
Submitted:
12 December 2023
Posted:
13 December 2023
You are already at the latest version
Abstract
Cannabis, renowned for its historical medicinal use, harbours various bioactive compounds—cannabinoids, terpenes, and flavonoids. While major cannabinoids like THC and CBD have received extensive scrutiny for their pharmacological properties, emerging evidence underscores the collaborative interactions among these constituents, suggesting a collective therapeutic potential. This comprehensive review explores the intricate relationships and synergies between cannabinoids, terpenes, and flavonoids in cannabis. Cannabinoids, pivotal in cannabis's bioactivity, exhibit well-documented analgesic, anti-inflammatory, and neuroprotective effects. Terpenes, aromatic compounds imbuing distinct flavours, not only contribute to cannabis's sensory profile but also modulate cannabinoid effects through diverse molecular mechanisms. Flavonoids, another cannabis component, demonstrate anti-inflammatory, antioxidant, and neuroprotective properties, particularly relevant to neuroinflammation. The entourage hypothesis posits that combined cannabinoid, terpene, and flavonoid action yields synergistic or additive effects, surpassing individual compound efficacy. Recognizing the nuanced interactions is crucial for unravelling cannabis's complete therapeutic potential. Tailoring treatments based on the holistic composition of cannabis strains allows optimization of therapeutic outcomes while minimizing potential side effects. This review underscores the imperative to delve into the intricate roles of cannabinoids, terpenes, and flavonoids, offering promising prospects for innovative therapeutic interventions and advocating continued research to unlock cannabis's full therapeutic potential within the realm of natural plant-based medicine.

Keywords:
Neuroinflammatory
; Neuroprotective diseases
; Phytochemicals
; Cannabis Sativa
; Entourage effects
; Synergistic effects
; Terpenes
; Flavonoids
; Cannabinoids
Introduction
Cannabis, known by various names such as marijuana, ganja, hashish, pot, and hemp, is an ancient plant cultivated and exploited for its various properties. It is a versatile plant, being used as a fibre source, food ingredient, and medicinal substance (Pisanti & Bifulco, 2019). This annual flowering herb can be classified into three primary species: Cannabis sativa, which is taller and more fibrous, and Cannabis indica, which is shorter and more psychoactive. Both species exist in both wild and cultivated forms. Additionally, some taxonomists propose including a third putative species, Cannabis ruderalis, which is solely wild (Pisanti & Bifulco, 2019).
Cannabis is a genus within the Cannabaceae plant family, including hops. A defining characteristic of all Cannabis plants is the presence of secondary substances called cannabinoids or phytocannabinoids (Schilling et al., 2020). At the same time, the genus comprises three species C. sativa, C. ruderalis, and C. indica. C. sativa is the most extensively studied species in terms of its medicinal potential, unlike C. ruderalis and C. indica, which require further elucidation regarding their therapeutic properties (Schilling et al., 2020).
C. sativa holds significant value as a medicinal plant and has garnered increasing interest in the research and manufacturing sectors. To date, over 150 cannabinoids and numerous other compounds, including terpenoids, flavonoids, and alkaloids, have been identified in C. sativa (Bonini et al., 2018; Hanuš et al., 2016; Jin et al., 2020). Many traditional medicinal uses of C. sativa have been studied (Jin et al., 2020). Furthermore, cannabis has historically been employed in treating various ailments, including pain, inflammation, and mental illnesses. However, it is important to note that discrepancies in terminology between historical texts and modern scientific literature, as well as potential nuances lost in translation between Chinese and English, may exist (Jin et al., 2020).
Combining biomechanics research and investigations into the therapeutic effects of specific substances can facilitate the development of applications utilising different plant parts (Amar, 2006). For instance, 9-THC, known for its antiemetic and appetite-stimulating properties, has been utilised in approved medications such as Marinol (dronabinol, synthetic 9-THC) and Cesamet to address chemotherapy-induced nausea or vomiting and anorexia associated with AIDS-related weight loss (nabilone, a THC derivative) (Amar, 2006).
Neuroinflammation is a multifaceted response in the brain following injury, involving the activation of glial cells, the release of inflammatory mediators like cytokines and chemokines, and the production of reactive oxygen and nitrogen species (Agostinho et al., 2010; Mosley et al., 2006). Although it is considered a secondary event to neuronal dysfunction or death, neuroinflammation plays a significant role in the onset and progression of neurodegenerative diseases such as Alzheimer's Disease (AD), Parkinson's Disease (PD), Multiple Sclerosis (MS), Chronic Traumatic Encephalopathy (CTE) (Agostinho et al., 2010; Mosley et al., 2006). Due to the limited efficacy of current treatments for these conditions, neuroinflammation has emerged as a promising therapeutic target in drug discovery (Agostinho et al., 2010; Mosley et al., 2006). Consequently, various in vivo and in vitro models of neuroinflammation have been developed to study its mechanisms and potential interventions.
2. Phytochemicals in medicinal cannabis
C. sativa contains a wide range of phytocannabinoids, which are oxygenated aromatic hydrocarbons derived from meroterpenoids with various substitutions in the resorcinol core (Figure 1) (Gülck et al., 2020; Hanuš et al., 2016). These phytocannabinoids often have alkyl side chains with an odd number of carbon atoms and are initially produced in their acid form (Figure 1). Through decarboxylation, they are converted into their active forms (Gülck et al., 2020). The two most abundant phytocannabinoids in C. sativa are cannabidiols (CBD) and Δ9-trans-tetrahydrocannabinol (Δ9-THCs) (Figure 1). Additionally, cannabigerol (CBG) and its acid form (CBGA) serve as core intermediates and provide phytocannabinolic acids (Figure 1) (Hanuš et al., 2016; Tahir et al., 2021).
Terpenes, which are the second-largest class of cannabis constituents after phytocannabinoids, are also present in C. sativa and many other non-cannabinoid plants such as tea, thyme, Spanish sage, and citrus fruits (ElSohly et al., 2017). The major terpenes in C. sativa include myrcene, alpha-pinene, linalool, and limonene (ElSohly et al., 2017). In addition, C. sativa also biosynthesises flavonoids, including cannflavins, which are prenylated (C5) and geranylated (C10) flavones (Ubeed et al., 2022).
Figure 1.
Chemical structures and molecular formula of the main selected (a) cannabinoids, (b) terpenes, and (c) prenylflavonoids in C. sativa.
Figure 1.
Chemical structures and molecular formula of the main selected (a) cannabinoids, (b) terpenes, and (c) prenylflavonoids in C. sativa.
3. The endocannabinoid system and neuroinflammation
Neuroinflammation refers to a broad spectrum of immune responses in the central nervous system that stem from peripheral inflammation (Lyman et al., 2014). Key cellular players in this process include microglia and astrocytes, which are primary cells involved in the immune reactions within the central nervous system (Lyman et al., 2014). The activation of a neuroinflammatory response occurs due to peripheral inflammation affecting various components, such as the blood-brain barrier (BBB), glial cells, and neurons (Lyman et al., 2014). Previously, it was widely believed that BBB, a specialised type of endothelium, ultimately separated the central nervous system from the peripheral immune system (Lyman et al., 2014). However, it has been discovered that the BBB can become permeable to pro-inflammatory molecules generated during peripheral inflammation and facilitate their release and transport into the brain (De Vries et al., 1996; Laflamme & Rivest, 1999). This neuroinflammatory reaction leads to synaptic dysfunction, neuronal loss, and exacerbation of various brain disorders (Kitazawa et al., 2005; Marquette et al., 1996; Micheau & Tschopp, 2003).
Microglial cells are a crucial component of the central nervous system (CNS) immune defence and maintenance of homeostasis (Aloisi, 2001; Cardona et al., 2006; Filiano et al., 2015; Wirenfeldt et al., 2011). They act as resident macrophages, responding to pathogenic invasion, tissue damage, and protein aggregates by recognising danger-associated molecular patterns (DAMPs) or pathogen-associated molecular patterns (PAMPs) through specific receptors (Filiano et al., 2015; Kettenmann et al., 2011; Wirenfeldt et al., 2011).
Microglia can migrate to the injury site and initiate an innate immune response when activated (Ji et al., 2013). Additionally, they play a critical role in preserving synaptic plasticity and contribute significantly to learning and memory processes by modifying synapses associated with learning (Parkhurst et al., 2013). Recent advances in single-cell RNA sequencing have revealed a distinct subtype of microglia known as disease-associated microglia (DAM), which has been implicated in the progression of Alzheimer's disease (AD) (Keren-Shaul et al., 2017). The blood-brain barrier (BBB), consisting of tight junctions between brain endothelial cells, restricts the entry of pathogenic microorganisms into the CNS. However, certain head injuries or infections can significantly change brain function and behaviour. Inflammatory responses involving pro-inflammatory cytokines are observed when brain tissue is damaged or infected, and microglial activation plays a key role in this process (Konsman, 2022).
In neurodegenerative diseases, microglia are associated with neuroinflammation by activating cell surface receptors, such as toll-like receptors (TLRs), scavenger receptors, and NLRP3 inflammasome (Bamberger et al., 2003; El Khoury et al., 2003; Fassbender et al., 2004; Kagan & Horng, 2013; Stewart et al., 2010). Impaired microglial phagocytic ability and reduced amyloid-beta (Aβ) clearance are observed in these conditions, characterised by altered expression of Aβ phagocytosis receptors and elevated cytokine levels. The dysregulation of immune receptors, such as TREM2 and CD33, further highlights the significant role of neuroinflammation in neurodegenerative diseases (Bradshaw et al., 2013; Griciuc et al., 2013; Guerreiro et al., 2013; Heneka et al., 2013; Hickman et al., 2008; Liu & Jiang, 2016; Mawuenyega et al., 2010; Saresella et al., 2016; Sheedy et al., 2013).
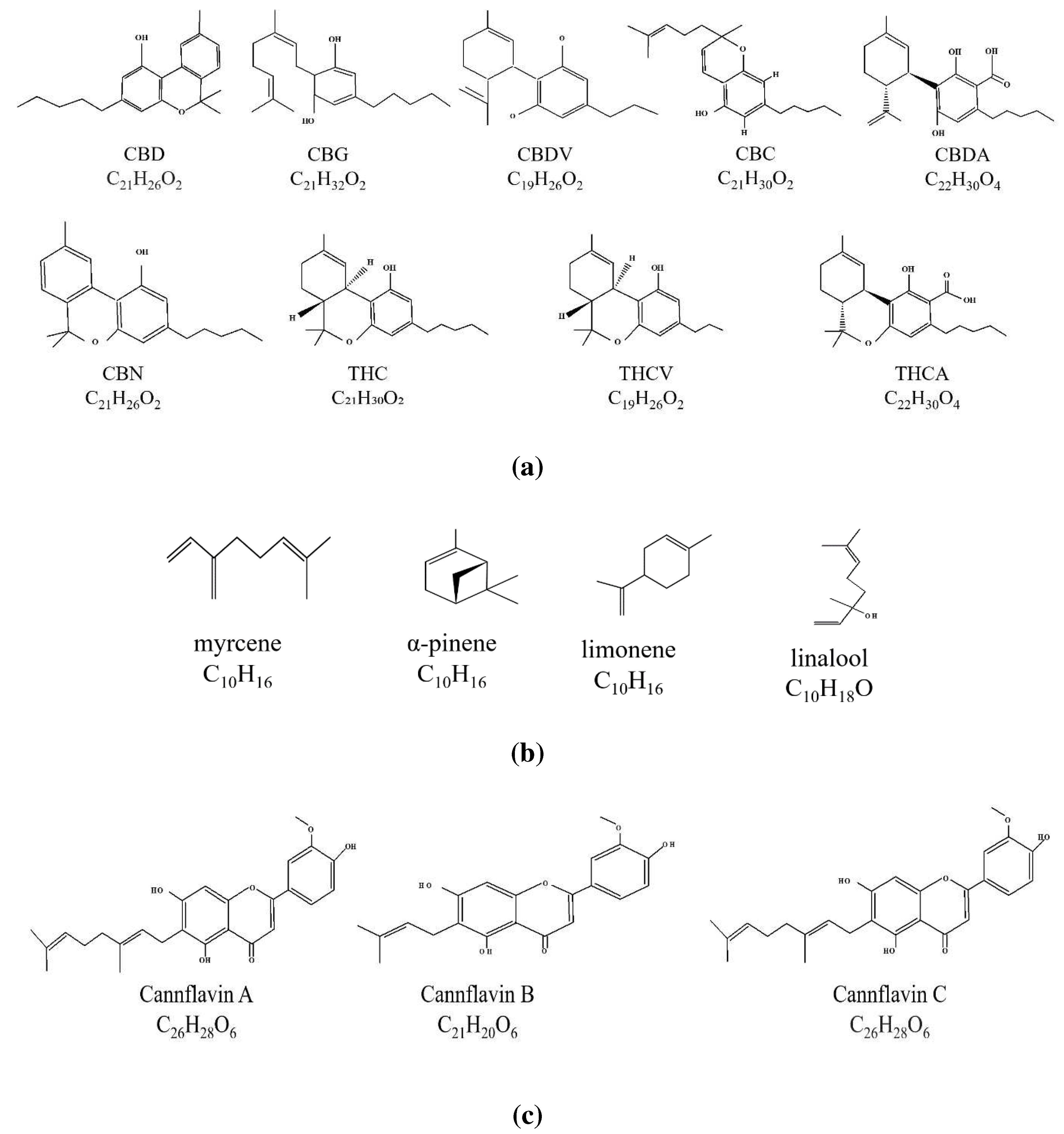
Tetrahydrocannabinol (THC) and cannabidiol (CBD) are phytocannabinoids in C. sativa. They exert their effects on neuroinflammation primarily through activating CB1 and CB2 cannabinoid receptors (Figure 2) (Baker et al., 2007). In addition to these receptors, the endocannabinoid system (ECS) includes proteins involved in the synthesis, inactivation and other endocannabinoid molecular targets. Key components of the ECS include endogenous ligands such as arachidonyl ethanolamide (AEA) and 2-arachidonylglycerol (2-AG), which are derivatives of the polyunsaturated fatty acid arachidonic acid (Piomelli et al., 1998).
