
Submitted:
20 December 2023
Posted:
20 December 2023
You are already at the latest version
Abstract
Trees, often referred to as the "lungs of the Earth," hold immense significance for both the natural world and human society. Their contributions span a multitude of ecological, environmental, economic, and social domains, making them essential components of a healthy and thriving planet. Phytoplasma association not only compromise tree aesthetics but also reduce photosynthetic capacity and overall vigour. Phytoplasma diseases also lead to increased tree mortality, altering ecosystem dynamics.This review article provides an in-depth exploration of phytoplasma diseases affecting trees worldwide, encompassing their epidemiology, symptomatology, impact and management strategies.
Keywords:
trees
; forest
; fruits
; phytoplasma
; symptoms
; worldwide
; economic losses
; top of form
Introduction
Out of the approximate 350,000 species of plants that exist, the roughly 59,000 documented types of trees (BGCI), [1] make up the majority of biomass and play a crucial role as ecosystem architects, providing shelter and sustenance for countless species [2,3,4]. Moreover, trees serve as invaluable assets to humanity, supporting the well-being of virtually every individual through their provision of resources such as timber, sustenance, shade, fuel, building materials, and carbon storage [5,6].Mollicutes are a special group of bacteria that are wall-less and unculturable in-vitro. Phytoplasma is one of the important mollicute that causes several hundreds of plant diseases worldwide [8]. It shows wide host range from grasses, herbs, and shrubs to higher plants. In recent years the association of phytoplasma with trees has increased in number and shows devastating effects either in timber production as well as yield losses in fruit trees. Although phytoplasma is well known for its typical symptoms in diseased hosts such as, witches’ broom, little leaf, yellowing, stunted growth, phyllody and virescence [8].All phytoplasmas are transmitted with the aid of phloem-feeding insects, in most cases leafhoppers, planthoppers, and psyllids. After 19th century, with the development of advanced technology, the identification and its characterization have been largely envisaged and proposed. Because of its non-culturable nature, the phytoplasmas genus is designated as ‘Candidatus Phytoplasma’[9]. And till date,forty four phytoplasma groups have been proposed mainly based on PCR analysis, restriction Polymorphism and sequence analysis.[10].
In the last few decades, several new technologies for the detection of plant pathogens have evolved butthe important thing is that these are rigorously authenticated regardingsensitivity and specificity, not only withpure DNA samples or pure cultures but also with diseased plant samples associated with the target plant pathogen. Before using a new technique in practice, it should be compared to more conventional methods and assessed for cost-effectiveness [11]. The current approaches to managing phytoplasmoses focus on producing planting material free of pathogens, eradication programmes, quarantine restrictions, and chemical control of insect vectors. Sensitive diagnostic instruments and a fundamental understanding of phytoplasma presence in fruit trees and their potential vectors in fruit-growing areas are prerequisites for the adoption of these procedures.
This review article provides an in-depth exploration of phytoplasma diseases affecting trees worldwide, encompassing their epidemiology, symptomatology, impact and management strategies.
A. Phytoplasmas Associated with Fruit Trees
Phytoplasma detection in fruit crops and in insect vectors is now mainly based on molecular tools as for all phytoplasma diseases. For a number of these species grown in temperate areas, phytoplasma-specific protocols that allow sensitive andreliable detection were developed and validated. Phytoplasmas have small genomes missing major metabolic pathways. More recent efforts of scientists have centered at the characterization of phytoplasma virulence factors. Proteins expressed via plasmids and plentiful cells floor proteins are virulence factors concerned in insect transmission. Phytoplasmas use the Sec-established protein translocation device for secretion of virulence elements, which includes effector proteins, into plant and insect hosts. several effector proteins of the AY-WB phytoplasma target nuclei of plant cells and are probably concerned with the manipulation of plant tactics. Extending the concept of effectors to the phytoplasmasfactors to novel studies strategies for unravelling pathogenicity mechanisms of those fascinating pathogens [12]. Phytoplasmas in the 16SrX group, specific primers [13,14] that usually also require Restriction Fragment Length Polymorphism analysis of the amplified fragments can be employed. Specific primer pairs are also available that target non-ribosomal genes [15]. Several qPCR assays based either on SybrGreen® [16,17] or TaqMan® [18,19,20] technologies have been developed; group specific qPCR systems [21] can also be employed. Finally, a LAMP-based technique has been developed recently [22]. Latterly, the low cost Illumina sequencing technology,is widely used and more than thirty phytoplasma draft genome sequences were published [23].
In the pome and stone fruits, several other non species-specific phytoplasmas were detected worldwide as summarized below:
1. Pome Fruit
Pome fruits belong to the botanical family Rosaceae, specifically the sub-family pomoideae. These fruits are characterized by their central core, which contains multiple small seeds, enveloped by a resilient membrane. This membrane is further enclosed within a layer of edible flesh. The trees bearing pome fruits are deciduous and undergo a period of dormancy during winter, necessitating cold temperatures for the tree to successfully emerge from dormancy in the spring season. The main phytoplasma historically linked with diseases in apples and pears are ‘Candidatus Phytoplasma mali’ and ‘Ca. Phytoplasma pyri’, respectively [24]. They both belong to 16SrX-A and 16SrX-C subgroups [25].Due to the ecological complexity of plant host and insect vectors in the biological cycle of ‘Ca. Phytoplasma mali’, various studies have been conducted focusing on the the identification of molecular markers prioritizing the distinction of strains linked to specific biological features. Furthermore, nucleotide sequence analyses of multiple genomic regions appear as a significant tool in identifying genetic diversity within ‘Ca. Phytoplasma mali’ populations [26,27]. The whole genome of ‘Ca. P. mali’ has already been sequenced, while for ‘Ca. Phytoplasma pyri’ it was estimated to be linear, as reported for the other available phytoplasma genomes. Full-length genome sequencing provides an access to identify genes coding AA`A+ ATPases and HflB/FstH proteases, suitable for distinguishing closely related strains that display a wide range of virulence during the infection [28].
