
Submitted:
15 December 2023
Posted:
19 December 2023
You are already at the latest version
Abstract
MicroRNAs (miRNAs) play a pivotal role in important biological processes by regulating posttranscriptional gene expression and exhibit differential expression patterns during development, immune responses, and stress challenges. The diamondback moth causes significant economic damage to crops worldwide. Despite substantial advancements in understanding the molecular biology of this pest, our knowledge regarding the role of miRNAs in regulating key immunity-related genes remains limited. In this study, we leveraged whole transcriptome resequencing data from P. xylostella infected with Metarhizium anisopliae to identify specific miRNAs targeting the prophenoloxidase-activating protease1 (PAP1) gene through bioinformatic analysis. Seven miRNAs (pxy-miR-375-5p, pxy-miR-4448-3p, pxy-miR-279a-3p, pxy-miR-3286-3p, pxy-miR-965-5p, pxy-miR-8799-3p and pxy-miR-14b-5p) were identified and validated. Luciferase reporter assays confirmed that pxy-miR-279a-3p binds to the open reading frame (ORF) and pxy-miR-965-5p to the 3’ untranslated region (3’ UTR) of PAP1. Our experiments demonstrated that injecting a pxy-miR-965-5p mimic significantly reduced PAP1 expression in P. xylostella larvae, suppressing PO activity and increasing larval mortality rate. Conversely, the injection of inhibitors obtained opposite results. Furthermore, we identified four LncRNAs (MSTRG.32910.1, MSTRG.7100.1, MSTRG.6802.1, and MSTRG.22113.1) that potentially interact with pxy-miR-965-5p. Interference assays using antisense oligonucleotides (ASO) revealed that silencing MSTRG.7100.1 and MSTRG.22113.1 increased the expression of pxy-miR-965-5p. These findings shed light on the potential role of pxy-miR-965-5p in the immune response of P. xylostella to M. anisopliae infection and provide a theoretical basis for biological control strategies targeting the immune system of this pest.
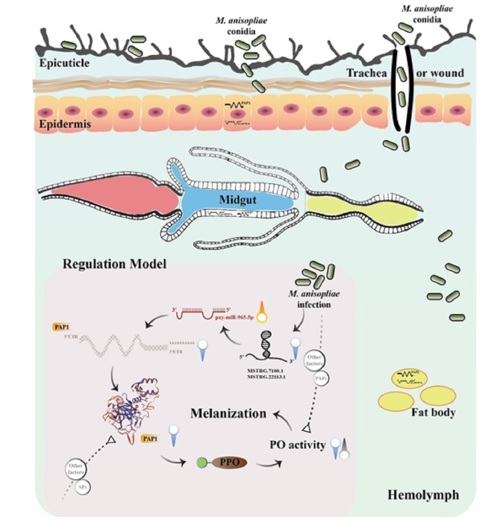
Keywords:
Diamondback moth
; noncoding RNAs
; melanization
; pest management.
Article
1. Introduction
Many RNA molecules are produced in eukaryotic cells, but only a minute fraction of the total RNA is converted into protein, and others devoid of protein-coding regions are called non-coding RNAs (ncRNAs). While the small ncRNAs (sncRNAs) such as microRNA (miRNA), small nuclear RNA (snRNA), small interfering RNA (siRNA), and PIWI-interacting RNAs (piRNAs) have a size <200 nucleotides, whereas the long ncRNAs (LncRNAs) have a length of >200 bp [1,2]. The LncRNAs have been found to play a crucial role in regulating gene expressions in various physiological, pathological, and immunological processes in higher organisms [3]. In insects, they play a key role in the regulation of the developmental (sex-determination, immunity, and morphogenesis) as well as behavioral (sleeping, foraging, courtship, etc) processes [2,4,5]. However, studies on insect LncRNAs are mostly limited to model species [2]. With the advancement of high throughput technology (HTS), many insects, including the nonmodal species, have been characterized and identified several novel non-coding RNAs (ncRNAs). For example, several novel LncRNAs have been identified in P. xylostella that play important roles in developing host resistance by acting as a precursor molecule for the production of miRNAs. The lnc-GSTu1-AS has been reported in chlorantraniliprole-resistant strains of P. xylostella, and its deletion significantly decreased the host resistance to chlorantraniliprole. It has been reported that the lnc-GSTu1-AS affects the expression of GSTu1 by targeting miR-8525-5p, leading to enhanced resistance to chlorantraniliprole [6]. The function of microRNAs (miRNAs) is mainly involved in the regulation of the genes' expression through binding on the mRNAs and modulates metabolism, immunity, and developmental processes via post-transcriptional regulation [7,8]. Synthetic miRNA mimics and inhibitors has been identified that target key genes and interfere with the phenotypic expression of the insect [9,10,11,12]. Although several miRNAs have been reported to have possible involvement in regulating immune-related processes, mechanistic studies on their role in insect immunity are largely limited [13].
The innate immunity of insects comprises cellular and humoral responses [14,15]. Cellular immunity primarily refers to phagocytosis and encapsulation by the hemocytes [16], while humoral immunity deals with the production of antimicrobial peptides [17] in the fat body and release rapidly into the hemolymph, leading to the activation of the prophenoloxidase (proPO) or the melanization cascade [18,19,20]. proPO is activated via serine proteases (SPs) cascades when insect pattern recognition receptors (PRRs) recognize the pathogen-associated molecular patterns (PAMPs). This activates of prophenoloxidase-activating proteinase (PAP) [21]. The PO cascade plays a crucial role during the fungal invasion by converting phenols to quinones and subsequently polymerizing to form melanin to defend against pathogens [22]. PAPs are the direct activator of proPO, while Serpins can prevent the SPs from excessively regulating melanization [23,24]. In addition to the SPs cascades, PAPs and PO activities are also regulated by miRNAs [25].
P. xylostella (Linnaeus) (Lepidoptera: Plutellidae) causes significant economic losses to cruciferous crops [26,27]. The present control strategy mainly involves the application of wide-spectrum insecticides, resulting in the development of resistance and environmental and health risks [28,29]. Entomopathogenic fungal insecticides are promising pest management tools that attach to the cuticle, germinate, and penetrate hemolymph, leading to host death [30,31,32]. For example, members under the genus Beauveria and Metarhizium have been successfully applied against Aedes albopictus (Skuse) [33], Bemisia tabaci (Gennadius) [34], Spodoptera frugiperda Smith [35] and P. xylostella [36]. However, insects respond to pathogenic infections by activating their innate immune responses that develop resistance against EPF [37]. The miRNAs play an essential role in regulating multiple vital biological processes (development, immunity, insecticide resistance, metamorphosis, reproduction etc.) of insects. Since miRNAs and their binding site sequences are target-specific, their application allows the killing of specific pests without interfering with non-target organisms. Application of miRNA-based pest control strategy can reduce the use of hazardous pesticides and associated resistant strain development and environmental pollution. Moreover, artificial regulation of the activity of specific miRNAs can help restore insecticide sensitivity so that existing pesticides can work effectively [38]. Therefore, miRNA-based strategy could be the key to formulating a new generation of effective, environment-friendly pest control agents [39]. In our previous study, three PAPs (PAP1, PAP3, and PAP3a) were identified in P. xylostella [40], and based on transcriptome data we predicted that non-coding RNAs might regulate them. Here, we aim to delineate the interaction mechanism between LncRNA and miRNA of P. xylostella infected with Metarhizium anisopliae, using in vitro and in vivo experiments. The identified candidate miRNA pxy-miR-965-5p that targets PAP1 could serve as a promising candidate for pest management strategies.
2. Results
2.1. The infection of P. xylostella by M. anisopliae
The conidia of M. anisopliae permeate the circulatory system via wounds and the tracheal system; alternatively, they adhere to the cuticle of P. xylostella (Figure 1A). The initiation of conidial germination and subsequent growth is observed at 24 hours post-infection (hpi), culminating in a significant mortality incidence in P. xylostella larvae from 36 hpi onwards. This phenomenon is ascribed to the rampant proliferation of mycelia within the larval body, which incrementally engulfs the entire body by 60 hpi. Typically, M. anisopliae engenders an augmented count of new conidia post 96 hpi. The presence of black patches in the integument of P. xylostella after M. anisopliae infection signifies an association with melanization (Figure 1B). Following infection, the PO activity was significantly upregulated by 2.4, 1.3, 1.2 times in the treated group as compared to the control at 12, 24, and 36 hpi, respectively (Figure 1C). These findings demonstrate that M. anisopliae infection can trigger the PO cascade involved in the melanization of P. xylostella.
2.2. The functional analysis of PAP1
The PAP1 and PAP1Xa proteins were expressed in vitro to investigate the function of PAP1. The operational mode of PAP1 and the IEGR mutant PAP1Xa is depicted in Figure 2A. The SDS-PAGE results revealed a molecular weight of 57.3 kDa for PAP1Xa, which could be cleaved by Factor Xa proteases to produce an activated protein with a molecular weight of 29.4 kDa (Figure 2B). Upon cleavage by Factor Xa proteases, the trypsin activity of PAP1Xa significantly increased. The results of PO activity demonstrated that both PAP1 and activated-PAP1Xa could significantly enhance PO activity in the plasma (Figure 2C). These findings indicated that PAP1 exhibits trypsin activity and can modulate the PO activity in the plasma of P. xylostella after M. anisopliae infection.
2.3. The targeting miRNA of PAP1
Seven miRNAs that may potentially target the PAP1 gene have been identified through predictive analysis. Among these, four specific miRNAs, namely pxy-miR-375-5p, pxy-miR-4448-3p, pxy-miR-279a-3p, and pxy-miR-3286-3p, were found to target the ORF of the PAP1 gene. The remaining three, pxy-miR-965-5p, pxy-miR-8799-3p, and pxy-miR-14b-5p, were associated with the 3’ UTR region of the PAP1 gene (Figure 3A). Subsequent dual-luciferase assays revealed that only two of these miRNAs, pxy-miR-279a-3p, and pxy-miR-965-5p, could significantly downregulate the relative fluorescence units (RFU) in 293T cells. Specifically, transfection with mimics of pxy-miR-279a-3p and pxy-miR-965-5p resulted in a downregulation of RFU by 29.55% and 30.77%, respectively (Figure 3B). This suggests that both pxy-miR-279a-3p and pxy-miR-965-5p can act as negative regulators of PAP1 in vitro.
2.4. The expression patterns of PAP1, pxy-miR-279a-3p and pxy-miR-965-5p
We utilized RT-qPCR to study the expression of PAP1, pxy-miR-279a-3p, and pxy-miR-965-5p across various tissues in P. xylostella. Without M. anisopliae, the expression patterns of PAP1, pxy-miR-279a-3p, and pxy-miR-965-5p in various tissues were similar. The highest expression was observed in the epidermis, followed by the fat body and midgut. Both pxy-miR-279a-3p and pxy-miR-965-5p exhibited low expression in the hemocytes and malpighian tubules (Figure 4A). Figure 4B illustrates the expression patterns of PAP1, pxy-miR-279a-3p, and pxy-miR-965-5p after M. anisopliae infection. The relative expression of pxy-miR-965-5p was significantly increased in the midgut and fat body of P. xylostella at 12 and 24 hpi and in the epidermis at 36 hpi. Correspondingly, the relative expression of PAP1 was significantly decreased. The expression patterns of PAP1 and pxy-miR-965-5p after M. anisopliae infection appeared to exhibit a negative regulatory trend. On the other hand, the relative expression of pxy-miR-279a-3p was significantly downregulated in the midgut of P. xylostella at 12 and 24 hpi, while it was significantly upregulated in the epidermis at these same time points. However, no significant pattern was observed in relation to the expression of PAP1.
2.5. pxy-miR-965-5p modulates the susceptibility of P. xylostella to M. anisopliae
To elucidate the in vivo function of these two miRNAs, we injected them separately into P. xylostella larvae, which were subsequently infected with M. anisopliae. The mRNA level of PAP1 significantly downregulated from 12 to 36 h post pxy-miR-965-5p mimic injection. Correspondingly, the mRNA level of PAP1 significantly increased from 12 to 36 h post pxy-miR-965-5p inhibitor injection (Figure 5A). However, the injection of both the pxy-miR-279a-3p mimic and inhibitor did not significantly impact the mRNA level of PAP1 (Figure 5B). Additionally, we verified that pxy-miR-965-5p could readily induce off-target effects on PAP1 following mutation at the 4th and 6th base in the pxy-miR-965-5p seed region (Figure S1). Furthermore, we evaluated the survival rates of P. xylostella under various treatment conditions. Notably, the survival rates of the group injected with the pxy-miR-965-5p mimic were significantly lower than those injected with the NC mimic. Conversely, the survival rates of the group injected with the pxy-miR-965-5p inhibitor were significantly higher than those of the NC mimic injection group (Figure 6A). A significant decrease in PO activity in the group injected with the pxy-miR-965-5p mimic compared to the group injected with the NC mimic was observed. The group injected with the pxy-miR-965-5p inhibitor exhibited a significant increase in PO activity compared to those injected with the NC mimic (Figure 6B). These findings suggested that pxy-miR-965-5p could modulate the PO cascade by targeting PAP1, thereby influencing the infectivity of M. anisopliae in P. xylostella.
