
Submitted:
14 December 2023
Posted:
15 December 2023
You are already at the latest version
Abstract
The study of trophic relationships among closely related species plays an important role in deepening our understanding of the resource utilization characteristics, differentiation patterns, and population dynamics of co-occurring species in the same habitat. This research uses two congeneric fish species, Pennahia pawak and Pennahia anea, as examples. Based on stomach content analysis and carbon-nitrogen stable isotope analysis, a comparative analysis of their feeding habits and trophic niches is conducted. Additionally, spatial niche analysis is employed to explore the coexistence and competitive mechanisms between these two closely related fish species. The results showed that both P. pawak and P. anea had similar feeding habits, mainly feeding on Pisces and Macrura, but there were some feeding preferences between the two species. Interspecific prey overlap was meaningful but not significant, with an overlap coefficient of 0.56. There were significant differences in feeding habits between the two species during different seasons. Based on stable isotope analyses, body length of both species had a significant positive correlation with δ15N (Pearson r=0.65, P<0.01; Pearson r=0.44, P<0.01), but not with δ13C (P>0.05). Standard ellipse corrected area (SEAc) of P. pawak and P. anea were 0.65 and 1.16, while the trophic niche overlap was only 0.29, which was not significant; the spatial niche width of the two was 1.19 and 0.58, respectively, and the spatial niche overlap was only 0.20, which was not significant. Intra-species competition was intense in P. pawak and P. anea, but reduced feeding specialization had a mitigating effect on intra-species competition. Interspecific competition was present but not significant, and was mitigated by trophic and spatial niche differentiation.
Keywords:
Pennahia pawak
; Pennahia anea
; stomach contents analysis
; stable isotope analysis
; Sympatonic coexistence
1. Introduction
Competition is the phenomenon of two organisms competing for a particular resource. Competition between different species for food and space is relatively common in communities, and the result is almost always a victory for one side and the exclusion of the other. Competition is most common between two species with similar food habits [1,2]. Intraspecific food competition, which can generally be defined as competition between two or more conspecifics for a critical and limited food resource with strong selective forces, is central to species biology and plays a crucial role in limiting populations, eliminating unsuitable genes, and driving evolution. Interspecific food competition is the basis for studying the trophic dynamics of ecological food webs [3]. In marine ecosystems, food linkages are one of the most important interrelationships between organisms. Different from terrestrial vertebrates, the competition for food of fish is more important than the competition for habitat, and they both depend on and compete with each other through hunting and being hunted, forming a dynamic food relationship [4]. Many scholars both domestically and internationally have conducted research on fish feeding relationships [5,6,7,8,9], but most of them have focused on relationships between different species or non-congeneric species. The interrelationships among closely related species are a particularly unique and significant aspect of species relationships [10]. With the development of theoretical research and scientific technology, research on food relationships among closely related species has started to receive more attention, and hope to reveal the mechanism of species coexistence more deeply [11,12,13].
Feeding relationship research tools mainly include the traditional stomach content analysis (SCA) and stable isotope analysis (SIA) [14,15]. SCA is a traditional method that has been used for a long time in marine feeding ecology, and its advantage is that it can accurately and intuitively reflect the composition and taxonomic characteristics of the prey consumed by fish [16]. However, traditional stomach contents analysis is time-consuming and labor-intensive, has the drawbacks of only being able to characterize transient feeding information as well as not being able to determine the fragmented prey organisms being digested, whereas stable isotopes of carbon and nitrogen in tissues, which can record feeding information on medium to long time scales, have been proved to be the best alternative and complement to stomach contents analysis [17], and have been widely used in the reconstruction of feeding habits of aquatic organisms. However, the stable isotope technique also has some limitations in that it is unable to visually obtain information on ingested food types and sizes, so SIA and SCA were combined as complementary techniques to more comprehensively and accurately reflect the food relationships of organisms in the study of feeding ecology.
Pennahia pawak and Pennahia anea have relatively important ecological niches as relatively stable resource fish species in the Beibu Gulf [18]. The studies on P. pawak and P. anea have mainly focused on fishery biology [19,20], growth and mortality [21], reproductive biology [22], population structural characteristics and spatial-temporal distribution [23,24], and feeding ecology [25], whereas the studies on the interspecies feeding relationship between P. pawak and P. anea have not been reported. The two species are not only closely related, but also have overlapping distribution areas [18], so there may be potential feeding competition between them. In this study, using stomach content analysis and carbon-nitrogen stable isotope analysis, we compared and analyzed the differences in the feeding habits and trophic niches of P. pawak and P. anea. We combined spatial niche analysis to elucidate the feeding relationship between these two closely related species, aiming to explore the mechanisms of coexistence of sympatric species in the Beibu Gulf.
2. Materials and methods
2.1. Sample collection and processing
Samples for trophic niche analysis were collected in 2020 from Guangdong Jianghong and Beihai fishing harbors, with 1506 and 1684 samples of P. pawak and P. anea, respectively, randomly sampled in four seasons. The samples collected in each season were frozen and sent back to the laboratory for the next step of processing. After thawing the samples in the laboratory, biological indicators such as total length, body length and weight were measured according to Specifications for oceanographic survey [26], and stomach contents samples were retained for freezing and preservation.

Table 1.
Sampling information of two Pennahia species in the Beibu Gulf.
| Species | Number of sample | |||
|---|---|---|---|---|
| Spring | Summer | Autumn | Winter | |
| P. pawak | 417 | 226 | 579 | 284 |
| P. anea | 317 | 241 | 788 | 388 |
2.2. Stomach contents analysis
After thawing the stomach contents samples, the food mass was removed and placed in a clean petri dish, and the excess water on the surface of the food mass was carefully blotted out with filter paper. Subsequently, the identification of the different species of prey in the food mass was continued using forceps to identify the prey organisms to the smallest possible taxonomic order, and the food mass was weighed (to the nearest 0.01 g) and the data recorded.
In this study, the repletion index (RI) and vacuity coefficient (VC) were used to determine the feeding intensity of fish [27]. The formulas are as follows:
where Wf is the total weight f of the fish, is the total weight f of all the food in the stomach of the fish and the vacuity coefficient (VC).
The importance of prey organisms was evaluated using the Index of Relative Importance percentage (IRI%) [28] with the following formula:
where N is the number of a given prey as a percentage of the total number of prey; W is the weight of a given prey as a percentage of the total weight of prey; and F is the percentage of frequency of occurrence of each prey.
The Shannon-Wiener diversity index (H') was used to evaluate the width of the trophic niche of the two fish species. The overlap of trophic niches between the two species [29,30] was also calculated by using Pianka's coefficient of ecosystem overlap (Oij) to evaluate the intensity of competition between the two species.
Hi' is the width of the trophic niche for fish i, and Pik is the percentage of the number of prey k in the food of fish i(N).
Where s is the total number of preys consumed by the two fish species, Pik and Pjk represent the number percentage (N) of prey k in the prey composition of fish i and j, respectively. The value of Oij ranges from 0 to 1. The larger the value, the higher the food similarity, that is, the fiercer the food competition. Oij> 0.3 means that the overlap is effective, and Oij>0.6 has significant overlap.
Individual specialization (IS) was used to evaluate the intensity of intraspecific competition between the two species [31], and Whin-individual component (WIC) to the width of its trophic niche (H') was calculated. The range was 0-1, and the smaller the ratio, the lower the individual overlap and the higher individual specialization [32]. The related calculation formula is as follows:
where Pi is the percentage of prey consumed by individual i to the population of prey in that population; Pik is the percentage of prey consumed by individual i to the population of prey in that population.
2.3. Carbon and nitrogen stable isotope analysis
A rectangular cut was made below the dorsal fin and above the lateral line of the fish, the fish epidermis was peeled off from the inner side of the skin, and an appropriate amount of muscle sample was taken from the notch in a 2 ml centrifuge tube and cryopreserved for subsequent processing.
The muscle sample was placed in the sample tray of the freeze-dryer (Christ, Germany, Alpha1-4/2-4LD Plus), freeze-dried for 48 hours at -48℃ until the muscle maintained constant weight. The sample was taken out and 2 small steel balls were put into each centrifuge tube containing the sample. Put in the homogenizer (BIOSPEC MiniBeadbeater-16) and grind for 1min. The ground muscle powder was embedded and sent to the Isotope Laboratory of the School of Marine Meteorology, Guangdong Ocean University, where it was measured using an EA Isolink Elemental Analyzer and a 253 Plus Isotope Mass Spectrometer (Thermo, USA).
The stable isotope ratios are expressed according to the internationally accepted δ-value [33], which is calculated by the formula:
Where δX denotes the carbon stable isotope ratio δ13C or nitrogen stable isotope ratio δ15N, Rsample denotes the ratio of carbon or nitrogen stable isotope in the sample, and Rstandard denotes the ratio of carbon or nitrogen stable isotope in the standard. Where the carbon isotope ratio is 13C/12C and the nitrogen stable isotope ratio is 15N/14N.
Trophic level formula [34]:
where TL denotes the trophic level of the fish, δ15Nsample denotes the nitrogen stable isotope signature of the sample; δ15N0 is the nitrogen stable isotope signature of the baseline organism; δ15Nc denotes the nitrogen isotope enrichment (3.4 ‰); and TLb denotes the trophic level of the baseline organism, and in this study, Amusium pleuronectes was selected as the baseline organism (8.89 ‰) [35], which had a trophic level of 2.
