
Submitted:
13 December 2023
Posted:
14 December 2023
You are already at the latest version
Abstract
Abiotic stress is an adverse environmental factor that severely affects plant growth and development, and plants have developed complex regulatory mechanisms to adapt to these unfavourable conditions through long-term evolution. In recent years, many transcription factor families of genes have been identified to regulate the ability of plants to respond to abiotic stresses. Among them, AP2/ERF (APETALA2/ethylene responsive factor) family is a large class of plant-specific that regulate plant response to abiotic stresses and can also play a role in regulating plant growth and development. This paper reviews the structural features and classification of AP2/ERF transcription factors, that transcriptional regulation, reciprocal proteins, downstream genes and hormone-dependent signalling and hormone-independent signalling pathways are involved in the response to abiotic stress. The study of AP2/ERF transcription factors and interacting proteins, as well as the identification of their downstream target genes, can provide us with a more comprehensive understanding of the mechanism of plant action in response to abiotic stress, which can improve the tolerance ability of plants in abiotic stress, and provide more theoretical basis for rice to obtain more yields under abiotic stress.
Keywords:
AP2/ERF transcription factor
; abiotic stress
; target gene
; hormone signalling
1. Introduction
Plants encounter a variety of stresses during their growth and development from seed germination to full maturity. One type of stress is biotic stress due to plant pathogens and the other is abiotic stress due to environmental factors. These two types of stress can trigger a series of morphological, physiological or biochemical changes in plants, affect plant growth and development, and can cause plant death in severe cases [1,2,3]. Abiotic stresses are the most important limiting factors for production, such as drought, high salt, low temperature, high temperature, nutrient stress and heavy metals. With the global climate change, the impact of these abiotic stresses on plants is increasing and has caused the reduction of global production of major crop and ultimately have a serious impact on global food security [4,5].
Abiotic stresses are environmental factors that can severely affect plant yield, so it is important to improve plant tolerance in response to abiotic stresses [6,7]. Plants have evolved a series of complex response mechanisms for normal growth in unfavourable environments, such as through the perception of abiotic stress signals, hormone -dependent signalling and hormone-independent signalling channel signal transduction, induction of abiotic stress gene expression and further activation of physiological and metabolic responses [7,8,9]. The gene products of plants in response to abiotic stresses are divided into two categories: The first type is regulatory proteins, which consist of protein factors that regulate downstream signal transduction and the expression of abiotic stress-responsive genes. The second type is functional proteins, which have a direct role in plant adaptation to abiotic stresses [10]. Regulatory proteins include protein kinases, phosphokinases and transcription factors (AP2, NAC, MYB, WRKY and bZIP transcription factors) [11,12,13]. Functional proteins include proline, soluble sugars, late embryogenesis abundant protein (LEA), dehydrins, Superoxide Dismutase (SOD), peroxidase (POD) and Catalase (CAT), water channel proteins etc. These proteins can mitigate damage to plant cells from various abiotic stresses by maintaining plant cell expansion pressure, scavenging reactive oxygen species and protecting the structure of intracellular biomolecules [14,15].
Transcription factors are proteins that regulate downstream gene expression by binding to specific sequences in DNA or other protein complexes. Transcriptional regulation refers to the combination of transcription factors with cis-acting elements upstream of stress-responsive genes to activate or repress gene expression during the process of gene expression [16,17]. Abiotic stresses such as drought, low temperature and high salt lead to an increase in abscisic acid (ABA) biosynthesis, which activates the binding of transcription factors to cis-acting elements in specific sequences in the promoter regions of the corresponding downstream stress-responsive genes, regulating gene expression and thus modulating plant tolerance in response to abiotic stresses [18,19]. As one of the plant-specific transcription factor families, the AP2/ERF family of transcription factors is of great significance for plant survival and development. Nowadays, with the release of the whole genome sequences of many plants, their functional regulatory networks in crops such as rice (Oryza sativa L.), maize (Zea mays L.), and soybean (Glycine max L.) will be gradually revealed. The article discusses the structural features of AP2/ERF transcription factor, binding elements, transcriptional regulation and interaction proteins, as well as the progress of research on AP2/ERF in regulating the response to abiotic stresses, and provides references for future research on AP2/ERF transcription factor to improve the tolerance of plants to cope with abiotic stresses [20,21,22].
1.1. Classification and structural classification of AP2/ERF transcription factors
The AP2/ERF transcription factor family is a large group of plant-specific transcription factors that regulate plant growth, development and abiotic stress response. Much AP2/ERF family genes were identified in the plant genome, they can be divided into five main groups based on the number of AP2/ERF structural domains, namely AP2 (APETALA2), ERF (ethylene responsive factor), DREB (dehydration-responsive element binding protein), Soloist and RAV (related to ABI3/VP1) (Supplemental Table S1). For example, AP2/ERF transcription factor genes OsDREB2B, OsRPH1, OsEATB, and OsAP2-39 are able to affect rice growth and development, and consequently plant height, by regulating the expression of Gibberellin (GA) metabolic genes [23,24,25,26]. Present-day studies are more extensive and in-depth for the AP2, DREB and ERF subfamilies, whereas very few studies have been reported on the Soloist subfamily, whose nucleotide sequences are known to be highly conserved in most plants [27] (Figure 1). The structure of AP2/ERF transcription factors is characterized by four major functional regions, namely the DNA-binding domain, the transcription regulation domain, the oligomerization site and the nuclear localization signal (NLS), and the AP2/ERF binding domain is highly conserved [28]. The AP2/ERF transcription factor structural domain contains 60-70 amino acid residues, forming a typical three-dimensional structure according to three β-folds and one α-helix. The YRG and RAYD elements in the AP2 domain play an important role in DNA binding activity. The YRG element at the N-terminal end of the domain consists of about 19-22 hydrophilic amino acid residues and promotes DNA binding through bases and hydrophilic groups; the RAYD element at the C-terminal end of the domain consists of about 42-43 residues and mediates protein-protein interactions through the α-helices or interacts with DNA through the hydrophobic surface of the α-helices [29]. Many studies have shown that AP2/ERF transcription factors are highly structurally similar, which may lead to a large amount of gene redundancy in the AP2/ERF family of transcription factors [23,24].
1.2. Cis-acting elements recognized by AP2/ERF transcription factors
In previous promoter region analyses of AP2/ERF transcription factor-regulated genes, scientists have identified much AP2/ERF transcription factors capable of specifically binding to stress-responsive gene initiator cis-elements in the promoter region [30,31,32]. ERF subfamily members can participate in the regulation of abiotic stress by binding to the ethylene response element (ERE) (GCC-box, core sequence AGCCGCC). Members of the DREB subfamily of transcription factors specifically recognize and bind to a DRE/CRT element (dehydration responsive element/C-repeat, core sequence A/GCCGAC) in the promoter region of another gene, regulating the expression of response genes related to drought, low temperature and salt stress [33,34,35,36]. AP2/ERF transcription factors bind not only to both DRE/CRT and ERE elements, but also to other cis-elements such as coupling element 1 (CE1, TGCCACCG), coupling element 3 like (CE-3like, CGCG) and hypoxia-responsive promoter element (HRPE). AP2/ERF transcription factors also bind to these cis-elements, for example CAACA, ATCTA, CATGCA, CGNCCA and ATCGAG [37,38,39,40].
2. AP2/ERF transcriptional regulation and interacting protein under abiotic stresses
2.1. AP2/ERF transcriptional regulation under abiotic stresses
Under abiotic stresses, some genes related to adversity in plants can bind to cis-acting elements conserved in AP2/ERF transcription factors to regulate the expression of their transcription factor gene, which include low-temperature responsive elements, heat shock responsive elements, and ABA responsive elements, etc. It has been shown that heat shock factor 1 (HSF1) and ABA responsive element binding protein (ABRE) can bind to the heat shock response element in the DREB2A promoter, respectively, and regulate the expression of their transcription factor gene, thereby altering the plant's ability to tolerate abiotic stresses [41,42,43,44]. Song et al. showed that the interaction of the APETALA2/ EREBP-type transcription factor AtERF7 with the protein kinase PKS3 is involved in the regulation of the plant ABA response. AtERF7 binds to the GCC box and is able to inhibit gene transcription. AtERF7 interacts with the transcriptional corepressor AtSin3, which in turn may interact with the histone deacetylase HDA19. HDA19 and AtSin3 enhance the transcriptional repression activity of AtERF7. Thus AP2/ERF transcription can be regulated by histone modifications such as phosphorylation, ubiquitination, methylation and acetylation by modulating the spatial state of chromatin [45,46,47,48,49]. Kavas et al. identified 180 AP2-ERF superfamily genes in Phaseolus vulgaris. Micro-RNA target transcript analyses identified in computer simulations identified almost all PvAP2-ERF genes as miRNA targets in 44 different plant species, the most abundant target gene was PvERF20, PvERF25, PvERF62, PvERF78, PvERF113, PvERF173 [50].
2.2. AP2/ERF interacting protein under abiotic stresses
Transcription factor (TF), also known as trans-acting factor, is a DNA-binding protein that specifically interacts with cis-acting elements of eukaryotic genes and activates or inhibits gene transcription. Many studies have shown that AP2/ERF transcription factors interact with other transcription factors to form protein complexes or can directly bind to the promoters of their downstream target genes to repress or promote gene expression [51,52]. AP2/ERF family transcription factor OsDREB2B was able to regulate the expression of OsAP2-39 by binding to its promoter, and OsDREB2B interacted with OsWRKY21 to regulate the expression of GA metabolism genes and inhibit GA synthesis, leading to a decrease in GA content and thus exerting a negative effect on rice growth and development [23]. Overexpression of the AP2/ERF transcription factor OsRPH1 resulted in reduced plant height and OsRPH1 interacted with the blue light receptor OsCRY1b [24]. Tiwari et al. identified a short "EDLL" motif that is present in AtERF98/TDR1 and other branch members from the same AP2 subfamily. This motif has a unique arrangement of acidic amino acids and hydrophobic leucines, and functions as a strong activation domain that partially overcomes the repression conferred by the AtHB2 transcription factor, which contains an ERF-associated amphiphilic repressor (EAR) motif [53]. AtERF7 overexpressed plants have reduced sensitivity of defence cells to ABA and increased water loss, ERF3 interacts with the subunit of the histone deacetylase complex SAP18 (SIN3 associated polypeptide P18), and co-recruits HDA19 to form a complex which inhibits the expression of the associated genes [54]. Franco-Zorrilla et al. analysed co-regulated gene and transcriptome data from transcription factor mutants. The results indicate that at least one target sequence for each transcription factor is functionally important and that the function of a transcription factor as an activator or a repressor can be predicted from specific DNA sequences [55]. So, genes co-regulated by AP2/ERFs transcription factors are also enriched for AP2-ERFs target genes, and future analyses of homologous sequences by scientists will help to identify putative targets of transcription factors and predict their biological functions.
3. AP2/ERF by participating in the regulation of hormone-mediated abiotic stresses
Phytohormone also known as plant natural hormones or plant endogenous hormones, refers to a number of organic compounds produced by the plant body in minute quantities that can regulate (promote, inhibit) their own physiological processes. The main role of phytohormones is to regulate the ability of plants to resist the adverse external environment and plant growth and development. Studies have shown that AP2/ERF transcription factors can synergies with hormone signaling to form a cross-regulatory network, e.g., by participating in growth and development and abiotic stress responses mediated by the plant hormones abscisic acid (ABA), gibberellin (GA), auxin (IAA), ethylene (ET), brassinosteroid (BR) and cytokinin (CTK) [56,57,58,59,60].
3.1. AP2/ERF involved in ABA-mediated stress response
The main role of ABA is to promote the shedding of organs such as plant leaves, fruits and calyxes and to regulate plant growth and development. ABA is one of the key hormones in response to abiotic stresses such as drought, salt, cold and heat. ABA promotes the synthesis of osmotic substances and protects against abiotic stresses by regulating stomatal opening and closure and root architecture [61,62]. Overexpression of OsERF71 plants reduced water loss resulting in increased tolerance to drought stress. OsERF71 was able to regulate the expression of ABA-responsive and proline biosynthesis genes under drought stress, resulting in increased sensitivity to exogenous ABA treatment and proline accumulation [63]. Cheng et al. identified 229 AP2/ERF genes in the latest maize reference genome, of which 32 ZmAP2/ERFs regulate biotic stress and 24 ZmAP2/ERFs are involved in abiotic stress response. ZmDREB39 and ZmDREB89 were up-regulated in response to ABA [64]. Xiong et al identified a total of 135 TkERF genes from Trichosanthes kirilowii. A co-expression network constructed using transcriptome data from different flowering stages showed that 67 of the AP2/ERF genes were associated with members of the abscisic acid signalling pathway. Seventeen genes were found to be up-regulated when tissue culture seedlings were treated with ABA, suggesting that some members of the TkERF gene family may be involved in phytohormone signalling pathways. Twenty genes were up-regulated under PEG treatment, suggesting that these selected genes may be involved in plant drought stress [65]. TaAP2-10 is able to be induced by abiotic stress and ABA hormone treatment [66]. The CBF (C-repeat binding factor) subfamily of transcription factors belongs to the AP2/ERF (Apetala 2/ethylene response factor) family of transcription factors. Research shows cis-acting elements in the promoter region of BpCBFs that are associated with environmental stress and hormones. Most of these transcription factors were found to be responsive to ABA or salt stress in different plant tissues after treatment using ABA or salt [67].