2.6. Interaction of LncRNAs with pxy-miR-965-5p
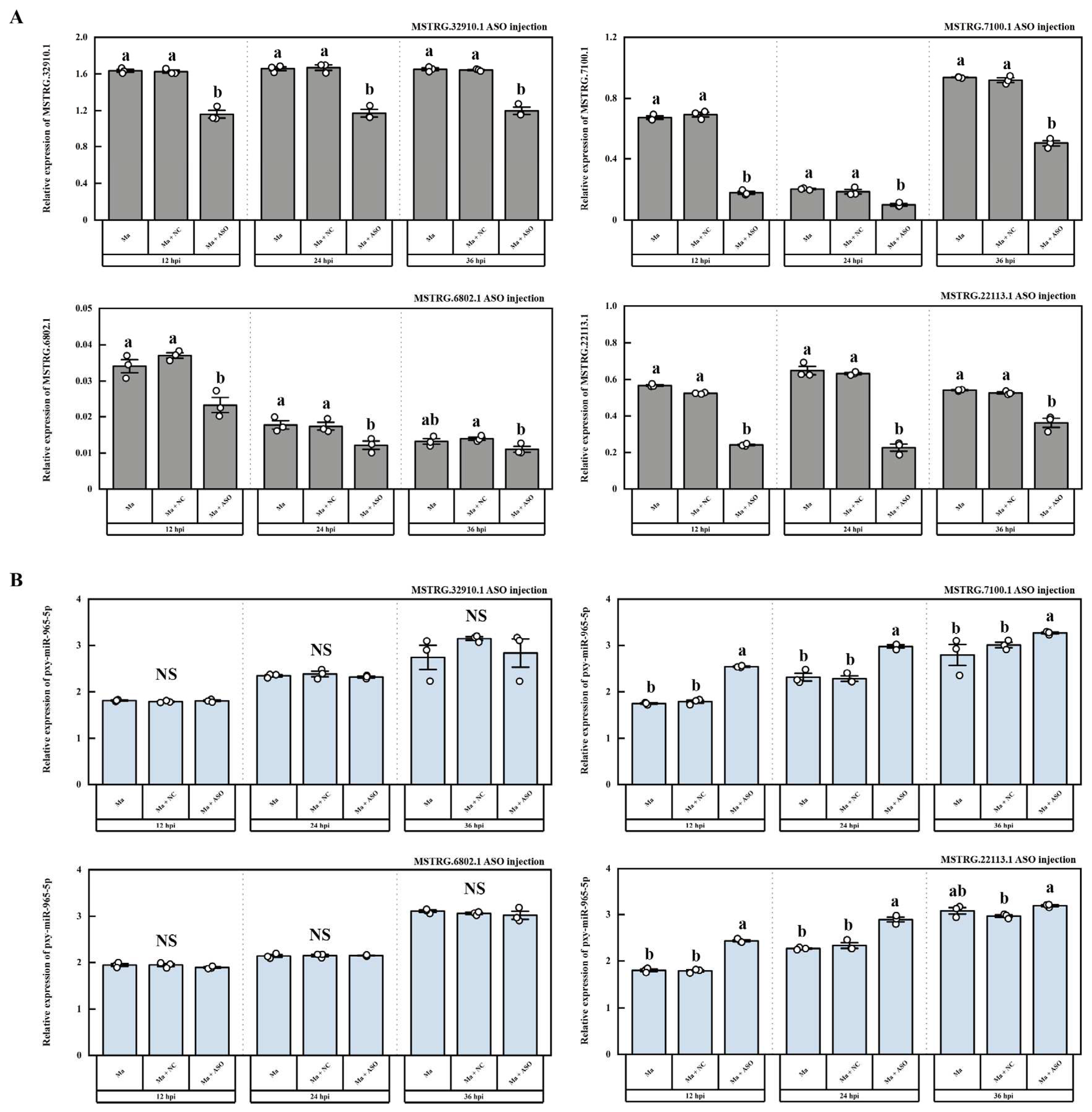
According to the previous whole transcriptome sequencing and dual-luciferase assays, we also identified four LncRNAs that potentially interact with pxy-miR-965-5p, namely MSTRG.32910.1, MSTRG.7100.1, MSTRG.6802.1, and MSTRG.22113.1, the interaction models were depicted in Figure S2. pxy-miR-965-5p was found to bind with the dual-luciferase plasmids containing the target fragments of these four LncRNAs, resulting in a decrease in RFU by 24.38%, 15.75%, 21.69%, and 10.63%, respectively (Figure 7). In vivo, we silenced these four LncRNAs through ASO injection. The results demonstrated that ASO injection effectively silenced these four LncRNAs (Figure 8A). When MSTRG.7100.1 and MSTRG.22113.1 were silenced, the relative expression of pxy-miR-965-5p correspondingly increased. However, the decrease in the relative expression of MSTRG.32910.1 and MSTRG.6802.1 did not significantly affect the relative expression of pxy-miR-965-5p (Figure 8B).
3. Discussion
Insects solely depend on the innate immune system for recognizing and targeting foreign particles since they don’t possess adaptive immunity, and the innate immunity largely depends on prophenoloxidase, which plays a key role in the cellular and humoral defense of insects [14,17,41]. The proPO system is strongly regulated by the different serine proteases such as PAPs, Serpins, and SPs/ Serine protease homologs (SPs/ SPHs). PAPs release active proPO at specific sites, mediating melanization. Although PO-mediated melanization can kill invading pathogens, overproduction of the intermediates can harm the insects. Serpin can prevent excessive melanism by inhibiting SPs/ SPHs activity and blocking proPO activation. SPs/ SPHs and Serpin rigorously regulate the activation of proPO in time and space, removing the invading exogenous substances and evading the detrimental effects of intermediates [42,43,44,45,46,47]. Therefore, the PO cascade is a good target for biological pest control. However, so far, the immune response to pathogens and the regulatory mechanism of PO production has only been preliminarily described at the protein level in some model insects, and the precise regulation of PO-mediated melanism is still not understood [45,48].
It has been described that the expression of Serpin-2, Serpin-4, Serpin-5 [49], PxSP1, PxSP2, PPO1, and PPO2 [50] were regulated after pathogenic infection in P. xylostella which triggered the PO activity. Similarly, an increased PO activity was reported in hemolymph at 24 - 48 hpi by M. anisopliae [49]. In the present study the PO activity increased in the plasma of P. xylostella at 12 to 36 h after M. anisopliae infection, (Figure 1C). The high PO activity may be due to the intense immune response in the early stages of infection that gradually declines as the infection progresses. Previous experiments showed that PPAF-II acted as a module for PO binding through the central cleft in Holotrichia Diomphalia Bates [51], and PAP-1, PAP-2, PAP-3 showed diverse abilities to activate PO activity in Manduca sexta (Linnaeus) [52,53,54,55]. Correspondingly, in this study, PAP1 activity was compared to trypsin activity (Figure 2A and 2C), and the activated PAP1 could activate the PO activity in P. xylostella (Figure 2C).
miRNAs targeted gene expression may play vital roles at the post-transcriptional level in maintaining homeostasis and plasticity of immunity. miRNA mimics or inhibitors were used to assess the function of miRNAs in insects [56,57]. For example, in Drosophila, bioinformatics was used to detect potential miRNA targets and express the profiles of miRNAs involved in regulating immune response [58]. miR-8 has been reported to target the transcripts of GNBP3 and then negatively regulate the expression of Drosomycin and Diptericin in Drosophila. It also indicated that the transcript levels of the AMPs were significantly elevated in the absence of the miR-8 in Drosophila larvae and flies [59]. The bioinformatic analysis predicted that 7 miRNAs might participate in immunity by targeting PAP1 in post M. anisopliae infection (Figure 3A). The results of the dual luciferase assay showed that only pxy-miR-279a-3p and pxy-miR-965-5p could significantly reduce the expression level of the PAP1 in 293T cells (Figure 3B).
The seed region (g2-g8) of miRNA is crucial for target recognition, and the mutations of seed sequence may disrupt the interaction between miRNAs and targeted mRNAs [60,61,62]. Experiments have depicted that the mismatches in positions 2–7 of miR-7 and miR-278 strongly reduced the magnitude of target regulation in cells [63]. The targeted site of let-7a mismatched at positions 3 and 4 or mismatched at positions 5 and 6 would reduce the repression effect on targeted mRNA in HeLa cells [64]. It also indicated that the ΔG value of this interaction is an important determinant of activity [65]. The findings of the current study proved that the targeted site in the seed region of pxy-miR-965-5p mismatched at positions 4 or 6, leading to a loss of ability to suppress PAP1 (Figure S1). Moreover, recent research also showed their involvement in recognition and regulation [65]. The mutations of a single nucleotide at g11–g16 lead to functional defects of miRNA let-7a [66].
Fungal infection can alter the expression of insect miRNAs [67]. For instance, pathogen infection leads to the upregulation of miR-92, which plays a major role in the developmental process of A. albopictus and Culex quinquefasciatus (Say) [68]. In the present study, an upregulated expression of pxy-miR-965-5p was observed upon M. anisopliae infection. The expression patterns of PAP1 and pxy-miR-965-5p after M. anisopliae infection appeared to exhibit a negative regulatory trend (Figure 4B). In addition, the tissue expression patterns of pxy-miR-279a-3p and pxy-miR-965-5p were similar to PAP1, which was upregulated in the epidermis and fat body (Figure 4A). Generally, miRNAs function by down-regulating the expression of targeted genes [69], and the expression pattern of most miRNAs and their targeted mRNAs had spatial-temporal specificity [70,71]. miR-92b can down-regulate Mef2 and were expressed in the heart and specific muscles in Drosophila [72]; miR-927 can down-regulate Kr-h1 in the fat body of Drosophila [73]; miR-184 can down-regulate the CYP303A1 in the integument of L. migratoria [74].
The expression of EcR was down-regulated by miR-34-5p in Helicoverpa armigera (Hübner), Spodoptera exigua Hübner, and P. xylostella, but the physiological effects were not quite the same [75]. JNK, regulated by miRNA-184, can regulate the ROS metabolism and PO activity but was not exactly a simple corresponding relationship in vivo in the agomir injection experiments [76]. We observed that PAP1 expression was down-regulated after the pxy-miR-965-5p mimic was injected into P. xylostella, leading to decreased PO activity; on the control, pxy-miR-965-5p inhibitor was injected into P. xylostella, leading to increased PO activity (Figure 5A, Figure 6B). The pathogen's infection would increase with the repression of the PO cascade. A repressed PO cascade increased the mortality of L. migratoria nymphs during M. anisopliae infection [77]. The activity of PO cascade can block baculovirus infection, but the inhibition can be rescued by serpin-9 [78]. The results of our study demonstrated that pxy-miR-965-5p could repress the PO cascade. It indicated that the overexpression of pxy-miR-965-5p was conducive to M. anisopliae infection (Figure 6A). Therefore, during M. anisopliae infection, pxy-miR-965-5p was upregulated, and the PO cascade was suppressed, resulting in increased mortality in P. xylostella (Figure 6).