CB1 receptors are predominantly found in the central nervous system, while CB2 receptors are primarily expressed peripherally in lymphoid organs, peripheral blood leukocytes, mast cells, and to a lesser extent in the pancreas(Howlett et al., 2019; Sinha et al., 1998). CB1 mRNA and protein expression have been observed in various immune cells, including B cells, NK cells, neutrophils, CD8+ T cells, monocytes, and CD4+ T cells, albeit in decreasing order, whereas CB2 is expressed at higher levels in these immune cells, approximately 10-100 times more than CB1 (Galiègue et al., 1995). Given their widespread expression in the immune system, these receptors may play a crucial role in immunomodulation.
Endocannabinoids, such as 2-AG and AEA, are produced in large quantities by microglia, macrophages, astrocytes, and neurons during inflammation. These endocannabinoids bind to CB receptors and have been shown to reduce neuronal damage by protecting the nervous system from excitotoxicity (Figure 2) (Table 1) (Eljaschewitsch et al., 2006; Marinelli et al., 2008; Walter et al., 2004; Walter et al., 2002).
Figure 2.
The mechanisms of activating CBD receptors through the endocannabinoid system and phytocannabinoid on neuroinflammation.
Figure 2.
The mechanisms of activating CBD receptors through the endocannabinoid system and phytocannabinoid on neuroinflammation.
Furthermore, cannabinoid treatment has been demonstrated to attenuate the inflammatory effects of IL-1 and protect glial cells from death (Aguado et al., 2006; Sheng et al., 2005). Overall, the cannabinoid system plays a protective role by combating CNS excitotoxicity and neuroinflammation. The evidence primarily supports the anti-inflammatory benefits of cannabis, although some studies suggest potential pro-inflammatory effects, creating a more nuanced understanding (Killestein et al., 2003; Maestroni, 2004).
The endocannabinoid system (ECS) modulates multiple physiological processes within the nervous system, and dysregulation of ECS has been associated with various pathological conditions, including neuroinflammation (Di Marzo & Piscitelli, 2015; Hillard, 2018). Therapeutic modulation of ECS activity has shown beneficial effects on medical conditions related to neuroinflammation (Ambrose & Simmons, 2019; Giacobbe et al., 2021). The ECS comprises multiple receptors, including peroxisome proliferator-activated receptors (PPARs) and ion channels (such as the transient receptor potential ankyrin [TRPA] family and the transient receptor potential vanilloid [TRPV] family), as well as cannabinoid receptor types 1 and 2 (CB1 and CB2, respectively) (Biringer, 2021). The ECS also involves endocannabinoids derived from arachidonic acid, receptor ligands, and enzymes responsible for endocannabinoid metabolism (Di Marzo & Piscitelli, 2015).
Endocannabinoids, the enzymes involved in their biosynthesis and degradation, and endocannabinoid receptors are expressed by most immune cells (Chiurchiù et al., 2015). CB1 and CB2 receptors are present in immune cells, with CB2 being expressed at higher levels than CB1 (Jean-Gilles et al., 2010; Rahaman & Ganguly, 2021). Activation of CB receptors regulates anti-inflammatory responses, as evidenced by increased release of the anti-inflammatory cytokine IL-10 and decreased release of pro-inflammatory cytokines IL-12 and IL-23 upon CB2 receptor activation in activated macrophages (Figure 2) (Correa et al., 2009; Correa et al., 2010). The CB2 receptor system has also been implicated in anxiety, depression, and substance abuse, suggesting its involvement in modulating dopamine reward pathways (Al Mansouri et al., 2014; Gertsch et al., 2008; Xi et al., 2011). Trans-caryophyllene has demonstrated neuroinflammatory inhibition and lipid regulation properties (Zhang et al., 2017).
4. Anti-neuroinflammatory activity of phytochemicals in C. Sativa
4.1. CBD
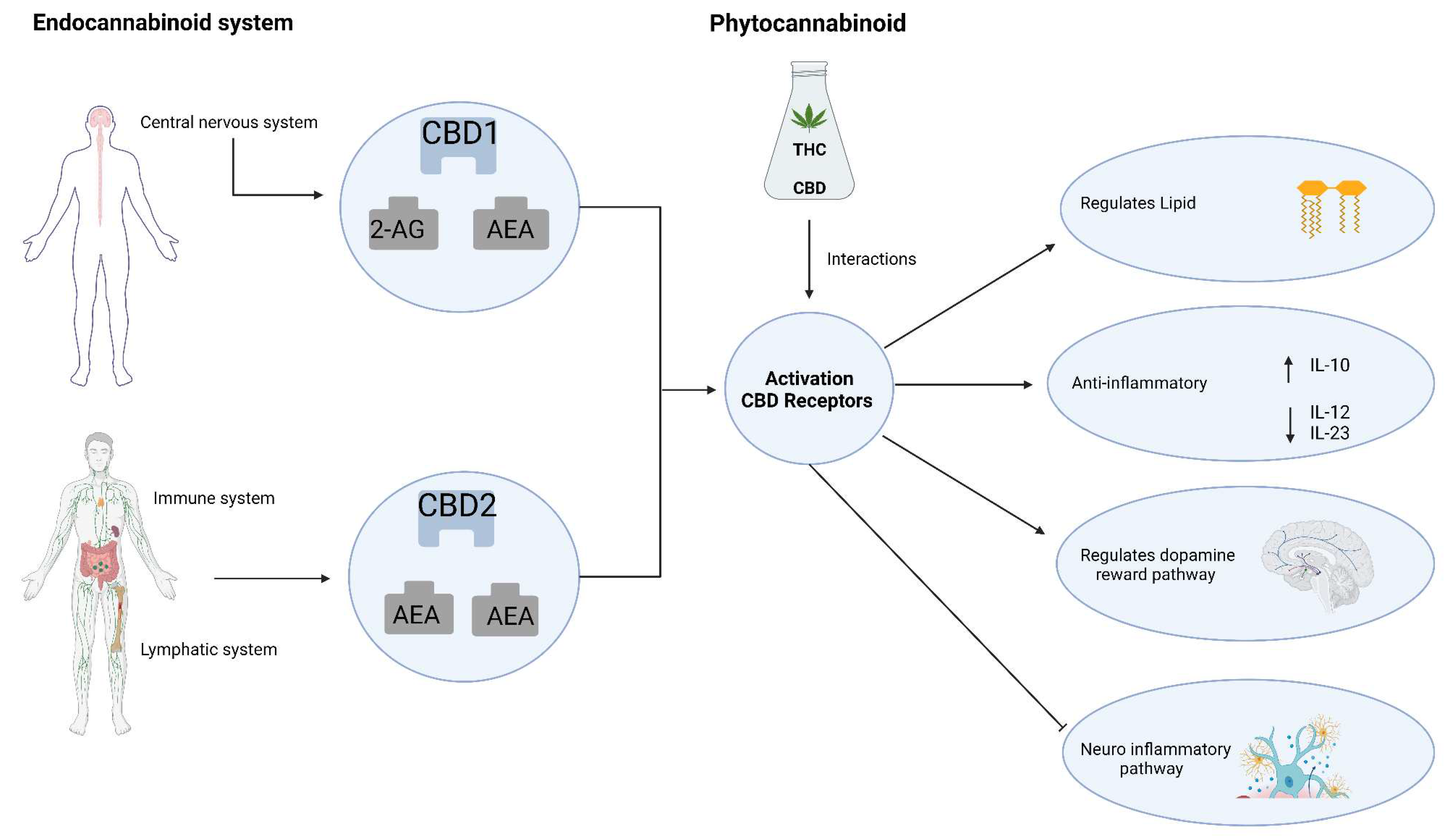
CBD has been extensively studied for its potential anti-neuroinflammatory properties in various in vitro and in vivo models of degenerative diseases (Table 1). However, the precise mechanism underlying its anti-neuroinflammatory activity still needs to be understood. In the context of hypoxic-ischemic (HI) immature brains in newborn mice, CBD treatment was found to significantly decrease the expression of inflammatory markers such as IL-6, TNF-α, COX-2, and iNOS in brain slices (Figure 3). It has been suggested that this effect may be mediated through the CB2 and adenosine A2A receptors (Castillo et al., 2010). Similarly, low doses of CBD were observed to reduce TNF-α production in mice treated with lipopolysaccharide, and this effect was abolished in mice lacking the A2A receptor and restored by an A2A adenosine receptor, indicating a potential modulation of adenosine signalling by CBD (Carrier et al., 2006).
Furthermore, CBD selectively inhibits GPR55, another G protein-coupled receptor in human macrophages. In microglial cells isolated from the retinas of newborn rats treated with endotoxin or LPS for acute ocular inflammation, CBD treatment inhibited TNF-α production via the p38 MAPK pathway. In rat retinas exposed to LPS, CBD administration prevented the development of macrophage accumulation, activated microglia, increased levels of reactive oxygen species (ROS) and nitrotyrosine, activated p38 MAPK, and neuronal apoptosis (Figure 3) (El-Remessy et al., 2008).
In LPS-activated microglial cells (BV-2 cells), CBD has been shown to reduce the production and release of inflammatory cytokines such as IL-1, IL-6, and IFN-β. This reduction is associated with a decrease in the activity of the NF-κB pathway and the levels of IL-1β and IL-6. Additionally, CBD downregulates the expression of the SOCS3 gene, which regulates cytokine and hormone signalling. CBD treatment leads to increased phosphorylation of the STAT3 transcription factor, which is required for activation. In contrast, CBD decreases the phosphorylation of STAT1, a transcription factor involved in IFN-β-dependent pro-inflammatory processes (Carow & Rottenberg, 2014; Kozela et al., 2010). NF-κB and STAT3 have important and sometimes overlapping roles in pro-inflammatory responses, while STAT1 plays a significant role in IFN-β-mediated inflammation (Carow & Rottenberg, 2014; Kozela et al., 2010).
Figure 3.
The anti-neuroinflammatory activity of CBD.
4.2. THC
Since its synthesis in 1964, delta-9-tetrahydrocannabinol (THC) has been the most extensively studied phytocannabinoid, primarily due to its pharmacological effects. THC primarily interacts with the endocannabinoid receptors CB1 and CB2, acting as a partial agonist at sub-micromolar doses. These receptors have been the focus of considerable research in understanding the psychoactive effects of THC. The development of synthetic high-affinity counterparts has facilitated the identification of the endocannabinoid system and its central nervous system targets (Gaston & Friedman, 2017). The metabolic precursor of THC, delta-9-tetrahydrocannabinolic acid (THCA), is present in high concentrations in cannabis plants. Upon drying or burning, THCA is decarboxylated to THC. THCA is believed to have less psychoactive properties than THC (Gaston & Friedman, 2017). However, at concentrations exceeding 10 µM, THC inhibits cyclooxygenases-1 and 2, as well as diacylglycerol lipase alpha (DLG), an essential enzyme in the biosynthesis of the endocannabinoid 2-arachidonoylglycerol (2-AG). In vitro, experiments have shown activation of TRPA1 and TRPV4 channels, while TRPM8 channels are blocked at low micromolar concentrations (Gaston & Friedman, 2017).
Another cannabinoid present in varying levels of cannabis is delta-9-tetrahydrocannabivarin (THCV). Similar to THC, THCV acts as a partial agonist of CB1/2 receptors and exhibits activity on GPR55, TRPA1, and TRPV1-4 receptors at sub-micromolar or low micromolar doses (Pertwee & Cascio, 2014). In vitro and in vivo animal models have demonstrated the anti-seizure effects of THCV in one study (Hill et al., 2010).
THC has been shown in numerous studies to possess anti-neuroinflammatory properties (Table 1). For instance, it increases the production of anti-inflammatory cytokines while decreasing pro-inflammatory cytokine production in multiple sclerosis (MS). THC also promotes apoptosis in T cell-driven inflammation and increases the population of FoxP3+ regulatory T cells through miRNA induction and epigenetic modifications (Figure 4) (Rao et al., 2014; Sido et al., 2016).
Figure 4.
The Anti-neuroinflammatory activity of THC.

Moreover, THC has been found to inhibit acetylcholine esterase (AchE)-induced aggregation of amyloid-beta (Aβ), improve motor coordination deficits in R6/2 mice, mitigate striatal atrophy and huntingtin aggregate accumulation, and exacerbate malonate lesions in Alzheimer's disease (AD) (Table 1) (Blazquez et al., 2011; Dowie et al., 2010; Eubanks et al., 2006; Lastres-Becker et al., 2003). THC, THCA, and the metabolite cannabinol (CBN) have been described to possess analgesic, anti-inflammatory, and neuroprotective effects (Baron, 2018; De Petrocellis et al., 2011; Pugazhendhi et al., 2021).
