
Submitted:
16 December 2023
Posted:
18 December 2023
You are already at the latest version
Abstract
Background: Although cerebral aneurysm (CA) is a defining complication of COL4A1/2-related vasculopathy, the specific factors influencing its onset remain uncertain. This study aimed to identify and analyze these factors. Methods: We described a family presenting with a novel variant in the COL4A1 gene complicated with CA. Concurrently, an exhaustive review of previously documented patients with COL4A1/2-related vasculopathy was conducted by sourcing data from PubMed, Web of Science, Google Scholar, and Ichushi databases. We compared the variant types and locations between patients with CA (positive group) and those without CA (negative group). Results: This investigation encompassed 53 COL4A1/2 variants in 76 patients. Except for one start codon variant, all the identified variants in CA were missense variants. Otherwise, CA was not associated with other clinical manifestations, such as small-vessel disease or other large-vessel abnormalities. A higher frequency of missense variants (95.5% vs 58.1%, p = 0.0035) was identified in the CA positive group. Conclusions: CA development appears to necessitate qualitative alterations in COL4A1/2, and the underlying mechanism seems independent of small vessel disease or other large vessel anomalies. Our findings suggest that a meticulous evaluation of CA is necessary when missense variants in COL4A1/2 are identified.
Keywords:
COL4A1
; COL4A2
; cerebral aneurysm
; small vessel disease
; large vessel abnormality
1. Introduction
Pathogenic variants are critical causes of juvenile small-vessel strokes and can manifest as large-vessel diseases beyond small-vessel disorders. Pathogenic variants in Collagen type IV alpha 1 (COL4A1) and alpha 2 (COL4A2) are recognized causes of small-vessel strokes, particularly intracerebral hemorrhage. Furthermore, COL4A1/2 variants can manifest as various clinical manifestations, including retinal arterial tortuosity (RAT), kidney disease, and muscle cramps [1,2]. Cerebral aneurysm (CA) is another notable clinical feature of COL4A1/2-related vasculopathy that requires careful assessment and follow-up in clinical contexts. The prevalence of CA is remarkably high; previous reviews have shown that 44.4% of patients with COL4A1/2 variants had CA, and 50.0% of them displayed multiple CAs. The most common site for CA is the internal carotid artery (ICA) [3], whereas CAs in locations other than the ICA are rare.
Several characteristic variants, such as substitution of glycine with other amino acids (AAs) in the triplet sequence of glycine (Gly-X-Y) or variants in the collagenous domain, have been reported to be associated with COL4A1/2-related vasculopathy [4]. Additionally, mouse models have demonstrated that clinical severity is influenced by the dosage effects of glycine substitutions in the triplet sequence [5]. Despite these insights, the genotype-phenotype correlation of CA in patients with COL4A1/2 variants remains unclear. Large vessel abnormalities other than CA have also been reported in patients with COL4A1/2-related vasculopathy [1,6]; however, the relationship between CA and those large vessel abnormalities in COL4A1/2 variants is yet to be clearly defined.
In this report, we present a family member carrying a novel variant in COL4A1/2 who exhibited multiple CAs accompanied by vertebrobasilar dolichoectasia. Through an extensive literature search, we investigated the genotype-phenotype correlation among CA, small vessel disease, and large vessel abnormalities in patients with COL4A1/2 variants.
2. Materials and Methods
- Genetic tests and exome sequencing
This study was approved by the ethics board of Niigata University (approval numbers 801, 802, and G2020-0032). Written informed consents were obtained from the patient (proband) and her mother. Genetic testing and exome sequencing (ES) were performed as described previously [7]. Briefly, genomic DNA was extracted from the proband's blood, and Sanger sequencing analysis was performed for NOTCH3 and HTRA1. After obtaining negative results for NOTCH3 and HTRA1, we used ES to investigate genes associated with monogenic small vessel disease. ES analysis was outsourced to Macrogen (Korea). Additionally, we used Sanger's method on the genomic DNA of the proband's mother, using a forward primer (5'-TGGGGGCCAAACACAATTTC-3') and a reverse primer (5'-GCGTTTCCCATCACCTCAAAC-3').
- Literature search
We conducted a comprehensive literature search on PubMed and Web of Sciences until February 2023, focusing on publications and conference abstracts related to pathogenic variants in the COL4A1/2 genes. Text words, phrases, and Medical Subject Headings corresponding to monogenic syndromes/diseases associated with variants in COL4A1/2 genes were used for the search (Supplementary materials). We also explored relevant publications using Google Scholar (https://scholar.google.jp/) and Ichushi, a Japanese publication database (https://login.jamas.or.jp/). All available publications, including those that only offered abstracts without the full text, were examined. We did not restrict our search to publications in English. For publications in languages other than English or Japanese, DeepL (https://www.deepl.com/translator) or ChatGPT (https://openai.com/) was used for translation. Following the initial search, we excluded studies in which the CA status was not available.
- Classifying the variants
Variants reported in the included publications as well as those identified in our patients were analyzed using PolyPhen2, SIFT, and Provean on the VaProS website (http://p4d-info.nig.ac.jp/vapros/). Additionally, the PHRED score of the combined annotation-dependent depletion (CADD) of each variant was calculated using the CADD website (https://cadd.gs.washington.edu/). The cutoff value of the PHRED score was defined as 20 [8]. Furthermore, if variants were involved in a splicing site, additional computational analysis was performed using SpliceAI [9] and MaxEntScan (MES) [10]. The cut-off values of the spliceDelta score in SpliceAI and the %variance of MES were defined as 0.5 and 15, respectively. The %variance of MES reflects the difference between the wild-type and mutant MES, expressed as a percentage of wild-type MES [11]. A splice-site pathogenic variant is defined as pathogenic when both the %variance of MES and spliceDelta score exceed the threshold value. The frequency of each variant was determined using the Genome Aggregation Database (gnomAD) (https://gnomad.broadinstitute.org/). We also evaluated the pathogenicity of the identified variants by using ClinVar (https://www.ncbi.nlm.nih.gov/clinvar/). The pathogenicity of each variant was classified according to the American College of Medical Genetics and Genomics (ACMG) standards and guidelines [12]. Variants classified as benign or likely benign according to ACMG criteria were excluded from further analysis.
Finally, to explore the three-dimensional locations of identified mutation in the probands, we generated 3D models of COL4A1 using alphafold2 software [13]. This model was employed to create images demonstrating the locations of variants specific to each group using PyMOL software, version 2.5.0 (Schrodinger, LLC, New York, NY, USA).