LncRNA is a non-coding single-stranded RNA (>200 bp) composed of exon or intron sequences. It can be clipped, capped, polyadenylate, and can play a regulatory role in the nucleus or cytoplasm [79]. They are involved in regulating the gene expression in the nucleus via controlling the local chromatin structure or recruiting regulatory factors to specific sites, such as acting as enhancers to regulate gene expression [80,81]. LncRNA regulates gene expression by altering the scaffold activity of chromatin modification proteins such as methyltransferase, demethylase, acetyltransferase, and deacetylase. And recruit these proteins to target sites by either cis-regulation (regulating transcription of nearby genes) or trans-regulation (regulating transcription of genes farther away in the genome) [82]. In the cytoplasm, LncRNA interacts with other types of RNA, affecting mRNA stability, mRNA translation and acting as a miRNA sponge for miRNA absorption [83]. miR-2834 can reduce the expression of vitellogenin (vg) in silkworms, thereby affecting the formation of eggs, while lncR26319 can act as a miRNA sponge to reduce miR-2834's silting of vg, thereby increasing vg expression [84]. At present, the functional research of LncRNA mainly focuses on the knockdown. The mechanisms of dsRNA and ASO are different. dsRNA is cleaved by Dicer enzyme and then combined with AGO to form RISC silencing complex, which is then complementary and paired with target RNA to perform functions, while ASO can directly bind to target RNA. The RNase-H enzyme is recruited in the nucleus or cytoplasm to cut the target RNA to perform the function [85]. In the present study, based on the whole transcriptome sequencing and dual-luciferase assays, four LncRNAs, MSTRG.32910.1, MSTRG.7100.1, MSTRG.6802.1, and MSTRG.22113.1 were identified (Figure S2). In vitro, we tested the activity of four LncRNAs by dual-luciferase (Figure 7). In vivo, we silenced these four LncRNAs through ASO injection. The results demonstrated that ASO injection can effectively silence four LncRNAs (Figure 8A). When MSTRG.7100.1 and MSTRG.22113.1 were silenced, the expression of pxy-miR-965-5p correspondingly increased. However, silence the MSTRG.32910.1 and MSTRG.6802.1 fails to alter the expression of pxy-miR-965-5p (Figure 8B).
4. Materials and Methods
4.1. Insects, pathogens and cell lines
The strain (susceptible) of P. xylostella was collected from the Engineering Research Center of Biological Control, SCAU, China, and reared for a minimum of 10 generations in an environment devoid of insecticides. The artificial diet with 10% honey was used to rear the insects at 25 ± 1°C, a photoperiod of 16:8 h (light: dark), and a relative humidity of 65 ± 5%. M. anisopliae was obtained from the Bahauddin Zakariya University, Pakistan, grown on potato dextrose agar (PDA), and harvested in a solution of 0.05% Tween-80 (Sigma-P1754). The concentration of the conidia was measured using a hemocytometer. The HEK 293T cell line was grown in Dulbecco’s Modified Eagle Medium (Gibco, USA), supplemented with 10% fetal bovine serum and 1% Penicillin-streptomycin (double antibody) (Gibco, USA). The cells were incubated at 37 ℃ in an environment with 5% CO2.
4.2. P. xylostella infection by M. anisopliae
4.3. PO activity assay
The plasma was obtained from the P. xylostella at 0, 12, 24, and 36 h post-treatment, and the protein content was analyzed using the BCA Protein Assay Kit (Sangon Biotech, CHN). For the PO activity assay, the plasma samples were treated with 200 μL substrate (2 mM dopamine in 10 mM sodium phosphate buffer [pH 7.4]), and the absorbance of the product was measured by iMark Absorbance Microplate Reader at 490 nm in 10 min. One unit of activity was defined as ΔA490 of 0.01 in one minute [86].
4.4. The prokaryotic expression and purification of recombinant proteins
The recombination proteins of PAP1 and PAP1Xa were expressed using the vector pET-32a and Escherichia coli BL21. The primers for plasmid preparation were shown in Table S1. The recombinant protein production and purification was carried out as per our previous publication, and the purified proteins were stored in a PBS buffer [40].
4.5. The enzyme activity of PAP1
0.4 μg of Factor Xa Protease (New England Biolabs, USA) and 2 μg of recombinant protein PAP1Xa were incubated at 25 ℃ for 6 h, resulting in the cleavage and activation of PAP1 Xa. The various treatment groups were separately added with 150 μL 1mM N-Benzoyl-L-arginine ethyl ester hydrochloride (BAEE) solution (Solarbio, CHN), and the trypsin activity was determined at 253 nm using SynergyTM H1 (BioTek, USA) over a duration of 30 min (30 s interval). In addition, the various treatment groups were separately incubated with plasma (24 h post M. anisopliae infection) at room temperature for 10 min, then treated with 200 μL substrate (2 mM dopamine in 10 mM sodium phosphate buffer [pH 7.4]), the PO activity as per the standard procedure [40].
4.6. Prediction of Interactions Among mRNA, miRNA, and LncRNAs
Three tools, such as miRanda, RNAhybrid, and TargetScan, were used to identify the miRNA and LncRNA targets. The target mRNA, miRNA, and LncRNAs were screened from the transcriptomics of P. xylostella infected by M. anisopliae, which was previously published by our laboratory [87,88,89,90]. The target fragments of miRNAs to PAP1, pxy-miR-965-5p to LncRNAs, and the mutant target fragments of pxy-miR-965-5p to PAP1 were cloned into the psiCHECK-2 plasmid (Promega, USA). The primer details for plasmid preparation were shown in Tables S3 and S4. HEK 293T cells were cultured at a density of 2 × 105 cells/well in 24-well culture plates and co-transfected with 0.8 μg of the luciferase reporter vector and 20 pmol of miRNA mimic using LipofectamineTM 2000 (Invitrogen, USA). Cells were cultured and transfected for 48 h to get dual luciferase. The activities were measured by Dual-Luciferase® Reporter Assay System (Promega, USA) using the multimode reader SynergyTM H1 (BioTek, USA).
4.7. The synthesis of miRNA mimic, miRNA inhibitors, and ASO for LncRNAs
Mimics of miRNA pxy-miR-965-5p, pxy-miR-375-5p, pxy-miR-4448-3p, pxy-miR-279a-3p, pxy-miR-3286-3p, pxy-miR-8799-3p, pxy-miR-14b-5p and ASO for LncRNAs MSTRG.32910.1, MSTRG.7100.1, MSTRG.6802.1, and MSTRG.22113.1 were all synthesized by Gene Pharma (GenePharma, CHN) (Tables S2 and S6).
4.8. Total RNA extraction and real-time quantitative PCR (RT-qPCR)
Total RNA was isolated using Total RNA Kit Ⅱ (Omega Bio-Tek, USA). The cDNA of mRNA and LncRNA was synthesized using the Prime script RT Master Kit (Takara, JPN), while the cDNA of miRNA was synthesized using the Mir-XTM miRNA First-Strand Synthesis Kit (Takara, JPN), all in accordance with the respective manufacturer’s protocols. RT-qPCR was conducted on the CFX96 Real-Time PCR Detection System (Bio-Rad, USA) using TB Green® Premix Ex Taq™ II (Takara, JPN). The primers for RT-qPCR were represented in Table S5. The data was referenced to the mRNA internal control RPS13 [91] and miRNA internal control U6 [92] using CT methods [93].
4.9. The microinjection of P. xylostella
P. xylostella larvae (L3D2) were selected for microinjection of miRNA mimic 20 μM (0.2 μL) per larva using FemtoJet 4x (Eppendorf, GER), and 0.2 μL NC mimic was injected as control. Larvae of the same age were injected with LncRNA ASO 20 μM (0.2 μL) per larva, and 0.2 μL NC ASO was injected as a control.
4.10. Statistical analysis
All the figures were drawn by Origin2022 and TBtools [94]. All statistical analyses were performed by using SPSS Statistics 26 Software. All the experiments were conducted in triplicate with three biological repeats.
5. Conclusions
The infection of M. anisopliae induced the PO cascade of P. xylostella larvae, but non-coding RNA played a key role in the regulation. This study demonstrated that the miRNA pxy-miR-965-5p could regulate the PO cascade by targeting PAP1, thus affecting the effect of M. anisopliae on infecting P. xylostella. In addition, we also verified that miRNA pxy-miR-965-5p was negatively regulated by LncRNA MSTRG.7100.1 and MSTRG.22113.1. The application of M. anisopliae in the presence of pxy-miR-965-5p mimic suggested that it could augment significant susceptibility to M. anisopliae. pxy-miR-965-5p contributes to melanization to resist fungi by targeting PAP1 in P. xylostella, which can provide a new approach to insect pest control.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org. Figure S1: The target sites verification of pxy-miR-965-5p to PAP1; Figure S2: The interaction modules of LncRNAs with pxy-miR-965-5p; Table S1: Primers for protein expression in the E. coli expression system; Table S2: The sequences of miRNA predicted to target PAP1; Table S3: Primers for Dual-luciferase assay (miRNAs to PAP1); Table S4: Primers for Dual-luciferase assay (pxy-miR-965-5p to LncRNAs); Table S5: Primers for RT-qPCR; Table S6 The sequences of ASO for LncRNAs.
Author Contributions
Conceptualization, X.X., F.J.; methodology, Z.Z., J.H., L.Z.; validation, Z.Z.; formal analysis, Z.Z., F.J., J.Z.; resources, X.X., F.J.; writing—original draft preparation, Z.Z., X.X., F.J.; writing—review and editing, Z.Z., X.X., F.J., J.Z., S.D.M.; supervision, F.J., X.X.; funding acquisition, F.J. and X.X. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by grants from the National Natural Science Foundation of China (32172498,31972345), Natural Science Foundation of Guangdong, China (2023A1515010305).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
We are very grateful to the National Key Laboratory of Green Pesticide, China, for support. We also thank anonymous referees for their invaluable comments and suggestions.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Ponting, C.P.; Oliver, P.L.; Reik, W. Evolution and functions of long non-coding RNAs. Cell 2009, 136, 629–641. [Google Scholar] [CrossRef] [PubMed]