3.2. AP2/ERF involved in GA-mediated stress response
The main physiological role of GA is to promote elongation of plant cells, increase plant height, promote growth and development of lateral buds, promote flower bud differentiation and flowering, and inhibit aging and shedding of leaves. Li et al. identified a total of 218 AP2/ERF genes in the sugarcane genome, and the presence of multiple cis-regulatory elements (CREs) in the SsAP2/ERF promoter was associated with abiotic stress, suggesting that SsAP2/ERF activity may contribute to the adaptation of sugarcane to environmental changes. SsSoloist4 was most significantly up-regulated in response to treatment with the exogenous hormone GA, suggesting that this gene may play a role in the response to GA-associated [68]. OsDREB2B is an AP2/ERF family transcription factor, overexpression of OsDREB2B in rice can significantly improve rice drought tolerance [69]. Ma et al. found that OsDREB2B overexpression resulted in shorter plant height, and the length of the second leaf sheath in overexpression plants was restored to that of the wild type by exogenous GA3 application. Expression of GA biosynthesis genes was altered in OsDREB2B-OE plants [23]. Fu et al. found that overexpression of ZmEREB20 in Arabidopsis thaliana enhanced ABA sensitivity and caused delayed seed germination under salt stress by regulating ABA and GA-related genes [70]. Zhang et al. found that about 49 genes containing complete AP2/ERF structural domains were identified from the Taxus x media (Yew tree) transcriptome database. 9 of these TmERF genes could respond to low temperature and hormone treatments, and the expression of TmERF5, TmERF14, and TmERF36 was elevated under GA treatment. These suggests that they may be involved in crosstalk between abiotic stress response signalling pathways [71].
3.3. AP2/ERF involved in IAA-mediated stress response
The main effects of IAA are to promote the division and elongation of plant cells, the growth of roots and stems, and to promote the development and ripening of fruits, to regulate the form and growth direction of plants. Cai et al. demonstrated a regulatory role for the APETALA2/ethylene response factor (AP2/ERF) transcription factor OsERF096 in the cold stress response. Targeted metabolomics analyses revealed that OsERF096 can respond to cold stress by regulating IAA accumulation and signalling [72]. Huang et al. found that overexpression ERF012 showed resistance to temperature, drought, salt and heavy metal stresses, respectively. Overexpression ERF012 inhibited root growth and promoted root hair development and leaf senescence. The application of exogenous IAA effectively mitigated this effect. ERF012 may down-regulate its target genes AtC4H and At4CL1 (key genes for phenylpropanoid metabolism and cell wall formation), resulting in reduced IAA accumulation leading to leaf senescence [73]. Overexpression SlERF.B1 showed significantly higher sensitivity to salt treatment at both phenotypic and physiological levels. Overexpression SlERF.B1 plants showed low tolerance to mannitol and drought stresses. SlERF.B1 expression was induced by salt, mannitol, cold, heat, and ACC treatments, but was inhibited by ABA, IAA, and 1-MCP treatments [74].
3.4. AP2/ERF involved in ET-mediated stress response
ET can promote fruit ripening and leaf senescence, induces adventitious roots and root hairs, breaks the dormancy of plant seeds and buds, and inhibits flowering in many plants (but induces and promotes flowering in pineapple and its congeners). Ethylene, one of the six plant hormones, also plays a role in a variety of stress responses such as salt, low temperature and flooding [75]. Sub1A is a rice flooding tolerance gene, the expression of which was up-regulated under flooding conditions, while ET treatment induced its expression, but GA treatment did not. Flood-tolerant rice varieties containing the Sub1A gene synthesised ET in the plants under flooded conditions, and ethylene promoted the degradation of ABA on the one hand, and the expression of Sub1A on the other [76,77]. The AP2/ERF transcription factor RAP2.2 in Arabidopsis thaliana belongs to the same subfamily as the rice (Oryza sativa) submergence tolerance gene SUB1A. RAP2.2 is induced by ET in shoots and functions in ET-controlled signal transduction pathways, overexpression RAP2.2 lines showed increased survival under hypoxia stress [78].
3.5. AP2/ERF involved in BR-mediated stress response
BR has an important role in plant growth and development and, together with other plant hormones, is involved in the regulation of many aspects of plant development, including stem and leaf growth, root growth, vascular tissue differentiation, fertility, seed germination, maintenance of apical dominance and plant photomorphogenesis [79]. Liu et al. show that an AP2/ERF family transcription factor ERF72 may be a candidate gene for cross-interaction between the BR signalling pathway and stress response. In Arabidopsis, ERF72/RAP2.3 antagonised BZR1 (brassinazole-resistant 1) and ARF6 (auxin responsive factor 6) and inhibited hypocotyl elongation [80]. Schmitz et al. found SUB1A differentially regulates the expression of BR synthesis-related genes during plant flooding, activates BR biosynthesis and signal transduction, induces the expression of GA2ox7, a key gene for GA degradation, and thus controls GA levels in rice plants [81].
3.6. AP2/ERF involved in CTK-mediated stress response
The main physiological role of CTK is to promote cell division and prevent leaf senescence. The senescence and yellowing of green plant leaves is due to the decomposition of proteins and chlorophyll in them; while CTK can maintain protein synthesis, thus keeping the leaves green, promoting the differentiation of buds, cell enlargement, the development of lateral buds and the removal of apical dominance [82]. Moreover, CTK also plays an important role in plant response to abiotic stresses [83]. AP2/ERF-N22(2) has an AP2 structural domain consisting of 55 amino acid residues and a set of acidic amino acid residues in the C-terminal region that can act as a trans-activating structural domain. Moreover, AP2/ERF-N22(2) belongs to group VI L. Members of group VI L Arabidopsis have been shown to be involved in the response to cytokinins under drought stress [84]. Cytokinin response factor (CRF) is an AP2/ERF family transcription factor that regulates plant cotyledon and embryo developmental processes and is also involved in cytokinin signalling [85,86]. In Arabidopsis, cytokinin response factor 6 (CRF6) is an AP2/ERF transcription factor that is induced by CTK. CRF6 is highly expressed in the veins of mature leaves, promotes CTK synthesis, and is induced by abiotic stress [87,88]. Zwack et al. showed that cytokinin response factor 4 (CRF4) can be induced by cold for a short period of time, especially in root and shoot tissues. altered transcriptional expression of genes of the cold signalling pathway in C-repeat binding factors (CBF) and COR15a in crf4 mutants and overexpression CRF4 lines suggests that CRF4 may be involved in this pathway [89]. The specific regulatory mechanism of CRF is still unclear, so future identification of CRF target genes and upstream signalling molecules could help explain this.
4. Role of AP2/ERF transcription factors in response to abiotic stresses(Not depend on hormone signalling pathway)
Stresses such as drought, high salt, high temperature, low temperature, nutrients and heavy metals are the major abiotic stress factors affecting plant growth and development, crop yield and quality. In the course of plant evolution, complex defence mechanisms have been developed to adapt to abiotic stresses, so that plants have improved their ability to tolerate abiotic stresses. Studies have shown that the AP2/ERF family of transcription factors is widely involved in regulating the plant response to various abiotic stresses [90] (Figure 2).
4.1. AP2/ERF transcription factors in response to drought stress
Water is a vital resource for the survival of all life and has played an important role in the evolution of life. Water is also one of the raw materials for photosynthesis in green plants and if there is a lack of water, the plant's photosynthesis will be weakened. Leaves will wilt and in severe cases can lead to the death of the plant [91]. Yu et al. found that overexpression of the AP2/ERF family gene TaERF-6-3A increased sensitivity to drought and salt stress in Arabidopsis. Expression of stress-related and antioxidant-related genes was down-regulated in TaERF-6-3A overexpressing plants [92]. Genome-wide analysis of the AP2/ERF genes in Pisum sativum (L.) identified 153 AP2/ERF genes. Among them, Jarambasa et al. found that DREB2A, DREB2C, DREB2E, and DREB2F were induced in leaves under drought stress [93]. Kumar et al. showed overexpressing Os-AP2/ERF-N22 line showed higher relative water content, membrane stability index, wax content, osmotic potential, stomatal conductance and transpiration rate activities [94]. Kabir et al. demonstrate a total of 119 CoAP2/ERF genes were identified from the dark jute genome. Among them, CoDREB-11, CoDREB-14 and CoRAV-01 genes were significantly up-regulated under salinity and drought stress conditions [95]. Kong et al. demonstrate that the osmotic stress-induced PtoERF15 and its target gene PtoMYC2b, which is involved in mediating blood vessel size, density and cell wall thickness in response to drought in poplar, were identified and characterised. overexpression of PtoERF15 contributes to the maintenance of stem water potential, thereby increasing drought tolerance [96].
4.2. Molecular mechanisms of AP2/ERF associated with salt stress
Soil salinity affects around 6% of the world's land and 23% of arable land, causing considerable economic losses through crop stress and reduced yields. Because salinity plays a vital role in plant growth, above a certain limit, excess soluble salts will have a toxic effect on plants. That can greatly affect the growth and development of plants, ultimately resulting in reduced yields [97]. The AP2/ERF transcription factor OsERF19 was identified in rice by Huang et al. OsERF19 expression was inhibited by salt stress. Overexpressing OsERF19 line increased the tolerance of plants to salt stress. In addition, OsLEA3, OsNHX1, OsHKT6, and OsOTS1 genes were up-regulated in overexpressing lines when plants were subjected to salt stress [98]. Within the AP2/ERF family of transcription factors AtERF71/HRE2 is known to be involved in hypoxia and osmotic stress responses [99,100]. Seok et al. dissected the HRE2 promoter and showed that the -116 to -2 region is responsible for hypoxia and salt stress responses. This region contains both positive and negative cis-regulatory elements that may regulate HRE2 expression under salt stress [101].
4.3. AP2/ERF involved in plant response to temperature stress
4.3.1. AP2/ERF and high temperature stress
Temperature is the main environmental factor affecting plant growth and development and the quality of life of the fruit after harvest, appropriate temperatures promote plant growth. The main damage of high temperature is to make the plant growth and development is hindered, the plant is short, the leaves and stems occur localised burns, the amount of flowers is reduced. High temperatures also encourage increased transpiration, disrupting the water balance and causing the plant to wilt and die [102]. He et al. treated the heat tolerance of T2 (Jinxibai) and T21 (Taizhongxianxuan 2 hao) at 45°C and found that T21 was sensitive to heat stress during the seedling stage. In which 60 transcription factors, including AP2/ERF, NAC, HSF, WRKY and C2H2 family members, were involved in the heat stress response [103]. Zhang et al. found that PlTOE3, a (AP2/ERF) transcription factor, can specifically activate the PlTDC promoter. High-temperature stress can affect the transcriptional level of PlTOE3. Overexpression of PlTOE3 in tobacco enhanced plant melatonin production and heat stress tolerance, whereas silencing of PlTDC expression gave the opposite results [104]. The SHN/WIN evolutionary branch of the AP2/ERF transcription factor family is involved in many important processes. Djemal et al. showed that overexpression of HvSHN1 increased tolerance to heat stress. Under heat stress, the HvSHN1 overexpression strain exhibited higher superoxide dismutase (SOD) and catalase (CAT) activities as well as lower levels of malonic dialdehyde (MDA) and hydrogen peroxide (H2O2) than the wild type [105].
4.3.2. AP2/ERF and low temperature stress
Symptoms of low temperature are discolouration of plant leaves, necrosis and the appearance of spots on the surface of the plant, making the plant grow slowly and other changes in morphological characteristics, photosynthetic rate is significantly reduced, resulting in lower yields and lower quality. However, low temperature stress not only leads to a reduction in plant yield, but in severe cases it can also cause plant death [106]. Xie et al. research shows that seventy AP2/ERF transcription factors were identified in Tetrastigma hemsleyanum, and ThERF5, ThERF31, ThERF46 and ThERF55 were found to be more sensitive to cold stress at low temperature. Overexpression of ThERF46 line and RNA interference strain indicated that ThERF46 has a specific tolerance to cold stress [107]. Zhao et al. through phylogenetic analysis, 184 AP2/ERF genes were identified in the genome of Juglans mandshurica. QRT-PCR expression profiles showed that 14 JmAP2/ERF genes were associated with cold resistance, of which seven and five were significantly up-regulated under cold stress [108]. Zhang et al. found the expression and roles of 27 AP2/ERF genes in Eribotrya japonica were investigated, of which EjERF27, EjERF30, EjERF36, and EjERF39 were significantly induced by storage at low temperature, but suppressed by cold-conditioning treatments [109]. Ren et al. identified RAP2.4 which is a member of the AP2/ERF transcription factors family in Chrysamthemum lavandulifolium. The activities of superoxide dismutase and peroxidase, and proline content in leaves in the four overexpression lines were higher than those in the wild type (WT), whereas the electrical conductivity and malondialdehyde content were decreased, indicating that the tolerance of plants with ClRAP2.4 overexpression to cold stress was increased. SOD and POD activities as well as proline content were higher in the overexpressing ClRAP2.4 strain than in the wild type, whereas conductivity and malondialdehyde content were reduced, suggesting that overexpression of ClRAP2.4 is increased in plants tolerant to cold stress [110].