- Clinical assessment and comparison between CA positive and negative variants
To elucidate the factors associated with CA in the context of COL4A1/2 vasculopathy, we segregated the variants into CA-positive and -negative groups. Subsequently, we compared the frequencies of the variant types and domains between the two groups. Furthermore, previous reports have demonstrated that substitutions in charged (Arg, Asp, Glu, and Lys) (https://proteinstructures.com/sequence/amino-acids/) or branched-chain AAs (Val, Leu, and Ile) [14] influence the clinical manifestations of COL4A1/2-related vasculopathy [4]. Therefore, we compared the frequencies of variants that were substituted with charged/branched-chain AAs.
Second, we explored whether the CA-positive phenotype correlated with other COL4A1/2-related phenotypes, including other large vessel abnormalities. Patients from the literature and our own were divided into two groups according to their CA status. We compared the clinical features and imaging findings between the two groups. Signs specific to hereditary angiopathy with nephropathy, aneurysms, and muscle cramps (HANAC) syndrome (OMIM: #611773), including RAT, muscle cramps, and nephropathy (hematuria or decreased renal function), were also investigated.
- Statistics
Statistical analyses were conducted using the MATLAB R2021a software (9.10.0.01739362) (MathWorks, Inc., Natick, MA, USA). Continuous variables, such as age at stroke onset, were compared using Wilcoxon rank-sum tests because of their non-normal distribution and unequal variance. Fisher’s exact test or Chi-squared tests was used to compare frequencies of variables, such as neurological symptoms/signs. Statistical significance was defined as P < 0.05. In instances where information was unavailable, such data were excluded from the statistical analysis.
3. Results
3.1. Case Report
The proband (III-1, Figure 1) had no vascular risk factors, such as hypertension, diabetes or smoking, but had migraine without aura. Upon awakening one morning at 32 years of age, she noticed difficulty in moving her right upper and lower extremities, and salivation from the right corner of her mouth. As there was no clear improvement in her symptoms, she visited Tane General Hospital the following day. Her blood pressure and heart rate were 115/44 mm Hg and 72 beats/min, respectively. Neurological examination revealed right facial palsy, dysarthria, and clumsiness of the right hand. Diffusion-weighted imaging (DWI) on the brain magnetic resonance imaging (MRI) revealed acute infarction in the left posterior limb of the internal capsule. Additionally, fluid-attenuated inversion recovery (FLAIR) images showed patchy periventricular white matter hyperintensities (WMHs) on each side. T2*-weighted images revealed multiple microbleeds (MBs). Brain MR angiography (MRA) and computed tomography angiography (CTA) revealed multiple CAs, including the lateral side of the basilar artery (BA) and the superior cerebellar artery (SCA), as well as vertebrobasilar dolichoectasia (Figure 1). The SCA aneurysm measured 10 × 10 × 12 mm.
The patient was admitted to our hospital for further evaluation and treatment with aspirin. Electrocardiography monitoring revealed no atrial fibrillation, and echocardiography showed no cardiac dysfunction. Laboratory test results, including those for protein C, S, and antiphospholipid antibodies, were negative. Urinalysis indicated hematuria, although renal function was normal. There were no remarkable ophthalmological findings, such as RAT. Her score on the Japanese edition of the Montreal Cognitive Assessment was 23, which was slightly lower than the normal threshold of 26. Cerebral angiography subsequently revealed a small ICA aneurysm at the C3 level (images not shown). The proband's neurological symptoms gradually improved, with only mild facial hemiparesis. The SCA aneurysm was subsequently treated with coiling,
The proband's mother (II-1, Figure 1) suffered an acute cerebellar ischemic infarction at 48 years of age, which was treated with clopidogrel. She had hypertension, but no history of diabetes, smoking or migraine. Laboratory data were not available. Brain MRI at the age of 48 revealed diffuse WMHs and a few lacunar infarctions. MRA of the brain at that time revealed a right ICA aneurysm (Figure 2).
The proband’s grandfather (I-1, Figure 1) also experienced an ischemic stroke at 54 years of age. Subsequently, he died of gastric cancer. Detailed clinical information such as medical history, laboratory data, and brain imaging was not available.
Owing to the young onset of stroke and family history, we suspected a monogenic stroke. After obtaining consent for genetic testing from the proband and her mother, we performed a sequence analysis. ES revealed a novel heterozygous variant in exon 42 of the COL4A1 gene (c.3734G>A, p.Gly1245Asp). The p.Gly1245 site is located within the glycine triplet sequence (Gly-X-Y) of the collagenous domain of COL4A1 (Figure 3). Using Sanger sequencing, we detected the same variant in the proband's mother (Figure 2).
3.2. Literature review
We conducted a comprehensive review of previous publications on COL4A1/2 variants. Initially, our search identified 470 publications from PubMed and 452 from Web of Science that matched our search terms. After eliminating duplicate publications, we added 6 publications from Ichushi and 13 from Google Scholar to our pool of resources. These publications were screened by title or abstract, and 444 publications were excluded. Subsequently, full-text screening was conducted, and 55 publications were ultimately included in our study (Supplementary Figure S1).
From these publications, 55 variants were identified. These variants were classified according to the ACMG criteria and three were found to be likely benign (p.Gln1150Lys, p.Glu1123Gly, and p.Ala1690Thr in COL4A2) [15,16,17] (Supplementary Table S3). These three variants and their corresponding publications were excluded from further analysis. Ultimately, our study included 74 patients, 52 variants from 52 publications, and two of our patients.
The variants included in this study are summarized in Supplementary Table S1 and S2. Of the 53 variants, 39 were missense variants. Other variants included three duplication/copy number variants (CNVs), two frameshift variants, four splice site variants, one start codon variant, and four point variants in the 3’-untranslated region (UTR). All four splice site variants were located at canonical splice sites, and three of the four variants were predicted to cause in-frame variants due to exon skipping [18,19,20,21].
- Characteristics of CA-positive Variants
First, we divided the included variants according to whether CA had been previously reported (Table 1 and Supplementary Table S4). Among 53 variants, CA was detected in 22. Among the variants in COL4A1, except for one start codon mutation, no variants other than missense variants were found in the CA-positive group. As compared with the CA negative group, the frequency of missense variants (94.4% vs 59.1%, p = 0.0126) and variants substituted with charged/branched-chain AAs in COL4A1 were significantly higher in the CA-positive group than in the negative group (77.8% vs. 40.9%, p = 0.0267), whereas glycine substitution variants in the triplet sequence (Gly-X-Y) was similar between the two groups (72.2% vs. 54.5%, p = 0.3319).
Although there were no significant differences in the frequency of variant types or domains due to the small sample size of the variants in COL4A2, all four CA-positive variants were located at the glycine site of the triplet sequence, in addition to charged/branched-chain AAs substitutions.
