
Submitted:
15 December 2023
Posted:
15 December 2023
You are already at the latest version
Abstract
Non-alcoholic fatty liver disease (NAFLD) has emerged as a significant liver ailment attributed to factors like obesity and diabetes. While ongoing research explores treatments for NAFLD, further investigation is imperative to address this escalating health concern. NAFLD manifests as hepatic steatosis, precipitating insulin resistance and metabolic syndrome. This study aims to validate the regenerative potential of chimeric fibroblast growth factor 21 (FGF21)-Hepatocyte Growth Factor Receptor (HGFR) in NAFLD-afflicted liver cells. AML12, a murine hepatocyte cell line, was utilized to gauge the regenerative effects of chimeric FGF21/HGFR expression. Polysaccharide accumulation was affirmed through periodic acid-Schiff (PAS) staining, while LDL uptake was microscopically observed with labeled LDL. mRNA analysis via RT-PCR revealed reduced expression of key NAFLD markers, including acetyl-CoA carboxylase 1 (ACC1) and sterol regulatory element-binding protein (SREBP) cleavage-activating protein (SCAP), alongside elevated levels of hepatocyte growth factor (HGF), hepatocyte nuclear factor 4 alpha (HNF4A), and albumin (ALB). These findings affirm the hepatoregenerative properties of chimeric FGF21/HGFR within AML12 cells, opening novel avenues for NAFLD therapeutic exploration.
Keywords:
keyword 1 alpha mouse liver 12(AML12)
; fibroblast growth factor 21 (FGF21)
; Hepatocyte Growth Factor Receptor (HGFR)
; chimeric FGF21/HGFR
; liver regeneration
1. Introduction
Non-alcoholic fatty liver disease (NAFLD) has become one of the leading causes of chronic liver disease globally, with its prevalence reaching epidemic proportions [1,2]. NAFLD encompasses a spectrum of liver pathologies, ranging from simple hepatic steatosis, characterized by excessive fat accumulation in hepatocytes, to non-alcoholic steatohepatitis (NASH), which involves inflammation, hepatocyte injury, and fibrosis [3]. The increasing prevalence of NAFLD is closely associated with obesity, insulin resistance, metabolic syndrome, and sedentary lifestyles [4]. Consequently, the therapeutic demand for NAFLD and its progressive forms has never been greater.
A liver affected by NAFLD due to various causes causes various metabolic problems compared to a normal liver [5]. The problem is that the fatty liver caused by NAFLD has insulin resistance properties, so insulin cannot affect liver cells, which causes problems with blood sugar control and fat absorption, as well as protein production and breakdown in liver cells [6]. In addition to these problems, fatty liver cells in NAFLD have a reduced ability to regenerate into normal liver cells, which can lead to other liver diseases. If the condition becomes more severe, it can lead to liver cancer [7]. Because NAFLD is a disease that occurs in the liver that can lead to serious consequences over time, research into treatments to regenerate the liver is needed before this process progresses.
The liver's regenerative capacity is a vital defense mechanism that allows it to respond to injuries and maintain functional integrity [8]. However, in the context of NAFLD, the regenerative capacity of hepatocytes is often compromised, leading to impaired liver regeneration and potential progression to more severe liver diseases [9]. Therefore, developing innovative strategies to enhance liver regeneration in NAFLD patients has become a significant research focus in hepatology and regenerative medicine.
Fibroblast Growth Factor 21 (FGF21) and Hepatocyte Growth Factor Receptor (HGFR, also known as c-Met) have emerged as promising therapeutic targets in liver regeneration research [10,11]. FGF21, a member of the fibroblast growth factor superfamily, has shown potent metabolic regulatory effects, including its ability to improve lipid metabolism and insulin sensitivity [12]. Importantly, recent studies have unveiled additional roles for FGF21 in tissue repair and regeneration beyond its metabolic functions [13]. HGFR, a receptor tyrosine kinase, is a critical player in hepatocyte proliferation, migration, and tissue repair processes during liver regeneration [14].
The concept of chimeric fusion proteins combining FGF21 and HGFR has introduced a novel therapeutic approach to harness the regenerative potential of these two molecules in a synergistic manner. The chimeric FGF21/HGFR fusion protein holds the promise of addressing both the metabolic imbalances associated with NAFLD and the impaired liver regeneration observed in these patients.
The objective of this thesis is to investigate and confirm the regeneration of non-alcoholic fatty liver cells using chimeric FGF21/HGFR as a novel therapeutic approach. By exploring the molecular mechanisms underlying liver regeneration, the specific roles of FGF21 and HGFR in tissue repair, and their potential synergistic effects as a chimeric fusion protein, we aim to shed light on its efficacy and translational potential as a therapeutic intervention for NAFLD.
2. Results
2.1. Construction of Stable AML12 Cell Line for Expression of chimeric FGF21/HGFR
In order to study the functionality of chimeric FGF21/HGFR, it is necessary to insert the vector into cells and express gene. For the expression of chimeric FGF21/HGFR in AML12, the sequence encoding FGF21/HGFR was cloned into the pcDNA3.1 expression vector. This vector was designed to include the GFP gene to confirm for expression (Figure 1).