4.3. CBG
While there is still a need for further research on the anti-neuroinflammatory effects of cannabigerol (CBG) compared to cannabidiol (CBD), several studies have discussed the neuroprotective properties of CBG against neuroinflammation (Table 1). For instance, in cultured motor neurons, CBG pretreatment was found to reduce the levels of pro-inflammatory cytokines such as IL-1β, TNFα, and IFN-γ, and prevent apoptosis in LPS-stimulated macrophages by inhibiting the expression of caspase-3 and Bax, while increasing Bcl-2 levels (Gugliandolo et al., 2018). Similarly, in an in vivo study using a 3-nitro propionate model to examine the effects of CBG on Huntington's disease pathology, treatment with CBG significantly attenuated the upregulation of COX-2, iNOS, and pro-inflammatory cytokines such as TNF-α and IL-6 (Figure 5) (Valdeolivas et al., 2015).
Figure 5.
The Anti-neuroinflammatory activity of CBG.
These findings highlight the potential of CBG as a neuroprotective agent against neuroinflammation, but further investigation is necessary to understand its mechanisms and therapeutic potential fully.

Table 1.
A summary of preclinical evidence of cannabinoids on microglial activation and neuroinflammatory signalling.
Table 1.
A summary of preclinical evidence of cannabinoids on microglial activation and neuroinflammatory signalling.
| Compound | Model | Concentration/ Dose |
indicated neurodegenerative diseases | Outcome | References |
|---|---|---|---|---|---|
| CBD | in vitro glutamate neuronal toxicity model | N/A | N/A | CBD was shown to be more protective than either α-tocopherol or vitamin C and comparable to butylated hydroxytoluene (BHT). | (Saito et al., 2012; Yousaf et al., 2022) |
| THC | in vivo in hemiparkinsonian rats | N/A | PD | neuroprotective effect | (Lastres-Becker et al., 2005) |
| CBD | in vivo in hemiparkinsonian rats | 3 mg/kg | PD | exhibited a potent neuroprotective effect in this rat model | (Lastres-Becker et al., 2005) |
| CBD | N/A | < 1 μM | N/A | inhibit activated microglial cell migration by antagonising the abnormal-cannabidiol (Abn-CBD)-sensitive receptor | (Walter et al., 2003) |
| CBD |
in vitro PC12 cells |
N/A | AD | Neuroprotective against the neuronal damage induced by the β-amyloid peptide (Aβ) inhibits Aβ-induced neurotoxicity. | (Esposito et al., 2006) |
| CBD |
In vivo mouse model |
N/A | AD | Attenuated the expression of several glial pro-inflammatory proteins, including glial fibrillary acidic protein, inducible nitric oxide synthase (iNOS) and interleukin 1β (IL-1β), which are major contributors to the propagation of neuroinflammation and oxidative stress. | (Esposito et al., 2007) |
| CBD |
in vivo mouse model |
100-200 mg/kg | Dravet Syndrome | It has beneficial effects on seizures and social deficits | (Kaplan et al., 2017) |
| CBD |
in vivo mouse model |
10 mg/kg twice- daily |
schizophrenia | improves the social and cognitive dysfunctions | (Osborne et al., 2017) |
| CBDV | Clinical trial | Single oral dose | ASD | It modulates glutamatergic but not γ-aminobutyric acid (GABA) neurotransmission in adult male patients, although the biological response may differ between autistic individuals. | (Pretzsch et al., 2019) |
| THCV |
in vivo mouse model |
<3 mg/kg | PD | alleviates motor inhibition in 6-OHDA-lesioned rodents by blocking CB1 receptors at low doses | (Espadas et al., 2020) |
| THC | N/A | N/A | PD | It reduced levodopa-induced dyskinesia | (Cristino et al., 2020) |
| CBN |
in vitro C6 glioma cells |
0.3–30000 nM EC50: 700 nM |
N/A | It inhibited NO production and iNOS expression | (Esposito et al., 2001) |
| CBN | N/A | N/A | MS | It may antagonise the 2-AG-induced recruitment of microglial cells and produces minimal palliative. | (Walter et al., 2003) |
| THC |
in vitro BV-2 murine microglial cell line |
10 μM | N/A | It decreases the production and release of proinflammatory cytokines, including interleukin-1β, interleukin-6, and interferon (IFN)β, from LPS-activated microglial cells. | (Kozela et al., 2010) |
| CBG | in vitro murine microglial cell line |
25 μM | MS | It has inhibited the microglia-driven inflammatory response, protected neurons from toxic insults in vitro, and restored motor function impairment by inhibiting the synthesis of IL-1β, IL-6, TNF-α, the chemokine, MIP-1α and prostaglandin E2 (PGE2). | (Carrillo-Salinas et al., 2014; Granja et al., 2012) |
| CBG |
in vitro NSC-34 motor neurons |
7.5 µM | N/A | CBG pre-treatment has REDUCED IL-1β, TNF-α, IFN-γ and PPARγ protein levels and reduced nitrotyrosine, SOD1 and iNOS protein levels and restored Nrf-2 levels. | (Gugliandolo et al., 2018) |
| CBG |
In vivo and in vitro |
N/A | PD | It shows a neuroprotective against inflammation-driven neuronal damage, acting through the activation of the canonic binding site in PPARγ receptors. |
(García et al., 2018) |
| CBG |
In vivo and In vitro neuroblastoma Neuro-2a (N2a) |
2 g/ 6.319 mM | HD | It has improved motor deficits, reactive astrogliosis and microglial activation, inhibiting the upregulation of proinflammatory markers and improving antioxidant defences in the brain. | (Díaz-Alonso et al., 2016) |
| CBDA |
In vitro Neuro-2a (N2a) cells |
25 μM | HD | CBDA shows potent neuroprotective activity by activating PPARγ with higher potency than their decarboxylated products | (Nadal et al., 2017) |
| CBDA | in vivo | 10 and 30 mg/kg | Dravet syndrome | It has an anticonvulsant against pentylenetetrazol-induced seizures and hyperthermia-induced seizures. | (Anderson et al., 2019) |
| CBDV |
in vivo mouse model |
CBDV | Rett syndrome (RTT), a rare neurological disorder affecting predominantly females | It improves behavioural and functional deficits. | (Hagberg et al., 2002; Ricceri et al., 2013; Vigli et al., 2018; Zamberletti et al., 2019) |
| CBC |
In vitro |
1 μM | N/A | CBC exert potential actions on brain health through effects on adult neural stem cells using whole brain-derived neural stem progenitor cells (NSPCs). | (Shinjyo & Di Marzo, 2013) |
| THC |
In vitro |
10 μM | N/A | THC reduces IL-1β, IL-6, and TNFα production in LPS-stimulated rat microglial cells. | (Puffenbarger et al., 2000) |
| THC |
In vitro |
0 – 15 μM | AD | It inhibits the enzyme acetylcholinesterase (AChE) and prevents AChE-induced amyloid β-peptide (Aβ) aggregation, which is considered the key pathological marker of Alzheimer's disease. | (Eubanks et al., 2006) |
| THC |
in vivo R6/1 mouse model |
10 mg/kg | HD | It inhibits acetylcholine esterase (AchE)-induced aggregation of Aβ and attenuates the motor coordination deficits of R6/1 mice. | (Dowie et al., 2010) |
| THCA |
In vitro N2a cells |
10 μM IC50 of 0.47 μM |
HD | It has neuroprotective activity by activate PPARγ transcriptional activity. | (Nadal et al., 2017) |
4.4. Terpenes
Terpenes and terpenoids, found in plant resins and essential oils, are significant components responsible for the pharmacological effects of various medicinal plants, including cannabis. Terpenes are hydrocarbons, while terpenoids contain additional functional groups derived from different chemical elements, making them the most abundant class of phytochemicals. In cannabis, there are approximately 200 unique terpenes, focusing on the primary terpenes found in the highest concentrations. These aromatic essential oils contribute to the distinctive aromas, flavours, and characteristics of different cannabis strains (Cox-Georgian et al., 2019; Ludwiczuk et al., 2017).
Terpenes have lipophilic properties and interact with various bodily targets, including neurotransmitter receptors, ion channels in muscles and neurons, G-protein receptors, enzymes, cell membranes, and second messenger systems. They work independently and synergistically with cannabinoids to produce various therapeutic effects. Additionally, terpenes can enhance the permeability of the blood-brain barrier, leading to the development of transdermal cannabinoid patches containing terpenes as permeation agents. They also influence the binding of THC to CB1 receptors, contributing to the analgesic effects of cannabinoids (Baron, 2018).
While terpenes have been associated with health benefits such as analgesia, anxiolytic and antidepressant effects, skin penetration enhancement, cancer chemoprevention, and antimicrobial activities, their anti-neuroinflammatory activities have not been extensively studied. It is important to note that most available data come from preclinical studies conducted using animal models or in vitro experiments. Some reported benefits of specific terpenes are based on studies evaluating whole essential oils or plants, where the specified terpene may be the most abundant constituent. Additionally, the potential therapeutic contributions of minor terpenes should be considered. Among the primary terpenes found in cannabis are -caryophyllene, myrcene, -pinene, humulene, linalool, limonene, terpinolene, terpineol, ocimene, valencene, and geraniol (Abd Rashed et al., 2021; He et al., 2022).
Myrcene is commonly found in aromatic plants such as sweet basil, bay leaves, lemongrass, and mango. It is utilized in the cosmetic industry due to its remarkable anti-inflammatory, analgesic, and anxiolytic properties (Van Cleemput et al., 2009). The analgesic effects of myrcene appear to be mediated through an opioid mechanism, as they were inhibited by naloxone (Lorenzetti et al., 1991). Additionally, myrcene exhibits muscle relaxant, hypnotic, sedative, sleep aid, and antioxidant properties (De-Oliveira et al., 1997).
Alpha-pinene contributes to the distinctive scent of fresh pine needles, conifers, and sage. It is also present in various herbs, including parsley, rosemary, basil, and dill, making it the most prevalent natural terpenes (Noma et al., 2010). Studies have demonstrated its antioxidant activity (Wang et al., 2008) and anti-inflammatory effects in human chondrocytes (Neves et al., 2010; Rufino et al., 2014), suggesting its potential for anti-osteoarthritic activity (Rufino et al., 2014). Alpha-pinene also acts as an acetylcholinesterase inhibitor, enhancing memory and counteracting the short-term memory loss caused by THC (Kennedy et al., 2011).
Extensive research indicates that linalool, a monoterpene, possesses anti-ischemic, antioxidant, and anti-inflammatory properties. It enhances the activities of antioxidant enzymes superoxide dismutase (SOD) and catalase in vitro, inhibits LPS-induced MCP-1 in airway epithelia, scavenges reactive oxygen species (ROS) in neurons after oxygen-glucose deprivation/reoxygenation, and inhibits MCP-1-induced microglia migration. Linalool also protects neurons from glutamate-induced oxidative stress by preventing mitochondrial ROS and calcium synthesis. Furthermore, it can potentially block LPS-induced PGE2 synthesis and NF-κB/TNF-α expression in macrophages and microglia (Downer, 2020).
Limonene, a monoterpene, exhibits significant anti-inflammatory and antioxidant effects both in vitro and in vivo. It reduces IL-1-induced nitric oxide synthesis in human chondrocytes and decreases the production of prostaglandin E2, nitric oxide, and TNF-α/IL-1 in macrophages stimulated with lipopolysaccharide (LPS). Moreover, in animal models of colitis, limonene has been shown to alleviate intestinal inflammation when administered in vivo. It also demonstrates nonprotective effects by targeting COX-2 and nitric oxide, preventing renal injury. Additionally, limonene enhances the activity of antioxidant enzymes superoxide dismutase (SOD), catalase, and glutathione in the central nervous system during cerebral ischemia models, while reducing the generation of IL-1 and reactive oxygen species (ROS), thus exhibiting its antioxidant potential (Downer, 2020).
4.5. Flavonoids
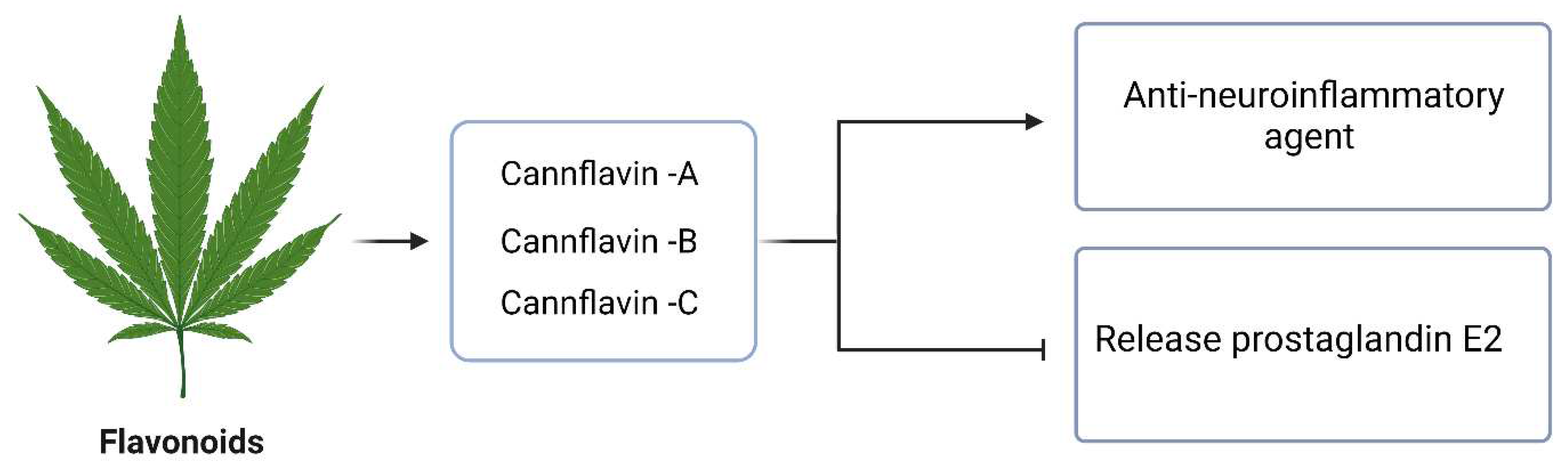
Flavonoids are a class of phenolic compounds characterized by the presence of a phenol ring in their molecular structure. These compounds are known to possess various health benefits, although most of the research conducted so far has been in preclinical models (Andre et al., 2016). Among the flavonoids found in cannabis, three cannflavins, namely cannflavin A (CFL-A), B (CFL-B), and C (CFL-C), have been identified. These cannflavins exhibit promising therapeutic properties, particularly as anti-neuroinflammatory agents (Erridge et al., 2020). In a series of studies conducted in the mid-1980s, Barret et al. investigated the ability of these compounds to inhibit the release of prostaglandin E2 (PGE2) from human rheumatoid synovial cells (Figure 6). The results showed that cannflavins were approximately 30 times more potent than aspirin in ex vivo experiments (Erridge et al., 2020).