- Choudhary, C.; Sharma, S.; Meghwanshi, K.K.; Patel, S.; Mehta, P.; Shukla, N.; Do, D.N.; Rajpurohit, S.; Suravajhala, P.; Shukla, J.N. Long Non-Coding RNAs in Insects. Animals (Basel) 2021, 11, 1118. [Google Scholar] [CrossRef] [PubMed]
- Fernandes, J.C.R.; Acuña, S.M.; Aoki, J.I.; Floeter-Winter, L.M.; Muxel, S.M. Long non-coding RNAs in the regulation of gene expression: physiology and disease. Non-coding RNA 2019, 5, 17. [Google Scholar] [CrossRef] [PubMed]
- Gomes, A.Q.; Nolasco, S.; Soares, H. Non-coding RNAs: multi-tasking molecules in the cell. International journal of molecular sciences 2013, 14, 16010–16039. [Google Scholar] [CrossRef]
- Liu, F.; Shi, T.; Qi, L.; Su, X.; Wang, D.; Dong, J.; Huang, Z.Y. lncRNA profile of Apis mellifera and its possible role in behavioural transition from nurses to foragers. Bmc Genomics 2019, 20, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Zhu, B.; Li, L.; Wei, R.; Liang, P.; Gao, X. , Regulation of GSTu1-mediated insecticide resistance in Plutella xylostella by miRNA and lncRNA. Plos Genetics 2021, 17, e1009888. [Google Scholar] [CrossRef] [PubMed]
- Ylla, G.; Piulachs, M.-D.; Belles, X. Comparative analysis of miRNA expression during the development of insects of different metamorphosis modes and germ-band types. BMC genomics 2017, 18, 1–13. [Google Scholar] [CrossRef]
- Jiang, S.; Wu, H.; Liu, H.; Zheng, J.; Lin, Y.; Chen, H. The overexpression of insect endogenous small RNAs in transgenic rice inhibits growth and delays pupation of striped stem borer (Chilo suppressalis). Pest Management Science 2017, 73, 1453–1461. [Google Scholar] [CrossRef] [PubMed]
- Etebari, K.; Asgari, S. Conserved microRNA miR-8 blocks activation of the Toll pathway by upregulating Serpin 27 transcripts. RNA Biology 2013, 10, 1356–1364. [Google Scholar] [CrossRef]
- Yang, M.; Wang, Y.; Jiang, F.; Song, T.; Wang, H.; Liu, Q.; Zhang, J.; Zhang, J.; Kang, L. miR-71 and miR-263 Jointly Regulate Target Genes Chitin synthase and Chitinase to Control Locust Molting. Plos Genetics 2016, 12, e1006257. [Google Scholar] [CrossRef]
- Liu, Z.; Ling, L.; Xu, J.; Zeng, B.; Huang, Y.; Shang, P.; Tan, A. MicroRNA-14 regulates larval development time in Bombyx mori. Insect Biochemistry and Molecular Biology 2018, 93, 57–65. [Google Scholar] [CrossRef]
- Wang, Y.; Cui, C.; Wang, G.; Li, Y.; Wang, S. Insects defend against fungal infection by employing microRNAs to silence virulence-related genes. Proceedings of the National Academy of Sciences 2021, 118, e2023802118. [Google Scholar] [CrossRef] [PubMed]
- Fullaondo, A.; Lee, S.Y. Regulation of Drosophila-virus interaction. Developmental & Comparative Immunology 2012, 36, 262–266. [Google Scholar] [CrossRef] [PubMed]
- Lemaitre, B.; Hoffmann, J. The Host Defense of Drosophila melanogaster. Annual Review of Immunology 2007, 25, 697–743. [Google Scholar] [CrossRef] [PubMed]
- Chae, J.-H.; Kurokawa, K.; So, Y.-I.; Hwang, H.O.; Kim, M.-S.; Park, J.-W.; Jo, Y.-H.; Lee, Y.S.; Lee, B.L. Purification and characterization of tenecin 4, a new anti-Gram-negative bacterial peptide, from the beetle Tenebrio molitor. Developmental & Comparative Immunology 2012, 36, 540–546. [Google Scholar] [CrossRef]
- Sackton, T.B.; Lazzaro, B.P.; Schlenke, T.A.; Evans, J.D.; Hultmark, D.; Clark, A.G. Dynamic evolution of the innate immune system in Drosophila. Nature Genetics 2007, 39, 1461–1468. [Google Scholar] [CrossRef] [PubMed]
- Hampson, G.F. The Fauna of British India, including Ceylon and Burma. Moths. Volume 1. The Fauna of British India, including Ceylon and Burma. Moths. Volume 1. 1892. [Google Scholar]
- Ferrandon, D.; Imler, J.-L.; Hetru, C.; Hoffmann, J.A. The Drosophila systemic immune response: sensing and signalling during bacterial and fungal infections. Nature Reviews Immunology 2007, 7, 862–874. [Google Scholar] [CrossRef] [PubMed]
- Lazzaro, B.P. Natural selection on the Drosophila antimicrobial immune system. Current Opinion in Microbiology 2008, 11, 284–289. [Google Scholar] [CrossRef] [PubMed]
- Jiravanichpaisal, P.; Lee, B.L.; Söderhäll, K. Cell-mediated immunity in arthropods: Hematopoiesis, coagulation, melanization and opsonization. Immunobiology 2006, 211, 213–236. [Google Scholar] [CrossRef]
- Wang, Q.; Ren, M.; Liu, X.; Xia, H.; Chen, K. Peptidoglycan recognition proteins in insect immunity. Molecular Immunology 2019, 106, 69–76. [Google Scholar] [CrossRef]
- Chen, X.; Yu, J.; Wang, W.; Lu, H.; Kang, L.; Cui, F.; Lemaitre, B. A Plant Virus Ensures Viral Stability in the Hemolymph of Vector Insects through Suppressing Prophenoloxidase Activation. mBio 2020, 11, e01453–01420. [Google Scholar] [CrossRef] [PubMed]
- Li, B.; Li, H.; Tian, Y.; Abro, N.A.; Nong, X.; Zhang, Z.; Wang, G. Molecular Identification and Immunity Functional Characterization of Lmserpin1 in Locusta migratoria manilensis. Insects 2021, 12, 178. [Google Scholar] [CrossRef] [PubMed]
- Shi, M.; Chen, X.-Y.; Zhu, N.; Chen, X.-X. Molecular identification of two prophenoloxidase-activating proteases from the hemocytes of Plutella xylostella (Lepidoptera: Plutellidae) and their transcript abundance changes in response to microbial challenges. Journal of Insect Science 2014, 14, 179. [Google Scholar] [CrossRef] [PubMed]
- Zhou, L.; Lim, M.Y.T.; Kaur, P.; Saj, A.; Bortolamiol-Becet, D.; Gopal, V.; Tolwinski, N.; Tucker-Kellogg, G.; Okamura, K. Importance of miRNA stability and alternative primary miRNA isoforms in gene regulation during Drosophila development. Elife 2018, 7, e38389. [Google Scholar] [CrossRef]
- Zhao, A.; Yuan, X.; Hu, D.; Leng, C.; Li, Y.; Wang, P.; Li, Y. The effect of host plant on the development and larval midgut protease activity of Plutella xylostella (Lepidoptera: Plutellidae). Phytoparasitica 2019, 47, 475–483. [Google Scholar] [CrossRef]
- You, M.S.; Ke, F.S.; You, S.J.; Wu, Z.Y.; Liu, Q.F.; He, W.Y.; Baxter, S.W.; Yuchi, Z.G.; Vasseur, L.; Gurr, G.M. , et al. Variation among 532 genomes unveils the origin and evolutionary history of a global insect herbivore. Nature Communications 2020, 11, 2321. [Google Scholar] [CrossRef] [PubMed]
- Sarauer, B.L.; Gillott, C.; Hegedus, D. Characterization of an intestinal mucin from the peritrophic matrix of the diamondback moth, Plutella xylostella. Insect Molecular Biology 2003, 12, 333–343. [Google Scholar] [CrossRef] [PubMed]
- Zhu, X.; Lei, Y.; Yang, Y.; Baxter, S.W.; Li, J.; Wu, Q.; Wang, S.; Xie, W.; Guo, Z.; Fu, W. , et al. Construction and characterisation of near-isogenic Plutella xylostella (Lepidoptera: Plutellidae) strains resistant to Cry1Ac toxin. Pest Management Science 2015, 71, 225–233. [Google Scholar] [CrossRef] [PubMed]
- Islam, W.; Adnan, M.; Shabbir, A.; Naveed, H.; Abubakar, Y.S.; Qasim, M.; Tayyab, M.; Noman, A.; Nisar, M.S.; Khan, K.A. , et al. Insect-fungal-interactions: A detailed review on entomopathogenic fungi pathogenicity to combat insect pests. Microbial Pathogenesis 2021, 159, 105122. [Google Scholar] [CrossRef]
- Peng, Y.; Tang, J.; Xie, J. Transcriptomic Analysis of the Brown Planthopper, Nilaparvata lugens, at Different Stages after Metarhizium anisopliae Challenge. Insects (2075-4450) 2020, 11, 139. [Google Scholar] [CrossRef]
- Shin, T.Y.; Lee, M.R.; Park, S.E.; Lee, S.J.; Kim, W.J.; Kim, J.S. Pathogenesis-related genes of entomopathogenic fungi. Archives of Insect Biochemistry and Physiology 2020, 105, e21747. [Google Scholar] [CrossRef] [PubMed]
- Shoukat, R.F.; Hassan, B.; Shakeel, M.; Zafar, J.; Li, S.; Freed, S.; Xu, X.; Jin, F. Pathogenicity and Transgenerational Effects of Metarhizium anisopliae on the Demographic Parameters of Aedes albopictus (Culicidae: Diptera). J Med Entomol 2020, 57, 677–685. [Google Scholar] [CrossRef]
- Zafar, J.; Freed, S.; Khan, B.A.; Farooq, M. Effectiveness of Beauveria bassiana against cotton whitefly, Bemisia tabaci (Gennadius)(Aleyrodidae: Homoptera) on different host plants. Pak. J. Zool 2016, 48, 91–99. [Google Scholar]
- Mwamburi, L.A. Endophytic fungi, Beauveria bassiana and Metarhizium anisopliae, confer control of the fall armyworm, Spodoptera frugiperda (JE Smith)(Lepidoptera: Noctuidae), in two tomato varieties. Egyptian Journal of Biological Pest Control 2021, 31, 1–6. [Google Scholar] [CrossRef]
- Zafar, J.; Shoukat, R.F.; Zhang, Y.; Freed, S.; Xu, X.; Jin, F. Metarhizium Anisopliae Challenges Immunity and Demography of Plutella xylostella. Insects 2020, 11, 694. [Google Scholar] [CrossRef]
- Evans, J.; Aronstein, K.; Chen, Y.P.; Hetru, C.; Imler, J.L.; Jiang, H.; Kanost, M.; Thompson, G.; Zou, Z.; Hultmark, D. Immune pathways and defence mechanisms in honey bees Apis mellifera. Insect Mol Biol. 2006, 15, 645–656. [Google Scholar] [CrossRef]
- Muthu Lakshmi Bavithra, C.; Murugan, M.; Pavithran, S.; Naveena, K. , Enthralling genetic regulatory mechanisms meddling insecticide resistance development in insects: role of transcriptional and post-transcriptional events. Front Mol Biosci 2023, 10, 1257859. [Google Scholar] [CrossRef]
- Zhang, Q.; Dou, W.; Taning, C. N. T.; Smagghe, G.; Wang, J. J. , Regulatory roles of microRNAs in insect pests: prospective targets for insect pest control. Curr Opin Biotechnol 2021, 70, 158–166. [Google Scholar] [CrossRef]
- Zhang, Z.; Zhu, S.; De Mandal, S.; Gao, Y.; Yu, J.; Zeng, L.; Huang, J.; Zafar, J.; Jin, F.; Xu, X. Combined transcriptomic and proteomic analysis of developmental features in the immune system of Plutella xylostella during larva-to-adult metamorphosis. Genomics 2022, 114, 110381. [Google Scholar] [CrossRef]
- Lavine, M.D.; Strand, M.R. Insect hemocytes and their role in immunity. Insect Biochemistry and Molecular Biology 2002, 32, 1295–1309. [Google Scholar] [CrossRef]
- Jiang, H.; Vilcinskas, A.; Kanost, M.R. Immunity in Lepidopteran insects. In Invertebrate Immunity, Soderhall, K., Ed. 2010; Vol. 708, pp. 181-204.
