
Submitted:
08 February 2024
Posted:
09 February 2024
You are already at the latest version
Abstract
Biotechnological potential of Thermoanaerobacter strain AK15 to produce ethanol and valuable alcohols from carbohydrates and amino acids were evaluated in present study. The strain is highly ethanologenic, producing a maximum of 1.57 mol ethanol per mol of glucose degraded at high liquid-gas phase ratios. The strain degrades most of the sugars tested as well as starch and pretreated hydrolysates of biopolymers (Whatman paper, newspaper, Timothy grass, Rhubarb leaves, and several macroalgae species) to ethanol mainly. The strains degraded serine and threonine when used as a single substrate, producing mainly acetate and ethanol as end products, and the branched-chain amino acids (leucine, isoleucine, valine) when cultivated in the presence of thiosulfate. The main end products from branched-chain amino acids were a mixture of their corresponding branched-chain fatty acids and alcohols. Finally, the strain was also shown to use butyrate as an electron sink during glucose degradation resulting in the reduced product butanol in addition to end products produced from glucose. Thus, strain AK15 is a promising candidate for ethanol and fine chemical production from a range of lignocellulosic and algal biomass.
Keywords:
Thermoanaerobacter carbohydrate
; protein
; sugar
; amino acid
; end products
1. Introduction
Production of both biofuels and fine chemicals from renewable sources is in more demand than ever before. The urge to replace fossil fuels with renewable alternatives is becoming more and more urgent because of the global climate changes on Earth as well as the geopolitical uncertainties associated with major oil exporting nations. As such, the United Nations Sustainable Development Goals place a great emphasis on climate action and sustainable development.
While there are several viable options for biofuels that can meet the needs of a liquid energy carrier to replace gasoline, bioethanol, and other alcohols are promising due to their ease of production, low toxicity, direct compatibility of blended fuels with many existing combustion engines, and high volatility [1]. Furthermore, bioethanol production has enjoyed a long history but its production as a biofuel at industrial scales has been primarily limited to using biomass containing starch and sucrose as a substrate which poses several problems, namely the environmental impact and negligible energy yield from cultivating these crops and increasing the price of food [2,3]. To this end, there has been a lot of interest in using other biomasses that do not compete with food utilization such as lignocellulosic and seaweed biomass. Lignocellulosic biomass is composed of cellulose, hemicellulose, and lignin [4] while seaweeds have a much more diverse composition of carbohydrates which often includes cellulose but can also include β-glucans such as laminarin, uronic-acid containing carbohydrates such as carrageenan, alginate, agarose, mannitol, and sulfated polysaccharides [5,6]. As with terrestrial biomass, the exact composition varies as a function of the type of seaweed in addition to other variables related to growth conditions. Both lignocellulosic and algal biomass present a challenge to the production of bioethanol given the greater diversity of carbohydrate building blocks present making finding an organism for bioprocessing more challenging.
The production of bioethanol from simple biomass has also led to the debate on the utilization of food and feed for fuel production. Therefore, the utilization of more complex biomass is a better alternative, but its use is more complex because of the complex structure and the pretreatment needed for their degradation to hexoses and pentoses. Additionally, the most common microorganism used for bioethanol production has a limited capacity of degrading the wide variety of sugars present in lignocellulosic biomass. This has led to a search for more feasible microorganisms with a broader substrate range than yeasts. To this end, thermophilic anaerobes have been promising candidates for this purpose and many studies in the past twenty years have focused on hydrogen and ethanol production from complex biomass [2,3,7,8]. Additionally, several species belonging to the genera of Thermococcus, Pyrococcus, Caldicellulosiruptor, and Thermotoga have been genetically manipulated to increase hydrogen and/or ethanol production [9,10,11,12]. Production of fine chemicals, like 1,2-propanediol, 1,3-propanediol, and branched-chain alcohols have historically been produced from non-renewable sources, like gasoline and gases [13]. More environmental routes to such chemicals are thus more feasible and have led to the investigation of the production of various fine chemicals [14]. Several thermophilic bacteria have been used for the production of various fine chemicals, e.g. Thermoanaerobacterium and Caldicellulosiruptor for 1,2 propanediol [15,16], Caloramator for 1,3 propanediol [17], and Thermoanaerobacter and Caldanaerobacter for production of branched-chain alcohols [18] Thermophiles have also been described as emerging metabolic platform organisms for the production of various industrial products apart from ethanol and hydrogen, e.g. for phytoene, acetoin, 3-hydroxypropionate, and butanol [19].
All species within Thermoanaerobacter are obligate anaerobes; ferment various carbohydrates to ethanol, acetate, lactate, hydrogen, and carbon dioxide [20] and originate from various habitats like hot springs, hydrothermal vents, and oil fields [21,22,23,24]. Species within the genus Thermoanaerobacter have received considerable attention due to their biotechnological potential, having a very broad substrate spectrum, especially among the sugars present in lignocellulosic biomass, and high ethanol yields from lignocellulosic biomasses [25,26]. Many Thermoanaerobacter species have been investigated for their ethanol production e.g. T. ethanolicus, T. mathranii, and T. pseudethanolicus [7,8,27]. Recent investigations have also shown the ability of some species within the genus to produce valuable high-carbon alcohols from specific amino acids [18] as well as being able to reduce fatty acids to their corresponding alcohol during growth on both sugars and amino acids [28]. It is of interest that the end products formed during fermentation can be manipulated by controlling environmental factors. Thus, a very good ethanol producing strain under specific conditions can be cultivated under conditions that may lead to that the main end product of the strain being acetate and hydrogen. This has been shown for Thermoanaerobacter strain AK5 where the ethanol and production varied from being 0.7 mol ethanol/mol glucose to 1.7 mol ethanol/mol glucose, simply by cultivating the strain at different liquid to gas phase ratios [29]. Similarly, the acetate produced changed from 0.14 to 0.74 mol acetate/mol glucose. This change was even more dramatic when the strain was cultivated in the presence of thiosulfate or in the presence of a hydrogenotrophic methanogen. Under these conditions, ethanol became only a minor end product but acetate, by far, was the major volatile end product. Due to the broad substrate range of Thermoanaerobacter, they have often been linked to biofuel production from complex biomass (lignocellulose). Several reports show that Thermoanaerobacter species produce between 3 to 6 mM ethanol g-1 dw of various biomass types, such as grass, hemp, barley straw, corn straw, and so forth [3 and references therein].
The present investigation is on both the carbohydrate and amino acid metabolism of Thermoanaerobacter strain AK15 as well as their potential to utilize complex biomass and macroalgae hydrolysates. Thermoanaerobacter strain AK15 is a strictly anaerobic bacterium and was isolated from an alkaline hot spring (pH 8.6, Temperature 60°C) in Viti in the Krafla area in NE-Iceland as previously described by Scully and co-workers [18].
2. Results and Discussion
2.1. Substrate Spectra
Thermoanaerobacter AK15 was cultivated on the main carbohydrates present in lignocellulosic and algal biomass, including hexoses (glucose, mannose, galactose), pentoses (arabinose, xylose), disaccharides (cellobiose), as well as several deoxysugars (L-fucose and L-rhamnose), and mannitol which is found in seaweeds. Additionally, strain AK15 was tested for growth on polymeric carbohydrate substances, all 20 of the proteogenic amino acids with and without the inclusion of thiosulfate (20 mM), and various proteins with and without thiosulfate (20 mM).
2.1.1. Degradation of Carbohydrates
Thermoanaerobacter AK15 degraded cellobiose and all tested monosaccharides except for the two deoxysugars, L-fucose and L-rhamnose (Figure 1). The main end product in all substrates that were degraded was ethanol (Figure 1) which resulted in optical densities between 0.35 to 0.48 and a decrease of pH for about 1.0 pH unit compared with the control (Supplementary Table S1). Some thermophiles have been shown to degrade deoxysugars like Caldicellulosiruptor species of which more than half of the type strains within the genus degraded L-rhamnose to 1,2-propanediol and three strains degraded L-fucose to 1,2-propanediol [16]. Other thermophilic anaerobes known to produce 1,2-propanediol are moderately thermophilic strains of Clostridium [30,31].
The strain degraded mannitol to ethanol mainly, as has also been shown for T. uzonensis, the closest relative to strain AK15 [32]. Mannitol utilization was not tested in the original characterization study of Wagner and coworkers describing T. uzonensis [33].
The original data on ethanol production from glucose for T. uzonensis showed 1.15 mol glucose per mol glucose degraded [33] while Thermoanaerobacter strain AK15 produces 1.10 ethanol mol glucose−1 (Figure 1). Other well-known species producing high amounts of ethanol from sugars within the genus are i.e. T. ethanolicus and T. pseudethanolicus [8,27]. The other end products are mainly acetate with traces of lactate; this was also the case for other carbohydrates tested. It is a well-known phenomenon that the ratio of end products is highly influenced by culture conditions, particularly hydrogen accumulation. Therefore, a simple experiment was performed on the strain by cultivating it in three different liquid–gas phase ratios: 0.09, 1.00, and 5.62. As shown in Figure 2, the end product profile changes dramatically during growth under different liquid-gas phase ratios for the strain.