The degree of overlap between SEAc was used to quantify the area of isotopic ecotope overlap between P. pawak and P. anea [34], and the overlap of the two species was expressed as the proportion of overlap in the maximum likelihood estimate of SEAc, which represents the point estimate of ecotope overlap between the two species, with overlap close to 0 indicating the two ellipsoids are separated, and a value close to 1 indicating that the two ellipsoids are completely overlapped. According to Schoener's D index and according to the results of other stable isotope niche area overlap studies [36], Bij is the niche overlap index, ranging from 0 to 1, with larger values indicating higher overlap, and Bij greater than 0.3 is regarded as meaningful overlap, and greater than 0.6 is regarded as significant overlap [37].
2.4. Spatial niche
The summer close season is from 12:00 a.m. May 1 to 12:00 a.m. August 16. The data used for the spatial niche analysis came from surveys of 26 stations in the Beibu Gulf before (April) and after (August) period of fishing moratorium. The survey vessel had a power of 441KW, and the net was a bottom trawl with a 20m wide net opening, a maximum mesh of 5cm, and a sac mesh of 2cm. 1 hour of trawling was conducted at each station. Samples were collected and preserved on ice and brought back to the laboratory for classification and biological determination. The number and weight of each species were recorded and converted to biomass (kg/km2) and abundance (ind/km2) per unit area. Sample sampling and analysis were performed according to Specifications for oceanographic survey [28].
(1) Resource intensity [38]
Where D is the resource density (kg·km-2); C is the catch rate (kg·h-1); E is the escape rate taking the empirical value (0.5) [23]; a is the hourly swept area (km2) of the survey vessel, the swept width is taken as 1/2 of the length of the float line, and the towing speed is taken as the average towing speed 3.0 kn.
(2) Spatial niche width and overlap
Spatial niche width and overlap were estimated based on catch, where spatial niche width was expressed using the Shannon-Wiener index (H') [39], and the spatial niche overlap index (Qij) was expressed using the Pianka index [29] with the following formula:
Where: Hʹ denotes the spatial ecological niche width, Pij denotes the mass percentage of fish i in the total biomass of the jth station. Bij is the ecological niche overlap index, which ranges from 0 to 1, with larger values indicating a higher degree of overlap, and Bij greater than 0.3 is regarded as meaningful overlap, and greater than 0.6 is regarded as significant overlap [37].
2.5. Data processing
Data were processed, analyzed and graphed using Excel 2016 and SPSS 25.0, and Pearson analysis was used to test for linear correlation between body length and trophic level for two Pennahia species. Trophic niche of δ13C and δ15N of the two Pennahia species was performed using the SIAR data package in R 3.6.3, and mean centrifugal distance (CD), carbon range (CR), nitrogen range (NR), mean nearest neighbor distance (MNND), standard deviation of nearest neighbor distance (SDNND), standard ellipse corrected area (SEAc), the area of the convex hull (TA), and other ecological indicators [34,37]. The distribution of resources was mapped using ArcGIS 10.8 software.
3. Results
This section may be divided by subheadings. It should provide a concise and precise description of the experimental results, their interpretation, as well as the experimental conclusions that can be drawn.
3.1. Feeding habit
3.1.1. Differences in feeding habits within species
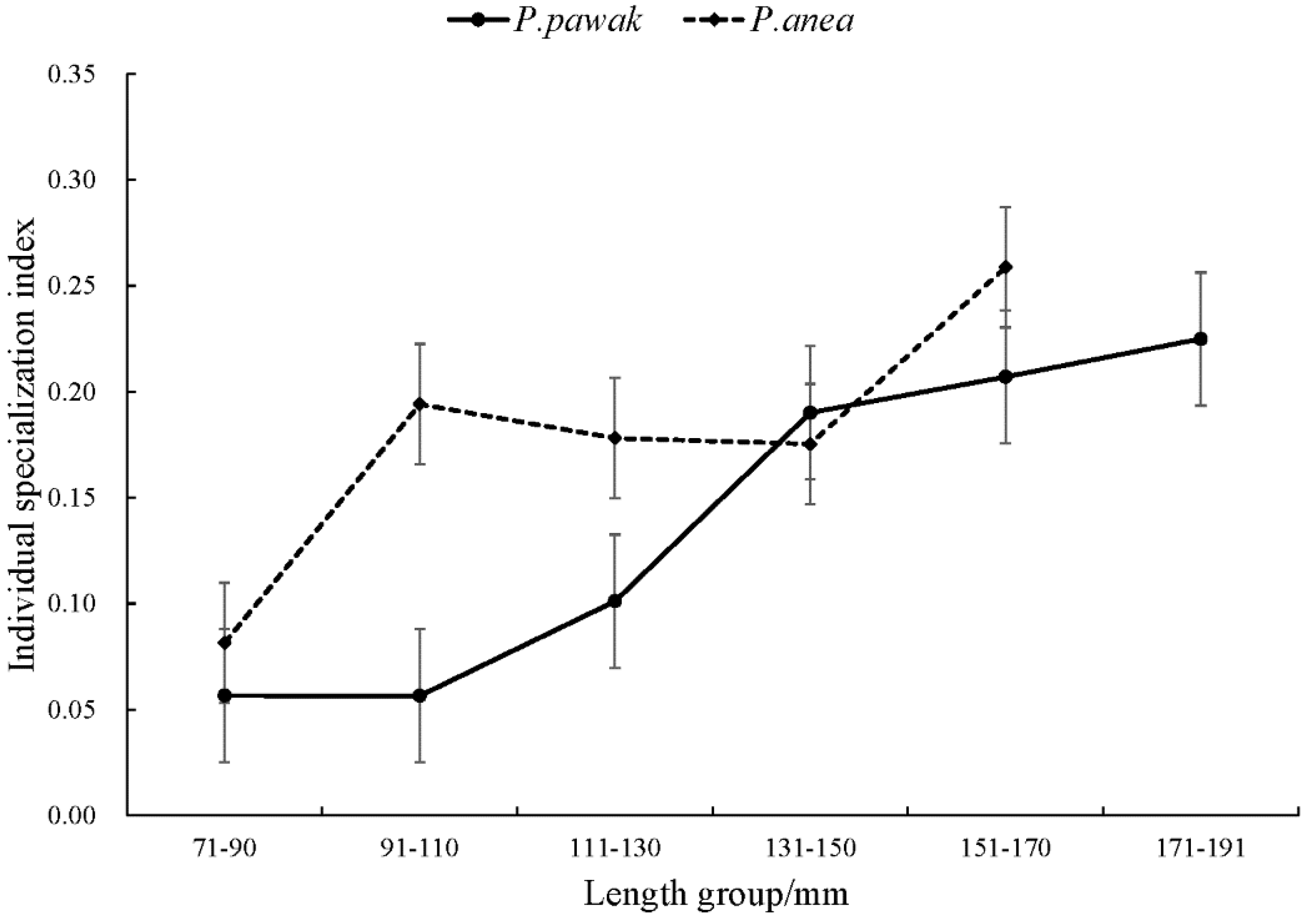
Intraspecific feeding habits of P. pawak varied significantly between seasons. In spring and winter, the stomach contents of P. pawak had the highest proportion of Macrura (77.99% and 63.78%, respectively), and in summer and autumn, the highest proportion of Pisces (47.37% and 34.73%, respectively); Gastropoda were also present only in spring. The prey overlap coefficients among the length groups of P. pawak were all greater than 0.6 (except for 171-191 mm), and there was significant food overlap. The prey overlap coefficients between the 71-80mm length group and the 91-110mm length group, the 111-130mm length group, and between the 91-110mm length group and the 111-130mm length group were close to 1, with almost complete overlap of prey species. Meanwhile, individual specialization index of the groups of different lengths ranged from 0.06 to 0.22, with an overall increasing trend with the increase of length.
Intraspecific feeding habits of P. anea differed markedly between seasons. The highest proportion of Macrura in the stomach contents of P. anea was found in spring and winter, and the highest proportion of Pisces was found in summer and autumn; in addition, gastropods were present only in spring. P. anea prey overlap coefficients were greater than 0.6 between length groups, except between 171-190mm length group and 111-130mm length group, 131-151mm length group, 151-170mm length group, which were less than 0.6, and there was significant food overlap. Among them, the highest prey overlap coefficient was found between 71-90mm length group and 171-190mm length group, followed by 131-150mm length group and 151-170mm length group. Meanwhile, the fluctuation range of individual specialization index of different length groups was 0.08-0.26, which showed a fluctuating upward trend with the increase of length.
Figure 1.
The dietary composition of P. pawak and P. anea by different seasons. Note: From the inside out are spring, summer, autumn and winter. On the left is P. pawak and on the right is P. anea.
Figure 1.
The dietary composition of P. pawak and P. anea by different seasons. Note: From the inside out are spring, summer, autumn and winter. On the left is P. pawak and on the right is P. anea.