- Cerenius, L.; Lee, B.L.; Söderhäll, K. The proPO-system: pros and cons for its role in invertebrate immunity. Trends in Immunology 2008, 29, 263–271. [Google Scholar] [CrossRef] [PubMed]
- An, C.J.; Ragan, E.J.; Kanost, M.R. Regulation of Manduca sexta hemolymph proteinase 8 by serpin-1 isoforms. Faseb Journal 2009, 23. [Google Scholar] [CrossRef]
- Meekins, D.A.; Kanost, M.R.; Michel, K. Serpins in arthropod biology. Seminars in Cell & Developmental Biology 2017, 62, 105–119. [Google Scholar] [CrossRef]
- Gao, G.; Xu, X.-X.; Yu, J.; Li, L.-M.; Ju, W.-Y.; Jin, F.-L.; Freed, S. Identification and molecular characterization of two serine proteases and their potential involvement in prophenoloxidase activation in Plutella xylostella. Archives of Insect Biochemistry and Physiology 2016, 93, 25–39. [Google Scholar] [CrossRef] [PubMed]
- Shakeel, M.; Xu, X.; De Mandal, S.; Jin, F. Role of serine protease inhibitors in insect-host-pathogen interactions. Archives of Insect Biochemistry and Physiology 2019, 102, e21556. [Google Scholar] [CrossRef] [PubMed]
- Yuan, C.; Xing, L.; Wang, M.; Wang, X.; Yin, M.; Wang, Q.; Hu, Z.; Zou, Z. Inhibition of melanization by serpin-5 and serpin-9 promotes baculovirus infection in cotton bollworm Helicoverpa armigera. Plos Pathogens 2017, 13, e1006645. [Google Scholar] [CrossRef] [PubMed]
- Han, P.F.; Fan, J.Q.; Liu, Y.; Cuthbertson, A.G.S.; Yan, S.Q.; Qiu, B.L.; Ren, S.X. RNAi-Mediated Knockdown of Serine Protease Inhibitor Genes Increases the Mortality of Plutella xylostella Challenged by Destruxin A. Plos One 2014, 9. [Google Scholar] [CrossRef] [PubMed]
- Dong, W.; Tang, B.Z.; Sonoda, S.; Liang, P.; Gao, X.W. Sequencing and characterization of two cDNAs putatively encoding prophenoloxidases in the diamondback moth, Plutella xylostella (L.) (Lepidoptera: Yponomeutidae). Applied Entomology and Zoology 2011, 46, 211–221. [Google Scholar] [CrossRef]
- Piao, S.; Song, Y.L.; Kim, J.H.; Park, S.Y.; Park, J.W.; Lee, B.L.; Oh, B.H.; Ha, N.C. Crystal structure of a clip-domain serine protease and functional roles of the clip domains. Embo Journal 2005, 24, 4404–4414. [Google Scholar] [CrossRef]
- Wang, Y.; Jiang, H. Purification and characterization of Manduca sexta serpin-6: a serine proteinase inhibitor that selectively inhibits prophenoloxidase-activating proteinase-3. Insect Biochemistry and Molecular Biology 2004, 34, 387–395. [Google Scholar] [CrossRef]
- Jiang, H.B.; Wang, Y.; Yu, X.Q.; Zhu, Y.F.; Kanost, M. Prophenoloxidase-activating proteinase-3 (PAP-3) from Manduca sexta hemolymph: a clip-domain serine proteinase regulated by serpin-1J and serine proteinase homologs. Insect Biochemistry and Molecular Biology 2003, 33, 1049–1060. [Google Scholar] [CrossRef] [PubMed]
- Gupta, S.; Wang, Y.; Jiang, H.B. Manduca sexta prophenoloxidase (proPO) activation requires proPO-activating proteinase (PAP) and serine proteinase homologs (SPHs) simultaneously. Insect Biochemistry and Molecular Biology 2005, 35, 241–248. [Google Scholar] [CrossRef] [PubMed]
- Jiang, H.B.; Wang, Y.; Yu, X.Q.; Kanost, M.R. Prophenoloxidase-activating proteinase-2 from hemolymph of Manduca sexta. Journal of Biological Chemistry 2003, 278, 3552–3561. [Google Scholar] [CrossRef] [PubMed]
- He, K.; Sun, Y.; Xiao, H.; Ge, C.; Li, F.; Han, Z. Multiple miRNAs jointly regulate the biosynthesis of ecdysteroid in the holometabolous insects, Chilo suppressalis. Rna 2017, 23, 1817–1833. [Google Scholar] [CrossRef] [PubMed]
- Jiang, J.; Ge, X.; Li, Z.; Wang, Y.; Song, Q.; Stanley, D.W.; Tan, A.; Huang, Y. MicroRNA-281 regulates the expression of ecdysone receptor (EcR) isoform B in the silkworm, Bombyx mori. Insect Biochem Mol Biol. 2013, 43, 692–700. [Google Scholar] [CrossRef]
- Fullaondo, A.; García-Sánchez, S.; Sanz-Parra, A.; Recio, E.; Lee, S.Y.; Gubb, D. Spn1 Regulates the GNBP3-Dependent Toll Signaling Pathway in Drosophila melanogaster. Molecular and Cellular Biology 2011, 31, 2960–2972. [Google Scholar] [CrossRef] [PubMed]
- Choi, I.K.; Hyun, S. Conserved microRNA miR-8 in fat body regulates innate immune homeostasis in Drosophila. Developmental & Comparative Immunology 2012, 37, 50–54. [Google Scholar] [CrossRef] [PubMed]
- Bhattacharya, A.; Cui, Y. Systematic Prediction of the Impacts of Mutations in MicroRNA Seed Sequences. Journal of Integrative Bioinformatics 2017, 14. [Google Scholar] [CrossRef] [PubMed]
- Salomon, W.E.; Jolly, Samson M.; Moore, Melissa J.; Zamore, Phillip D.; Serebrov, V. Single-Molecule Imaging Reveals that Argonaute Reshapes the Binding Properties of Its Nucleic Acid Guides. Cell 2015, 162, 84–95. [Google Scholar] [CrossRef]
- Schirle, N.T.; Sheu-Gruttadauria, J.; MacRae, I.J. Structural basis for microRNA targeting. Science 2014, 346, 608–613. [Google Scholar] [CrossRef]
- Brennecke, J.; Stark, A.; Russell, R.B.; Cohen, S.M. Principles of microRNA-target recognition. PLoS Biol 2005, 3, e85. [Google Scholar] [CrossRef] [PubMed]
- Doench, J.G.; Sharp, P.A. Specificity of microRNA target selection in translational repression. Genes Dev 2004, 18, 504–511. [Google Scholar] [CrossRef] [PubMed]
- Chipman, L.B.; Pasquinelli, A.E. miRNA Targeting: Growing beyond the Seed. Trends in Genetics 2019, 35, 215–222. [Google Scholar] [CrossRef] [PubMed]
- Duan, Y.; Veksler-Lublinsky, I.; Ambros, V. Critical contribution of 3′ non-seed base pairing to the in vivo function of the evolutionarily conserved let-7a microRNA. Cell Reports 2022, 39, 110745. [Google Scholar] [CrossRef] [PubMed]
- Vilcinskas, A. The role of epigenetics in host–parasite coevolution: lessons from the model host insects Galleria mellonella and Tribolium castaneum. Zoology 2016, 119, 273–280. [Google Scholar] [CrossRef] [PubMed]
- Skalsky, R.L.; Vanlandingham, D.L.; Scholle, F.; Higgs, S.; Cullen, B.R. Identification of microRNAs expressed in two mosquito vectors, Aedes albopictus and Culex quinquefasciatus. BMC Genomics 2010, 11, 119. [Google Scholar] [CrossRef] [PubMed]
- Mohr, A.M.; Mott, J.L. Overview of MicroRNA Biology. Semin Liver Dis 2015, 35, 003–011. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez, A.; Griffiths-Jones, S.; Ashurst, J.L.; Bradley, A. Identification of mammalian microRNA host genes and transcription units. Genome Research 2004, 14, 1902–1910. [Google Scholar] [CrossRef] [PubMed]
- Achkar, N.P.; Cambiagno, D.A.; Manavella, P.A. miRNA Biogenesis: A Dynamic Pathway. Trends in Plant Science 2016, 21, 1034–1044. [Google Scholar] [CrossRef]
- Chen, Z.M.; Liang, S.S.; Zhao, Y.; Han, Z. miR-92b regulates Mef2 levels through a negative-feedback circuit during Drosophila muscle development. Development 2012, 139, 3543–3552. [Google Scholar] [CrossRef]
- He, Q.; Zhang, Y.; Dong, W. MicroRNA miR-927 targets the juvenile hormone primary response geneKruppel homolog1to controlDrosophiladevelopmental growth. Insect Molecular Biology 2020, 29, 545–554. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.-L.; Wu, L.-X.; Li, H.-Y.; Wen, X.-Q.; Ma, E.-B.; Zhu, K.-Y.; Zhang, J.-Z. The microRNA miR-184 regulates the CYP303A1 transcript level to control molting of Locusta migratoria. Insect Science 2021, 28, 941–951. [Google Scholar] [CrossRef]
- Li, L.; Zhu, B.; Sun, X.; Zheng, K.; Liang, P.; Gao, X. miR-34-5p, a novel molecular target against lepidopteran pests. Journal of Pest Science, 2022. [Google Scholar] [CrossRef]
- Ma, L.; Liu, L.; Zhao, Y.J.; Yang, L.; Chen, C.H.; Li, Z.F.; Lu, Z.Q. JNK pathway plays a key role in the immune system of the pea aphid and is regulated by microRNA-184. Plos Pathogens 2020, 16. [Google Scholar] [CrossRef] [PubMed]
- Han, P.; Gong, Q.; Fan, J.; Zhang, M.; Abbas, M.; Zhu, W.; Deng, S.; Xing, S.; Zhang, J. 20-Hydroxyecdysone regulates the prophenoloxidase cascade to immunize Metarhizium anisopliae in Locusta migratoria. Pest Management Science 2020, 76, 3149–3158. [Google Scholar] [CrossRef]
- Wang, Q.; Yin, M.; Yuan, C.; Liu, X.; Hu, Z.; Zou, Z.; Wang, M. Identification of a Conserved Prophenoloxidase Activation Pathway in Cotton Bollworm Helicoverpa armigera. Frontiers in Immunology 2020, 11. [Google Scholar] [CrossRef]
- Sanbonmatsu, K.Y. Towards structural classification of long non-coding RNAs. Biochim Biophys Acta 2016, 1859, 41–45. [Google Scholar] [CrossRef]
- Long, Y.; Wang, X.; Youmans, D.T.; Cech, T.R. How do lncRNAs regulate transcription? Sci Adv 2017, 3, eaao2110. [Google Scholar] [CrossRef] [PubMed]
- Sanli, I.; Lalevée, S.; Cammisa, M.; Perrin, A.; Rage, F.; Llères, D.; Riccio, A.; Bertrand, E.; Feil, R. Meg3 Non-coding RNA Expression Controls Imprinting by Preventing Transcriptional Upregulation in cis. Cell Rep 2018, 23, 337–348. [Google Scholar] [CrossRef] [PubMed]
- Engreitz, J.M.; Ollikainen, N.; Guttman, M. Long non-coding RNAs: spatial amplifiers that control nuclear structure and gene expression. Nat Rev Mol Cell Biol 2016, 17, 756–770. [Google Scholar] [CrossRef]
- Kondo, Y.; Shinjo, K.; Katsushima, K. Long non-coding RNAs as an epigenetic regulator in human cancers. Cancer Sci 2017, 108, 1927–1933. [Google Scholar] [CrossRef]
- Wang, Y.; Fu, Y.; Cheng, H.; Zhao, C.; Huang, Q.; Chang, M.; Qiu, W.; Shen, Y.; Li, D. lncR26319/miR-2834/EndophilinA axis regulates oogenesis of the silkworm, Bombyx mori. Insect Sci 2023, 30, 65–80. [Google Scholar] [CrossRef] [PubMed]
- Drozd, V.S.; Eldeeb, A.A.; Kolpashchikov, D.M.; Nedorezova, D.D. Binary Antisense Oligonucleotide Agent for Cancer Marker-Dependent Degradation of Targeted RNA. Nucleic Acid Ther 2022, 32, 412–420. [Google Scholar] [CrossRef] [PubMed]
- Ashida, M. Purification and characterization of pre-phenoloxidase from hemolymph of the silkworm Bombyx mori. Archives of Biochemistry & Biophysics 1971, 144, 749–762. [Google Scholar]
- Shakeel, M.; Xu, X.; Xu, J.; Li, S.; Yu, J.; Zhou, X.; Xu, X.; Hu, Q.; Yu, X.; Jin, F. Genome-Wide Identification of Destruxin A-Responsive Immunity-Related MicroRNAs in Diamondback Moth, Plutella xylostella. Frontiers in Immunology 2018, 9, 185. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Xu, X.; Li, S.; Wang, S.; Xu, X.; Zhou, X.; Yu, J.; Yu, X.; Shakeel, M.; Jin, F. Genome-Wide Profiling of Plutella xylostella Immunity-Related miRNAs after Isaria fumosorosea Infection. Frontiers in Physiology 2017, 8, 1054. [Google Scholar] [CrossRef]
- Li, S.; Xu, X.; Zheng, Z.; Zheng, J.; Shakeel, M.; Jin, F. MicroRNA expression profiling of Plutella xylostella after challenge with B. thuringiensis. Developmental & Comparative Immunology 2019, 93, 115–124. [Google Scholar] [CrossRef] [PubMed]
- Zafar, J.; Zhang, Y.; Huang, J.; Freed, S.; Shoukat, R.F.; Xu, X.; Jin, F. Spatio-Temporal Profiling of Metarhizium anisopliae-Responsive microRNAs Involved in Modulation of Plutella xylostella Immunity and Development. Journal of fungi (Basel, Switzerland) 2021, 7. [Google Scholar] [CrossRef] [PubMed]
- Fu, W.; Xie, W.; Zhang, Z.; Wang, S.L.; Wu, Q.J.; Liu, Y.; Zhou, X.M.; Zhou, X.G.; Zhang, Y.J. Exploring Valid Reference Genes for Quantitative Real-time PCR Analysis in Plutella xylostella (Lepidoptera: Plutellidae). Int. J. Biol. Sci. 2013, 9, 792–802. [Google Scholar] [CrossRef]
- Yang, Y.; Li, Z.; Cao, J.; Li, Y.; Li, H.; Yang, Q.; Zhang, Q.; Liu, X. Identification and Evaluation of Suitable Reference Genes for Normalization of MicroRNA Expression in Helicoverpa armigera (Lepidoptera: Noctuidae) Using Quantitative Real-Time PCR. J Insect Sci 2017, 17. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of Relative Gene Expression Data Using Real-Time Quantitative PCR and the 2−ΔΔCT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Chen, C.J.; Chen, H.; Zhang, Y.; Thomas, H.R.; Frank, M.H.; He, Y.H.; Xia, R. TBtools: An Integrative Toolkit Developed for Interactive Analyses of Big Biological Data. Mol Plant 2020, 13, 1194–1202. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
The infection of P. xylostella by M. anisopliae. (A) The phenomena of infection. (B) The black patch in the integument of P. xylostella. (C) Determination of PO activity. “Control” indicates the group treated by conidia germinating medium, and “Ma” indicates the group infected by M. anisopliae. The column represented the PO activity, and data in the figure are mean ± SE (LSD and Duncan analysis: α = 0.05, n=3, letters on the columns indicate significant differences among groups, p < 0.05).
