMPK, leading to decreased gluconeogenesis and glucose output from the liver [349]. It also activates AMPK in the SKM, which leads to an increase in glucose uptake and fatty acid oxidation [350]. Research suggests that ALA activates AMPK in the liver and SKM through increased intracellular calcium ion concentration, and not through LKB1 activation [314]. Surprisingly, ALA was found to inhibit hypothalamic AMPK, leading to a reduction in food intake and body weight [295]. These metabolic effects of ALA have been tested in pre-diabetic volunteers in a randomized, placebo-controlled pilot study [351]. Twelve volunteers who were eligible and met the criteria for prediabetes were included in the study and took ALA supplementation (600 mg/day) for 30 days. ALA improved glycemic control and insulin sensitivity in pre-diabetic volunteers (assessed by HOMA-IR and fasting serum insulin); it, however, had no effect on the lipid profile. In another study, diabetic individuals who orally consumed 600 mg of ALA supplementation (twice a day) also showed improved insulin sensitivity, assessed by a 2-hour manual hyper-insulinemic euglycemic clamp technique, expressed as a glucose disposal rate and insulin sensitivity index [352]. Moreover, patients with diabetic nephropathy were shown to benefit from the oral administration of ALA [353].
5.2.2. Flavonoids as a Metabolic Modulator in T2D
Flavonoids have demonstrated beneficial effects in T2D via metabolic reprogramming in pancreatic β-cells, hepatocytes, adipocytes, and SKM [297].
In an in vitro study by Kyriakis et al., two flavonoids, gallic acid and its dimer ellagic acid, were found to bind to glycogen phosphorylase and inhibit its action, thus decreasing glycogen metabolism and glucose production [320]. Therefore, it was suggested that gallic acid and ellagic acid could be administered as antihyperglycemic agents.
Lagouge et al. showed that mice fed with a high-fat diet and administered a flavonoid called resveratrol did not develop obesity or insulin resistance. Resveratrol induced mitochondrial OXPHOS and improved muscle respiratory capacity by activating peroxisome proliferator-activated receptor gamma coactivator 1-alpha (PGC-1α) through Sirtuin-1 (SIRT-1)-mediated deacetylation [354].
Recently, the therapeutic effects of flavonoids were further confirmed by Meng et al. [355]. This study showed that flavonoids extracted from mulberry leaves activated AMPK in mice with spontaneous T2D and enhanced glucose uptake and OXPHOS in L6 SKM cell line. Flavonoids also induced the expression of PGC-1α and the upregulation of GLUT4 [355].
Generally, flavonoids were reported to enhance insulin sensitivity, decrease ROS, and mitigate inflammation in SKM and adipose tissues [297]. Flavonoids could enhance insulin secretion by pancreatic β-cells and reduce apoptosis in these cells. They also enhanced glucose uptake by SKM and white adipose tissue [297]. These encouraging results need to be further navigated and confirmed in clinical trials to be administered for therapeutic purposes in patients with T2D.
5.2.3. Effects of glutamine supplementation on T2D
Glutamine pathway plays a protective role in T2D. Studies have shown that glutamine metabolism plays an important role in insulin signaling and glucose metabolism [356]. Specifically, the TCA cycle intermediates generated from glutamine metabolism can stimulate insulin secretion and enhance insulin sensitivity in various tissues, including the liver, muscle, and adipose tissue [357,358,359,360]. For example, α-ketoglutarate has been shown to stimulate insulin secretion in pancreatic beta cells. α-ketoglutarate is converted to succinyl-CoA, which in turn activates the ATP-sensitive potassium channel, leading to depolarization of the cell membrane and subsequent calcium influx. The calcium influx triggers insulin secretion from the beta cells [361]. In addition, other TCA cycle intermediates such as citrate and malate have also been shown to stimulate insulin secretion. Citrate can enhance insulin secretion by activating the exocytotic machinery in pancreatic β cells [362], while malate can increase ATP production and stimulate insulin secretion [363].
Interestingly, α-ketoglutarate and succinate can activate the mTOR signaling pathway, which in turn enhances insulin signaling and glucose uptake [364,365]. The activation of mTOR stimulates the activity of IRS-1, a key signaling molecule in the insulin signaling pathway. This enhances the translocation of GLUT4 to the membrane and increases glucose uptake in insulin-sensitive tissues such as skeletal muscle and adipose tissue [366].
In addition to mTOR signaling, TCA cycle intermediates can also enhance insulin sensitivity by regulating the activity of key metabolic enzymes. For example, α-ketoglutarate can induce the activity of PDH thus enhancing glucose oxidation and improving insulin sensitivity [367]. Likewise, succinate has been shown to inhibit the activity of HIF-1α, adding to its role in cancer, it plays a role in glucose metabolism and insulin sensitivity [368,369].
Previous studies indicate a possible impact of glutamine on oxidative stress and inflammatory markers. In animal studies, supplementation with glutamine demonstrated a notable elevation in antioxidant proteins such as superoxide dismutase, glutathione peroxidase (GPx), and catalase levels [370,371,372,373], along with significant improvements in inflammatory markers levels such as c-reactive protein, interleukins 6 and 23, and monocyte chemoattractant protein-1 [374]. The antioxidant effect of glutamine may be attributed to its involvement in glutathione synthesis, leading to increased enzymatic activity of GPx and a reduction in ROS production [370,371].
A comprehensive systematic review conducted by Maleki's team in 2020 revealed interesting findings. Among the 19 examined studies, nine highlighted a significant rise GLP-1 levels in the sera. Furthermore, eight studies showed a reduction in fasting blood sugar levels, with four studies reporting decreases in postprandial blood sugar and triglyceride levels after glutamine supplementation. While seven studies demonstrated a significant increase in insulinemia with glutamine, the outcomes regarding Hb-A1c levels were inconclusive [375].
Overall, the TCA cycle intermediates generated from glutamine metabolism can enhance insulin sensitivity by regulating multiple signaling pathways and metabolic enzymes. Understanding the complex interplay between glutamine metabolism and insulin signaling may provide insights into the development of new therapies for insulin resistance and related metabolic disorders.
Discussion:
Targeting the metabolic paradigms of cancer and those of diabetes exhibits a yin-yang relationship. The yin-yang concept from Chinese philosophy symbolizes two opposing yet interconnected forces. Metformin is a common therapeutic target in both T2D and cancer. Several meta-analysis studies over the last decade have reported that diabetic patients receiving metformin have lower risk to develop cancer [376,377,378]. Moreover, metformin was able to improve survival and response to treatment in cancer patients [377,378,379,380]. These studies corroborated previous in vitro and in vivo studies in animal models that showed metformin exhibiting anti-cancer effects [190,377,378,381]. For instance, Noto et al, 2012 conducted a systematic meta-analysis on 6 studies (4 cohort studies, 2 RCTs) with a total of 210,829 diabetic patients data [376]. They found that diabetic patients taking metformin had significantly lower risk of cancer incidence and cancer mortality using pooled relative risk measures. One of the earliest meta-analysis studies done by DeCensi et al. in 2010, showed a 31% reduction in overall relative risk of cancer incidence in subjects receiving metformin compared to other anti-diabetic treatments [382]. Another meta-analysis study by Xie et al, (2014), performed on data from 13 observational studies (10 cohort, 3 case control), found that the use of metformin was associated with reduced risk of pancreatic cancer in T2D patients. In another observational study by Lee et al. (2020), involving a Korean cohort of 323,430 individuals with a median follow-up of 12.7 years, data were extracted from national health records spanning 2002 to 2015. The findings indicated that diabetic individuals undergoing Metformin treatment had a reduced risk of cancer incidence compared to diabetic patients not receiving Metformin, with an incidence percentage of 10.3% in Metformin users compared to 11.1% in non-Metformin users [383]. Similar results have been reported in other meta-analysis retrospective studies [384,385,386,387,388,389]. However, a study conducted in the UK showed no protective effect of Metformin against cancer incidence in diabetic patients [390]. An insightful review by Moradi-Kor et al. (2019) aimed to explain the mechanisms by which Metformin exerts its beneficial effects in cancer [377]. The review also encompassed clinical trials conducted to confirm the beneficial effects of Metformin on cancer. Based on this analysis, several clinical trials took place in non-diabetic patients to test the effects of Metformin, but the results were inconclusive in proving a protective anti-cancer effect in non-diabetic patients. Therefore, further investigations are needed.
Although Metformin is the most commonly studied as an AMPK activator, there are other physiological and pharmacological activators of AMPK. Acting either directly or indirectly to activate AMPK, they raise the question of whether other AMPK activators exhibit the same anti-cancer preventive effects in diabetic patients. Thiazolidinediones (TZDs), including troglitazone, pioglitazone, and rosiglitazone, represent another class of anti-diabetic drugs known to activate AMPK.
Other studies showed absence of any significant association between cancer risk and taking TZD’s in diabetic patients [391,392,393]. Other studies have shown that T2D patients who are taking TZD’s had lower cancer risk in certain cancer types [391,393,394,395]. Interestingly some clinical studies have shown that patients taking TZD’s have an increased risk of cancer [396,397]. Thus, observations and associations have been conflicting and inconclusive in meta-analysis studies. This is attributed to methodological variations within these studies and the intricacy of the disease [398].Thus, more studies are needed. However, the evidence that associates metformin use in T2D patients with lower risk of cancer is stronger and more consistent among studies [398]. Although Metformin (belongs to Biguanides) and TZD’s can activate indirectly AMPK through inhibiting complex-1 in the mitochondrial respiratory chain (ETC cycle), Metformin also acts in a non-AMPK dependent manner [399]. Metformin's impact on the liver are mediated by antagonizing glucagon signaling through cyclic AMP and PKA, operating independently of AMPK [400,401]. In contrast, TZD’s activate AMPK by targeting the nuclear hormone receptor peroxisome proliferator- activated receptors (PPARs) which in turn stimulate the secretion of adiponectin and consequently activates AMPK [402]. Other AMPK activators such as polyphenols and 5-Aminoimidazole-4-carboxamide riboside (AICARs) have not been studies in the context of anti-diabetic drugs and the risk of cancer. Yet, it is worth noting that similar anti-cancer effects to those of metformin have also been observed with other AMPK activators in-vitro and in-vivo. This suggests that metformin may not be the only drug with dual effects and other AMPK activators might exhibit promising anti-cancer effects as well as anti-diabetic ones [402].
6. Conclusion
Both cancer and T2D are intricate metabolic disorders characterized by distinct yet seemingly opposite metabolic shifts. Cancer, being predominantly glycolytic, opposes the OXPHOS-based metabolism observed in T2D. Multiple factors, such as mitochondrial dysfunction, low AMPK, elevated PDK levels, LDH, HIF-1α, decreased PDC levels, NADH, mutations in oncogenes and tumor suppressor genes, and other tumor microenvironmental influences, can drive the dominant bioenergetic shift in tumors towards the Warburg effect. The presence of one or more of these factors can dictate the course of the bioenergetic profile of the tumor. In contrast to cancer, T2D exhibits intact mitochondrial OXPHOS machinery, high ROS levels, high hepatic gluconeogenesis, and insulin resistance. Targeted metabolic therapies aimed at rectifying the metabolic imbalance in cancer and diabetes exhibit promising outcomes in in vivo and in vitro preclinical analyses, along with several ongoing clinical trials. Further investigations into the efficacy and safety of potential nutrient adjuvants and therapies for patients with cancer and diabetes are much warranted. These metabolic corrective therapies have also demonstrated preventive and protective effects against cancer and diabetes, aspects that should not be overlooked.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org.
Author Contributions
All authors participated in writing the manuscript in descending order. FAM conceived the idea, while all participating authors guided the research study, provided material support, reviewed, edited, and critically commented on the manuscript.
Funding
This work is supported by the Kuwait Foundation for the Advancement of Sciences (KFAS), with grant numbers P11613MG02 and RA HM-2021-001.
Data Availability Statement
This review did not rely on any specific dataset. All referenced data is accounted for in the references section. The information presented in supplementary Table S1 was obtained from ClinicalTrials.gov on November 23, 2023, and any alterations made to the data are explicitly outlined in the legend in accordance with the ClinicalTrials.gov "Use of Data" policy.
Acknowledgments
All figures were created with BioRender.com.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- DeBeradinis R, Thompson C. Cellular Metabolism And Disease: What Do Metabolic Outliers Teach Us? NIH Public Access. 2012;148:1132–44.
- O’Connor C, Adams JU. Essentials of cell biology. 2001.
- Xian Zhang P, Qi Xing Y, Dong Niu Y. Fundamentals Of The Warburg Effect In Cancer. Journal of Nutritional Oncology. 2019;4:108–14.
- O. Warburg. On The Origin Of Cancer Cells. Science. 1956;123:309–14. [CrossRef]
- Warburg O. The Metabolism Of Carcinoma Cells. The Journal of Cancer Research. 1925;9:148–63. [CrossRef]
- Warburg O, Minami S. Experiments On Surviving Carcinoma Tissue. Journal Of Moelcular Medicine. 1923;2:776–7.
- Koukourakis MI, Giatromanolaki A, Bougioukas G, Sivridis E. Lung Cancer: A Comparative Study Of Metabolism Related Protein Expression In Cancer Cells And Tumor Associated Stroma. Cancer Biology & Therapy. 2007;6:1472–5.
- Isidoro A, Casado E, Redondo A, Acebo P, Espinosa E, Alonso AM, Cejas P, Hardisson D, Fresno Vara JA, Belda-Iniesta C, González-Barón M, Cuezva JM. Breast Carcinomas Fulfill The Warburg Hypothesis And Provide Metabolic Markers Of Cancer Prognosis. Carcinogenesis. 2005;26:2095–104. [CrossRef]
- Leung CO ning, Wong CC lui, Fan DN yin, Kai AK lun, Tung EK kwan, Xu IM jing, Ng IO lin, Lo RC lam. PIM1 Regulates Glycolysis And Promotes Tumor Progression In Hepatocellular Carcinoma. Oncotarget. 2015;6:10880–92.
- Fan P, Wang B, Meng Z, Zhao J, Jin X. PES1 Is Transcriptionally Regulated By BRD4 And Promotes Cell Proliferation And Glycolysis In Hepatocellular Carcinoma. International Journal of Biochemistry and Cell Biology. 2018;104:1–8. [CrossRef]
- Marie SKN, Shinjo SMO. Metabolism And Brain Cancer. Clinics. 2011;66:33–43. [CrossRef]
- Martiny PB, Alcoba DD, Neto BS, Carvalho PC, Brum IS. A Proteomic Glimpse Into The Oncogenesis Of Prostate Cancer. Journal of Applied Biomedicine. 2018;16:328–36. [CrossRef]
- Kobayashi Y, Banno K, Kunitomi H, Takahashi T, Takeda T, Nakamura K, Tsuji K, Tominaga E, Aoki D. Warburg Effect In Gynecologic Cancers. Journal of Obstetrics and Gynaecology Research. 2019;45:542–8.
- Zhou W, Capello M, Fredolini C, Racanicchi L, Piemonti L, Liotta LA, Novelli F, Petricoin EF. Proteomic Analysis Reveals Warburg Effect And Anomalous Metabolism Of Glutamine In Pancreatic Cancer Cells. Journal of Proteome Research. 2012;11:554–63. [CrossRef]
- Altenberg B, Greulich KO. Genes Of Glycolysis Are Ubiquitously Overexpressed In 24 Cancer Classes. Genomics. 2004;84:1014–20. [CrossRef]
- Kim TM, Paeng JC, Chun IK, Keam B, Jeon YK, Lee SH, Kim DW, Lee DS, Kim CW, Chung JK, Kim IH, Heo DS. Total Lesion Glycolysis In Positron Emission Tomography Is A Better Predictor Of Outcome Than The International Prognostic Index For Patients With Diffuse Large B Cell Lymphoma. Cancer. 2013;119:1195–202. [CrossRef]
- Guo B, Tan X, Ke Q, Cen H. Prognostic Value Of Baseline Metabolic Tumor Volume And Total Lesion Glycolysis In Patients With Lymphoma: A Meta-Analysis. PLoS ONE. 2019;14:1–17. [CrossRef]
- Suganuma K, Miwa H, Imai N, Shikami M, Gotou M, Goto M, Mizuno S, Takahashi M, Yamamoto H, Hiramatsu A, Wakabayashi M, Watarai M, Hanamura I, Imamura A, Mihara H, Nitta M. Energy Metabolism Of Leukemia Cells: Glycolysis Versus Oxidative Phosphorylation. Leukemia and Lymphoma. 2010;51:2112–9.
- Chen L, Hu N, Wang C, Zhao H. HOTAIRM1 Knockdown Enhances Cytarabine-Induced Cytotoxicity By Suppression Of Glycolysis Through The Wnt/β-Catenin/PFKP Pathway In Acute Myeloid Leukemia Cells. Archives of Biochemistry and Biophysics. 2020;680:108244. [CrossRef]
- Whitaker-Menezes D, Martinez-Outschoorn UE, Flomenberg N, Birbe RC, Witkiewicz AK, Howell A, Pavlides S, Tsirigos A, Ertel A, Pestell RG, Broda P, Minetti C, Lisanti MP, Sotgia F. Hyperactivation Of Oxidative Mitochondrial Metabolism In Epithelial Cancer Cells In Situ: Visualizing The Therapeutic Effects Of Metformin In Tumor Tissue. Cell Cycle. 2011;10:4047–64.
- Viale A, Corti D, Draetta GF. Tumors And Mitochondrial Respiration: A Neglected Connection. Cancer Research. 2015;75:3685–6. [CrossRef]
- Simonnet H. Low Mitochondrial Respiratory Chain Content Correlates With Tumor Aggressiveness In Renal Cell Carcinoma. Carcinogenesis. 2002;23:759–68. [CrossRef]
- Faure-Vigny H, Heddi A, Giraud S, Chautard D, Stepien G. Expression Of Oxidative Phosphorylation Genes In Renal Tumors And Tumoral Cell Lines. Molecular Carcinogenesis. 1996;16:165–72. [CrossRef]
- Hayashi JI, Takemitsu M, Nonaka I. Recovery Of The Missing Tumorigenicity In Mitochondrial DNA-Less HeLa Cells By Introduction Of Mitochondrial DNA From Normal Human Cells. Somatic Cell and Molecular Genetics. 1992;18:123–9. [CrossRef]
- Cavalli LR, Varella-Garcia M, Liang BC. Diminished Tumorigenic Phenotype After Depletion Of Mitochondrial DNA. Cell Growth and Differentiation. 1997;8:1189–98.
- Tan AS et al. Mitochondrial Genome Acquisition Restores Respiratory Function And Tumorigenic Potential Of Cancer Cells Without Mitochondrial DNA. Cell Metabolism. 2015;21:81–94.