At high pH2, the main end product is ethanol, or 33.3 mM which corresponds to 1.57 mol/mol glucose degraded (controls subtracted) but drops to 0.44 mM at the lowest pH2. Similarly, acetate and hydrogen decrease at high pH2 conditions. The higher acetate concentrations at low liquid – gas phase rations resulted in lower pH, but the optical density was similar in all conditions (Supplementary Table S2). This highlights the importance of the culture conditions on end-product formation and has been reported for various thermophilic bacteria in recent studies [29,34]. Thus, the strain is highly ethanologenic and can be manipulated to convert more than 75% of the glucose to ethanol as has been reported for several other Thermoanaerobacter strains [35]. Also, being versatile concerning the utilization of a wide variety of carbohydrates, strain AK15 may be considered as a good candidate for bioethanol production from lignocellulosic biomass. To investigate this further the strains were cultivated on several lignocellulosic and algal biomass pretreated with dilute acid and cellulases.
2.1.2. Degradation of Polymeric Carbohydrate Substrates
The ability of microorganisms to be good candidates for bioprocessing of lignocellulosic or algal biomass should include the capability of degrading a wide range of glycosidic bonds. The ability of strain AK15 to degrade β-1,3-O glycosidic bonds were evaluated using lichenan and laminarin. The ability of strains to utilize the hemicellulose fraction was evaluated using xylan, mannan (β-O-(1-4) linked mannose), galactan, and rhamnan. The strain did not utilize any of these substrates. Of the polymeric carbohydrates tested, strain AK15 only showed end product formation from starch (Figure 3). T. uzonensis is however reported as starch negative [33] although many of members of the genus degrade starch as the sole carbon source. The main end product from starch was ethanol (12.3 mM). No growth was observed when the strain was cultivated on cellulose, as previously reported for Thermoanaerobacter species [20] nor did the strain degrade laminarin, chitosan, lichenan, keratin, mannan, galactan, or rhamnan. This is also confirmed by no increase in optical densities or lowering of pH in the cultures (Supplementary Table S3). There are only several species within the genus that are known to degrade keratin with the best-known being “Thermoanaerobacter keratinophilus” [36], although this strain likely falls within the modern genus of Caldanaerobacter.
2.1.3. Degradation of Biomass Hydrolysates
The strain was tested on several complex biomass types, both lignocellulosic biomass (grass, newspaper, rhubarb leaves) and macroalgae types (Ascophyllum nodusum, Palmaria palmata, Laminaria digitata, Ulva lactuca) as shown in Figure 4. Whatman paper was used as a reference in the same concentration as for the biomass samples.
As for the end product formation from individual sugars, the main end product from the biomass hydrolysates used was ethanol. The strain was positive for end-product formation (Figure 4) and growth (Supplementary Table S4). Glucose degradation under the same conditions resulted in the formation of 23.8 mM of ethanol, or 1.15 mol/mol glucose degraded. Theoretical yields of glucose from 2.5% w/v Whatman paper that is completely hydrolysed is 15.4 mM. The amount of ethanol produced from Whatman paper hydrolysates is 15.4 mM, or 1.0 mol ethanol/mol glucose which corresponds to 5.6 mM/g glucose, assuming all glucose is released during hydrolysis (control ethanol production subtracted). Not surprisingly, ethanol yields were lower in lignocellulosic biomasses (newspaper, grass, rhubarb) because its more complex nature. Yields on newspaper, grass, and rhubarb were 3.7, 4.4, and 3.9 mM g-1 dw). The highest yields of ethanol from complex biomass by thermophilic bacteria is by Thermoanaerobacter strain BG1L1 on wheat straw and corn stover, or 8.5-9.2 mM ethanol g-1 dw [8,37]. Yields from grass are relatively good compared with other similar species. Thermoanaerobacter AK5 produces 4.1 mM g-1 dw grass [29], and Thermoanaerobacterium strain AK17 produces 5.5 mM g-1 dw [38]. In all cases the grass hydrolysates were pretreated with acid, as well as with enzymatic pretreatment as performed in the present study. Many other reports on thermophiles capable of good ethanol yields from lignocellulose have also been presented in recent years [39,40,41,42,43]. Thus, Thermoanaerobacter strain AK15 seems to be suitable for bioethanol production from complex biomass with good ethanol yields.
Macroalgae is another type of biomass that may be of future use as a potential biomass for bioethanol and bioactive compound production [44,45,46,47]. In the present study, four types of macroalgae were used, two brown algae (Ascophyllum nodusum, Laminaria digitata), one red algae (Palmaria palmata), and one green algae (Ulva lactuca). Ethanol yields for these macroalgae were in most cases lower compared with lignocellulosic biomass. The highest yields were from Laminaria digitata, or 2.8 mM g-1 dw which corresponds to 25% of theoretical yields. Most data on ethanol production from various macroalgae species in literature originate from yeast fermentation of hydrolysates produced by a wide variety of pretreatment methods. Hou and coworkers reported the production 14.7 gL-1 of ethanol using an SSF approach involving cellulase and alginate lyase of L. digitata (10 g/L) using S. cerevisiae as a fermenting microorganism [48]. Similarly, Li and co-workers achieved an ethanol titer of 6.65 gL-1 of ethanol from a cellulase hydrolysate of S. japonica (60 g/L) using S. cerevisiae strain DK 410362 [49] while others have achieved a 14.0 gL-1 ethanol titer from S. japonica waste [50]. A report by Wang et al used Gracilaria salicornia, an invasive brown algae, to achieve a modest ethanol titer of 1.57 g/L using E. coli strain KO11 (ATCC 55124) [51]. A recent investigation of the fermentation of Laminaria digitata hydrolysates, pretreated with enzymatic hydrolysis showed 0.30 gg-1 of consumed substrate (59% of theoretical yields) [52].
2.1.4. Degradation of Amino Acids
The is a huge amount of data concerning sugar metabolism of thermophilic bacteria in general, with most focus on ethanol production from complex biomass. Much less attention has been on the capacity of thermophiles to degrade amino acids. Most of the information on amino acid catabolism among Clostridia has been gained from well-known proteolytic members of Clostridium sporogenes [53], Clostridium botulinum [54] and Clostridium sticklandii [55]. The degradation of amino acids is a complex process involving several oxidation and reduction steps, and only possible under specific conditions and unlike carbohydrates, many amino acids are not degraded as single substrates because of the thermodynamics involved in the reactions [56]. There are various pathways to degrade the 20 amino acids present in proteins, some of which are highly reduced and are not degraded unless the electrons produced in the oxidative step are scavenged [57,58,59]. The most common way to degrade amino acids is to use a two-step mechanism, first an oxidative deamination of the amino acid, yielding a corresponding keto acid, which is oxidatively decarboxylated to give one carbon shorter fatty acids [56]. This is possible under anaerobic conditions only for amino acids with high oxidation stages [56]. The so-called reduced amino acids, e.g. the branched-chain amino acids (leucine, isoleucine, valine) and alanine. In the 1990s, several investigations showed that the reduced amino acids could only be degraded when the amino acid-degrading bacteria could dispose of the electrons produced during the oxidation of these amino acids to an external electron acceptor. This could be done either by co-cultivating the amino acid degrading bacterium with a hydrogenotrophic bacterium, i.e. methanogens, via interspecies hydrogen transfer, or by using thiosulfate as a chemical electron acceptor. This was shown by Thermoanaerobacter brockii during growth on the branched-chain amino acids [60] where leucine, isoleucine, and valine were degraded to 3-methylbutyrate, 2-methylbutyrate and 2-methylpropionate, in the presence of thiosulfate, respectively. Later investigations revealed that the branched-chain amino acids were not only degraded to their corresponding fatty acid but to a mixture of their corresponding fatty acid and alcohol by Thermoanaerobacter brockii, and Caldanaerobacter subterraneous subsp. yonseiensis, when thiosulfate was added into the culture medium [61]. In the present investigation, serine and threonine were the only amino acids that were degraded without the addition of thiosulfate, mainly to ethanol and acetate. In the presence of thiosulfate, these two amino acids were also degraded with a change in the proportion of the end product formation (see equations 1 and 2 below). In the case of serine degradation, acetate became the main end product and ethanol minor product. Similarly, acetate became the main end product in the presence of thiosulfate during threonine degradation, but ethanol increased as well.
Equations 1A and 1B show the stoichiometry of serine degradation without and with thiosulfate, and equations 2A and 2B for threonine, respectively.
20 mM Serine → 6.91 mM Ethanol + 11.33 mM Acetate (Equation 1A)
20 mM Serine + 20 mM S2O3 → 1.21 mM Ethanol + 21.41 mM Acetate (Equation 1B)
20 mM Threonine → 3.42 mM Ethanol + 6.15 mM Acetate (Equation 2A)
20 mM Threonine + 20 mM S2O3 →10.68 mM Ethanol + 16.70 mM Acetate (Equation 2A)
Similarly, Thermoanaerobacter brockii shifts its end product formation during growth on serine in the presence of electron acceptor, pushing end product towards more oxidized end product like acetate and less reduced end products, like ethanol, were produced [49].
In present study, the thiosulfate addition resulted in the degradation of the branched-chain amino acids (leucine, isoleucine, and valine) (Figure 6). As previously reported for other Thermoanaerobacter strains [18], Thermoanaerobacter strain AK15 produced a mixture of branched-chain fatty acids (BCFA) and branched-chain alcohols (BCOH) from branched-chain amino acids (BCAA) and the concentration of the acid was always higher than the alcohol. When the strain was cultivated without the addition of thiosulfate, no growth was observed (Supplementary Table S5).
Figure 5.