1.1. Apple
Apple (Malus domestica; Family: Rosaceae) trees yield a wide variety of apple cultivars, which are consumed fresh, used in cooking, pressed for apple juice and cider, and made into pies and sauces. They are also utilized for the making of apple-based products like applesauce and apple butter.
Apple proliferation is a devastating disease affecting apple trees (Malus domestica Borkh.) leading to huge economic losses in apple production. Apple's proliferation (AP) disease is well known to cause a huge economic loss in some of the biggest apple growing regions in Europe of about € 50 mill. in the high-incident years 2006 and 2013 [29].'Ca. Phytoplasma mali' is a phytoplasma associated with Proliferation of Apple disease that led to significant yield losses in as grow apples for commercial use[24].
Diseased trees show symptoms of small fruits with pale colour that appear unripe [30]. Few other symptoms of Apple proliferation (AP) are witches’ brooms,early bud break in spring, premature reddening of the foliage, late bloom, chlorosis, and enlarged stipules with dents [31]. Enlarged stipules and witches’ brooms appear to be the most significant symptoms for apple proliferation detection in apple orchads. Witches’ brooms arise during the summers and are most clearly visible in autumn. So far, no curative treatment for AP disease is reported. Hence, the disease spread could be possibly checked only by phytosanitary measures and eradication of diseased trees.However, the most efficient method for determining a tree's infection status at any point throughout the vegetative season is the detection of pathogen in tree roots using a (PCR) based method that detects phytoplasma-specific DNA. Spectroscopic techniques in combination with multivariate data analysis and statistical approaches are few trustworthy tools providing economical, efficient, fast and non-destructive detection of plant diseases [31]. Apple proliferation (AP) disease is related to ‘Ca. P. mali’, one of the most important diseases of apple trees. In case of intensive cultivation, disease is the main limiting factor for the crop sustainability. ‘Ca. P. mali’ is considered as a quarantine pathogen for both EPPO in Europe and NAPPO in North America, it weakens the productivity of trees whose fruit cannot be commercialized because of their poor organoleptic qualities.AP disease was first described in 1950 in Italy and has been reported in several other countries in European region such as, Norway, Germany, Czech Republic, Albania, Romania, Slovenia, Switzerland, Austria, Bulgaria, Croatia, France, Greece, Hungary, Moldova, the Netherlands, Poland, Southern Russia, Serbia and Montenegro, Slovakia, Spain, and Ukraine. In Asia, AP disease is only reported in Turkey. The epidemics of the disease is studied in various Italian northern regions that demonstrate Malus domestica cultivars as the main hosts of ‘Ca. Phytoplasma mali’ and the majority of cultivars being susceptible. The reported host range include, periwinkle, oriental lily, apricot, pear, apple, bindweed, hazelnut, Bermuda grass, magnolia, cherry, plum, Japanese plum and rose [32,33]. In extreme conditions, AP can result in productivity losses ranging from 50% to 80%, contingent upon the type, rootstock, surrounding circumstances, and agronomic management practices employed [34]. The most characteristic symptom is proliferation of the buds, which generally appear near the apex of the plant as a result of axillary buds’ development.The initial symptom develops a year after the infection, when the presence of phytoplasma encourages the formation of secondary shoots that are sensitive to powdery mildew and subsequently to witches’ brooms. The disease leads to loss of vitality and the flowers exhibit phyllody in spring. The leaves are smaller, roll downward, and become brittle; also, they are delicate, show irregularly serrations, sometimes display signs of early autumn.Psyllids transfer "Ca. P. mali" from symptomatic plants to healthy plant species. The main vector of "Ca. Phytoplasma mali" in Northwestern areas of Italy is Cacopsyllamelanoneura (Förster) [35], whereas the main vector of "Ca. P. mali" in Germany is C. picta (Förster) [36,37] in the northeastern areas [38]. The disease shows graft transmission with varying efficacy rates; moreover, the transmission rates vary up to 30% in response to the seasonal variation in colonization of aerial parts of apple trees. The tools available for proper management of the disease are not reliable and effective for apple proliferation. Therefore, in specific areas of interest, regular application of pesticides is essential for management of insect vectors in addition to eradication of infected symptomatic plants.
1.2. Pear
Pear Pyrus communisdecline is an important disease on pear trees described in Europe, North America, South America, Middle East, Chile and also Taiwan [39,40,41,42]. Pear decline (PD) is a devastating disease of pear (Pyrus communis L.) caused by 'Candidatus Phytoplasma pyri', which assigned to the subgroup 16SrX-C of the apple proliferation (AP) group of phytoplasmas first time reported in Canada [42]. The significance of "Ca. P. pyri" in pear production is continuously increasing, and there has been a proposal to regulate it as a quarantine organism in the European Union's A2 List (EPPO Global Database 2009). The Ca. P. pyri, transmitted by pear psylla, causes pear decline. Cacopsylla bidens, which may be another vector of PD in Bulgaria, has been found to carry "Ca. P. pyri" [43]. This phytoplasma can also be transmitted through grafting and budding. Trees with rootstocks of Pyrus ussuriensis or P. pyrifolia are more susceptible to decline than those propagated on domestic P. communis roots. Pear decline is characterized by two phases: quick decline and slow decline. Infected trees may wilt, scorch, and die in a few weeks, or lose vigor over several seasons, with sparse foliage and little or no terminal growth. Leaves turn red early due to starch accumulation in the upper tree. Yellowing symptoms , reddening, loss of vigor, early blooming, and, in some cases, a general collapse are exhibited by pear trees infected by this pathogen. The rootstock and the stage of the disease significantly influence disease development and symptoms expressed [44]. "Ca. P. pyri" has also been associated with peach yellow leaf roll (PYLR) diseases in North America, Europe, and Argentina [39]
Among the genes of pears involved in the metabolism of carbohydrates examined in this study, in PD-infected leaves of selection 8824-1 a significant up-regulated expression of both a sucrose synthase and an acid invertase gene was noticed , even though the PD population was similar to that of selection 9328-1 in leaves. The study selected pear sucrose synthase and acid invertase genes based on their up-regulation in grapevine leaves infected with FD, BN, and Stolbur phytoplasmas, as well as periwinkle and tomato leaves infected by Stolbur phytoplasma. The cleavage of sucrose by these enzymes results in fructose and UDP-glucose or fructose and glucose. The localized increase in sucrose synthase and vacuolar invertase can lead to a decrease in sucrose levels and an upgradation in fructose and glucose levels. Phytoplasmas require an upgrade in host sucrose synthase and vacuolar invertase gene expression due to their inability to import and phosphorylate sugars, such as sucrose, glucose, and fructose, as they lack the enzymes in phosphotransferase systems.Thus, increasing sucrose synthase and vacuolar invertase activities in host cells will allow for greater breakdown of host sucrose into fructose and glucose for phytoplasma growth [45,46].