In combination with variants in COL4A1/2 and duplication/CNV, the frequencies of missense variants (95.5% vs. 58.1%, p = 0.0035), glycine substitution variants in the glycine triplet sequence (77.3% vs. 48.4%, p = 0.0475), variants substituted by charged/branched-chain AAs (81.8% vs. 38.7%, p = 0.0022), and variants in the triple-helical domain (90.9% vs. 64.5%, p = 0.0497) were significantly higher in the CA-positive group than in the negative group.
Next, we demonstrated the distribution of variants resulting in substitution with charged/branched-chain AAs in COL4A1 (Figure 4). Variants in the CA-positive group were primarily concentrated in exons 24 and 25, and then in exon 27 and 42 of COL4A1.
- Characteristics of CAs in patients with COL4A1/2 variants
The patients were categorized into positive and negative groups according to the presence of CA. Among the 76 patients with COL4A1/2 variants, 25 had CA. The age at stroke onset and frequency of family or medical history of stroke were similar in both groups. No patients with subarachnoid hemorrhage (SAH) were found in the CA-positive group, whereas one patient in the CA-negative group had a history of SAH resulting from traumatic injury [6]. Geographic distribution was similar between the two groups. In addition, imaging findings of small-vessel disease were equally similar in the two groups. The frequencies of other clinical findings such as RAT, muscle cramps, and nephropathy were also similar. However, the frequency of migraines was significantly higher in the CA-positive group (Supplementary Table S5). The proportion of modalities used to evaluate cerebral vessels was as follows for the CA-positive and the CA-negative group, respectively: MRA was 68.0% and 72.5%, CTA was 20.0% and 13.7%, and cerebral angiography was 28.0% and 15.7%.