- Lebleu VS, O’Connell JT, Gonzalez Herrera KN, Wikman H, Pantel K, Haigis MC, De Carvalho FM, Damascena A, Domingos Chinen LT, Rocha RM, Asara JM, Kalluri R. PGC-1α Mediates Mitochondrial Biogenesis And Oxidative Phosphorylation In Cancer Cells To Promote Metastasis. Nature Cell Biology. 2014;16:992–1003. [CrossRef]
- Yang L, Venneti S, Nagrath D. Glutaminolysis: A Hallmark Of Cancer Metabolism. Annual Review of Biomedical Engineering. 2017;19:163–94. [CrossRef]
- Moreadith RW, Lehninger AL. The Pathways Of Glutamate And Glutamine Oxidation By Tumor Cell Mitochondria. Role Of Mitochondrial NAD(P)+-Dependent Malic Enzyme. Journal of Biological Chemistry. 1984;259:6215–21. [CrossRef]
- Schiliro C, Firestein BL. Mechanisms Of Metabolic Reprogramming In Cancer Cells Supporting Enhanced Growth And Proliferation. Cells 2021, Vol 10, Page 1056. 2021;10:1056. [CrossRef]
- Fan J, Kamphorst JJ, Mathew R, Chung MK, White E, Shlomi T, Rabinowitz JD. Glutamine-Driven Oxidative Phosphorylation Is A Major ATP Source In Transformed Mammalian Cells In Both Normoxia And Hypoxia. Molecular Systems Biology. 2013;9:712. [CrossRef]
- Tardito S et al. Glutamine Synthetase Activity Fuels Nucleotide Biosynthesis And Supports Growth Of Glutamine-Restricted Glioblastoma. Nature Cell Biology. 2015;17:1556–68. [CrossRef]
- WILLIAMS JIMC, KIZAKI H, WEBER G, MORRIS HP. Increased CTP Synthetase Activity In Cancer Cells. Nature. 1978;271:71–3. [CrossRef]
- DeFronzo RA, Tripathy D. Skeletal Muscle Insulin Resistance Is The Primary Defect In Type 2 Diabetes. Diabetes Care. 2009;32:S157–63. [CrossRef]
- Weir GC, Leahy JL, Bonner-Weiner S. B-Cell Dysfunction Induced By Chronic Hyperglycemia. Diabetes Care. 1992;15:442–55.
- Robertson R, Zhou H, Zhang T, Harmon JS. Chronic Oxidative Stress As A Mechanism For Glucose Toxicity Of The Beta Cell In Type 2 Diabetes. Cell Biochemistry and Biophysics. 2007;48:139–46. [CrossRef]
- Sakai K, Matsumoto K, Nishikawa T, Suefuji M, Nakamaru K, Hirashima Y, Kawashima J, Shirotani T, Ichinose K, Brownlee M, Araki E. Mitochondrial Reactive Oxygen Species Reduce Insulin Secretion By Pancreatic β-Cells. Biochemical and Biophysical Research Communications. 2003;300:216–22. [CrossRef]
- Lortz S, Tiedge M. Glucose Toxicity In B-Cells: Type 2 Diabetes, Good Radicals Gone Bad, And The Glutathione Connection. Free Radical Biology and Medicine. 2003;34:581–7.
- Larsen S, Scheede-Bergdahl C, Whitesell T, Boushel R, Bergdahl A. Increased Intrinsic Mitochondrial Respiratory Capacity In Skeletal Muscle From Rats With Streptozotocin-Induced Hyperglycemia. Physiological Reports. 2015;3:1–10. [CrossRef]
- Boirie Y. Insulin Regulation Of Mitochondrial Proteins And Oxidative Phosphorylation In Human Muscle. Trends in Endocrinology and Metabolism. 2003;14:393–4. [CrossRef]
- Zhang Y, Ye J. Mitochondrial Inhibitor As A New Class Of Insulin Sensitizer. Acta Pharmaceutica Sinica B. 2012;2:341–9. [CrossRef]
- Stump CS, Short KR, Bigelow ML, Schimke JM, Nair KS. Effect Of Insulin On Human Skeletal Muscle Mitochondrial ATP Production, Protein Synthesis, And MRNA Transcripts. Proceedings of the National Academy of Sciences of the United States of America. 2003;100:7996–8001.
- Špaček T, Šantorová J, Zacharovová K, Berková Z, Hlavatá L, Saudek F, Ježek P. Glucose-Stimulated Insulin Secretion Of Insulinoma INS-1E Cells Is Associated With Elevation Of Both Respiration And Mitochondrial Membrane Potential. International Journal of Biochemistry and Cell Biology. 2008;40:1522–35. [CrossRef]
- Ahmad W, Ijaz B, Shabbiri K, Ahmed F, Rehman S. Oxidative Toxicity In Diabetes And Alzheimer’s Disease: Mechanisms Behind ROS/ RNS Generation. Journal of Biomedical Science. 2017;24:1–10. [CrossRef]
- Bhansali S, Bhansali A, Walia R, Saikia UN, Dhawan V. Alterations In Mitochondrial Oxidative Stress And Mitophagy In Subjects With Prediabetes And Type 2 Diabetes Mellitus. Frontiers in Endocrinology. 2017;8. [CrossRef]
- Bitar MS, Al-Saleh E, Al-Mulla F. Oxidative Stress - Mediated Alterations In Glucose Dynamics In A Genetic Animal Model Of Type II Diabetes. Life Sciences. 2005;77:2552–73. [CrossRef]
- Petersen KF, Dufour S, Befroy D, Garcia R, Shulman GI. Impaired Mitochondrial Activity In The Insulin-Resistant Offspring Of Patients With Type 2 Diabetes. New England Journal of Medicine. 2004;350:664–71. [CrossRef]
- Kelley DE, He J, Menshikova E V., Ritov VB. Dysfunction Of Mitochondria In Human Skeletal Muscle In Type 2 Diabetes. Diabetes. 2002;51:2944–50. [CrossRef]
- Frederiksen CM, Højlund K, Hansen L, Oakeley EJ, Hemmings B, Abdallah BM, Brusgaard K, Beck-Nielsen H, Gaster M. Transcriptional Profiling Of Myotubes From Patients With Type 2 Diabetes: No Evidence For A Primary Defect In Oxidative Phosphorylation Genes. Diabetologia. 2008;51:2068–77. [CrossRef]
- Boushel R, Gnaiger E, Schjerling P, Skovbro M, Kraunsøe R, Dela F. Patients With Type 2 Diabetes Have Normal Mitochondrial Function In Skeletal Muscle. Diabetologia. 2007;50:790–6. [CrossRef]
- Fisher-Wellman KH, Weber TM, Cathey BL, Brophy PM, Gilliam LAA, Kane CL, Maples JM, Gavin TP, Houmard JA, Neufer PD. Mitochondrial Respiratory Capacity And Content Are Normal In Young Insulin-Resistant Obese Humans. Diabetes. 2014;63:132–41. [CrossRef]
- Misu H, Takamura T, Matsuzawa N, Shimizu A, Ota T, Sakurai M, Ando H, Arai K, Yamashita T, Honda M, Yamashita T, Kaneko S. Genes Involved In Oxidative Phosphorylation Are Coordinately Upregulated With Fasting Hyperglycaemia In Livers Of Patients With Type 2 Diabetes. Diabetologia. 2006;50:268–77. [CrossRef]
- Buchner DA, Yazbek SN, Solinas P, Burrage LC, Morgan MG, Hoppel CL, Nadeau JH. Increased Mitochondrial Oxidative Phosphorylation In The Liver Is Associated With Obesity And Insulin Resistance. Obesity. 2011;19:917–24. [CrossRef]
- Ciapaite J, Bakker SJL, Van Eikenhorst G, Wagner MJ, Teerlink T, Schalkwijk CG, Fodor M, Ouwens DM, Diamant M, Heine RJ, Westerhoff H V., Krab K. Functioning Of Oxidative Phosphorylation In Liver Mitochondria Of High-Fat Diet Fed Rats. Biochimica et Biophysica Acta - Molecular Basis of Disease. 2007;1772:307–16. [CrossRef]
- Petersen KF, Befroy D, Dufour S, Dziura J, Ariyan C, Rothman DL, DiPietro L, Cline GW, Shulman GI. Mitochondrial Dysfunction In The Elderly: Possible Role In Insulin Resistance. Science. 2003;300:1140–2. [CrossRef]
- Chen H, Fang Y, Liang L, Wang C. Insulin Resistance Leads to Mitochondrial Dysfunction in Hepatocyte. Eur. Soc. Paediatr. Endocrinol., vol. 92, 2019.
- Sasaki M, Fujimoto S, Sato Y, Nishi Y, Mukai E, Yamano G, Sato H, Tahara Y, Ogura K, Nagashima K, Inagaki N. Reduction Of Reactive Oxygen Species Ameliorates Metabolism-Secretion Coupling In Islets Of Diabetic GK Rats By Suppressing Lactate Overproduction. Diabetes. 2013;62:1996–2003. [CrossRef]
- Cantley J, Biden TJ. Sweet And Sour β-Cells: ROS And Hif1α Induce Warburg-Like Lactate Production During Type 2 Diabetes. Diabetes. 2013;62:1823–5. [CrossRef]
- Choi CS, Kim YB, Lee FN, Zabolotny JM, Kahn BB, Youn JH. Lactate Induces Insulin Resistance In Skeletal Muscle By Suppressing Glycolysis And Impairing Insulin Signaling. American Journal of Physiology - Endocrinology and Metabolism. 2002;283:233–40. [CrossRef]
- Akram M. Citric Acid Cycle And Role Of Its Intermediates In Metabolism. Cell Biochemistry and Biophysics. 2014;68:475–8. [CrossRef]
- Akram M. Mini-Review On Glycolysis And Cancer. Journal of Cancer Education. 2013;28:454–7. [CrossRef]
- Nolfi-Donegan D, Braganza A, Shiva S. Mitochondrial Electron Transport Chain: Oxidative Phosphorylation, Oxidant Production, And Methods Of Measurement. Redox Biology. 2020:101674. [CrossRef]
- Simonnet H, Demont J, Pfeiffer K, Guenaneche L, Bouvier R, Brandt U, Schägger H, Godinot C. Mitochondrial Complex I Is Deficient In Renal Oncocytomas. Carcinogenesis. 2003;24:1461–6. [CrossRef]
- Baracca A, Chiaradonna F, Sgarbi G, Solaini G, Alberghina L, Lenaz G. Mitochondrial Complex I Decrease Is Responsible For Bioenergetic Dysfunction In K-Ras Transformed Cells. Biochimica et Biophysica Acta - Bioenergetics. 2010;1797:314–23. [CrossRef]
- Bonora E, Porcelli AM, Gasparre G, Biondi A, Ghelli A, Carelli V, Baracca A, Tallini G, Martinuzzi A, Lenaz G, Rugolo M, Romeo G. Defective Oxidative Phosphorylation In Thyroid Oncocytic Carcinoma Is Associated With Pathogenic Mitochondrial DNA Mutations Affecting Complexes I And III. Cancer Research. 2006;66:6087–96. [CrossRef]
- Bellance N, Benard G, Furt F, Begueret H, Smolková K, Passerieux E, Delage JP, Baste JM, Moreau P, Rossignol R. Bioenergetics Of Lung Tumors: Alteration Of Mitochondrial Biogenesis And Respiratory Capacity. International Journal of Biochemistry and Cell Biology. 2009;41:2566–77. [CrossRef]
- Liberti M V, Locasale JW. The Warburg Effect: How Does It Benefit Cancer Cells? Trends in Biochemical Sciences. 2016;41:287.
- Lee M, Yoon J. Metabolic Interplay Between Glycolysis And Mitochondrial Oxidation: The Reverse Warburg Effect And Its Therapeutic Implication. World Journal of Biological Chemistry. 2015;6:148–62. [CrossRef]
- Kim SY. Cancer Energy Metabolism: Shutting Power Off Cancer Factory. Biomolecules and Therapeutics. 2018;26:39–44. [CrossRef]
- Fu Y, Liu S, Yin S, Niu W, Xiong W. The Reverse Warburg Effect Is Likely To Be An Achilles ’ Heel Of Cancer That Can Be Exploited For Cancer Therapy 2017;8:57813–25. [CrossRef]
- Luis C, Duarte F, Faria I, Jarak I, Oliveira PF, Alves MG, Soares R, Fernandes R. Warburg Effect Inversion: Adiposity Shifts Central Primary Metabolism In MCF-7 Breast Cancer Cells. Life Sciences. 2019;223:38–46. [CrossRef]
- Slane BG, Aykin-Burns N, Smith BJ, Kalen AL, Goswami PC, Domann FE, Spitz DR. Mutation Of Succinate Dehydrogenase Subunit C Results In Increased O 2.-, Oxidative Stress, And Genomic Instability. Cancer Research. 2006;66:7615–20.
- Alam NA, Olpin S, Rowan A, Kelsell D, Leigh IM, Tomlinson IPM, Weaver T. Missense Mutations In Fumarate Hydratase In Multiple Cutaneous And Uterine Leiomyomatosis And Renal Cell Cancer. Journal of Molecular Diagnostics. 2005;7:437–43. [CrossRef]
- Gross S, Cairns RA, Minden MD, Driggers EM, Bittinger MA, Jang HG, Sasaki M, Jin S, Schenkein DP, Su SM, Dang L, Fantin VR, Mak TW. Cancer-Associated Metabolite 2-Hydroxyglutarate Accumulates In Acute Myelogenous Leukemia With Isocitrate Dehydrogenase 1 And 2 Mutations. Journal of Experimental Medicine. 2010;207:339–44. [CrossRef]
- Grassian AR et al. IDH1 Mutations Alter Citric Acid Cycle Metabolism And Increase Dependence On Oxidative Mitochondrial Metabolism. Cancer Research. 2014;74:3317–31.
- Chen JQ, Russo J. Dysregulation Of Glucose Transport, Glycolysis, TCA Cycle And Glutaminolysis By Oncogenes And Tumor Suppressors In Cancer Cells. Biochimica et Biophysica Acta - Reviews on Cancer. 2012;1826:370–84.
- Bourgeron T, Rustin P, Chretien D, Birch-machin M, Bourgeois M, Viegas-pequignot E, Munnich A, Rotig A. Mutation Of A Nuclear Succinate Dehydrogenase Gene Results In Mitochondrial Respiratory Chain Deficiency. Nature Genetics. 1995;11:144–9. [CrossRef]
- Böttcher M, Renner K, Berger R, Mentz K, Thomas S, Cardenas-Conejo ZE, Dettmer K, Oefner PJ, Mackensen A, Kreutz M, Mougiakakos D. D-2-Hydroxyglutarate Interferes With HIF-1α Stability Skewing T-Cell Metabolism Towards Oxidative Phosphorylation And Impairing Th17 Polarization. OncoImmunology. 2018;7:1–12. [CrossRef]
- Ferro F, Servais S, Besson P, Roger S, Dumas JF, Brisson L. Autophagy And Mitophagy In Cancer Metabolic Remodelling. Seminars in Cell and Developmental Biology. 2020;98:129–38. [CrossRef]
- Gonzalez CD, Alvarez S, Ropolo A, Rosenzvit C, Gonzalez Bagnes MF, Vaccaro MI. Autophagy, Warburg, And Warburg Reverse Effects In Human Cancer. BioMed Research International. 2014;2014. [CrossRef]
- Bhattacharya B, Mohd Omar MF, Soong R. The Warburg Effect And Drug Resistance. British Journal of Pharmacology. 2016;173:970–9. [CrossRef]
- Das CK, Mandal M, Kögel D. Pro-Survival Autophagy And Cancer Cell Resistance To Therapy. Cancer and Metastasis Reviews. 2018;37:749–66. [CrossRef]
- Poulogiannis G, McIntyre RE, Dimitriadi M, Apps JR, Wilson CH, Ichimura K, Luo F, Cantley LC, Wyllie AH, Adams DJ, Arends MJ. PARK2 Deletions Occur Frequently In Sporadic Colorectal Cancer And Accelerate Adenoma Development In Apc Mutant Mice. Proceedings of the National Academy of Sciences of the United States of America. 2010;107:15145–50.
- Veeriah S et al. Somatic Mutations Of The Parkinson’s Disease-Associated Gene PARK2 In Glioblastoma And Other Human Malignancies. Nature Genetics. 2010;42:77–82. [CrossRef]
- Hu HH et al. PARKIN Inactivation Links Parkinson’s Disease To Melanoma. Journal of the National Cancer Institute. 2016;108:1–8.
- Lee SB, She J, Deng B, Kim JJ, de Andrade M, Na J, Sun Z, Wampfler JA, Cunningham JM, Wu Y, Limper AH, Aubry MC, Wendt C, Biterman P, Yang P, Lou Z. Multiple-Level Validation Identifies PARK2 In The Development Of Lung Cancer And Chronic Obstructive Pulmonary Disease. Oncotarget. 2016;7:44211–23. [CrossRef]
- Letessier A, Garrido-Urbani S, Ginestier C, Fournier G, Esterni B, Monville F, Adélaïde J, Geneix J, Xerri L, Dubreuil P, Viens P, Charafe-Jauffret E, Jacquemier J, Birnbaum D, Lopez M, Chaffanet M. Correlated Break At PARK2/FRA6E And Loss Of AF-6/Afadin Protein Expression Are Associated With Poor Outcome In Breast Cancer. Oncogene. 2007;26:298–307.
- Sreedhar A, Petruska P, Miriyala S, Panchatcharam M, Zhao Y. UCP2 Overexpression Enhanced Glycolysis Via Activation Of PFKFB2 During Skin Cell Transformation. Oncotarget. 2017;8:95504–15. [CrossRef]
- Xu XD, Shao SX, Jiang HP, Cao YW, Wang YH, Yang XC, Wang YL, Wang XS, Niu HT. Warburg Effect Or Reverse Warburg Effect? A Review Of Cancer Metabolism. Oncology Research and Treatment. 2015;38:117–22. [CrossRef]
- Brandi J, Cecconi D, Cordani M, Torrens-Mas M, Pacchiana R, Dalla Pozza E, Butera G, Manfredi M, Marengo E, Oliver J, Roca P, Dando I, Donadelli M. The Antioxidant Uncoupling Protein 2 Stimulates HnRNPA2/B1, GLUT1 And PKM2 Expression And Sensitizes Pancreas Cancer Cells To Glycolysis Inhibition. Free Radical Biology and Medicine. 2016;101:305–16. [CrossRef]
- Pons DG, Nadal-Serrano M, Torrens-Mas M, Valle A, Oliver J, Roca P. UCP2 Inhibition Sensitizes Breast Cancer Cells To Therapeutic Agents By Increasing Oxidative Stress. Free Radical Biology and Medicine. 2015;86:67–77. [CrossRef]
- Mycielska ME, Broke-Smith TP, Palmer CP, Beckerman R, Nastos T, Erguler K, Djamgoz MBA. Citrate Enhances In Vitro Metastatic Behaviours Of PC-3M Human Prostate Cancer Cells: Status Of Endogenous Citrate And Dependence On Aconitase And Fatty Acid Synthase. International Journal of Biochemistry and Cell Biology. 2006;38:1766–77. [CrossRef]
- Arceci RJ. An Integrated Genomic Analysis Of Human Glioblastoma Multiforme. Yearbook of Oncology. 2009;2009:185–6. [CrossRef]
- Yan H, Parsons DW, Jin G, McLendon R, Rasheed BA, Yuan W, Kos I, Batinic-Haberle I, Jones S, Riggins GJ, Friedman H, Friedman A, Reardon D, Herndon J, Kinzler KW, Velculescu VE, Vogelstein B, Bigner DD. IDH1 And IDH2 Mutations In Gliomas. New England Journal of Medicine. 2009;360:765–73.
- Ward PS, Patel J, Wise DR, Abdel-Wahab O, Bennett BD, Coller HA, Cross JR, Fantin VR, Hedvat C V., Perl AE, Rabinowitz JD, Carroll M, Su SM, Sharp KA, Levine RL, Thompson CB. The Common Feature Of Leukemia-Associated IDH1 And IDH2 Mutations Is A Neomorphic Enzyme Activity Converting α-Ketoglutarate To 2-Hydroxyglutarate. Cancer Cell. 2010;17:225–34. [CrossRef]
- Kranendijk M et al. IDH2 Mutations In Patients With D-2-Hydroxyglutaric Aciduria. Science. 2010;330:7. [CrossRef]
- Dang L et al. Cancer-Associated IDH1 Mutations Produce 2-Hydroxyglutarate. Nature. 2009;462:739–44.
- Bardella C, Pollard PJ, Tomlinson I. SDH Mutations In Cancer. Biochimica et Biophysica Acta - Bioenergetics. 2011;1807:1432–43.