2. Stone Fruit
A global report by Zirak et al.[47] revealed that stone fruit trees are vulnerable to numerous phytoplasma strains, which are classified under at least 14 distinct 'Candidatus Phytoplasma species'.Stone fruits are extensively cultivated worldwide, occupying a vast area of 5.07 million hectares and yielding a production of 35.24 million tons. Prunus is a genus of trees and shrubs, which includes the fruits cherries, peaches, plums, nectarines, apricots, and almonds. Native to the North American temperate regions, the neotropics of South America, and the paleotropics of Asia and Africa, [48]. In India, a total of 43,000 hectares are dedicated to the cultivation of apricots, peaches and nectarines, plums, and cherries, with an annual production of approximately 0.25 million tons (FAO, 2008). The cultivation of these fruits can be leveraged for profitable purposes, particularly through the production of fruit wines, which can generate employment opportunities and provide better returns to the orchardists [49,50].
Prunus sp. are associated with several phytoplasmas diseases among which European stone fruit yellows (ESFY) disease is the most frequent and is caused by ‘Candidatus Phytoplasma prunorum’. European stone fruit yellows disease causes huge economic losses in Prunus species throughout Asia and Europe. Phytoplasma associated stone fruit diseases for example Diseases of stone fruits associated with phytoplasma infection including apricot chlorotic leaf roll, Plum leptonecrosis peach yellowing and declining of plum , almond and peach [51,52,53,54], were detected to have a similar epidemiology. That’s why for all stone fruit diseases a common name was proposed as European stone fruit yellows (ESFY) disease [55]. ‘Candidatus Phytoplasma prunorum’, is the main causal organism of stone fruits diseases [24]. European and Mediterranean Plant Protection Organization (EPPO) included this phytoplasma agent as quarantine pathogen due to huge losses caused by this pathogen in fruit trees worldwide. Cultivated and wild Prunus sp. act as the natural hosts of ‘Candidatus Phytoplasma prunorum’ [54,56,57,58]. Carraro et al. [58] further proved that ‘Ca. P. prunorum’ primarily infects cherry plum (P. cerasifera Ehrh.), apricot (Prunus armeniaca L.), peach (P. persica L.), European plum (P. domestica L.), blackthorn (P. spinosa L.) and Japanese plum (P. salicina Lindley). Mahaleb (P. mahaleb L.) and sweet cherry (P. avium L.) were recognized as uncommon natural hosts. The causal agent was also identified in wild growing ash (Fraxinus excelsior L.), and hackberry (Celtis australis L.), dog rose (Rosa canina L.) in France [59], as well as in grapevine in Hungary [60] and Serbia [61]. However, earlier reports of European stone fruit yellows phytoplasma presence in Ribes rubrum in the Czech Republic were not substantiated [62].
2.1. Almond
The almond tree, scientifically known as Prunus dulcis, bears an edible seed. It is native to southwestern Asia and is a significant crop tree that is primarily cultivated in Mediterranean climates between 28° and 48° N and between 20° and 40° S. California is the leading producer, accounting for nearly 80 percent of the world's almond supply. The most commonly observed phytoplasma diseases in almond trees include almond witches' broom, Peach yellows, stone fruit yellows and X-disease [63]. The occurrence of Almond witches' broom (AlmWB) disease is linked to the presence of a phytoplasma known as 'Candidatus Phytoplasma phoenicium', specifically the 16SrIX-B strain [64]. The first outbreak of this lethal and devastating disease in almond trees (Prunus dulcis) was observed in the southern region of Lebanon during the early 1990s. Subsequently, it was also reported in northern Lebanon and Iran, starting in 1995 [65,66,67].
During the past two decades, the emergence of AlmWB has resulted in a rapid decline of almond trees in northern regions and the Bekaa Valley in Lebanon [66,68], as well as in Fars province and other southern provinces in Iran [69]. In past ten years, the disease rapidly spread throughout Lebanonfrom coastal areas to high mountainous regions, lead to the death of around 100,000 trees.In southern Lebanon, in year 2009 'Ca. P. phoenicium' was reported to be in association with serious disease affecting peach (P. persica) and nectarine (P. persica var. nucipersica) trees [70]. In 16 out of 24 Lebanese districts, over 40,000 newly diseased almond trees were observed in year 2010 [67].