Figure 6.
The Anti-neuroinflammatory activity of Flavonoids in Cannabis.
5. Entourage effects among the phytochemicals in C. Sativa
In 1998, a groundbreaking study conducted by Mechoulam et al. unveiled a pair of monoacylglycerols that influenced the activity of the endogenous cannabinoid 2-arachidonoyl-glycerol through inhibiting its metabolism (Ben-Shabat et al., 1998; Mechoulam & Ben-Shabat, 1999). Despite being pharmacologically inert on their own, these compounds exhibited a significant impact on the activity of the target compound, giving rise to the concept known as the "entourage effect." This effect refers to modifying the pharmacological properties of individual molecules through interactions with co-existing metabolites, even if these metabolites lack inherent pharmacological activity (Cogan, 2020).
Throughout history, cannabis has been utilized as a medicinal plant, and its crude extracts have been found to contain various phytomolecules, such as flavonoids, terpenes, and phytocannabinoids. Recent research has emphasized the preference for combining these phytomolecules in medical therapies due to the observed entourage effect. This phenomenon encompasses two types of interactions: "intra-entourage," arising from interactions among phytocannabinoids or terpenes, and "inter-entourage," resulting from interactions between phytocannabinoids and terpenes (Koltai & Namdar, 2020). Investigating the combinations of phytomolecules exhibiting entourage effects is crucial for developing novel drugs (Koltai & Namdar, 2020).
5.1. The preclinical and clinical evidence
Preclinical studies have demonstrated the interaction between phytocannabinoids and terpenes, suggesting that the enhanced medical benefits of full-spectrum cannabis extracts, compared to isolated molecules, can be attributed to the entourage effect (Mazuz et al., 2020; Namdar et al., 2019). However, it is essential to note that unfavorable interactions, referred to as the "parasitage effect," can also occur in specific in vitro molecular interactions (Namdar et al., 2019).
Careful selection of active phytomolecules and reduction of inactive or potentially pro-inflammatory compounds hold promise for optimizing therapeutic activity. Research has shown that the THCA-rich fraction of a cannabis strain exhibits superior anti-inflammatory activity compared to the crude extract, suggesting the potential benefit of selectively choosing compounds (Nallathambi et al., 2018).
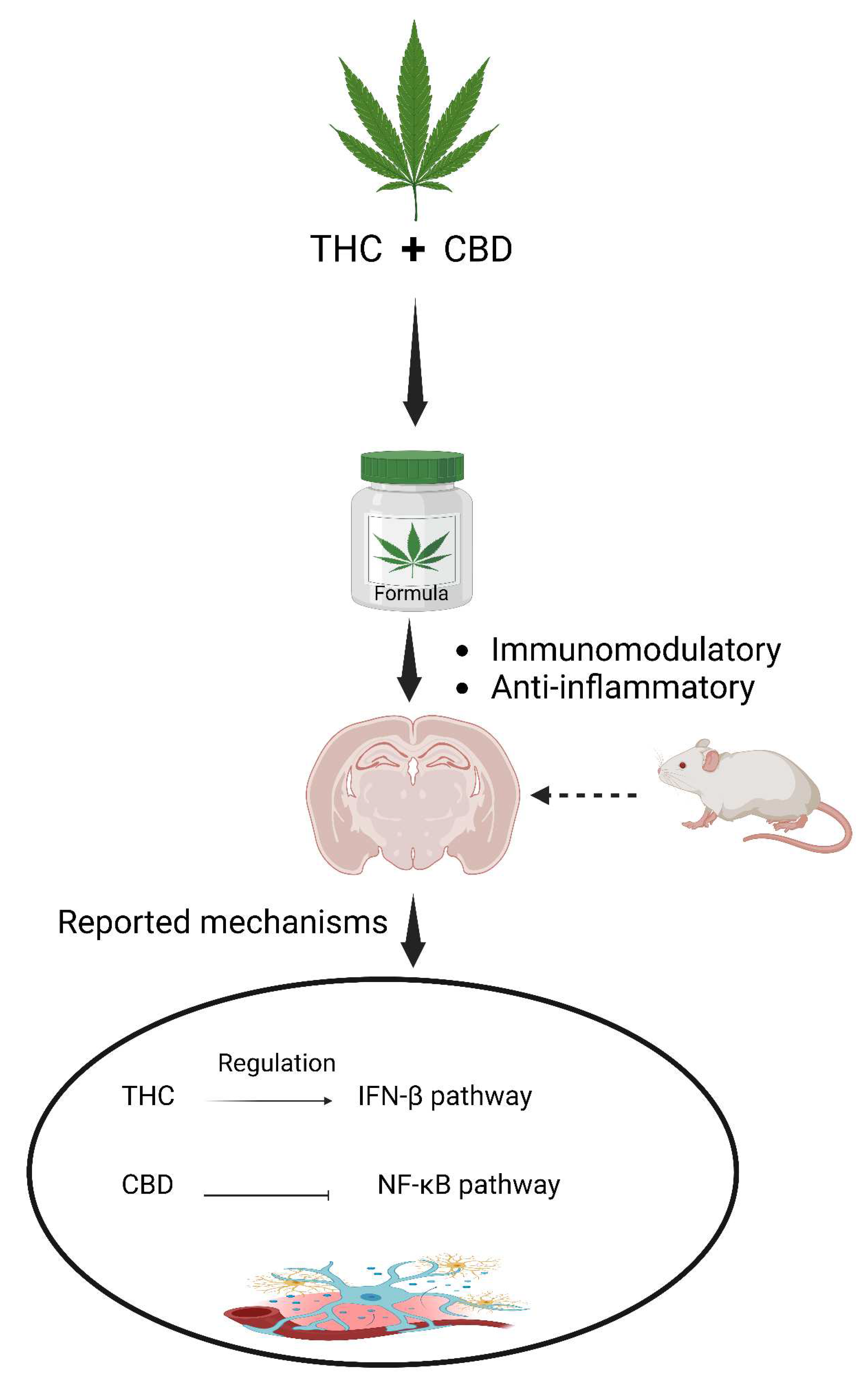
Moreover, recent studies have demonstrated the suppressive effect of a combination of THC and CBD on neuroinflammation in animal models of multiple sclerosis (Feliú et al., 2015; Moreno-Martet et al., 2015). Phytocannabinoids, including THC and CBD, exhibit immunomodulatory and anti-inflammatory properties, acting through distinct signaling pathways. For example, in LPS-activated microglial cells, THC and CBD were found to exert different mechanisms of action, with THC controlling the IFNβ pathway activity and CBD inhibiting the NF-κB-dependent pathway (Figure 7) (Kozela et al., 2010).
Figure 7.
The activity of THC and CBD combination in mediating anti-neuroinflammatory properties.
5.2. The entourage effects in the context of neuroinflammation
The entourage effect of cannabis in the context of neuroinflammation and neurodegenerative disorders is a fascinating phenomenon that underscores the complex interplay between various phytochemicals found in C. Sativa (Hazzah et al., 2020). Extensive research has demonstrated that the therapeutic potential of cannabis extends beyond the individual effects of its primary cannabinoids, such as cannabidiol (CBD) and tetrahydrocannabinol (THC) (Marsh, 2022). Instead, it is the combined action of these cannabinoids, along with a diverse array of terpenes and flavonoids, contributing to the entourage effect, leading to a more comprehensive and robust therapeutic response (Marsh, 2022).
The endocannabinoid system (ECS) is central to the impact of the entourage effect on neuroinflammation and neuroprotection (ECS), a crucial physiological system involved in maintaining homeostasis throughout the body (Soundara Rajan et al., 2017). Cannabinoids, such as CBD and THC, interact with the ECS receptors, CB1 and CB2, modulating inflammatory responses and exerting neuroprotective effects (Soundara Rajan et al., 2017).
In addition to cannabinoids, terpenes play a pivotal role in the entourage effect by enhancing the overall therapeutic potential of cannabis. Terpenes, responsible for the plant's distinctive aroma and flavor, have been found to possess anti-inflammatory, analgesic, and anxiolytic properties (Chacon et al., 2022; Fidyt et al., 2016). For example, β-caryophyllene, a common terpene in Cannabis sativa, has been identified as a selective CB2 receptor agonist with potential anti-inflammatory effects (Fidyt et al., 2016). Moreover, these compounds can influence the blood-brain barrier's permeability, potentially facilitating the passage of cannabinoids into the brain and central nervous system, where they can exert their neuroprotective effects more effectively.
5.3. The mechanisms underpin the entourage effects
The therapeutic synergies between phytocannabinoids and various cannabis phytochemicals remain inadequately investigated, with a limited understanding of the underlying mechanisms and pharmacological basis. Santiago et al. (2019) demonstrated that the dominant terpenes in Cannabis sativa, namely α-pinene, β-pinene, β-caryophyllene, linalool, limonene, and myrcene, either individually or in combinations, did not impact the hyperpolarization induced by Δ9-THC, suggesting that if phytocannabinoid synergies exist, they do not operate through CB1R or CB2R activation (Santiago et al., 2019). However, Cheng et al. (2014) reported that β-caryophyllene prefers binding to CB2R, potentially contributing to synergistic effects within the phytochemical matrix of C. sativa to mitigate Alzheimer's disease (AD)-related neurotoxicity (Cheng et al., 2014).
In enhancing bioavailability, the role of terpenoids, particularly their interaction with phytocannabinoids, warrants further exploration. Namdar et al. (2019) highlighted the need for a comprehensive understanding of potential synergistic actions (Namdar et al., 2019). Terpenes like limonene serve as permeation enhancers for lipophilic compounds through the skin. At the same time, linalool demonstrated the ability to enhance the permeability of hydrophilic compounds via the same route (El Maghraby et al., 2004). Moreover, myrcene's potential to enhance the transport of Δ9-THC across the blood-brain barrier presents a promising avenue for developing centrally penetrant AD therapeutics (Hartsel et al., 2016). The bioavailability of hydrophobic bioactives, such as phytocannabinoids, is notably low through ingestion compared to smoking. Goulle et al. (2008) reported ingestion rates of 6–7%, whereas smoking exhibited higher bioavailability ranging from 10–35% (Goulle et al., 2008). Co-ingestion of triglycerides, particularly long-chain fatty acids, has been identified as a strategy to improve the absorption of ingested lipophilic compounds through the gastrointestinal tract (Wu et al., 2018). Additionally, flavonoids, alkaloids, and other polyphenols have shown potential in increasing phytocannabinoid bioavailability by inhibiting major drug-metabolizing enzymes of the cytochrome P450 family, reducing Phase II-metabolism through inhibition of uridine 5′-diphospho-glucuronosyltransferase, and inhibiting P-glycoprotein 1 efflux pumps (Cherniakov et al., 2017).
Furthermore, flavonoids, another group of phytochemicals in cannabis, have gained increasing attention for their antioxidative and neuroprotective properties. These compounds have shown promise in combating oxidative stress and neurodegeneration, making them valuable contributors to the entourage effect's neuroprotective capabilities (Wang et al., 2021). A review published in the journal Frontiers in Aging Neuroscience highlighted the neuroprotective effects of various flavonoids, including quercetin and apigenin, which have been shown to attenuate neuroinflammation and reduce neurodegenerative processes (Costa et al., 2016).
Positive potentiating interactions, commonly referred to as synergies, occur when the combined effects of compounds exceed the anticipated benefits derived from individual constituents (Caesar & Cech, 2019). The botanical synergies of C. sativa phytochemicals, colloquially known as 'entourage effects,' purportedly demonstrate greater efficacy clinically, in vivo, and in vitro compared to a single or predominant phytocannabinoid molecule (Russo, 2019). Numerous studies have highlighted beneficial combinations for AD prevention. In a mouse model of tauopathy, Sativex (1:1 THC/CBD) reduced Aβ and tau deposition in the hippocampus and cerebral cortex (Casarejos et al., 2013). A CBD-THC combination in the APPxPS1 mouse model also decreased soluble Aβ42 and plaque composition, whereas CBD and THC individually did not (Aso & Ferrer, 2014). Another study using APPxPS1 mice demonstrated that a combination of CBD and THC may improve cognition in aged transgenic AD mice by normalizing synaptosome-associated protein 25, glutamate receptors 2 and 3, and γ-aminobutyric acid receptor A subunit α1 expression (Aso & Ferrer, 2016). Schubert and colleagues also demonstrated significant synergistic in vitro enhancement of neuroprotection between Δ9-THC and CBN in an oxytosis cell death assay (Schubert et al., 2019).