2.2. FGF21/HGFR on AML12 cells are closely related glycogen accumulation ability.
AML12 (alpha mouse liver 12) cells are hepatocytes isolated from the normal liver. To confirm the recovery of hepatocyte function in AML12, the cells were transfected with chimeric FGF21/HGFR and PAS staining confirmed that glycogen was accumulated in AML12 expressing chimeric FGF21/HGFR. In order to confirm glycogen expression in AML12 transfected with chimeric FGF21/HGFR, Schiff's reagent was used to stain the glycogen inside the cells, and cell nuclei were stained using Mayer's Hematoxylin. In AML12 cells expressing chimeric FGF21/HGFR, it was confirmed that magenta-stained glycogen was produced, and in the ctrl group, glycogen was not observed, as shown in Figure 2
2.3. FGF21/HGFR promotes LDL uptake on AML12 cells.
This study confirmed the liver regeneration ability of chimeric FGF21/HGFR at the cell level through LDL uptake and regulation. AML12 was transformed with chimeric FGF21/HGFR and confirmed through fluorescence microscopy. To observe chimeric FGF21/HGFR induced lipid accumulation and lipid uptake, AML12 cells treated with 10 ug/ml of labeled LDL for 3 hours were observed under a fluorescence microscope. We confirmed that AML12 uptakes a large amount of LDL in the FGF21/HGFR group compared to the control group, and these data suggest that the function of hepatocytes in AML12 was enhanced (Figure 3).
2.4. Evaluation of the expression for hepatocyte function-related genes.
The induction of NAFLD is closely related to free fatty acids, of which saturated fatty acids, of which palmitate is an important component. RT-PCR was performed to confirm regeneration at the mRNA level when chimeric FGF21/HGFR was involved in AML12 by treating palmitate. The results showed a significant decrease in Acetyl-CoA carboxylase 1 (Acc1) (Figure 4A) and Sterol regulatory element binding protein (SREBP) cleavage activating protein (Scap) at the mRNA level in AML12 transfected with FGF21/HGFR compared to the control group in the NAFLD state, and upregulation of Hepatocyte growth factor (Hgf), and Albumin (Figure 4B). These data indicate that chimeric FGF21/HGFR may be helpful in studying hepatocyte proliferation and liver regeneration mechanisms in NAFLD.
3. Disscusion
The liver, a highly metabolic organ, possesses remarkable regenerative capabilities. However, the surge in obesity rates has led to an alarming increase in non-alcoholic fatty liver disease (NAFLD) [15]. NAFLD's impact on liver regenerative capacity poses a pressing concern, prompting our investigation into the regenerative potential of chimeric fibroblast growth factor 21 (FGF21)/Hepatocyte Growth Factor Receptor (HGFR) within the context of NAFLD. This study aims to unravel its effects on hepatocyte function and gene expression, with the ultimate goal of shedding light on novel avenues for NAFLD treatment.
Before delving into the core experiments, we meticulously confirmed the expression of chimeric FGF21/HGFR within AML12 cells. The insertion of a GFP tag into pcDNA3.1 and subsequent fluorescent gene expression in AML12 cells enabled us to visually affirm the co-expression of chimeric FGF21/HGFR. This preliminary step ensured that subsequent experiments were conducted exclusively on cells displaying fluorescence, establishing a robust foundation for our investigation.
Glycogen synthesis serves as a vital marker of hepatocyte function, with profound implications for liver regeneration [16]. Our investigation, employing the periodic acid-Schiff (PAS) staining assay, revealed a striking contrast between control and chimeric FGF21/HGFR-expressing cells. The notable accumulation of glycogen in the latter signifies an enhanced capacity for glucose conversion, pointing toward substantial functional improvement induced by chimeric FGF21/HGFR.
Effective LDL uptake stands as a pivotal indicator of hepatocyte function [17]. Our findings in the LDL uptake assay corroborate the hypothesis of functional enhancement attributed to liver regeneration. In contrast to control cells, those expressing chimeric FGF21/HGFR exhibited a substantial increase in LDL uptake. This heightened capacity for lipoprotein metabolism underscores the potential of chimeric FGF21/HGFR to alleviate lipid dysregulation associated with NAFLD.
To comprehensively characterize liver regeneration and growth in AML12 cells under the influence of chimeric FGF21/HGFR, we scrutinized mRNA levels of key markers. These included Acc1 and Scap, associated with lipid metabolism, whose expressions were notably influenced by both NAFLD induction and chimeric FGF21/HGFR expression. Furthermore, our analysis unveiled upregulation of hepatocyte growth factor (HGF) and Albumin, suggesting enhanced hepatocyte functionality and liver regeneration potential, in concurrence with previous NAFLD research findings.
The findings of this study align closely with existing NAFLD research, bolstering the notion that chimeric FGF21/HGFR holds promise in improving liver regeneration function in NAFLD-afflicted AML12 cells [18-21]. These observations not only provide a deeper understanding of NAFLD but also open doors to novel perspectives in liver regeneration treatment.