The ICA is the most affected artery, with 86.4% of patients exhibiting ICA aneurysms. In addition to ICA, other CA are rare. A single instance of aneurysm in the middle cerebral artery (MCA) [6], anterior communicating artery (Acom) [22], posterior communicating artery (Pcom) [23], and superior cerebellar artery (SCA) were observed, and two patients had basilar artery (BA) aneurysms. The BA aneurysm was located at the top of the BA in one patient [24], whereas our proband had a CA on the lateral side of the BA in addition to SCA. All patients with CA in arteries other than the ICA carried missense variants in COL4A1, and four out of five of these variants involved substitutions of glycine with charged/branched-chain AAs in the glycine triplet sequence (Table 2). The prevalence of multiple CA was 54.5%. All identified variants in cases of multiple CA involved charged/branched-chain AA substitutions and glycine substitutions in the glycine triplet sequence (Gly498Val, Gly501Asp, Gly525Leu, Gly528Glu, Gly755Arg, Gly835Glu, Gly888Arg, Gly1245Asp in COL4A1, and Gly702Asp and Gly1389Arg in COL4A2), except for Pro116Leu in COL4A1. Notably, ten of the 12 patients had multiple ICA aneurysms.
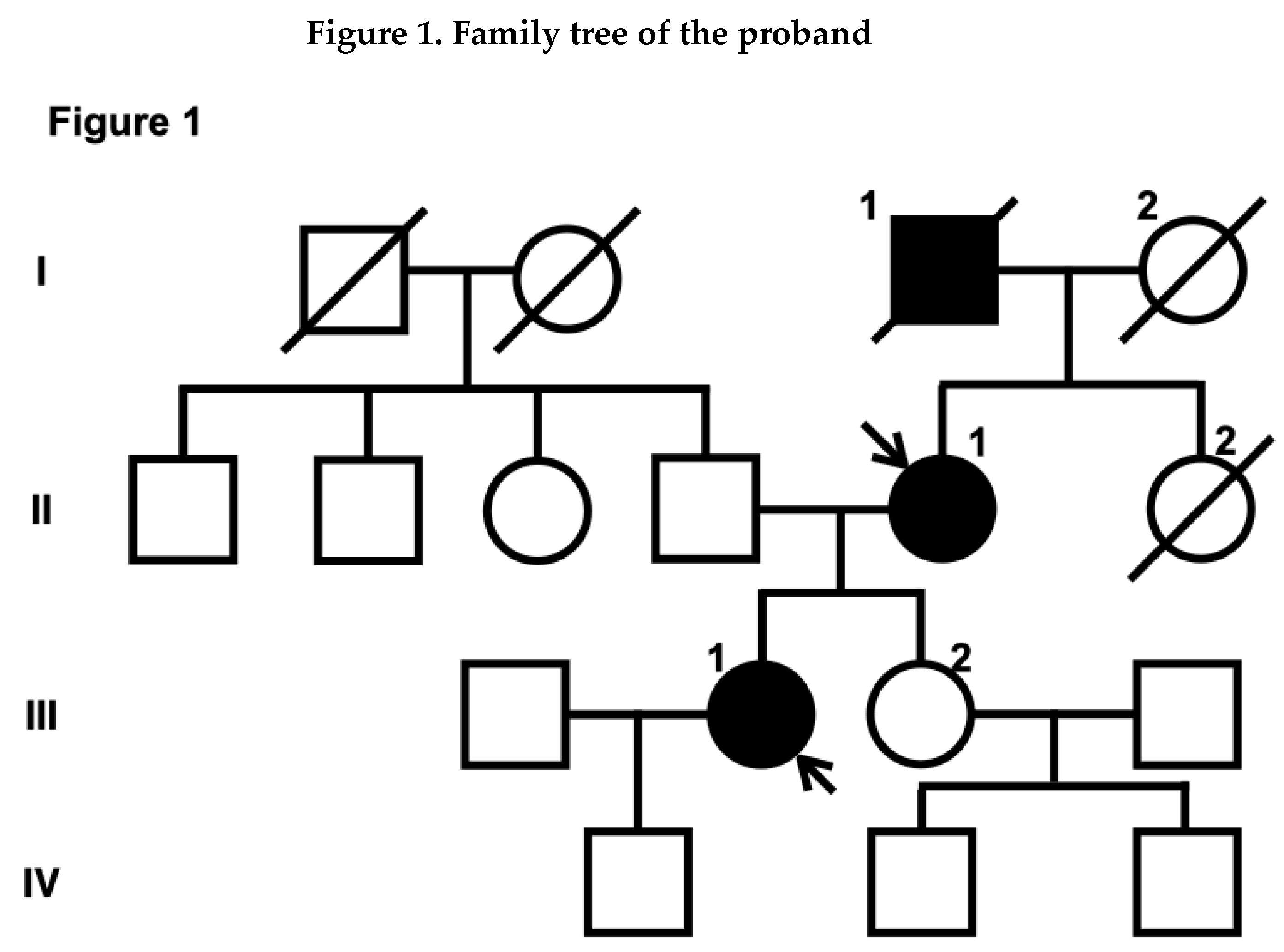
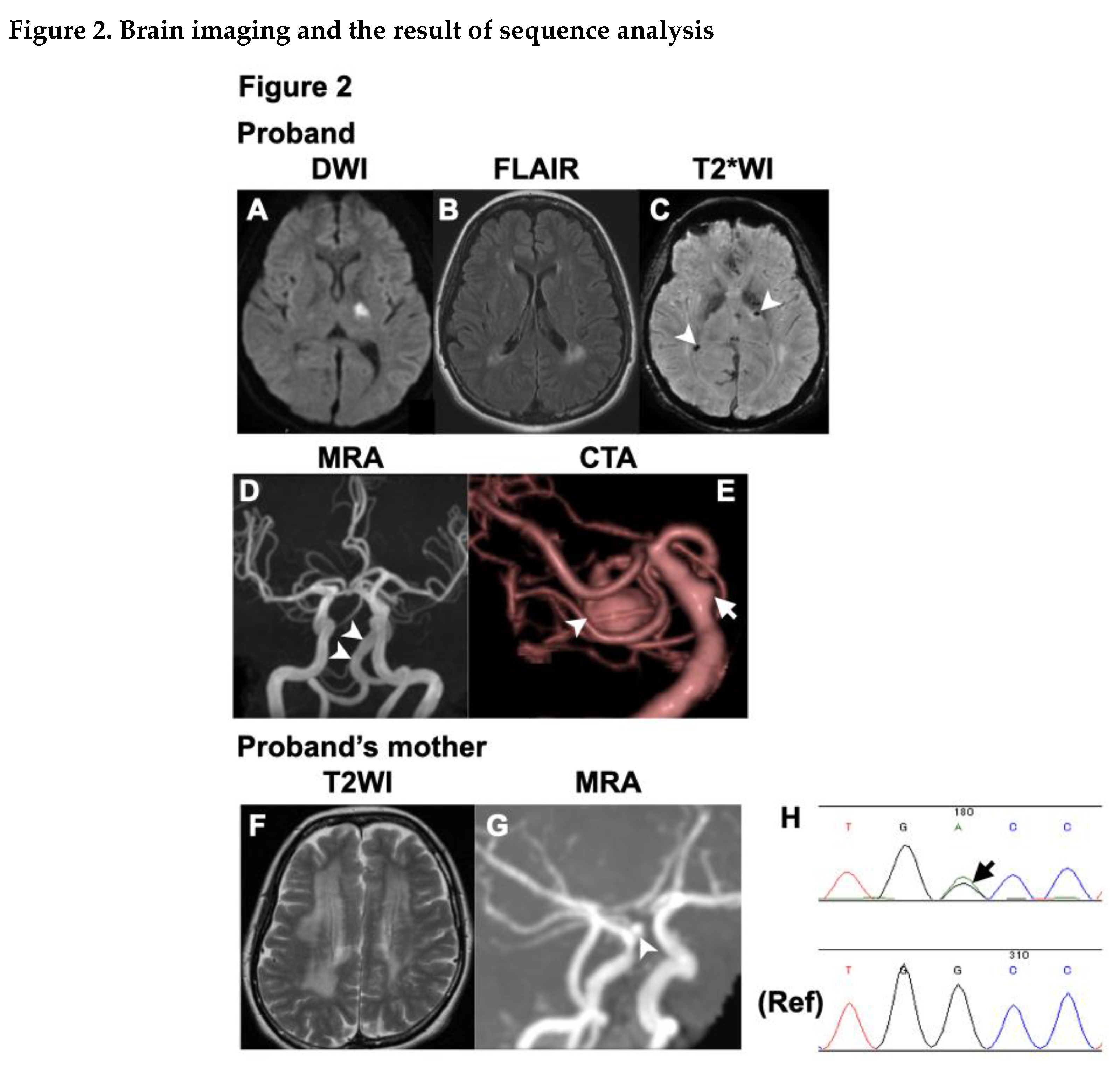
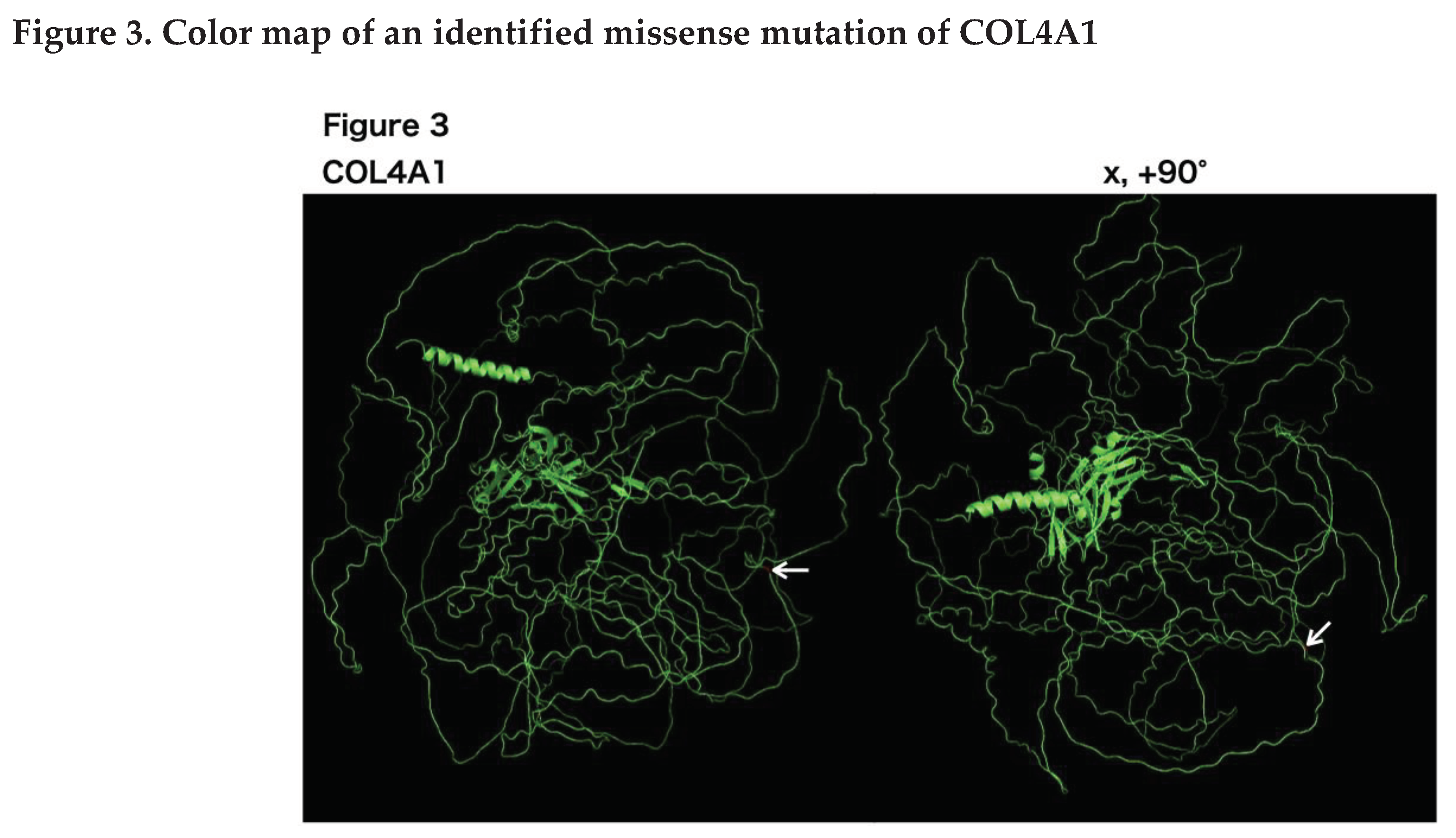
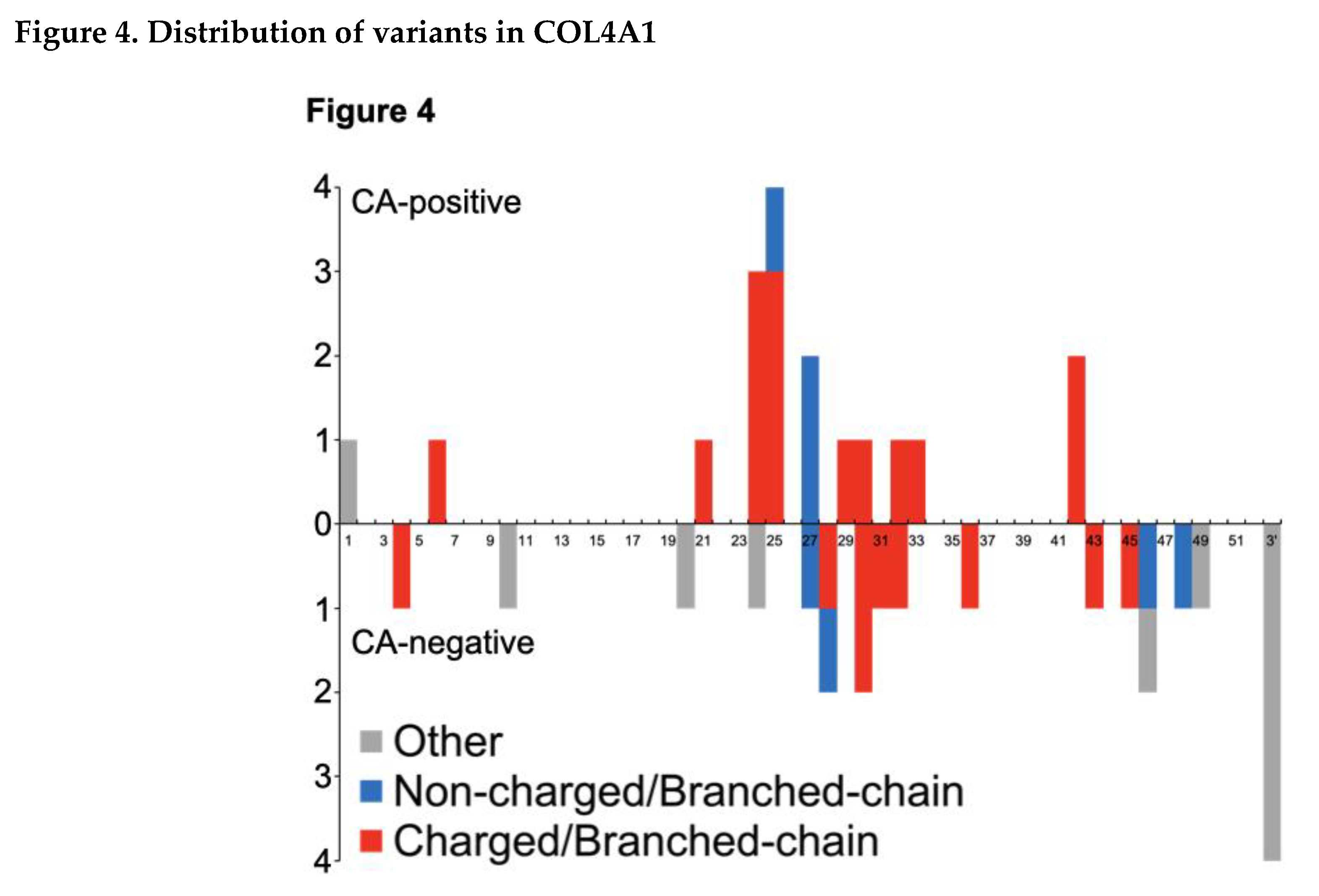
Figure 1.
Box and circle symbols represent male and female individuals, respectively. Filled symbols indicate individuals with a history of stroke. Diagonal lines were used to mark deceased family members. Arrows indicate the proband (III-1) and the proband's mother (II-1).
Figure 1.
Box and circle symbols represent male and female individuals, respectively. Filled symbols indicate individuals with a history of stroke. Diagonal lines were used to mark deceased family members. Arrows indicate the proband (III-1) and the proband's mother (II-1).
Figure 2.