Clinically, a randomized controlled trial showed that oral mucosal C. sativa-based extracts, specifically Δ9-THC and CBD-based whole plant extract, were more effective than placebo or Δ9-THC-predominant extract for treating cancer pain (Johnson et al., 2010). Observations in patients with severe epilepsy indicated notable improvements with lower CBD extract doses than purified CBD (Pamplona et al., 2018). In mice with seizures induced by pentylenetetrazol, the botanical synergy of minor phytocannabinoids was statistically relevant for treating tonic-clonic seizures and improving survival rates (Berman et al., 2018). In non-neurogenic therapeutic areas, Blasco-Benito and colleagues demonstrated that C. sativa extract treatment was more efficient than pure Δ9-THC in producing antitumor responses in vitro and in vivo (Blasco-Benito et al., 2018), while humulene was shown to synergize with β-caryophyllene for enhanced anticancer activities (Legault & Pichette, 2007).
Recent evidence by Finlay and colleagues suggested that terpenoids did not alter the binding of Δ9-THC, CBD, and CBR radioligand ([3H]-CP55,940) or exert functional effects on CB1R or CB2R, indicating that phytocannabinoid synergies may involve pathways beyond direct effects on these receptors (Finlay et al., 2020).
To further understand entourage pathways, investigations into the effects of terpenoids on cannabinoid metabolism and distribution are warranted, as current studies primarily focus on CB1R and CB2R signalling through the Gi/o protein-coupled receptors pathway (Santiago et al., 2019). Notably, Δ9-THC may influence signalling at non-cannabinoid receptor targets (Banister et al., 2019).
Regular consumption of C. sativa seeds may elevate endocannabinoid levels due to their high linoleic acid content (Callaway, 2004; Maccarrone et al., 2010). with potential neuroprotective effects explored in preclinical studies (Bilkei-Gorzo, 2012). This consumption may also be an absorption enhancer due to the high phytochemical content in seeds, sprouts, and leaves (Frassinetti et al., 2018). Cannflavin A, a neuroprotective prenylflavonoid in C. sativa, has a prolonged elimination half-life, suggesting that regular hemp sprouts may extend their presence in plasma and tissues (Werz et al., 2014). Further studies are needed to explore the potential synergies of whole plant C. sativa extracts in preventing neuroinflammatory diseases.
The significance of the entourage effect in the context of neuroinflammation and neuroprotective disorders offers a novel perspective for developing therapeutic interventions. By harnessing the collective strength of various phytochemicals present in C. Sativa, researchers and medical practitioners can explore innovative treatment approaches that capitalise on the synergistic interactions of these compounds. Furthermore, understanding the entourage effect can guide the development of targeted cannabis-based formulations tailored to specific neuroinflammatory conditions and neuroprotective disorders, potentially leading to more effective and well-tolerated treatments for those in need.
6. Conclusion
In cannabis science, cannabinoids, terpenes, and flavonoids have often been overlooked, with much of the literature focusing predominantly on the major cannabinoids THC and CBD. However, emerging evidence suggests that these constituents, particularly cannabinoids and terpenes, play a substantial role in interacting and collaborating. This interplay gives rise to the diverse effects, benefits, and side effects observed among different cannabis strains, which can vary in the ratios of these components (Moulin et al., 2014). Moreover, they are both interact with the endocannabinoid system and exert various effects on the body, including analgesic, anti-inflammatory, and neuroprotective actions. However, it is becoming increasingly clear that their effects are not solely attributed to their actions but are modulated by other compounds in the plant.
Terpenes, aromatic compounds found in cannabis and other plants, contribute to the distinct flavours and aromas associated with different strains. They have been shown to have pharmacological properties and can interact with neurotransmitter receptors, enzymes, and cell membranes, among other targets. Moreover, terpenes can influence the pharmacokinetics and pharmacodynamics of cannabinoids, potentially enhancing or modulating their effects. The concept of the entourage effect suggests that the combined action of cannabinoids and terpenes may result in a synergistic or additive therapeutic effect greater than the sum of their individual effects.
Flavonoids, another class of compounds found in cannabis, have also demonstrated therapeutic potential. Although research on cannabis flavonoids is limited, studies have suggested their anti-inflammatory, antioxidant, and neuroprotective properties. Furthermore, specific flavonoids, such as cannflavins, have shown potent anti-inflammatory effects, particularly in neuroinflammation.
Understanding the intricate interplay between cannabinoids, terpenes, and flavonoids is paramount for realizing the full therapeutic benefits of cannabis. This paper outlines critical research directions and identifies key evidence gaps necessitating immediate attention.
Firstly, elucidating the synergistic effects and underlying mechanisms of cannabinoids, terpenes, and flavonoids demands a focused investigation.
Secondly, comprehending the intricacies of cannabis phytochemical production and accumulation mechanisms, particularly under varying lighting conditions, is pivotal for advancing medicinal applications.
Thirdly, conducting comprehensive phytochemical characterization of cannabis strains, including their distinct ratios of cannabinoids, terpenes, and flavonoids, holds promise for refining treatment strategies. Such endeavours can pave the way for developing more personalized and productive medicinal interventions.
Moreover, addressing regulatory barriers obstructing cannabis research is imperative. Overcoming these obstacles, stemming from the classification of cannabis as a Schedule I substance is crucial to expanding access to cannabis products for research purposes. Furthermore, this would enable a more comprehensive exploration of the therapeutic and adverse effects of cannabis and cannabinoids, fostering informed decision-making in public health initiatives.
Finally, recognizing the value of non-phytocannabinoid compounds, such as terpenes and flavonoids, in therapeutic development necessitates a broader research focus. Exploring these compounds' biosynthesis, bioactivities, and biotechnological applications is pivotal for harnessing their therapeutic potential and diversifying treatment options.
In conclusion, a comprehensive exploration of the synergies between cannabinoids, terpenes, and flavonoids, coupled with advancements in phytochemical research and the removal of regulatory barriers, holds the key to unlocking the full therapeutic potential of cannabis. Addressing these gaps is crucial for advancing the field and fostering evidence-based, personalized treatment modalities.
References
- Abd Rashed, A., Abd Rahman, A. Z., & Rathi, D. N. G. (2021). Essential oils as a potential neuroprotective remedy for age-related neurodegenerative diseases: A review. Molecules, 26(4), 1107. [CrossRef]
- Agostinho, P., A Cunha, R., & Oliveira, C. (2010). Neuroinflammation, oxidative stress and the pathogenesis of Alzheimer's disease. Current pharmaceutical design, 16(25), 2766-2778.
- Aguado, T., Palazuelos, J., Monory, K., Stella, N., Cravatt, B., Lutz, B., Marsicano, G., Kokaia, Z., Guzmán, M., & Galve-Roperh, I. (2006). The endocannabinoid system promotes astroglial differentiation by acting on neural progenitor cells. Journal of Neuroscience, 26(5), 1551-1561. [CrossRef]
- Al Mansouri, S., Ojha, S., Al Maamari, E., Al Ameri, M., Nurulain, S. M., & Bahi, A. (2014). The cannabinoid receptor 2 agonist, β-caryophyllene, reduced voluntary alcohol intake and attenuated ethanol-induced place preference and sensitivity in mice. Pharmacology Biochemistry and Behavior, 124, 260-268.
- Aloisi, F. (2001). Immune function of microglia. Glia, 36(2), 165-179. [CrossRef]
- Amar, M. B. (2006). Cannabinoids in medicine: A review of their therapeutic potential. Journal of ethnopharmacology, 105(1-2), 1-25.
- Ambrose, T., & Simmons, A. (2019). Cannabis, cannabinoids, and the endocannabinoid system—is there therapeutic potential for inflammatory bowel disease? Journal of Crohn's and Colitis, 13(4), 525-535.
- Anderson, L. L., Low, I. K., Banister, S. D., McGregor, I. S., & Arnold, J. C. (2019). Pharmacokinetics of Phytocannabinoid Acids and Anticonvulsant Effect of Cannabidiolic Acid in a Mouse Model of Dravet Syndrome. Journal of Natural Products, 82(11), 3047-3055. [CrossRef]
- Andre, C. M., Hausman, J.-F., & Guerriero, G. (2016). Cannabis sativa: the plant of the thousand and one molecules. Frontiers in plant science, 19. [CrossRef]
- Aso, E., & Ferrer, I. (2014). Cannabinoids for treatment of Alzheimer’s disease: moving toward the clinic. Frontiers in pharmacology, 5, 37.
- Aso, E., & Ferrer, I. (2016). CB2 cannabinoid receptor as potential target against Alzheimer's disease. Frontiers in neuroscience, 10, 243.
- Baker, D., Jackson, S., & Pryce, G. (2007). Cannabinoid control of neuroinflammation related to multiple sclerosis. British journal of pharmacology, 152(5), 649-654. [CrossRef]
- Bamberger, M. E., Harris, M. E., McDonald, D. R., Husemann, J., & Landreth, G. E. (2003). A cell surface receptor complex for fibrillar β-amyloid mediates microglial activation. Journal of Neuroscience, 23(7), 2665-2674. [CrossRef]
- Banister, S. D., Arnold, J. C., Connor, M., Glass, M., & McGregor, I. S. (2019). Dark classics in chemical neuroscience: Δ9-tetrahydrocannabinol. ACS Chemical Neuroscience, 10(5), 2160-2175.
- Baron, E. P. (2018). Medicinal properties of cannabinoids, terpenes, and flavonoids in cannabis, and benefits in migraine, headache, and pain: an update on current evidence and cannabis science. Headache: The Journal of Head and Face Pain, 58(7), 1139-1186. [CrossRef]
- Ben-Shabat, S., Fride, E., Sheskin, T., Tamiri, T., Rhee, M.-H., Vogel, Z., Bisogno, T., De Petrocellis, L., Di Marzo, V., & Mechoulam, R. (1998). An entourage effect: inactive endogenous fatty acid glycerol esters enhance 2-arachidonoyl-glycerol cannabinoid activity. European journal of pharmacology, 353(1), 23-31. [CrossRef]
- Berman, P., Futoran, K., Lewitus, G. M., Mukha, D., Benami, M., Shlomi, T., & Meiri, D. (2018). A new ESI-LC/MS approach for comprehensive metabolic profiling of phytocannabinoids in Cannabis. Scientific Reports, 8(1), 14280. [CrossRef]
- Bilkei-Gorzo, A. (2012). The endocannabinoid system in normal and pathological brain ageing. Philosophical Transactions of the Royal Society B: Biological Sciences, 367(1607), 3326-3341.
- Biringer, R. G. (2021). Endocannabinoid signaling pathways: Beyond CB1R and CB2R. Journal of cell communication and signaling, 15(3), 335-360. [CrossRef]
- Blasco-Benito, S., Seijo-Vila, M., Caro-Villalobos, M., Tundidor, I., Andradas, C., García-Taboada, E., Wade, J., Smith, S., Guzmán, M., & Pérez-Gómez, E. (2018). Appraising the “entourage effect”: Antitumor action of a pure cannabinoid versus a botanical drug preparation in preclinical models of breast cancer. Biochemical pharmacology, 157, 285-293. [CrossRef]
- Blazquez, C., Chiarlone, A., Sagredo, O., Aguado, T., Pazos, M. R., Resel, E., Palazuelos, J., Julien, B., Salazar, M., & Boerner, C. (2011). Loss of striatal type 1 cannabinoid receptors is a key pathogenic factor in Huntington’s disease. Brain, 134(1), 119-136. [CrossRef]
- Bonini, S. A., Premoli, M., Tambaro, S., Kumar, A., Maccarinelli, G., Memo, M., & Mastinu, A. (2018). Cannabis sativa: A comprehensive ethnopharmacological review of a medicinal plant with a long history. Journal of ethnopharmacology, 227, 300-315. [CrossRef]
- Bradshaw, E. M., Chibnik, L. B., Keenan, B. T., Ottoboni, L., Raj, T., Tang, A., Rosenkrantz, L. L., Imboywa, S., Lee, M., & Von Korff, A. (2013). CD33 Alzheimer's disease locus: altered monocyte function and amyloid biology. Nature neuroscience, 16(7), 848-850.