4.4. Role of Plant AP2/ERF in Response to Nutritional element stress
Nitrogen (N), phosphorus (P), and potassium (K), which are essential nutrients for plants, thay play very important physiological roles in plant growth and development. N is found in both proteins and nucleic acids, and proteins in turn are the basic substances that make up protoplasm [111].
N is also a constituent of chlorophyll, which is indispensable for photosynthesis in higher green plants, and is therefore essential for plant photosynthesis [112]. RNA-seq analysis by Joshi et al. showed that N stress induced most of the transcriptome changes in spinach roots , identifying 1,346 differential expressed genes (DEGs). in the presence of high N in leaf tissues, a subset of AP2/ERF family member transcription factors were all overexpressed in tissues in response to N perturbation [113]. Bacterial accommodation within living plant cells was restricted to nitrogen-fixing rhizobium symbiosis, bacterial uptake is mediated by tubular structures called infection threads. Cerri et al. identified an AP2/ERF family of transcription factor genes ERN1. Rhizobium primordia were formed following mutation of the ern1 gene in Lotus japonicus, but the majority remained uninfected and bacterial entry into the root epidermis via infection threads was eliminated [114].
P is a constituent of the nucleus and nucleic acids, which have a special role in plant life and hereditary processes [115]. Most terrestrial plants establish symbiotic relationships with arbuscular mycorrhizal fungi (AMF), providing them with lipids and sugars in exchange for phosphorus and nitrogen. zhang et al. identified the AP2/ERF transcription factor MtERM1 as being able to bind directly to the AW-box and AW-box-like cis-elements in the MtSTR2 and MtSTR promoters, which are required for host lipid efflux and tuft development [116]. Chen et al. identified an AP2/ERF transcription factor gene, PalERF2, from Populus alba var. pyramidalis, and PalERF2 overexpression lines enhanced tolerance to Pi deficiency, but knockout PalERF2 lines showed the opposite phenotype . In addition, overexpression of PalERF2 upregulated the expression level of phosphate starvation-induced (PSI) genes and increased phosphate uptake under drought conditions [117].
K is an activator of many of the enzymes of photosynthesis and enhances the activity of the enzymes and hence promotes photosynthesis. K also functions to control the opening and closing of the stomata of the plant and hence facilitates the conservation of water by the plant [118]. Kim et al. showed that the AP2/ERF family transcription factor RAP2.11 was identified as a component of the low potassium response. RAP2.11 regulates the expression of the high-affinity K+ uptake transporter protein AtHAK5 under low K+ conditions and also contributes to a coordinated response to low potassium conditions by regulating other genes in the low K+ signalling cascade [119]. Chen et al. demonstrated that the AP2/ERF family transcription factor OsERF106 was expressed in germinating seeds, primary roots, and developing flowers. Overexpression of OsERF106 resulted in stunted growth, relatively high levels of MDA and reactive oxygen species (ROS), reduced CAT activity, and excessive accumulation of sodium (Na+) and potassium (K+) ions in transgenic rice [120].
4.5. AP2/ERF Involved in Plant Response to heavy metals stress
Heavy metals in soil are a stress factor for plants, affecting their survival in various ways, including growth, development and reproduction. Excessive amounts of heavy metals affect photosynthesis in plants, thus causing symptoms such as yellowing and wilting of plant leaves, and reducing the efficiency of light energy utilisation in plants. High concentrations of heavy metals in the soil affect the ability of plants to absorb and utilise nutrients, leading to a reduction in the number of leaves, changes in the root system and a reduction in stem expansion [121,122,123,124]. Karanja et al. identified a total of 247 ERF family genes in the radish genome, and a portion of the AP2/ERF genes were preferentially expressed under drought and heat stress, whereas they were repressed under heavy metal stress [125]. Chen et al. identified a large number of key heavy metal Cd-induced DEGs containing transcription factors such as AP2/ERF, MYB, NAC and WRKY. DEGs are involved in pathways including antioxidants, heavy metal transport or detoxification, and AP2/ERF family transcription factors were suggested to play crucial roles in kenaf Cd tolerance [126]. Exposure of Solanum tuberosum L. to the heavy metals Cd/Pb/Zn/Ni/Cu by Tian et al. identified 181 potential StAP2/ERF genes, and the StAP2/ERF075, AP2/ERF077 and AP2/ERF126 genes were able to promote Cd accumulation and yeast growth (Cd detoxification phenotype) [127].
5. Conclusion and Prospects
Nowadays, there are many reports on the involvement of AP2/ERFs in the regulation of hormone signalling-mediated stress response. When plant is sufferred to abiotic stress, the inducing hormones (ABA and ET) and growth-promoting hormones (GAs, IAA, CTK, and BRs) carry out their defence against the adverse external environment through a mechanism regulated by AP2/ERFs (Figure 3) [128].
ABA Pathway: Under abiotic stress conditions when plants are subjected to water deficit, the rate-limiting ABA biosynthetic enzyme Nine-cisEpoxycarotenoid Dioxygenase (NCED) is rapidly up-regulated to promote ABA biosynthesis [129]. Subsequently ABA is sensed by ABA receptor (PYR/PLY/RCAR) that interact with Protein Phosphatases 2Cs (PP2Cs) as a ternary complex releasing the inhibition of SnRK2 kinases (SnRK2s) by PP2Cs. Active SnRK2s phosphorylates downstream substrate proteins, including AREBs/ABFs, ion channels, and nicotinamide adenine dinucleotide phosphate (NADPH) oxidases, thereby inducing ABA response [130,131]. ERF18/ORA47 activates the PP2C family phosphatase gene ABI2. Meanwhile, ABI1 acts upstream of ORA47 to activate ORA47, forming an ABI1-ORA47-ABI2 regulatory loop that inhibits ABA signalling as well as drought tolerance [39]. RAV1 inhibits root growth sensitivity to ABA by repressing ABI3, ABI4, and ABI5 expression. SnRK2.2, SnRK2.3, and SnRK2.6 also interact and phosphorylate RAV1 to inhibit transcriptional repression of target genes by RAV1 [132]. GA Pathway: In the absence of GA, DELLA inhibits GA responses. Abiotic stress leads to reduced GA content and signalling by inhibiting AP2/ERF-mediated GA biosynthesis enzymes. DREB1E and DREB1F lead to salt stress-induced growth retardation mainly by inhibiting GA20oxes [133]. Overexpressing CBF1 and ERF6 plants are sensitive to stress-induced growth retardation due to increased expression of GA2oxs and accumulation of DELLA. ERF11 promotes plant internode elongation by activating GA biosynthesis, and the expression of GA3ox1 and GA20oxs is increased in overexpressing ERF11 plants [134]. IAA Pathway: Huang et al. found that overexpression of ERF012 showed resistance to temperature, drought, salt and heavy metal stresses, respectively. Overexpression of ERF012 inhibited root growth and promoted root hair development and leaf senescence. The application of exogenous IAA effectively mitigated this effect. ERF012 may down-regulate its target genes AtC4H and At4CL1 (key genes for phenylpropanoid metabolism and cell wall formation), resulting in reduced IAA accumulation [73]. CTK Pathway: More than half of the CTK-responsive genes are regulated by both CRF and B-type ARRs (typical cytokinin-responsive transcription factors), and CRF6 also cooperates with CTK signalling to inhibit stress-induced leaf senescence through a common subset of CTK-regulated genes. CRF6 also represses CTK-related target genes involved in CTK biosynthesis, signalling and transport to mitigate the adverse effects of CTK on abiotic stresses [87,88]. BR Pathway: BR is sensed by the plasma membrane receptor kinase BRASSINOSTEROID INSENSITIVE 1 (BR1), which represses the negative regulator BRASSINOSTEROID INSENSITIVE 2 (BIN2), leading to the accumulation of the transcription factor BRASSINAZOLE-RESISTANT 2/BRI1-EMS-SUPPRESSOR 1 (BES1/BZR1) to regulate the response genes involved in the BR in response to plant growth and stress responses. BR positively regulates cold tolerance partly through the CBF-mediated cold response pathway, in which BZR1 binds and promotes CBF1/CBF2 expression in response to cold. Cold stress also promotes the accumulation of the unphosphorylated active form of BZR1 through unknown mechanisms [135,136]. ET Pathway: ET is synthesised by the rate-limiting enzyme ACC synthase (ACS), a major target for regulating ET production under stress conditions [137]. Binding of ET to its receptor ethylene-insensitive 1 (ETR1) inactivates constitutive triple response 1 (CTR1) kinase activity, releasing CTR1 from its inhibition of ethylene-insensitive 2 (EIN2). The C-terminus of EIN2 then translocates to the nucleus, activating ethylene insensitive 3 (EIN3) and the transcriptional cascade of ethylene-regulated genes [138,139]. ET represses CBF to negatively regulate cold stress, ET positively regulates ERF-VII-mediated flooding and inundation, and enhances salt tolerance through activation of ERF1 and ESE. The ET-insensitive mutants etr1, ein2, and ein3 show enhanced freezing tolerance. EIN3 represses CBF expression by directly binding to its promoter. Flooding causes hypoxia, which promotes ET production and activates the expression of a group of ERF-VIIs; however, ERF-VIIs regulate the hypoxic response partly through an ET-independent pathway. ERF-VII may also negatively regulate ET signalling and homeostasis through feedback regulation [140]. The involvement of AP2/ERFs in the regulation of hormone signalling-mediated stress response has been widely reported. But we do not know the exact regulatory mechanism, more scientific exploration is needed. Question 1: How do AP2/ERFs feedback regulate hormone biosynthesis and metabolism when they receive hormone signals? Question 2: How AP2/ERFs regulate the expression of downstream related genes by synergizing or antagonizing multiple hormone signal transduction groups. Question 3: Whether AP2/ERFs can interact with other family transcription factors to jointly regulate the expression of downstream related genes through hormone signal transduction. Explaining these three issues will help to systematically explain the regulatory network of AP2/ERFs in response to abiotic stress conditions in plants, and provide a theoretical basis for subsequent plant stress tolerance breeding projects.
Plants respond to abiotic stress not only through the above hormone signalling networks, but also a series of other complex signalling pathways and gene expression regulatory networks have been established during the long-term evolution of plants. When plants are affected by external stress, they are able to regulate their growth and development and their ability to cope with tolerance under abiotic stress. AP2/ERF family transcription factors can play a role in abiotic stress response through hormone-dependent or hormone-independent signalling pathways. In recent years, there have been many reports on the involvement of AP2/ERF family transcription factors in the response of plants to abiotic stresses (Table 1). But they are still focused on abiotic stresses, such as drought, high salt, high temperature, low temperature, the molecular mechanisms of AP2/ERF transcription factors in response to chemical reagent stress are rarely reported, for example, pesticides, car exhaust and haze etc. For exhaust and haze, the main component of which is sulphide, can cause plants to develop leaf scorch and impede growth and development. Excessive use of pesticides can damage the ecosystem and lead to a decline in crop yields. Therefore, it is important to study AP2/ERF in response to chemical stress in plants. Therefore, our in-depth study of the AP2/ERF transcription factors molecular mechanisms of plant responses to abiotic stresses, such as stress signal perception and transmission, transcriptional regulation and expression of response genes, can ensure the normal growth and development of plants in response to abiotic stresses, and ensure food production, which will contribute to the global food security and the improvement of the quality of life of humans.
Supplementary Materials
The following are available online at www.mdpi.com/xxx/s1.
Author Contributions
All the authors contributed to the present form of the manuscript. Z.M collected the data, drafted the manuscript; Z.M. and W.Z. edited; Z.M. and L.J. created figures and tables; Z.M. and L.J. and W.Z. supervised; Z.M. and L.J. and W.Z. finalized and approved the final version of manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by the National Natural Science Foundation of China (Grant No. 32201695). Open Access funding provided by the Max Planck Society.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no competing interests.