End product formation from leucine, isoleucine, and valine (all individually at 20 mM concentrations) in the presence of thiosulfate (40 mM). For simplicity formation of acetate, ethanol and lactate is not shown but was in similar concentrations as in control bottles. Standard deviations are presented as error bars.
Figure 5.
End product formation from leucine, isoleucine, and valine (all individually at 20 mM concentrations) in the presence of thiosulfate (40 mM). For simplicity formation of acetate, ethanol and lactate is not shown but was in similar concentrations as in control bottles. Standard deviations are presented as error bars.
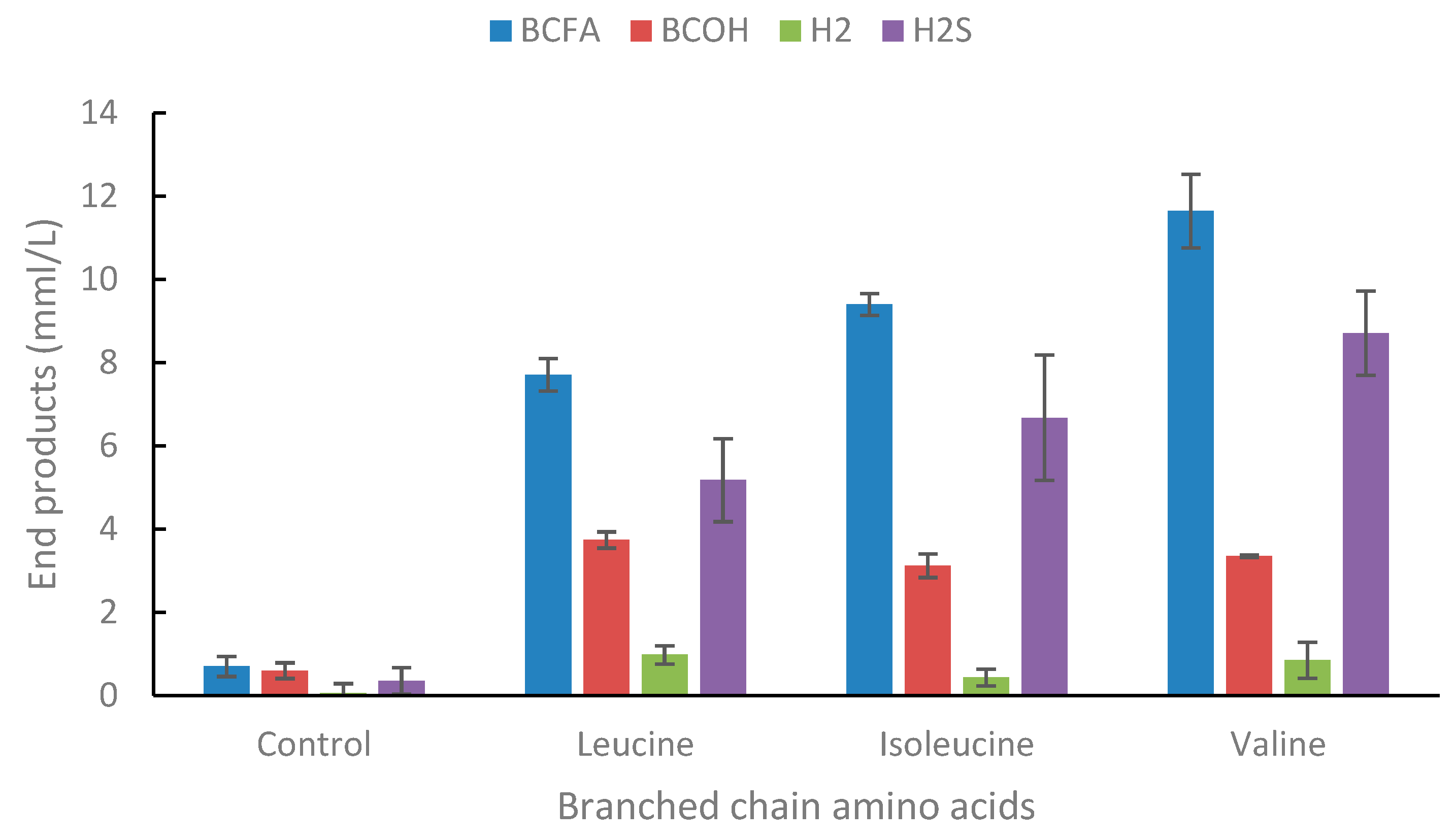
In the presence of thiosulfate leucine was degraded to 3-methylbutyrate and 3-methylbutanol, isoleucine to 2-methylbutyrate and 2-methylbutanol, and valine to 2-methylpropionate and 2-methylpropanol. BCAA have been reported to be degraded to BCFA and BCOH under anaerobic conditions, mostly by Lactobacillus and yeasts that use the Ehrlich pathway. Usually, the concentration of these compounds is of importance for the flavor of foods and beverages [62]. Early studies of thermophilic anaerobic bacteria were performed in several investigations on Thermoanaerobacter brockii. This bacterium was described to degrade the BCAAs to their corresponding BCFAs, but only when thiosulfate was added to the culture medium, acting as a hydrogen scavenger. The thermodynamics in the deamination of leucine to its corresponding keto acid is ΔG + 51.5 kJ/mol whereas the ΔG of the amino acid to 2-methybutyrate is + 4.2 kJ/ Recent work in our laboratory has since then shown that indeed the main bottleneck for the degradation of these reduced amino acids is the first energy-demanding deamination step. Original investigations also showed that both Thermoanaerobacter brockii and Caldanaerobacter subterraneous subsp. yonseiensis degraded the branched-chain amino acids to only their corresponding BCFAs when the strains were co-cultivated in the presence of a hydrogenotrophic methanogen, but to a mixture of their BCFA and BCOH when cultivated alone in the presence of thiosulfate [61]. Thus, it seems to be important how effective the electron acceptor is in determining the product distribution during the degradation of BCAAs. Later studies on other strains within the genera of Thermoanaerobacter and Caldanaerobacter showed that this ability to produce a mixture of alcohols and acids from BCAA was common among both genera [18]. Investigations to understand in more detail the reaction pathway these bacteria use to produce both the acid and the alcohol have been done with Thermoanaerobacter strain AK85 which is closely related to Thermoanaerobacter uzonensis. This study showed that indeed the partial pressure of hydrogen was of great importance for the ratio of end products formed [63]. Finally, it was demonstrated by NMR studies that these bacteria first produce the BCFA which in turn is converted to their corresponding BCOH, both for Thermoanaerobacter strain AK85 and Thermoanaerobacter pseudethanolicus [63,64].
2.1.5. Degradation of Proteins
Protein degradation by thermophilic anaerobic bacteria has received much less attention as compared with carbohydrates. Mesophilic anaerobic protein degrading bacteria have however got much more attention, mainly because of the high number of pathogens that are proteolytic. Studies on thermophilic anaerobes have been limited to several genera like Caloramator and Thermoanaerobacter. The importance of electron acceptors for protein and amino acid degradation has been known for some time now and is important knowledge to understand the role of thermophilic bacteria in hot environments [65].
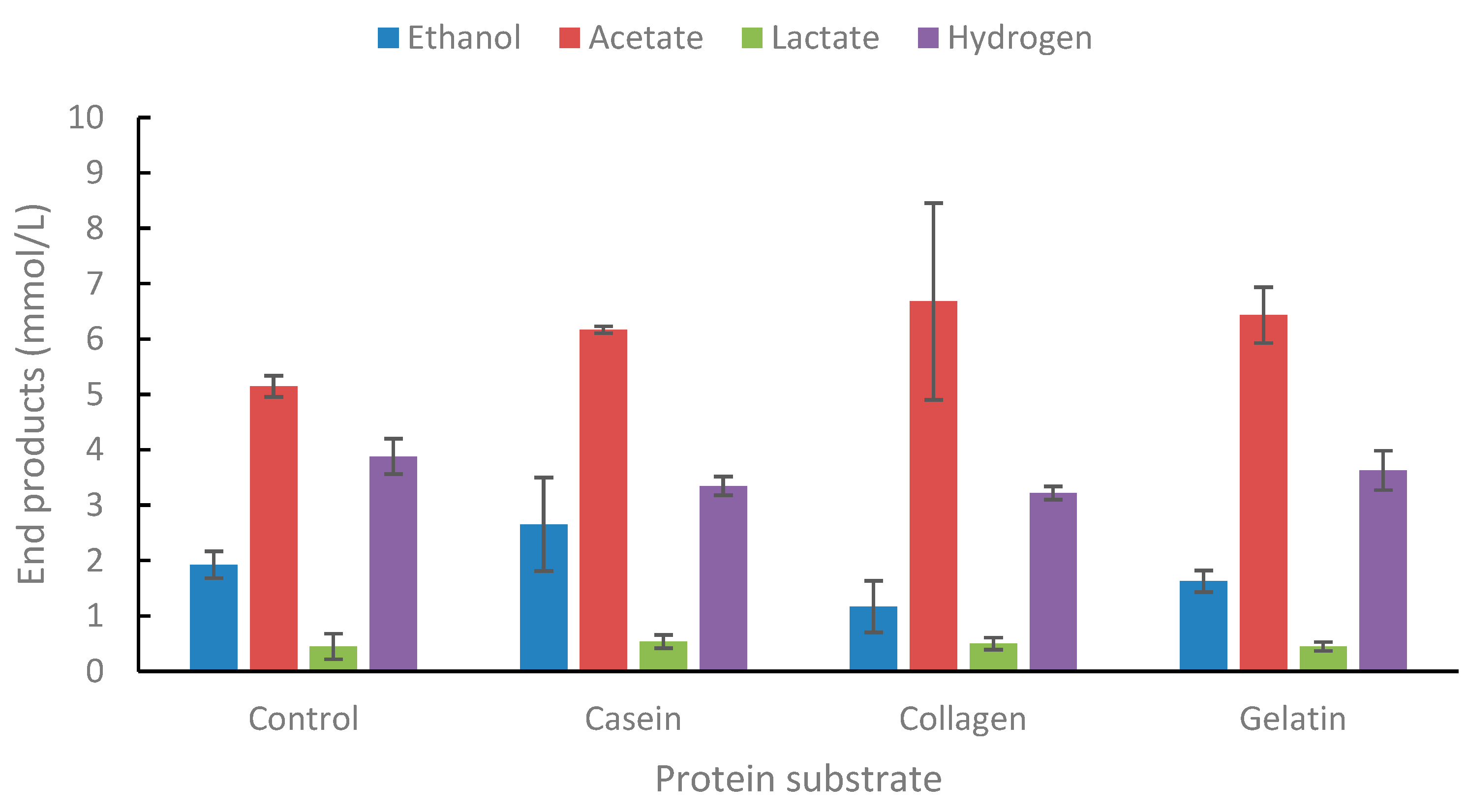
The strain was tested for growth on three types of proteins, casein, collagen, and gelatin (Figure 6). A slight increase was observed in acetate from casein and collagen as compared to controls. However, the addition of peptone and yeast extract enhanced end-product formation by the strain (results not shown).