- Caesar, L. K., & Cech, N. B. (2019). Synergy and antagonism in natural product extracts: when 1+ 1 does not equal 2. Natural product reports, 36(6), 869-888. [CrossRef]
- Callaway, J. (2004). Hempseed as a nutritional resource: An overview. Euphytica, 140, 65-72. [CrossRef]
- Cardona, A. E., Huang, D., Sasse, M. E., & Ransohoff, R. M. (2006). Isolation of murine microglial cells for RNA analysis or flow cytometry. Nature protocols, 1(4), 1947-1951. [CrossRef]
- Carow, B., & Rottenberg, M. E. (2014). SOCS3, a Major Regulator of Infection and Inflammation. Front Immunol, 5, 58. [CrossRef]
- Carrier, E. J., Auchampach, J. A., & Hillard, C. J. (2006). Inhibition of an equilibrative nucleoside transporter by cannabidiol: a mechanism of cannabinoid immunosuppression. Proceedings of the National Academy of Sciences, 103(20), 7895-7900. [CrossRef]
- Carrillo-Salinas, F. J., Navarrete, C., Mecha, M., Feliú, A., Collado, J. A., Cantarero, I., Bellido, M. L., Muñoz, E., & Guaza, C. (2014). A cannabigerol derivative suppresses immune responses and protects mice from experimental autoimmune encephalomyelitis. PloS one, 9(4), e94733. [CrossRef]
- Casarejos, M. J., Perucho, J., Gomez, A., Munoz, M. P., Fernandez-Estevez, M., Sagredo, O., Fernandez Ruiz, J., Guzman, M., de Yebenes, J. G., & Mena, M. A. (2013). Natural cannabinoids improve dopamine neurotransmission and tau and amyloid pathology in a mouse model of tauopathy. Journal of Alzheimer's Disease, 35(3), 525-539. [CrossRef]
- Castillo, A., Tolón, M., Fernández-Ruiz, J., Romero, J., & Martinez-Orgado, J. (2010). The neuroprotective effect of cannabidiol in an in vitro model of newborn hypoxic–ischemic brain damage in mice is mediated by CB2 and adenosine receptors. Neurobiology of disease, 37(2), 434-440. [CrossRef]
- Chacon, F. T., Raup-Konsavage, W. M., Vrana, K. E., & Kellogg, J. J. (2022). Secondary Terpenes in Cannabis sativa L.: Synthesis and Synergy. Biomedicines, 10(12). [CrossRef]
- Cheng, Y., Dong, Z., & Liu, S. (2014). β-Caryophyllene ameliorates the Alzheimer-like phenotype in APP/PS1 Mice through CB2 receptor activation and the PPARγ pathway. Pharmacology, 94(1-2), 1-12. [CrossRef]
- Cherniakov, I., Izgelov, D., Domb, A. J., & Hoffman, A. (2017). The effect of Pro NanoLipospheres (PNL) formulation containing natural absorption enhancers on the oral bioavailability of delta-9-tetrahydrocannabinol (THC) and cannabidiol (CBD) in a rat model. European Journal of Pharmaceutical Sciences, 109, 21-30. [CrossRef]
- Chiurchiù, V., Leuti, A., & Maccarrone, M. (2015). Cannabinoid Signaling and Neuroinflammatory Diseases: A Melting pot for the Regulation of Brain Immune Responses. J Neuroimmune Pharmacol, 10(2), 268-280. [CrossRef]
- Cogan, P. S. (2020). The ‘entourage effect’ or ‘hodge-podge hashish’: the questionable rebranding, marketing, and expectations of cannabis polypharmacy. Expert Review of Clinical Pharmacology, 13(8), 835-845. [CrossRef]
- Correa, F., Docagne, F., Mestre, L., Clemente, D., Hernangómez, M., Loría, F., & Guaza, C. (2009). A role for CB2 receptors in anandamide signalling pathways involved in the regulation of IL-12 and IL-23 in microglial cells. Biochemical pharmacology, 77(1), 86-100. [CrossRef]
- Correa, F., Hernangómez, M., Mestre, L., Loría, F., Spagnolo, A., Docagne, F., Di Marzo, V., & Guaza, C. (2010). Anandamide enhances IL-10 production in activated microglia by targeting CB2 receptors: Roles of ERK1/2, JNK, and NF-κB. Glia, 58(2), 135-147.
- Costa, L., Garrick, J., Roquè, P., & Pellacani, C. (2016). Mechanisms of neuroprotection by quercetin: counteracting oxidative stress and more. Oxid Med Cell Longev 2016: 2986796. In: Epub 2016/02/24. [CrossRef]
- Cox-Georgian, D., Ramadoss, N., Dona, C., & Basu, C. (2019). Therapeutic and medicinal uses of terpenes. Medicinal plants: from farm to pharmacy, 333-359.
- Cristino, L., Bisogno, T., & Di Marzo, V. (2020). Cannabinoids and the expanded endocannabinoid system in neurological disorders. Nature Reviews Neurology, 16(1), 9-29. [CrossRef]
- De-Oliveira, A., Ribeiro-Pinto, L., Otto, S., Gonçalves, A., & Paumgartten, F. (1997). Induction of liver monooxygenases by β-myrcene. Toxicology, 124(2), 135-140. [CrossRef]
- De Petrocellis, L., Ligresti, A., Moriello, A. S., Allarà, M., Bisogno, T., Petrosino, S., Stott, C. G., & Di Marzo, V. (2011). Effects of cannabinoids and cannabinoid-enriched Cannabis extracts on TRP channels and endocannabinoid metabolic enzymes. British journal of pharmacology, 163(7), 1479-1494. [CrossRef]
- De Vries, H. E., Blom-Roosemalen, M. C., Van Oosten, M., De Boer, A. G., Van Berkel, T. J., Breimer, D. D., & Kuiper, J. (1996). The influence of cytokines on the integrity of the blood-brain barrier in vitro. Journal of neuroimmunology, 64(1), 37-43.
- Di Marzo, V., & Piscitelli, F. (2015). The endocannabinoid system and its modulation by phytocannabinoids. Neurotherapeutics, 12, 692-698.
- Díaz-Alonso, J., Paraíso-Luna, J., Navarrete, C., del Río, C., Cantarero, I., Palomares, B., Aguareles, J., Fernández-Ruiz, J., Bellido, M. L., Pollastro, F., Appendino, G., Calzado, M. A., Galve-Roperh, I., & Muñoz, E. (2016). VCE-003.2, a novel cannabigerol derivative, enhances neuronal progenitor cell survival and alleviates symptomatology in murine models of Huntington’s disease. Scientific Reports, 6(1), 29789. [CrossRef]
- Dowie, M., Howard, M., Nicholson, L., Faull, R., Hannan, A., & Glass, M. (2010). Behavioural and molecular consequences of chronic cannabinoid treatment in Huntington's disease transgenic mice. Neuroscience, 170(1), 324-336.
- Downer, E. J. (2020). Anti-inflammatory Potential of Terpenes Present in Cannabis sativa L. ACS Chemical Neuroscience, 11(5), 659-662. [CrossRef]
- El-Remessy, A. B., Tang, Y., Zhu, G., Matragoon, S., Khalifa, Y., Liu, E., Liu, J., Hanson, E., Mian, S., & Fatteh, N. (2008). Neuroprotective effects of cannabidiol in endotoxin-induced uveitis: critical role of p38 MAPK activation. Molecular vision, 14, 2190.
- El Khoury, J. B., Moore, K. J., Means, T. K., Leung, J., Terada, K., Toft, M., Freeman, M. W., & Luster, A. D. (2003). CD36 mediates the innate host response to β-amyloid. The Journal of experimental medicine, 197(12), 1657-1666. [CrossRef]
- El Maghraby, G., Williams, A. C., & Barry, B. (2004). Interactions of surfactants (edge activators) and skin penetration enhancers with liposomes. International journal of pharmaceutics, 276(1-2), 143-161. [CrossRef]
- Eljaschewitsch, E., Witting, A., Mawrin, C., Lee, T., Schmidt, P. M., Wolf, S., Hoertnagl, H., Raine, C. S., Schneider-Stock, R., & Nitsch, R. (2006). The endocannabinoid anandamide protects neurons during CNS inflammation by induction of MKP-1 in microglial cells. Neuron, 49(1), 67-79. [CrossRef]
- ElSohly, M. A., Radwan, M. M., Gul, W., Chandra, S., & Galal, A. (2017). Phytochemistry of Cannabis sativa L. Phytocannabinoids: unraveling the complex chemistry and pharmacology of Cannabis sativa, 1-36.
- Erridge, S., Mangal, N., Salazar, O., Pacchetti, B., & Sodergren, M. H. (2020). Cannflavins – From plant to patient: A scoping review. Fitoterapia, 146, 104712. [CrossRef]
- Espadas, I., Keifman, E., Palomo-Garo, C., Burgaz, S., García, C., Fernández-Ruiz, J., & Moratalla, R. (2020). Beneficial effects of the phytocannabinoid Δ9-THCV in L-DOPA-induced dyskinesia in Parkinson's disease. Neurobiology of disease, 141, 104892. [CrossRef]
- Esposito, G., De Filippis, D., Carnuccio, R., Izzo, A. A., & Iuvone, T. (2006). The marijuana component cannabidiol inhibits β-amyloid-induced tau protein hyperphosphorylation through Wnt/β-catenin pathway rescue in PC12 cells. Journal of molecular medicine, 84, 253-258. [CrossRef]
- Esposito, G., Scuderi, C., Savani, C., Steardo Jr, L., De Filippis, D., Cottone, P., Iuvone, T., Cuomo, V., & Steardo, L. (2007). Cannabidiol in vivo blunts β-amyloid induced neuroinflammation by suppressing IL-1β and iNOS expression. British journal of pharmacology, 151(8), 1272-1279.
- Eubanks, L. M., Rogers, C. J., Beuscher IV, A. E., Koob, G. F., Olson, A. J., Dickerson, T. J., & Janda, K. D. (2006). A molecular link between the active component of marijuana and Alzheimer's disease pathology. Molecular pharmaceutics, 3(6), 773-777.
- Fassbender, K., Walter, S., Kühl, S., Landmann, R., Ishii, K., Bertsch, T., Stalder, A., Muehlhauser, F., Liu, Y., & Ulmer, A. (2004). The LPS receptor (CD14) links innate immunity with Alzheimer's disease. The FASEB Journal, 18(1), 203-205.
- Feliú, A., Moreno-Martet, M., Mecha, M., Carrillo-Salinas, F., De Lago, E., Fernández-Ruiz, J., & Guaza, C. (2015). AS ativex®-like combination of phytocannabinoids as a disease-modifying therapy in a viral model of multiple sclerosis. British journal of pharmacology, 172(14), 3579-3595.
- Fidyt, K., Fiedorowicz, A., Strządała, L., & Szumny, A. (2016). β-caryophyllene and β-caryophyllene oxide—natural compounds of anticancer and analgesic properties. Cancer medicine, 5(10), 3007-3017.
- Filiano, A. J., Gadani, S. P., & Kipnis, J. (2015). Interactions of innate and adaptive immunity in brain development and function. Brain research, 1617, 18-27. [CrossRef]
- Finlay, D. B., Sircombe, K. J., Nimick, M., Jones, C., & Glass, M. (2020). Terpenoids from cannabis do not mediate an entourage effect by acting at cannabinoid receptors. Frontiers in pharmacology, 11, 359. [CrossRef]
- Frassinetti, S., Moccia, E., Caltavuturo, L., Gabriele, M., Longo, V., Bellani, L., Giorgi, G., & Giorgetti, L. (2018). Nutraceutical potential of hemp (Cannabis sativa L.) seeds and sprouts. Food chemistry, 262, 56-66. [CrossRef]
- Galiègue, S., Mary, S., Marchand, J., Dussossoy, D., Carrière, D., Carayon, P., Bouaboula, M., Shire, D., LE Fur, G., & Casellas, P. (1995). Expression of central and peripheral cannabinoid receptors in human immune tissues and leukocyte subpopulations. European journal of biochemistry, 232(1), 54-61. [CrossRef]
- García, C., Gómez-Cañas, M., Burgaz, S., Palomares, B., Gómez-Gálvez, Y., Palomo-Garo, C., Campo, S., Ferrer-Hernández, J., Pavicic, C., Navarrete, C., Luz Bellido, M., García-Arencibia, M., Ruth Pazos, M., Muñoz, E., & Fernández-Ruiz, J. (2018). Benefits of VCE-003.2, a cannabigerol quinone derivative, against inflammation-driven neuronal deterioration in experimental Parkinson’s disease: possible involvement of different binding sites at the PPARγ receptor. Journal of Neuroinflammation, 15(1), 19. [CrossRef]
- Gaston, T. E., & Friedman, D. (2017). Pharmacology of cannabinoids in the treatment of epilepsy. Epilepsy & Behavior, 70, 313-318. [CrossRef]
- Gertsch, J., Leonti, M., Raduner, S., Racz, I., Chen, J.-Z., Xie, X.-Q., Altmann, K.-H., Karsak, M., & Zimmer, A. (2008). Beta-caryophyllene is a dietary cannabinoid. Proceedings of the National Academy of Sciences, 105(26), 9099-9104. [CrossRef]
- Giacobbe, J., Marrocu, A., Di Benedetto, M. G., Pariante, C. M., & Borsini, A. (2021). A systematic, integrative review of the effects of the endocannabinoid system on inflammation and neurogenesis in animal models of affective disorders. Brain, behavior, and immunity, 93, 353-367. [CrossRef]
- Goulle, J., Saussereau, E., & Lacroix, C. (2008). Delta-9-tetrahydrocannabinol pharmacokinetics. Annales pharmaceutiques francaises.
- Granja, A. G., Carrillo-Salinas, F., Pagani, A., Gómez-Cañas, M., Negri, R., Navarrete, C., Mecha, M., Mestre, L., Fiebich, B. L., & Cantarero, I. (2012). A cannabigerol quinone alleviates neuroinflammation in a chronic model of multiple sclerosis. Journal of neuroimmune pharmacology, 7(4), 1002-1016. [CrossRef]
- Griciuc, A., Serrano-Pozo, A., Parrado, A. R., Lesinski, A. N., Asselin, C. N., Mullin, K., Hooli, B., Choi, S. H., Hyman, B. T., & Tanzi, R. E. (2013). Alzheimer’s disease risk gene CD33 inhibits microglial uptake of amyloid beta. Neuron, 78(4), 631-643. [CrossRef]
- Guerreiro, R., Wojtas, A., Bras, J., Carrasquillo, M., Rogaeva, E., Majounie, E., Cruchaga, C., Sassi, C., Kauwe, J. S., & Younkin, S. (2013). TREM2 variants in Alzheimer's disease. New England Journal of Medicine, 368(2), 117-127.