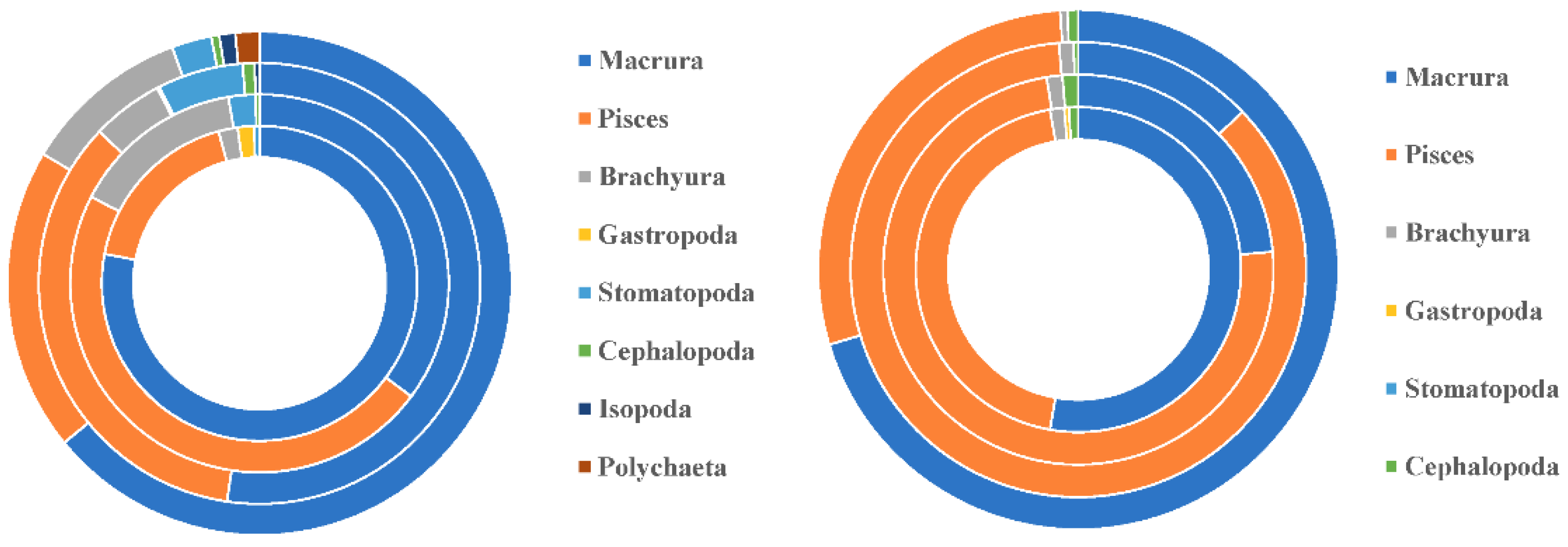
Table 2.
The diet overlap coefficient of P. pawak and P. anea in each length group.
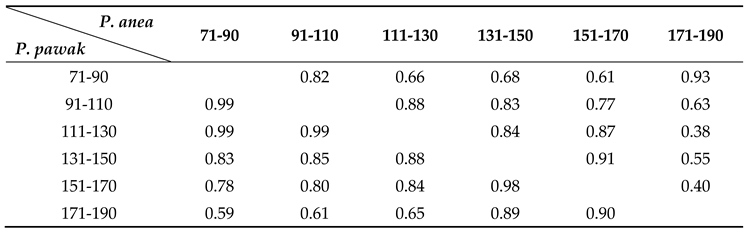
Figure 2.
Individual specialization index of P. pawak and P. anea in different length groups.
3.1.2. Differences in feeding habits between species
P. pawak and P. anea have similar feeding habits and some feeding preferences. The following are some of the most common species in the genus, although both species of Pennahia genus take Pisces and Macrura as prey, the proportion of Pisces in the prey of P. anea is obviously higher than that of P. pawak; there are also some differences in specific species of the two species of Pennahia genus, among which P. anea are dominated by Alpheus, Bregmaceros, and Stolephorus, whereas P. pawak, in addition to Alpheus and Bregmaceros, Metapenaeopsis barbata and Alpheus bisincisus also accounted for a certain proportion. The trophic niche widths for P. pawak and P. anea were 2.21 and 2.29, respectively, and the prey overlap coefficient was 0.56, reaching the level of meaningful overlap.
Table 3.
Composition of food items of P. pawak and P. anea in the Beibu Gulf in 2020.
| Prey | P. pawak | P. anea | ||||||
|---|---|---|---|---|---|---|---|---|
| Science name | W% | N% | F% | IRI% | W% | N% | F% | IRI% |
| Pisces | ||||||||
| Leiognathidae | 0.46 | 0.22 | 0.56 | 0.02 | 0.42 | 0.92 | 3.86 | 0.03 |
| Trichiuridae | 0.87 | 0.88 | 2.22 | 0.18 | 0.47 | 0.23 | 0.97 | + |
| Thrissa dussumieri | 2.90 | 0.44 | 1.11 | 0.17 | 3.02 | 0.69 | 2.90 | 0.07 |
| Champsodon atridorsalis | 0.35 | 0.66 | 1.67 | 0.08 | 0.10 | 0.12 | 0.48 | + |
| Photopectoralis bindus | 1.98 | 2.84 | 7.22 | 1.60 | 1.09 | 0.69 | 2.90 | 0.03 |
| Stolephorus heterolobas | 0.66 | 0.44 | 1.11 | 0.06 | 0.23 | 0.12 | 0.48 | + |
| Secutor ruconius | 0.23 | 0.22 | 0.56 | 0.01 | 0.83 | 0.35 | 1.45 | 0.01 |
| Bregmaceros rarisquamosus | 1.54 | 6.35 | 16.11 | 5.82 | 8.07 | 23.44 | 98.07 | 19.14 |
| Apogonidae | 4.52 | 6.13 | 15.56 | 7.60 | 0.21 | 0.12 | 0.48 | + |
| Bregmaceros | 3.25 | 7.88 | 20.00 | 10.20 | 9.07 | 28.52 | 119.32 | 27.79 |
| Gobiidae | 2.44 | 3.50 | 8.89 | 2.42 | 0.05 | 0.12 | 0.48 | + |
| Atherinidae | 0.33 | 0.22 | 0.56 | 0.01 | 1.55 | 0.46 | 1.93 | 0.02 |
| Bregmaceros nectabanus | 3.08 | 3.94 | 10.00 | 3.22 | 6.20 | 7.51 | 31.40 | 2.67 |
| Stolephorus indicus | 0.23 | 0.22 | 0.56 | 0.01 | 1.26 | 0.35 | 1.45 | 0.01 |
| Stolephorus | 2.75 | 2.41 | 6.11 | 1.45 | 47.16 | 26.10 | 109.18 | 49.55 |
| Champsodon | 0.63 | 0.88 | 2.22 | 0.15 | - | - | - | - |
| Jaydia striata | 0.11 | 0.22 | 0.56 | 0.01 | - | - | - | - |
| Trypauchen vagina | 0.94 | 0.66 | 1.67 | 0.12 | - | - | - | - |
| Thryssa | 0.21 | 0.22 | 0.56 | 0.01 | - | - | - | - |
| Bregmaceros macclellandii | 0.59 | 0.22 | 0.56 | 0.02 | - | - | - | - |
| Diaphus knappi | 0.11 | 0.22 | 0.56 | 0.01 | - | - | - | - |
| Parachaeturichthys polynema | 2.56 | 1.53 | 3.89 | 0.73 | - | - | - | - |
| Carangidae | 0.54 | 0.22 | 0.56 | 0.02 | - | - | - | - |
| Leiognathus berbis | 0.67 | 0.22 | 0.56 | 0.02 | - | - | - | - |
| Sirembo imberbis | 0.22 | 0.22 | 0.56 | 0.01 | - | - | - | - |
| Callionymus octostigmatus | 1.15 | 0.22 | 0.56 | 0.03 | - | - | - | - |
| Sirembo | 0.04 | 0.22 | 0.56 | 0.01 | - | - | - | - |
| Hypoatherina valenciennei | - | - | - | - | 0.64 | 0.12 | 0.48 | + |
| Leiognathus nuchalis | - | - | - | - | 0.08 | 0.12 | 0.48 | + |
| Stolephorus commersonnii | - | - | - | - | 4.59 | 1.62 | 6.76 | 0.26 |
| Stolephorus zollingeri | - | - | - | - | 0.60 | 0.23 | 0.97 | + |
| Sardinella | - | - | - | - | 6.18 | 0.92 | 3.86 | 0.17 |
| Stolephorus chinensis | - | - | - | - | 0.56 | 0.35 | 1.45 | 0.01 |
| Macrura | ||||||||
| Alpheus | 5.16 | 16.19 | 41.11 | 40.26 | 1.05 | 1.50 | 6.28 | 0.10 |
| Metapenaeopsis palmensis | 0.85 | 1.09 | 2.78 | 0.25 | 0.48 | 0.23 | 0.97 | + |
| Alpheus bisincisus | 3.57 | 5.47 | 13.89 | 5.76 | 0.70 | 0.35 | 1.45 | 0.01 |
| Metapenaeopsis barbata | 9.47 | 7.00 | 17.78 | 13.43 | 0.52 | 0.12 | 0.48 | - |
| Solenocera crassicornis | 2.55 | 1.75 | 4.44 | 0.88 | 0.46 | 0.12 | 0.48 | - |
| Metapenaeopsis | 0.70 | 1.31 | 3.33 | 0.31 | - | - | - | - |
| Alpheus brevicristatus | 0.84 | 1.31 | 3.33 | 0.33 | - | - | - | - |
| Penaeidae | 1.59 | 1.31 | 3.33 | 0.44 | - | - | - | - |
| Parapenaeopsis | 0.10 | 0.44 | 1.11 | 0.03 | - | - | - | - |
| Parapenaeopsis | 0.51 | 0.22 | 0.56 | 0.02 | - | - | - | - |
| Metapenaeopsis acclivis | 0.56 | 0.22 | 0.56 | 0.02 | - | - | - | - |
| Parapenaeus sextuberculatus | 0.43 | 0.44 | 1.11 | 0.04 | - | - | - | - |
| Parapenaeus | 0.17 | 0.22 | 0.56 | 0.01 | - | - | - | - |