2.1.6. Conversion of fatty acids to alcohols
Recent investigations have shown that bacteria within the genera of Thermoanaerobacter and Caldanaerobacter can dispose their electrons produced during glucose (and other sugars) oxidations not only to pyruvate to produce ethanol or lactate but may also use other electron acceptors like fatty acids which are converted to their corresponding alcohols [28,65,66]. This was tested for strain AK15 by cultivating the strain on glucose only and on glucose in the presence of butyrate. The strain degraded glucose to a mixture of ethanol, acetate, and lactate according to equation 3:
1 Glucose → 1.24 Ethanol + 0.48 Acetate + 0.05 Lactate + 0.54 H2 (eq. 3)
When the strain was cultivated on glucose with the addition of 20 mM of butyrate the reaction stoichiometry changed according to equation 4.
1 Glucose + 1 Butyrate → 0.78 Ethanol + 0.87 Acetate + 0.58 Butyrate + 0.36 Butanol (eq. 4)
Thus, as expected, the strain produces less ethanol and more acetate in the presence of butyrate as an electron acceptor, and the fatty acid is partially converted to its corresponding alcohol, butanol. No lactate and hydrogen were observed as end products at end of fermentation in the presence of butyrate This was also investigated for other volatile fatty acids, like propionate, branched chain fatty acids and pentanol, with similar results of the conversion of the fatty acid to their corresponding alcohol (results not shown). Thermoanaerobacter pseudethanolicus has recently been shown to convert fatty acids to alcohols, during sugar degradation [66]. Production of high carbon alcohols from complex biomass by adding cheap volatile fatty acids to the fermentation broth of the hydrolysates is indeed a new way of biofuel production that may be of great importance in the near future.
3. Materials and Methods
3.1. Chemicals
All chemicals were obtained from Sigma-Aldrich unless otherwise noted. L-fucose and fucoidan were obtained from Dextra (Reading, UK). Starch was from corn. Rhamnogalacturonan (from soy), mannan (from ivory nut), and galactan (from lupin) were obtained from Megazyme. Keratin was locally obtained from milled and used without further preparation.
3.2. Culture Medium and Preparation
Thermoanaerobacter strain AK15 was cultivated in Basal Mineral (BM) medium prepared as previously described [67]; the medium consisted of (per liter): NaH2PO4 2.34 g, Na2HPO4 3.33 g, NH4Cl 2.2 g, NaCl 3.0 g, CaCl2 8.8 g, MgCl2 x 6 H2O 0.8 g, yeast extract 2.0 g, resazurin 1 mg, trace element solution 1 mL, vitamin solution (DSM141) 1 mL, and NaHCO3 0.8 g. The trace element solution consisted of the following on a per liter basis: FeCl2 x 4H2O 2.0g, EDTA 0.5 g, CuCl2 0.03 g, H3BO3, ZnCl2, MnCl2 x 4H2O, (NH4)Mo7O24, AlCl3, CoCl2 x 6H2O, NiCl2, and 0.05 g, Na2S x 9H2O 0.3 g, and 1 mL of concentrated HCl. The medium was prepared by adding the buffer to distilled water containing resazurin and boiled for 10 min and cooling under nitrogen flushing. The mixture was then transferred to serum bottles using the Hungate technique [68] and autoclaved (121°C) for 60 min. All other components of the medium were added separately through filter (0.45 µm) sterilized solutions. All experiments were conducted at 65°C and at pH of 7.0 with a liquid-gas (L-G) ratio of 1:1 unless otherwise noted. In all cases, experiments were performed in triplicate.
3.3. Bacterial Strain
Thermoanaerobacter strain AK15 was isolated from an alkaline hot spring, Viti (pH 8.6; temperature 60°C) in NE-Iceland according to Scully and coworkers [18]. The strain has been sequenced for 16S rRNA and is most closely related to Thermoanaerobacter uzonensis [18]. The strain was persevered in rigorously degassed by sonication under vacuum BM medium supplemented with 30% v/v glycerol and stored at -20°C. All cultivations were conducted at pH 7.0 at 65°C. All inoculation stocks of the strain were taken from frozen (-20°C) culture with rigorously degassed 30% (v/v) glycerol and reactivated on BM medium containing glucose (20 mM). Reactivated cultures were inoculated (2% v/v) from exponential growth phase to 25 mL serum bottles (liquid-gas ratio 1:1). Cultures were grown for five days and screened for end-product formation.
3.4. Substrate Utilization Spectrum
The ability of strains to utilize selected hexoses (D-glucose, D-mannose, D-galactose), pentoses (D-xylose, D-arabinose, D-ribose), methypentoses (L-fucose and L-rhamnose), sugar alcohols (mannitol), and cellobiose were evaluated at a concentration of 20 mM except for cellobiose (10 mM). Amino acids were tested at 20 mM concentration in the absence and presence of thiosulfate. Polymeric substrates (starch, cellulose, laminarin, xylan, chitosan, chitin, casein, keratin, collagen, lichenan, pectin, and keratin), were evaluated at a concentration of 0.2% (w/v) except laminarin (0.1% (w/v)). Cultures were incubated for a period of 5 days at which time end products were analysed. Experiments were done in 25 mL serum bottles with liquid–gas phase of 1.0 ratio. Control bottles were only with yeast extract (2 g L-1).
Samples of cellulose and complex biomass were obtained from Whatman paper, printed newspaper, grass (Phleum pratense), rhubarb (Rheum rhabarbarum), Ascophyllum nodosum, Palmaria palmata, Laminaria digitata, and Ulva lactuca. The biomass was pretreated as previously described with 0.1% H2SO4 and enzymes (CelluclastR and Novozyme 188) as previously described [32], rendering hydrolysates that were diluted to a concentration of 2.5% (w/v).
3.5. Influence of Liquid-Gas Phase Ratio
The influence of partial hydrogen pressure (ρH2) on end product formation was investigated with different ratios of liquid and gas phases when grown on 20 mM glucose. The liquid phase varied from 5.0, 29.45, and 50 mL in serum bottles with a total volume of 57 mL; thus, the L-G volume ratio varied from 0,09, 1.00, and 5.62, respectively. A control experiment at each L-G consisted of BM medium containing yeast extract (0.2% w/v) only. After 5 days of incubation, end products were quantified by GC.
3.6. Analytical Methods
Hydrogen was analyzed by Perkin Elmer Auto System XL gas chromatograph according to Orlygsson and Baldursson [67]. Alcohols and volatile fatty acids were measured by gas chromatography using a Perkin-Elmer Clarus 580 gas chromatograph as previously described [67]. Lactate was quantified colorimetrically according to the method of Taylor [69] with modification according to Scully and Orlygsson [63]. Optical density was determined by measuring absorbance at 600 nm by a Shimadzu UV-1800 UV-Vis spectrophotometer in a quartz cuvette with a pathlength of 1 cm. Hydrogen sulfide was analysed as described by Cline [70].
4. Conclusions
The data presented shows that the thermophilic anaerobic bacterium Thermoanaerobacter strain AK15 is a potentially good organism for biotechnological applications. The bacterium is highly ethanologenic, producing more than 1.57 mol ethanol per mol glucose degraded and good ethanol yields from complex biomass hydrolysates but not from most of the untreated biopolymers and proteins applied. The strain is also interesting concerning producing fine chemicals like branched-chain alcohols from branched-chain amino acids and converting volatile fatty acids to high-carbon fuel molecules. Finally, the strain possesses many enzymes of importance for bioprocessing purposes.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org. Supplementary Tables S1-S5.