Figure 1.
The infection of P. xylostella by M. anisopliae. (A) The phenomena of infection. (B) The black patch in the integument of P. xylostella. (C) Determination of PO activity. “Control” indicates the group treated by conidia germinating medium, and “Ma” indicates the group infected by M. anisopliae. The column represented the PO activity, and data in the figure are mean ± SE (LSD and Duncan analysis: α = 0.05, n=3, letters on the columns indicate significant differences among groups, p < 0.05).
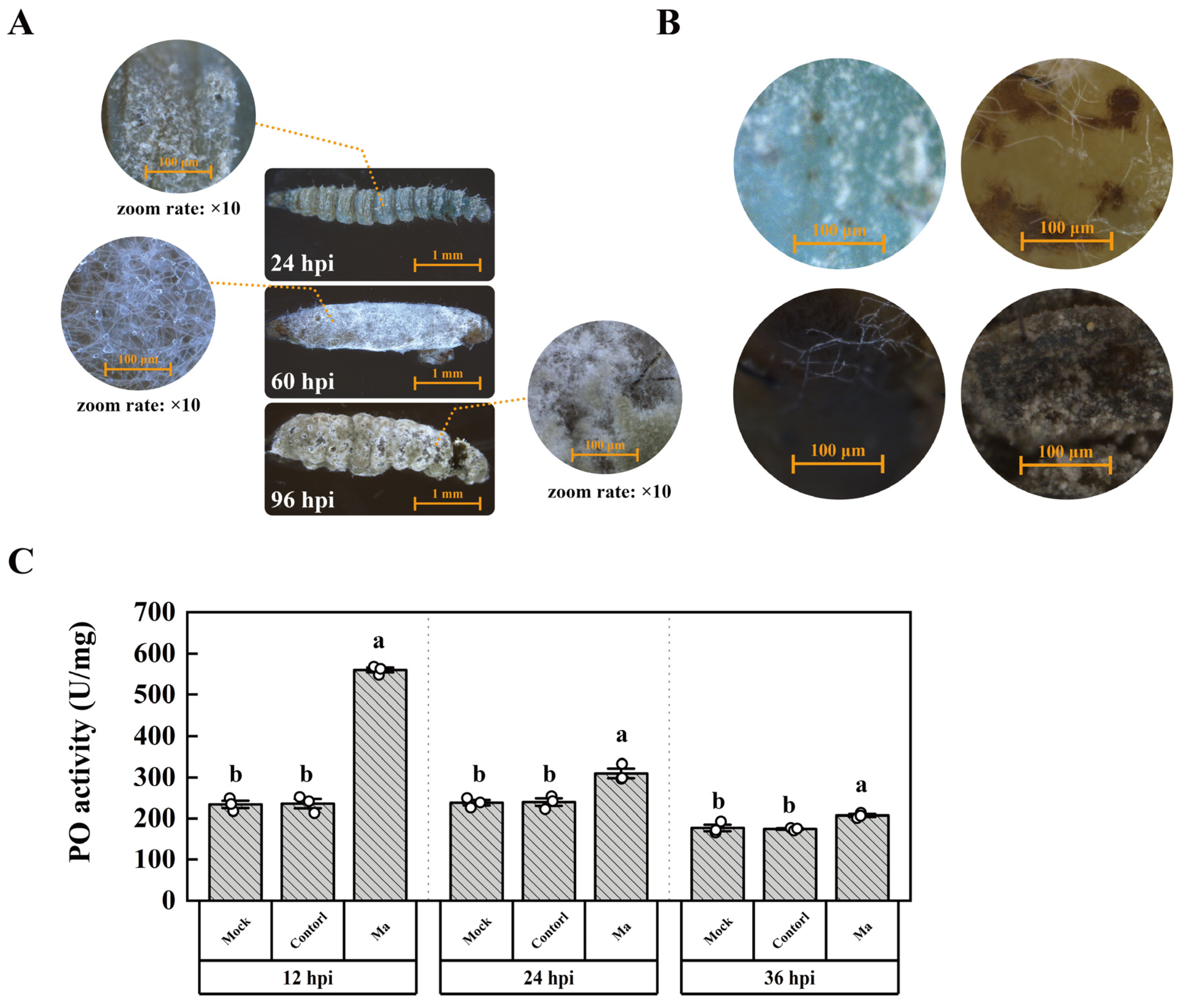
Figure 2.
PAP1 activated P. xylostella melanization in vitro. (A) The mode of PAP1 and IEGR mutant PAP1Xa. (B). The cleavage of PAP1Xa by Factor Xa, monoclonal antibody 6×His was used as the primary antibody, marker was on the left; protein PAP1Xa was on the line 57.3 kDa and protein activated PAP1Xa after cleaved was on line 29.4 kDa. (C) The enzyme activity of PAP1. The column represented the enzyme activity, data in the figure are mean ± SE (LSD and Duncan analysis: α = 0.05, n=3, letters on the columns indicate significant differences among groups, p < 0.05).
Figure 2.
PAP1 activated P. xylostella melanization in vitro. (A) The mode of PAP1 and IEGR mutant PAP1Xa. (B). The cleavage of PAP1Xa by Factor Xa, monoclonal antibody 6×His was used as the primary antibody, marker was on the left; protein PAP1Xa was on the line 57.3 kDa and protein activated PAP1Xa after cleaved was on line 29.4 kDa. (C) The enzyme activity of PAP1. The column represented the enzyme activity, data in the figure are mean ± SE (LSD and Duncan analysis: α = 0.05, n=3, letters on the columns indicate significant differences among groups, p < 0.05).
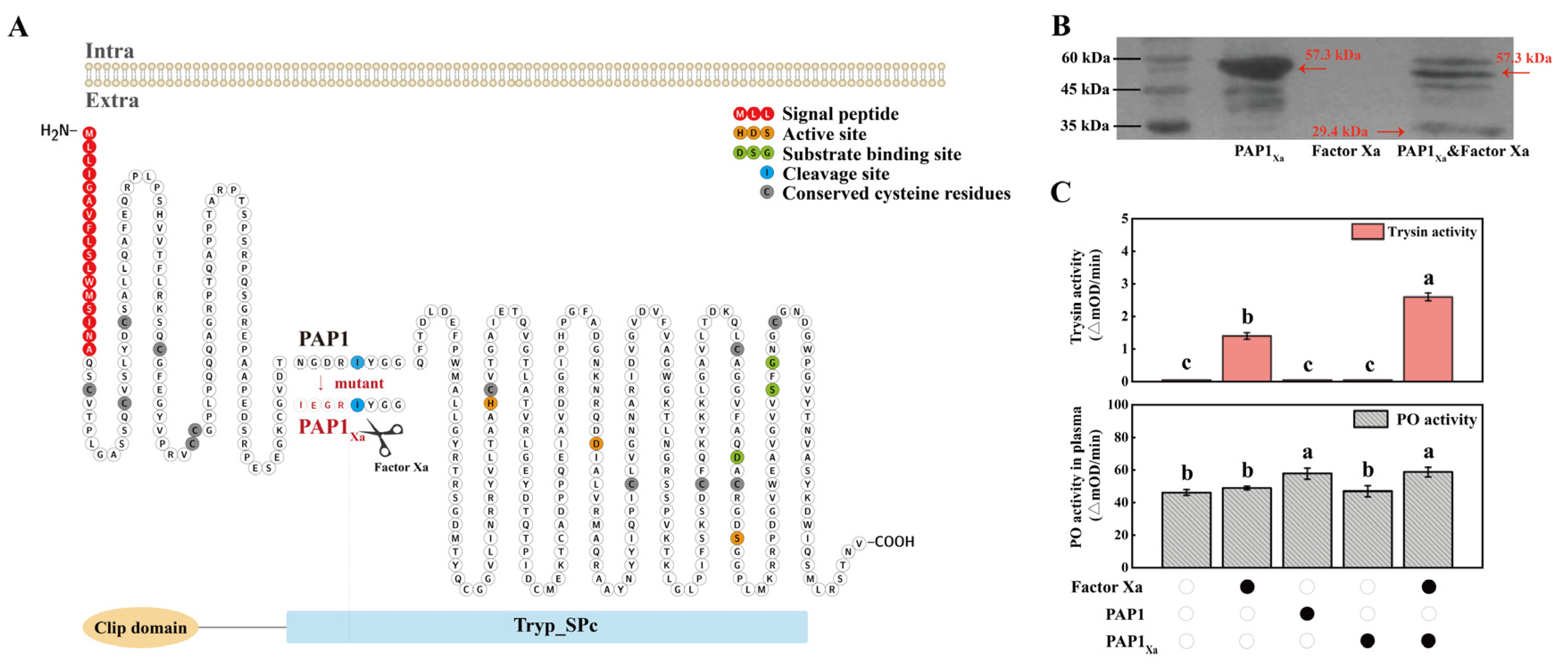
Figure 3.
The miRNA target prediction and verification of PAP1. (A) Sites on PAP1 targeted by miRNAs. 5’ UTR, 5’ Untranslated Region; ORF, Open Reading Frame; 3’ UTR, 3’ Untranslated Region. (B) The dual luciferase reporter assay of miRNAs to PAP1. The column represented the RFU (hRluc/hluc+) fold change of miRNA mimic treatment groups compared to negative control groups, data in the figure are mean ± SE (Two-tailed Student’s t-tests: α = 0.05, n=3, asterisks indicated significant differences between treatment and control groups, *p < 0.05, **p < 0.01, ***p < 0.001, NS = no significant differences).
Figure 3.
The miRNA target prediction and verification of PAP1. (A) Sites on PAP1 targeted by miRNAs. 5’ UTR, 5’ Untranslated Region; ORF, Open Reading Frame; 3’ UTR, 3’ Untranslated Region. (B) The dual luciferase reporter assay of miRNAs to PAP1. The column represented the RFU (hRluc/hluc+) fold change of miRNA mimic treatment groups compared to negative control groups, data in the figure are mean ± SE (Two-tailed Student’s t-tests: α = 0.05, n=3, asterisks indicated significant differences between treatment and control groups, *p < 0.05, **p < 0.01, ***p < 0.001, NS = no significant differences).
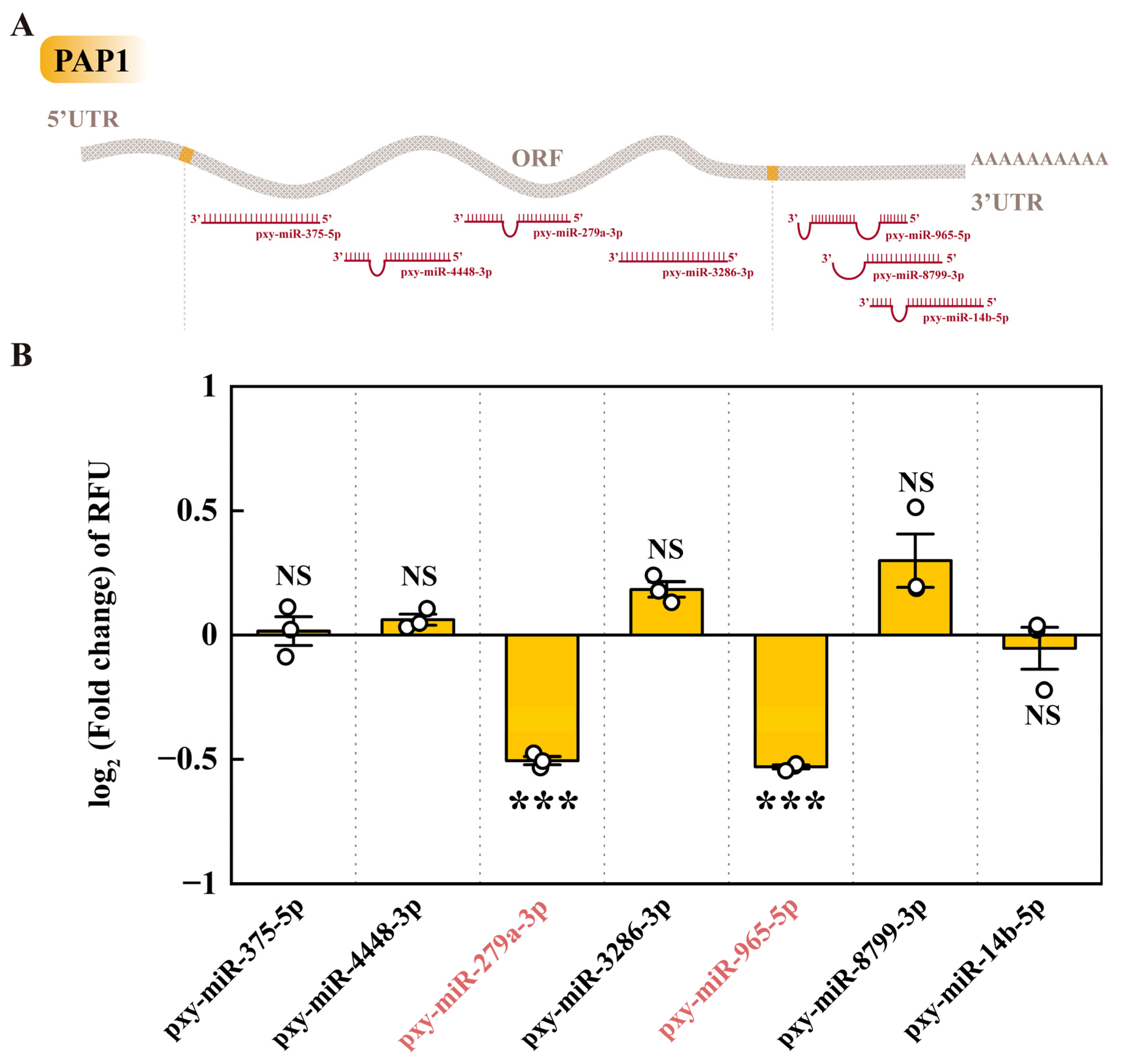
Figure 4.