In conclusion, this study underscores the regenerative potential of chimeric FGF21/HGFR in the context of NAFLD, offering promising prospects for therapeutic interventions. By enhancing hepatocyte function and influencing gene expression patterns, chimeric FGF21/HGFR emerges as a potential candidate for further exploration in the quest to address NAFLD and advance liver regeneration research.
4. Materials and Methods
4.1. Construction of chimeric FGF21/HGFR receptor
To generate the chimeric FGF21/HGFR receptor construct, a synthetic HGFR sequence encompassing the intracellular domain and transmembrane domain from amino acid (aa) 251 to 712 of HGFR including signal peptide sequence was synthesized (Genotech, Daejeon, Korea). This sequence was then inserted into the pCDNA3.1 expression vector (Invitrogen) using the restriction enzymes NheI and XhoI. In addition, a synthetic FGF21 sequence, encoding the full length protein from aa 28 to 208 of FGF21 including green fluorescent protein GFP (238 aa) was produced from Genotech (Daejeon, Korea) was fused to the transmembrane domain of HGFR using the restriction enzymes BglII and BamHI.
4.2. Cell culture and Treatment
AML12 cells were cultured in Dulbecco Modified Eagle Medium/F12 (DMEM/F12) (Gibco, #11320033) containing 10% (v/v) heat-inactivated Fetal bovine serum (Corning, 35-015-CV), 1% Insulin-Transferrin-selenium 100X (Gibco, #41400-045) and 40ng/ml Dexamethasone (Sigma-Aldrich, D4902-25MG) at 37 °C in a humidified atmosphere with 5% CO2.
For transfection, AML12 cells are seeded in 6-well plate (SPL, #30006). at 8*104 cells/ml overnight. After that, they were transfected with chimeric FGF21/HGFR DNA and control DNA. This transfection assay was performed according to the Lipofectamine 3000 (Invitrogen, #L3000001) protocol.
4.3. Periodic acid-schiff (PAS) staining
In the control group and FGF21 group (transfected chimeric FGF21/HGFR), the AML12 were seeding at a density of 3*105 cells/well and incubated for overnight at 37 °C. The medium was removed from the plate, cells rinsed with PBS two times and fixed with 4% paraformaldehyde (Tech-innovation, #BPP-9004-001L) for 10 min at room temperature, washed 2 times with PBS, oxidized for 5 min with 1% periodic acid, and washed 2 times PBS. Cells were then stained with Schiff’s reagent (Abcam, #395B) for 15 min, washed 2 times with PBS. Mayer's Hematoxylin for 2 min and washed 3 times with PBS before microscopic examination.
4.4. LDL uptake assay
This study was conducted using Image-iT™ Low Density Lipoprotein Uptake Kit, pHrodo™ Red (Invitrogen, #I34360). Seeding 30–40% confluency AML12 Cells in a confocal dish (SPL, #211350) 24h. Serum starvation media consisting of 10ml of FluoroBrite DMEM (Gibco, A1896701) and 200mg of 2% bovine serum albumin (BSA) (Solmate, #SM-BOV-100) was additionally added to the dish and incubated for 14 hours. Add 10ug/ml of Labeled LDL to the dish and incubate for 3h. Hoechst was added and incubated for 10 min to stain the nuclei, followed by PBS washing. Stained cells were observed under a microscope and fluorescence was observed.
4.5. Reverse transcription (RT)-PCR and real-time PCR
Total RNA was extracted using Ribospin™ (Geneall, #304-150) and cDNA was synthesized (Applied Biosystems, #4368814). mRNA expression of genes (Acc1, Scap, Hnf4a, Hgf, and Albumin) was examined by quantitative real-time PCR. The primer sequences of these genes used in this experiment are listed in Table 1. All real-time PCR experiments were analyzed on an ABI Step One real-time PCR system (Applied Biosystem, Foster City, USA). Each real-time PCR reaction was performed in a 20μL reaction mixture containing 0.1 μM of each primer, 10 μL of SYBR Hi-ROX (bioline, #QT605-05), and 5µL of template cDNA. The real-time PCR cycles were as follows: 1 cycle at 95˚C for 10 min (enzyme activation), 40 cycles at 95˚C for 15 sec (denaturation), 60˚C for 15 sec (annealing), and 72˚C for 30 sec (extension), 1 cycle at 95˚C for 15 sec, 60˚C for 1 min, 95˚C for 15 sec (Melt curve stage)
4.6. Statistical analysis
Experiments were performed three times each in triplicate. Results are expressed as mean ± standard deviation. Statistical analysis was done using the Student's t-test. (*p<0.05, **p<0.01, and *** p < 0.001).
5. Conclusions
In this study, we investigated the regenerative potential of chimeric FGF21/HGFR as a therapeutic strategy for non-alcoholic fatty liver disease (NAFLD). Through cell transfection and analysis, we observed that chimeric FGF21/HGFR has the effect of regenerating normal hepatocellular properties in non-alcoholic fatty liver. These findings suggest chimeric FGF21/HGFR's potential as a therapeutic intervention for NAFLD.