- Gugliandolo, A., Pollastro, F., Grassi, G., Bramanti, P., & Mazzon, E. (2018). In Vitro Model of Neuroinflammation: Efficacy of Cannabigerol, a Non-Psychoactive Cannabinoid. International Journal of Molecular Sciences, 19(7), 1992. https://www.mdpi.com/1422-0067/19/7/1992. [CrossRef]
- Gülck, T., Booth, J., Carvalho, Â., Khakimov, B., Crocoll, C., Motawia, M., Møller, B., Bohlmann, J., & Gallage, N. (2020). Synthetic biology of cannabinoids and cannabinoid glucosides in Nicotiana benthamiana and Saccharomyces cerevisiae. Journal of Natural Products, 83(10), 2877-2893. [CrossRef]
- Hagberg, B., Hanefeld, F., Percy, A., & Skjeldal, O. (2002). An update on clinically applicable diagnostic criteria in Rett syndrome. Comments to Rett syndrome clinical criteria consensus panel satellite to European Paediatric neurology society meeting, Baden Baden, Germany, 11 September 2001. European journal of paediatric neurology: EJPN: official journal of the European Paediatric Neurology Society, 6(5), 293-297. [CrossRef]
- Hanuš, L. O., Meyer, S. M., Muñoz, E., Taglialatela-Scafati, O., & Appendino, G. (2016). Phytocannabinoids: a unified critical inventory. Natural product reports, 33(12), 1357-1392. [CrossRef]
- Hartsel, J. A., Eades, J., Hickory, B., & Makriyannis, A. (2016). Cannabis sativa and Hemp. In Nutraceuticals (pp. 735-754). Elsevier.
- Hazzah, T., Andre, C., Richter, G., McGrath, S., & Collins, F. (2020). Cannabis in veterinary medicine: a critical review. AHVMA, 61, 25.
- He, B., Chen, Y., Yu, S., Hao, Y., Wang, F., & Qu, L. (2022). Food plant extracts for sleep-related skin health: mechanisms and prospects. Food Bioscience, 101951. [CrossRef]
- Heneka, M. T., Kummer, M. P., Stutz, A., Delekate, A., Schwartz, S., Vieira-Saecker, A., Griep, A., Axt, D., Remus, A., & Tzeng, T.-C. (2013). NLRP3 is activated in Alzheimer’s disease and contributes to pathology in APP/PS1 mice. Nature, 493(7434), 674-678. [CrossRef]
- Hickman, S. E., Allison, E. K., & El Khoury, J. (2008). Microglial dysfunction and defective β-amyloid clearance pathways in aging Alzheimer's disease mice. Journal of Neuroscience, 28(33), 8354-8360.
- Hill, A. J., Weston, S. E., Jones, N. A., Smith, I., Bevan, S. A., Williamson, E. M., Stephens, G. J., Williams, C. M., & Whalley, B. J. (2010). Δ9-Tetrahydrocannabivarin suppresses in vitro epileptiform and in vivo seizure activity in adult rats. Epilepsia, 51(8), 1522-1532.
- Hillard, C. J. (2018). Circulating endocannabinoids: from whence do they come and where are they going? Neuropsychopharmacology, 43(1), 155-172.
- Howlett, A. C., Evans, D., & Houston, D. (2019). The cannabinoid receptor. Marijuana/cannabinoids, 35-72.
- Jean-Gilles, L., Gran, B., & Constantinescu, C. S. (2010). Interaction between cytokines, cannabinoids and the nervous system. Immunobiology, 215(8), 606-610. [CrossRef]
- Ji, K., Akgul, G., Wollmuth, L. P., & Tsirka, S. E. (2013). Microglia actively regulate the number of functional synapses. PloS one, 8(2), e56293. [CrossRef]
- Jin, D., Dai, K., Xie, Z., & Chen, J. (2020). Secondary metabolites profiled in cannabis inflorescences, leaves, stem barks, and roots for medicinal purposes. Scientific Reports, 10(1), 1-14. [CrossRef]
- Johnson, J. R., Burnell-Nugent, M., Lossignol, D., Ganae-Motan, E. D., Potts, R., & Fallon, M. T. (2010). Multicenter, double-blind, randomized, placebo-controlled, parallel-group study of the efficacy, safety, and tolerability of THC: CBD extract and THC extract in patients with intractable cancer-related pain. Journal of pain and symptom management, 39(2), 167-179.
- Kagan, J. C., & Horng, T. (2013). NLRP3 inflammasome activation: CD36 serves double duty. Nature immunology, 14(8), 772-774. [CrossRef]
- Kaplan, J. S., Stella, N., Catterall, W. A., & Westenbroek, R. E. (2017). Cannabidiol attenuates seizures and social deficits in a mouse model of Dravet syndrome. Proceedings of the National Academy of Sciences, 114(42), 11229-11234. [CrossRef]
- Kennedy, D. O., Dodd, F. L., Robertson, B. C., Okello, E. J., Reay, J. L., Scholey, A. B., & Haskell, C. F. (2011). Monoterpenoid extract of sage (Salvia lavandulaefolia) with cholinesterase inhibiting properties improves cognitive performance and mood in healthy adults. Journal of Psychopharmacology, 25(8), 1088-1100. [CrossRef]
- Keren-Shaul, H., Spinrad, A., Weiner, A., Matcovitch-Natan, O., Dvir-Szternfeld, R., Ulland, T. K., David, E., Baruch, K., Lara-Astaiso, D., & Toth, B. (2017). A unique microglia type associated with restricting development of Alzheimer’s disease. Cell, 169(7), 1276-1290. e1217. [CrossRef]
- Kettenmann, H., Hanisch, U.-K., Noda, M., & Verkhratsky, A. (2011). Physiology of microglia. Physiological reviews, 91(2), 461-553. [CrossRef]
- Killestein, J., Hoogervorst, E., Reif, M., Blauw, B., Smits, M., Uitdehaag, B., Nagelkerken, L., & Polman, C. (2003). Immunomodulatory effects of orally administered cannabinoids in multiple sclerosis. Journal of neuroimmunology, 137(1-2), 140-143.
- Kitazawa, M., Oddo, S., Yamasaki, T. R., Green, K. N., & LaFerla, F. M. (2005). Lipopolysaccharide-induced inflammation exacerbates tau pathology by a cyclin-dependent kinase 5-mediated pathway in a transgenic model of Alzheimer's disease. Journal of Neuroscience, 25(39), 8843-8853.
- Koltai, H., & Namdar, D. (2020). Cannabis Phytomolecule 'Entourage': From Domestication to Medical Use. Trends in Plant Science, 25(10), 976-984. [CrossRef]
- Konsman, J. P. (2022). Cytokines in the Brain and Neuroinflammation: We Didn’t Starve the Fire! Pharmaceuticals, 15(2), 140. https://www.mdpi.com/1424-8247/15/2/140. [CrossRef]
- Kozela, E., Pietr, M., Juknat, A., Rimmerman, N., Levy, R., & Vogel, Z. (2010). Cannabinoids Δ9-Tetrahydrocannabinol and Cannabidiol Differentially Inhibit the Lipopolysaccharide-activated NF-κB and Interferon-β/STAT Proinflammatory Pathways in BV-2 Microglial Cells*. Journal of Biological Chemistry, 285(3), 1616-1626. [CrossRef]
- Laflamme, N., & Rivest, S. (1999). Effects of systemic immunogenic insults and circulating proinflammatory cytokines on the transcription of the inhibitory factor κBα within specific cellular populations of the rat brain. Journal of neurochemistry, 73(1), 309-321. [CrossRef]
- Lastres-Becker, I., Bizat, N., Boyer, F., Hantraye, P., Brouillet, E., & Fernández-Ruiz, J. (2003). Effects of cannabinoids in the rat model of Huntington's disease generated by an intrastriatal injection of malonate. Neuroreport, 14(6), 813-816. [CrossRef]
- Lastres-Becker, I., Molina-Holgado, F., Ramos, J. A., Mechoulam, R., & Fernández-Ruiz, J. (2005). Cannabinoids provide neuroprotection against 6-hydroxydopamine toxicity in vivo and in vitro: relevance to Parkinson's disease. Neurobiology of disease, 19(1-2), 96-107.
- Legault, J., & Pichette, A. (2007). Potentiating effect of β-caryophyllene on anticancer activity of α-humulene, isocaryophyllene and paclitaxel. Journal of Pharmacy and Pharmacology, 59(12), 1643-1647.
- Liu, G., & Jiang, Q. (2016). Alzheimer’s disease CD33 rs3865444 variant does not contribute to cognitive performance. Proceedings of the National Academy of Sciences, 113(12), E1589-E1590. [CrossRef]
- Lorenzetti, B. B., Souza, G. E., Sarti, S. J., Santos Filho, D., & Ferreira, S. H. (1991). Myrcene mimics the peripheral analgesic activity of lemongrass tea. Journal of ethnopharmacology, 34(1), 43-48. [CrossRef]
- Ludwiczuk, A., Skalicka-Woźniak, K., & Georgiev, M. (2017). Terpenoids. In Pharmacognosy (pp. 233-266). Elsevier.
- Lyman, M., Lloyd, D. G., Ji, X., Vizcaychipi, M. P., & Ma, D. (2014). Neuroinflammation: The role and consequences. Neuroscience Research, 79, 1-12. [CrossRef]
- Maccarrone, M., Rossi, S., Bari, M., De Chiara, V., Rapino, C., Musella, A., Bernardi, G., Bagni, C., & Centonze, D. (2010). Abnormal mGlu 5 receptor/endocannabinoid coupling in mice lacking FMRP and BC1 RNA. Neuropsychopharmacology, 35(7), 1500-1509. [CrossRef]
- Maestroni, G. J. (2004). The endogenous cannabinoid 2-arachidonoyl glycerol as in vivo chemoattractant for dendritic cells and adjuvant for Th1 response to a soluble protein. The FASEB Journal, 18(15), 1914-1916. [CrossRef]
- Marinelli, S., Pacioni, S., Bisogno, T., Di Marzo, V., Prince, D. A., Huguenard, J. R., & Bacci, A. (2008). The endocannabinoid 2-arachidonoylglycerol is responsible for the slow self-inhibition in neocortical interneurons. Journal of Neuroscience, 28(50), 13532-13541. [CrossRef]
- Marquette, C., Van Dam, A.-M., Ceccaldi, P.-E., Weber, P., Haour, F., & Tsiang, H. (1996). Induction of immunoreactive interleukin-1β and tumor necrosis factor-α in the brains of rabies virus infected rats. Journal of neuroimmunology, 68(1-2), 45-51. [CrossRef]
- Marsh, D. T. (2022). Exploring the Bioactivity and Therapeutic Potential of Structurally Diverse Phytochemicals in Neurodegenerative and Gastrointestinal Disease.
- Mawuenyega, K. G., Sigurdson, W., Ovod, V., Munsell, L., Kasten, T., Morris, J. C., Yarasheski, K. E., & Bateman, R. J. (2010). Decreased clearance of CNS β-amyloid in Alzheimer’s disease. Science, 330(6012), 1774-1774.
- Mazuz, M., Tiroler, A., Moyal, L., Hodak, E., Nadarajan, S., Vinayaka, A. C., Gorovitz-Haris, B., Lubin, I., Drori, A., & Drori, G. (2020). Synergistic cytotoxic activity of cannabinoids from cannabis sativa against cutaneous T-cell lymphoma (CTCL) in-vitro and ex-vivo. Oncotarget, 11(13), 1141.
- Mechoulam, R., & Ben-Shabat, S. (1999). From gan-zi-gun-nu to anandamide and 2-arachidonoylglycerol: the ongoing story of cannabis. Natural product reports, 16(2), 131-143. [CrossRef]
- Micheau, O., & Tschopp, J. (2003). Induction of TNF receptor I-mediated apoptosis via two sequential signaling complexes. Cell, 114(2), 181-190. [CrossRef]
- Moreno-Martet, M., Feliú, A., Espejo-Porras, F., Mecha, M., Carrillo-Salinas, F. J., Fernández-Ruiz, J., Guaza, C., & de Lago, E. (2015). The disease-modifying effects of a Sativex-like combination of phytocannabinoids in mice with experimental autoimmune encephalomyelitis are preferentially due to Δ9-tetrahydrocannabinol acting through CB1 receptors. Multiple sclerosis and related disorders, 4(6), 505-511. [CrossRef]
- Mosley, R. L., Benner, E. J., Kadiu, I., Thomas, M., Boska, M. D., Hasan, K., Laurie, C., & Gendelman, H. E. (2006). Neuroinflammation, oxidative stress, and the pathogenesis of Parkinson’s disease. Clinical neuroscience research, 6(5), 261-281.