Author Contributions
Conceptualization, J.O and S.M.S.; Methodology, J.O and S.M.S.; Software, J.O and S.M.S.; Validation, J.O.; Formal analysis, J.O and S.M.S.; Investigation, S.M.S.; Resources, J.O.; Data curation, J.O and S.M.S..; Writing—original draft preparation, J.O.; Writing—review and editing, J.O and S.M.S.; Visualization, J.O. and S.M.S.; Supervision, J.O.; Project administration, J.O. Both authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Demain, A. Biosolutions to the energy problem. J. Ind. Microbiol. Biot. 2009, 36, 319–332. [Google Scholar] [CrossRef] [PubMed]
- Taylor, M.P.; Eley, K.L.; Martin, S.; Tuffin, M.I.; Burton, S.G.; Cowan, D.A. Thermophilic ethanologenesis: future prospects for second-generation bioethanol production. Trends Biotechnol. 2009, 27, 398–405. [Google Scholar] [CrossRef] [PubMed]
- Scully, S.M.; Orlygsson, J. Recent Advances in Second Generation Ethanol Production by Thermophilic Bacteria. Energies 2015, 8, 1–30. [Google Scholar] [CrossRef]
- Hahn-Hagerdal, B.; Galbe, M.; Gorwa-Grauslund, M.F.; Liden, G.; Zacchi, G. Bioethanol - the future of tomorrow from the residues of today. Trends Biotechnol. 2006, 12, 549–556. [Google Scholar] [CrossRef]
- Kelly, M.S.; Dworjanyn, S. The potential of marine biomass for anaerobic biogas production; Wrown Estate: London, UK, 2008. [Google Scholar]
- Tedesco, S.; Stokes, J. Valorisation to biogas of macroalgal waste streams: A circular approach to bioproducts and bioenergy in Ireland. Chem. Pap. 2017, 71, 721–728. [Google Scholar]
- Wiegel, J.; Ljungdahl, L. Thermoanaerobacter ethanolicus gen. nov., spec. nov., a new, extreme thermophilic, anaerobic bacterium. Arch. Microbiol 1981, 128, 343–348. [Google Scholar] [CrossRef]
- Georgieva, T.I.; Mikkelsen, M.J.; Ahring, B.K. Ethanol production from wet-exploded wheat straw hydrolysate by thermophilic anaerobic bacterium Thermoanaerobacter BG1L1 in a continuous immobilized reactor. Appl. Biochem. Biotech. 2008, 145, 99–110. [Google Scholar] [CrossRef]
- Santangelo, T.J.; Cubonova, L.; Reeve, J.N. Deletion of alternative pathways for reductant recycling of Thermococcus kodakarensis increases hydrogen production. Mol. Microbiol. 2011, 81, 897–911. [Google Scholar] [CrossRef]
- Lipscomb, G.L.; Stirrett, K.; Schut, G.J.; Yang, F.; Jenney, F.E.; Scott, R.A.; Adams, M.W.W.; Westpheling, J. Pyrococcus furiosus facilitates genetic manipulation: Construction of markerless deletions of genes encoding the two cytoplasmic hydrogenases. Appl. Environ. Microb. 2011, 77, 2232–2238. [Google Scholar] [CrossRef]
- Cha, M.; Chung, D.W.; Elkins, J.G.; Guss, A.M.; Westpheling, J. Metabolic engineering of Caldicellulosiruptor bescii yields increased hydrogen production from lignocellulosic biomass. Biotechnol. Biofuels. 2013, 6, 85. [Google Scholar] [CrossRef]
- Pradhan, N.; Dipasquale, L.; d’Ippolito, G.; Panico, A.; Lens, P.N.L.; Esposito, G.; Fontana, A. Hydrogen and lactic acid synthesis by the wild-type and laboratory strain of the hyperthermophilic bacterium Thermotoga neapolitana DSMZ 4359T under capnophilic lactic fermentation conditions. Int. J. Hydrogen Energ. 2017, 42, 16023–16030. [Google Scholar] [CrossRef]
- Martin, A.; Murphy, F.H. Propylene glycols. In Kirk-Othmer Encyclopedia of Chemical Technology, 4th edition; Kroschwitz, J.I. (Ed.)., Ed.; Wiley: New York, 1994; Vol. 17, pp. 715–726. [Google Scholar]
- Saxena, R.; Anand, P.; Saran, S.; Isar, J.; Agarwal, L. Microbial production and applications of 1,2-propanediol. Ind. J. Microbiol. 2010, 50, 2–11. [Google Scholar] [CrossRef]
- Cameron, D.C.; Cooney, C.L. A novel fermentation: The production of ®-1,2-propanediol and acetol by Clostridium thermosaccharolyticum. Bioresource Technol. 1986, 4, 651–654. [Google Scholar] [CrossRef]
- Ingvadottir, E.M.; Scully, S.M.; Orlygsson, J. Evaluation of the genus of Caldicellulosiruptor for production of 1,2-propanediol from methylpentoses. Anaerobe 2017, 47, 86–88. [Google Scholar] [CrossRef]
- Seyfried, M.; Lyon, D.; Rainey, F.A.; Wiegel, J. Caloramator viterbensis sp. nov., a novel thermophilic glycerol-fermenting bacterium isolated from a hot spring in Italy. Int. J. Syst. Evol. Micr. 2002, 52, 1177–1184. [Google Scholar]
- Scully, S.M.; Iloranta, P.; Myllymaki, P.; Orlygsson, J. Branched-chain alcohol formation by thermophilic bacteria within the genera of Thermoanaerobacter and Caldanaerobacter. Extremophiles 2015, 19, 809–18. [Google Scholar] [CrossRef] [PubMed]
- Crosby, J.R.; Laemthong, T.; Lewis, A.M.; Straub, C.T.; Adams, M.W.W.; Kelly, R.M. Extreme thermophiles as emerging metabolic platforms. Curr. Opin. Biotech. 2019, 59, 55–64. [Google Scholar] [CrossRef]
- Stackebrandt, E. The family Thermoanaerobacteraceae. In The Prokaryotes - Firmicutes and Tenericutes; Rosenberg, E., et al., Eds.; Springer-Verlag: Berlin, 2014. [Google Scholar]
- Collins, M.D.; Lawson, P.A.; Willems, A.; Cordoba, J.J.; Fernandezgarayzabal, J.; Garcia, P. The phylogeny of the genus Clostridium - proposal of 5 new genera and 11 new species combinations. Int. J. Syst. Bacteriol. 1994. 44, 812–826.
- Larsen, L.; Nielsen, P. & Ahring, B.K. Thermoanaerobacter mathranii sp nov, an ethanol-producing, extremely thermophilic anaerobic bacterium from a hot spring in Iceland. Arch. Microbiol.
- Lee, Y.E.; Jain, M.K.; Lee, C.Y.; Lowe, S.E.; Zeikus, J.G. Taxonomic distinction of saccharolytic thermophilic anaerobes - description of Thermoanaerobacterium xylanolyticum gen-nov, sp-nov, and Thermoanaerobacterium saccharolyticum gen-nov, sp-nov - reclassification of Thermoanaerobium brockii, Clostridium thermosulfurogenes, and Clostridium thermohydrosulfuricum E100-69 as Thermoanaerobacter brockii comb-nov, Thermoanaerobacterium thermosulfurigenes comb-nov, and Thermoanaerobacter thermohydrosulfuricus comb-nov, respectively - and transfer of Clostridium thermohydrosulfuricum 39E to Thermoanaerobacter ethanolicus. Int. J. Syst. Bacteriol. 1993, 43, 41–51. [Google Scholar]
- Slobodkin, A.I.; Tourova, T.P.; Kuznetsov, B.B.; Kostrikina, N.A.; Chernyh, N.A.; Bonch-Osmolovskaya, E.A. Thermoanaerobacter siderophilus sp. nov., a novel dissimilatory Fe(III)-reducing, anaerobic thermophilic bacterium. Int. J. Syst. Bacteriol. 1999, 49, 1471–1478. [Google Scholar] [CrossRef]
- Lacis. L.S.; Lawford, H.G. Ethanol production from xylose by Thermoanaerobacter ethanolicus in batch and continuous culture. Arch. Microbiol. 1988, 150, 48–55. [Google Scholar] [CrossRef]
- Avci, A.; Donmez, S. Effect of zinc on ethanol production by two Thermoanaerobacter strains. Process Biochem. 2009, 41, 984–989. [Google Scholar] [CrossRef]
- Onyenwoke, R.U.; Kevbrin, V.V.; Lysenko, A.M.; Wiegel, J. Thermoanaerobacter pseudethanolicus sp. nov., a thermophilic heterotrophic anaerobe from Yellowstone National Park. Int. J. Syst. Evol. Microbiol 2007, 57, 2191–2193. [Google Scholar] [CrossRef]
- Hitschler, L.; Kuntz, M.; Langschied, F.; Basen, M. Thermoanaerobacter species differ in their potential to reduce organic acids to their corresponding alcohols. Appl. Microbiol. Biot. 2018, 102, 8465–8476. [Google Scholar] [CrossRef] [PubMed]
- Brynjarsdottir, H.; Wawiernia, B.; Orlygsson, J. Ethanol production from sugars and complex biomass by Thermoanaerobacter AK5: The effect of electron-scavenging systems on end-product formation. Energ. Fuel 2012, 26, 4568–4574. [Google Scholar] [CrossRef]
- Ingvadottir, E.M.; Scully, S.M.; Orlygsson, J. Production of (S)-1,2-prpanediol from rhamnose using the moderate thermophilic Clostridium strain AK1. Anaerobe 2018, 54, 26–30. [Google Scholar] [CrossRef] [PubMed]
- Altaras, N.E.; Etzel, M.R.; Cameron, D.C. Conversion of sugars to 1,2-propanediol by Thermoanaerobacterium thermosaccharolyticum HG-8. Biotechnol. Progr. 2001, 17, 52–56. [Google Scholar] [CrossRef]
- Chades, T.; Scully, S.M.; Ingvadottir, E.M.; Orlygsson, J. Fermentation of Mannitol Extracts From Brown Macro Algae by Thermophilic Clostridia. Front. Microbiol. 2018, 9, 1–13. [Google Scholar] [CrossRef]
- Wagner, I.D.; Zhao, W.; Zhang, C.L.; Romanek, C.S.; Rohde, M.; Wiegel, J. Thermoanaerobacter uzonensis sp. nov., an anaerobic thermophilic bacterium isolated from a hot spring within the Uzon Caldera, Kamchatka, Far East Russia. Int. J. Syst. Evol. Microbiol. 2008, 58, 2565–2573. [Google Scholar] [CrossRef]
- Rydzak, T.; Levin, D.B.; Cicek, N.; Sparling, R. End-product induced metabolic shifts in Clostridium thermocellum ATCC 27405. Appl. Microbiol Biotechnol. 2011, 2011. 92, 199–209. [Google Scholar] [CrossRef]