Brain magnetic resonance images (MRIs) of the proband (A-E), including magnetic resonance angiography (MRA) (D) and computed tomography angiography (CTA) (E) as well as MRI and MRA (F, G) of the proband’s mother are presented. (A) Diffusion-weighted image shows acute infarction in the left posterior limb of the internal capsule. (B) Fluid-attenuated inversion recovery image (FLAIR) shows patchy white matter hyperintensities (WMHs) around the lateral ventricles. (C) T2* weighted image displays multiple microbleeds (arrowheads). (D) Brain magnetic resonance angiography (MRA) shows vertebrobasilar dolichoectasia. (E) Computed tomography angiography (CTA) reveals a large superior cerebellar artery (SCA) aneurysm (marked by an arrowhead) and an aneurysm on the lateral side of the basilar artery (marked by an arrow). (F) T2 weighted image shows diffuse WMHs, and (G) MRA shows an aneurysm of the right ICA just above the ophthalmic artery. (H) Sequence analysis reveals a heterozygous variant, c.3734G>A, in COL4A1.
Figure 2.
Brain magnetic resonance images (MRIs) of the proband (A-E), including magnetic resonance angiography (MRA) (D) and computed tomography angiography (CTA) (E) as well as MRI and MRA (F, G) of the proband’s mother are presented. (A) Diffusion-weighted image shows acute infarction in the left posterior limb of the internal capsule. (B) Fluid-attenuated inversion recovery image (FLAIR) shows patchy white matter hyperintensities (WMHs) around the lateral ventricles. (C) T2* weighted image displays multiple microbleeds (arrowheads). (D) Brain magnetic resonance angiography (MRA) shows vertebrobasilar dolichoectasia. (E) Computed tomography angiography (CTA) reveals a large superior cerebellar artery (SCA) aneurysm (marked by an arrowhead) and an aneurysm on the lateral side of the basilar artery (marked by an arrow). (F) T2 weighted image shows diffuse WMHs, and (G) MRA shows an aneurysm of the right ICA just above the ophthalmic artery. (H) Sequence analysis reveals a heterozygous variant, c.3734G>A, in COL4A1.