In Iran, the phytoplasma 'Ca. P. phoenicium' has been detected in other plant species such as GF-677 (P. amygdalus × P. persica) and wild almond (P. scoparia) [71]. The manifestation of symptoms typically leads to a complete loss of production within 1-2 years of initial appearance [66].It has been noted that AlmWB infects solitary wild trees, abandoned orchards, and well-managed orchards.Characteristic symptoms in almond trees include (i) a proliferation of shoots on the main trunk leading to witches' broom; (ii) the perpendicular development of many axillary buds with small, yellowish leaves; and (iii) a general tree decline that eventually results in dieback. Using housekeeping gene sequence analyses, multiple gene typing analyses of "Ca. P. phoenicium" strains infecting almond, peach, and nectarine trees in Lebanon have shown a significant genetic homogeneity within the analyzed phytoplasma populations. Additionally, the IMP (integral membrane protein) gene sequence assay has enabled the detection of distinct phytoplasma strains associated with AlmWB from a variety of host plants.
This evidence in correlation to other reports of multiple insects’ vectors indicate that AlmWB might be associated to phytoplasma strains that are adapted from an original strain to various hosts [72].Strains of 'Ca. P. phoenicium' are considered as genetic variants of 16SrIX-B subgroup because of shared biological characteristics and are classified among 16SrIX-F and 16SrIX-G subgroups [73].
The primary measures employed for containing AlmWB involve the use of healthy plant material and vector control. Other almond phytoplasma diseases, known as AlmWB-like diseases, have been reported worldwide, causing broomings in Iran [64] in association with phytoplasmas classified in subgroup 16SrIX-C [69]. One of the most prevalent phytoplasma infecting Prunus dulcis, resulting in aster yellows disease, is 'Candidatus Phytoplasma asteris'. In recent times, almond plantations in the Nauni region of India have been consistently impacted by phytoplasma. This has been confirmed through the observation of visible symptoms, fluorescent microscopic studies, and molecular characterization conducted byGupta et al. [63].
2.2. Apricot
Apricot (Prunus armeniaca) trees produce small, orange fruits that are enjoyed fresh, dried, or used in jams and desserts. The phytoplasma associated with the disease known as apricot chlorotic leaf roll assigned to the 16SrX-B subgroup, which is also referred to as stone fruit yellows (ESFY) or 'Ca. P. prunorum' [22]. This pathogen is considered to be the most destructive to apricot trees [54,59,74,75,76,77,78]. ESFY has been found to cause high mortality rates in apricot trees, with a study performed in France showing an average annual mortality rate of 5% in infected plants [79]. The prevalence of infected plants in each orchard is influenced by factors such as the variety of apricot, rootstock, age of the orchard, environmental conditions, infecting strain, and populations of insect vectors. In certain orchards in Germany, the prevalence of infected plants has reached up to 80% [59]. Prevalence values range from 5% to 40% in different studies [74,80], while in Italy and Switzerland, the prevalence rates are 93% and 4% respectively [81,82]. During the spring, infected plants initially exhibit leaf production instead of flowers. From the end of summer until the middle of autumn, the main symptoms include yellowing and upward rolling of the leaves, followed by leaf reddening, branch and phloem necrosis, and overall decline. The decline of apricot trees in European countries, such as Hungary, is resulting in increasing economic losses caused by 'Candidatus Phytoplasma prunorum'. This study utilized nested-PCR to identify the pathogen in plant tissues and insects, which were identified through morphology and molecular methods. The incidence of the disease was recorded to be 29.6% in randomly selected apricot trees, with most infected trees exhibiting symptoms dying within a year.These discoveries show that phytoplasma are characteristically present and cause damage in examined plantations. Plum psyllid, Cacopsylla pruni, is regarded as the sole insect vector of this phytoplasma, regular encounters were common in the surveyed apricot orchards and the surrounding. According to a two-year study, several adult vectors among the sampled specimens carried the pathogen confing the association of these insect vectors in phytoplasma transmisssion. In addition to this, several adult species of Cacopsylla crataegi were also found associated with the pathogen in samples.In the two psyllid species' adult male and female, the frequencies of the phytoplasma seem to be similar. Later, under examination genetic differences among the C. crataegi individuals were observed and from the specimens involved in a previous investigation [83].
2.3. Peach and Nectarine
The ESFY phytoplasma in the Mediterranean region is associated with yellows and decline diseases [75,84,85,86,87,88,89,90,91]. Peach species, along with apricot and plum, are highly susceptible to ESFY [92]. Infected plants exhibit reduced productivity and vigor, and may succumb to the disease within a few years. In Spanish peach orchards, the prevalence of infected plants ranged from 5% to 25%. Peach (Prunus persica) is highly susceptible to phytoplasma infection. Numerous peach diseases have been reported worldwide in association with genetically and biologically distinct phytoplasmas belonging to at least eleven16Sr taxonomic groups (16SrI, II, III, V, VI, VII, IX, X, XII, XV, and XVII) [93].In case of North America, peach production is adversely affected by destructive phytoplasma diseases such as, peach rosette, western X-disease and peach yellows. These diseases are linked with 'Candidatus Phytoplasma pruni' subgroup 16SrIII-B [28,93,94]. In contrast, peach yellow leaf roll is related with a 'Ca. Phytoplasma pyri' subtype belonging to subgroup 16SrX-C. In Europe, 'Ca. P. prunorum', subgroup 16SrX-B associated with European stone fruit yellows (ESFY) of Prunus species, like peach, is an essential phytoplasma disease. In Lebanon, 'Ca. P. phoenicium', subgroup 16SrX-B (an etiological agent of almond witches'-broom disease) is related to the most devastating disease of peach [28,59]. In Iran, grafting transmission trials under controlled environment conditions proved that 'Ca. P. phoenicium' from almond-infected scions can be successfully transmitted and induce symptoms in peach trees [95].