References
- Gupta, B.; Shrestha, J. Editorial: Abiotic stress adaptation and tolerance mechanisms in crop plants. Front Plant Sci. 2023, 14, 1278895. [Google Scholar] [CrossRef]
- Ma, Z.; Hu, L. MicroRNA: A Dynamic Player from Signalling to Abiotic Tolerance in Plants. Int J Mol Sci. 2023, 24, 11364. [Google Scholar] [CrossRef]
- Mittler, R.; Zandalinas, SI.; Fichman, Y.; Van Breusegem, F. Reactive oxygen species signalling in plant stress responses. Nat Rev Mol Cell Biol. 2022, 23, 663–679. [Google Scholar] [CrossRef]
- Colombage, R.; Singh, M.B.; Bhalla, P.L. Melatonin and Abiotic Stress Tolerance in Crop Plants. Int J Mol Sci. 2023, 24, 7447. [Google Scholar] [CrossRef]
- Shoji, T.; Yuan, L. ERF Gene Clusters: Working Together to Regulate Metabolism. Trends Plant Sci. 2021, 26, 23–32. [Google Scholar] [CrossRef] [PubMed]
- Nakabayashi, R.; Saito, K. Integrated metabolomics for abiotic stress responses in plants. Curr Opin Plant Biol. 2015, 24, 10–6. [Google Scholar] [CrossRef] [PubMed]
- Nadarajah, K.; Abdul, H.; Abdul, R.N. SA-Mediated Regulation and Control of Abiotic Stress Tolerance in Rice. Int J Mol Sci. 2021, 22, 5591. [Google Scholar] [CrossRef] [PubMed]
- Habibpourmehraban, F.; Atwell, B.J.; Haynes, P.A. Unique and Shared Proteome Responses of Rice Plants (Oryza sativa) to Individual Abiotic Stresses. Int J Mol Sci. 2022, 23, 15552. [Google Scholar] [CrossRef] [PubMed]
- Habibpourmehraban, F.; Wu, Y.; Masoomi-Aladizgeh, F.; Amirkhani, A.; Atwell, B.J.; Haynes, P.A. Pre-Treatment of Rice Plants with ABA Makes Them More Tolerant to Multiple Abiotic Stress. Int J Mol Sci. 2023, 24, 9628. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Zhu, J.; Gong, Z.; Zhu, J.K. Abiotic stress responses in plants. Nat Rev Genet. 2022, 23, 104–119. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.H.; Jeong, J.C.; Park, S.; Lee, H.S.; Kwak, S.S. Molecular characterization of two ethylene response factor genes in sweetpotato that respond to stress and activate the expression of defense genes in tobacco leaves. J Plant Physiol. 2012, 169, 1112–20. [Google Scholar] [CrossRef] [PubMed]
- Shinozaki, K.; Yamaguchi-Shinozaki, K. Gene networks involved in drought stress response and tolerance. J Exp Bot. 2007, 58, 221–7. [Google Scholar] [CrossRef] [PubMed]
- Xiao, M.; Li, Z.; Zhu, L.; Wang, J.; Zhang, B.; Zheng, F.; Zhao, B.; Zhang, H.; Wang, Y.; Zhang, Z. The Multiple Roles of Ascorbate in the Abiotic Stress Response of Plants: Antioxidant, Cofactor, and Regulator. Front Plant Sci. 2021, 12, 598173. [Google Scholar] [CrossRef] [PubMed]
- Khan, M.I.R.; Kumari, S.; Nazir, F.; Khanna, R.R.; Gupta, R.; Chhillar, H. Defensive Role of Plant Hormones in Advancing Abiotic Stress-Resistant Rice Plants. Rice Sci. 2023, 30, 15–35. [Google Scholar] [CrossRef]
- Saijo, Y.; Loo, E.P. Plant immunity in signal integration between biotic and abiotic stress responses. New Phytol. 2020, 225, 87–104. [Google Scholar] [CrossRef]
- Lim, J.; Lim, C.W.; Lee, S.C. Role of pepper MYB transcription factor CaDIM1 in regulation of the drought response. Front Plant Sci. 2022, 13, 1028392. [Google Scholar] [CrossRef]
- Hussain, S.; Cheng, Y.; Li, Y.; Wang, W.; Tian, H.; Zhang, N.; Wang, Y.; Yuan, Y.; Hussain, H.; Lin, R.; Wang, C.; Wang, T.; Wang, S. AtbZIP62 Acts as a Transcription Repressor to Positively Regulate ABA Responses in Arabidopsis. Plants (Basel). 2022, 11, 3037. [Google Scholar] [CrossRef]
- Siemiatkowska, B.; Chiara, M.; Badiger, B.G.; Riboni, M.; D'Avila, F.; Braga, D.; Salem, M.A.A.; Martignago, D.; Colanero, S.; Galbiati, M.; Giavalisco, P.; Tonelli, C.; Juenger, T.E.; Conti, L. GIGANTEA Is a Negative Regulator of Abscisic Acid Transcriptional Responses and Sensitivity in Arabidopsis. Plant Cell Physiol. 2022, 63, 1285–1297. [Google Scholar] [CrossRef]
- Vanitha, P.A.; Vijayaraghavareddy, P.; Uttarkar, A.; Dawane, A.; Sujitha, D.; Ashwin, V.; Babitha, K.C.; Niranjan, V.; Sheshshayee, M.S.; Anuradha, C.V.; et al. Novel small molecules targeting bZIP23 TF improve stomatal conductance and photosynthesis under mild drought stress by regulating ABA. FEBS J. 2022, 289, 6058–6077. [Google Scholar]
- Michaud, O.; Krahmer, J.; Galbier, F.; Lagier, M.; Galvão, V.C.; Ince, Y.Ç.; Trevisan, M.; Knerova, J.; Dickinson, P.; Hibberd, J.M.; Zeeman, S.C.; Fankhauser, C. Abscisic acid modulates neighbor proximity-induced leaf hyponasty in Arabidopsis. Plant Physiol. 2023, 191, 542–557. [Google Scholar] [CrossRef]
- Finkelstein, R.R.; Lynch, T.J. Overexpression of ABI5 Binding Proteins Suppresses Inhibition of Germination Due to Overaccumulation of DELLA Proteins. Int J Mol Sci. 2022, 23, 5537. [Google Scholar] [CrossRef] [PubMed]
- Gupta, A.; Upadhyay, RK.; Prabhakar, R.; Tiwari, N.; Garg, R.; Sane, V.A.; Sane, A.P. SlDREB3, a negative regulator of ABA responses, controls seed germination, fruit size and the onset of ripening in tomato. Plant Sci. 2022, 19, 111249. [Google Scholar] [CrossRef]
- Ma, Z.; Jin, Y.M.; Wu, T.; Hu, L.; Zhang, Y.; Jiang, W.; Du, X. OsDREB2B, an AP2/ERF transcription factor, negatively regulates plant height by conferring GA metabolism in rice. Front. Plant Sci. 2022, 13, 1007811. [Google Scholar] [CrossRef]
- Ma, Z.; Wu, T.; Huang, K.; Jin, Y.M.; Li, Z.; Chen, M.; Yun, S.; Zhang, H.; Yang, X.; Chen, H.; et al. A Novel AP2/ERF Transcription Factor, OsRPH1, Negatively Regulates Plant Height in Rice. Front. Plant Sci. 2020, 11, 709. [Google Scholar] [CrossRef]
- Qi, W.; Sun, F.; Wang, Q.; Chen, M.; Huang, Y.; Feng, Y.Q.; Luo, X.; Yang, J. Rice ethylene-response AP2/ERF factor OsEATB restricts internode elongation by down-regulating a gibberellin biosynthetic gene. Plant Physiol. 2011, 157, 216–28. [Google Scholar] [CrossRef] [PubMed]
- Yaish, M.W.; El-Kereamy, A.; Zhu, T.; Beatty, P.H.; Good, A.G.; Bi, Y.M.; Rothstein, S.J. The APETALA-2-like transcription factor OsAP2-39 controls key interactions between abscisic acid and gibberellin in rice. PLoS Genet. 2010, 6, e1001098. [Google Scholar] [CrossRef]
- Sun, Z.M.; Zhou, M.L.; Wang, D.; Tang, Y.X.; Lin, M.; Wu, Y.M. Overexpression of the lotus corniculatus soloist gene LcAP2/ERF107 enhances tolerance to salt stress. Protein Peptide Lett. 2016, 23, 442–449. [Google Scholar] [CrossRef] [PubMed]
- Abiri, R.; Shaharuddin, N.A.; Maziah, M.; Yusof, Z.N.B.; Atabaki, N.; Sahebi, M.; Valdiani, A.; Kalhori, N.; Azizi, P.; Hanafi, M. M. Role of ethylene and the APETALA 2/ethylene response factor superfamily in rice under various abiotic and biotic stress conditions. Environmental and Experimental Botany 2017, 134, 33–44. [Google Scholar] [CrossRef]
- Xie, W.; Ding, C.; Hu, H.; Dong, G.; Zhang, G.; Qian, Q.; Ren, D. Molecular Events of Rice AP2/ERF Transcription Factors. Int J Mol Sci. 2022, 23, 12013. [Google Scholar] [CrossRef]
- Lee, S.J.; Park, J.H.; Lee, M.H.; Yu, J.H.; Kim, S.Y. Isolation and functional characterization of CE1 binding proteins. BMC Plant Biol. 2010, 10, 277. [Google Scholar] [CrossRef]
- Lee, S.B.; Lee, S.J.; Kim, S.Y. AtERF15 is a positive regulator of ABA response. Plant Cell Rep. 2015, 34, 71–81. [Google Scholar] [CrossRef]
- Ritonga, F.N.; Ngatia, J.N.; Wang, Y.; Khoso, M.A.; Farooq, U.; Chen, S. AP2/ERF, an important cold stress-related transcription factor family in plants: A review. Physiol Mol Biol Plants. 2021, 27, 1953–1968. [Google Scholar] [CrossRef]
- Reboledo, G.; Agorio, A.; Ponce, D.L.I. Moss transcription factors regulating development and defense responses to stress. J Exp Bot. 2022, 73, 4546–4561. [Google Scholar] [CrossRef]
- Javed, T.; Shabbir, R.; Ali, A.; Afzal, I.; Zaheer, U.; Gao, S.J. Transcription Factors in Plant Stress Responses: Challenges and Potential for Sugarcane Improvement. Plants (Basel). 2020, 9, 491. [Google Scholar] [CrossRef] [PubMed]
- Jan, R.; Asaf, S.; Numan, M.; Lubna; Kim, K.-M. Plant Secondary Metabolite Biosynthesis and Transcriptional Regulation in Response to Biotic and Abiotic Stress Conditions. Agronomy. 2021, 11, 968. [Google Scholar] [CrossRef]
- Hussain, Q.; Asim, M.; Zhang, R.; Khan, R.; Farooq, S.; Wu, J. Transcription Factors Interact with ABA through Gene Expression and Signaling Pathways to Mitigate Drought and Salinity Stress. Biomolecules. 2021, 11, 1159. [Google Scholar] [CrossRef] [PubMed]
- Gasch, P.; Fundinger, M.; Müller, J.T.; Lee, T.; Bailey, S.J.; Mustroph, A. Redundant ERF-VII Transcription Factors Bind to an Evolutionarily Conserved cis-Motif to Regulate Hypoxia-Responsive Gene Expression in Arabidopsis. Plant Cell. 2016, 28, 160–80. [Google Scholar] [CrossRef] [PubMed]
- Park, C.S.; Go, Y.S.; Suh, M.C. Cuticular wax biosynthesis is positively regulated by WRINKLED4, an AP2/ERF-type transcription factor, in Arabidopsis stems. Plant J. 2016, 88, 257–270. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.Y.; Hsieh, E.J.; Cheng, M.C.; Chen, C.Y.; Hwang, S.Y.; Lin, T.P. ORA47 (octadecanoid-responsive AP2/ERF-domain transcription factor 47) regulates jasmonic acid and abscisic acid biosynthesis and signaling through binding to a novel cis-element. New Phytol. 2016, 211, 599–613. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.Y.; Hwang, E.Y.; Seok, H.Y.; Tarte, V.N.; Jeong, M.S.; Jang, S.B.; Moon, Y.H. Arabidopsis AtERF71/HRE2 functions as transcriptional activator via cis-acting GCC box or DRE/CRT element and is involved in root development through regulation of root cell expansion. Plant Cell Rep. 2015, 34, 223–31. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.S.; Mizoi, J.; Yoshida, T.; Fujita, Y.; Nakajima, J.; Ohori, T.; Todaka, D.; Nakashima, K.; Hirayama, T.; Shinozaki, K.; Yamaguchi-Shinozaki, K. An ABRE promoter sequence is involved in osmotic stress-responsive expression of the DREB2A gene, which encodes a transcription factor regulating drought-inducible genes in Arabidopsis. Plant Cell Physiol. 2011, 52, 2136–46. [Google Scholar] [CrossRef]
- Liu, H.C.; Liao, H.T.; Charng, Y.Y. The role of class A1 heat shock factors (HSFA1s) in response to heat and other stresses in Arabidopsis. Plant Cell Environ. 2011, 34, 738–51. [Google Scholar] [CrossRef]
- Owji, H.; Hajiebrahimi, A.; Seradj, H.; Hemmati, S. Identification and functional prediction of stress responsive AP2/ERF transcription factors in Brassica napus by genome-wide analysis. Comput Biol Chem. 2017, 71, 32–56. [Google Scholar] [CrossRef] [PubMed]