Author Contributions
Sung-Jun Kim performed the experiments and wrote the manuscripts; So Jung Kim, Jeongeun Hyun and Hae-won Kim performed the experiment; Jun-Hyeog Jang organized this work and contributed to writing the manuscript.
Acknowledgments
This work was supported by the National Research Foundation of Korea (NRF) grants funded by the Korean government (MSIT) (RS-2023-00208339 to J.H.J., 2021R1C1C1003904 to J.H.) and INHA UNIVERSITY Research Grant.
Conflicts of Interest
The authors have no conflicts of interest to declare.
References
- Swinburn, B.A.; Sacks, G.; Hall, K.D.; et al. The global obesity pandemic: shaped by global drivers and local environments. Lancet 2011, 378, 804-814. [CrossRef]
- Younossi, Z.M.; Koenig, A.B.; Abdelatif, D.; Fazel, Y.; Henry, L.; Wymer, M. Global epidemiology of nonalcoholic fatty liver disease—Meta-analytic assessment of prevalence, incidence, and outcomes. Hepatology 2016, 64, 73-84. [CrossRef]
- European Association for the Study of the Liver (EASL); European Association for the Study of Diabetes (EASD); European Association for the Study of Obesity (EASO). EASL-EASD-EASO Clinical Practice Guidelines for the Management of Non-Alcoholic Fatty Liver Disease. Obes Facts 2016, 9, 65-90. [CrossRef]
- Halmos, T.; Suba, I. A nem alkoholos zsírmáj mint a metabolikus szindróma komponense és kauzális kapcsolatai egyéb kórképekkel [Non-alcoholic fatty liver disease, as a component of the metabolic syndrome, and its causal correlations with other extrahepatic diseases]. Orv Hetil 2017, 158, 2051-2061. [CrossRef]
- Zeigerer, A. NAFLD - A rising metabolic disease. Mol Metab. 2021, 50, 101274. [CrossRef]
- Tanase, D.M.; Gosav, E.M.; Costea, C.F.; et al. The Intricate Relationship between Type 2 Diabetes Mellitus (T2DM), Insulin Resistance (IR), and Nonalcoholic Fatty Liver Disease (NAFLD). J Diabetes Res 2020, 2020, 3920196. [CrossRef]
- He, X.L.; Chen, J.Y.; Feng, Y.L.; et al. Single-cell RNA sequencing deciphers the mechanism of sepsis-induced liver injury and the therapeutic effects of artesunate. Acta Pharmacol Sin 2023, 44, 1801-1814. [CrossRef]
- Angulo, P.; Kleiner, D.E.; Dam-Larsen, S.; et al. Liver Fibrosis, but No Other Histologic Features, Is Associated With Long-term Outcomes of Patients With Nonalcoholic Fatty Liver Disease. Gastroenterology 2015, 149, 389-397.e10. [CrossRef]
- Markan, K.R.; Naber, M.C.; Ameka, M.K.; et al. Circulating FGF21 is liver derived and enhances glucose uptake during refeeding and overfeeding. Diabetes 2014, 63, 4057-4063. [CrossRef]
- Drescher, H.K.; Schumacher, F.; Schenker, T.; et al. c-Met Signaling Protects from Nonalcoholic Steatohepatitis- (NASH-) Induced Fibrosis in Different Liver Cell Types. Oxid Med Cell Longev 2018, 2018, 6957497. [CrossRef]
- Falamarzi, K.; Malekpour, M.; Tafti, M.F.; Azarpira, N.; Behboodi, M.; Zarei, M. The role of FGF21 and its analogs on liver associated diseases. Front Med (Lausanne) 2022, 9, 967375. [CrossRef]
- Attia, S.L.; Softic, S.; Mouzaki, M. Evolving Role for Pharmacotherapy in NAFLD/NASH. Clin Transl Sci 2021, 14, 11-19. [CrossRef]
- Yang, C.; Lu, W.; Lin, T.; et al. Activation of Liver FGF21 in hepatocarcinogenesis and during hepatic stress. BMC Gastroenterol 2013, 13, 67. [CrossRef]
- Baier, P.; Wolf-Vorbeck, G.; Hempel, S.; Hopt, U.T.; von Dobschuetz, E. Effect of liver regeneration after partial hepatectomy and ischemia-reperfusion on expression of growth factor receptors. World J Gastroenterol 2006, 12, 3835-3840. [CrossRef]
- Ma, C.; Avenell, A.; Bolland, M.; Hudson, J.; Stewart, F.; Robertson, C.; et al. Effects of weight loss interventions for adults who are obese on mortality, cardiovascular disease, and cancer: systematic review and meta-analysis. BMJ 2017, 359, j4849. [CrossRef]
- Roach, P.J.; Depaoli-Roach, A.A.; Hurley, T.D.; Tagliabracci, V.S. Glycogen and its metabolism: some new developments and old themes. Biochem J 2012, 441, 763-787. [CrossRef]
- Perla, F.M.; Prelati, M.; Lavorato, M.; Visicchio, D.; Anania, C. The Role of Lipid and Lipoprotein Metabolism in Non-Alcoholic Fatty Liver Disease. Children (Basel) 2017, 4, 46. [CrossRef]
- Higuchi, N.; Kato, M.; Shundo, Y.; et al. Liver X receptor in cooperation with SREBP-1c is a major lipid synthesis regulator in nonalcoholic fatty liver disease. Hepatol Res 2008, 38, 1122-1129. [CrossRef]
- Moon, Y.A. The SCAP/SREBP Pathway: A Mediator of Hepatic Steatosis. Endocrinol Metab (Seoul) 2017, 32, 6-10. [CrossRef]
- Pan, X.; Zhang, Y. Hepatocyte Nuclear Factor 4α in the Pathogenesis of Non-Alcoholic Fatty Liver Disease. Chin Med J (Engl) 2022, 135, 1172-1181. [CrossRef]
- Sun, L.; Wang, Q.; Liu, M.; et al. Albumin binding function is a novel biomarker for early liver damage and disease progression in non-alcoholic fatty liver disease. Endocrine 2020, 69, 294-302. [CrossRef]
Figure 1.