| Alpheus japonicus | 0.22 | 0.44 | 1.11 | 0.03 | - | - | - | - |
| Alpheus distinguendus | 1.21 | 1.75 | 4.44 | 0.60 | - | - | - | - |
| Metapenaeus | 0.06 | 0.22 | 0.56 | 0.01 | - | - | - | - |
| Trachypenaeus curvirostris | 0.26 | 0.44 | 1.11 | 0.04 | - | - | - | - |
| Palaemonidae | 0.03 | 0.22 | 0.56 | 0.01 | - | - | - | - |
| Miyadiella podophthalmus | 0.02 | 0.44 | 1.11 | 0.02 | - | - | - | - |
| Parapenaeus longipes | 0.68 | 0.88 | 2.22 | 0.16 | - | - | - | |
| Parapenaeopsis cornuta | - | - | - | - | 0.54 | 0.46 | 1.93 | 0.01 |
| Trachypenaeus pescadoreensis | - | - | - | - | 0.52 | 0.12 | 0.48 | + |
| Parapenaeopsis incisa | - | - | - | - | 0.21 | 0.23 | 0.97 | + |
| Brachyura | ||||||||
| Portunus | 0.18 | 1.31 | 3.33 | 0.23 | 0.03 | 0.12 | 0.48 | - |
| Eucrate alcocki | 0.06 | 0.22 | 0.56 | 0.01 | - | - | - | - |
| Typhlocarcinops canaliculata | 0.05 | 0.22 | 0.56 | 0.01 | - | - | - | - |
| Charybdis hellerii | 0.21 | 0.44 | 1.11 | 0.03 | - | - | - | - |
| Charybdis variegata brevispinosa | 0.55 | 0.66 | 1.67 | 0.09 | - | - | - | - |
| Lissocarcinus laevis | 0.03 | 0.22 | 0.56 | 0.01 | - | - | - | - |
| Portunus hastatoides | 0.58 | 2.19 | 5.56 | 0.70 | - | - | - | - |
| Jonas | 0.04 | 0.22 | 0.56 | 0.01 | - | - | - | - |
| Eucrate solaris | 0.12 | 0.22 | 0.56 | 0.01 | - | - | - | - |
| Charybdis | 0.25 | 0.88 | 2.22 | 0.11 | - | - | - | - |
| Petrolisthes | 0.05 | 0.22 | 0.56 | 0.01 | - | - | - | - |
| Portunus argentatus | 1.43 | 2.41 | 6.11 | 1.07 | - | - | - | - |
| Charybdis truncata | 0.07 | 0.22 | 0.56 | 0.01 | - | - | - | - |
| Carcinoplax purpurea | 0.10 | 0.22 | 0.56 | 0.01 | - | - | - | - |
| Charybdis vadorum | 0.06 | 0.22 | 0.56 | 0.01 | - | - | - | - |
| Charybdis variegata | 0.10 | 0.44 | 1.11 | 0.03 | - | - | - | - |
| Typhlocarcinus villosusStimpson | - | - | - | - | 0.03 | 0.12 | 0.48 | - |
| Stomatopoda | ||||||||
| Gryllotalpidae | 0.16 | 0.22 | 0.56 | 0.01 | 0.09 | 0.12 | 0.48 | + |
| Oratosquillina interrupta | 0.48 | 0.44 | 1.11 | 0.05 | - | - | - | - |
| Oratosquilla oratoria | 0.35 | 0.44 | 1.11 | 0.04 | - | - | - | - |
| Anchisquilla fasciata | 0.16 | 0.22 | 0.56 | 0.01 | - | - | - | - |
| Oratosquillina | 0.28 | 0.44 | 1.11 | 0.04 | - | - | - | - |
| kempina stridulans | 0.77 | 0.66 | 1.67 | 0.11 | - | - | - | - |
| Squillidae | - | - | - | - | 0.23 | 0.92 | 3.86 | 0.03 |
| Cephalopoda | ||||||||
| Cephalopoda | 0.17 | 1.53 | 3.89 | 0.30 | 2.00 | 0.69 | 2.90 | 0.05 |
| Isopoda | ||||||||
| isopoda | 0.15 | 0.88 | 2.22 | 0.10 | 0.04 | 0.23 | 0.97 | + |
| Gastropoda | ||||||||
| Turritella terebra | 0.32 | 1.53 | 3.89 | 0.33 | - | - | - | - |
| Polychaeta | ||||||||
| Nereis | 0.14 | 0.22 | 0.56 | 0.01 | - | - | - | - |
| Sipunculs nudus | 0.23 | 0.44 | 1.11 | 0.03 | - | - | - | - |
Note: “+” indicates that the ratio was smaller than 0.01 % ,“-” indicates absence.
3.2. Isotopic characteristics and trophic levels
The δ13C and δ15N isotopic characteristics of P. pawak and P. anea are different to some extent. The δ13C of P. pawak ranged from -18.08 to -15.22‰, with a mean value of -16.57‰, and the δ15N ranged from 14.68 to 16.40‰, with a mean value of 15.60‰. After Pearson analysis, length of P. pawak had a significant positive correlation with δ15N (Pearson r=0.65 , P<0.01), but no significant correlation with δ13C (P>0.05); δ13C of P. anea ranged from -18.12 to -15.10‰, with a mean value of -16.58‰, and δ15N ranged from 13.65 to 16.63‰, with a mean value of 15.18‰. After Pearson analysis, length of P. anea had a significant positive correlation with δ15N (Pearson r=0.44, P<0.01), while there was no significant correlation with δ13C (P>0.05). The δ13C ranges of P. pawak and P. anea were similar, with P. anea being slightly higher, while P. pawak had a larger δ15N range than P. anea.
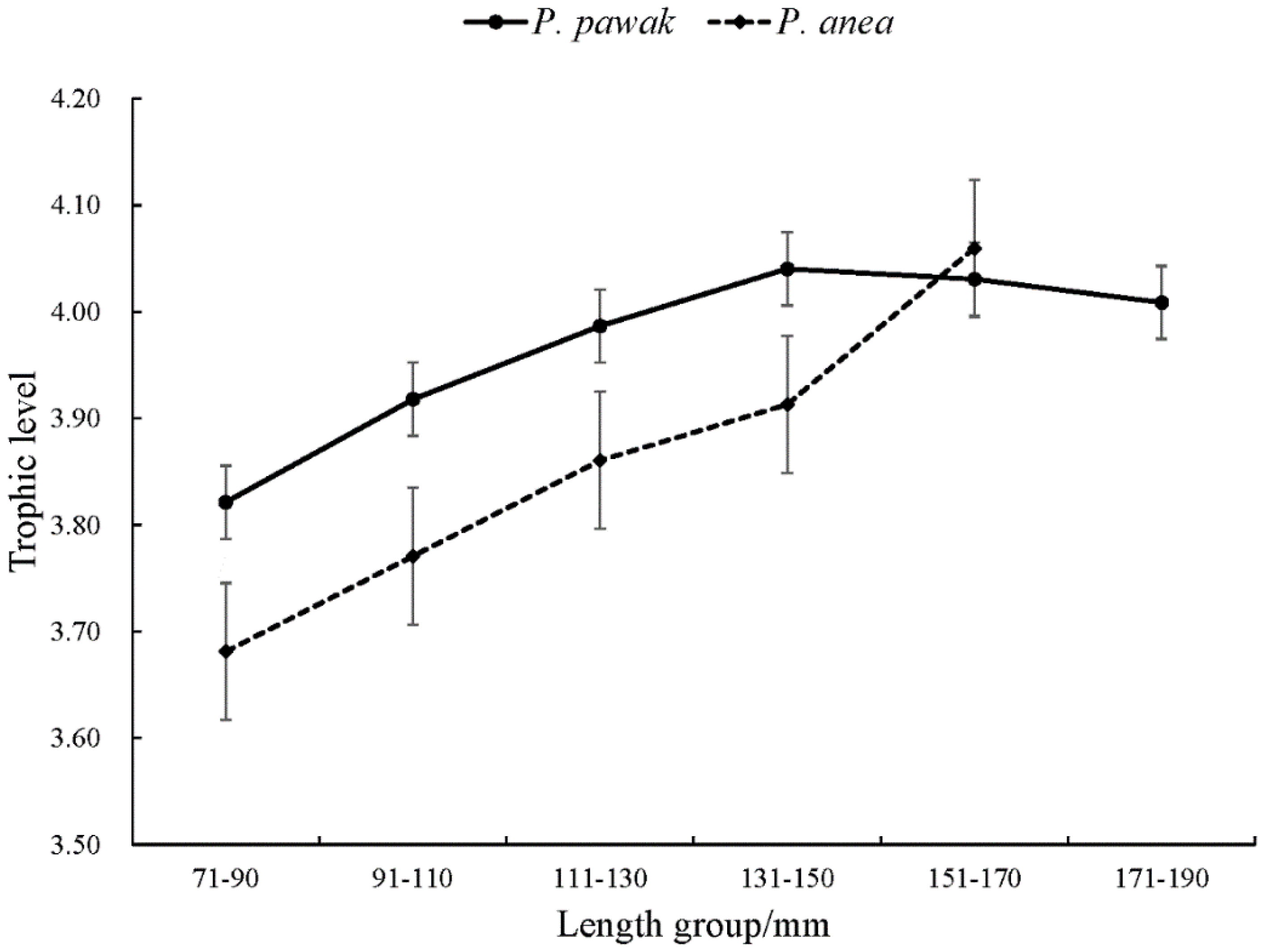
The trophic levels of P. pawak and P. anea are different to some extent. The trophic levels of P. pawak ranged from 3.70 to 4.21 based on the carbon and nitrogen stable isotope ratios, with an average of 3.97 and a trophic span of 0.51. The trophic levels of the P. anea ranged from 3.30 to 4.28, with an average of 3.85 and a trophic span of 0.7. The trophic level of the P. pawak showed a rising trend with the growth of the length, with a decreasing trend from the 131-150 mm length group. The trophic level of P. anea gradually increased with body length. The trophic span of P. pawak is smaller than that of P. anea. With the increase of length group, the trophic level of P. pawak increased continuously, while that of P. anea decreased when it increased to a certain extent.