- Mizoi, J.; Kanazawa, N.; Kidokoro, S.; Takahashi, F.; Qin, F.; Morimoto, K.; Shinozaki, K.; Yamaguchi-Shinozaki, K. Heat-induced inhibition of phosphorylation of the stress-protective transcription factor DREB2A promotes thermotolerance of Arabidopsis thaliana. J Biol Chem. 2019, 294, 902–917. [Google Scholar] [CrossRef] [PubMed]
- Song, C.P.; Agarwal, M.; Ohta, M.; Guo, Y.; Halfter, U.; Wang, P.; Zhu, J.K. Role of an Arabidopsis AP2/EREBP-type transcriptional repressor in abscisic acid and drought stress responses. Plant Cell. 2005, 17, 2384–96. [Google Scholar] [CrossRef] [PubMed]
- Pfluger, J.; Wagner, D. Histone modifications and dynamic regulation of genome accessibility in plants. Curr Opin Plant Biol. 2007, 10, 645–52. [Google Scholar] [CrossRef] [PubMed]
- Rando, O.J.; Ahmad, K. Rules and regulation in the primary structure of chromatin. Curr Opin Cell Biol. 2007, 19, 250–6. [Google Scholar] [CrossRef]
- Klose, R.J.; Bird, A.P. Genomic DNA methylation: the mark and its mediators. Trends Biochem Sci. 2006, 31, 89–97. [Google Scholar] [CrossRef]
- Zemach, A.; Grafi, G. Methyl-CpG-binding domain proteins in plants: interpreters of DNA methylation. Trends Plant Sci. 2007, 12, 80–5. [Google Scholar] [CrossRef]
- Kavas, M.; Kizildogan, A.; Gökdemir, G.; Baloglu, M.C. Genome-wide investigation and expression analysis of AP2-ERF gene family in salt tolerant common bean. EXCLI J. 2015, 14, 1187–206. [Google Scholar]
- Jisha, V.; Dampanaboina, L.; Vadassery, J.; Mithöfer, A.; Kappara, S.; Ramanan, R. Overexpression of an AP2/ERF Type Transcription Factor OsEREBP1 Confers Biotic and Abiotic Stress Tolerance in Rice. PLoS One. 2015, 10, e0127831. [Google Scholar] [CrossRef] [PubMed]
- Nakano, T.; Suzuki, K.; Fujimura, T.; Shinshi, H. Genome-wide analysis of the ERF gene family in Arabidopsis and rice. Plant Physiol. 2006, 140, 411–32. [Google Scholar] [CrossRef]
- Tiwari, S.B.; Belachew, A.; Ma, S.F.; Young, M.; Ade, J.; Shen, Y.; Marion, C.M.; Holtan, H.E.; Bailey, A.; Stone, J.K.; Edwards, L.; Wallace, A.D.; Canales, R.D.; Adam, L.; Ratcliffe, O.J.; Repetti, P.P. The EDLL motif: a potent plant transcriptional activation domain from AP2/ERF transcription factors. Plant J. 2012, 70, 855–65. [Google Scholar] [CrossRef] [PubMed]
- Song, C.P.; Galbraith, D.W. AtSAP18, an orthologue of human SAP18, is involved in the regulation of salt stress and mediates transcriptional repression in Arabidopsis. Plant Mol Biol. 2006, 60, 241–57. [Google Scholar] [CrossRef] [PubMed]
- Franco-Zorrilla, J.M.; López-Vidriero, I.; Carrasco, J.L.; Godoy, M.; Vera, P.; Solano, R. DNA-binding specificities of plant transcription factors and their potential to define target genes. Proc Natl Acad Sci U S A. 2014, 111, 2367–72. [Google Scholar] [CrossRef]
- Waadt, R.; Seller, C.A.; Hsu, P.K.; Takahashi, Y.; Munemasa, S.; Schroeder, J.I. Plant hormone regulation of abiotic stress responses. Nat Rev Mol Cell Biol. 2022, 23, 680–694. [Google Scholar] [CrossRef]
- Xie, Z.; Nolan, T.M.; Jiang, H.; Yin, Y. AP2/ERF Transcription Factor Regulatory Networks in Hormone and Abiotic Stress Responses in Arabidopsis. Front Plant Sci 2019, 28, 228. [Google Scholar] [CrossRef]
- Yoon, Y.; Seo, D.H.; Shin, H.; Kim, H.J.; Kim, C.M.; Jang, G. The Role of Stress-Responsive Transcription Factors in Modulating Abiotic Stress Tolerance in Plants. Agronomy. 2020, 10, 788. [Google Scholar] [CrossRef]
- Mohanty, B. Genomic architecture of promoters and transcriptional regulation of candidate genes in rice involved in tolerance to anaerobic germination. Curr. Plant Biol. 2022, 29, 100236. [Google Scholar] [CrossRef]
- Zwack, P.J.; Rashotte, A.M. Interactions between cytokinin signalling and abiotic stress responses. J Exp Bot. 2015, 66, 4863–71. [Google Scholar] [CrossRef]
- Chen, M.; Yan, T.; Shen, Q.; Lu, X.; Pan, Q.; Huang, Y.; Tang, Y.; Fu, X.; Liu, M.; Jiang, W.; Lv, Z.; Shi, P.; Ma, Y.N.; Hao, X.; Zhang, L.; Li, L.; Tang, K. GLANDULAR TRICHOME-SPECIFIC WRKY 1 promotes artemisinin biosynthesis in Artemisia annua. New Phytol. 2017, 214, 304–316. [Google Scholar] [CrossRef]
- Li, Z.; Sheerin, D.J.; von Roepenack-Lahaye., E.; Stahl, M.; Hiltbrunner, A. The phytochrome interacting proteins ERF55 and ERF58 repress light-induced seed germination in Arabidopsis thaliana. Nat Commun 2022, 13, 1656. [Google Scholar] [CrossRef]
- Li, J.; Guo, X.; Zhang, M.; Wang, X.; Zhao, Y.; Yin, Z.; Zhang, Z.; Wang, Y.; Xiong, H.; Zhang, H.; Todorovska, E.; Li, Z. OsERF71 confers drought tolerance via modulating ABA signaling and proline biosynthesis. Plant Sci. 2018, 270, 131–139. [Google Scholar] [CrossRef] [PubMed]
- Cheng, C.; An, L.; Li, F.; Ahmad, W.; Aslam, M.; Ul Haq, M.Z.; Yan, Y.; Ahmad, R.M. Wide-Range Portrayal of AP2/ERF Transcription Factor Family in Maize (Zea mays L.) Development and Stress Responses. Genes (Basel) 2023, 14, 194. [Google Scholar] [CrossRef] [PubMed]
- Xiong, R.; Chu, Z.; Peng, X.; Cui, G.; Li, W.; Dong, L. Transcript-wide identification and expression pattern analysis to comprehend the roles of AP2/ERF genes under development and abiotic stress in Trichosanthes kirilowii. BMC Plant Biol. 2023, 23, 354. [Google Scholar] [CrossRef]
- Hu, Z.; Wang, X.; Wei, L.; Wansee, S.; Rabbani Nasab, H.; Chen, L.; Kang, Z.; Wang, J. TaAP2-10, an AP2/ERF transcription factor, contributes to wheat resistance against stripe rust. J Plant Physiol. 2023, 288, 154078. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Yu, J.; Wang, R.; Liu, W.; Chen, S.; Wang, Y.; Yu, Y.; Qu, G.; Chen, S. Genome-Wide Identification and Expression Profiles of C-Repeat Binding Factor Transcription Factors in Betula platyphylla under Abiotic Stress. Int J Mol Sci. 2023, 24, 10573. [Google Scholar] [CrossRef] [PubMed]
- Li, P.; Chai, Z.; Lin, P.; Huang, C.; Huang, G.; Xu, L.; Deng, Z.; Zhang, M.; Zhang, Y.; Zhao, X. Genome-wide identification and expression analysis of AP2/ERF transcription factors in sugarcane (Saccharum spontaneum L.). BMC Genomics 2020, 21, 685. [Google Scholar] [CrossRef]
- Chen, J.Q.; Meng, X.P.; Zhang, Y.; Xia, M.; Wang, X.P. Over-expression of OsDREB genes lead to enhanced drought tolerance in rice. Biotechnol Lett. 2008, 30, 2191–8. [Google Scholar] [CrossRef]
- Fu, J.; Zhu, C.; Wang, C.; Liu, L.; Shen, Q.; Xu, D.; Wang, Q. Maize transcription factor ZmEREB20 enhanced salt tolerance in transgenic Arabidopsis. Plant Physiol Biochem. 2021, 159, 257–267. [Google Scholar] [CrossRef]
- Zhang, K.; Jiang, L.; Wang, X.; Han, H.; Chen, D.; Qiu, D.; Yang, Y. Transcriptome-wide analysis of AP2/ERF transcription factors involved in regulating taxol biosynthesis in Taxus× media. Ind. Crops Prod. 2021, 171, 113972. [Google Scholar] [CrossRef]
- Cai, X.; Chen, Y.; Wang, Y.; Shen, Y.; Yang, J.; Jia, B.; Sun, X.; Sun, M. A comprehensive investigation of the regulatory roles of OsERF096, an AP2/ERF transcription factor, in rice cold stress response. Plant Cell Rep. 2023, 42, 2011–2022. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.; Liu, L.; Hu, H.; Tang, N.; Shi, L.; Xu, F.; Wang, S. Arabidopsis ERF012 Is a Versatile Regulator of Plant Growth, Development and Abiotic Stress Responses. Int J Mol Sci. 2022, 23, 6841. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.Q.; Xia, D.N.; Li, W.Q.; Cao, X.Y.; Ma, F.; Wang, Q.Q.; Zhan, X.Q.; Hu, T.X. Overexpression of a tomato AP2/ERF transcription factor SlERF.B1 increases sensitivity to salt and drought stresses. Sci. Hortic. 2022, 304, 111332. [Google Scholar] [CrossRef]
- Kazan, K. Diverse roles of jasmonates and ethylene in abiotic stress tolerance. Trends Plant Sci. 2015, 20, 219–29. [Google Scholar] [CrossRef] [PubMed]
- Jung, K.H.; Seo, Y.S.; Walia, H.; Cao, P.; Fukao, T.; Canlas, P.E.; Amonpant, F.; Bailey-Serres, J.; Ronald, P.C. The submergence tolerance regulator Sub1A mediates stress-responsive expression of AP2/ERF transcription factors. Plant Physiol. 2010, 152, 1674–92. [Google Scholar] [CrossRef] [PubMed]
- Peña-Castro, J.M.; van, Zanten. M.; Lee, S.C.; Patel, M.R.; Voesenek, L.A.; Fukao, T.; Bailey-Serres, J. Expression of rice SUB1A and SUB1C transcription factors in Arabidopsis uncovers flowering inhibition as a submergence tolerance mechanism. Plant J. 2011, 67, 434–46. [Google Scholar] [CrossRef] [PubMed]
- Hinz, M.; Wilson, I.W.; Yang, J.; Buerstenbinder, K.; Llewellyn, D.; Dennis, E.S.; Sauter, M.; Dolferus, R. Arabidopsis RAP2.2: an ethylene response transcription factor that is important for hypoxia survival. Plant Physiol 2010, 153, 757–72. [Google Scholar] [CrossRef]
- Altamura, M.M.; Piacentini, D.; Della Rovere., F.; Fattorini, L.; Falasca, G.; Betti, C. New Paradigms in Brassinosteroids, Strigolactones, Sphingolipids, and Nitric Oxide Interaction in the Control of Lateral and Adventitious Root Formation. Plants (Basel) 2023, 12, 413. [Google Scholar] [CrossRef]
- Liu, K.; Li, Y.; Chen, X.; Li, L.; Liu, K.; Zhao, H.; Wang, Y.; Han, S. ERF72 interacts with ARF6 and BZR1 to regulate hypocotyl elongation in Arabidopsis. J Exp Bot. 2018, 69, 3933–3947. [Google Scholar] [CrossRef]
- Schmitz, A.J.; Folsom, J.J.; Jikamaru, Y.; Ronald, P.; Walia, H. SUB1A-mediated submergence tolerance response in rice involves differential regulation of the brassinosteroid pathway. New Phytol. 2013, 198, 1060–1070. [Google Scholar] [CrossRef] [PubMed]
- Kurepa, J.; Shull, T.E.; Smalle, J.A. Friends in Arms: Flavonoids and the Auxin/Cytokinin Balance in Terrestrialization. Plants (Basel). 2023, 12, 517. [Google Scholar] [CrossRef]
- Gu, C.; Guo, Z.H.; Hao, P.P.; Wang, G.M.; Jin, Z.M.; Zhang, S.L. Multiple regulatory roles of AP2/ERF transcription factor in angiosperm. Bot Stud. 2017, 58, 6. [Google Scholar] [CrossRef] [PubMed]
- Mawlong, I.; Ali, K.; Tyagi, A. Cloning and characterization of a water deficit stress responsive transcription factor gene from Oryza sativa L. Indian J Exp Biol. 2016, 54, 26–36. [Google Scholar]
- Rashotte, A.M.; Mason, M.G.; Hutchison, C.E.; Ferreira, F.J.; Schaller, G.E.; Kieber, J.J. A subset of Arabidopsis AP2 transcription factors mediates cytokinin responses in concert with a two-component pathway. Proc Natl Acad Sci U S A. 2006, 103, 11081–5. [Google Scholar] [CrossRef] [PubMed]