- Oyedotun KS, Lemire BD. The Quaternary Structure Of The Saccharomyces Cerevisiae Succinate Dehydrogenase: Homology Modeling, Cofactor Docking, And Molecular Dynamics Simulation Studies. Journal of Biological Chemistry. 2004;279:9424–31.
- Baysal BE. On The Association Of Succinate Dehydrogenase Mutations With Hereditary Paraganglioma. Trends in Endocrinology and Metabolism. 2003;14:453–9. [CrossRef]
- Neumann HPH, Pawlu C, Peczkowska M, Bausch B, McWhinney SR, Muresan M, Buchta M, Franke G, Klisch J, Bley TA, Hoegerle S, Boedeker CC, Opocher G, Schipper J, Januszewicz A, Eng C. Distict Clinical Features Of Paraganglioma Syndromes Associated With SDHB And SDHD Gene Mutations. The Journal of the American Medical Association. 2004;292:943–52.
- Pollard PJ, Wortham NC, Tomlinson IPM. The TCA Cycle And Tumorigenesis: The Examples Of Fumarate Hydratase And Succinate Dehydrogenase. Annals of Medicine. 2003;35:634–5. [CrossRef]
- Habano W, Sugai T, Nakamura SI, Uesugi N, Higuchi T, Terashima M, Horiuchi S. Reduced Expression And Loss Of Heterozygosity Of The SDHD Gene In Colorectal And Gastric Cancer. Oncology Reports. 2003;10:1375–80. [CrossRef]
- Gimenez-Roqueplo AP, Favier J, Rustin P, Mourad JJ, Plouin PF, Corvol P, Rötig A, Jeunemaitre X. The R22X Mutation Of The SDHD Gene In Hereditary Paraganglioma Abolishes The Enzymatic Activity Of Complex II In The Mitochondrial Respiratory Chain And Activates The Hypoxia Pathway. American Journal of Human Genetics. 2001;69:1186–97. [CrossRef]
- Gimenez-Roqueplo AP, Favier J, Rustin P, Rieubland C, Crespin M, Nau V, Van Kien PK, Corvol P, Plouin PF, Jeunemaitre X. Mutations In The SDHB Gene Are Associated With Extra-Adrenal And/Or Malignant Phaeochromocytomas. Cancer Research. 2003;63:5615–21.
- Selak MA, Armour SM, MacKenzie ED, Boulahbel H, Watson DG, Mansfield KD, Pan Y, Simon MC, Thompson CB, Gottlieb E. Succinate Links TCA Cycle Dysfunction To Oncogenesis By Inhibiting HIF-α Prolyl Hydroxylase. Cancer Cell. 2005;7:77–85. [CrossRef]
- Jaramillo MC, Zhang DD. The Emerging Role Of The Nrf2-Keap1 Signaling Pathway In Cancer. Genes and Development. 2013;27:2179–91. [CrossRef]
- Tomlinson IPM et al. Germline Mutations In FH Predispose To Dominantly Inherited Uterine Fibroids, Skin Leiomyomata And Papillary Renal Cell Cancer The Multiple Leiomyoma Consortium. Nature Genetics. 2002;30:406–10.
- Martinez-Mir A, Glaser B, Chuang GS, Horev L, Waldman A, Engler DE, Gordon D, Spelman LJ, Hatzibougias I, Green J, Christiano AM, Zlotogorski A. Germline Fumarate Hydratase Mutations In Families With Multiple Cutaneous And Uterine Leiomyomata. Journal of Investigative Dermatology. 2003;121:741–4. [CrossRef]
- Bardella C et al. Aberrant Succination Of Proteins In Fumarate Hydratase-Deficient Mice And HLRCC Patients Is A Robust Biomarker Of Mutation Status. Journal of Pathology. 2011;225:4–11. [CrossRef]
- Ooi A, Furge KA. Fumarate Hydratase Inactivation In Renal Tumors: HIF1α, NRF2 And “Cryptic Targets” Of Transcription Factors. Chinese Journal of Cancer. 2012;31:413–20.
- Goetzman ES, Prochownik E V. The Role For Myc In Coordinating Glycolysis, Oxidative Phosphorylation, Glutaminolysis, And Fatty Acid Metabolism In Normal And Neoplastic Tissues. Frontiers in Endocrinology. 2018;9. [CrossRef]
- Levine AJ, Puzio-Kuter AM. The Control Of The Metabolic Switch In Cancers By Oncogenes And Tumor Suppressor Genes. Science. 2010;330:1340–4. [CrossRef]
- Li F, Wang Y, Zeller KI, Potter JJ, Wonsey DR, O’Donnell KA, Kim J -w., Yustein JT, Lee LA, Dang C V. Myc Stimulates Nuclearly Encoded Mitochondrial Genes And Mitochondrial Biogenesis. Molecular and Cellular Biology. 2005;25:6225–34.
- Song HP, Zhang L, Dang YM, Yan H, Chu ZG, Huang YS. The Phosphatidylinositol 3-Kinase-Akt Pathway Protects Cardiomyocytes From Ischaemic And Hypoxic Apoptosis Via Mitochondrial Function. Clinical and Experimental Pharmacology and Physiology. 2010;37:598–604. [CrossRef]
- Goo CK, Lim HY, Ho QS, Too HP, Clement MV, Wong KP. PTEN/Akt Signaling Controls Mitochondrial Respiratory Capacity Through 4E-BP1. PLoS ONE. 2012;7:1–12. [CrossRef]
- Lebedeva MA, Eaton JS, Shadel GS. Loss Of P53 Causes Mitochondrial DNA Depletion And Altered Mitochondrial Reactive Oxygen Species Homeostasis. Biochimica et Biophysica Acta - Bioenergetics. 2009;1787:328–34. [CrossRef]
- Matoba S, Kang JG, Patino WD, Wragg A, Boehm M, Gavrilova O, Hurley PJ, Bunz F, Hwang PM. P53 Regulates Mitochondrial Respiration. Science. 2006;312:1650–3. [CrossRef]
- Goan YG, Wu WT, Liu CI, Neoh CA, Wu YJ. Involvement Of Mitochondrial Dysfunction, Endoplasmic Reticulum Stress, And The PI3K/AKT/MTOR Pathway In Nobiletin-Induced Apoptosis Of Human Bladder Cancer Cells. Molecules. 2019;24. [CrossRef]
- Luo J, Manning BD, Cantley LC. Targeting The PI3K-Akt Pathway In Human Cancer: Rationale And Promise. Cancer Cell. 2003;4:257–62. [CrossRef]
- Rubio-Patiño C, Trotta AP, Chipuk JE. MDM2 And Mitochondrial Function: One Complex Intersection. Biochemical Pharmacology. 2019;162:14–20. [CrossRef]
- Haq R, Shoag J, Andreu-Perez P, Yokoyama S, Edelman H, Rowe GC, Frederick DT, Hurley AD, Nellore A, Kung AL, Wargo JA, Song JS, Fisher DE, Arany Z, Widlund HR. Oncogenic BRAF Regulates Oxidative Metabolism Via PGC1α And MITF. Cancer Cell. 2013;23:302–15. [CrossRef]
- Chun SY, Johnson C, Washburn JG, Cruz-Correa MR, Dang DT, Dang LH. Oncogenic KRAS Modulates Mitochondrial Metabolism In Human Colon Cancer Cells By Inducing HIF-1α And HIF-2α Target Genes. Molecular Cancer. 2010;9:1–11. [CrossRef]
- Pylayeva-Gupta Y, Grabocka E, Bar-Sagi D. RAS Oncogenes: Weaving A Tumorigenic Web. Nature Reviews Cancer. 2011;11:761–74. [CrossRef]
- Mukhopadhyay S, Vander Heiden MG, McCormick F. The Metabolic Landscape Of RAS-Driven Cancers From Biology To Therapy. Nature Cancer. 2021;2:271–83. [CrossRef]
- Moss DY, McCann C, Kerr EM. Rerouting The Drug Response: Overcoming Metabolic Adaptation In KRAS-Mutant Cancers. Science Signaling. 2022;15:eabj3490. [CrossRef]
- Holmström KM, Kostov R V., Dinkova-Kostova AT. The Multifaceted Role Of Nrf2 In Mitochondrial Function. Current Opinion in Toxicology. 2016;2:80–91. [CrossRef]
- Nisr RB, Shah DS, Ganley IG, Hundal HS. Proinflammatory NFkB Signalling Promotes Mitochondrial Dysfunction In Skeletal Muscle In Response To Cellular Fuel Overloading. Cellular and Molecular Life Sciences. 2019;76:4887–904. [CrossRef]
- Demory ML, Boerner JL, Davidson R, Faust W, Miyake T, Lee I, Hüttemann M, Douglas R, Haddad G, Parsons SJ. Epidermal Growth Factor Receptor Translocation To The Mitochondria. Journal of Biological Chemistry. 2009;284:36592–604. [CrossRef]
- Logan S, Pharaoh GA, Marlin MC, Masser DR, Matsuzaki S, Wronowski B, Yeganeh A, Parks EE, Premkumar P, Farley JA, Owen DB, Humphries KM, Kinter M, Freeman WM, Szweda LI, Van Remmen H, Sonntag WE. Insulin-Like Growth Factor Receptor Signaling Regulates Working Memory, Mitochondrial Metabolism, And Amyloid-β Uptake In Astrocytes. Molecular Metabolism. 2018;9:141–55. [CrossRef]
- Grazette LP, Boecker W, Matsui T, Semigran M, Force TL, Hajjar RJ, Rosenzweig A. Inhibition Of ErbB2 Causes Mitochondrial Dysfunction In Cardiomyocytes: Implications For Herceptin-Induced Cardiomyopathy. Journal of the American College of Cardiology. 2004;44:2231–8. [CrossRef]
- Bui T, Thompson CB. Cancer’s Sweet Tooth. Cancer Cell. 2006;9:419–20.
- Kim JW, Tchernyshyov I, Semenza GL, Dang C V. HIF-1-Mediated Expression Of Pyruvate Dehydrogenase Kinase: A Metabolic Switch Required For Cellular Adaptation To Hypoxia. Cell Metabolism. 2006;3:177–85. [CrossRef]
- Papandreou I, Cairns RA, Fontana L, Lim AL, Denko NC. HIF-1 Mediates Adaptation To Hypoxia By Actively Downregulating Mitochondrial Oxygen Consumption. Cell Metabolism. 2006;3:187–97. [CrossRef]
- Wasylenko TM, Ahn WS, Stephanopoulos G. The Oxidative Pentose Phosphate Pathway Is The Primary Source Of NADPH For Lipid Overproduction From Glucose In Yarrowia Lipolytica. Metabolic Engineering. 2015;30:27–39. [CrossRef]
- De Preter G, Neveu MA, Danhier P, Brisson L, Payen VL, Porporato PE, Jordan BF, Sonveaux P, Gallez B. Inhibition Of The Pentose Phosphate Pathway By Dichloroacetate Unravels A Missing Link Between Aerobic Glycolysis And Cancer Cell Proliferation. Oncotarget. 2016;7:2910–20. [CrossRef]
- DeBerardinis RJ, Mancuso A, Daikhin E, Nissim I, Yudkoff M, Wehrli S, Thompson CB. Beyond Aerobic Glycolysis: Transformed Cells Can Engage In Glutamine Metabolism That Exceeds The Requirement For Protein And Nucleotide Synthesis. Proceedings of the National Academy of Sciences of the United States of America. 2007;104:19345–50. [CrossRef]
- Locasale JW et al. Phosphoglycerate Dehydrogenase Diverts Glycolytic Flux And Contributes To Oncogenesis. Nature Genetics. 2011;43:869–74. [CrossRef]
- Luengo A, Li Z, Gui DY, Sullivan LB, Zagorulya M, Spranger S, Matheson NJ, Heiden MG Vander. Increased Demand For NAD+ Relative To ATP Drives Aerobic Glycolysis. BioRxiv. 2020:1–58. [CrossRef]
- White KA, Grillo-Hill BK, Barber DL. Cancer Cell Behaviors Mediated By Dysregulated PH Dynamics At A Glance. Journal of Cell Science. 2017;130:663–9. [CrossRef]
- Luengo A, Li Z, Gui DY, Sullivan LB, Zagorulya M, Do BT, Ferreira R, Naamati A, Ali A, Lewis CA, Thomas CJ, Spranger S, Matheson NJ, Vander Heiden MG. Increased Demand For NAD+ Relative To ATP Drives Aerobic Glycolysis. Molecular Cell. 2020:1–17. [CrossRef]
- Heiden MGV the warburg effect: T metabolic requirements of cell proliferation, Cantley LC, Thompson CB. Understanding The Warburg Effect: The Metabolic Requirements Of Cell Proliferation. Science. 2009;324:1029–33. [CrossRef]
- Kondoh H, Lleonart ME, Nakashima Y, Yokode M, Tanaka M, Bernard D, Gil J, Beach D. A High Glycolytic Flux Supports The Proliferative Potential Of Murine Embryonic Stem Cells. Antioxidants & Redox Signaling. 2006;0:061221112325011.
- Ciavardelli D, Rossi C, Barcaroli D, Volpe S, Consalvo A, Zucchelli M, De Cola A, Scavo E, Carollo R, D’Agostino D, Forlì F, D’Aguanno S, Todaro M, Stassi G, Di Ilio C, De Laurenzi V, Urbani A. Breast Cancer Stem Cells Rely On Fermentative Glycolysis And Are Sensitive To 2-Deoxyglucose Treatment. Cell Death and Disease. 2014;5:1–12. [CrossRef]
- Ravera S, Podestà M, Sabatini F, Fresia C, Columbaro M, Bruno S, Fulcheri E, Ramenghi LA, Frassoni F. Mesenchymal Stem Cells From Preterm To Term Newborns Undergo A Significant Switch From Anaerobic Glycolysis To The Oxidative Phosphorylation. Cellular and Molecular Life Sciences. 2018;75:889–903. [CrossRef]
- Da Veiga Moreira J, Hamraz M, Abolhassani M, Bigan E, Pérès S, Paulevé L, Nogueira ML, Steyaert JM, Schwartz L. The Redox Status Of Cancer Cells Supports Mechanisms Behind The Warburg Effect. Metabolites. 2016;6:1–12. [CrossRef]
- Pouysségur J, Franchi A, L’Allemain G, Paris S. Cytoplasmic PH, A Key Determinant Of Growth Factor-Induced DNA Synthesis In Quiescent Fibroblasts. FEBS Letters. 1985;190:115–9.
- Aerts RJ, Durston AJ, Moolenaar WH. Cytoplasmic PH And The Regulation Of The Dictyostelium Cell Cycle. Cell. 1985;43:653–7. [CrossRef]
- Birkeland ES, Koch LM, Dechant R. Another Consequence Of The Warburg Effect? Metabolic Regulation Of Na+/H+ Exchangers May Link Aerobic Glycolysis To Cell Growth. Frontiers in Oncology. 2020;10:1–9.
- Calderón Montaño J, Burgos Morón E, Pérez Guerrero M, Salvador Bofill F, Robles Frías A, López Lázaro M. Role Of The Intracellular PH In The Metabolic Switch Between Oxidative Phosphorylation And AerobicGlycolysis - Relevance To Cancer 2011:1–10.
- Icard P, Shulman S, Farhat D, Steyaert JM, Alifano M, Lincet H. How The Warburg Effect Supports Aggressiveness And Drug Resistance Of Cancer Cells? Drug Resistance Updates. 2018;38:1–11.
- Alreshidi MM, Dunstan RH, Gottfries J, Macdonald MM, Crompton MJ, Ang CS, Williamson NA, Roberts TK. Changes In The Cytoplasmic Composition Of Amino Acids And Proteins Observed In Staphylococcus Aureus During Growth Under Variable Growth Conditions Representative Of The Human Wound Site. PLoS ONE. 2016;11:1–19.
- Reshkin SJ, Bellizzi A, Caldeira S, Albarani V, Malanchi I, Poignee M, Alunni-Fabbroni M, Casavola V, Tommasino M. Na + /H + Exchanger-dependent Intracellular Alkalinization Is An Early Event In Malignant Transformation And Plays An Essential Role In The Development Of Subsequent Transformation-associated Phenotypes. The FASEB Journal. 2000;14:2185–97.
- Riddle SR, Ahmad A, Ahmad S, Deeb SS, Malkki M, Schneider BK, Allen CB, White CW. Hypoxia Induces Hexokinase II Gene Expression Inhuman Lung Cell Line A549. American Journal of Physiology - Lung Cellular and Molecular Physiology. 2000;278:L407–16.
- Bergeron M, Yu AY, Solway KE, Semenza GL, Sharp FR. Induction Of Hypoxia-Inducible Factor-1 (HIF-1) And Its Target Genes Following Focal Ischaemia In Rat Brain. European Journal of Neuroscience. 1999;11:4159–70.
- Cota-Ruiz K, Leyva-Carrillo L, Peregrino-Uriarte AB, Valenzuela-Soto EM, Gollas-Galván T, Gómez-Jiménez S, Hernández J, Yepiz-Plascencia G. Role Of HIF-1 On Phosphofructokinase And Fructose 1, 6-Bisphosphatase Expression During Hypoxia In The White Shrimp Litopenaeus Vannamei. Comparative Biochemistry and Physiology -Part A : Molecular and Integrative Physiology. 2016;198:1–7.
- Pelletier J, Bellot G, Gounon P, Lacas-Gervais S, Pouysségur J, Mazure NM. Glycogen Synthesis Is Induced In Hypoxia By The Hypoxia-Inducible Factor And Promotes Cancer Cell Survival. Frontiers in Oncology. 2012;2:1–9. [CrossRef]
- Semenza GL, Jiang BH, Leung SW, Passantino R, Concordat JP, Maire P, Giallongo A. Hypoxia Response Elements In The Aldolase A, Enolase 1, And Lactate Dehydrogenase A Gene Promoters Contain Essential Binding Sites For Hypoxia-Inducible Factor 1. Journal of Biological Chemistry. 1996;271:32529–37. [CrossRef]
- Wigfield SM, Winter SC, Giatromanolaki A, Taylor J, Koukourakis ML, Harris AL. PDK-1 Regulates Lactate Production In Hypoxia And Is Associated With Poor Prognosis In Head And Neck Squamous Cancer. British Journal of Cancer. 2008;98:1975–84. [CrossRef]
- Rosafio K, Pellerin L. Oxygen Tension Controls The Expression Of The Monocarboxylate Transporter MCT4 In Cultured Mouse Cortical Astrocytes Via A Hypoxia-Inducible Factor-1α-Mediated Transcriptional Regulation. Glia. 2014;62:477–90. [CrossRef]
- McClelland GB, Brooks GA. Changes In MCT 1, MCT 4, And LDH Expression Are Tissue Specific In Rats After Long-Term Hypobaric Hypoxia. Journal of Applied Physiology. 2002;92:1573–84. [CrossRef]
- Ullah MS, Davies AJ, Halestrap AP. The Plasma Membrane Lactate Transporter MCT4, But Not MCT1, Is Up-Regulated By Hypoxia Through A HIF-1α-Dependent Mechanism. Journal of Biological Chemistry. 2006;281:9030–7.