- Moulin, D., Boulanger, A., Clark, A., Clarke, H., Dao, T., Finley, G., Furlan, A., Gilron, I., Gordon, A., & Morley-Forster, P. K. (2014). Pharmacological management of chronic neuropathic pain: revised consensus statement from the Canadian Pain Society. Pain Research and Management, 19, 328-335. [CrossRef]
- Nadal, X., del Río, C., Casano, S., Palomares, B., Ferreiro-Vera, C., Navarrete, C., Sánchez-Carnerero, C., Cantarero, I., Bellido, M. L., Meyer, S., Morello, G., Appendino, G., & Muñoz, E. (2017). Tetrahydrocannabinolic acid is a potent PPARγ agonist with neuroprotective activity. British journal of pharmacology, 174(23), 4263-4276. [CrossRef]
- Nallathambi, R., Mazuz, M., Namdar, D., Shik, M., Namintzer, D., Vinayaka, A. C., Ion, A., Faigenboim, A., Nasser, A., Laish, I., Konikoff, F. M., & Koltai, H. (2018). Identification of Synergistic Interaction Between Cannabis-Derived Compounds for Cytotoxic Activity in Colorectal Cancer Cell Lines and Colon Polyps That Induces Apoptosis-Related Cell Death and Distinct Gene Expression. Cannabis Cannabinoid Res, 3(1), 120-135. [CrossRef]
- Namdar, D., Voet, H., Ajjampura, V., Nadarajan, S., Mayzlish-Gati, E., Mazuz, M., Shalev, N., & Koltai, H. (2019). Terpenoids and phytocannabinoids co-produced in Cannabis sativa strains show specific interaction for cell cytotoxic activity. Molecules, 24(17), 3031. [CrossRef]
- Neves, Â., Rosa, S., Gonçalves, J., Rufino, A., Judas, F., Salgueiro, L., Lopes, M. C., Cavaleiro, C., & Mendes, A. F. (2010). Screening of five essential oils for identification of potential inhibitors of IL-1-induced NF-κB activation and NO production in human chondrocytes: characterization of the inhibitory activity of α-pinene. Planta medica, 76(03), 303-308.
- Noma, Y., Hashimoto, T., Uehara, S., & Asakawa, Y. (2010). Erratum: Microbial transformation of isopinocampheol and caryophyllene oxide. Flavour and fragrance journal, 25(4), 257-266. [CrossRef]
- Osborne, A. L., Solowij, N., Babic, I., Huang, X.-F., & Weston-Green, K. (2017). Improved social interaction, recognition and working memory with cannabidiol treatment in a prenatal infection (poly I: C) rat model. Neuropsychopharmacology, 42(7), 1447-1457.
- Pamplona, F. A., Da Silva, L. R., & Coan, A. C. (2018). Potential clinical benefits of CBD-rich cannabis extracts over purified CBD in treatment-resistant epilepsy: observational data meta-analysis. Frontiers in neurology, 9, 759.
- Parkhurst, C. N., Yang, G., Ninan, I., Savas, J. N., Yates III, J. R., Lafaille, J. J., Hempstead, B. L., Littman, D. R., & Gan, W.-B. (2013). Microglia promote learning-dependent synapse formation through brain-derived neurotrophic factor. Cell, 155(7), 1596-1609.
- Piomelli, D., Beltramo, M., Giuffrida, A., & Stella, N. (1998). Endogenous cannabinoid signaling. Neurobiology of disease, 5(6), 462-473. [CrossRef]
- Pisanti, S., & Bifulco, M. (2019). Medical Cannabis: A plurimillennial history of an evergreen. Journal of cellular physiology, 234(6), 8342-8351. [CrossRef]
- Pretzsch, C. M., Voinescu, B., Lythgoe, D., Horder, J., Mendez, M. A., Wichers, R., Ajram, L., Ivin, G., Heasman, M., & Edden, R. A. (2019). Effects of cannabidivarin (CBDV) on brain excitation and inhibition systems in adults with and without Autism Spectrum Disorder (ASD): a single dose trial during magnetic resonance spectroscopy. Translational psychiatry, 9(1), 313. [CrossRef]
- Puffenbarger, R. A., Boothe, A. C., & Cabral, G. A. (2000). Cannabinoids inhibit LPS-inducible cytokine mRNA expression in rat microglial cells. Glia, 29(1), 58-69. [CrossRef]
- Pugazhendhi, A., Suganthy, N., Chau, T. P., Sharma, A., Unpaprom, Y., Ramaraj, R., Karuppusamy, I., & Brindhadevi, K. (2021). Cannabinoids as anticancer and neuroprotective drugs: Structural insights and pharmacological interactions—A review. Process Biochemistry, 111, 9-31. [CrossRef]
- Rahaman, O., & Ganguly, D. (2021). Endocannabinoids in immune regulation and immunopathologies. Immunology, 164(2), 242-252. [CrossRef]
- Rao, R., Nagarkatti, P., & Nagarkatti, M. (2014). Staphylococcal enterotoxin B-induced microRNA-155 targets SOCS1 to promote acute inflammatory lung injury. Infection and immunity, 82(7), 2971-2979. [CrossRef]
- Ricceri, L., De Filippis, B., & Laviola, G. (2013). Rett syndrome treatment in mouse models: searching for effective targets and strategies. Neuropharmacology, 68, 106-115. [CrossRef]
- Rufino, A. T., Ribeiro, M., Judas, F., Salgueiro, L., Lopes, M. C., Cavaleiro, C., & Mendes, A. F. (2014). Anti-inflammatory and chondroprotective activity of (+)-α-pinene: structural and enantiomeric selectivity. Journal of Natural Products, 77(2), 264-269. [CrossRef]
- Russo, E. B. (2019). The case for the entourage effect and conventional breeding of clinical cannabis: no “strain,” no gain. Frontiers in plant science, 9, 1969.
- Saito, V. M., Rezende, R. M., & Teixeira, A. L. (2012). Cannabinoid modulation of neuroinflammatory disorders. Curr Neuropharmacol, 10(2), 159-166. [CrossRef]
- Santiago, M., Sachdev, S., Arnold, J. C., McGregor, I. S., & Connor, M. (2019). Absence of entourage: terpenoids commonly found in Cannabis sativa do not modulate the functional activity of Δ9-THC at human CB1 and CB2 receptors. Cannabis and cannabinoid research, 4(3), 165-176.
- Saresella, M., La Rosa, F., Piancone, F., Zoppis, M., Marventano, I., Calabrese, E., Rainone, V., Nemni, R., Mancuso, R., & Clerici, M. (2016). The NLRP3 and NLRP1 inflammasomes are activated in Alzheimer’s disease. Molecular neurodegeneration, 11(1), 1-14. 10.1186/s13024-016-0088-1.
- Schilling, S., Melzer, R., & McCabe, P. F. (2020). Cannabis sativa. Current Biology, 30(1), R8-R9.
- Schubert, D., Kepchia, D., Liang, Z., Dargusch, R., Goldberg, J., & Maher, P. (2019). Efficacy of cannabinoids in a pre-clinical drug-screening platform for Alzheimer’s disease. Molecular neurobiology, 56, 7719-7730. [CrossRef]
- Sheedy, F. J., Grebe, A., Rayner, K. J., Kalantari, P., Ramkhelawon, B., Carpenter, S. B., Becker, C. E., Ediriweera, H. N., Mullick, A. E., & Golenbock, D. T. (2013). CD36 coordinates NLRP3 inflammasome activation by facilitating intracellular nucleation of soluble ligands into particulate ligands in sterile inflammation. Nature immunology, 14(8), 812-820. [CrossRef]
- Sheng, W. S., Hu, S., Min, X., Cabral, G. A., Lokensgard, J. R., & Peterson, P. K. (2005). Synthetic cannabinoid WIN55, 212-2 inhibits generation of inflammatory mediators by IL-1β-stimulated human astrocytes. Glia, 49(2), 211-219.
- Shinjyo, N., & Di Marzo, V. (2013). The effect of cannabichromene on adult neural stem/progenitor cells. Neurochemistry International, 63(5), 432-437. [CrossRef]
- Sido, J. M., Jackson, A. R., Nagarkatti, P. S., & Nagarkatti, M. (2016). Marijuana-derived Δ-9-tetrahydrocannabinol suppresses Th1/Th17 cell-mediated delayed-type hypersensitivity through microRNA regulation. Journal of molecular medicine, 94(9), 1039-1051.
- Sinha, D., Bonner, T. I., Bhat, N. R., & Matsuda, L. A. (1998). Expression of the CB1 cannabinoid receptor in macrophage-like cells from brain tissue: immunochemical characterization by fusion protein antibodies. Journal of neuroimmunology, 82(1), 13-21. [CrossRef]
- Soundara Rajan, T., Giacoppo, S., Scionti, D., Diomede, F., Grassi, G., Pollastro, F., Piattelli, A., Bramanti, P., Mazzon, E., & Trubiani, O. (2017). Cannabidiol activates neuronal precursor genes in human gingival mesenchymal stromal cells. Journal of cellular biochemistry, 118(6), 1531-1546. [CrossRef]
- Stewart, C. R., Stuart, L. M., Wilkinson, K., Van Gils, J. M., Deng, J., Halle, A., Rayner, K. J., Boyer, L., Zhong, R., & Frazier, W. A. (2010). CD36 ligands promote sterile inflammation through assembly of a Toll-like receptor 4 and 6 heterodimer. Nature immunology, 11(2), 155-161. [CrossRef]
- Tahir, M. N., Shahbazi, F., Rondeau-Gagné, S., & Trant, J. F. (2021). The biosynthesis of the cannabinoids. Journal of cannabis research, 3(1), 1-12. [CrossRef]
- Ubeed, H. M. S. A., Bhuyan, D. J., Alsherbiny, M. A., Basu, A., & Vuong, Q. V. (2022). A Comprehensive Review on the Techniques for Extraction of Bioactive Compounds from Medicinal Cannabis. Molecules, 27(3). [CrossRef]
- Valdeolivas, S., Navarrete, C., Cantarero, I., Bellido, M. L., Muñoz, E., & Sagredo, O. (2015). Neuroprotective properties of cannabigerol in Huntington’s disease: studies in R6/2 mice and 3-nitropropionate-lesioned mice. Neurotherapeutics, 12(1), 185-199.
- Van Cleemput, M., Cattoor, K., De Bosscher, K., Haegeman, G., De Keukeleire, D., & Heyerick, A. (2009). Hop (Humulus lupulus)-derived bitter acids as multipotent bioactive compounds. Journal of Natural Products, 72(6), 1220-1230.
- Vigli, D., Cosentino, L., Raggi, C., Laviola, G., Woolley-Roberts, M., & De Filippis, B. (2018). Chronic treatment with the phytocannabinoid Cannabidivarin (CBDV) rescues behavioural alterations and brain atrophy in a mouse model of Rett syndrome. Neuropharmacology, 140, 121-129. [CrossRef]
- Walter, L., Dinh, T., & Stella, N. (2004). ATP induces a rapid and pronounced increase in 2-arachidonoylglycerol production by astrocytes, a response limited by monoacylglycerol lipase. Journal of Neuroscience, 24(37), 8068-8074. [CrossRef]
- Walter, L., Franklin, A., Witting, A., Möller, T., & Stella, N. (2002). Astrocytes in culture produce anandamide and other acylethanolamides. Journal of Biological Chemistry, 277(23), 20869-20876. [CrossRef]
- Walter, L., Franklin, A., Witting, A., Wade, C., Xie, Y., Kunos, G., Mackie, K., & Stella, N. (2003). Nonpsychotropic cannabinoid receptors regulate microglial cell migration. Journal of Neuroscience, 23(4), 1398-1405. [CrossRef]
- Wang, Q., Dong, X., Zhang, R., & Zhao, C. (2021). Flavonoids with potential anti-amyloidogenic effects as therapeutic drugs for treating Alzheimer’s disease. Journal of Alzheimer's Disease, 84(2), 505-533. [CrossRef]
- Wang, W., Wu, N., Zu, Y., & Fu, Y. (2008). Antioxidative activity of Rosmarinus officinalis L. essential oil compared to its main components. Food chemistry, 108(3), 1019-1022. [CrossRef]
- Werz, O., Seegers, J., Schaible, A. M., Weinigel, C., Barz, D., Koeberle, A., Allegrone, G., Pollastro, F., Zampieri, L., & Grassi, G. (2014). Cannflavins from hemp sprouts, a novel cannabinoid-free hemp food product, target microsomal prostaglandin E2 synthase-1 and 5-lipoxygenase. PharmaNutrition, 2(3), 53-60. [CrossRef]
- Wirenfeldt, M., Babcock, A. A., & Vinters, H. V. (2011). Microglia–insights into immune system structure, function, and reactivity in the central nervous system. Histology and histopathology, Vol. 26, nº 4 (2011).
- Wu, W., Zhang, R., McClements, D. J., Chefetz, B., Polubesova, T., & Xing, B. (2018). Transformation and speciation analysis of silver nanoparticles of dietary supplement in simulated human gastrointestinal tract. Environmental science & technology, 52(15), 8792-8800. [CrossRef]
- Xi, Z.-X., Peng, X.-Q., Li, X., Song, R., Zhang, H.-Y., Liu, Q.-R., Yang, H.-J., Bi, G.-H., Li, J., & Gardner, E. L. (2011). Brain cannabinoid CB2 receptors modulate cocaine's actions in mice. Nature neuroscience, 14(9), 1160-1166.
- Yousaf, M., Chang, D., Liu, Y., Liu, T., & Zhou, X. (2022). Neuroprotection of Cannabidiol, Its Synthetic Derivatives and Combination Preparations against Microglia-Mediated Neuroinflammation in Neurological Disorders. Molecules, 27(15), 4961. https://mdpi-res.com/d_attachment/molecules/molecules-27-04961/article_deploy/molecules-27-04961.pdf?version=1659606660. [CrossRef]
- Zamberletti, E., Gabaglio, M., Piscitelli, F., Brodie, J. S., Woolley-Roberts, M., Barbiero, I., Tramarin, M., Binelli, G., Landsberger, N., & Kilstrup-Nielsen, C. (2019). Cannabidivarin completely rescues cognitive deficits and delays neurological and motor defects in male Mecp2 mutant mice. Journal of Psychopharmacology, 33(7), 894-907. [CrossRef]
- Zhang, Z., Yang, C., Dai, X., Ao, Y., & Li, Y. (2017). Inhibitory effect of trans-caryophyllene (TC) on leukocyte-endothelial attachment. Toxicology and Applied Pharmacology, 329, 326-333. [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