Figure 3.
Three-dimensional COL4A1 monomer structures generated by PyMol are shown. COL4A1 monomers are shown as green ribbons. The COL4A1 structure reference data were generated using the alphafold2. On the left side are images from the front view, and x, 90 indicate the degree of rotation along the x-axes, respectively. Arrows indicate Gly1245 in COL4A1, which was identified in our patient.
Figure 3.
Three-dimensional COL4A1 monomer structures generated by PyMol are shown. COL4A1 monomers are shown as green ribbons. The COL4A1 structure reference data were generated using the alphafold2. On the left side are images from the front view, and x, 90 indicate the degree of rotation along the x-axes, respectively. Arrows indicate Gly1245 in COL4A1, which was identified in our patient.
Figure 4.
In the graph, the exon positions of COL4A1 are presented along the horizontal axis, and the number of mutations is depicted on the vertical axis. Missense mutations in COL4A1 that give rise to charged/branch-chain amino acid substitutions are highlighted in red. These missense mutations are particularly noteworthy as our findings suggest their relevance to cerebral aneurysms (CA). Mutations that resulted in substitutions other than the charged/branch chain amino acids are denoted in blue, while other mutation types are indicated in gray. For splice site mutations, the exons predicted to undergo skipping are shown. The 3' untranslated region (3' UTR) is denoted as 3’.
Figure 4.
In the graph, the exon positions of COL4A1 are presented along the horizontal axis, and the number of mutations is depicted on the vertical axis. Missense mutations in COL4A1 that give rise to charged/branch-chain amino acid substitutions are highlighted in red. These missense mutations are particularly noteworthy as our findings suggest their relevance to cerebral aneurysms (CA). Mutations that resulted in substitutions other than the charged/branch chain amino acids are denoted in blue, while other mutation types are indicated in gray. For splice site mutations, the exons predicted to undergo skipping are shown. The 3' untranslated region (3' UTR) is denoted as 3’.
4. Discussion
In this study, we presented two patients carrying a novel COL4A1 variant, and one of them had multiple CA, including an SCA aneurysm. Thus, we extended our investigation to a review of previous publications on COL4A1/2 variants, and demonstrated that CAs outside the ICA are rare. Although there have been no reports of SAH caused by aneurysmal rupture among patients with COL4A1/2 variants, we chose to treat the SCA aneurysm in the proband because the risk of rupture was presumed to be high [25].
This review presents several novel findings. First, all mutations except for a start codon mutation, in the CA group are missense variants. In addition, the frequency of missense mutations with charged/branched side-chain AA substitution were higher in CA-positive patients. No patients with frameshift, splice site, duplication/CNV, or 3’-UTR variants had CAs. To date, the role of COL4A1/2 in aneurysm development remains unclear. A recent report showed that loss of COL4A1 expression is associated with the development of abdominal aneurysms [26]. However, our results suggest that quantitative changes are not sufficient and that qualitative changes of COL4A1/2 proteins are necessary for CA development.
COL4A1/2 are network-forming extracellular proteins that interact with several extracellular matrix proteins such as integrins and laminins. Variants in HANAC syndrome are clustered in amino acids 498-528, which are encoded by exons 24 and 25 of COL4A1. Gly1245Asp are located outside the clustered site [27]. Therefore, the pathomechanism of Gly1245Asp may differ from that of mutations in clustered sites. A recent report showed that a soluble form of the COL4A1 protein, induced by X-box binding protein 1, promotes the migration of vascular progenitor cells to injured vessels [28]. This soluble form of COL4A1 is a transcript variant generated by joining the front part of exon 4 with the rear part of exon 42 and contains the Gly1239 and Gly1245 sequences of COL4A1. According to the results of this study, these two variants may cause functional impairment of the soluble form of COL4A1, resulting in insufficient vessel repair.
Next, we demonstrated that CA could develop irrespective of the status of small-vessel disease or other large vessel abnormalities. We investigated clinical findings or signs related to small-vessel disease, including stroke, RAT, or nephropathy, and found that the frequency of these clinical findings was similar between the two groups. Similarly, the frequency of large vessel abnormalities other than CA was comparable between the two groups. These results suggest that the mechanism of aneurysm formation is distinct from that of small-vessel disease or other large vessel abnormalities. Although various factors, such as inflammatory response and shear stress, can contribute to CA formation, the precise mechanism remains elusive. Generally, CAs preferentially develop in the MCA, Acom, Pcom, and BA because of high local shear stress [29]. However, the preferential site of CA in COL4A1/2-related vasculopathy differs from that in general CAs. Our review revealed that more than 80% of patients had ICA aneurysms, and 10 of 25 patients had multiple ICA aneurysms. These characteristic predispositions among patients with COL4A1/2 variants suggest that the underlying mechanisms beyond shear stress play a significant role in CA development.
This study has several limitations. First, the extracted publications often provided insufficient descriptions of COL4A1/2-related symptoms or signs, leading to several instances of missing data, making it difficult to interpret clinical and imaging findings. Second, CA was defined as the presence of an aneurysm, regardless of its size or shape. Finally, as a literature review, our study is subject to several biases, such as selection bias or reporting bias.
5. Conclusions
Here, we report two patients of a family with a novel COL4A1 variant. Furthermore, our results suggested that CA in patients with COL4A1/2-related vasculopathy is caused by missense variants. CAs can develop irrespective of other vasculopathies, such as small-vessel stroke or other large-vessel complications. Our findings underscore the need for the careful evaluation of CAs if missense variants in COL4A1/2 are identified.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org.
Author Contributions
Masahiro Uemura: Conceptualization; Data curation; Formal analysis; Investigation; Methodology (genetic tests and searching publications); Software; Visualization; Writing – original draft; Writing – review and editing. Natsuki Tanaka: Investigation (collecting the patient’s information); Writing – review and editing. Shoichiro Ando: Visualization; Writing - review, and editing. Takehiko Yanagihara: Supervision; Writing – review and editing. Osamu Onodera: Supervision; Funding acquisition; Writing – review and editing.