- Baumann MU, Zamudio S, Illsley NP. Hypoxic Upregulation Of Glucose Transporters In BeWo Choriocarcinoma Cells Is Mediated By Hypoxia-Inducible Factor-1. American Journal of Physiology - Cell Physiology. 2007;293:477–85. [CrossRef]
- Hervouet E, Cízková A, Demont J, Vojtísková A, Pecina P, Franssen-van Hal NLW, Keijer J, Simonnet H, Ivének R, Kmoch S, Godinot C, Houstek J. HIF And Reactive Oxygen Species Regulate Oxidative Phosphorylation In Cancer. Carcinogenesis. 2008;29:1528–37. [CrossRef]
- Hervouet E, Demont J, Pecina P, Vojtísková A, Houstek J, Simonnet H, Godinot C. A New Role For The Von Hippel-Lindau Tumor Suppressor Protein: Stimulation Of Mitochondrial Oxidative Phosphorylation Complex Biogenesis. Carcinogenesis. 2005;26:531–9. [CrossRef]
- Fukuda R, Zhang H, Kim J whan, Shimoda L, Dang C V., Semenza GLL. HIF-1 Regulates Cytochrome Oxidase Subunits To Optimize Efficiency Of Respiration In Hypoxic Cells. Cell. 2007;129:111–22. [CrossRef]
- Kang JH, Lee SH, Lee JS, Nam B, Seong TW, Son J, Jang H, Hong KM, Lee C, Kim SY. Aldehyde Dehydrogenase Inhibition Combined With Phenformin Treatment Reversed NSCLC Through ATP Depletion. Oncotarget. 2016;7:49397–410. [CrossRef]
- Kang JH, Lee SH, Hong D, Lee JS, Ahn HS, Ahn JH, Seong TW, Lee CH, Jang H, Hong KM, Lee C, Lee JH, Kim SY. Aldehyde Dehydrogenase Is Used By Cancer Cells For Energy Metabolism. Experimental & Molecular Medicine. 2016;48:e272. [CrossRef]
- Hong SM, Hwang SW, Wang T, Park CW, Ryu YM, Jung JH, Shin JH, Kim SY, Lee JL, Kim CW, Yoon G, Kim KH, Myung SJ, Choi KY. Increased Nicotinamide Adenine Dinucleotide Pool Promotes Colon Cancer Progression By Suppressing Reactive Oxygen Species Level. Cancer Science. 2019;110:629–38. [CrossRef]
- Rempel A, Mathupala SP, Griffin CA, Hawkins AL, Pedersen PL. Glucose Catabolism In Cancer Cells: Amplification Of The Gene Encoding Type II Hexokinase. Cancer Research. 1996;56:2468–71.
- Chen Z, Zhang H, Lu W, Huang P. Role Of Mitochondria-Associated Hexokinase II In Cancer Cell Death Induced By 3-Bromopyruvate. Biochimica et Biophysica Acta - Bioenergetics. 2009;1787:553–60. [CrossRef]
- Anderson M, Marayati R, Moffitt R, Yeh JJ. Hexokinase 2 Promotes Tumor Growth And Metastasis By Regulating Lactate Production In Pancreatic Cancer. Oncotarget. 2017;8:56081–94. [CrossRef]
- Yi W, Clark PM, Mason DE, Keenan MC, Hill C, William A, Iii G, Peters EC, Driggers EM, Hsieh-wilson LC. PFK1 Glycosylation Is A Key Regulator Of Cancer Cell Growth And Central Metabolic Pathways Wen. Science. 2013;337:975–80.
- Zancan P, Sola-Penna M, Furtado CM, Da Silva D. Differential Expression Of Phosphofructokinase-1 Isoforms Correlates With The Glycolytic Efficiency Of Breast Cancer Cells. Molecular Genetics and Metabolism. 2010;100:372–8. [CrossRef]
- Sun CM, Xiong DB, Yan Y, Geng J, Liu M, Yao XD. Genetic Alteration In Phosphofructokinase Family Promotes Growth Of Muscle-Invasive Bladder Cancer. International Journal of Biological Markers. 2016;31:e286–93. [CrossRef]
- Mazurek S, Boschek CB, Hugo F, Eigenbrodt E. Pyruvate Kinase Type M2 And Its Role In Tumor Growth And Spreading. Seminars in Cancer Biology. 2005;15:300–8. [CrossRef]
- Prakasam G, Iqbal MA, Bamezai RNK, Mazurek S. Posttranslational Modifications Of Pyruvate Kinase M2: Tweaks That Benefit Cancer. Frontiers in Oncology. 2018;8:1–12. [CrossRef]
- Zahra K, Dey T, Ashish, Mishra SP, Pandey U. Pyruvate Kinase M2 And Cancer: The Role Of PKM2 In Promoting Tumorigenesis. Frontiers in Oncology. 2020;10:1–9. [CrossRef]
- Shiroki T, Yokoyama M, Tanuma N, Maejima R, Tamai K, Yamaguchi K, Oikawa T, Noguchi T, Miura K, Fujiya T, Shima H, Sato I, Murata-Kamiya N, Hatakeyama M, Iijima K, Shimosegawa T, Satoh K. Enhanced Expression Of The M2 Isoform Of Pyruvate Kinase Is Involved In Gastric Cancer Development By Regulating Cancer-Specific Metabolism. Cancer Science. 2017;108:931–40. [CrossRef]
- Kolobova E, Tuganova A, Boulatnikov I, Popov KM. Regulation Of Pyruvate Dehydrogenase Activity Through Phosphorylation At Multiple Sites. Biochemistry. 2001;77:69–77.
- Dai ZJ, Pan S, Chen C, Cao L, Li X, Chen X, Su X, Lin S. Down-Regulation Of Succinate Dehydrogenase Subunit B And Up-Regulation Of Pyruvate Dehydrogenase Kinase 1 Predicts Poor Prognosis In Recurrent Nasopharyngeal Carcinoma. Tumor Biology. 2016;37:5145–52. [CrossRef]
- Wang JJ, Siu MK, Jiang YX, Leung TH, Chan DW, Cheng RR, Cheung AN, Ngan HY, Chan KK. Aberrant Upregulation Of PDK1 In Ovarian Cancer Cells Impairs CD8+ T Cell Function And Survival Through Elevation Of PD-L1. OncoImmunology. 2019;8. [CrossRef]
- Herzig S, Shaw RJ. AMPK: Guardian Of Metabolism And Mitochondrial Homeostasis. Nature Reviews Molecular Cell Biology. 2018;19:121–35. [CrossRef]
- Hong SP, Leiper FC, Woods A, Carling D, Carlson M. Activation Of Yeast Snf1 And Mammalian AMP-Activated Protein Kinase By Upstream Kinases. Proceedings of the National Academy of Sciences of the United States of America. 2003;100:8839–43. [CrossRef]
- Shaw RJ, Kosmatka M, Bardeesy N, Hurley RL, Witters LA, DePinho RA, Cantley LC. The Tumor Suppressor LKB1 Kinase Directly Activates AMP-Activated Kinase And Regulates Apoptosis In Response To Energy Stress. Proceedings of the National Academy of Sciences of the United States of America. 2004;101:3329–35. [CrossRef]
- Xiao B, Sanders MJ, Underwood E, Heath R, Mayer F V., Carmena D, Jing C, Walker PA, Eccleston JF, Haire LF, Saiu P, Howell SA, Aasland R, Martin SR, Carling D, Gamblin SJ. Structure Of Mammalian AMPK And Its Regulation By ADP. Nature. 2011;472:230–3. [CrossRef]
- Liu Q, Gauthier M, Sun L, Ruderman N, Lodish H. Activation Of AMP-activated Protein Kinase Signaling Pathway By Adiponectin And Insulin In Mouse Adipocytes: Requirement Of Acyl-CoA Synthetases FATP1 And Acsl1 And Association With An Elevation In AMP/ATP Ratio. The FASEB Journal. 2010;24:4229–39. [CrossRef]
- Hardie DG. Molecular Pathways: Is AMPK A Friend Or A Foe In Cancer? Clinical Cancer Research. 2015;21:3836–40.
- Li N, Huang D, Lu N, Luo L. Role Of The LKB1/AMPK Pathway In Tumor Invasion And Metastasis Of Cancer Cells (Review). Oncology Reports. 2015;34:2821–6.
- Yung MMH, Chan DW, Liu VWS, Yao KM, Ngan HYS. Activation Of AMPK Inhibits Cervical Cancer Cell Growth Through AKT/FOXO3a/FOXM1 Signaling Cascade. BMC Cancer. 2013;13. [CrossRef]
- Zhou J, Huang W, Tao R, Ibaragi S, Lan F, Ido Y, Wu X, Alekseyev YO, Lenburg ME, Hu GF, Luo Z. Inactivation Of AMPK Alters Gene Expression And Promotes Growth Of Prostate Cancer Cells. Oncogene. 2009;28:1993–2002. [CrossRef]
- Mack HID, Zheng B, Asara JM, Thomas SM. AMPK-Dependent Phosphorylation Of ULK1 Regulates ATG9 Localization. Autophagy. 2012;8:1197–214. [CrossRef]
- Petherick KJ, Conway OJL, Mpamhanga C, Osborne SA, Kamal A, Saxty B, Ganley IG. Pharmacological Inhibition Of ULK1 Kinase Blocks Mammalian Target Of Rapamycin (MTOR)-Dependent Autophagy. Journal of Biological Chemistry. 2015;290:11376–83.
- Nardo A Di, Wertz MH, Kwiatkowski E, Tsai PT, Leech JD, Greene-Colozzi E, Goto J, Dilsiz P, Talos DM, Clish CB, Kwiatkowski DJ, Sahin M. Neuronal Tsc1/2 Complex Controls Autophagy Through AMPK-Dependent Regulation Of ULK1. Human Molecular Genetics. 2014;23:1–10.
- Faubert B, Boily G, Izreig S, Griss T, Samborska B, Dong Z, Dupuy F, Chambers C, Fuerth BJ, Viollet B, Mamer OA, Avizonis D, Deberardinis RJ, Siegel PM, Jones RG. AMPK Is A Negative Regulator Of The Warburg Effect And Suppresses Tumor Growth In Vivo. Cell Metabolism. 2013;17:113–24. [CrossRef]
- Inoki K, Zhu T, Guan K-LG. TSC2 Mediates Cellular Energy Response To Control Cell Growth And Survival. Cell Press. 2003;115:577–90. [CrossRef]
- Inoki K, Li Y, Zhu T, Wu J, Guan KL. TSC2 Is Phosphorylated And Inhibited By Akt And Suppresses MTOR Signalling. Nature Cell Biology. 2002;4:648–57. [CrossRef]
- Inoki K, Li Y, Xu T, Guan KL. Rheb GTpase Is A Direct Target Of TSC2 GAP Activity And Regulates MTOR Signaling. Genes and Development. 2003;17:1829–34. [CrossRef]
- Sun Q et al. Mammalian Target Of Rapamycin Up-Regulation Of Pyruvate Kinase Isoenzyme Type M2 Is Critical For Aerobic Glycolysis And Tumor Growth. Proceedings of the National Academy of Sciences of the United States of America. 2011;108:4129–34.
- Ling NXY, Kaczmarek A, Hoque A, Davie E, Ngoei KRW, Morrison KR, Smiles WJ, Forte GM, Wang T, Lie S, Dite TA, Langendorf CG, Scott JW, Oakhill JS, Petersen J. MTORC1 Directly Inhibits AMPK To Promote Cell Proliferation Under Nutrient Stress. Nature Metabolism. 2020;2:41–9. [CrossRef]
- Guo X, Li H, Xu H, Woo S, Dong H, Lu F, Lange AJ, Wu C. Glycolysis In The Control Of Blood Glucose Homeostasis. Acta Pharmaceutica Sinica B. 2012;2:358–67. [CrossRef]
- Praiss M, Cerasi E. Immediate And Time-Dependent Effects Of Gluoce An Insulin Release: Differential Calcium Requirements. Acta Endocrinologica. 1988;117:409–16.
- Nesher R, Praiss M, Cerasi E. Immediate And Time-Dependent Effects Of Glucose On Insulin Release: Differential Calcium Requirements. Acta Endocrinologica. 1988;117:409–16. [CrossRef]
- Pottout V, Robertson RP. Secondary B-Cell Failure In Type 2 Diabetes-A Convergence Of Glucotoxicity And Lipotoxicity. Endocrinology. 2002;143:339–42.
- Rossetti L, Shulman GI, Zawalich W, DeFronzo RA. Effect Of Chronic Hyperglycemia On In Vivo Insulin Secretion In Partially Pancreatectomized Rats. Journal of Clinical Investigation. 1987;80:1037–44. [CrossRef]
- Gerber PA, Rutter GA. The Role Of Oxidative Stress And Hypoxia In Pancreatic Beta-Cell Dysfunction In Diabetes Mellitus. Antioxidants and Redox Signaling. 2017;26:501–18.
- Noguchi R, Kubota H, Yugi K, Toyoshima Y, Komori Y, Soga T, Kuroda S. The Selective Control Of Glycolysis, Gluconeogenesis And Glycogenesis By Temporal Insulin Patterns. Molecular Systems Biology. 2013;9:1–12. [CrossRef]
- Bitar MS, Al-Mulla F. ROS Constitute A Convergence Nexus In The Development Of IGF1 Resistance And Impaired Wound Healing In A Rat Model Of Type 2 Diabetes. DMM Disease Models and Mechanisms. 2012;5:375–88. [CrossRef]
- Al-Lahham R, Deford JH, Papaconstantinou J. Mitochondrial-Generated ROS Down Regulates Insulin Signaling Via Activation Of The P38MAPK Stress Response Pathway. Molecular and Cellular Endocrinology. 2016;419:1–11. [CrossRef]
- Ogihara T, Asano T, Katagiri H, Sakoda H, Anai M, Shojima N, Ono H, Fujishiro M, Kushiyama A, Fujita Y, M. F, Kikuchi N, Noguchi I, Aburatani H, Gotoh Y, Komuro T. Oxidative Stress Induces Insulin Resistance By Activating The Nuclear Factor-Kappa B Pathway And Disrupting Normal Subcellular Distribution Of Phosphatidylinositol 3-Kinase. Diabetologia. 2004;47:794–805.
- Newsholme P, Haber EP, Hirabara SM, Rebelato ELO, Procopio J, Morgan D, Oliveira-Emilio HC, Carpinelli AR, Curi R. Diabetes Associated Cell Stress And Dysfunction: Role Of Mitochondrial And Non-Mitochondrial ROS Production And Activity. Journal of Physiology. 2007;583:9–24. [CrossRef]
- Hoeks J, Van Herpen NA, Mensink M, Moonen-Kornips E, Van Beurden D, Hesselink MKC, Schrauwen P. Prolonged Fasting Identifies Skeletal Muscle Mitochondrial Dysfunction As Consequence Rather Than Cause Of Human Insulin Resistance. Diabetes. 2010;59:2117–25. [CrossRef]
- Bonnard C, Durand A, Peyrol S, Chanseaume E, Chauvin MA, Morio B, Vidal H, Rieusset J. Mitochondrial Dysfunction Results From Oxidative Stress In The Skeletal Muscle Of Diet-Induced Insulin-Resistant Mice. Journal of Clinical Investigation. 2008;118:789–800. [CrossRef]
- Mootha VK et al. PGC-1α-Responsive Genes Involved In Oxidative Phosphorylation Are Coordinately Downregulated In Human Diabetes. Nature Genetics. 2003;34:267–73. [CrossRef]
- Asmann YW, Stump CS, Short KR, Coenen-Schimke JM, Guo ZK, Bigelow ML, Nair KS. Skeletal Muscle Mitochondrial Functions, Mitochondrial DNA Copy Numbers, And Gene Transcript Profiles In Type 2 Diabetic And Nondiabetic Subjects At Equal Levels Of Low Or High Insulin And Euglycemia. Diabetes. 2006;55:3309–19. [CrossRef]
- Sparks LM, Xie H, Koza RA, Mynatt R, Hulver MW, Bray GA, Smith SR. A High-Fat Diet Coordinately Downregulates Genes Required For Mitochondrial Oxidative Phosphorylation In Skeletal Muscle. Diabetes. 2005;54:1926–33. [CrossRef]
- Mogensen M, Sahlin K, Fernström M, Glintborg D, Vind BF, Beck-Nielsen H, Højlund K. Mitochondrial Respiration Is Decreased In Skeletal Muscle Of Patients With Type 2 Diabetes. Diabetes. 2007;56:1592–9. [CrossRef]
- Poussin C, Ibberson M, Hall D, Ding J, Soto J, Abel ED, Thorens B. Oxidative Phosphorylation Flexibility In The Liver Of Mice Resistant To High-Fat Diet-Induced Hepatic Steatosis. Diabetes. 2011;60:2216–24. [CrossRef]
- Pospisilik JA, Knauf C, Joza N, Benit P, Orthofer M, Cani PD, Ebersberger I, Nakashima T, Sarao R, Neely G, Esterbauer H, Kozlov A, Kahn CR, Kroemer G, Rustin P, Burcelin R, Penninger JM. Targeted Deletion Of AIF Decreases Mitochondrial Oxidative Phosphorylation And Protects From Obesity And Diabetes. Cell. 2007. [CrossRef]
- Roden M. Muscle Triglycerides And Mitochondrial Function: Possible Mechanisms For The Development Of Type 2 Diabetes. International Journal of Obesity. 2005;29:S111–5. [CrossRef]
- Fazakerley DJ, Minard AY, Krycer JR, Thomas KC, Stöckli J, Harney DJ, Burchfield JG, Maghzal GJ, Caldwell ST, Hartley RC, Stocker R, Murphy MP, James DE. Mitochondrial Oxidative Stress Causes Insulin Resistance Without Disrupting Oxidative Phosphorylation. Journal of Biological Chemistry. 2018;293:7315–28.
- Lewis MT, Kasper JD, Bazil JN, Frisbee JC, Wiseman RW. Quantification Of Mitochondrial Oxidative Phosphorylation In Metabolic Disease: Application To Type 2 Diabetes. International Journal of Molecular Sciences. 2019;20. [CrossRef]
- El-Najjar N, Chatila M, Moukadem H, Vuorela H, Ocker M, Gandesiri M, Schneider-Stock R, Gali-Muhtasib H. Reactive Oxygen Species Mediate Thymoquinone-Induced Apoptosis And Activate ERK And JNK Signaling. Apoptosis. 2010;15:183–95. [CrossRef]
- Schreck R, Rieber P, Baeuerle PA. Reactive Oxygen Intermediates As Apparently Widely Used Messengers In The Activation Of The NF-ΚB Transcription Factor And HIV-1. EMBO Journal. 1991;10:2247–58.
- Gloire G, Legrand-Poels S, Piette J. NF-ΚB Activation By Reactive Oxygen Species: Fifteen Years Later. Biochemical Pharmacology. 2006;72:1493–505.
- Dabrowski A, Boguslowicz C, Dabrowska M, Tribillo I, Gabryelewicz A. Reactive Oxygen Species Activate Mitogen-Activated Protein Kinases In Pancreatic Acinar Cells. Pancreas. 2000;21:376–84. [CrossRef]
- Ringvold HC, Khalil RA. Chapter Six - Protein Kinase C as Regulator of Vascular Smooth Muscle Function and Potential Target in Vascular Disorders. Vascul. Pharmacol., vol. 78, Academic Press; 2017, p. 203–301.
- Bitar MS, Al-Mulla F. Upregulation Of CREM/ICER Suppresses Wound Endothelial CRE-HIF-1α-VEGF-Dependent Signaling And Impairs Angiogenesis In Type 2 Diabetes. DMM Disease Models and Mechanisms. 2015;8:65–80.