- Striberny, B.; Melton, A.E.; Schwacke, R.; Krause, K.; Fischer, K.; Goertzen, L.R.; Rashotte, A.M. Cytokinin Response Factor 5 has transcriptional activity governed by its C-terminal domain. Plant Signal Behav. 2017, 12, e1276684. [Google Scholar] [CrossRef]
- Zwack, P.J.; Robinson, B.R.; Risley, M.G.; Rashotte, A.M. Cytokinin response factor 6 negatively regulates leaf senescence and is induced in response to cytokinin and numerous abiotic stresses. Plant Cell Physiol. 2013, 54, 971–81. [Google Scholar] [CrossRef]
- Zwack, P.J.; De Clercq, I.; Howton, T.C.; Hallmark, H.T.; Hurny, A.; Keshishian, E.A.; Parish, A.M.; Benkova, E.; Mukhtar, M.S.; Van, Breusegem. F.; Rashotte, A.M. Cytokinin Response Factor 6 Represses Cytokinin-Associated Genes during Oxidative Stress. Plant Physiol 2016, 172, 1249–1258. [Google Scholar] [CrossRef]
- Zwack, P.J.; Compton, M.A.; Adams, C.I.; Rashotte, A.M. Cytokinin response factor 4 (CRF4) is induced by cold and involved in freezing tolerance. Plant Cell Rep. 2016, 35, 573–84. [Google Scholar] [CrossRef]
- Golldack, D.; Li, C.; Mohan, H.; Probst, N. Tolerance to drought and salt stress in plants: Unraveling the signaling networks. Front Plant Sci. 2014, 5, 151. [Google Scholar] [CrossRef]
- Kim, Y.; Chung, Y.S.; Lee, E.; Tripathi, P.; Heo, S.; Kim, K.H. Root Response to Drought Stress in Rice (Oryza sativa L.). Int J Mol Sci 2020, 21, 1513. [Google Scholar] [CrossRef]
- Yu, Y.; Yu, M.; Zhang, S.; Song, T.; Zhang, M.; Zhou, H.; Wang, Y.; Xiang, J.; Zhang, X. Transcriptomic Identification of Wheat AP2/ERF Transcription Factors and Functional Characterization of TaERF-6-3A in Response to Drought and Salinity Stresses. Int J Mol Sci. 2022, 23, 3272. [Google Scholar] [CrossRef]
- Jarambasa, T.; Regon, P.; Jyoti, S.Y.; Gupta, D.; Panda, S.K.; Tanti, B. Genome-wide identification and expression analysis of the Pisum sativum (L.) APETALA2/ethylene-responsive factor (AP2/ERF) gene family reveals functions in drought and cold stresses. Genetica 2023, 151, 225–239. [Google Scholar] [CrossRef]
- Kumar, V.; Kumar, A.; Tewari, K.; Garg, N.K.; Changan, S.S.; Tyagi, A. Isolation and characterization of drought and ABA responsive promoter of a transcription factor encoding gene from rice. Physiol Mol Biol Plants. 2022, 28, 1813–1831. [Google Scholar] [CrossRef]
- Kabir, S.M.T.; Hossain, M.S.; Bashar, K.K.; Honi, U.; Ahmed, B.; Emdad, E.M.; Alam, M.M.; Haque, M.S.; Islam, M.S. Genome-wide identification and expression profiling of AP2/ERF superfamily genes under stress conditions in dark jute (Corchorus olitorius L.). Ind. Crops Prod 2021, 166, 113469. [Google Scholar] [CrossRef]
- Kong, L.; Song, Q.; Wei, H.; Wang, Y.; Lin, M.; Sun, K.; Zhang, Y.; Yang, J.; Li, C.; Luo, K. The AP2/ERF transcription factor PtoERF15 confers drought tolerance via JA-mediated signaling in Populus. New Phytol. 2023, 240, 1848–1867. [Google Scholar] [CrossRef] [PubMed]
- Litalien, A.; Zeeb, B. Curing the earth: A review of anthropogenic soil salinization and plant-based strategies for sustainable mitigation. Sci Total Environ. 2020, 698, 134235. [Google Scholar] [CrossRef]
- Huang, S.; Ma, Z.; Hu, L.; Huang, K.; Zhang, M.; Zhang, S.; Jiang, W.; Wu, T.; Du, X. Involvement of rice transcription factor OsERF19 in response to ABA and salt stress responses. Plant Physiol Biochem. 2021, 167, 22–30. [Google Scholar] [CrossRef] [PubMed]
- Licausi, F.; van Dongen, J.T.; Giuntoli, B.; Novi, G.; Santaniello, A.; Geigenberger, P.; Perata, P. HRE1 and HRE2, two hypoxia-inducible ethylene response factors, affect anaerobic responses in Arabidopsis thaliana. Plant J. 2010, 62, 302–15. [Google Scholar] [CrossRef]
- Park, H.Y.; Seok, H.Y.; Woo, D.H.; Lee, S.Y.; Tarte, V.N.; Lee, E.H.; Lee, C.H.; Moon, Y.H. AtERF71/HRE2 transcription factor mediates osmotic stress response as well as hypoxia response in Arabidopsis. Biochem Biophys Res Commun. 2011, 414, 135–41. [Google Scholar] [CrossRef]
- Seok, H.Y.; Tran, H.T.; Lee, S.Y.; Moon, Y.H. AtERF71/HRE2, an Arabidopsis AP2/ERF Transcription Factor Gene, Contains Both Positive and Negative Cis-Regulatory Elements in Its Promoter Region Involved in Hypoxia and Salt Stress Responses. Int J Mol Sci. 2022, 23, 5310. [Google Scholar] [CrossRef]
- Posch, B.C.; Kariyawasam, B.C.; Bramley, H.; Coast, O.; Richards, R.A.; Reynolds, M.P.; Trethowan, R.; Atkin, O.K. Exploring high temperature responses of photosynthesis and respiration to improve heat tolerance in wheat. J Exp Bot. 2019, 70, 5051–5069. [Google Scholar] [CrossRef]
- He, Y.; Guan, H.; Li, B.; Zhang, S.; Xu, Y.; Yao, Y.; Yang, X.; Zha, Z.; Guo, Y.; Jiao, C.; Cai, H. Transcriptome Analysis Reveals the Dynamic and Rapid Transcriptional Reprogramming Involved in Heat Stress and Identification of Heat Response Genes in Rice. Int J Mol Sci. 2023, 24, 14802. [Google Scholar] [CrossRef]
- Zhang, T.; Tang, Y.; Luan, Y.; Cheng, Z.; Wang, X.; Tao, J.; Zhao, D. Herbaceous peony AP2/ERF transcription factor binds the promoter of the tryptophan decarboxylase gene to enhance high-temperature stress tolerance. Plant Cell Environ. 2022, 45, 2729–2743. [Google Scholar] [CrossRef]
- Djemal, R.; Khoudi, H. The barley SHN1-type transcription factor HvSHN1 imparts heat, drought and salt tolerances in transgenic tobacco. Plant Physiol Biochem. 2021, 164, 44–53. [Google Scholar] [CrossRef]
- Kidokoro, S.; Shinozaki, K.; Yamaguchi-Shinozaki, K. Transcriptional regulatory network of plant cold-stress responses. Trends Plant Sci. 2022, 27, 922–935. [Google Scholar] [CrossRef]
- Xie, Z.; Yang, C.; Liu, S.; Li, M.; Gu, L.; Peng, X.; Zhang, Z. Identification of AP2/ERF transcription factors in Tetrastigma hemsleyanum revealed the specific roles of ERF46 under cold stress. Front Plant Sci. 2022, 13, 936602. [Google Scholar] [CrossRef] [PubMed]
- Zhao, M.; Li, Y.; Zhang, X.; You, X.; Yu, H.; Guo, R.; Zhao, X. Genome-Wide Identification of AP2/ERF Superfamily Genes in Juglans mandshurica and Expression Analysis under Cold Stress. Int J Mol Sci. 2022, 23, 15225. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Yin, X.R.; Li, H.; Xu, M.; Zhang, M.X.; Li, S.J.; Liu, X.F.; Shi, Y.N.; Grierson, D.; Chen, K.S. ETHYLENE RESPONSE FACTOR39-MYB8 complex regulates low-temperature-induced lignification of loquat fruit. J Exp Bot. 2020, 71, 3172–3184. [Google Scholar] [CrossRef] [PubMed]
- Ren, M.; Yang, W.; Zhang, J.; Zhao, L.; Quan, Y.; He, Z.; Xu, Y.; Zhang, F.; Yin, M.; Wang, Y.; Gao, R. Overexpression of ClRAP2.4 in Chrysanthemum enhances tolerance to cold stress. Funct Plant Biol. 2023, 50, 470–481. [Google Scholar] [CrossRef] [PubMed]
- Yahaya, S.M.; Mahmud, A.A.; Abdullahi, M.; Haruna, A. Recent advances in the chemistry of N, P, K as fertilizer in soil: A review. Pedosphere. 2022, 35, 385–406. [Google Scholar]
- Waqas, M.; Hawkesford, M.J.; Geilfus, C.M. Feeding the world sustainably: Efficient nitrogen use. Trends Plant Sci. 2023, 28, 505–508. [Google Scholar] [CrossRef]
- Joshi, V.; Joshi, M.; Penalosa, A. Comparative analysis of tissue-specific transcriptomic responses to nitrogen stress in spinach (Spinacia oleracea). PLoS One. 2020, 15, e0232011. [Google Scholar] [CrossRef]
- Cerri, M.R.; Wang, Q.; Stolz, P.; Folgmann, J.; Frances, L.; Katzer, K.; Li, X.; Heckmann, A.B.; Wang, T.L.; Downie, J.A.; Klingl, A.; de Carvalho-Niebel, F.; Xie, F.; Parniske, M. The ERN1 transcription factor gene is a target of the CCaMK/CYCLOPS complex and controls rhizobial infection in Lotus japonicus. New Phytol. 2017, 215, 323–337. [Google Scholar] [CrossRef]
- Jezek, M.; Allan, A.C.; Jones, J.J.; Geilfus, C.M. Why do plants blush when they are hungry? New Phytol. 2023, 239, 494–505. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Wang, S.; Xie, Q.; Xia, Y.; Lu, L.; Wang, M.; Wang, G.; Long, S.; Cai, Y.; Xu, L.; Wang, E.; Jiang, Y. Control of arbuscule development by a transcriptional negative feedback loop in Medicago. Nat Commun. 2023, 14, 5743. [Google Scholar] [CrossRef] [PubMed]
- Chen, N.; Qin, J.; Tong, S.; Wang, W.; Jiang, Y. One AP2/ERF Transcription Factor Positively Regulates Pi Uptake and Drought Tolerance in Poplar. Int J Mol Sci. 2022, 23, 5241. [Google Scholar] [CrossRef]
- Parakkunnel, R.; Naik, K.B.; Vanishree G, C.S.; Purru, S.; Bhaskar, K.U.; Bhat, K.V.; Kumar, S. Gene fusions, micro-exons and splice variants define stress signaling by AP2/ERF and WRKY transcription factors in the sesame pan-genome. Front Plant Sci. 2022, 13, 1076229. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.J.; Ruzicka, D.; Shin, R.; Schachtman, D.P. The Arabidopsis AP2/ERF transcription factor RAP2.11 modulates plant response to low-potassium conditions. Mol Plant 2012, 5, 1042–57. [Google Scholar] [CrossRef]
- Chen, H.C.; Chien, T.C.; Chen, T.Y.; Chiang, M.H.; Lai, M.H.; Chang, M.C. Overexpression of a Novel ERF-X-Type Transcription Factor, OsERF106MZ, Reduces Shoot Growth and Tolerance to Salinity Stress in Rice. Rice (N Y) 2021, 14, 82. [Google Scholar] [CrossRef]
- Gavrilescu, M. Enhancing phytoremediation of soils polluted with heavy metals. Curr. Opin. Biotechnol. 2022, 74, 21–31. [Google Scholar] [CrossRef] [PubMed]
- Chot, E.; Reddy, M.S. Role of Ectomycorrhizal Symbiosis Behind the Host Plants Ameliorated Tolerance Against Heavy Metal Stress. Front. Microbiol. 2022, 13, 855473. [Google Scholar] [CrossRef] [PubMed]
- Asgher, M.; Rehaman, A.; Islam, S.N.u.; Arshad, M.; Khan, N.A. Appraisal of Functions and Role of Selenium in Heavy Metal Stress Adaptation in Plants. Agriculture. 2023, 13, 1083. [Google Scholar] [CrossRef]
- Pandey, A.K.; Zori´c, L.; Sun, T.; Karanovi´c, D.; Fang, P.; Borišev, M.; Wu, X.; Lukovi´c, J.; Xu, P. The Anatomical Basis of Heavy Metal Responses in Legumes and Their Impact on Plant-Rhizosphere Interactions. Plants. 2022, 11, 2554. [Google Scholar] [CrossRef]
- Karanja, B.K.; Xu, L.; Wang, Y.; Tang, M.; M'mbone Muleke, E.; Dong, J.; Liu, L. Genome-wide characterization of the AP2/ERF gene family in radish (Raphanus sativus L.): Unveiling evolution and patterns in response to abiotic stresses. Gene 2019, 718, 144048. [Google Scholar] [CrossRef]
- Chen, P.; Li, Z.; Luo, D.; Jia, R.; Lu, H.; Tang, M.; Hu, Y.; Yue, J.; Huang, Z. Comparative transcriptomic analysis reveals key genes and pathways in two different cadmium tolerance kenaf (Hibiscus cannabinus L.) cultivars. Chemosphere 2021, 263, 128211. [Google Scholar] [CrossRef]