- Verbeke, T.J.; Zhang, X.; Henrissat, B.; Spicer, B.; Tydzak, T.; Krokhin, O.V.; Fristensky, B.; Levin, D.B.; Sparling, R. Genomic evaluation of Thermoanerobacter spp. for the construction of designer co-cultures to improve lignocellulosic biofuel production. PLoS ONE 2023, 8, 3. [Google Scholar]
- Riessen, S.; Antranikian, G. Isolation of Thermoanaerobacter keratinophilus sp. nov., a novel thermophilic, anaerobic bacterium with keratinolytic activity. Extremophiles 2001, 5, 399–408. [Google Scholar] [CrossRef] [PubMed]
- Georgieva, T.I.; Ahring, B.K. Evaluation of continuous ethanol fermentation of dilute-acid corn stover hydrolysate using thermophilic anaerobic bacterium Thermoanaerobacter BG1L1. Appl. Microbiol. Biot. 2007, 77, 61–8. [Google Scholar] [CrossRef] [PubMed]
- Almarsdottir, A.R.; Sigurbjornsdottir, M.A.; Orlygsson, J. Effect of various factors on ethanol yields from lignocellulosic biomass by Thermoanaerobacterium AK 17. Biotechnol. Bioeng. 2012, 109, 686–694. [Google Scholar] [CrossRef] [PubMed]
- Kaparaju, P.; Serrano, M.; Thomsen, A.B.; Kongjan, P.; Angelidaki, I. Bioethanol, biohydrogen and biogas production from wheat straw in a biorefinery concept. Bioresource Technol. 2008, 100, 2562–2568. [Google Scholar] [CrossRef] [PubMed]
- Beluhan, S.; Mihajlovski, K.; Santek, B.; Santek, M.I. The production of bioethanol from lignocellulosic biomass: Pretreatment methods, fermentation, and downstream processing. Energies 2023, 16, 19. [Google Scholar] [CrossRef]
- Tomas, A.F.; Karagöz, P.; Karakashev, D.; Angelidaki, I. Extreme thermophilic ethanol production from rapeseed straw: Using the newly isolated Thermoanaerobacter pentosaceus and combining with Saccharomyces cerevisae in a two-step process. Biotechnol. Bioeng. 2013, 110, 1574–1582. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Y.J.; Xin, F.X.; Lu, J.S.; Dong, W.L.; Zhang, W.M.; Zhang, M.; Wu, H.; Ma, J.F.; Jiang, M. State of the art review of biofuels production from lignocellulose by thermophilic bacteria. Bioresource Technol. 2017, 245, 1498–1506. [Google Scholar] [CrossRef]
- Zuliani, L.; Serpico, A.; Di Simone, M.; Frison, N.; Fusco, S. Biorefinery gets hot: Thermophilic enzymes and microorganisms for second generation bioethanol production. Processes 2021, 9, 9. [Google Scholar] [CrossRef]
- Tong, K.T.X.; Tan, I.S.; Foo, H.C.Y.; Lam, M.K.; Lim, S.; Lee, K.T. Advancement of biorefinery-derived platform chemicals from macroalgae: a perspective for bioethanol and lactic acid. Biomass Conversion and Biorefinery 2022. [Google Scholar] [CrossRef]
- Shibasaki, S.; Ueda, M. Utilization of macroalgae for the production of bioactive compounds and bioprocesses using microbial biotechnology. Microorganisms. 2023, 11, 1499. [Google Scholar] [CrossRef] [PubMed]
- Priyadharsini, P.; Dawn, S.S.; Arun, J.; Ranjan, A.; Jayaprabakar, J. Recent advances in conventional and genetically modified macroalgal biomass as substrates in bioethanol production: a review. Biofuels UK. 2023, 2023. 14, 1103–1118. [Google Scholar]
- Zheng, Y.Z; Li, Y.P.; Yang, Y.Y.; Zhang, Y.; Wang, D.; Wang, P.Y.; Wong, A.C.Y.; Hsieh, Y.S.Y.; Wang, D.M. Recent advances in bioutilization of marine macroalgae carbohydrates: Degradation, metabolism, and fermentation. J. Agr. Food Chem. 2022, 2022. 70, 1438–1453. [Google Scholar] [CrossRef]
- Hou, X.; Hansen, J.H.; Bjerre, A.B. Integrated bioethanol and protein production from brown seaweed Laminaria digitata. Bioresource. Technol. 2015, 197, 310–317. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.Y.; Li, P.; Lee, J.; Ryu, H.J.; Oh, K.K. Ethanol production from Saccharina japonica using an optimized extremely low acid pretreatment followed by simultaneous saccharification and fermentation. Bioresource Technol. 2013, 127, 127,119–125. [Google Scholar] [CrossRef] [PubMed]
- Ge, L.; Wang, P.; Mou, H. Study on saccharification techniques of seaweed wastes for the transformation of ethanol. Renew. Energ 2011, 36, 84–89. [Google Scholar] [CrossRef]
- Kim, N.J.; Li, H.; Jung, K.; Chang, H.N.; Lee, P.C. Ethanol production from marine algal hydrolysates using Escherichia coli KO11, Bioresource. Technol. 2011, 10, 7466–7469. [Google Scholar] [CrossRef]
- Moenaert, A.; Lopez-Contreras, A.M.; Budde, M.; Allahgholi, L.; Xiaoru, H.; Bjerre, A.-B.; Orlygsson, J.; Karlsson, E.N.; Fridjonsson, O.H.; Hreggvidsson, G.O. Evaluation of Laminaria digitata hydrolysate for the production of bioethanol and butanol by fermentation. Fermentation 2023, 9, 59. [Google Scholar] [CrossRef]
- Stickland, L.H. CCXXXII. Studies in the metabolism of the strict anaerobes (genus Clostridium). The chemical reactions by which Cl. sporogenes obtains its energy. Biochem. J. 1934, 28, 1746–1759. [Google Scholar] [CrossRef]
- Deklevat, M.L.; Dasgupta, B.R. Purification and characterization of a protease from Clostridium botulinum Type A that nicks single-chain type A botulinum neurotoxin into the Di-Chain form. J. Bacteriol. 1990, 172, 2498–2503. [Google Scholar] [CrossRef]
- Elsden, S.R.; Hilton, M.G. The end products of metabolism of aromatic amino acids by clostridia. Arch. Microbiol. 1976, 107, 283–288. [Google Scholar] [CrossRef] [PubMed]
- Scully, S.M. Amino acid and related catabolisms of Thermoanaerobacter species. PhD thesis. University of Iceland. 2019. [Google Scholar]
- Elsden, S.R.; Hilton, M.G. Volatile acid production from threonine, valine, leucine and isoleucine by clostridia. Arch. Microbiol. 1978, 117, 165–172. [Google Scholar] [CrossRef] [PubMed]
- Laanbroek, H.H.; Smit, A.J.; Nulend, G.K.; Veldkamp, H. Competition for glutamate between specialized and versatile Clostridium species. Arch. Microbiol. 1979, 66, 61–66. [Google Scholar] [CrossRef] [PubMed]
- Toda, Y.; Saiki, T.; Uozumi, T.; Beppu, T. Isolation and characterization of protease-producing, thermophilic, anaerobic bacterium, Thermobacterides leptospartum sp. nov. Agr. Biol. Chem. 1988, 52, 1339–1344. [Google Scholar] [CrossRef]
- Fardeau, M.-L.; Patel, B.K.C.; Magot, M.; Ollivier, B. Utilization of serine, leucine, isoleucine, and valine by Thermoanaerobacter brockii in the presence of thiosulfate or Methanobacterium sp. as electron acceptors. Anaerobe 1997, 3, 405–410. [Google Scholar] [CrossRef] [PubMed]
- Scully, S.M.; Orlygsson, J. Branched-chain alcohol formation from branched-chain amino acids by Thermoanaerobacter brockii and Thermoanaerobacter yonseiensis. Anaerobe. 2014, 30, 82–84. [Google Scholar] [CrossRef]
- Smit, G.; Smit, B.A.; Engels, W.J.M. Flavour formation by lactic acid bacteria and biochemical flavor profiling of cheese products. FEMS Microbiol. Rev. 2005, 29, 591–610. [Google Scholar] [CrossRef]
- Scully, S.M.; Orlygsson, J. Branched-chain amino acid catabolism of Thermoanaerobacter strain AK85 and the influence of culture conditions on branched-chain alcohol formation. Amino acids 2019, 51, 1039–1054. [Google Scholar] [CrossRef]
- Scully, S.M.; Orlygsson, J. Branched-chain amino acid catabolism of Thermoanaerobacter pseudethanolicus reveals potential route to branched-chain alcohol formation. Extremophiles. 2020, 24, 121–133. [Google Scholar] [CrossRef]
- Faudon, C.; Fardeau, M.L.; Heim, J.; Patel, B.; Magot, M.; Ollivier, B. Peptide and amino acid oxidation in the presence of thiosulfate by members of the genus Thermoanaerobacter. Curr. Microbiol. 1995, 31, 152–157. [Google Scholar] [CrossRef]
- Scully, S.M.; Brown, A.E.; Mueller-Hilger, Y.; Ross, A.B.; Orlygsson, J. Influence of culture conditions on the bioreduction of organic acids to alcohols by Thermoanaerobacter pseudethanolicus. Microorganisms 2021, 9, 162. [Google Scholar] [CrossRef]
- Orlygsson, J.; Baldursson, S.R.B. Phylogenetic and physiological studies of four hydrogen-producing thermoanareobes. Iceland Agr. Sci. 2007, 20, 93–105. [Google Scholar]
- Hungate, R.E. A roll tube method for cultivation of strict anaerobes. In Methods in microbiology; Norris, J.R., Ribbons, D.W. (eds)., Eds.; Academic Press: New York, 1969; vol 3B, pp. 117–132. [Google Scholar]
- Taylor, K.A.C.C. A simple colorimetric assay for muramic acid and lactic acid. Appl. Biochem. Biotech. 1996, 56, 49–58. [Google Scholar] [CrossRef]
- Cline, J.D. Spectrophotometric determination of hydrogen sulfide in natural waters. Limnol Oeanogr. 1969, 14, 454–458. [Google Scholar] [CrossRef]
Figure 1.