- Bitar MS, Wahid S, Mustafa S, Al-Saleh E, Dhaunsi GS, Al-Mulla F. Nitric Oxide Dynamics And Endothelial Dysfunction In Type II Model Of Genetic Diabetes. European Journal of Pharmacology. 2005;511:53–64. [CrossRef]
- Bitar MS, Al-Mulla F. A Defect In Nrf2 Signaling Constitutes A Mechanism For Cellular Stress Hypersensitivity In A Genetic Rat Model Of Type 2 Diabetes. American Journal of Physiology - Endocrinology and Metabolism. 2011;301. [CrossRef]
- Akhter N, Madhoun A, Arefanian H, Wilson A, Kochumon S, Thomas R, Shenouda S, Al-Mulla F, Ahmad R, Sindhu S. Oxidative Stress Induces Expression Of The Toll-Like Receptors (TLRs) 2 And 4 In The Human Peripheral Blood Mononuclear Cells: Implications For Metabolic Inflammation. Cellular Physiology and Biochemistry. 2019;53:1–18. [CrossRef]
- Ben Djoudi Ouadda A, Levy E, Ziv E, Lalonde G, Sané AT, Delvin E, Elchebly M. Increased Hepatic Lipogenesis In Insulin Resistance And Type 2 Diabetes Is Associated With AMPK Signalling Pathway Up-Regulation In Psammomys Obesus. Bioscience Reports. 2009;29:283–92.
- Ji J, Petropavlovskaia M, Khatchadourian A, Patapas J, Makhlin J, Rosenberg L, Maysinger D. Type 2 Diabetes Is Associated With Suppression Of Autophagy And Lipid Accumulation In β-Cells. Journal of Cellular and Molecular Medicine. 2019;23:2890–900. [CrossRef]
- Luo Z, Zhang Y, Li F, He J, Ding H, Yan L, Cheng H. Resistin Induces Insulin Resistance By Both AMPK-Dependent And AMPK-Independent Mechanisms In HepG2 Cells. Endocrine. 2009;36:60–9. [CrossRef]
- Fujii N, Ho RC, Manabe Y, Jessen N, Toyoda T, Holland WL, Summers SA, Hirshman MF, Goodyear LJ. Ablation Of AMP-Activated Protein Kinase α 2 Activity Exacerbates Insulin Resistance Induced By High-Fat Feeding Of Mice. Diabetes. 2008;57:2958–66. [CrossRef]
- Kraegen EW, Saha AK, Preston E, Wilks D, Hoy AJ, Cooney GJ, Ruderman NB. Increased Malonyl-CoA And Diacylglycerol Content And Reduced AMPK Activity Accompany Insulin Resistance Induced By Glucose Infusion In Muscle And Liver Of Rats. American Journal of Physiology - Endocrinology and Metabolism. 2006;290:471–9.
- Lee JM, Seo WY, Song KH, Chanda D, Kim YD, Kim DK, Lee MW, Ryu D, Kim YH, Noh JR, Lee CH, Chiang JYL, Koo SH, Choi HS. AMPK-Dependent Repression Of Hepatic Gluconeogenesis Via Disruption Of CREB·CRTC2 Complex By Orphan Nuclear Receptor Small Heterodimer Partner. Journal of Biological Chemistry. 2010;285:32182–91. [CrossRef]
- Lu J. The Warburg Metabolism Fuels Tumor Metastasis. Cancer and Metastasis Reviews. 2019;38:157–64. [CrossRef]
- Hsu CC, Tseng LM, Lee HC. Role Of Mitochondrial Dysfunction In Cancer Progression. Experimental Biology and Medicine. 2016;241:1281–95. [CrossRef]
- Chaudhary AK, Bhat TA, Kumar S, Kumar A, Kumar R, Underwood W, Koochekpour S, Shourideh M, Yadav N, Dhar S, Chandra D. Mitochondrial Dysfunction-Mediated Apoptosis Resistance Associates With Defective Heat Shock Protein Response In African-American Men With Prostate Cancer. British Journal of Cancer. 2016;114:1090–100. [CrossRef]
- Esner M, Graifer D, Lleonart ME, Lyakhovich A. Targeting Cancer Cells Through Antibiotics-Induced Mitochondrial Dysfunction Requires Autophagy Inhibition. Cancer Letters. 2017;384:60–9. [CrossRef]
- Sun J, Hoshino H, Takaku K, Nakajima O, Muto A, Suzuki H, Tashiro S, Takahashi S, Shibahara S, Alam J, Taketo MM, Yamamoto M, Igarashi K. Hemoprotein Bach1 Regulates Enhancer Availability Of Heme Oxygenase-1 Gene. EMBO Journal. 2002;21:5216–24. [CrossRef]
- Dohi Y, Ikura T, Hoshikawa Y, Katoh Y, Ota K, Nakanome A, Muto A, Omura S, Ohta T, Ito A, Yoshida M, Noda T, Igarashi K. Bach1 Inhibits Oxidative Stress-Induced Cellular Senescence By Impeding P53 Function On Chromatin. Nature Structural and Molecular Biology. 2008;15:1246–54. [CrossRef]
- Lee J et al. Effective Breast Cancer Combination Therapy Targeting BACH1 And Mitochondrial Metabolism. Nature. 2019;568:254–8.
- Pomatto LCD, Davies KJA. The Role Of Declining Adaptive Homeostasis In Ageing. Journal of Physiology. 2017;595:7275–309. [CrossRef]
- Wiel C, Le Gal K, Ibrahim MX, Jahangir CA, Kashif M, Yao H, Ziegler D V., Xu X, Ghosh T, Mondal T, Kanduri C, Lindahl P, Sayin VI, Bergo MO. BACH1 Stabilization By Antioxidants Stimulates Lung Cancer Metastasis. Cell. 2019;178:330-345.e22. [CrossRef]
- Mansoori B, Dehghan R, Duijf P, Mohammadi A. MiR - 142 - 3p As Tumor Suppressor MiRNA In The Regulation Of Tumorigenicity, Invasion And Migration Of Human Breast Cancer By Targeting Bach - 1 Expression 2018.
- Id SS, Mansoori B, Mohammadi A, Shajari N, Duijf PHG, Najafi S. MiR-330 Regulates Colorectal Cancer Oncogenesis By Targeting BACH1. Tabriz University of Medical Sciences. 2020;10:444–51. [CrossRef]
- Zhou L, Zhang H, Davies KJA, Forman HJ. Aging-Related Decline In The Induction Of Nrf2-Regulated Antioxidant Genes In Human Bronchial Epithelial Cells. Redox Biology. 2018;14:35–40. [CrossRef]
- Sun RC, Fadia M, Dahlstrom JE, Parish CR, Board PG, Blackburn AC. Reversal Of The Glycolytic Phenotype By Dichloroacetate Inhibits Metastatic Breast Cancer Cell Growth In Vitro And In Vivo. Breast Cancer Research and Treatment. 2010;120:253–60. [CrossRef]
- De Rosa V, Iommelli F, Monti M, Fonti R, Votta G, Stoppelli MP, Del Vecchio S. Reversal Of Warburg Effect And Reactivation Of Oxidative Phosphorylation By Differential Inhibition Of EGFR Signaling Pathways In Non-Small Cell Lung Cancer. Clinical Cancer Research. 2015;21:5110–20. [CrossRef]
- Zhuang Y, Keith WK. Cell Cycle Arrest In Metformin Treated Breast Cancer Cells Involves Activation Of AMPK, Downregulation Of Cyclin D1, And Requires P27Kip1 Or P21Cip1. Journal of Molecular Signaling. 2008;3:1–11.
- Liang J, Shao SH, Xu ZX, Hennessy B, Ding Z, Larrea M, Kondo S, Dumont DJ, Gutterman JU, Walker CL, Slingerland JM, Mills GB. The Energy Sensing LKB1-AMPK Pathway Regulates P27 Phosphorylation Mediating The Decision To Enter Autophagy Or Apoptosis. Nature Cell Biology. 2007;9:218–24. [CrossRef]
- Yang J, Nie J, Ma X, Wei Y, Peng Y, Wei X. Targeting PI3K In Cancer: Mechanisms And Advances In Clinical Trials. Molecular Cancer. 2019;18:1–28. [CrossRef]
- Khan KH, Wong M, Rihawi K, Bodla S, Morganstein D, Banerji U, Molife LR. Hyperglycemia And Phosphatidylinositol 3-Kinase/Protein Kinase B/Mammalian Target Of Rapamycin (PI3K/AKT/MTOR) Inhibitors In Phase I Trials: Incidence, Predictive Factors, And Management. The Oncologist. 2016;21:855–60. [CrossRef]
- Cui Y, Zhou J, Rong F. Combination Of Metformin And RG7388 Enhances Inhibition Of Growth And Induction Of Apoptosis Of Ovarian Cancer Cells Through The PI3K/AKT/MTOR Pathway. Biochemical and Biophysical Research Communications. 2020;533:665–71. [CrossRef]
- Candido S, Abrams SL, Steelman L, Lertpiriyapong K, Martelli AM, Cocco L, Ratti S, Follo MY, Murata RM, Rosalen PL, Lombardi P, Montalto G, Cervello M, Gizak A, Rakus D, Suh P-G, Libra M, McCubrey JA. Metformin Influences Drug Sensitivity In Pancreatic Cancer Cells. Advances in Biological Regulation. 2018;68:13–30. [CrossRef]
- Cantley LC. The Phosphoinositide 3-Kinase Pathway. Science. 2002;296:1655–7.
- Lennartz MR. Phospholipases And Phagocytosis: The Role Of Phospholipid-Derived Second Messengers In Phagocytosis. The International Journal of Biochemistry & Cell Biology. 1999;31:415–30. [CrossRef]
- Fang Y, Park IH, Wu AL, Du G, Huang P, Frohman MA, Walker SJ, Brown HA, Chen J. PLD1 Regulates MTOR Signaling And Mediates Cdc42 Activation Of S6K1. Current Biology. 2003;13:2037–44. [CrossRef]
- O’neil TK, Duffy LR, Frey JW, Hornberger TA. The Role Of Phosphoinositide 3-Kinase And Phosphatidic Acid In The Regulation Of Mammalian Target Of Rapamycin Following Eccentric Contractions. Journal of Physiology. 2009;587:3691–701. [CrossRef]
- Hong-Brown LQ, Brown CR, Navaratnarajah M, Lang CH. Activation Of AMPK/TSC2/PLD By Alcohol Regulates MTORC1 And MTORC2 Assembly In C2C12 Myocytes. Alcoholism: Clinical and Experimental Research. 2013;37:1849–61.
- Standaert ML, Avignon A, Yamada K, Bandyopadhyay G, Farese R V. The Phosphatidylinositol 3-Kinase Inhibitor, Wortmannin, Inhibits Insulin-Induced Activation Of Phosphatidylcholine Hydrolysis And Associated Protein Kinase C Translocation In Rat Adipocytes. Biochemical Journal. 1996;313:1039–46. [CrossRef]
- Lee JS, Kim JH, Jang IH, Kim HS, Han JM, Kazlauskas A, Yagisawa H, Suh P-G, Ryu SH. Phosphatidylinositol (3,4,5)-Trisphosphate Specifically Interacts With The Phox Homology Domain Of Phospholipase D1 And Stimulates Its Activity. Journal of Cell Science. 2005;118:4405–13. [CrossRef]
- Toschi A, Lee E, Xu L, Garcia A, Gadir N, Foster DA. Regulation Of MTORC1 And MTORC2 Complex Assembly By Phosphatidic Acid: Competition With Rapamycin. Molecular and Cellular Biology. 2009;29:1411–20. [CrossRef]
- Franke TF, Kaplan DR, Cantley LC, Toker A. Direct Regulation Of The Akt Proto-Oncogene Product By Phosphatidylinositol-3,4-Bisphosphate. Science. 1997;275:665–8.
- Frame S, Cohen P, Biondi RM. A Common Phosphate Binding Site Explains The Unique Substrate Specificity Of GSK3 And Its Inactivation By Phosphorylation. Molecular Cell. 2001;7:1321–7. [CrossRef]
- Salazar M, Rojo AI, Velasco D, de Sagarra RM, Cuadrado A. Glycogen Synthase Kinase-3β Inhibits The Xenobiotic And Antioxidant Cell Response By Direct Phosphorylation And Nuclear Exclusion Of The Transcription Factor Nrf2*. Journal of Biological Chemistry. 2006;281:14841–51. [CrossRef]
- Kensler TW, Wakabayashi N, Biswal S. Cell Survival Responses To Environmental Stresses Via The Keap1-Nrf2-ARE Pathway. Annual Review of Pharmacology and Toxicology. 2007;47:89–116. [CrossRef]
- Wang X-J, Sun Z, Villeneuve NF, Zhang S, Zhao F, Li Y, Chen W, Yi X, Zheng W, Wondrak GT, Wong PK, Zhang DD. Nrf2 Enhances Resistance Of Cancer Cells To Chemotherapeutic Drugs, The Dark Side Of Nrf2. Carcinogenesis. 2008;29:1235–43. [CrossRef]
- Mitsuishi Y, Taguchi K, Kawatani Y, Shibata T, Nukiwa T, Aburatani H, Yamamoto M, Motohashi H. Nrf2 Redirects Glucose And Glutamine Into Anabolic Pathways In Metabolic Reprogramming. Cancer Cell. 2012;22:66–79. [CrossRef]
- DeNicola GM, Karreth FA, Humpton TJ, Gopinathan A, Wei C, Frese K, Mangal D, Yu KH, Yeo CJ, Calhoun ES, Scrimieri F, Winter JM, Hruban RH, Iacobuzio-Donahue C, Kern SE, Blair IA, Tuveson DA. Oncogene-Induced Nrf2 Transcription Promotes ROS Detoxification And Tumorigenesis. Nature. 2011;475:106–9. [CrossRef]
- Wang H et al. NRF2 Activation By Antioxidant Antidiabetic Agents Accelerates Tumor Metastasis. Science Translational Medicine. 2016;8:334ra51-334ra51.
- Padmanabhan B, Tong KI, Ohta T, Nakamura Y, Scharlock M, Ohtsuji M, Kang M-I, Kobayashi A, Yokoyama S, Yamamoto M. Structural Basis For Defects Of Keap1 Activity Provoked By Its Point Mutations In Lung Cancer. Molecular Cell. 2006;21:689–700. [CrossRef]
- Mukhopadhyay S, Goswami D, Adiseshaiah PP, Burgan W, Yi M, Guerin TM, Kozlov S V, Nissley D V, McCormick F. Undermining Glutaminolysis Bolsters Chemotherapy While NRF2 Promotes Chemoresistance In KRAS-Driven Pancreatic Cancers. Cancer Research. 2020;80:1630–43.
- Hayes JD, Dinkova-Kostova AT. The Nrf2 Regulatory Network Provides An Interface Between Redox And Intermediary Metabolism. Trends in Biochemical Sciences. 2014;39:199–218. [CrossRef]
- Taguchi K, Motohashi H, Yamamoto M. Molecular Mechanisms Of The Keap1–Nrf2 Pathway In Stress Response And Cancer Evolution. Genes to Cells. 2011;16:123–40. [CrossRef]
- Joza N, Oudit GY, Brown D, Bénit P, Kassiri Z, Vahsen N, Benoit L, Patel MM, Nowikovsky K, Vassault A, Backx PH, Wada T, Kroemer G, Rustin P, Penninger JM. Muscle-Specific Loss Of Apoptosis-Inducing Factor Leads To Mitochondrial Dysfunction, Skeletal Muscle Atrophy, And Dilated Cardiomyopathy. Molecular and Cellular Biology. 2005;25:10261–72. [CrossRef]
- Susin SA, Lorenzo HK, Zamzami N, Marzo I, Snow BE, Brothers GM, Mangion J, Jacotot E, Costantini P, Loef¯er M, Larochette N, Goodlett DR, Aebersold R, Siderovski DP, M.Penninger J, Kroemer G. Molecular Characterization Of Mitochondrial Apoptosis-Inducing Factor. Letters to Nature. 1999;353:441–6.
- Vahsen N et al. AIF Deficiency Compromises Oxidative Phosphorylation. EMBO Journal. 2004;23:4679–89. [CrossRef]
- Cheung ECC, Joza N, Steenaart NAE, McClellan KA, Neuspiel M, McNamara S, MacLaurin JG, Rippstein P, Park DS, Shore GC, McBride HM, Penninger JM, Slack RS. Dissociating The Dual Roles Of Apoptosis-Inducing Factor In Maintaining Mitochondrial Structure And Apoptosis. EMBO Journal. 2006;25:4061–73. [CrossRef]
- Berg J, Tymoczko J, Stryer L. Gluconeogenesis and Glycolysis Are Reciprocally Regulated. In: Freeman WH, editor. Biochemistry. fifth, New York: 2002, p. 1–4.
- Pizarro-Delgado J, Deeney JT, Corkey BE, Tamarit-Rodriguez J. Direct Stimulation Of Islet Insulin Secretion By Glycolytic And Mitochondrial Metabolites In KCL-Depolarized Islets. PLoS ONE. 2016;11:1–18.
- OWEN MR, DORAN E, HALESTRAP AP. Evidence That Metformin Exerts Its Anti-Diabetic Effects Through Inhibition Of Complex 1 Of The Mitochondrial Respiratory Chain. Biochemical Journal. 2000;348:607–14.
- Wheaton WW, Weinberg SE, Hamanaka RB, Soberanes S, Sullivan LB, Anso E, Glasauer A, Dufour E, Mutlu GM, Scott Budigner GR, Chandel NS. Metformin Inhibits Mitochondrial Complex I Of Cancer Cells To Reduce Tumorigenesis. ELife. 2014;2014:1–18.
- Anedda A, Rial E, Gonza MM. Metformin Induces Oxidative Stress In White Adipocytes And Raises Uncoupling Protein 2 Levels. Journal of Endocrinology. 2008;199:33–40. [CrossRef]
- Huang X, Liu G, Guo J, Su ZQ. The PI3K/AKT Pathway In Obesity And Type 2 Diabetes. International Journal of Biological Sciences. 2018;14:1483–96. [CrossRef]
- Kousteni S. FoxO1, The Transcriptional Chief Of Staff Of Energy Metabolism. Bone. 2012;50:437–43.
- O-Sullivan I, Zhang W, Wasserman DH, Liew CW, Liu J, Paik J, Depinho RA, Stolz DB, Kahn CR, Schwartz MW, Unterman TG. FoxO1 Integrates Direct And Indirect Effects Of Insulin On Hepatic Glucose Production And Glucose Utilization. Nature Communications. 2015;6. [CrossRef]
- Hay N. Interplay Between FOXO, TOR, And Akt. Biochimica et Biophysica Acta - Molecular Cell Research. 2011;1813:1965–70.
- Cross DAE, Alessi DR, Cohen P, Andjelkovich M, Hemmings BA. Inhibition Of Glycogen Synthase Kinase-3 By Insulin Mediated By Protein Kinase B. Nature. 1995;378:785–9. [CrossRef]
- Krycer JR, Sharpe LJ, Luu W, Brown AJ. The Akt-SREBP Nexus: Cell Signaling Meets Lipid Metabolism. Trends in Endocrinology and Metabolism. 2010;21:268–76.
- Ortega-Molina A, Lopez-Guadamillas E, Mattison JA, Mitchell SJ, Muñoz-Martin M, Iglesias G, Gutierrez VM, Vaughan KL, Szarowicz MD, González-García I, López M, Cebrián D, Martinez S, Pastor J, De Cabo R, Serrano M. Pharmacological Inhibition Of PI3K Reduces Adiposity And Metabolic Syndrome In Obese Mice And Rhesus Monkeys. Cell Metabolism. 2015;21:558–70. [CrossRef]
- Wang C et al. FAM3A Activates PI3K P110$α$/Akt Signaling To Ameliorate Hepatic Gluconeogenesis And Lipogenesis. Hepatology. 2014;59:1779–90.