Construction of chimeric FGF21/HGFR receptor. For the expression of chimeric FGF21/HGFR in AML12, the sequence encoding FGF21/HGFR was cloned into the pcDNA3.1 expression vector.
Figure 1.
Construction of chimeric FGF21/HGFR receptor. For the expression of chimeric FGF21/HGFR in AML12, the sequence encoding FGF21/HGFR was cloned into the pcDNA3.1 expression vector.
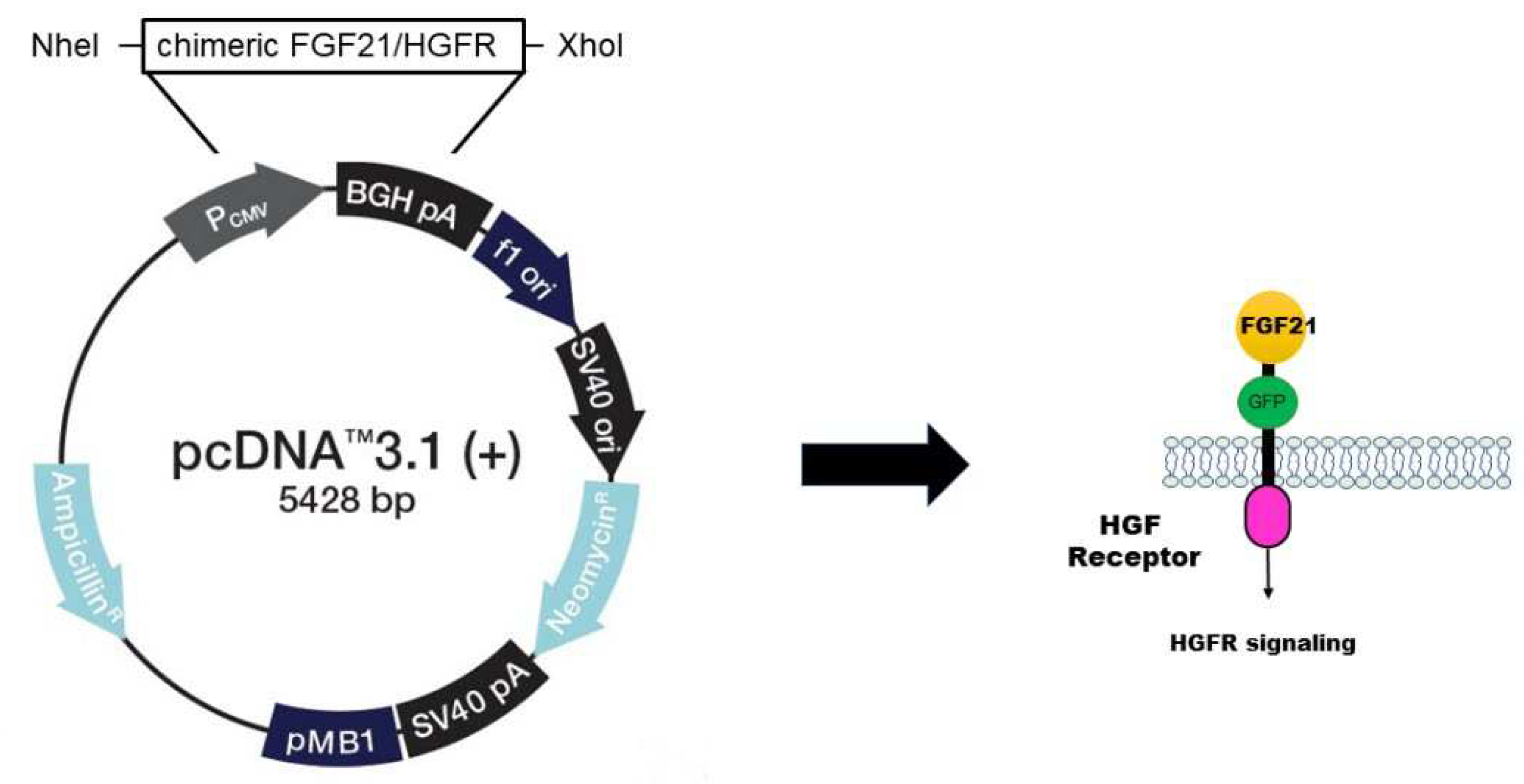
Figure 2.