2.4. Plum
Plum trees produce sweet and juicy fruits that are consumed fresh, dried into prunes, used in jams, and baked in desserts.Among the phytoplasmas affecting plum in Europe, 'Ca. P. prunorum' is the most significant in terms of economic impact. It causes significant damage to Japanese plum [96] and is a major constraint in the production of this crop in Mediterranean countries such as Spain, France, Italy, and the Balkans, where Japanese plum and other susceptible Prunus species are widely cultivated. The high percentage of naturally infected trees, often leading to a high mortality rate, renders orchards unproductive within 8-10 years of planting. In some cases, infection rates of up to 100% have been observed in susceptible Japanese plum cultivars Ozark Premier and Shiro after 7 years of growth [38]. However, certain Japanese plum varieties, such as Bragialla, Brarossa, Fortune, and Ruby Crunch, have shown resistance to 'Ca. P. prunorum' infection under natural pressure from ESFY disease, as observed over a 5-year period [97]. The impact of ESFY disease on European plum varies, with severe symptoms resembling those seen in Japanese plum observed in Italy on the "Susina di Dro" variety [98]. A 5-year monitoring period has provided valuable insights into the effects of ESFY disease on European plum. During a 5-year monitoring period conducted in orchards situated in a natural and severely ESFY-affected region, it was observed that only 6 out of 39 cultivars/ selections were found to be infected, with 4 of these exhibiting only mild symptoms (Landi et al. 2010). Phloem necrosis may also occur, particularly following winter frost. A decrease in fruit production is evident, with smaller fruits that ripen later than those from healthy trees and are prone to premature falling. Additionally, an earlier defoliation typically takes place in the summer or early autumn. Initially, the disease may affect a single branch, but within 2-3 years, the entire plant can become affected and perish.Abnormal appearance of suckers from the rootstock is typically observed, which can continue to grow even after the aboveground section of the tree has died. Among the Prunus species, psyllid Cacopsylla pruni (Scopoli) is the only known vector of 'Ca. P. prunorum' [38], moreover the two distinct genetic groups of C. pruni [99] represent divergent biological species [100]. During winters 'Ca. P. prunorum' persist in the stem, therefore it can be transmitted efficiently throughout the year by grafting (Seemüller et al. 1998). As per the monitoring of ESFY in an Italian plum-growingregions, myrobolan rootstocks infected with 'Ca. P. prunorum' resulted in 100% infection of newly grafted scions during single winter season [101].
The presence of 'Ca. P. solani' in plum in Jordan was confirmed through virtual RFLP. These findings indicate that 'Ca. P. solani' may pose a potential threat to other susceptible stone fruit hosts, including peach, almond, and cherry, which have already been reported to be susceptible to this pathogen. The presence of bindweed in Jordan [102], a known natural reservoir for phytoplasma [89], along with potential insect vectors such as Hyalesthes obsoletus [72] and the use of vegetatively propagated plant material [103], may have contributed to the infection of plum in Jordan [102].
2.5. Sweet and Sour Cherry
Cherry trees produce delicious fruits that are eaten fresh, used in pies, jams, and various desserts. Some cherry varieties are also utilized in the formation of wine and liqueurs. The disease known as "Molière decline", which is associated with ESFY, was initially observed in sweet cherry in France [104]. However, a definitive link between the presence of 'Ca. P. prunorum' and cherry "Molière decline" was not established. The symptoms of this disease include yellowing of leaves, decline of shoots, and ultimately, death of the plant. The destructive nature of 'Ca. P. prunorum' was first noted by Kison and Seemüller [80], who observed that flowering cherry used as scion on rootstocks showed symptoms of leaf yellowing and reduced vigor, ultimately resulting in the death of the entire plant or scion. 'Ca. P. prunorum' has since been detected in sweet and sour cherry in Poland and Hungary, and only in sour cherry in the Czech Republic. In addition, an ESFY-infected sour cherry plant in the same country exhibited stunting, leaf rolling, and yellowing.
The United States has also experienced widespread epidemics of X-disease in cherry trees, which have been associated to the presence of 'Ca. P. pruni', a member of the ribosomal subgroup 16SrIII-A [105]. Initially, the disease was reported in sweet cherry plants in California and later in peach trees in Connecticut. X-disease has only been detected in peach trees in Canada, in addition to the United States [106]. The latter species serves as an important reservoir for the phytoplasma, along with certain spontaneous herbaceous species such as Erigeron canadensis, Solidago rugosa, Medicago hispida, and Asclepias syriaca [106].The severity of X-disease is contingent upon climatic conditions, with its impact being more pronounced in California than in Washington State. Conversely, sweet cherry scions that are grafted on P. mahaleb exhibit a rapid decline due to the hypersensitivity reaction of this rootstock, which is purportedly resistant to the pathogen. The phytoplasma can be transmitted by several species of leafhopper, including Colladonas clitellarius, C. montanus, C. geminatus, Euscelidius variegatus, Fieberiella florii, Graphocephala confluens, Gyponana lamina, Keonella confluens, Norvellina seminuda, Osbornellus borealis, Paraphlepsius irroratus, Scaphytopius delonghi, and S. acutus[107]. 'Ca. P. pruni' was detected in symptomatic trees in British Columbia. A total of 118 samples of leaves and fruit stems were collected from individual symptomatic trees before harvest, originating from nine cherry orchards and one nectarine orchard in the Okanagan and Similkameen Valleys in BC. The observed symptoms included the small and misshapen fruit with inadequate color development. 'Ca. P. Pruni' presence was confirmed in sweet cherry samples from British Columbia. Previous reports have indicated the occurrence of 'Ca. P. Pruni'-related strains in commercial poinsettias (Euphorbia pulcherrima) in Canada [108].