- Golbidi S, Badran M, Laher I. Diabetes And Alpha Lipoic Acid. Frontiers in Pharmacology. 2011;2 NOV:1–15. [CrossRef]
- Schwartz L, Seyfried T, Alfarouk KO, Da Veiga Moreira J, Fais S. Out Of Warburg Effect: An Effective Cancer Treatment Targeting The Tumor Specific Metabolism And Dysregulated PH. Seminars in Cancer Biology. 2017;43:134–8. [CrossRef]
- Babu PVA, Liu D, Gilbert ER. Recent Advances In Understanding The Anti-Diabetic Actions Of Dietary Flavonoids. Journal of Nutritional Biochemistry. 2013;24:1777–89. [CrossRef]
- Sak K. Cytotoxicity Of Dietary Flavonoids On Diffeernt Human Cancer Types. Pharmacognosy Review. 2014;8:1–36.
- Feuerecker B, Pirsig S, Seidl C, Aichler M, Feuchtinger A, Bruchelt G, Senekowitsch-Schmidtke R. Lipoic Acid Inhibits Cell Proliferation Of Tumor Cells In Vitro And In Vivo. Cancer Biology and Therapy. 2012;13:1425–35. [CrossRef]
- Na MH, Seo EY, Kim WK. Effects Of α-Lipoic Acid On Cell Proliferation And Apoptosis In MDA-MB-231 Human Breast Cells. Nutrition Research and Practice. 2009;3:265. [CrossRef]
- Jeon MJ, Kim WG, Lim S, Choi HJ, Sim S, Kim TY, Shong YK, Kim WB. Alpha Lipoic Acid Inhibits Proliferation And Epithelial Mesenchymal Transition Of Thyroid Cancer Cells. Molecular and Cellular Endocrinology. 2016;419:113–23. [CrossRef]
- Konrad T, Vicini P, Kusterer K, Höflich A, Assadkhani A, Böhles HJ, Sewell A, Tritschler HJ, Cobelli C, Usadel KH. A-Lipoic Acid Treatment Decreases Serum Lactate And Pyruvate Concentrations And Improves Glucose Effectiveness In Lean And Obese Patients With Type 2 Diabetes. Diabetes Care. 1999;22:280–7.
- Korotchkina LG, Sidhu S, Patel MS. R-Lipoic Acid Inhibits Mammalian Pyruvate Dehydrogenase Kinase. Free Radical Research. 2004;38:1083–92. [CrossRef]
- Gandhi VM, Wagh SS, Natraj C V., Menon KKG. Lipoic Acid And Diabetes II: Mode Of Action Of Lipoic Acid. Journal of Biosciences. 1985;9:117–27. [CrossRef]
- Wenzel U, Nickel A, Daniel H. α-Lipoic Acid Induces Apoptosis In Human Colon Cancer Cells By Increasing Mitochondrial Respiration With A Concomitant O2-.-Generation. Apoptosis. 2005;10:359–68.
- Mantovani G, Macciò A, Madeddu C, Mura L, Gramignano G, Lusso MR, Murgia V, Camboni P, Ferreli L, Mocci M, Massa E. The Impact Of Different Antioxidant Agents Alone Or In Combination On Reactive Oxygen Species, Antioxidant Enyzmes And Cytokines In A Series Of Advanced Cancer Patients At Different Sites: Correlation With Disease Progression. Free Radical Research. 2003;37:213–23.
- Mantovani G, Macciò A, Madeddu C, Mura L, Massa E, Gramignano G, Lusso MR, Murgia V, Camboni P, Ferreli L. Reactive Oxygen Species, Antioxidant Mechanisms And Serum Cytokine Levels In Cancer Patients: Impact Of An Antioxidant Treatment. Journal of Cellular and Molecular Medicine. 2002;6:570–82. [CrossRef]
- Dörsam B, Fahrer J. The Disulfide Compound α-Lipoic Acid And Its Derivatives: A Novel Class Of Anticancer Agents Targeting Mitochondria. Cancer Letters. 2016;371:12–9. [CrossRef]
- Dörsam B, Göder A, Seiwert N, Kaina B, Fahrer J. Lipoic Acid Induces P53-Independent Cell Death In Colorectal Cancer Cells And Potentiates The Cytotoxicity Of 5-Fluorouracil. Archives of Toxicology. 2015;89:1829–46. [CrossRef]
- Kafara P, Icard P, Guillamin M, Schwartz L, Lincet H. Lipoic Acid Decreases Mcl-1, Bcl-XL And Up Regulates Bim On Ovarian Carcinoma Cells Leading To Cell Death. Journal of Ovarian Research. 2015;8:1–13.
- Piotrowski P, Wierzbicka K, Śmiałek M. Neuronal Death In The Rat Hippocampus In Experimental Diabetes And Cerebral Ischaemia Treated With Antioxidants. Folia Neuropathologica. 2001;39:147–54.
- Pierce RH, Campbell JS, Stephenson AB, Franklin CC, Chaisson M, Poot M, Kavanagh TJ, Rabinovitch PS, Fausto N. Disruption Of Redox Homeostasis In Tumor Necrosis Factor-Induced Apoptosis In A Murine Hepatocyte Cell Line. American Journal of Pathology. 2000;157:221–36. [CrossRef]
- Lin HY, Han HW, Sun WX, Yang YS, Tang CY, Lu GH, Qi JL, Wang XM, Yang YH. Design And Characterization Of α-Lipoic Acyl Shikonin Ester Twin Drugs As Tubulin And PDK1 Dual Inhibitors. European Journal of Medicinal Chemistry. 2018;144:137–50. [CrossRef]
- Shen QW, Zhu MJ, Tong J, Ren J, Du M. Ca2+/Calmodulin-Dependent Protein Kinase Kinase Is Involved In AMP-Activated Protein Kinase Activation By α-Lipoic Acid In C2C12 Myotubes. American Journal of Physiology - Cell Physiology. 2007;293:1395–403.
- Dozio E, Ruscica M, Passafaro L, Dogliotti G, Steffani L, Pagani A, Demartini G, Esposti D, Fraschini F, Magni P. The Natural Antioxidant Alpha-Lipoic Acid Induces P27Kip1-Dependent Cell Cycle Arrest And Apoptosis In MCF-7 Human Breast Cancer Cells. European Journal of Pharmacology. 2010;641:29–34. [CrossRef]
- Yadav VR, Prasad S, Sung B, Kannappan R, Aggarwal BB. Targeting Inflammatory Pathways By Triterpenoids For Prevention And Treatment Of Cancer. Toxins. 2010;2:2428–66. [CrossRef]
- Wei J, Huang Q, Bai F, Lin J, Nie J, Lu S, Lu C, Huang R, Lu Z, Lin X. Didymin Induces Apoptosis Through Mitochondrial Dysfunction And Up-Regulation Of RKIP In Human Hepatoma Cells. Chemico-Biological Interactions. 2017;261:118–26. [CrossRef]
- Zhao Y, Zhou Y, Wang M. Brosimone I, An Isoprenoid-Substituted Flavonoid, Induces Cell Cycle G1 Phase Arrest And Apoptosis Through ROS-Dependent Endoplasmic Reticulum Stress In HCT116 Human Colon Cancer Cells. Food and Function. 2019;10:2729–38.
- Al-Mulla F. Novel Flavonoid Didymin Inhibits Neuroblastomas - Letter. Cancer Prevention Research. 2012;5:883. [CrossRef]
- Kyriakis E, Stravodimos GA, Kantsadi AL, Chatzileontiadou DSM, Skamnaki VT, Leonidas DD. Natural Flavonoids As Antidiabetic Agents. The Binding Of Gallic And Ellagic Acids To Glycogen Phosphorylase B. FEBS Letters. 2015;589:1787–94. [CrossRef]
- Samec M et al. Flavonoids Against The Warburg Phenotype—Concepts Of Predictive, Preventive And Personalised Medicine To Cut The Gordian Knot Of Cancer Cell Metabolism. EPMA Journal. 2020;11:377–98. [CrossRef]
- Shan S, Shi J, Yang P, Jia B, Wu H, Zhang X, Li Z. Apigenin Restrains Colon Cancer Cell Proliferation Via Targeted Blocking Of Pyruvate Kinase M2-Dependent Glycolysis. Journal of Agricultural and Food Chemistry. 2017;65:8136–44. [CrossRef]
- Wei R, Mao L, Xu P, Zheng X, Hackman RM, MacKenzie GG, Wang Y. Suppressing Glucose Metabolism With Epigallocatechin-3-Gallate (EGCG) Reduces Breast Cancer Cell Growth In Preclinical Models. Food and Function. 2018;9:5682–96. [CrossRef]
- Feng J, Wu L, Ji J, Chen K, Yu Q, Zhang J, Chen J, Mao Y, Wang F, Dai W, Xu L, Wu J, Guo C. PKM2 Is The Target Of Proanthocyanidin B2 During The Inhibition Of Hepatocellular Carcinoma. Journal of Experimental and Clinical Cancer Research. 2019;38:1–15. [CrossRef]
- Chen J, Xie J, Jiang Z, Wang B, Wang Y, Hu X. Shikonin And Its Analogs Inhibit Cancer Cell Glycolysis By Targeting Tumor Pyruvate Kinase-M2. Oncogene. 2011;30:4297–306. [CrossRef]
- Zhao X, Zhu Y, Hu J, Jiang L, Li L, Jia S, Zen K. Shikonin Inhibits Tumor Growth In Mice By Suppressing Pyruvate Kinase M2-Mediated Aerobic Glycolysis. Scientific Reports. 2018;8:1–8. [CrossRef]
- Jia L, Huang S, Yin X, Zan Y, Guo Y, Han L. Quercetin Suppresses The Mobility Of Breast Cancer By Suppressing Glycolysis Through Akt-MTOR Pathway Mediated Autophagy Induction. Life Sciences. 2018;208:123–30.
- Liu W, Li W, Liu H, Yu X. Xanthohumol Inhibits Colorectal Cancer Cells Via Downregulation Of Hexokinases Ii-Mediated Glycolysis. International Journal of Biological Sciences. 2019;15:2497–508. [CrossRef]
- Deng X, Liu R, Li J, Li Z, Liu J, Xiong R, Lei X, Zheng X, Xie Z, Tang G. Design, Synthesis, And Preliminary Biological Evaluation Of 3′,4′,5′-Trimethoxy Flavonoid Salicylate Derivatives As Potential Anti-Tumor Agents. New Journal of Chemistry. 2019;43:1874–84.
- Guo Y, Wei L, Zhou Y, Lu N, Tang X, Li Z, Wang X. Flavonoid GL-V9 Induces Apoptosis And Inhibits Glycolysis Of Breast Cancer Via Disrupting GSK-3β-Modulated Mitochondrial Binding Of HKII. Free Radical Biology and Medicine. 2020;146:119–29. [CrossRef]
- Zhou Y, Lu N, Qiao C, Ni T, Li Z, Yu B, Guo Q, Wei L. FV-429 Induces Apoptosis And Inhibits Glycolysis By Inhibiting Akt-Mediated Phosphorylation Of Hexokinase II In MDA-MB-231 Cells. Molecular Carcinogenesis. 2016;55:1317–28. [CrossRef]
- Tao L, Liu Y, Ding Y, Liu X, Zhang X, Hu R, Wei L, Wang X, Yao Y, Lu J, Wang Q. Gen-27, a newly synthesized flavonoid, inhibits glycolysis and induces cell apoptosis via suppression of hexokinase II in human breast cancer cells. vol. 125. 2017. [CrossRef]
- Li W, Hao J, Zhang L, Cheng Z, Deng X, Shu G. Astragalin Reduces Hexokinase 2 Through Increasing MiR-125b To Inhibit The Proliferation Of Hepatocellular Carcinoma Cells In Vitro And In Vivo. Journal of Agricultural and Food Chemistry. 2017;65:5961–72. [CrossRef]
- Mazlaghaninia M, Atri MS, Seyedalipour B. Scopoletin And Morin Inhibit Lactate Dehydrogenase Enzyme Activity Is Critical For Cancer Metabolism. Hormozgan Medical Journal. 2019;In Press:1–6. [CrossRef]
- Liu Y, Venna CK, Morgan JB, Mohammed KA, Jekabsons MB, Nagle DG, Zhou YD. Methylalpinumisoflavone Inhibits Hypoxia-Inducible Factor-1 (HIF-1) Activation By Simultaneously Targeting Multiple Pathways. Journal of Biological Chemistry. 2009;284:5859–68. [CrossRef]
- Wei L, Zhou Y, Qiao C, Ni T, Li Z, You Q, Guo Q, Lu N. Oroxylin A Inhibits Glycolysis-Dependent Proliferation Of Human Breast Cancer Via Promoting SIRT3-Mediated SOD2 Transcription And HIF1α Destabilization. Cell Death and Disease. 2015;6:1–12. [CrossRef]
- Chen F, Zhuang M, Zhong C, Peng J, Wang X, Li J, Chen Z, Huang Y. Baicalein Reverses Hypoxia-Induced 5-FU Resistance In Gastric Cancer AGS Cells Through Suppression Of Glycolysis And The PTEN/Akt/HIF-1α Signaling Pathway. Oncology Reports. 2015;33:457–63. [CrossRef]
- Wang H, Zhao L, Zhu LT, Wang Y, Pan D, Yao J, You QD, Guo QL. Wogonin Reverses Hypoxia Resistance Of Human Colon Cancer HCT116 Cells Via Downregulation Of HIF-1α And Glycolysis, By Inhibiting PI3K/Akt Signaling Pathway. Molecular Carcinogenesis. 2014;53:1–12.
- Zhao Y, Zhang L, Wu Y, Dai Q, Zhou Y, Li Z, Yang L, Guo Q, Lu N. Selective Anti-Tumor Activity Of Wogonin Targeting The Warburg Effect Through Stablizing P53. Pharmacological Research. 2018;135:49–59. [CrossRef]
- Li J, Zou Y, Pei M, Zhang Y, Jiang Y. Berberine Inhibits The Warburg Effect Through TET3/MiR-145/HK2 Pathways In Ovarian Cancer Cells. Journal of Cancer. 2021;12:207–16.
- Li Z, Li H, Lu Y, Yang P, Li Z. Berberine Inhibited The Proliferation Of Cancer Cells By Suppressing The Activity Of Tumor Pyruvate Kinase M2. Natural Product Communications. 2017;12:1415–8. [CrossRef]
- Li W, Ma X, Li N, Liu H, Dong Q, Zhang J, Yang C, Liu Y, Liang Q, Zhang S, Xu C, Song W, Tan S, Rong P, Wang W. Resveratrol Inhibits Hexokinases II Mediated Glycolysis In Non-Small Cell Lung Cancer Via Targeting Akt Signaling Pathway. Experimental Cell Research. 2016;349:320–7. [CrossRef]
- Saunier E, Antonio S, Regazzetti A, Auzeil N, Laprévote O, Shay JW, Coumoul X, Barouki R, Benelli C, Huc L, Bortoli S. Resveratrol Reverses The Warburg Effect By Targeting The Pyruvate Dehydrogenase Complex In Colon Cancer Cells. Scientific Reports. 2017;7:1–16. [CrossRef]
- Omidian K, Rafiei H, Bandy B. Increased Mitochondrial Content And Function By Resveratrol And Select Flavonoids Protects Against Benzo[A]Pyrene-Induced Bioenergetic Dysfunction And ROS Generation In A Cell Model Of Neoplastic Transformation. Free Radical Biology and Medicine. 2020;152:767–75. [CrossRef]
- Samec M, Liskova A, Koklesova L, Mersakova S, Strnadel J, Kajo K, Pec M, Zhai K, Smejkal K, Mirzaei S, Hushmandi K, Ashrafizadeh M, Saso L, Brockmueller A, Shakibaei M, Büsselberg D, Kubatka P. Flavonoids Targeting HIF-1: Implications On Cancer Metabolism. Cancers. 2021;13:130. [CrossRef]
- Bisol Â, de Campos PS, Lamers ML. Flavonoids As Anticancer Therapies: A Systematic Review Of Clinical Trials. Phytotherapy Research. 2020;34:568–82. [CrossRef]
- Thilakarathna SH, Vasantha Rupasinghe HP. Flavonoid Bioavailability And Attempts For Bioavailability Enhancement. Nutrients. 2013;5:3367–87. [CrossRef]
- dos Santos Lima B, Shanmugam S, de Souza Siqueira Quintans J, Quintans-Júnior LJ, de Souza Araújo AA. Inclusion Complex With Cyclodextrins Enhances The Bioavailability Of Flavonoid Compounds: A Systematic Review. Phytochemistry Reviews. 2019;18:1337–59. [CrossRef]
- Foretz M, Ancellin N, Andreelli F, Saintillan Y, Grondin P, Kahn A, Thorens B, Vaulont S, Viollet B. Short-Term Overexpression Of A Constitutively Active Form Of AMP-Activated Protein Kinase In The Liver Leads To Mild Hypoglycemia And Fatty Liver. Diabetes. 2005;54:1331–9. [CrossRef]
- Woo JL, Song KH, Eun HK, Jong CW, Hyoun SK, Park HS, Kim MS, Kim SW, Lee KU, Park JY. α-Lipoic Acid Increases Insulin Sensitivity By Activating AMPK In Skeletal Muscle. Biochemical and Biophysical Research Communications. 2005;332:885–91. [CrossRef]
- Gosselin LE, Chrapowitzky L, Rideout TC. Metabolic Effects Of α-Lipoic Acid Supplementation In Pre-Diabetics: A Randomized, Placebo-Controlled Pilot Study. Food and Function. 2019;10:5732–8.
- Kamenova P. Improvement Of Insulin Sensitivity In Patients With Type 2 Diabetes Mellitus After Oral Administration Of Alpha-Lipoic Acid. Hormones. 2006;5:251–8. [CrossRef]
- Ziegler D, Hanefeld M, Ruhnau KJ, Mei\ner HP, Lobisch M, Schütte K, Gries FA. Treatment Of Symptomatic Diabetic Peripheral Neuropathy With The Anti-Oxidant α-Lipoic Acid - A 3-Week Multicentre Randomized Controlled Trial (ALADIN Study). Diabetologia. 1995;38:1425–33.
- Lagouge M, Argmann C, Gerhart-Hines Z, Meziane H, Lerin C, Daussin F, Messadeq N, Milne J, Lambert P, Elliott P, Geny B, Laakso M, Puigserver P, Auwerx J. Resveratrol Improves Mitochondrial Function And Protects Against Metabolic Disease By Activating SIRT1 And PGC-1α. Cell. 2006;127:1109–22. [CrossRef]
- Meng Q, Qi X, Fu Y, Chen Q, Cheng P, Yu X, Sun X, Wu J, Li W, Zhang Q, Li Y, Wang A, Bian H. Flavonoids Extracted From Mulberry (Morus Alba L.) Leaf Improve Skeletal Muscle Mitochondrial Function By Activating AMPK In Type 2 Diabetes. Journal of Ethnopharmacology. 2020;248:112326. [CrossRef]
- Han G, Takahashi H, Murao N, Gheni G, Yokoi N, Hamamoto Y, Asahara S-I, Seino Y, Kido Y, Seino S. Glutamate Is An Essential Mediator In Glutamine-Amplified Insulin Secretion. J Diabetes Investig. 2021;12:920–30. [CrossRef]
- Satapati S, Sunny NE, Kucejova B, Fu X, He TT, Méndez-Lucas A, Shelton JM, Perales JC, Browning JD, Burgess SC. Elevated TCA Cycle Function In The Pathology Of Diet-Induced Hepatic Insulin Resistance And Fatty Liver. Journal of Lipid Research. 2012;53:1080–92. [CrossRef]
- Maurer J, Hoene M, Weigert C. Signals From The Circle: Tricarboxylic Acid Cycle Intermediates As Myometabokines. Metabolites. 2021;11. [CrossRef]
- Gilbert M. Role Of Skeletal Muscle Lipids In The Pathogenesis Of Insulin Resistance Of Obesity And Type 2 Diabetes. Journal of Diabetes Investigation. 2021;12:1934–41. [CrossRef]
- Nagao H, Nishizawa H, Bamba T, Nakayama Y, Isozumi N, Nagamori S, Kanai Y, Tanaka Y, Kita S, Fukuda S, Funahashi T, Maeda N, Fukusaki E, Shimomura I. Increased Dynamics Of Tricarboxylic Acid Cycle And Glutamate Synthesis In Obese Adipose Tissue: IN VIVO METABOLIC TURNOVER ANALYSIS. The Journal of Biological Chemistry. 2017;292:4469–83.