Glycogen accumulation capacity was increased in chimeric FGF21/HGFR on AML12. (A, C) The GFP gene was expressed in AML12 and observed by fluorescence. (B, D) Accumulation of glycogen was observed through a microscope.
Figure 2.
Glycogen accumulation capacity was increased in chimeric FGF21/HGFR on AML12. (A, C) The GFP gene was expressed in AML12 and observed by fluorescence. (B, D) Accumulation of glycogen was observed through a microscope.
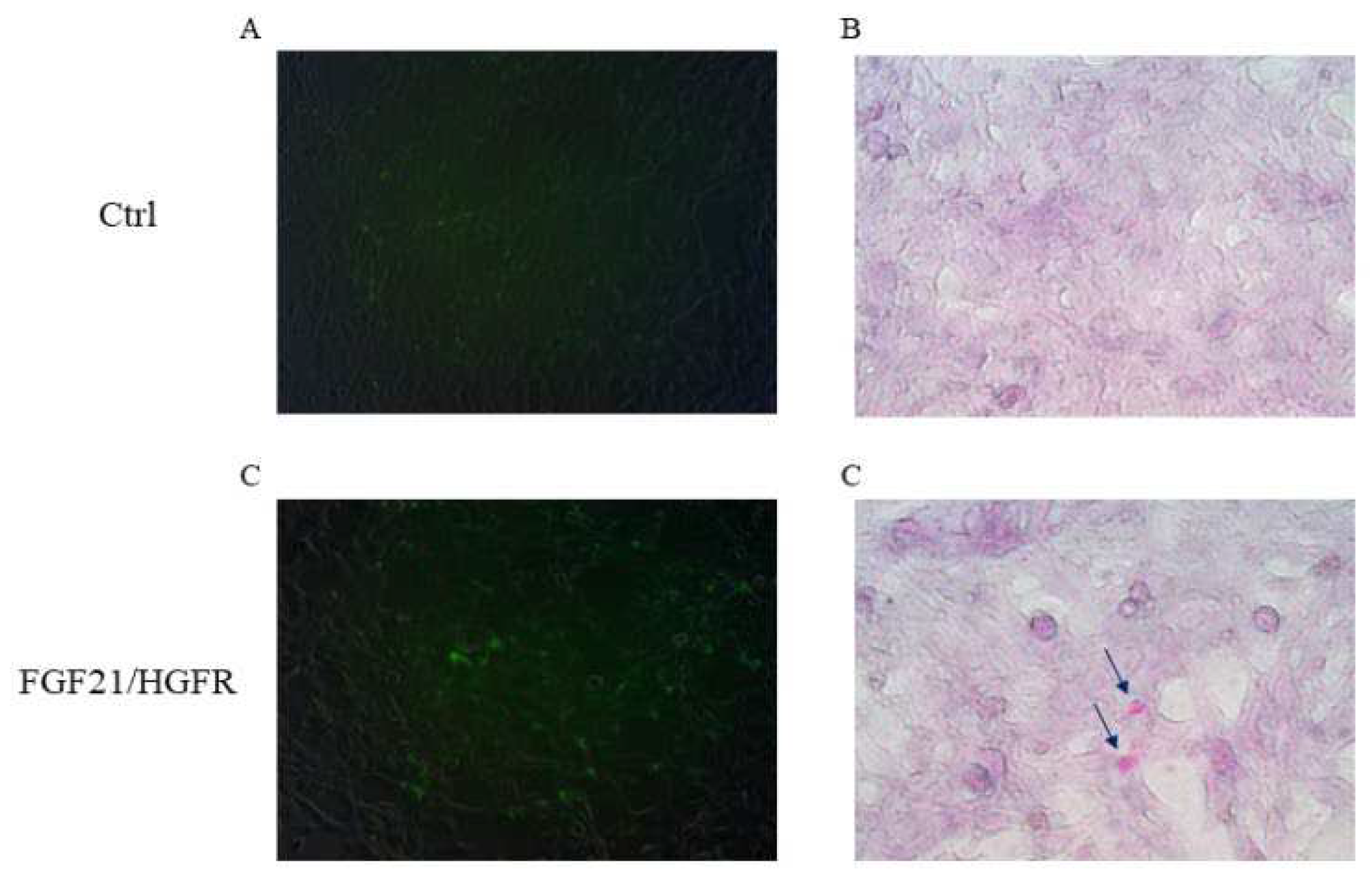
Figure 3.
LDL uptake was upregulated in chimeric FGF21/HGFR on AML12. The AML12 cells with labeled-LDL were detected by immunofluorescence staining.
Figure 3.