B. Phytoplasmas Associated with Forest Trees
Forest trees are essential for both environmental and economical necessities. They are important for global biodiversity, pollution reduction, and subsequent reduction in effects of climate change, in addition to serve as an alternative source of bioenergy. Generally, forest products including lumber show significant impact on revenue generation of many governments worldwide. An increase in demands for timber along with related products is predicted to be continued for near future.
However, diseases, adverse weather and insect pests are certain potent threats to sustainable supply of forest products. Few important severe epidemics of tree disease reported in last century include, Dutch elm disease (ED), chestnut blight, heterobasidion root and stem rot, sudden oak death (SOD), eucalyptus myrtle rust, and the most recent ash dieback [109].
The genetic and bioecological diversity of 'Ca. Phytoplasma' taxa is particularly evident in phytoplasmas that are associated with trees. In these obligate bacteria, their diversity plays a crucial role in the forest ecosystem [110]. The occurrence of phytoplasmas in urban areas is primarily linked to human activities such as plant propagation, horticulture, transportation, and trade, thereby serving as a constant source for the dissemination of these pathogens. Regrettably, there is a lack of comprehensive research on various aspects of phytoplasma presence in both forest and urban ecosystems. The diversity of phytoplasmas associated with different tree species exhibits significant epidemiological characteristics, ranging from severe impacts on infected host plants at a large regional level [111] to subtle symptoms of discoloration or even the absence of any visible signs [112]. Phytoplasma associated with Forest trees worldwide are described below:
3.1. Sandal
Sandalwood (Santalum spp)trees are famous for their aromatic heartwood, which is used to extract sandalwood oil. This oil is used in perfumes, cosmetics, and incense. Sandalwood is also used in traditional medicine and religious rituals.Sandal, is affected by a devastating disease called sandal spike (SAS), which is prevalent in southern India, particularly in the states of Karnataka, Tamil Nadu, and Kerala, but is not found elsewhere [113,114,115]. Over the years, SAS disease has spread progressively, causing significant damage to huge forest areas and posing a threat to the sandal industry in south India, where the production of sandalwood oil is of great importance. The most distinctive symptoms of SAS include shortened internodes, small and extremely thin leaves, and phyllody. The affected leaves, which are pale green or yellow, stand rigidly upright from the shoots, resembling spikes. The severely infected trees generally die within 2 or 3 years from the onset of symptoms. In southern Karnataka, a disease incidence of up to 55% has been recorded [116]. SAS is associated with the aster yellows agent 'Ca. P. asteris,' subgroup 16SrI-B [114,117,118]. The SAS phytoplasma has been transmitted from diseased sandal trees to Catharanthus and then back to sandal trees through dodder (C. subinclusa) bridges. Additionally, experimental transmission from sandal to sandal has been achieved through grafting. The leafhopper Coelidia indica, initially identified as Jassus indicus, has been reported as a natural vector of SAS [119]. However, no insect vectors have been confirmed to be responsible for spreading SSD at this time [120].
3.2. Eucalyptus
Eucalyptus (Eucalyptus spp)trees are valued for their timber, essential oils, and medicinal properties. They are used in the production of paper, as a source of eucalyptus oil, and in traditional medicine for their antimicrobial properties.Eucalyptus little-leaf is a disease caused by phytoplasma that affects various species of eucalyptus. It was initially observed in India in 1971 and was initially believed to be caused by a virus [121]. Subsequently, further researchers examined phytoplasma presence in diseased eucalyptus through the use of electron and light microscope observations [123]. This disease has also been documented in other countries such as Italy, China, Sudan, Iran, and Brazil [122,123,124,125,126,127]. The symptoms reported in different geographic regions and eucalyptus species are similar. These symptoms include the presence of small leaves, yellowing, and the growth of broom-like structures from axillary buds. Affected trees or their symptomatic parts do not produce fruits, exhibit stunted growth, and experience die-back. Severely affected trees show a decline in health. In Italy, a phytoplasma of the 16SrV group was identified in ELL-affected trees [126]. Moreover, the titre of phytoplasma in these infected trees was so low, nevertheless the infections could be analyzed by nested PCR assays. In addition to this, 16SrI-B and 16SrI-C group phytoplasmas were identified in affected eucalyptus trees in Italy, whereas 16SrI-B phytoplasma was detected in affected eucalyptus (E. camaldulensis) trees in Iran [122,125]. In Iran, phytoplasmas belonging to other taxonomic groups were also reported. Azimi et al. [126] in Ahvaz, southwestern Iran reported the detection of 'Ca. Phytoplasma aurantifolia' related strain belonging to 16SrII-D subgroup, in yellows-diseased E. camaldulensis trees. Disease incidence of 5% was recorded in affected trees in this area. Baghaee-Ravari et al. [127] conducted a study in Iran to detect a strain related to 'Ca. Phytoplasma solani', specifically subgroup 16SrXII-Ainyellows, which causes disease in Eucalyptus camaldulensis trees. The provinces of Fars and Khozestan were included in the study. The researchers employed nested PCR assays to identify the strain and found it in 14 out of 22 symptomatic trees, representing approximately 64% of the sample. In a separate study conducted in Brazil, a strain related to 'Ca. Phytoplasma aurantifolia', subgroup 16SrII-C, was detected in diseased Eucalyptus urophylla trees. Similarly, nested PCR assays were used, and 14 out of 22 symptomatic eucalyptus trees tested positive, accounting for approximately 64% of the sample [127]. However, the molecular methods used to determine the identity of phytoplasmas infecting eucalyptus trees in India, China, and Sudan have not been established [127].The eucalyptus trees associated with phytoplasma showing excessive shoot proliferation , reduced leaf size, and shortened internodes, has been reported in India, Sudan, China and Italy and the disease is known as little-leaf disease [112]. De Souza et al. [128] first reported phytoplasma associated with eucalyptus trees showing witches’ broom and little leaf symptoms in Brazil.