Figure 5.
Stable isotopic distribution of δ13C and δ15N in P. pawak and P. anea.
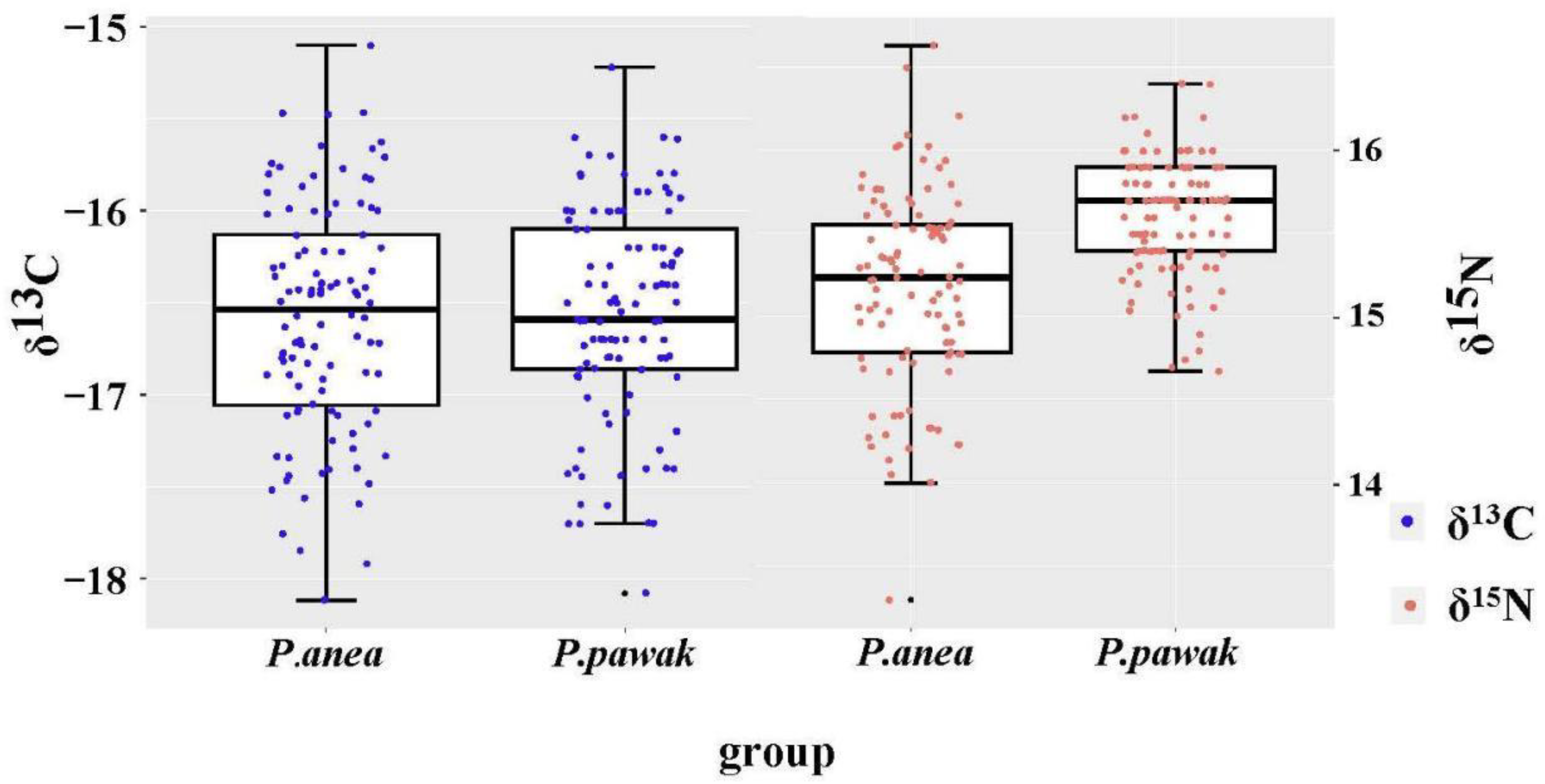
Figure 6.
Change of trophic levels of P. pawak and P. anea in different length.
3.3. Trophic niche
There are certain differences in the trophic niche between the P. pawak and P. anea, but the overlap is not obvious. Based on the carbon and nitrogen stable isotope analysis of the trophic niche constructed by P. pawak and P. anea, the trophic niche indexes of the two species of Pennahia genus showed that (Table 4), the level of centroid distance (CD) of P. pawak was 0.59, carbon range (CR) was 2.86, nitrogen range (NR) was 1.72, the overall density (MNND) was 0.09, the distribution range of trophic niche (SDNND) was 0.09, and the core ecological niche (SEAc) was 0.09, and total area of niche (TA) was 3.70. The level of trophic diversity (CD) of P. anea was 0.76, carbon range (CR) was 3.02, nitrogen range (NR) was 3.32, overall density (MNND) was 0.13, distribution range (SDNND) was 0.15, core niche (SEAc) was 1.16, and total niche area (TA) was 6.09. It can be seen that the basal food source, food chain length, level of trophic diversity, overall density, range of trophic niches, space of core niches, and total niche area of P. anea were larger than those of P. pawak. From the trophic niche constructed by carbon and nitrogen stable isotopes (Figure 6), it can be seen that the trophic niche of P. pawak and P. anea overlapped to some extent, but the degree of overlap was only 0.29, which was not significant.
Figure 7.
Trophic niche of two Pennahia species.
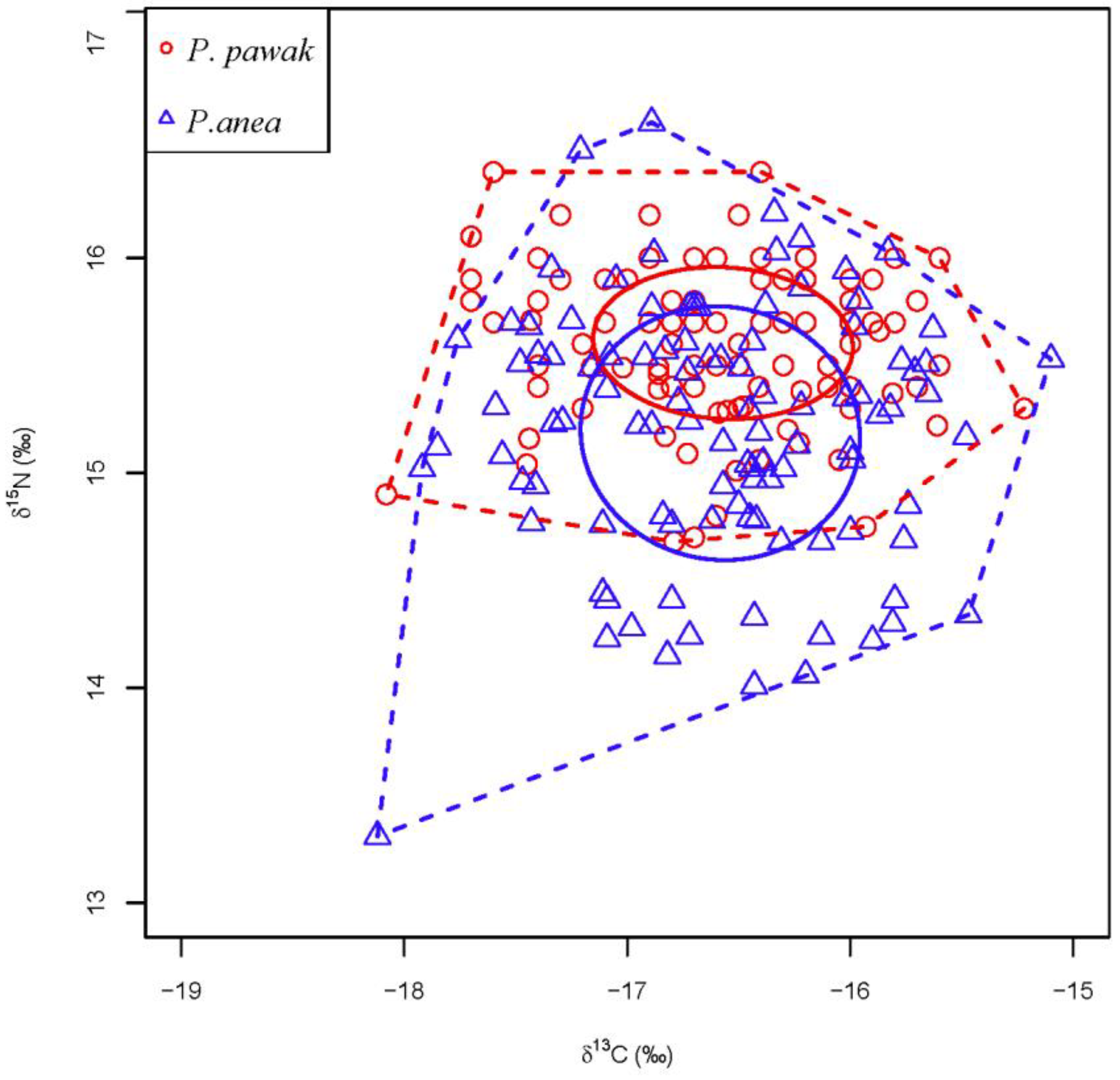
Table 4.