LDL uptake was upregulated in chimeric FGF21/HGFR on AML12. The AML12 cells with labeled-LDL were detected by immunofluorescence staining.
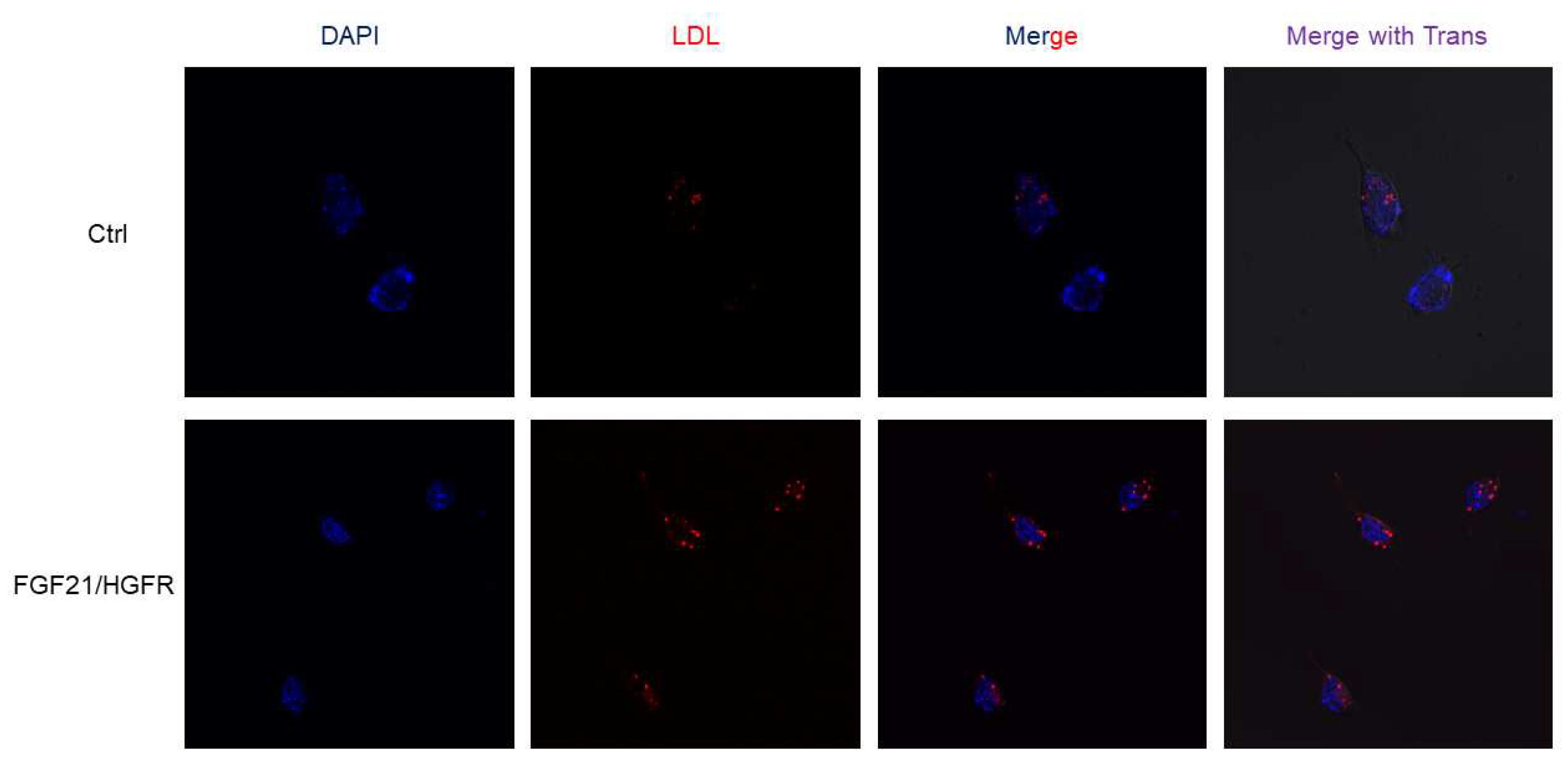
Figure 4.
In AML12 where steatosis was induced by palmitate, chimeric FGF21/HGFR showed an indicator of liver regeneration. (A) The mRNA levels of Acc1 and Scap were analyzed by real-time PCR, and the comparative mRNA levels of each gene were assessed relative to the mRNA levels of the housekeeping gene, Gapdh. (B) The mRNA levels of Hnf4a, Hgf, and Albumin were analyzed by real-time PCR, and the comparative mRNA levels of each gene were assessed relative to the mRNA levels of the housekeeping gene, Gapdh.
Figure 4.
In AML12 where steatosis was induced by palmitate, chimeric FGF21/HGFR showed an indicator of liver regeneration. (A) The mRNA levels of Acc1 and Scap were analyzed by real-time PCR, and the comparative mRNA levels of each gene were assessed relative to the mRNA levels of the housekeeping gene, Gapdh. (B) The mRNA levels of Hnf4a, Hgf, and Albumin were analyzed by real-time PCR, and the comparative mRNA levels of each gene were assessed relative to the mRNA levels of the housekeeping gene, Gapdh.