3.3. Mulberry
Mulberry (Morus spp.) trees are primarily grown for their sweet and juicy berries, which are used in cooking, making jams, and eaten fresh. Mulberry leaves are also used as food for silkworms in the silk industry. it is widely distributed in Africa, Asia and Europe and has diverse beneficial characteristics [129].Mulberry dwarf (MD) is a severe disease that affects various species of Morus (mulberry), including M. bombycis , M. alba and M. multicaulis. It has been documented in Japan and Korea [130]. The presence of phytoplasma in association with this disease was first confirmed in 1967 by Doi et al.[131], who observed numerous wall-less, pleomorphic bodies in the phloem sieve tube elements of mulberry plants affected by yellows disease. These bodies exhibited morphological and ultrastructural similarities to mycoplasmas, which are known to cause diseases in animals and humans. Around the same time, Ishiie et al. [132] successfully alleviated the symptoms of diseased mulberry plants by applying tetracyclines. The most notable symptoms of MD include yellowing of the leaves, phyllody, stunting, proliferation, and witches' brooms. The disease is associated with the presence of the aster yellows agent 'Ca. P. asteris,' subgroup 16SrI-B [130]. The MD phytoplasma is naturally spread by leafhoppers, specifically Hishimonoidessellatus and H. sellatiformis, with the former being a more efficient vector. In experimental settings, H. sellatus has been shown to transmit the MD phytoplasma to five herbaceous species, including periwinkle, white clover, Ladino clover, red clover, and Chinese milk vetch [133]. Certain cultivars of M. alba have been found to possess natural resistance to the MD phytoplasma, which is attributed to the concentration of phytoalexins in the cortex of the mulberry tree. Resistant cultivars have been found to contain four times higher levels of a specific group of compounds isolated through thin-layer chromatography compared to susceptible cultivars [134].
The causal agent of mulberry yellow dwarf (MYD) disease, known as 'Ca, P. asteris', is responsible for significant economic losses in the sericulture industry. Despite limited genomic resources, the molecular pathogenesis of 'Ca. P. asteris' remains poorly understood. In an effort to address this knowledge gap, a phytoplasma strain called MDGZ-01 was isolated from infected mulberry leaves in Guangdong, China. Utilizing Illumina NovaSeq sequencing technology, we conducted a comprehensive analysis of the MDGZ-01 genome. The resulting genome sequence revealed a circular chromosome spanning 622,358 bases, with a GC content of 29.10%. This valuable genomic resource offers an opportunity to investigate the mechanisms underlying MYD disease [135].
3.4. Paulownia
Paulownia (Paulownia tomentosa)trees are known for their fast growth and lightweight wood. They are used in the production of furniture, musical instruments, and as a source of biomass for paper and biofuel production.Several species of Paulownia trees are affected by a plant disease known as paulownia witches' broom (PaWB), which was one of the first phytoplasma diseases to be described. In trees affected by PaWB, growth and vitality are significantly reduced, and the leaves on affected branches appear yellowish, deformed, and smaller in size. Additionally, flower clusters, if produced, exhibit varying degrees of distortion and abnormal coloration, and may also be sterile. The emergence of slender shoots from the main branches gives rise to the characteristic appearance of witches' brooms. Affected branches display necrosis in the phloem and an irregular arrangement of cells in the woody cylinder, particularly around the vessel. Severely affected trees experience premature death. The wood from infected trees is of low quality and often unsuitable for commercial use. In China, the rapid spread of PaWB was facilitated by the extensive expansion of paulownia plantations in the 1970s, which involved the widespread use of PaWB-infected root cuttings for propagation. In the early 1970s, the incidence of the disease in northern China was 10-20%, which increased to over 70% in the 1980s. A separate survey conducted in China reported an incidence of 5-10% at the seedling stage, 10-20% in 1-year-old saplings, 50% at the middle age stage, and 70-100% in trees of several years old. PaWB is associated with a specific member of the aster yellows phytoplasma group, known as subgroup 16SrI-D [117,118,136].Paulownia is the exclusive natural host of PaWB and was experimentally transmitted to periwinkle by dodder in a study conducted by Doi and Asuyama [137]. Another study by Zhao et al. in 2016 found that there were minimal genetic variations in genes other than the 16S rRNA gene. The PaWB agent is known to be transmitted by three species of heteropteran insects from the family Pentatomidae, namely Halyomorpha mista, H. halys, and H. picus. This information was reported by Hiruki [138] and Weintraub and Beanland [139].
Paulownia witches’ broom (PaWB), caused by phytoplasmas, is a highly destructive infectious disease that affects Paulownia trees. While a few MADS-box transcription factors have been identified as being involved in the development of PaWB, there has been limited research on the entire MADS-box gene family in Paulownia. To address this gap, bioinformatics software was utilized to identify, characterize, localize, and analyze the phylogenetic relationships of the MADS-box genes. Additionally, conserved motifs, gene structures, cis-elements, and protein-protein interaction networks were predicted. The expression patterns of PfMADS-box genes in different tissues were analyzed using quantitative real-time polymerase chain reaction (qRT-PCR). By integrating transcriptome data and protein interaction network predictions, potential genes associated with PaWB formation were identified.89 MADS-box genes, classified into 14 subfamilies were found in P. fortunei genome by Deng et al. [140]. The comprehensive analysis revealed that segment duplication events largely contribute to the evolution of the PfMADS-box gene family; identical motif distribution of proteins in the same subfamily; the promoter regions were highly enriched in cis-elements related to stress and phytohormone-responsive development.The tissue expression pattern of PfMADS-box genes revealed significant subfunctional differentiation they undergo.PfMADS3, PfMADS57, and PfMADS87 are the three genes that might be associated with the presence of PaWB.These investigations led to a useful resource for investigating the possible activities and functions of PfMADS-box genes and establish a strong basis for comprehending the functions of PfMADS-box genes involved in interactions between Paulownia and phytoplasmas.