Trophic niche indicators in two Pennahia species.
| Species | CD | CR | NR | MNND | SDNND | SEAc | TA | Overlap index |
|---|---|---|---|---|---|---|---|---|
| P. pawak | 0.59 | 2.86 | 1.72 | 0.09 | 0.09 | 0.65 | 3.70 | 0.29 |
| P. anea | 0.76 | 3.02 | 3.32 | 0.13 | 0.15 | 1.16 | 6.09 |
3.4. Spatial niche
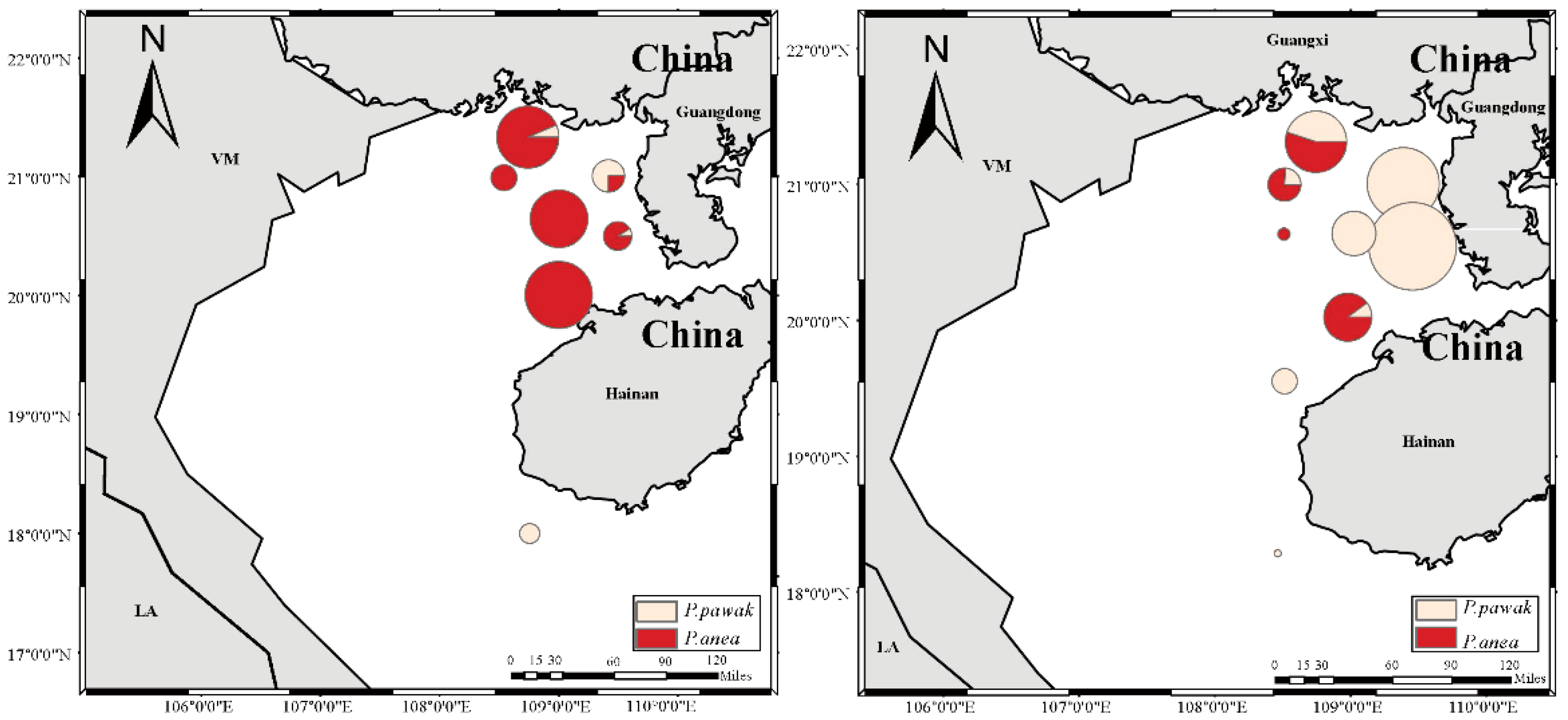
The resources of the Beibu Gulf of P. pawak and P. anea have obvious spatial distribution characteristics, and both of them are mainly distributed in the inner waters of the Beibu Gulf, while the P. pawak is also distributed in the middle and the mouth of the Gulf. P. pawak is distributed in the area of 108.46°~109.50°E, 18.00°~21.33°N, while P. anea is distributed in the area of 108.51°~109.50°E, 20.00°~21.33°N. Resource densities of the two species of Pennahia genus from 0.08 to 109.52 kg/m2 and 0.28 to 29.65 kg/m2, respectively, with the average resource density of P. pawak (21.19 kg/m2) being greater than that of P. anea (8.74 kg/m2). The spatial niche widths of P. pawak and P. anea differs before and after summer close season with P. anea (0.85) having a greater niche width than P. pawak (0.10) before summer close season, while P. pawak by (1.22) had a greater spatial niche width than P. anea (0.50) after summer close season. The spatial niche overlap index of the two Pennahia species was 0.20, which was present but not significant. Neither of the two Pennahia species reached a meaningful level of overlap before (0.16) or after summer close season (0.13).
Table 5.
Spatial niche width and overlap index.
| Total | Before SCS | After SCS | ||
|---|---|---|---|---|
| Spatial niche width | P. pawak | 1.19 | 0.10 | 1.22 |
| P. anea | 0.58 | 0.85 | 0.50 | |
| Overlap index | 0.20 | 0.16 | 0.13 |
Note: SCS indicates summer close season.
Figure 8.
Resource Density Distribution of P. pawak and P. anea. Note: On the left is before SCS and on the right is after SCS.
Figure 8.
Resource Density Distribution of P. pawak and P. anea. Note: On the left is before SCS and on the right is after SCS.
4. Discussion
The feeding ecology of marine fishes is an important part of marine food web research, through which we can understand the feeding habits of fishes and the food relationships among different fishes in marine ecosystems [16,40]. Food competition is one of the important manifestations of food relationships, including intraspecific and interspecific food competition. Intraspecific food competition is generally defined as competition between individuals of the same species for the same food resource, which is intense and largely unavoidable. In the short term, competing individuals may adopt different behavioral strategies and feeding patterns, or establish dominance hierarchies and feeding areas; in the long term, intraspecific food competition can drive ecological role turnover [41]. Studying the similarity in the composition of prey organisms consumed by different organisms (overlapping feeding niches) can help to understand food competition between species; the higher the degree of overlap in food between fish species, which leads to competition for food; on the contrary, if there is little food overlap between fish species, it means that the greater the differences in prey, the less competition between species. When the prey organisms in the environment are limited, fish with similar niches will have severe food competition, which may lead to turnover of dominant species and changes in community structure [42]. The study of interspecific food relationships among fishes of the same genus in the same sea area can reflect the status of fish utilization of food resources and the existence of competition in the feeding process, and is the basis for studying the trophic dynamics of food webs [43].
In this study, potential intraspecific competition was found to exist in both P. pawak and P. anea, which was mainly characterized by significant food overlap between individual length groups. Optimal Foraging Theory suggests that individual feeding differences depend on the phenotype (size, sex, or developmental stage) and prey availability of the individual [4]. More precisely, although individuals may consume a wide variety of prey, they adopt more specialized feeding habits to avoid intraspecific competition, depending on the state of the individual and the bait resources available in the habitat. At low population densities, individuals compete for the same preferred prey, but as population densities increase, this prey resource becomes scarce and individuals begin to feed on other, different prey, suggesting that increasing population densities lead to increased intraspecific competition, which in turn increases the degree of feeding specialization [44]. Xia et al. [45] used stomach content analysis and stable isotopes analysis to study the feeding habits of Megalobrama terminalis, and the results showed that the decrease in intraspecific competition was mainly due to individual feeding specialization. In this study, the degree of feeding specialization in both P. pawak and P. anea at the stage of 71-190 body length showed a decrease with the growth of body length, indicating that the intensity of intraspecific competition decreases with the growth of individuals during this stage.