The expression patterns of PAP1 and miRNA. (A) The expression patterns in different tissues. The column represented the relative expression of PAP1, pxy-miR-965-5p, and pxy-miR-279a-3p, data in the figure are mean ± SE (LSD and Duncan analysis: α = 0.05, n=3, letters on the columns indicated significant differences among different tissues, p < 0.05). (B) The expression patterns after M. anisopliae infection. The column represented the relative expression fold change of treatment groups compared to control groups; data in the figure are mean ± SE (Two-tailed Student’s t-tests: α = 0.05, n=3, asterisks indicated significant differences between treatment and control groups, *p < 0.05, **p < 0.01, ***p < 0.001, NS = no significant differences).
Figure 4.
The expression patterns of PAP1 and miRNA. (A) The expression patterns in different tissues. The column represented the relative expression of PAP1, pxy-miR-965-5p, and pxy-miR-279a-3p, data in the figure are mean ± SE (LSD and Duncan analysis: α = 0.05, n=3, letters on the columns indicated significant differences among different tissues, p < 0.05). (B) The expression patterns after M. anisopliae infection. The column represented the relative expression fold change of treatment groups compared to control groups; data in the figure are mean ± SE (Two-tailed Student’s t-tests: α = 0.05, n=3, asterisks indicated significant differences between treatment and control groups, *p < 0.05, **p < 0.01, ***p < 0.001, NS = no significant differences).
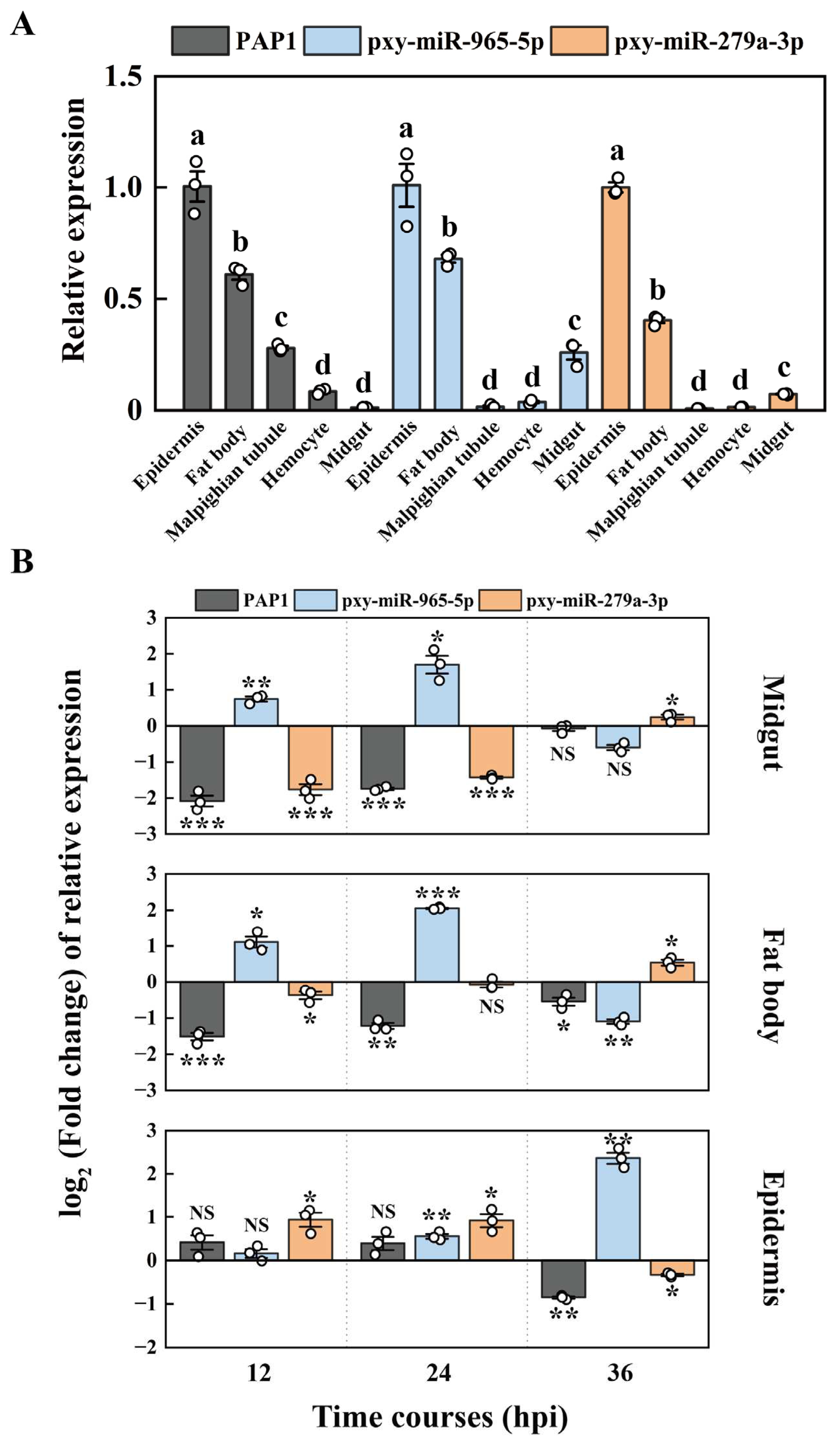
Figure 5.
The expression of PAP1 in P. xylostella under miRNA treatments. (A) The expression of PAP1 under pxy-miR-965-5p treatments. Ma, the group infected by M. anisopliae; Ma + NC, the group infected by M. anisopliae after mimics negative control injection; Ma + mimic, the group infected by M. anisopliae after pxy-miR-965-5p mimic injection; Ma + inhibitor, the group infected by M. anisopliae after pxy-miR-965-5p inhibitor injection. The column represented the relative expression of PAP1, data in the figure are mean ± SE (LSD and Duncan analysis: α = 0.05, n=3, letters on the columns indicate significant differences among groups, p < 0.05). (B) The expression of PAP1 under pxy-miR-279a-3p treatments. Ma, the group infected by M. anisopliae; Ma + NC, the group infected by M. anisopliae after mimics negative control injection; Ma + mimic, the group infected by M. anisopliae after pxy-miR-279a-3p mimic injection; Ma + inhibitor, the group infected by M. anisopliae after pxy-miR-279a-3p inhibitor injection. The column represented the relative expression of PAP1, data in the figure are mean ± SE (LSD and Duncan analysis: α = 0.05, n=3, letters on the columns indicate significant differences among groups, p < 0.05).
Figure 5.
The expression of PAP1 in P. xylostella under miRNA treatments. (A) The expression of PAP1 under pxy-miR-965-5p treatments. Ma, the group infected by M. anisopliae; Ma + NC, the group infected by M. anisopliae after mimics negative control injection; Ma + mimic, the group infected by M. anisopliae after pxy-miR-965-5p mimic injection; Ma + inhibitor, the group infected by M. anisopliae after pxy-miR-965-5p inhibitor injection. The column represented the relative expression of PAP1, data in the figure are mean ± SE (LSD and Duncan analysis: α = 0.05, n=3, letters on the columns indicate significant differences among groups, p < 0.05). (B) The expression of PAP1 under pxy-miR-279a-3p treatments. Ma, the group infected by M. anisopliae; Ma + NC, the group infected by M. anisopliae after mimics negative control injection; Ma + mimic, the group infected by M. anisopliae after pxy-miR-279a-3p mimic injection; Ma + inhibitor, the group infected by M. anisopliae after pxy-miR-279a-3p inhibitor injection. The column represented the relative expression of PAP1, data in the figure are mean ± SE (LSD and Duncan analysis: α = 0.05, n=3, letters on the columns indicate significant differences among groups, p < 0.05).
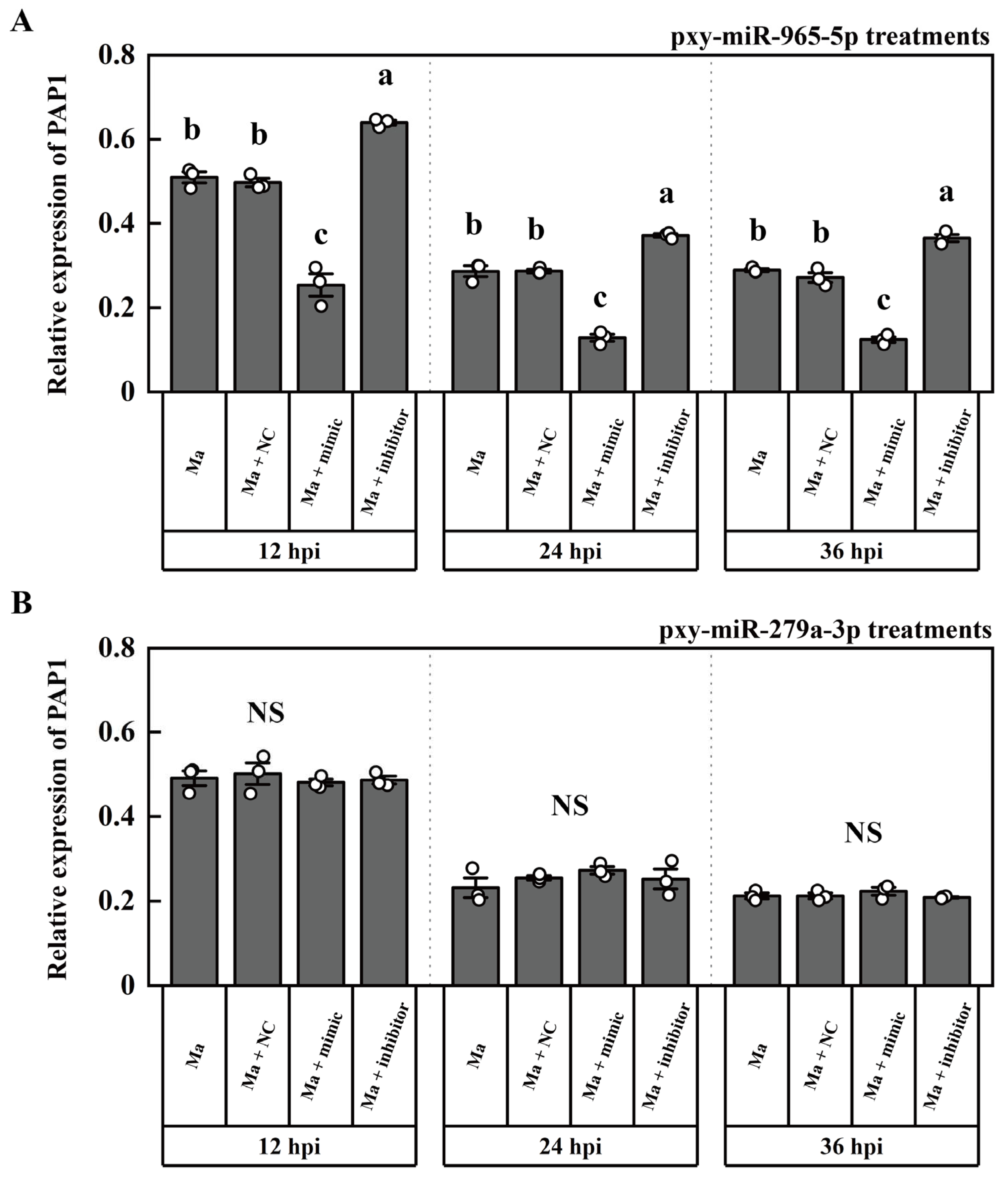
Figure 6.