- Newsholme P, Rowlands J, Rose’meyer R, Cruzat V. Metabolic Adaptions/Reprogramming In Islet Beta-Cells In Response To Physiological Stimulators-What Are The Consequences. Antioxidants (Basel, Switzerland). 2022;11. [CrossRef]
- Zhang GF, Jensen M V., Gray SM, El K, Wang Y, Lu D, Becker TC, Campbell JE, Newgard CB. Reductive TCA Cycle Metabolism Fuels Glutamine- And Glucose-Stimulated Insulin Secretion. Cell Metabolism. 2021;33:804-817.e5. [CrossRef]
- Heart E, Cline GW, Collis LP, Pongratz RL, Gray JP, Smith PJS. Role For Malic Enzyme, Pyruvate Carboxylation, And Mitochondrial Malate Import In Glucose-Stimulated Insulin Secretion. American Journal of Physiology Endocrinology and Metabolism. 2009;296. [CrossRef]
- Wang L, Yi D, Hou Y, Ding B, Li K, Li B, Zhu H, Liu Y, Wu G. The Journal Of Nutrition Nutrient Physiology, Metabolism, And Nutrient-Nutrient Interactions Dietary Supplementation With A-Ketoglutarate Activates MTOR Signaling And Enhances Energy Status In Skeletal Muscle Of Lipopolysaccharide-Challenged Piglets 1,2 n.d.
- Yao K, Yin Y, Li X, Xi P, Wang J, Lei J, Hou Y, Wu G. Alpha-Ketoglutarate Inhibits Glutamine Degradation And Enhances Protein Synthesis In Intestinal Porcine Epithelial Cells. Amino Acids. 2012;42:2491–500. [CrossRef]
- Fazakerley DJ, Krycer JR, Kearney AL, Hocking SL, James DE. Muscle And Adipose Tissue Insulin Resistance: Malady Without Mechanism? Journal of Lipid Research. 2019;60:1720–32.
- Tretter L, Adam-Vizi V. Alpha-Ketoglutarate Dehydrogenase: A Target And Generator Of Oxidative Stress. Philosophical Transactions of the Royal Society of London Series B, Biological Sciences. 2005;360:2335–45. [CrossRef]
- Selak MA, Armour SM, MacKenzie ED, Boulahbel H, Watson DG, Mansfield KD, Pan Y, Simon MC, Thompson CB, Gottlieb E. Succinate Links TCA Cycle Dysfunction To Oncogenesis By Inhibiting HIF-α Prolyl Hydroxylase. Cancer Cell. 2005;7:77–85. [CrossRef]
- Gabryelska A, Karuga FF, Szmyd B, Białasiewicz P. HIF-1α As A Mediator Of Insulin Resistance, T2DM, And Its Complications: Potential Links With Obstructive Sleep Apnea. Frontiers in Physiology. 2020;11. [CrossRef]
- Badole SL, Chaudhari SM, Bagul PP, Mahamuni SP, Khose RD. Effect Of Concomitant Administration Of L-Glutamine And Cycloart-23-Ene-3b, 25-Diol (B2) With Sitagliptin In GLP-1 (7-36) Amide Secretion, Biochemical And Oxidative Stress In Streptozotocin-Nicotinamide Induced Diabetic Sprague Dawley Rats. PLoS ONE. 2013;8:72817.
- Badole SL, Jangam GB, Chaudhari SM, Ghule AE, Zanwar AA. L-Glutamine Supplementation Prevents The Development Of Experimental Diabetic Cardiomyopathy In Streptozotocin-Nicotinamide Induced Diabetic Rats. PLOS ONE. 2014;9:e92697. [CrossRef]
- Tsai PH, Liu JJ, Yeh CL, Chiu WC, Yeh SL. Effects Of Glutamine Supplementation On Oxidative Stress-Related Gene Expression And Antioxidant Properties In Rats With Streptozotocin-Induced Type 2 Diabetes. The British Journal of Nutrition. 2012;107:1112–8. [CrossRef]
- Tsai PH, Liu JJ, Chiu WC, Pai MH, Yeh SL. Effects Of Dietary Glutamine On Adhesion Molecule Expression And Oxidative Stress In Mice With Streptozotocin-Induced Type 1 Diabetes. Clinical Nutrition. 2011;30:124–9. [CrossRef]
- Tsai PH, Yeh CL, Liu JJ, Chiu WC, Yeh SL. Effects Of Dietary Glutamine On Inflammatory Mediator Gene Expressions In Rats With Streptozotocin-Induced Diabetes. Nutrition. 2012;28:288–93. [CrossRef]
- Jafari-Vayghan H, Varshosaz P, Hajizadeh-Sharafabad F, Razmi HR, Amirpour M, Tavakoli-Rouzbehani OM, Alizadeh M, Maleki V. A Comprehensive Insight Into The Effect Of Glutamine Supplementation On Metabolic Variables In Diabetes Mellitus: A Systematic Review. Nutrition & Metabolism. 2020;17:80. [CrossRef]
- Noto H, Goto A, Tsujimoto T, Noda M. Cancer Risk In Diabetic Patients Treated With Metformin: A Systematic Review And Meta-Analysis. PLoS ONE. 2012;7:1–9. [CrossRef]
- Saraei P, Asadi I, Kakar MA, Moradi-Kor N. The Beneficial Effects Of Metformin On Cancer Prevention And Therapy: A Comprehensive Review Of Recent Advances. Cancer Management and Research. 2019;11:3295–313. [CrossRef]
- Mallik R, Chowdhury TA. Metformin In Cancer. Diabetes Research and Clinical Practice. 2018;143:409–19.
- Chen TM, Lin CC, Huang PT, Wen CF. Metformin Associated With Lower Mortality In Diabetic Patients With Early Stage Hepatocellular Carcinoma After Radiofrequency Ablation. Journal of Gastroenterology and Hepatology (Australia). 2011;26:858–65. [CrossRef]
- Doherty JR et al. Blocking Lactate Export By Inhibiting The Myc Target MCT1 Disables Glycolysis And Glutathione Synthesis. Cancer Research. 2014;74:908–20.
- Buzzai M, Jones RG, Amaravadi RK, Lum JJ, DeBerardinis RJ, Zhao F, Viollet B, Thompson CB. Systemic Treatment With The Antidiabetic Drug Metformin Selectively Impairs P53-Deficient Tumor Cell Growth. Cancer Research. 2007;67:6745–52.
- DeCensi A, Puntoni M, Goodwin P, Cazzaniga M, Gennari A, Bonanni B, Gandini S. Metformin And Cancer Risk In Diabetic Patients: A Systematic Review And Meta-Analysis. Cancer Prevention Research. 2010;3:1451–61. [CrossRef]
- Kim YS, Choi EA, woo Lee J, Kim Y, You HS, Han YE, Kim HS, Bae YJ, Kang HT, Kim J. Metformin Use Reduced The Overall Risk Of Cancer In Diabetic Patients: A Study Based On The Korean NHIS-HEALS Cohort. Nutrition, Metabolism and Cardiovascular Diseases. 2020;30:1714–22. [CrossRef]
- Kim J, Hyun HJ, Choi EA, Kim Y, Bae YJ, Kang HT. Metformin Use Reduced The Risk Of Stomach Cancer In Diabetic Patients In Korea: An Analysis Of Korean NHIS-HEALS Database. Gastric Cancer. 2020;23:1075–83. [CrossRef]
- Shuai Y, Li C, Zhou X. The Effect Of Metformin On Gastric Cancer In Patients With Type 2 Diabetes: A Systematic Review And Meta-Analysis. Clinical and Translational Oncology. 2020;22:1580–90. [CrossRef]
- Hu J, Fan HD, Gong JP, Mao QS. The Relationship Between The Use Of Metformin And The Risk Of Pancreatic Cancer In Patients With Diabetes: A Systematic Review And Meta-Analysis. BMC Gastroenterology. 2023;23:1–21. [CrossRef]
- Zhang K, Bai P, Dai H, Deng Z. Metformin And Risk Of Cancer Among Patients With Type 2 Diabetes Mellitus: A Systematic Review And Meta-Analysis. Primary Care Diabetes. 2021;15:52–8. [CrossRef]
- Mekuria AN, Ayele Y, Tola A, Mishore KM. Monotherapy With Metformin Versus Sulfonylureas And Risk Of Cancer In Type 2 Diabetic Patients: A Systematic Review And Meta-Analysis. Journal of Diabetes Research. 2019;2019. [CrossRef]
- Cunha V, Cotrim HP, Rocha R, Carvalho K, Lins-Kusterer L. Metformin In The Prevention Of Hepatocellular Carcinoma In Diabetic Patients: A Systematic Review. Annals of Hepatology. 2020;19:232–7. [CrossRef]
- Farmer RE, Ford D, Mathur R, Chaturvedi N, Kaplan R, Smeeth L, Bhaskaran K. Metformin Use And Risk Of Cancer In Patients With Type 2 Diabetes: A Cohort Study Of Primary Care Records Using Inverse Probability Weighting Of Marginal Structural Models. International Journal of Epidemiology. 2019;48:527–37. [CrossRef]
- Govindarajan R, Ratnasinghe L, Simmons DL, Siegel ER, Midathada M V, Kim L, Kim PJ, Owens RJ, Lang NP. Thiazolidinediones And The Risk Of Lung, Prostate, And Colon Cancer In Patients With Diabetes. Journal of Clinical Oncology. 2007;25:1476–81. [CrossRef]
- Du R, Lin L, Cheng D, Xu Y, Xu M, Chen Y, Wang W, Bi Y, Li D, Lu J. Thiazolidinedione Therapy And Breast Cancer Risk In Diabetic Women: A Systematic Review And Meta-Analysis. Diabetes/Metabolism Research and Reviews. 2018;34:9–11. [CrossRef]
- Chang CH, Lin JW, Wu LC, Lai MS, Chuang LM, Arnold Chan K. Association Of Thiazolidinediones With Liver Cancer And Colorectal Cancer In Type 2 Diabetes Mellitus. Hepatology. 2012;55:1462–72. [CrossRef]
- Lin HC, Hsu YT, Kachingwe BH, Hsu CY, Uang YS, Wang LH. Dose Effect Of Thiazolidinedione On Cancer Risk In Type 2 Diabetes Mellitus Patients: A Six-Year Population-Based Cohort Study. Journal of Clinical Pharmacy and Therapeutics. 2014;39:354–60. [CrossRef]
- Liao KF, Lin CL, Lai SW. Association Between Colorectal Cancer And Thiazolidinediones Administration In A Case-Control Study. BioMedicine (France). 2019;9:31–6. [CrossRef]
- Ramos-Nino ME, MacLean CD, Littenberg B. Association Between Cancer Prevalence And Use Of Thiazolidinediones: Results From The Vermont Diabetes Information System. BMC Medicine. 2007;5:1–7. [CrossRef]
- Bsch INC, Bowker SL, Majumdar SR, Johnson JA. Use Of Thiazolidinediones And The Risk Of Bladder Cancer Among People With Type 2 Diabetes: A Meta-Analysis. Canadian Medical Association Journal. 2012;184:675–83.
- Dankner R, Roth J. More Recent, Better Designed Studies Have Weakened Links Between Antidiabetes Medications And Cancer Risk. Diabetic Medicine. 2019;37:194–202. [CrossRef]
- Kim J, Yang G, Kim Y, Kim J, Ha J. AMPK Activators: Mechanisms Of Action And Physiological Activities. Experimental and Molecular Medicine. 2016;48:1–12. [CrossRef]
- Miller RA, Chu Q, Xie J, Foretz M, Viollet B, Birnbaum MJ. Biguanides Suppress Hepatic Glucagon Signaling By Decreasing Production Of Cyclic AMP. Physiology & Behavior. 2019;176:139–48. [CrossRef]
- Foretz M, Hébrard S, Leclerc J, Zarrinpashneh E, Soty M, Mithieux G, Sakamoto K, Andreelli F, Viollet B. Metformin Inhibits Hepatic Gluconeogenesis In Mice Independently Of The LKB1/AMPK Pathway Via A Decrease In Hepatic Energy State. The Journal of Clinical Investigation. 2010;120:2355–69.
- Coughlan KA, Valentine RJ, Ruderman NB, Saha AK. AMPK Activation: A Therapeutic Target For Type 2 Diabetes? Diabetes, Metabolic Syndrome and Obesity: Targets and Therapy. 2014:7–241. [CrossRef]
Figure 1.
Glycolytic metabolic shift in tumor cells compared to the oxidative phosphorylation (OXPHOS) metabolic dominance in normal non-proliferating cells. 1) Pyruvate can enter the mitochondria to be converted to acetyl-CoA, which enters the Krebs cycle (tricarboxylic acid cycle or citric acid cycle). NADH and FADH2 produced from the Krebs cycle enter the electron transport chain (ETC) cycle to generate the energy molecule, adenosine triphosphate (ATP). OXPHOS, also called the ETC cycle, occurs in the inner membrane of the mitochondria and generates approximately 32 ATP energy molecules from a single glucose molecule. 2), the pyruvate, generated in the cytoplasm from the breakdown of one glucose molecule, is utilized by the Warburg aerobic glycolysis or the anaerobic glycolysis to produce lactate and a net of two ATPs. Abbreviations: ADP—adenosine diphosphate; ATP—adenosine triphosphate; ETC—electron transport chain; GLUT4—glucose transporter 4; NADH—nicotinamide adenine dinucleotide; O2—oxygen; TCA—tricarboxylic acid. Created with Biorender.com.
Figure 1.
Glycolytic metabolic shift in tumor cells compared to the oxidative phosphorylation (OXPHOS) metabolic dominance in normal non-proliferating cells. 1) Pyruvate can enter the mitochondria to be converted to acetyl-CoA, which enters the Krebs cycle (tricarboxylic acid cycle or citric acid cycle). NADH and FADH2 produced from the Krebs cycle enter the electron transport chain (ETC) cycle to generate the energy molecule, adenosine triphosphate (ATP). OXPHOS, also called the ETC cycle, occurs in the inner membrane of the mitochondria and generates approximately 32 ATP energy molecules from a single glucose molecule. 2), the pyruvate, generated in the cytoplasm from the breakdown of one glucose molecule, is utilized by the Warburg aerobic glycolysis or the anaerobic glycolysis to produce lactate and a net of two ATPs. Abbreviations: ADP—adenosine diphosphate; ATP—adenosine triphosphate; ETC—electron transport chain; GLUT4—glucose transporter 4; NADH—nicotinamide adenine dinucleotide; O2—oxygen; TCA—tricarboxylic acid. Created with Biorender.com.
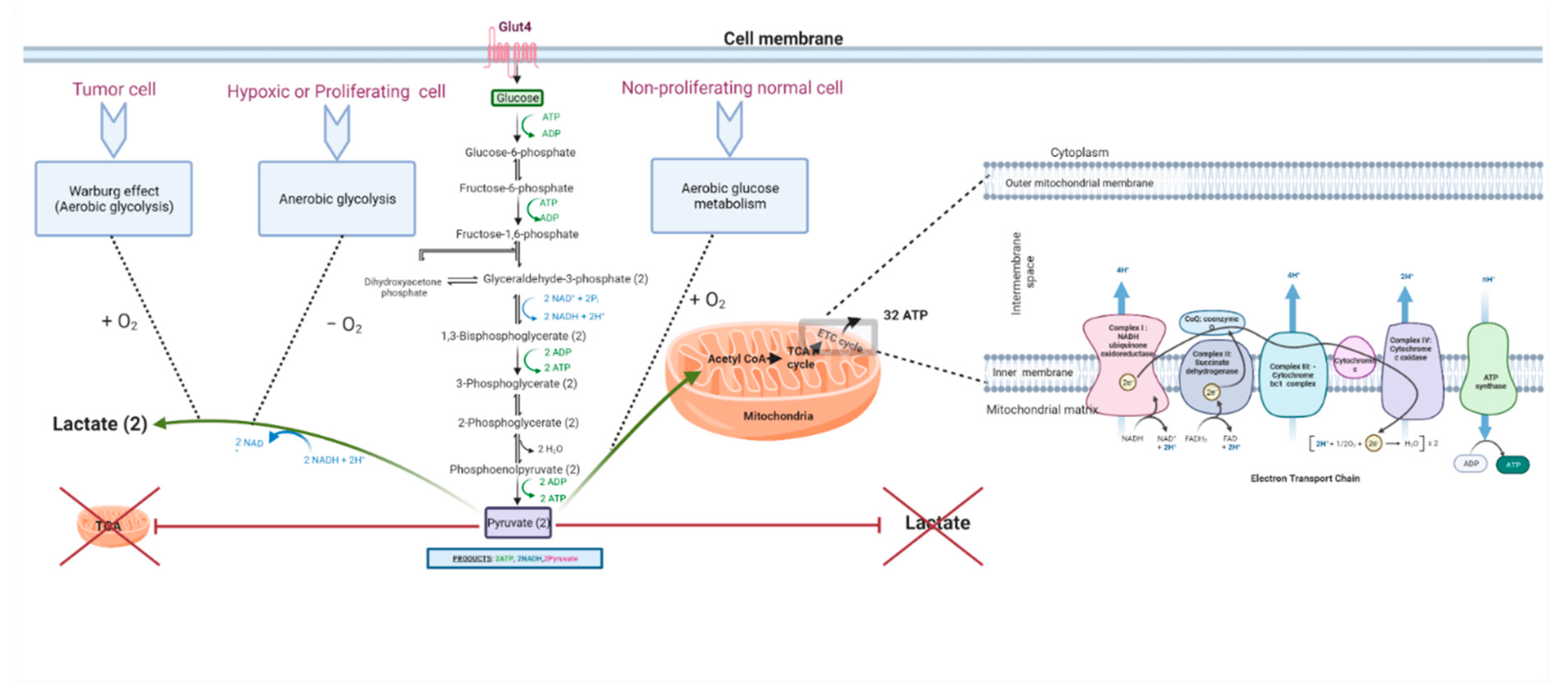
Figure 2.
Different theories behind the Warburg effect in cancer. Abbreviations: ATP—adenosine triphosphate; HIF-1—hypoxia-inducible factor-1; OXPHOS—oxidative phosphorylation; PDC—pyruvate dehydrogenase complex; PDK—pyruvate dehydrogenase kinase; UCP—uncoupling protein 2. Created with Biorender.com.
Figure 2.
Different theories behind the Warburg effect in cancer. Abbreviations: ATP—adenosine triphosphate; HIF-1—hypoxia-inducible factor-1; OXPHOS—oxidative phosphorylation; PDC—pyruvate dehydrogenase complex; PDK—pyruvate dehydrogenase kinase; UCP—uncoupling protein 2. Created with Biorender.com.

Figure 3.
Mechanisms underlying mitochondrial metabolic dysfunction in cancer. Abbreviations: mtDNA—mitochondrial deoxyribonucleic acid; ncDNA—nuclear deoxyribonucleic acid; ROS—reactive oxygen species. Created with Biorender.com.
Figure 3.
Mechanisms underlying mitochondrial metabolic dysfunction in cancer. Abbreviations: mtDNA—mitochondrial deoxyribonucleic acid; ncDNA—nuclear deoxyribonucleic acid; ROS—reactive oxygen species. Created with Biorender.com.