Funding
The study was funded by a grant-in-aid for Scientific Research on Innovative Areas (Brain Protein Aging and Dementia Control; 26117006) from MEXT, a grant-in-aid for Practical Research Project for Rare/Intractable Diseases (17928469) from AMED, a grant-in-aid for Scientific Research (A) (16804840), a grant-in-aid for Medical Research from the Takeda Science Foundation, and a grant-in-aid for Research on Intractable Disease (21FC0201) from the Japanese Ministry of Health, Labor, and Welfare, Japan.
Institutional Review Board Statement
This study was approved by the ethics board of Niigata University (approval numbers801, 802, and G2020-0032).
Informed Consent Statement
Written informed consent was obtained from our patient (Proband#1) and her mother (Proband#2), including the consent for publication.
Data Availability Statement
For appropriate reasons, the data supporting this research will be made available by the corresponding author.
Acknowledgments
We are grateful to Dr. Noriko Matsumoto (Department of Neurology, Nippon Medical School) for her cooperation in the collection of publications. We also extend our appreciation to Dr. Takahiro Nagai (Department of Neurology, Brain Research Institutes, Niigata University) for generating 3D models of COL4A1 using Alphafold2.
Conflicts of Interest
Osamu Onodera received a speaker honorarium from Kyowa Hakko Kirin Co., Ltd., Bristol-Myers Squibb, Ono Pharmaceutical Co., Ltd., Mitsubishi Tanabe Pharm, Takeda, Daiichi-Sankyo, FUJIFILM, SANOFI, and FP-pharm. Other authors have no conflicts of interest to declare.
References
- Plaisier, E.; Gribouval, O.; Alamowitch, S.; Mougenot, B.; Prost, C.; Verpont, M.C.; Marro, B.; Desmettre, T.; Cohen, S.Y.; Roullet, E.; et al. COL4A1 mutations and hereditary angiopathy, nephropathy, aneurysms, and muscle cramps. New England Journal of Medicine 2007, 357, 2687-2695. [CrossRef]
- Yoneda, Y.; Haginoya, K.; Arai, H.; Yamaoka, S.; Tsurusaki, Y.; Doi, H.; Miyake, N.; Yokochi, K.; Osaka, H.; Kato, M.; et al. De Novo and Inherited Mutations in COL4A2, Encoding the Type IV Collagen alpha 2 Chain Cause Porencephaly. American Journal of Human Genetics 2012, 90, 86-90. [CrossRef]
- Lanfranconi, S.; Markus, H.S. COL4A1 Mutations as a Monogenic Cause of Cerebral Small Vessel Disease A Systematic Review. Stroke 2010, 41, E513-E518. [CrossRef]
- Jeanne, M.; Gould, D.B. Genotype-phenotype correlations in pathology caused by collagen type IV alpha 1 and 2 mutations. Matrix Biology 2017, 57-58, 29-44. [CrossRef]
- Jeanne, M.; Jorgensen, J.; Gould, D.B. Molecular and Genetic Analyses of Collagen Type IV Mutant Mouse Models of Spontaneous Intracerebral Hemorrhage Identify Mechanisms for Stroke Prevention. Circulation 2015, 131, 1555-U1564. [CrossRef]
- Alamowitch, S.; Plaisier, E.; Favrole, P.; Prost, C.; Chen, Z.; Van Agtmael, T.; Marro, B.; Ronco, P. Cerebrovascular disease related to COL4A1 mutations in HANAC syndrome. Neurology 2009, 73, 1873-1882. [CrossRef]
- Uemura, M.; Hatano, Y.; Nozaki, H.; Ando, S.; Kondo, H.; Hanazono, A.; Iwanaga, A.; Murota, H.; Osakada, Y.; Osaki, M.; et al. High frequency of HTRA1 AND ABCC6 mutations in Japanese patients with adult-onset cerebral small vessel disease. Journal of Neurology Neurosurgery and Psychiatry 2023, 94, 74-81. [CrossRef]
- Garcia, F.A.O.; de Andrade, E.S.; Palmero, E.I. Insights on variant analysis in silico tools for pathogenicity prediction. Front Genet 2022, 13, 1010327. [CrossRef]
- Jaganathan, K.; Kyriazopoulou Panagiotopoulou, S.; McRae, J.F.; Darbandi, S.F.; Knowles, D.; Li, Y.I.; Kosmicki, J.A.; Arbelaez, J.; Cui, W.; Schwartz, G.B.; et al. Predicting Splicing from Primary Sequence with Deep Learning. Cell 2019, 176, 535-548 e524. [CrossRef]
- Yeo, G.; Burge, C.B. Maximum entropy modeling of short sequence motifs with applications to RNA splicing signals. J Comput Biol 2004, 11, 377-394. [CrossRef]
- Houdayer, C.; Caux-Moncoutier, V.; Krieger, S.; Barrois, M.; Bonnet, F.; Bourdon, V.; Bronner, M.; Buisson, M.; Coulet, F.; Gaildrat, P.; et al. Guidelines for splicing analysis in molecular diagnosis derived from a set of 327 combined in silico/in vitro studies on BRCA1 and BRCA2 variants. Hum Mutat 2012, 33, 1228-1238. [CrossRef]
- Richards, S.; Aziz, N.; Bale, S.; Bick, D.; Das, S.; Gastier-Foster, J.; Grody, W.W.; Hegde, M.; Lyon, E.; Spector, E.; et al. Standards and guidelines for the interpretation of sequence variants: a joint consensus recommendation of the American College of Medical Genetics and Genomics and the Association for Molecular Pathology. Genet Med 2015, 17, 405-424. [CrossRef]
- Jumper, J.; Evans, R.; Pritzel, A.; Green, T.; Figurnov, M.; Ronneberger, O.; Tunyasuvunakool, K.; Bates, R.; Zidek, A.; Potapenko, A.; et al. Highly accurate protein structure prediction with AlphaFold. Nature 2021, 596, 583-589. [CrossRef]
- Neinast, M.; Murashige, D.; Arany, Z. Branched Chain Amino Acids. Annu Rev Physiol 2019, 81, 139-164. [CrossRef]
- Fernandes, C.S.; Gens, H.; Machado, R.; Silva, A.R.; Baldeiras, I.; Almeida, M.R.; Santo, G. COL4A2 gene mutations as cause of cerebral small vessel disease, hemorrhagic stroke and intracranial vessels dolichoectasia. European Journal of Neurology 2020, 27, 1048-1048.