End product formation from various substrates. From left to right, acetate, ethanol, lactate, and hydrogen. Standard deviations are presented as error bars.
Figure 1.
End product formation from various substrates. From left to right, acetate, ethanol, lactate, and hydrogen. Standard deviations are presented as error bars.
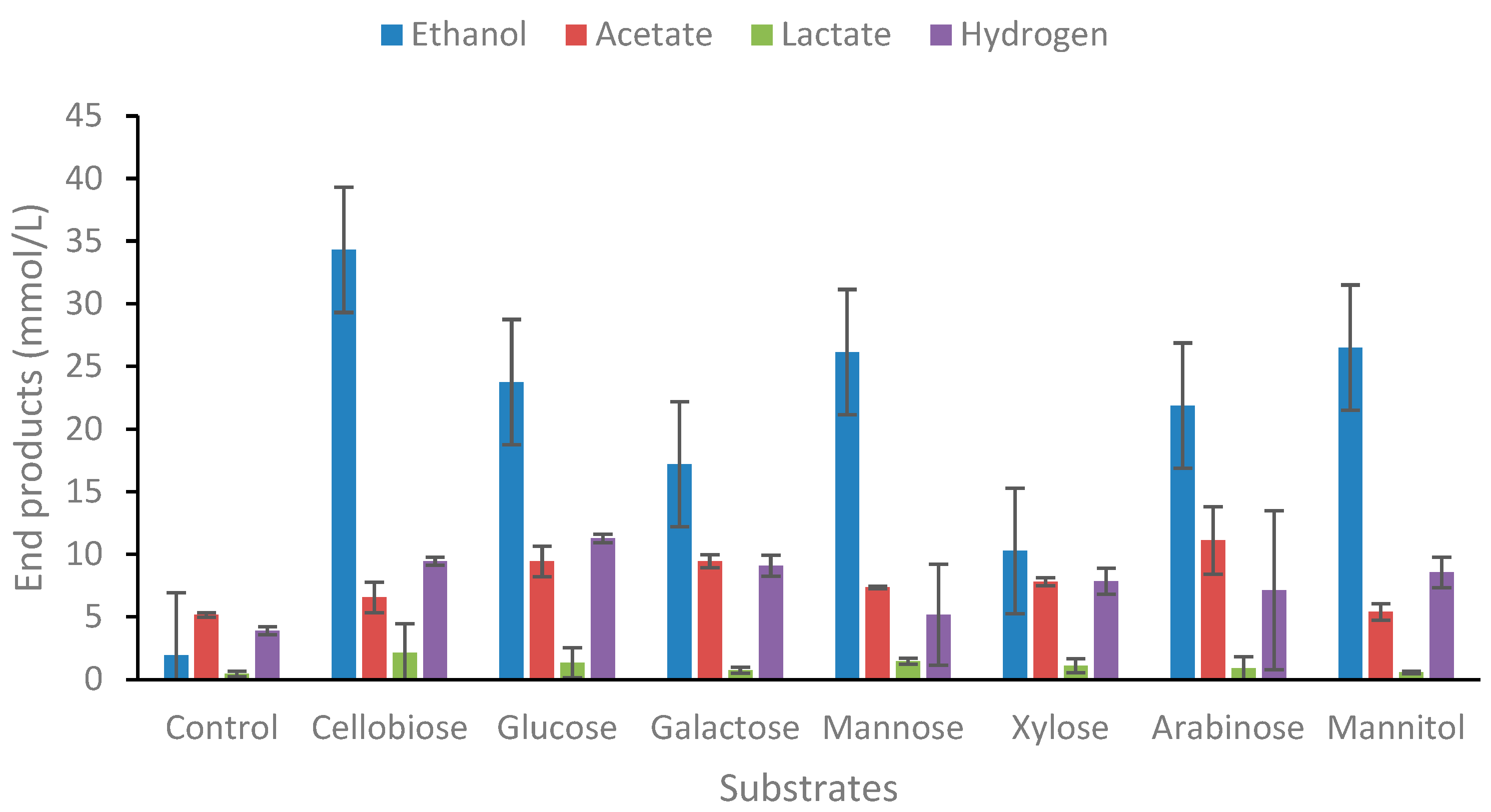
Figure 2.
End product formation from glucose (20 mM) at different liquid – gas phase ratios. Standard deviations are presented as error bars.
Figure 2.
End product formation from glucose (20 mM) at different liquid – gas phase ratios. Standard deviations are presented as error bars.
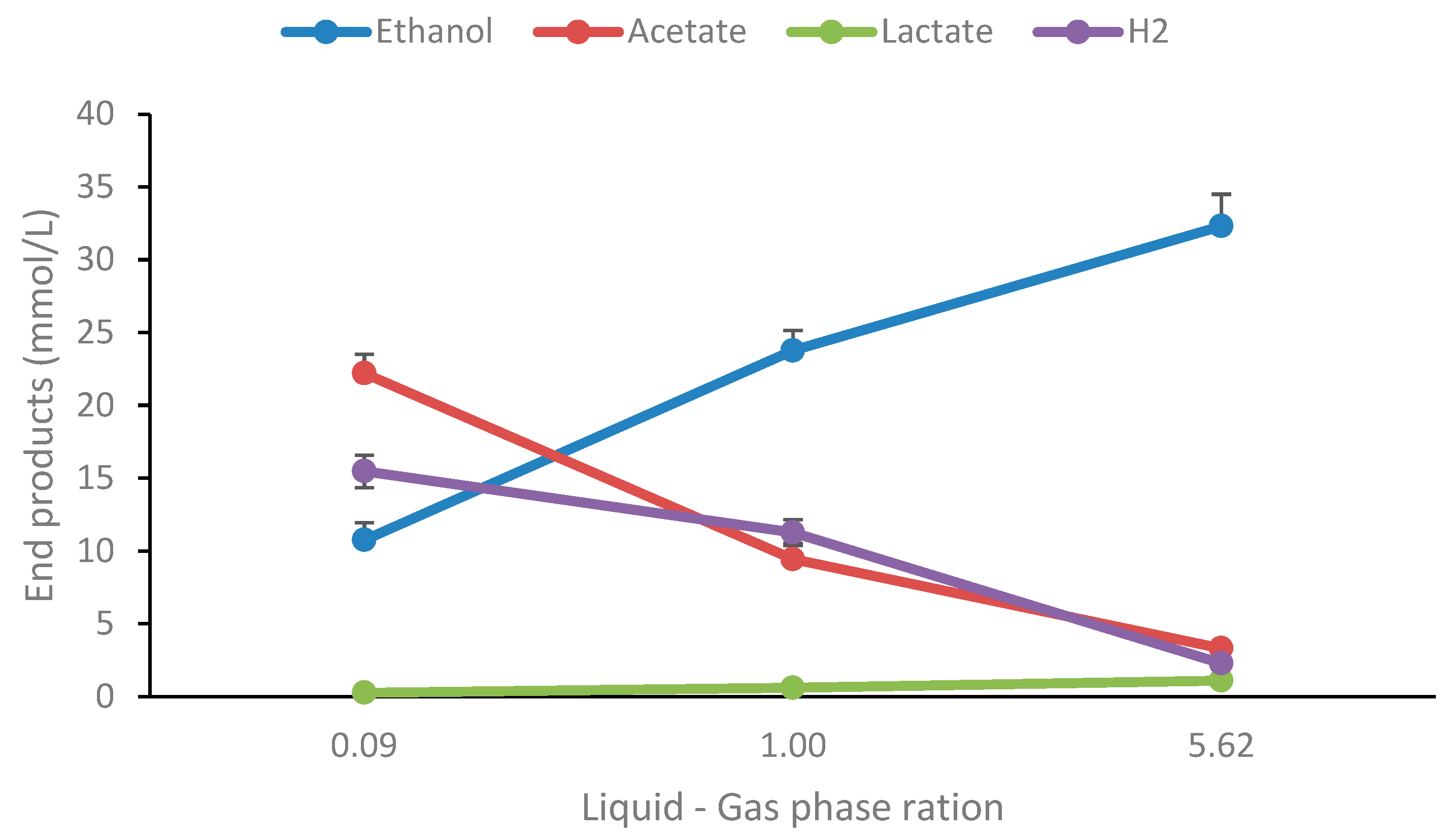
Figure 3.