- Tian, W.; Huang, Y.; Li, D.; Meng, L.; He, T.; He, G. Identification of StAP2/ERF genes of potato (Solanum tuberosum) and their multiple functions in detoxification and accumulation of cadmium in yest: Implication for Genetic-based phytoremediation. Sci Total Environ. 2022, 810, 152322. [Google Scholar] [CrossRef]
- Verma, V.; Ravindran, P.; Kumar, P.P. Plant hormone-mediated regulation of stress responses. BMC Plant Biol. 2016, 16, 86. [Google Scholar] [CrossRef]
- Takino, J.; Kozaki, T.; Ozaki, T.; Liu, C.; Minami, A.; Oikawa, H. Elucidation of biosynthetic pathway of a plant hormone abscisic acid in phytopathogenic fungi. Biosci Biotechnol Biochem. 2019, 83, 1642–1649. [Google Scholar] [CrossRef]
- Finkelstein, R. Abscisic Acid synthesis and response. Arabidopsis Book. 2013, 11, e0166. [Google Scholar] [CrossRef]
- Sah, S.K.; Reddy, K.R.; Li, J. Abscisic Acid and Abiotic Stress Tolerance in Crop Plants. Front Plant Sci. 2016, 7, 571. [Google Scholar] [CrossRef] [PubMed]
- Feng, C.Z.; Chen, Y.; Wang, C.; Kong, Y.H.; Wu, W.H.; Chen, Y.F. Arabidopsis RAV1 transcription factor, phosphorylated by SnRK2 kinases, regulates the expression of ABI3, ABI4, and ABI5 during seed germination and early seedling development. Plant J. 2014, 80, 654–68. [Google Scholar] [CrossRef] [PubMed]
- Magome, H.; Yamaguchi, S.; Hanada, A.; Kamiya, Y.; Oda, K. dwarf and delayed-flowering 1, a novel Arabidopsis mutant deficient in gibberellin biosynthesis because of overexpression of a putative AP2 transcription factor. Plant J. 2004, 37, 720–9. [Google Scholar] [CrossRef] [PubMed]
- Zhou, X.; Zhang, Z.L.; Park, J.; Tyler, L.; Yusuke, J.; Qiu, K.; Nam, E.A.; Lumba, S.; Desveaux, D.; McCourt, P.; Kamiya, Y.; Sun, T.P. The ERF11 Transcription Factor Promotes Internode Elongation by Activating Gibberellin Biosynthesis and Signaling. Plant Physiol. 2016, 171, 2760–70. [Google Scholar] [CrossRef] [PubMed]
- Guo, H.; Li, L.; Aluru, M.; Aluru, S.; Yin, Y. Mechanisms and networks for brassinosteroid regulated gene expression. Curr Opin Plant Biol. 2013, 16, 545–53. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Ye, K.; Shi, Y.; Cheng, J.; Zhang, X.; Yang, S. BZR1 Positively Regulates Freezing Tolerance via CBF-Dependent and CBF-Independent Pathways in Arabidopsis. Mol Plant. 2017, 10, 545–559. [Google Scholar] [CrossRef] [PubMed]
- Tao, J.J.; Chen, H.W.; Ma, B.; Zhang, W.K.; Chen, S.Y.; Zhang, J.S. The Role of Ethylene in Plants Under Salinity Stress. Front Plant Sci. 2015, 6, 1059. [Google Scholar] [CrossRef]
- Qiao, H.; Shen, Z.; Huang, S.S.; Schmitz, R.J.; Urich, M.A.; Briggs, S.P.; Ecker, J.R. Processing and subcellular trafficking of ER-tethered EIN2 control response to ethylene gas. Science. 2012, 338, 390–3. [Google Scholar] [CrossRef]
- Müller, M.; Munné-Bosch, S. Ethylene Response Factors: A Key Regulatory Hub in Hormone and Stress Signaling. Plant Physiol. 2015, 169, 32–41. [Google Scholar] [CrossRef]
- Yang, C.Y.; Hsu, F.C.; Li, J.P.; Wang, N.N.; Shih, M.C. The AP2/ERF transcription factor AtERF73/HRE1 modulates ethylene responses during hypoxia in Arabidopsis. Plant Physiol. 2011, 156, 202–12. [Google Scholar] [CrossRef]
- Wei, T.; Deng, K.; Liu, D.; Gao, Y.; Liu, Y.; Yang, M.; Zhang, L.; Zheng, X.; Wang, C.; Song, W.; Chen, C.; Zhang, Y. Ectopic Expression of DREB Transcription Factor, AtDREB1A, Confers Tolerance to Drought in Transgenic Salvia miltiorrhiza. Plant Cell Physiol. 2016, 57, 1593–609. [Google Scholar] [CrossRef]
- Mizoi, J.; Shinozaki, K.; Yamaguchi-Shinozaki, K. AP2/ERF family transcription factors in plant abiotic stress responses. Biochim Biophys Acta. 2012, 1819, 86–96. [Google Scholar] [CrossRef]
- Li, X.P.; Tian, A.G.; Luo, G.Z.; Gong, Z.Z.; Zhang, J.S.; Chen, S.Y. Soybean DRE-binding transcription factors that are responsive to abiotic stresses. Theor Appl Genet. 2005, 110, 1355–62. [Google Scholar] [CrossRef]
- Xing, L.; Di, Z.; Yang, W.; Liu, J.; Li, M.; Wang, X.; Cui, C.; Wang, X.; Wang, X.; Zhang, R.; Xiao, J.; Cao, A. Overexpression of ERF1-V from Haynaldia villosa Can Enhance the Resistance of Wheat to Powdery Mildew and Increase the Tolerance to Salt and Drought Stresses. Front Plant Sci. 2017, 8, 1948. [Google Scholar] [CrossRef]
- Feng, W.; Li, J.; Long, S.; Wei, S. A DREB1 gene from zoysiagrass enhances Arabidopsis tolerance to temperature stresses without growth inhibition. Plant Sci. 2019, 278, 20–31. [Google Scholar] [CrossRef]
- Liu, J.; Wang, F.; Yu, G.; Zhang, X.; Jia, C.; Qin, J.; Pan, H. Functional Analysis of the Maize C-Repeat/DRE Motif-Binding Transcription Factor CBF3 Promoter in Response to Abiotic Stress. Int J Mol Sci. 2015, 16, 12131–46. [Google Scholar] [CrossRef]
- Yang, Y.; Shi, J.; Chen, L.; Xiao, W.; Yu, J. ZmEREB46, a maize ortholog of Arabidopsis WAX INDUCER1/SHINE1, is involved in the biosynthesis of leaf epicuticular very-long-chain waxes and drought tolerance. Plant Sci. 2022, 321, 111256. [Google Scholar] [CrossRef]
- Zhu, Y.; Liu, Y.; Zhou, K.; Tian, C.; Aslam, M.; Zhang, B.; Liu, W.; Zou, H. Overexpression of ZmEREBP60 enhances drought tolerance in maize. J Plant Physiol. 2022, 275, 153763. [Google Scholar] [CrossRef]
- Liu, S.; Wang, X.; Wang, H.; Xin, H.; Yang, X.; Yan, J.; Li, J.; Tran, L.S.; Shinozaki, K.; Yamaguchi-Shinozaki, K.; Qin, F. Genome-wide analysis of ZmDREB genes and their association with natural variation in drought tolerance at seedling stage of Zea mays L. PLoS Genet. 2013, 9, e1003790. [Google Scholar] [CrossRef]
- Yu, F.; Liang, K.; Fang, T.; Zhao, H.; Han, X.; Cai, M.; Qiu, F. A group VII ethylene response factor gene, ZmEREB180, coordinates waterlogging tolerance in maize seedlings. Plant Biotechnol J. 2019, 17, 2286–2298. [Google Scholar] [CrossRef]
- Saleh, A.; Lumbreras, V.; Lopez, C.; Dominguez-Puigjaner, E.; Kizis, D.; Pagès, M. Maize DBF1-interactor protein 1 containing an R3H domain is a potential regulator of DBF1 activity in stress responses. Plant J. 2006, 46, 747–57. [Google Scholar] [CrossRef]
- Wang, Z.; Zhao, X.; Ren, Z.; Abou-Elwafa, S.F.; Pu, X.; Zhu, Y.; Dou, D.; Su, H.; Cheng, H.; Liu, Z.; Chen, Y.; Wang, E.; Shao, R.; Ku, L. ZmERF21 directly regulates hormone signaling and stress-responsive gene expression to influence drought tolerance in maize seedlings. Plant Cell Environ. 2022, 45, 312–328. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Ren, Y.; Liu, X.; Luo, S.; Zhang, X.; Liu, X.; Lin, Q.; Zhu, S.; Wan, H.; Yang, Y.; Zhang, Y.; Lei, B.; Zhou, C.; Pan, T.; Wang, Y.; Wu, M.; Jing, R.; Xu, Y.; Han, M.; Wu, F.; Lei, C.; Guo, X.; Cheng, Z.; Zheng, X.; Wang, Y.; Zhao, Z.; Jiang, L.; Zhang, X.; Wang, Y.F.; Wang, H.; Wan, J. Transcriptional activation and phosphorylation of OsCNGC9 confer enhanced chilling tolerance in rice. Mol Plant. 2021, 14, 315–329. [Google Scholar] [CrossRef]
- Figueiredo, D.D.; Barros, P.M.; Cordeiro, A.M.; Serra, T.S.; Lourenço, T.; Chander, S.; Oliveira, M.M.; Saibo, N.J. Seven zinc-finger transcription factors are novel regulators of the stress responsive gene OsDREB1B. J Exp Bot. 2012, 63, 3643–56. [Google Scholar] [CrossRef]
- Mao, D.; Chen, C. Colinearity and similar expression pattern of rice DREB1s reveal their functional conservation in the cold-responsive pathway. PLoS One. 2012, 7, e47275. [Google Scholar] [CrossRef]
- Tian, X.H.; Li, X.P.; Zhou, H.L.; Zhang, J.S.; Gong, Z.Z.; Chen, S.Y. OsDREB4 Genes in Rice Encode AP2-Containing Proteins that Bind Specifically to the Dehydration-Responsive Element. J. Integr. Plant Biol. 2005, 47, 467–476. [Google Scholar] [CrossRef]
- Wang, Q.; Guan, Y.; Wu, Y.; Chen, H.; Chen, F.; Chu, C. Overexpression of a rice OsDREB1F gene increases salt, drought, and low temperature tolerance in both Arabidopsis and rice. Plant Mol Biol. 2008, 67, 589–602. [Google Scholar] [CrossRef]
- "Zhou, X.Y.; Jenks, M.A.; Liu, J.; Liu, A.L.; Zhang, X.W.; Xiang, J.H.; Zou, J.; Peng, Y.; Chen, X.B. Overexpression of Transcription Factor OsWR2 Regulates Wax and Cutin Biosynthesis in Rice and Enhances its Tolerance to Water Deficit. Plant Mol. Biol. Rep. 2014, 32, 719–731. [Google Scholar]
- Lee, D.K.; Jung, H.; Jang, G.; Jeong, J.S.; Kim, Y.S.; Ha, S.H.; Do, C.Y.; Kim, J.K. Overexpression of the OsERF71 Transcription Factor Alters Rice Root Structure and Drought Resistance. Plant Physiol. 2016, 172, 575–88. [Google Scholar] [CrossRef]
- Xiong, H.; Yu, J.; Miao, J.; Li, J.; Zhang, H.; Wang, X.; Liu, P.; Zhao, Y.; Jiang, C.; Yin, Z.; Li, Y.; Guo, Y.; Fu, B.; Wang, W.; Li, Z.; Ali, J.; Li, Z. Natural Variation in OsLG3 Increases Drought Tolerance in Rice by Inducing ROS Scavenging. Plant Physiol. 2018, 178, 451–467. [Google Scholar] [CrossRef]
- Ramegowda, V.; Basu, S.; Krishnan, A.; Pereira, A. Rice GROWTH UNDER DROUGHT KINASE is required for drought tolerance and grain yield under normal and drought stress conditions. Plant Physiol. 2014, 166, 1634–45. [Google Scholar] [CrossRef]
- Tang, Y.; Liu, K.; Zhang, J.; Li, X.; Xu, K.; Zhang, Y.; Qi, J.; Yu, D.; Wang, J.; Li, C. JcDREB2, a Physic Nut AP2/ERF Gene, Alters Plant Growth and Salinity Stress Responses in Transgenic Rice. Front Plant Sci. 2017, 8, 306. [Google Scholar] [CrossRef]
- Serra, T.S.; Figueiredo, D.D.; Cordeiro, A.M.; Almeida, D.M.; Lourenço, T.; Abreu, I.A.; Sebastián, A.; Fernandes, L.; Contreras-Moreira, B.; Oliveira, M.M.; Saibo, N.J. OsRMC, a negative regulator of salt stress response in rice, is regulated by two AP2/ERF transcription factors. Plant Mol Biol. 2013, 82, 439–55. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, R.; Schippers, J.H.; Mieulet, D.; Watanabe, M.; Hoefgen, R.; Guiderdoni, E.; Mueller-Roeber, B. SALT-RESPONSIVE ERF1 is a negative regulator of grain filling and gibberellin-mediated seedling establishment in rice. Mol Plant. 2014, 7, 404–21. [Google Scholar] [CrossRef] [PubMed]
- Cheng, X.; Zhang, S.; Tao, W.; Zhang, X.; Liu, J.; Sun, J.; Zhang, H.; Pu, L.; Huang, R.; Chen, T. INDETERMINATE SPIKELET1 Recruits Histone Deacetylase and a Transcriptional Repression Complex to Regulate Rice Salt Tolerance. Plant Physiol. 2018, 178, 824–837. [Google Scholar] [CrossRef] [PubMed]
- Liu, D.; Chen, X.; Liu, J.; Ye, J.; Guo, Z. The rice ERF transcription factor OsERF922 negatively regulates resistance to Magnaporthe oryzae and salt tolerance. J Exp Bot. 2012, 63, 3899–911. [Google Scholar] [CrossRef]
Figure 1.