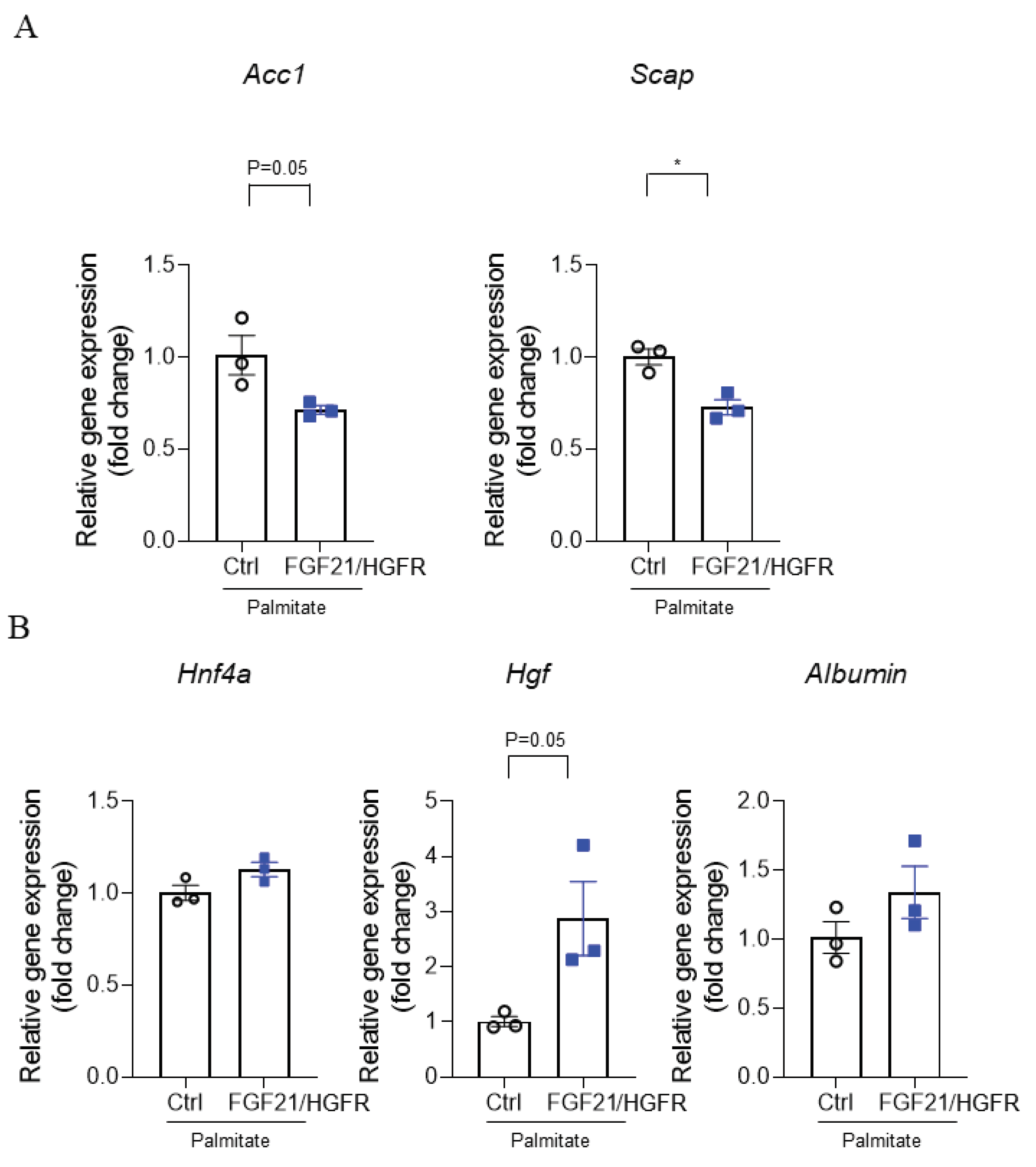

Table 1.
Sequence of primer used for quantitative real-time PCR.
| Genes | Forward primer | Reverse primer |
|---|---|---|
| GAPDH | 5'-CTGCACCACCAACTGCTTAG-3' | 5'-GTCTTCTGGGTGGCAGTGAT-3' |
| Hnf4a | 5'-TGCGAACTCCTTCTGGATGACC-3' | 5'-CAGCACGTCCTTAAACACCATGG-3' |
| Albumin | 5'-CAGTGTTGTGCAGAGGCTGACA-3' | 5'-GGAGCACTTCATTCTCTGACGG-3' |
| Scap | 5'-AGAATTCCACAGGTCCCGTT-3' | 5'-CTGCGCATCCTATCCAATTC-3' |
| Acc1 | 5'-TGACAGACTGATCGCAGAGAAAG-3' | 5'-TGGAGAGCCCCACACACA-3' |
| Hgf | 5'-GTCCTGAAGGCTCAGACTTGGT-3' | 5'-CCAGCCGTAAATACTGCAAGTGG-3' |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