- Ilinca, A.; Martinez-Majander, N.; Samuelsson, S.; Piccinelli, P.; Truve, K.; Cole, J.; Kittner, S.; Soller, M.; Kristoffersson, U.; Tatlisumak, T.; et al. Whole-Exome Sequencing in 22 Young Ischemic Stroke Patients With Familial Clustering of Stroke. Stroke 2020, 51, 1056-1063. [CrossRef]
- Kumai, T.; Sadato, A.; Kurahashi, H.; Kato, T.; Adachi, K.; Hirose, Y. Coexistence of RASA1 and COL4A2 variants caused pial arteriovenous fistula (AVF) in a patient with capillary malformation-arteriovenous malformation. Clin Neurol Neurosurg 2021, 204, 106612. [CrossRef]
- Faure, C.; Castrale, C.; Benabed, A.; Cognard, P.; Leze, R.; Castro-Farias, D.; Gerard, M.; Louapre, C.; Paques, M. Structural and functional analysis of retinal vasculature in HANAC syndrome with a novel intronic COL4A1 mutation. Microvascular Research 2023, 145. [CrossRef]
- Kellett, S.; Lemaire, M.; Miller, S.P.; Licht, C.; Yoon, G.; Dlamini, N.; Noone, D. Neonatal stroke and haematuria: Questions and Answers. Pediatric Nephrology 2018, 33, 805-811. [CrossRef]
- Shan, L.D.; Peng, J.; Xiao, H.; Wu, L.W.; Duan, H.L.; Pang, N.; Miriam, K.; Yin, F. Clinical features and COL4A1 genotype of a toddler with hereditary angiopathy with nephropathy, aneurysms and muscle cramps syndrome (Chinese). Zhongguo Dang Dai Er Ke Za Zhi 2019, 21, 754-760. [CrossRef]
- Anna, A.; Monika, G. Splicing mutations in human genetic disorders: examples, detection, and confirmation. J Appl Genet 2018, 59, 253-268. [CrossRef]
- Giorgio, E.; Vaula, G.; Bosco, G.; Giacone, S.; Mancini, C.; Calcia, A.; Cavalieri, S.; Di Gregorio, E.; De Longrais, R.R.; Leombruni, S.; et al. Two families with novel missense mutations in COL4A1: When diagnosis can be missed. Journal of the Neurological Sciences 2015, 352, 99-104. [CrossRef]
- Tee, T.Y.; Tan, Y.Y.; Ngu, L.H.; Husin, M.; Nasir, M.N.M.; Ibrahim, K.A.; Aziz, Z.A. Case report of COL4A1 mutation as monogenic cause of cerebral small vessel disease (Abstract). International Journal of Stroke 2022, 17, 183-183.
- Sibon, I.; Coupry, I.; Menegon, P.; Boucher, J.P.; Gorry, P.; Burgelin, I.; Calvas, P.; Orignac, I.; Dousset, V.; Lacombe, D.; et al. COL4A1 mutation in Axenfeld-Rieger anomaly with leukoencephalopathy and stroke. Annals of Neurology 2007, 62, 177-184. [CrossRef]
- Investigators, U.J.; Morita, A.; Kirino, T.; Hashi, K.; Aoki, N.; Fukuhara, S.; Hashimoto, N.; Nakayama, T.; Sakai, M.; Teramoto, A.; et al. The natural course of unruptured cerebral aneurysms in a Japanese cohort. N Engl J Med 2012, 366, 2474-2482. [CrossRef]
- Steffensen, L.B.; Stubbe, J.; Lindholt, J.S.; Beck, H.C.; Overgaard, M.; Bloksgaard, M.; Genovese, F.; Holm, N.S.; Tha, M.L.T.; Bang-Moeller, S.K.; et al. Basement membrane collagen IV deficiency promotes abdominal aortic aneurysm formation. Scientific Reports (Nature Publisher Group) 2021, 11. [CrossRef]
- Parkin, J.D.; San Antonio, J.D.; Pedchenko, V.; Hudson, B.; Jensen, S.T.; Savige, J. Mapping structural landmarks, ligand binding sites, and missense mutations to the collagen IV heterotrimers predicts major functional domains, novel interactions, and variation in phenotypes in inherited diseases affecting basement membranes. Hum Mutat 2011, 32, 127-143. [CrossRef]
- Angbohang, A.; Huang, L.; Li, Y.; Zhao, Y.; Gong, Y.; Fu, Y.; Mao, C.; Morales, J.; Luo, P.; Ehteramyan, M.; et al. X-box binding protein 1-mediated COL4A1s secretion regulates communication between vascular smooth muscle and stem/progenitor cells. J Biol Chem 2021, 296, 100541. [CrossRef]
- Jung, K.H. New Pathophysiological Considerations on Cerebral Aneurysms. Neurointervention 2018, 13, 73-83. [CrossRef]

Table 1.
Comparison between CA positive and negative variants.
| Positive | Negative | p-value | |
| COL4A1+COL4A2+Duplications/CNV | n = 22 | n = 31 | |
| Variant type | |||
| Missense | 21 (95.5) | 18 (58.1) | 0.0035 |
| G-M-Y | 1 ( 4.5) | 1 ( 3.2) | 1 |
| G-X-M | 2 ( 9.1) | 0 (0) | 0.16763 |
| M-X-Y | 17 (77.3) | 15 (48.4) | 0.0475 |
| Charged/Branched-chain AAs | 18 (81.8) | 12 (38.7) | 0.0022 |
| Other than missense variants | |||
| 3'UTR | 0 (0) | 4 (12.9) | 0.1324 |
| Duplication/CNV | 0 (0) | 3 ( 9.7) | 0.2576 |
| Frameshift | 0 (0) | 2 ( 6.5) | 0.5051 |
| Splice Site | 0 (0) | 4 (12.9) | 0.1324 |
| Start Codon | 1 ( 4.5) | 0 (0) | 0.4151 |
| Domain | |||
| 7S | 1 ( 4.5) | 1 (3.2) | 1 |
| NC1 | 0 (0) | 3 (9.7) | 0.2576 |
| Signal | 1 ( 4.5) | 0 (0) | 0.4151 |
| Triple-helical | 20 (90.9) | 20 (64.5) | 0.0497 |
Table 1. G-M-Y indicates that the variant is located at the amino acid position after the glycine residue in the glycine triplet sequence. G-X-M indicates that the variant is located at amino acid position two after the glycine residue in the glycine triplet sequence. M-X-Y indicates that the variant is located at the glycine position in the glycine triplet sequence. The UTR represents the untranslated region. NC1 represents the non-collagenous domain. AAs are amino acids. CNV is an abbreviation for copy number variant.
Table 2.
Variants found in patients with CA outside the ICA.
| CA positive | Ref | |
| MCA | COL4A1, Gly498Val | [6] |
| Acom | COL4A1, Gly417Arg | [22] |
| Pcom | COL4A1, Pro648Ala | [23] |
| BA |
COL4A1, Gly1245Asp COL4A1, Gly720Asp |
[24] ours |
| SCA | COL4A1, Gly1245Asp | ours |
Table 2. CA indicates cerebral aneurysm; ICA, internal carotid artery; MCA, middle cerebral artery; Acom, anterior communicating artery; Pcom, posterior communicating artery; SCA, superior cerebellar artery.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