The δ13C value is less variable during food chain transmission and can indicate its food source, while the δ15N value is relatively enriched in the organism and is generally used to determine the trophic level of the study object [34]. It has been shown that the carbon and nitrogen stable isotope compositions of fish are not constant at different growth stages [45]. In this study, the carbon stable isotope values of the two species did not vary much with body length, which may be related to the small enrichment of carbon stable isotopes in the living body on the one hand [46], and on the other hand, although there was a shift in feeding, the main feeding taxa, especially the final trophic sources (elements), were relatively stable. In addition, nitrogen stable isotopes and trophic levels showed an overall increasing trend with body length, which may be related to the increased feeding capacity of individuals. This pattern of change has been observed in many fish species [47,48], with the growth of individuals, the feeding and digestive organs of fish continue to improve, the swimming ability continues to increase, the range of prey selection is wider, and the prey of fish is shifted from a low trophic level to a higher trophic level. δ13C and δ15N values also reflect the niche change of organisms, and the niches of two species showed an expanding trend with the growth of individual development. The fish mainly fed on Pisces and Macrura during the small individual stage, but intraspecific competition prompted individuals to occupy a wider niche by feeding more on Brachyura, Cephalopoda, Gastropoda and other prey during growth. In conclusion, at the intraspecific level, intraspecific competition exists in both P. pawak and P. anea, but they avoid intense intraspecific competitive relationships by shifting their feeding to promote the expansion of the population niche.
Niche width represents the utilization of all available resources by a species to reduce the degree of intraspecific individual competition [49], and niche overlap represents the frequency of encounters between two species on the same spatial resource, which can also reflect the potential competitive relationship between species [50]. In this study, the niche overlap indexes (Oij = 0.56 and Bij = 0.29) for both P. pawak and P. anea did not reach the significant level. Although niche overlap indicates potential interspecific competition, its intensity also depends on consumer and resource abundance [51]. Strong interspecific competition causes species to reduce feeding on conspecific prey, thereby mitigating interactions with other species [52]. In the present study, it was found that although Macrura organisms were the main prey for two Pennahia species, P. anea predominantly consumed Alpheus. While P. pawak consumed a percentage of Metapenaeopsis barbata and Alpheus bisincisus in addition to Alpheus. This suggests that P. pawak and P. anea, when they have the same prey base, show different feeding preferences by enhancing their feeding on different types of prey. This is consistent with Evolutionarily Stable Strategy, in which closely related species inhabiting the same sea area will coexist by regulating their own feeding habits to attenuate food competition between species, thus maximizing equilibrium [12]. Based on carbon and nitrogen stable isotope analysis, the difference in δ13C variation between P. pawak and P. anea was not significant, but there was no significant niche overlap between the two species, and the total niche width (TA) and the core niche (SEAc) of P. anea were larger than that of P. pawak. Therefore, combined with the analysis of food composition, although the nutrient sources of the two species are basically the same, the niche overlap of the two species is not significant because of their different feeding preferences. This reflects that there is some interspecific competition between P. pawak and P. anea, but the two species avoid intense competition through allocation of prey resources and niche differentiation.
The width of the spatial niche reflects the spatial distribution of the species and its ability to utilize spatial resources [53]. The spatial niche width of P. pawak increased while that of P. anea decreased after the summer close season. P. pawak spawns in April-August [24], and the summer close season (May 1) facilitates the protection of the spawning population and juveniles, thus increasing its resource density; while P. anea spawns in March-June [22], which is two months earlier than the summer close season, and the protection of the spawning population and juveniles is insufficient, and the density of its resource is on a decreasing trend. Differences in the width of spatial niche is one of the important conditions for species to realize coexistence [53], and this change has a certain mitigating effect on the pressure of spatial resource competition between the two. The spatial niche overlap index reflects the similarity of species in spatial distribution [54]. The overlap between P. pawak and P. anea did not reach a significant level, indicating that the existence of competition between the two is not intense. Trophic niche analysis showed that the base food source (CR), food chain length (NR), level of trophic diversity (CD), overall density (MNND), trophic niche range (SDNND), core niche area (SEAc) and total niche area (TA) of P. pawak were larger than those of the P. anea, which suggests that when the two Pennahia species coexist in the same domain, P. pawak has a greater ability to utilize the resources than P. anea. Therefore, when the two species coexist in the same area, P. pawak will migrate to the area with fewer P. anea in order to reduce the competitive disadvantage. In terms of the spatial distribution of resources, there was some spatial overlap in the distribution of the two species during the same season, but there were also obvious differences in distribution, indicating that spatial heterogeneity in the distribution of resources is an important factor in mitigating the interspecific competition between the two Pennahia species.
Intraspecific competition promotes individual niche diversification within species niche, but interspecific competition prevents further species niche expansion and limits individual niche variation [55]. Intraspecific competition is intense between the P. pawak and P. anea, but both species mitigate intraspecific competition by reducing the degree of individual feeding specialization, promoting trophic niche expansion. On the other hand, interspecific competition existed between the two Pennahia species, but there were some differences in their preferred feeding prey, which reduced interspecific interactions, and they also mitigated interspecific interactions through trophic niche differentiation. Classical ecological theory suggests that the niche occupied by a species is limited by a variety of ecological factors, among which biological factors include its own population size, feeding and being fed on, and food competition, and that an increase in population size leads to increased competition [56]. Organisms with similar ecological status can reduce competition for limited food resources through dietary differentiation or selection of different habitat spaces, and dominant species will preferentially select prey organisms or choose other habitat spaces to reduce interspecific competition for food. In marine ecosystems, marine organisms inhabiting the same sea area mainly reduce interspecific competition by realizing the differentiation of trophic, spatial, and temporal niches, among which the differentiation of trophic and spatial ecological niches is particularly important [57]. From the present study, two species of Pennahia genus inhabiting the Beibu Gulf have similar ecological habits and competitive relationships, but their specificity in trophic and spatial niches reduces interspecific competition and achieves coexistence; in addition, the trophic and spatial niches of P. pawak are more advantageous than those of P. anea, and it has a stronger ability to utilize resources and ecological adaptability.
5. Conclusions
The results of this study indicated that there was intense intraspecific competition between P. pawak and P. anea, while the interspecific competition was not significant. Intraspecific competition was mitigated by feeding shifts and interspecific competition was reduced by trophic and spatial niche differentiation. This result elucidates the coexistence mechanism of the two species, which have similar morphology and ecological habits, and provides basic data for food web studies at the trophodynamic level in Beibu Gulf. In addition, abiotic factors such as water temperature, salinity, currents and light can affect the distribution of prey organisms and the feeding habits of fish, and thus have a certain impact on the feeding habits of fish. In the future, we can combine the abiotic factors to study the relationship between the feeding habits of fish and abiotic factors, so as to analyze the feeding ecology of fish in a more comprehensive way.
Author Contributions
Conceptualization, X.H., Y.Y.; formal analysis, K.L., Y.Z.; investigation, K.L., X.Y., J.W.; data curation, K.L., X.Y., C.Z.; writing-original draft preparation, K.L., X.H.; writing—review and editing, K.L., X.H. and Y.Y.; supervision, K.L., X.H. and Y.Y. All authors have read and agreed to the published version of the manuscript.
Funding
National Natural Science Foundation of China (U20A2087), The start-up funds of Guangdong Ocean University (No. 060302022104).
Institutional Review Board Statement
Not applicable. All specimens were collected from commercial fishery catches, and data were obtained from dead fishes that were not endangered or protected. The experimental protocols concerning handling of fishes were in accordance with the requirements of the Institutional Animal Care and Use Committee of Guangdong Ocean University, China.
Informed Consent Statement
Not applicable.
Data Availability Statement
To obtain data from this study, please contact author Konglan Luo (email: luokonlan@gmail.com).
Acknowledgments
The authors would like to thank Fengming Liu, Xiao Zhou, Kangwen Qiu, Sibiao Liu, Yujian Deng, and other people who assisted with the sampling and experiments.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Liu, J. Advanced aquatic biology; Science Press: Beijing, China, 1999.
- Brönmark, C., Hansson, L. The biology of lakes and ponds; Oxford University Press: New York, USA, 2005.
- Tang, Q. Strategies of research on marine food web and trophodynamics between high trophic levels. Progress in Fishery Sciences 1999, 2: 1-6.
- Schoener, T. Resource partitioning in ecological communities: research on how similar species divide resources helpsreveal the natural regulation of species diversity. Science 1974, 185: 27-39.
- Wang, J., Su, Y., Liu, J., et al. The feeding habits of five sciaenids fishes in Louyuan Bay. Journal of Jimei University (Natural Science) 1994, 2: 34-39.
- Li, Z., Dai, Q., Zuo, T., et al. Studies on food competition between Pseudosciaena polyactis and Johnius belengerii from changjian estuary and adjacent southern yellow sea in autumn. Journal of Hydroecology 2009, 30: 67-72.
- Matley, J., Heupel, M., Fisk, A., et al. Measuring niche overlap between co-occurring plectropomus spp. Using acoustic telemetry and stable isotopes. Mar Freshwater Res 2017, 68: 1468. [CrossRef]
- Wang, Q., Liu, M., Zhu, F., et al. Comparative study of three species of schizothoracine on feeding and digestive organs in Upper Nujiang River. Chinese Journal of Zoology 2019, 54: 207-221.