End product formation from various polymeric substrates. Standard deviations are presented as error bars.
Figure 3.
End product formation from various polymeric substrates. Standard deviations are presented as error bars.
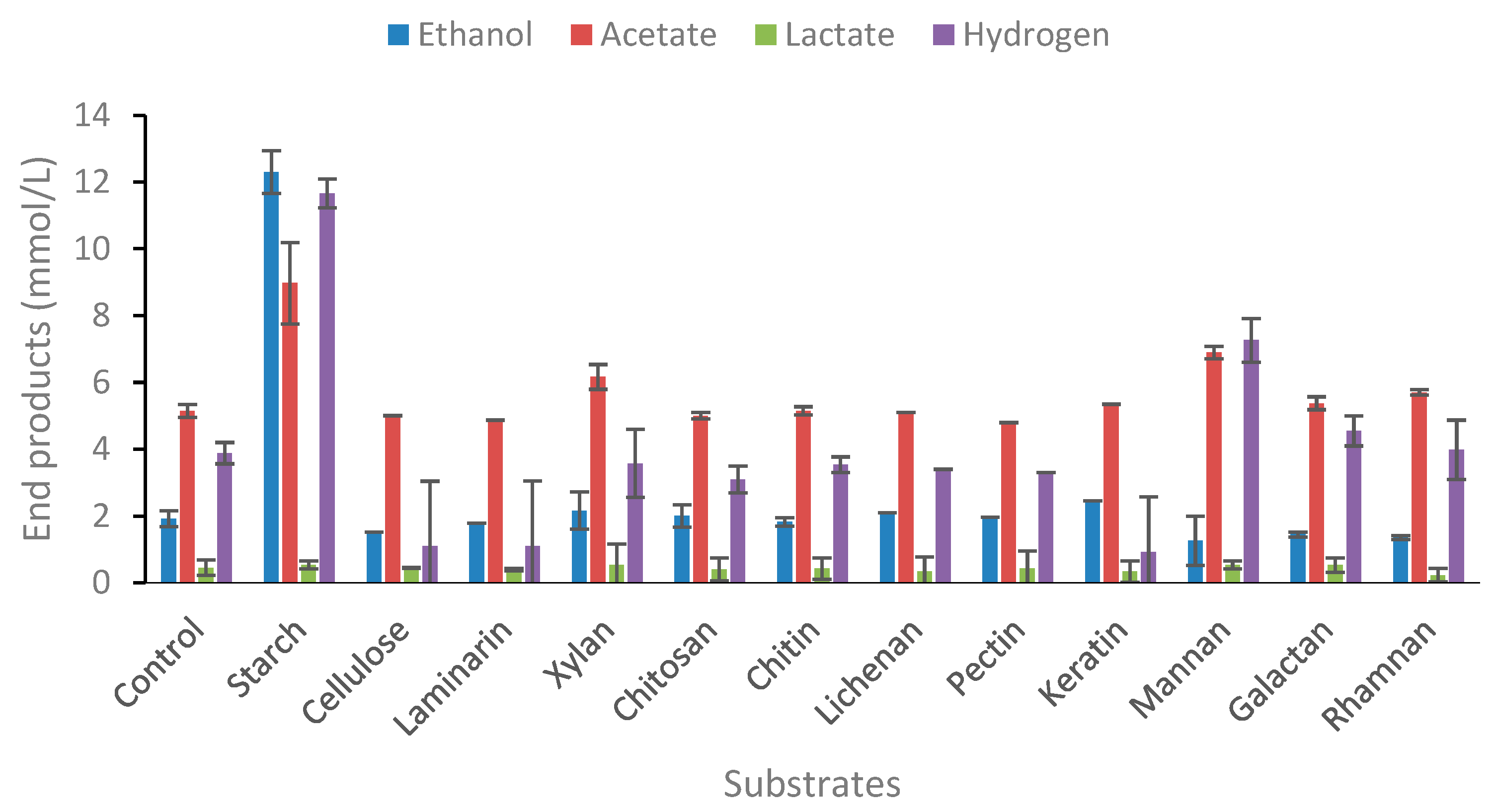
Figure 4.
End product formation from hydrolysates made from lignocellulosic and macroalgae biomasses. The concentration of hydrolysates was 2.5% (v/v). Standard deviations are presented as error bars.
Figure 4.
End product formation from hydrolysates made from lignocellulosic and macroalgae biomasses. The concentration of hydrolysates was 2.5% (v/v). Standard deviations are presented as error bars.
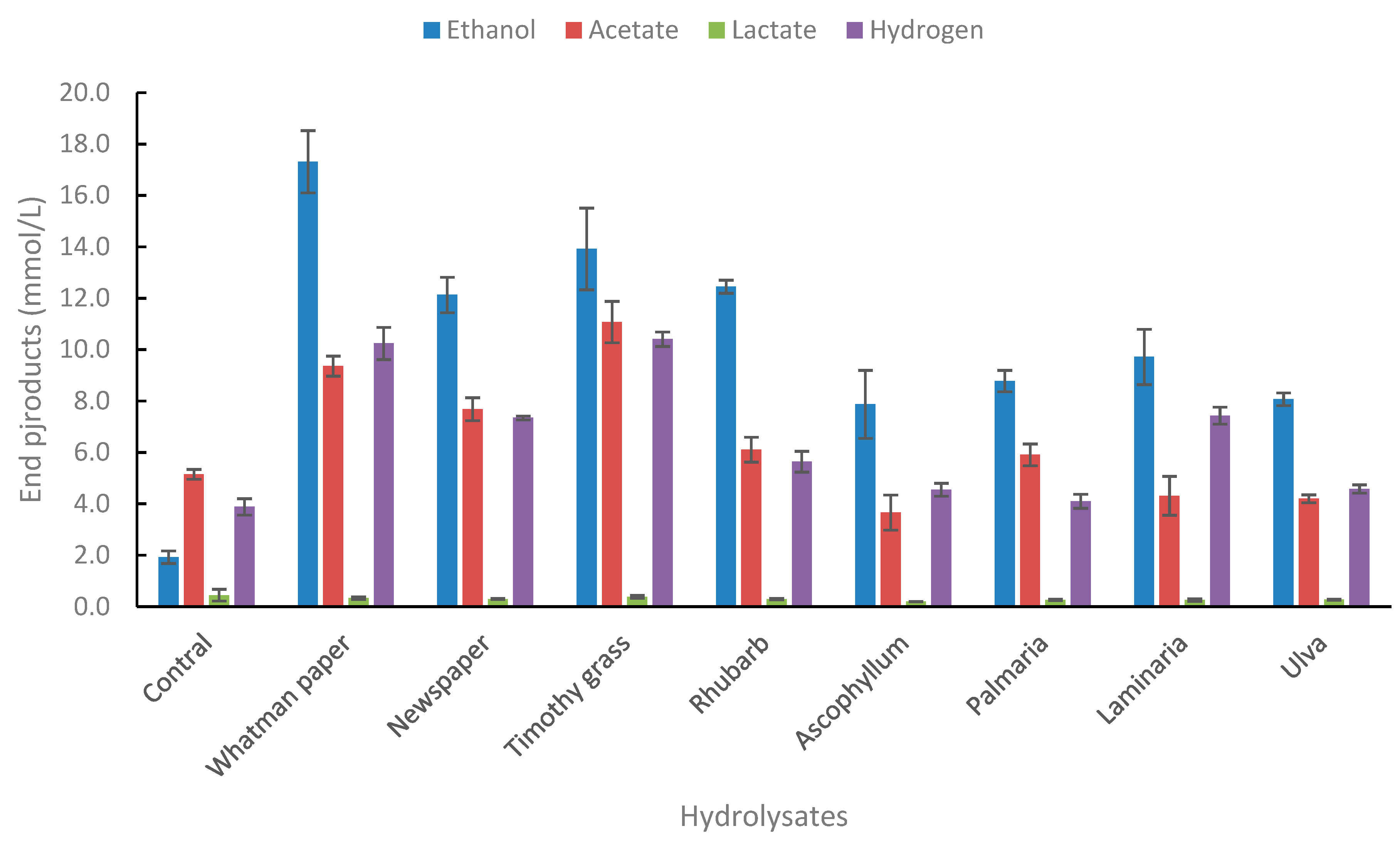
Figure 6.
End product formation from casein, collagen and gelatin. Concentration is 0.2% (w/v). Standard deviations are presented as error bars.
Figure 6.
End product formation from casein, collagen and gelatin. Concentration is 0.2% (w/v). Standard deviations are presented as error bars.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