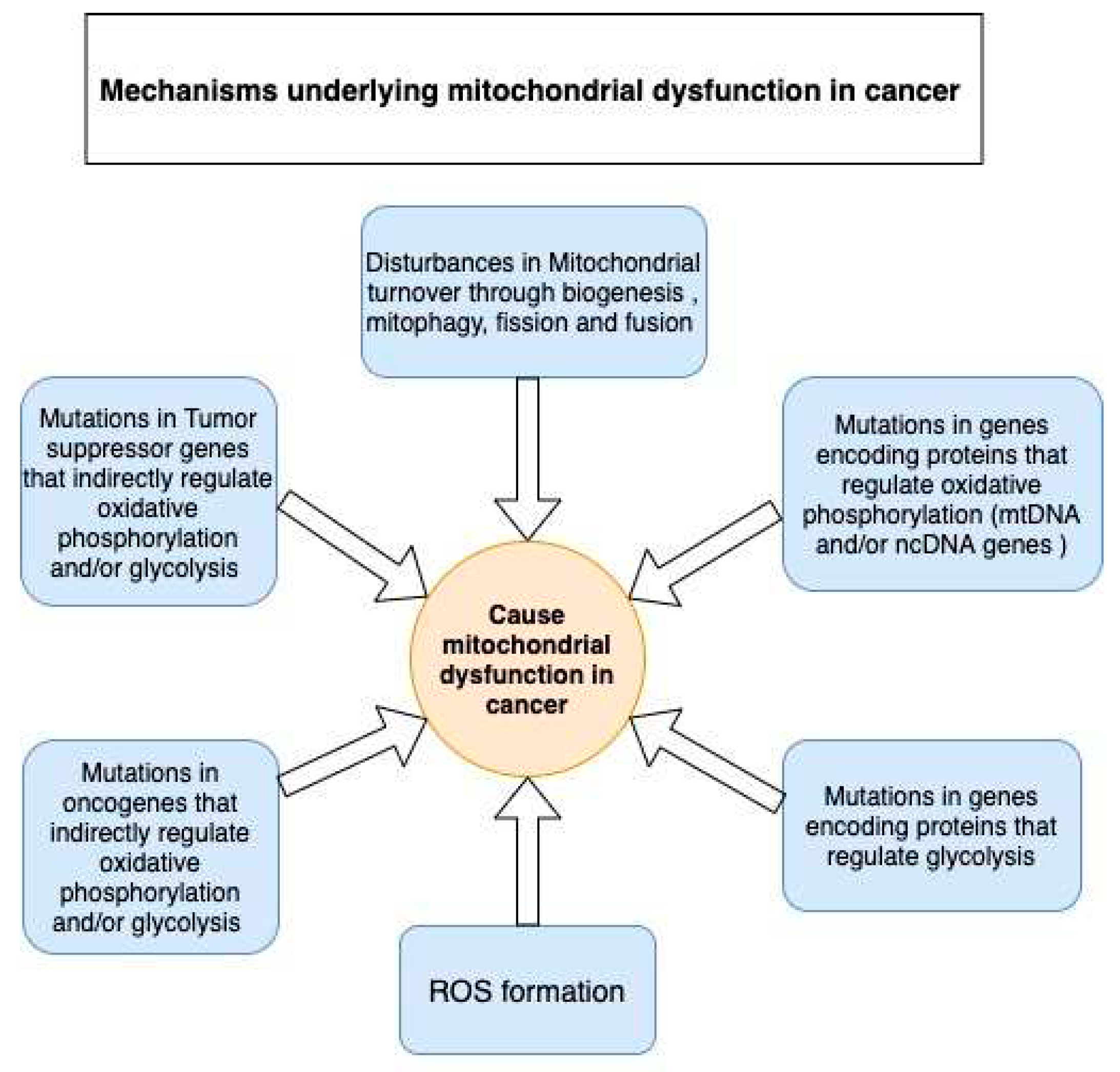
Figure 4.
The activation of AMPK. Abbreviations: AICAR— 5-aminoimidazole-4-carboxamide-1-β-D-ribofuranoside; AMP—adenosine monophosphate; AMPK—AMP-activated protein kinase; ATP—adenosine triphosphate; FFA—free fatty acids; LKB1—liver kinase B1; Thr172—threonine 172. Created with Biorender.com.
Figure 4.
The activation of AMPK. Abbreviations: AICAR— 5-aminoimidazole-4-carboxamide-1-β-D-ribofuranoside; AMP—adenosine monophosphate; AMPK—AMP-activated protein kinase; ATP—adenosine triphosphate; FFA—free fatty acids; LKB1—liver kinase B1; Thr172—threonine 172. Created with Biorender.com.
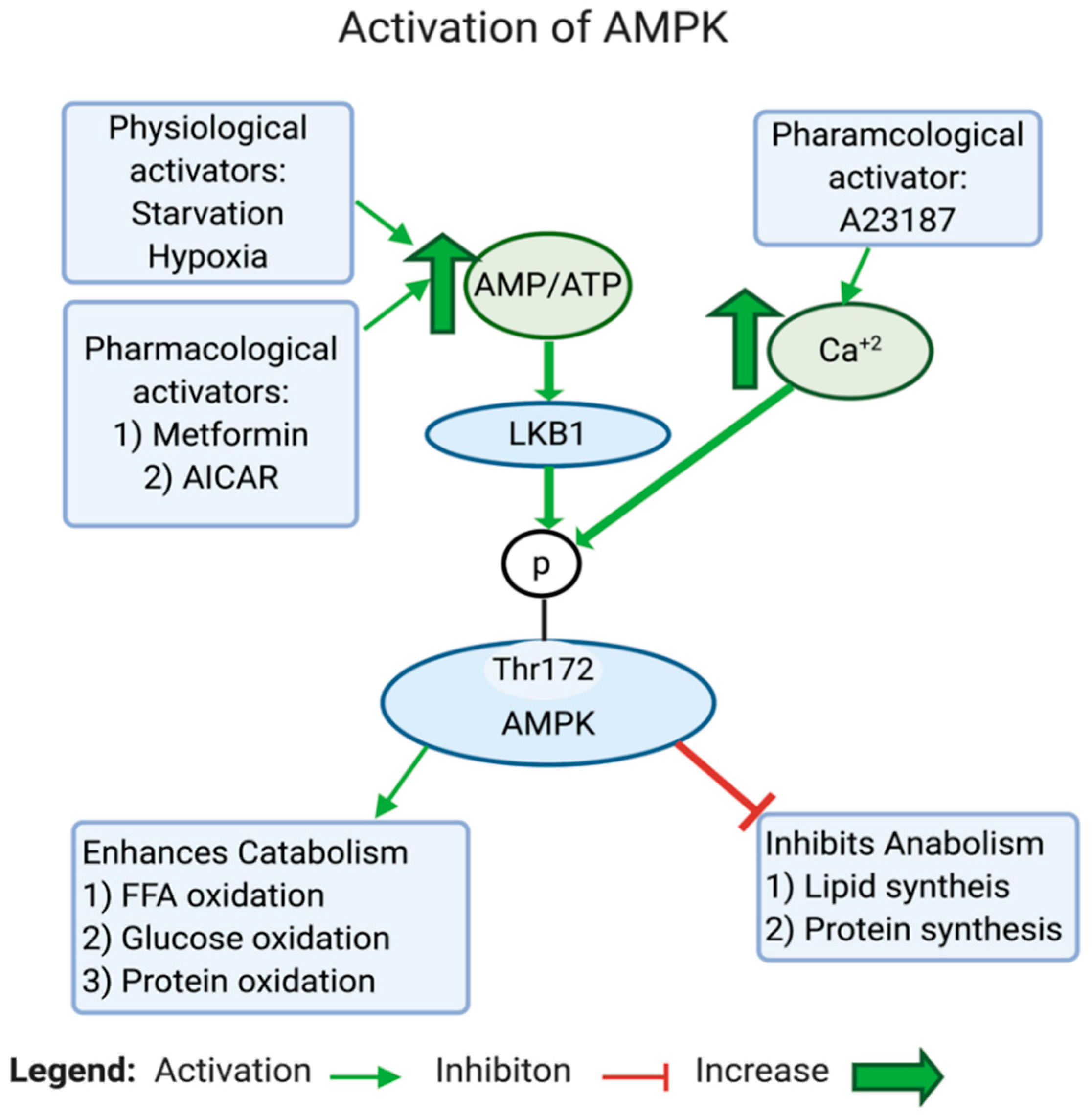
Figure 5.
The effects of AMPK inhibition on the metabolic switch in cancer. The inhibition of AMPK promotes the shift towards the Warburg effect away from OXPHOS. Abbreviations: AMPK—AMP-activated protein kinase; HIF-1 alpha—hypoxia-inducible factor-1 alpha; mTOR—mammalian target of rapamycin; OXPHOS—oxidative phosphorylation; TSC2—tuberous sclerosis complex 2; ULK1—Unc-51 like autophagy activating kinase 1. Created with Biorender.com.
Figure 5.
The effects of AMPK inhibition on the metabolic switch in cancer. The inhibition of AMPK promotes the shift towards the Warburg effect away from OXPHOS. Abbreviations: AMPK—AMP-activated protein kinase; HIF-1 alpha—hypoxia-inducible factor-1 alpha; mTOR—mammalian target of rapamycin; OXPHOS—oxidative phosphorylation; TSC2—tuberous sclerosis complex 2; ULK1—Unc-51 like autophagy activating kinase 1. Created with Biorender.com.
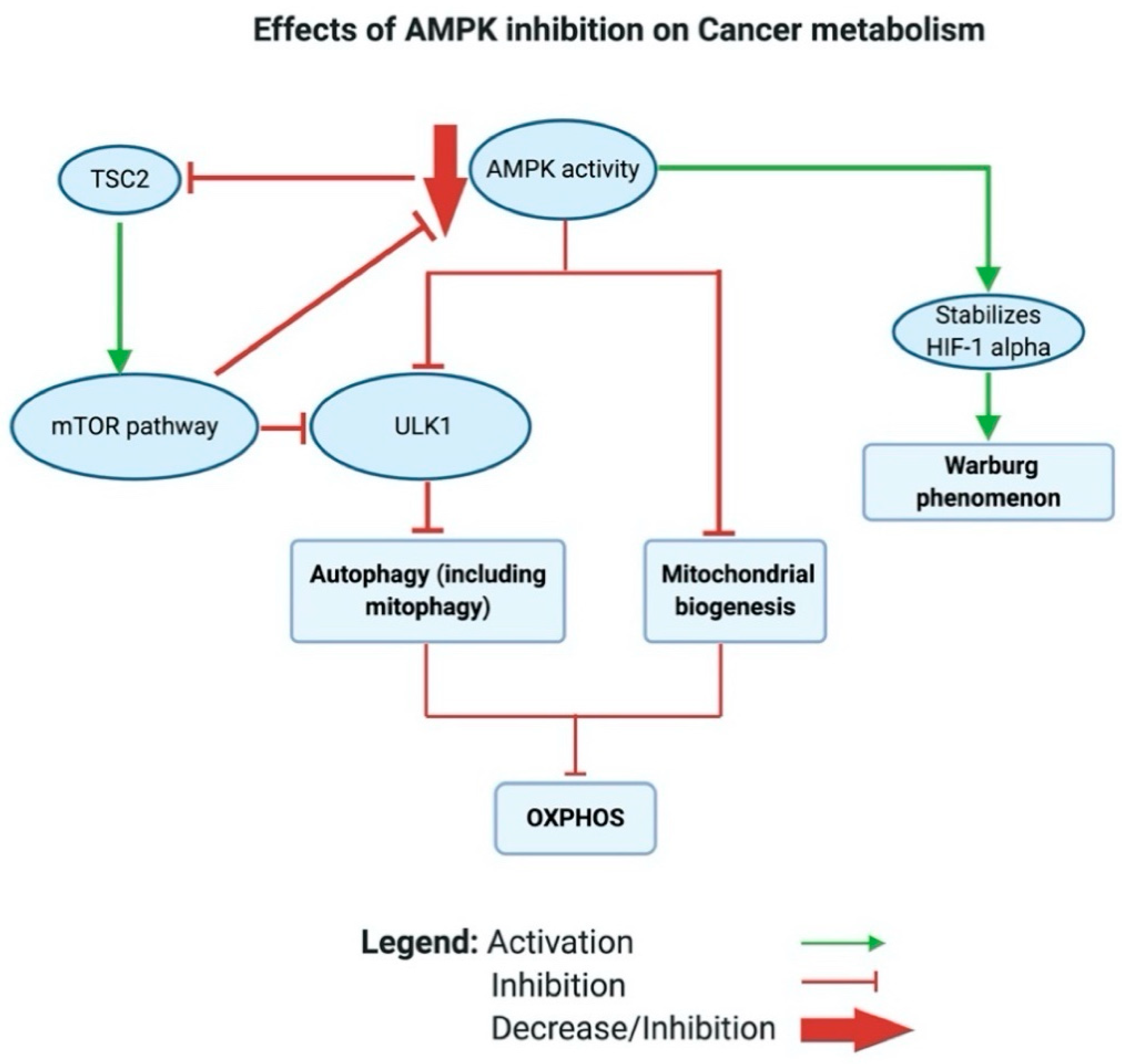
Figure 6.
Effect of low AMPK activity in T2D. The main metabolic alterations caused by low AMPK activity in T2D, which promotes insulin resistance.Abbreviations: AMPK—AMPK-activated protein kinase; FFA—free fatty acid; GLUT1—glucose transporter 1; GLUT4—glucose transporter 4. Created with Biorender.com.
Figure 6.
Effect of low AMPK activity in T2D. The main metabolic alterations caused by low AMPK activity in T2D, which promotes insulin resistance.Abbreviations: AMPK—AMPK-activated protein kinase; FFA—free fatty acid; GLUT1—glucose transporter 1; GLUT4—glucose transporter 4. Created with Biorender.com.
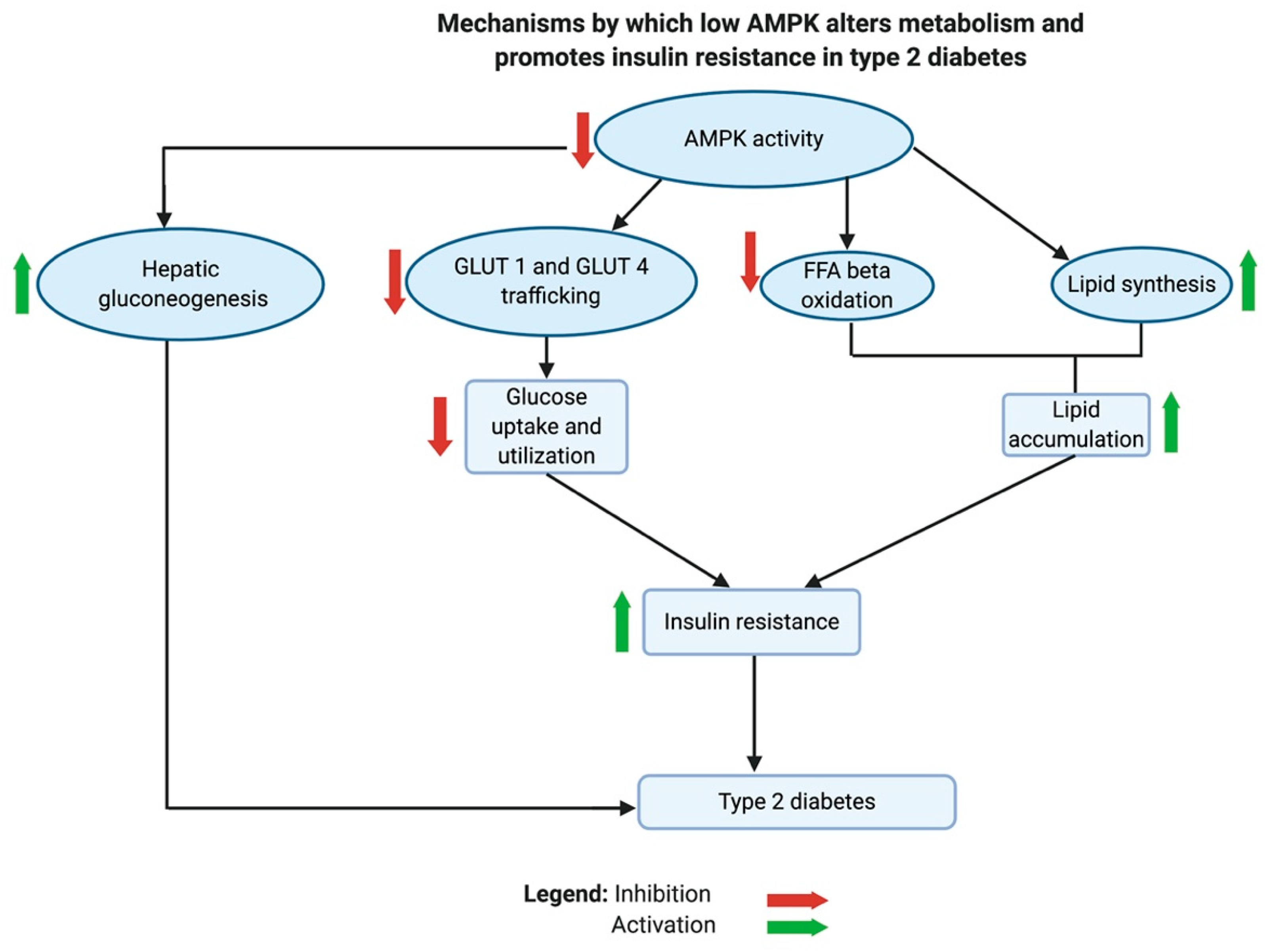
Table 4.
Summary of a group of flavonoids reported to reverse Warburg glycolysis towards mitochondrial respiration in preclinical studies.
Table 4.
Summary of a group of flavonoids reported to reverse Warburg glycolysis towards mitochondrial respiration in preclinical studies.
| Flavonoid’s name | Flavonoid subfamily | Mechanism of targeting Warburg glycolysis | Warburg glycolytic target | References |
|---|---|---|---|---|
| Apegenin | Flavones |
|
PKM2 | [322] |
| epigallocatechin-3-gallate (EGCG) | Flavan-3-ols |
|
PKM2 | [323] |
|
HK2 HIF-1a |
[323] [323] |
||
| Proanthocyanidin B2 (PB2) | Anthocyanidins |
|
PKM2 | [324] |
| Shikonin (SHI) | Naphthoquinone flavonoid |
|
PKM2 | [325,326] |
| Quercetin (QUE) | Flavonol |
|
PKM2 | [327] |
|
HK2 | [327] | ||
|
LDH | [327] | ||
| Xanthohumol (XA) | Prenylated Flavonoid |
|
HK2 | [328] |
| 10v | Synthetic flavonoid |
|
HK2 | [329] |
| GL-V9 | Synthetic flavonoid |
|
HK2 | [330] |
| FV-429 | Synthetic flavonoid |
|
HK2 | [331] |
| Gen-27 | Synthetic flavonoid |
|
HK2 | [332] |
| Astragalin (ASG) | O-glycoside flavonoid |
|
HK2 | [333] |
| Morin (MO) | Flavonol |
|
LDH | [334] |
| Methylalpinumisoflavon (MF) | Isoflavone |
|
HIF-1α | [335] |
| Oroxylin A (OX-A) | Flavone |
|
HIF-1α | [336] |
| Baicalein (BA) | Flavone |
|
HIF-1α | [337] |
| Wogonin | O-methylated flavone |
|
HIF-1α | [338] [339] |
| Berberine (BBR) | Isoquinoline flavonoid |
|
HK2 | [340] [341] |
| Resveratrol |
|
HK2 PDH complex |
[342] [343] [344] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