Survival rates and PO activity of P. xylostella under miRNA treatments. (A) Survival rates of P. xylostella under miRNA treatments. Ma, the group infected by M. anisopliae; Ma + NC, the group infected by M. anisopliae after mimics negative control injection; Ma + pxy-miR-965-5p mimic, the group infected by M. anisopliae after pxy-miR-965-5p mimic injection; Ma + pxy-miR-965-5p inhibitor, the group infected by M. anisopliae after pxy-miR-965-5p inhibitor injection. The curve represented the survival rates (Log-rank test: α = 0.05, n=100, letters on the columns indicate significant differences among groups, p < 0.05). (B) PO activity of P. xylostella under miRNA treatments. (B) PO activity of P. xylostella under miRNA treatments. Ma, the group infected by M. anisopliae; Ma + NC, the group infected by M. anisopliae after mimics negative control injection; Ma + mimic, the group infected by M. anisopliae after pxy-miR-965-5p mimic injection; Ma + inhibitor, the group infected by M. anisopliae after pxy-miR-965-5p inhibitor injection. The column represented the PO activity, data in the figure are mean ± SE (LSD and Duncan analysis: α = 0.05, n=3, letters on the columns indicate significant differences among groups, p < 0.05).
Figure 6.
Survival rates and PO activity of P. xylostella under miRNA treatments. (A) Survival rates of P. xylostella under miRNA treatments. Ma, the group infected by M. anisopliae; Ma + NC, the group infected by M. anisopliae after mimics negative control injection; Ma + pxy-miR-965-5p mimic, the group infected by M. anisopliae after pxy-miR-965-5p mimic injection; Ma + pxy-miR-965-5p inhibitor, the group infected by M. anisopliae after pxy-miR-965-5p inhibitor injection. The curve represented the survival rates (Log-rank test: α = 0.05, n=100, letters on the columns indicate significant differences among groups, p < 0.05). (B) PO activity of P. xylostella under miRNA treatments. (B) PO activity of P. xylostella under miRNA treatments. Ma, the group infected by M. anisopliae; Ma + NC, the group infected by M. anisopliae after mimics negative control injection; Ma + mimic, the group infected by M. anisopliae after pxy-miR-965-5p mimic injection; Ma + inhibitor, the group infected by M. anisopliae after pxy-miR-965-5p inhibitor injection. The column represented the PO activity, data in the figure are mean ± SE (LSD and Duncan analysis: α = 0.05, n=3, letters on the columns indicate significant differences among groups, p < 0.05).
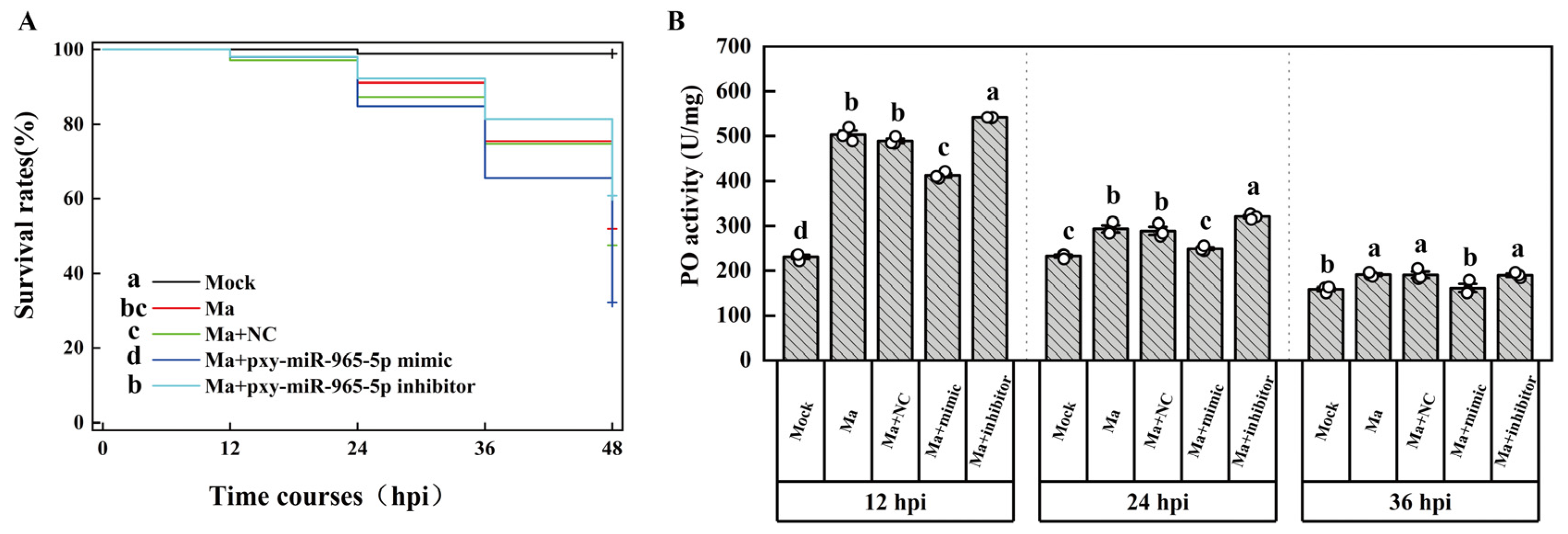
Figure 7.
The dual luciferase reporter assay of LncRNA to pxy-miR-965-5p. The column represented the RFU (hRluc/hluc+) fold change of miRNA mimic treatment groups compared to negative control groups, data in the figure are mean ± SE (Two-tailed Student’s t-tests: α = 0.05, n=3, asterisks indicated significant differences between treatment and control groups, **p < 0.01).
Figure 7.
The dual luciferase reporter assay of LncRNA to pxy-miR-965-5p. The column represented the RFU (hRluc/hluc+) fold change of miRNA mimic treatment groups compared to negative control groups, data in the figure are mean ± SE (Two-tailed Student’s t-tests: α = 0.05, n=3, asterisks indicated significant differences between treatment and control groups, **p < 0.01).
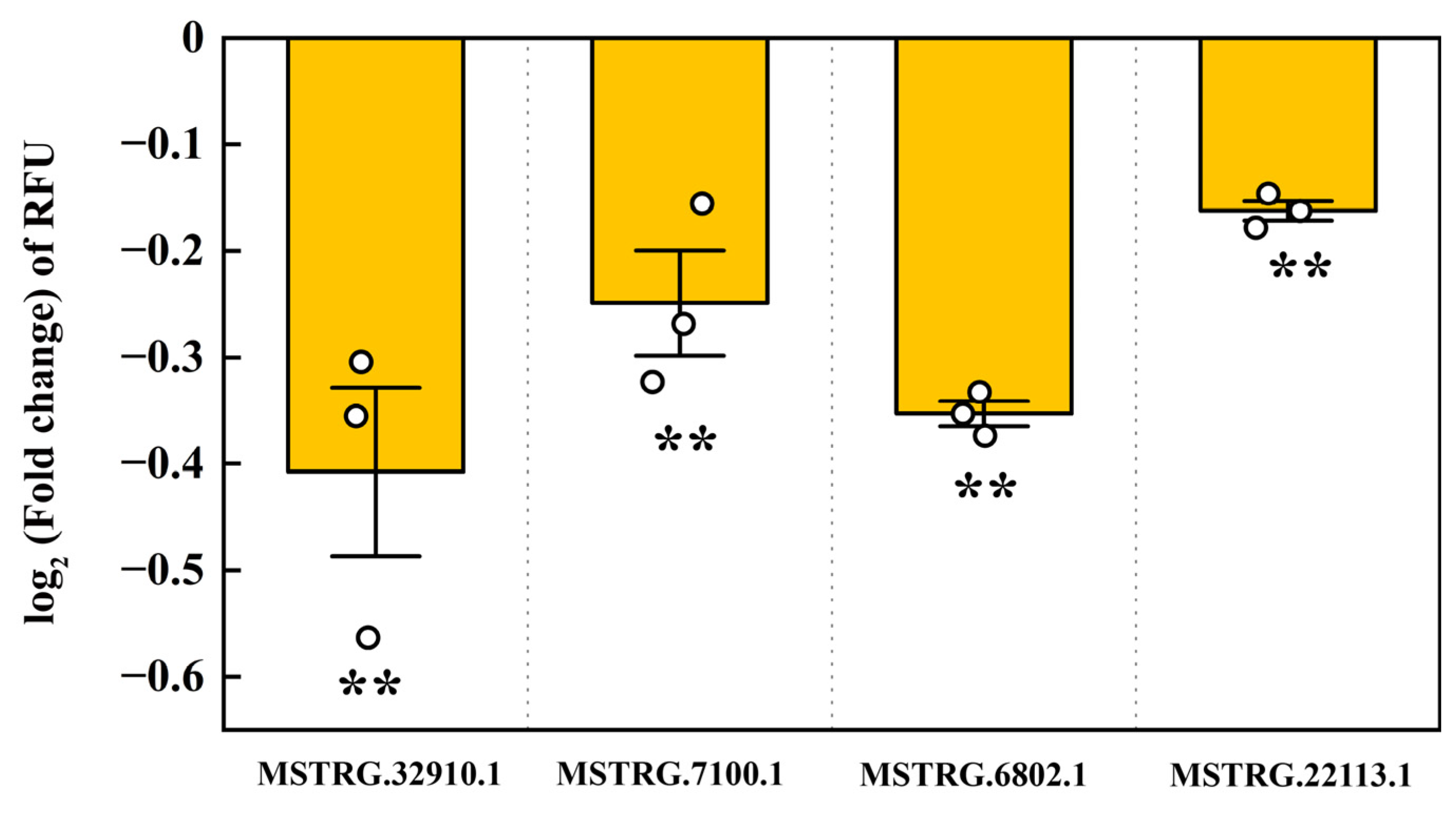
Figure 8.
The expression of LncRNA and miRNA in P. xylostella after ASO injection. (A) The expression of LncRNA after ASO injection. Ma, the group infected by M. anisopliae; Ma + NC, the group infected by M. anisopliae after ASO negative control injection; Ma + ASO, the group infected by M. anisopliae after ASO injection. The column represented the relative expression of LncRNA; data in figure are mean ± SE (LSD and Duncan analysis: α = 0.05, n=3, letters on the columns indicate significant differences among groups, p < 0.05). (B) The expression of pxy-miR-965-5p after ASO injection. Ma, the group infected by M. anisopliae; Ma + NC, the group infected by M. anisopliae after ASO negative control injection; Ma + ASO, the group infected by M. anisopliae after ASO injection. The column represented the relative expression of LncRNA; data in the figure are mean ± SE (LSD and Duncan analysis: α= 0.05, n=3, letters on the columns indicate significant differences among groups, p < 0.05).
Figure 8.
The expression of LncRNA and miRNA in P. xylostella after ASO injection. (A) The expression of LncRNA after ASO injection. Ma, the group infected by M. anisopliae; Ma + NC, the group infected by M. anisopliae after ASO negative control injection; Ma + ASO, the group infected by M. anisopliae after ASO injection. The column represented the relative expression of LncRNA; data in figure are mean ± SE (LSD and Duncan analysis: α = 0.05, n=3, letters on the columns indicate significant differences among groups, p < 0.05). (B) The expression of pxy-miR-965-5p after ASO injection. Ma, the group infected by M. anisopliae; Ma + NC, the group infected by M. anisopliae after ASO negative control injection; Ma + ASO, the group infected by M. anisopliae after ASO injection. The column represented the relative expression of LncRNA; data in the figure are mean ± SE (LSD and Duncan analysis: α= 0.05, n=3, letters on the columns indicate significant differences among groups, p < 0.05).
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