AP2/ERF transcription factor family classification and structural features. They can be divided into five main groups based on the number of AP2/ERF structural domains, namely AP2 (APETALA2), DREB (dehydration-responsive element binding protein), ERF (ethylene responsive factor), RAV (related to ABI3/VP1) and Soloist. AP2 domain contains two AP2 structural domains, DREB and ERF domain contains an AP2 structural domain, RAV domain contains a AP2 structural domain and a B3 structural domain, Soloist domain contains a AP2-like structural domain.
Figure 1.
AP2/ERF transcription factor family classification and structural features. They can be divided into five main groups based on the number of AP2/ERF structural domains, namely AP2 (APETALA2), DREB (dehydration-responsive element binding protein), ERF (ethylene responsive factor), RAV (related to ABI3/VP1) and Soloist. AP2 domain contains two AP2 structural domains, DREB and ERF domain contains an AP2 structural domain, RAV domain contains a AP2 structural domain and a B3 structural domain, Soloist domain contains a AP2-like structural domain.
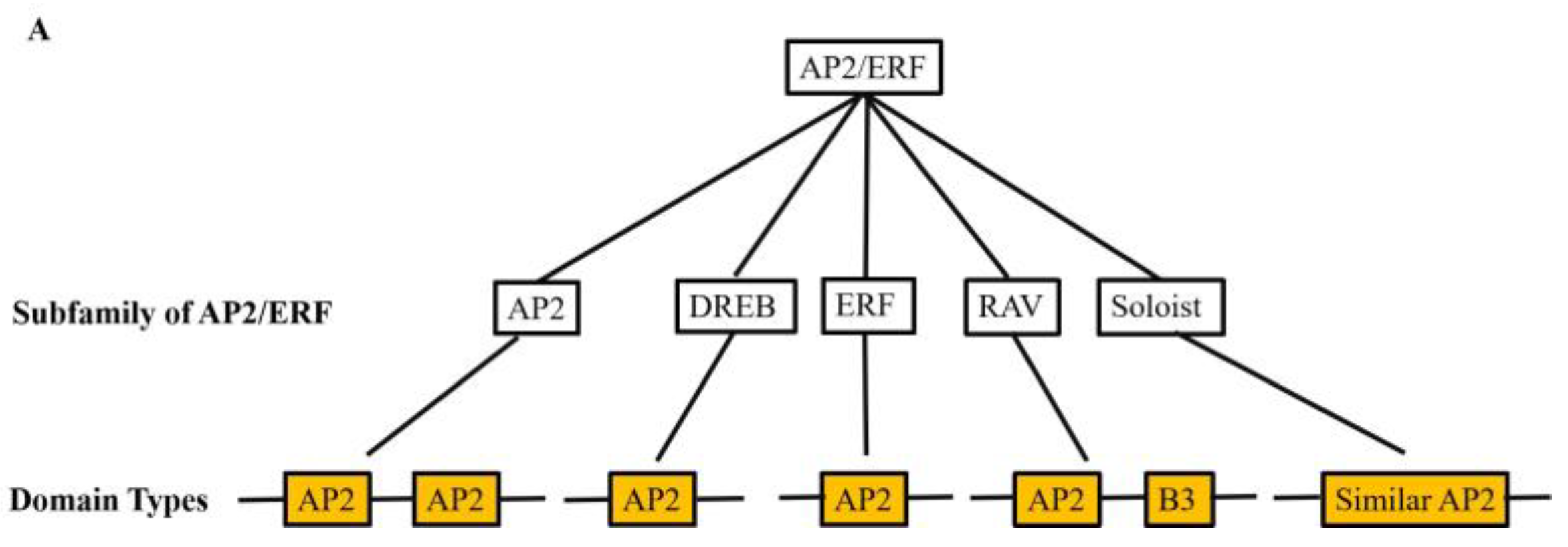
Figure 2.
AP2/ERF transcription factor involved in the regulation of multiple abiotic stress responses in plants. For example, drought stress, salt stress, temperature stress, nutritional element stress and heavy metals stress.
Figure 2.
AP2/ERF transcription factor involved in the regulation of multiple abiotic stress responses in plants. For example, drought stress, salt stress, temperature stress, nutritional element stress and heavy metals stress.
Figure 3.
AP2/ERF transcription factor by participating in the regulation of hormone-mediated response networks during abiotic stresses. Abiotic stress can alter the production and distribution of plant hormones and subsequently mediate stress responses through AP2/ERF family transcription factors and hormone signalling components. ABA: abscisic acid; GA: gibberellin; IAA: auxin; BR: oleuropein lactone; CTK: cytokinin; ET: ethylene. Arrows ends indicate activation effect, bar ends indicate repression effect.
Figure 3.
AP2/ERF transcription factor by participating in the regulation of hormone-mediated response networks during abiotic stresses. Abiotic stress can alter the production and distribution of plant hormones and subsequently mediate stress responses through AP2/ERF family transcription factors and hormone signalling components. ABA: abscisic acid; GA: gibberellin; IAA: auxin; BR: oleuropein lactone; CTK: cytokinin; ET: ethylene. Arrows ends indicate activation effect, bar ends indicate repression effect.
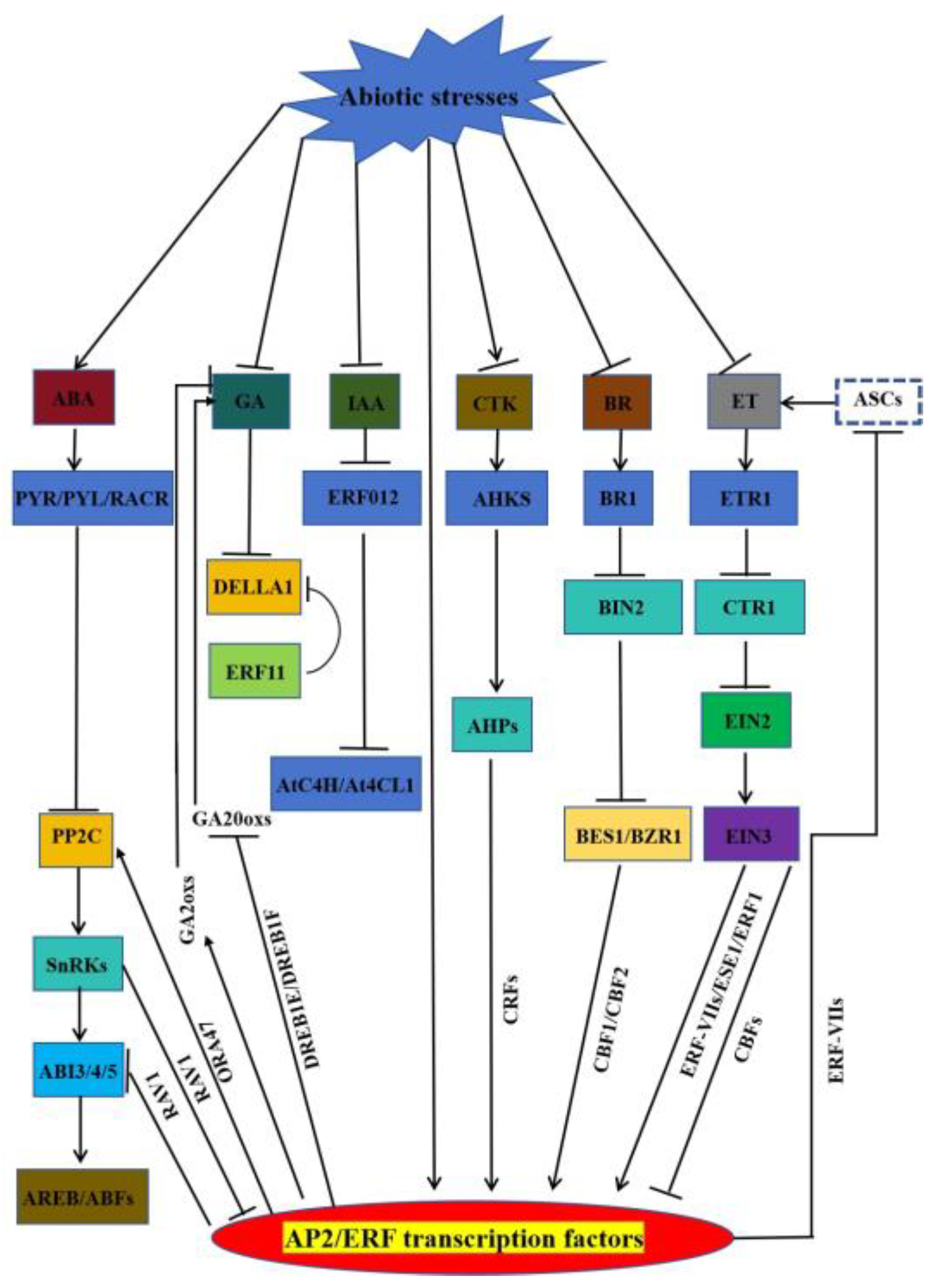

Table 1.
Abiotic stress responsive AP2/ERF transcription factors: their regulations and target genes in plants.
Table 1.
Abiotic stress responsive AP2/ERF transcription factors: their regulations and target genes in plants.
| Abiotic Stress Type | AP2/ERF transcription factors | Species | Reference |
| Drought | AtDREB1A | Arabidopsis thaliana L. | [141] |
| Cold | DREB1/CBF | Arabidopsis thaliana L. | [142] |
| Cold | DREB2 | Arabidopsis thaliana L. | [142] |
| Cold, salt, drought | GmDREBa | Soybean (Glycine max L.) | [143] |
| Cold, salt, drought | GmDREBc | Soybean (Glycine max L.) | [143] |
| Drought | ERF1-V | Wheat (Triticum aestivum) | [144] |
| Temperature | ZjDREB1.4 | Zoysiagrass (Zoysia japonica S.) | [145] |
| Cold, salt, drought | ZmEREB3 | Maize (Zea mays L.) | [146] |
| Salt | ZmEREB20 | Maize (Zea mays L.) | [70] |
| Drought | ZmEREB46 | Maize (Zea mays L.) | [147] |
| Drought | ZmEREB60 | Maize (Zea mays L.) | [148] |
| Drought | ZmEREB137 | Maize (Zea mays L.) | [149] |
| Waterlogging | ZmEREB180 | Maize (Zea mays L.) | [150] |
| Osmotic | ZmEREB204 | Maize (Zea mays L.) | [151] |
| Drought | ZmEREB240 | Maize (Zea mays L.) | [152] |
| Cold | OsDREB1A | Rice (Oryza sativa) | [153] |
| Temperature | OsDREB1B | Rice (Oryza sativa) | [154] |
| Cold, salinity | OsDREB1D | Rice (Oryza sativa) | [155] |
| Drought | OsDREB1E | Rice (Oryza sativa) | [155] |
| Drought | OsDREB1G; OsDREB1I | Rice (Oryza sativa) | [69] |
| Drought | OsDREB2B | Rice (Oryza sativa) | [69] |
| Salt, drought, temperature | OsDREB4-1 | Rice (Oryza sativa) | [156] |
| Salt, drought, temperature | OsDREB1F | Rice (Oryza sativa) | [157] |
| Temperature | OsWR2 | Rice (Oryza sativa) | [158] |
| Drought | OsERF71 | Rice (Oryza sativa) | [159] |
| Drought | OsLG3; OsERF62; OsRAF | Rice (Oryza sativa) | [160] |
| Drought | OsAP37 | Rice (Oryza sativa) | [161] |
| Salt | OsAP23 | Rice (Oryza sativa) | [162] |
| Salt, drought, temperature | OsEREBP2 | Rice (Oryza sativa) | [163] |
| Salt | SERF1 | Rice (Oryza sativa) | [164] |
| Salt | OsIDS1 | Rice (Oryza sativa) | [165] |
| Salt | OsERF922 | Rice (Oryza sativa) | [166] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