- Zhang, Y., Xu, B., Zhang, C., et al. Spatial heterogeneity in the feeding habits and feeding ground distribution of johnius belangerii in Haizhou Bay during spring. Journal of Fishery Sciences of China 2020, 27: 315-326.
- Olesen, J. M., Bascompte, J., Dupont, Y. L., et al. The modularity of pollination networks. P Natl Acad Sci USA 2007, 104: 19891-19896. [CrossRef]
- Raul, C. P., MaRcio, S. A., Souza, F. L., et al. Competition and resource breadth shape niche variation and overlap in multiple trophic dimensions. Proc Biol Sci 2019, 1902: 20190369.
- Li, Y., Wang, H., Chen, X., et al. Trophic niches and interspecific relationships between closely related ommastrephidae species, dosidicus gigas and sthenoteuthis oualaniensis. Acta Ecologica Sinica 2020, 40: 5418-5423.
- Xiao, Y., Jiang, R., Yin, R., et al. Trophic niche and interspecific relationship of five eels in the waters of the zhoushan islands. Journal of Fisheries of China 2023, 47: 65-74.
- Yang, Z., Chen, X., Zhao, N., et al. The effect of different habitat types and ontogenetic stages on the diet shift of a critically endangered fish species, coreius guichenoti (sauvage and dabry de thiersant, 1874). Int J Environ Res Public Health 2018, 15: 2204. [CrossRef]
- Hyslop, E. J. Stomach contents analysis-a review of methods and their application. J Fish Biol 1980, 17: 411-429. [CrossRef]
- Xue, Y., Jin, X. Review of the study on feeding habits of fishes and food webs. Progress in Fishery Sciences 2003, 2: 76-87.
- Dixon, H., Dempson, J., Sheehan, T., et al. Assessing the diet of North American Atlantic salmon (Salmo salar l.) off the West Greenland coast using gut content and stable isotope analyses. Fisheries Oceanography 2017, 26: 555-568. [CrossRef]
- Wang, X., Qiu, Y., Du, F., et al. Dynamics of demersal fish species diversity and biomass of dominant species in autumn in the Beibu Gulf, northwestern South Shina Sea. Acta Ecologica Sinica 2012, 32: 333-342.
- Yi, X., Qiu, K., Zhou, X., et al. Analysis of fishery biology of pennahia pawak in Beibu Gulf. Journal of Shanghai Ocean University 2021, 30: 515-524.
- Wang, X., Du, F., Qiu, Y. Length-weight relationships of important commercial fishes in Northern South China Sea. Journal of Applied Oceanography 2006, 2: 262-266.
- Wagiyo, K., Tirtadanu, Chodriyah, U. Biology characteristic, abundance index and fishing aspect of donkey croaker (pennahia anea bloch,1793) in the Tangerang Waters. E3S Web of Conferences 2020, 4: 1011.
- Tuuli, C., de Mitcheson, Y., Liu, M. Reproductive biology of the greyfin croaker pennahia anea in the northern South China Sea. Ichthyol Res 2011, 58: 302-309. [CrossRef]
- He, X., Tao, Y., Hou, G., et al. Population structure and spatio-temporal distribution of pennahia pawak in the Beibu Gulf, South China Sea. Journal of Guangdong Ocean University 2015, 35: 35-42.
- Yan, Y., Hou, G., Lu, H., et al. Age and growth of pawak croaker Pennahia pawak in Beibu Gulf. Journal of Fishery Sciences of China 2011, 18: 145-155. [CrossRef]
- Yi, X. Feeding ecology of pawak croaker (pennahia pawak) in the Beibu Gulf. Master Thesis, Guangdong Ocean University, Zhanjiang, China, 2021.
- Third Institute of Oceanography State Oceanic Administration. Specifications for oceanographic survey—part 6: marine biological survey; State General Administration of the People"s Republic of China for Quality Supervision and Inspection and Quarantine; Standardization Administration of the People'sRepublic of China: Beijing, China, 2007.
- Figueiredo, M., Morato, T., Barreiros, J. P., et al. Feeding ecology of the white seabream, Diplodus sargus, and the ballan wrasse, Labrus bergylta, in the azores. Fish Res 2005, 75: 107-119. [CrossRef]
- Dou, S. Fish-stomach content analysis: methods and application. Marine Science Bulletin 1992, 2: 28-31.
- Pianka, E. The structure of lizard communities. Annu Rev Ecol Evol S 1973, 4: 53-74. [CrossRef]
- Shannon, C. The mathematical theory of communication. 1963. Md Comput 1997, 14: 306-317.
- Roughgarden, J. Evolution of niche width. The American Naturalist 1972, 106: 683-718.
- Bolnick, D. I., Yang, L. H., Fordyce, J. A., et al. Measuring individual-level resource specialization. Ecology 2002, 83: 2936-2941.
- McKinney, C., McCrea, J., Epstein, S., et al. Improvements in mass spectrometers for the measurement of small differences in isotope abundance ratios. Rev Sci Instrum 1950, 21: 724-730. [CrossRef]
- Post, D. Using stable isotopes to estimate trophic position: models, methods, and assumptions. Ecology (Durham), 2002, 83: 703-718.
- He, X., Zhu, D., Zhao, C., et al. Feeding habit of asian moon scallop (amusium pleuronectes) and as an isotopic baseline indicator in the Beibu Gulf, South China Sea. J Shellfish Res 2019, 38: 245-252. [CrossRef]
- Jackson, A., Inger, R., Parnell, A., et al. Comparing isotopic niche widths among and within communities: siber - stable isotope bayesian ellipses in R. J Anim Ecol 2011, 80: 595-602. [CrossRef]
- Schoener, T. W. Sizes of feeding territories among birds. Ecology 1968, 49: 123-141. [CrossRef]
- Zhan, B. Fishery resources assessment; China Agriculture Press: Beijing, China, 1995, pp 26-27.
- Krebs, C. Ecological methodology; Harper Collins Publishers: New York, USA, 1989.
- Layman, C., Post, D. Can stable isotope ratios provide for community-wide measures of trophic structure? Reply. Ecology 2008, 89: 2358-2359. [CrossRef]
- Ward, A., Webster, M., Hart, P. Intraspecific food competition in fishes. Fish Fish, 2006,7(4):231-261. [CrossRef]
- Deng, J., Jiang, W., Yang, J., et al. Species interaction and food web of major predatory species in the Bohai Sea. Journal of Fishery Sciences of China 1997, 4: 2-8.
- Tang, Q., Fan, Y., Lin, H. Initial inquiring into the developmental strategy of chinese ocean ecosystem dynamics research. Advances in Earth Science 1996, 2: 160-168.
- Svanbäck, R., Bolnick, D. I. Intraspecific competition affects the strength of individual specialization: an optimal diet theory method. Evol Ecol Res 2005, 7: 993-1012.
- Xia, Y., Li, Y., Zhu, S., et al. Individual dietary specialization reduces intraspecific competition, rather than feeding activity, in black amur bream (megalobrama terminalis). Sci Rep 2020, 10: 17961. [CrossRef]
- Zhang, B., Yuan, W., Dai, F. Study on feeding ecology of fish community in Laoshan Bay during summer using stable carbon and nitrogen isotopes. Journal of Fisheries of China 2016, 40: 585-594.
- Ji, W., Chen, X., Jiang, Y., et al. Stable isotope analysis of some representative nektonic organisms in the central and northern part of East China Sea. Marine Fisheries 2011, 33: 241-250.
- Gao, Y., Sui, H., Ren, X., et al. Feeding habits of saurida elongata in Haizhou Bay, Shandong, China, based on stomach contents and stable isotope. Chinese Journal of Applied Ecology 2020, 31: 4277-4283.
- Wang, F., Ju, R., Li, Z., et al. Niche concept and its application in insect ecology. Chinese Journal of Ecology 2006, 10: 1280-1284.
- Yang, X., Ma, J. A review on some terms related to niche and their measurements. Chinese Journal of Ecology 1992, 2: 46-51.
- Hurlbert, S. H. The measurement of niche overlap and some relatives. Ecology 1978, 1: 66-77. [CrossRef]
- Abrams, P. Some comments on measuring niche overlap. Ecology 1980, 61: 44-49. [CrossRef]
- Yu, Z., Jin, X., Li, X. Analysis of ecological niche for major fish species in the central and southern Yellow Sea. Progress in Fishery Sciences 2010, 31: 1-8.
- Han, D., Xue, Y., Ji, Y., et al. Trophic and spatial niche of five gobiid fishes in Jiaozhou Bay. Journal of Fishery Sciences of China 2013, 20: 148-156. [CrossRef]
- Cachera, M., Ernande, B., Villanueva, M. C., et al. Individual diet variation in a marine fish assemblage: optimal foraging theory, niche variation hypothesis and functional identity. J Sea Res 2017, 120: 60-71. [CrossRef]
- Svanback, R., Bolnick, D. I. Intraspecific competition drives increased resource use diversity within a natural population. Proc Biol Sci 2007, 274: 839-844. [CrossRef]
- Ross, S. Resource partitioning in fish assemblages: a review of field studies. Copeia 1986, 2: 352-388. [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