3.5. Poplar
Poplar (Populus spp) trees are often grown for their wood, which is used in making furniture, paper, and various wood products. They are also used in landscaping and for providing shade.The poplar witches’ broom (PopWB) phytoplasma disease was characteristically discovered in Bulgaria on P. nigra cv Italica trees (Lombardy or black poplar) alongwith P. canadensis (Canadian poplar) in 1973 [141]. Eventually, the first record of the disease was from Netherlands on P. canescens (gray poplar), Lombardy poplar and P. alba (white poplar) [142], similarly from France on white and Lombardy poplar, Germany on P. tremula (aspen)Lombardy and white poplar, and then from Hungary on white poplar, and Croatia and Serbia on Lombardy poplar [143,144,145,146,147,148,149].
The disease is primarily identified by witches' broom, but on P. nigra cv Italica trees, only nonspecific symptoms like yellowing and undersized leaves, sparse foliage, stunting, dieback, and decline may be observed. On aspen trees, the disease is often linked to foliar reddening, yellowing, and decline symptoms, while witches' broom may occasionally develop on vigorous shoots.PopWB is related to ‘Candidatus Phytoplasma asteris,’ subgroup 16SrI-B [146,151] as a result of subsequent infection of 16SrI-A and 16SrI-P subgroup phytoplasma strains in PoWB-affected P. nigra cv Italica trees observed in Croatia and Serbia [147].Phytoplasma strains infecting P. nigra cv Italica, P. alba, and P. tremula in Germany, France, and Hungary showed three variable RFLP profiles on digestion with AluI restriction enzyme as a result of RFLP analyses of 16S rDNA and 16S–23S rDNA spacer region sequences [146]. Consequent differences between German and French strains infecting P. nigra cv Italica trees were found through Heteroduplex mobility assays of 16S rDNA and 16S–23S rDNA spacer region sequences [149]. 16SrI and 16SrVII group phytoplasmas were observed in infected P. nigra trees in Colombia [150]. Leafhoppers, Tremulicerus vitreus and Rhytidodus decimusquartus feeding on Populus speciesare reported as vector of PopWB in France [145,149].
3.6. Elm
Ulmus sp.commonly called elm belong to family Ulmaceae, mostly used for making furniture and boats it shows good resistance against water. Elm trees are infected by Elm yellows (EY) a common disease in Europe and North America [151] . Formely elm yellows disease was called elm phloem necrosis due to necrosis of phloem cells in the elm infected trees occurred. EY was first reported in Ohio [152]. Later on it was found that EY disease were caused by ‘Ca. Phytoplasma ulmi’ , assigned to the EY phytoplasma or 16SrV -A subgroup. And the genetic diversity of EY phyoplasma have been identified in other gene sequences like rpIV, rpsC, secY and imp gene [112]. Another phytoplasma group has been found to be associated with EY was reported in Illinois , USA named ILEY phytoplasma associated with 16SrVI-C subgroup , i.e., clover proliferation group [153]. Along with these two phytoplasma groups two more groups were reported on Elm trees are stolbur group (16SrXII) and aster yellows group (16SrI ) from Italy . , The important symptoms of EY are premature casting of the leaves, yellowing, witches broom , early bud break, brown discoloration of the phloem tissue, and tree death. In china brooming and stunting are typical symptoms of EY-affected elm trees. In several other European and Asian elm genotypes. Yellowing or reddening, reduced vigor, witches’ brooms, and die-back but not phloem discoloration are known to occur. The presence of ‘Ca. Phytoplasma ulmi’ are also many times reported in asymptomatic trees belonging to some Asian and European elm genotypes [58]. In North America leafhopper Scaphoideus luteolus is known to transmit ‘Ca. Phytoplasma ulmi’. Other vectors are also responsible for EY phytoplasma transmission as many insects are recorded on Elm trees for example Allygus atomarious and the spittlebug Philaenus spumarius [154]. Resistant Ulmus spp. Of asia and europ , either naturally infected or graft-inoculated, develop witches' brooms symptom and grow slowly and decline, but no phloem necrosis symptoms were observed. There are no practical methods for the prevention or cure of the mentioned condition. Nevertheless, there are available tolerant Ulmus species and cultivars. These tolerant species comprise Ulmus glabra, U. laevis, U. macrocarpa, U. minor, U. pumila, and U. wilsoniana [155].

Table 1.
Summarized the trees associated with phytoplasma group , their botanical and common names and distribution worldwide.
Table 1.
Summarized the trees associated with phytoplasma group , their botanical and common names and distribution worldwide.
| S. No. | Name of Tree species | Common name | Location | Accession No. with Phytoplasma Group/ Subgroup |
References |
|---|---|---|---|---|---|
| 1. |
Malus domestica Borkh. (Rosaceae) |
Apple | Dossenheim, Germany | AJ542541 Candidatus Phytoplasma mali (16SrX) |
Seemüller and Schneider, 2004 |
| 2. |
Pyrus communis L. (Rosaceae) |
Pear | Chile | KX644930 Candidatus Phytoplasma pyri (16SrX-C) |
Facundo et al., 2017 |
| 3. |
Prunus dulcis(Mill.) D.A. Webb (Rosaceae) |
Almond | Lebanon | KM275491 Candidatus Phytoplasma phoenicium (16SrIX-B) |
Quaglino et al., 2015 |
| 4. |
Prunus armeniaca (Rosaceae) |
Apricot |
Czech Republic | Y11933 Candidatus Phytoplasma prunorum (16SrX) |
Seemüller & Schneider, 2004 |
| 5. |
Prunus persica (Rosaceae) |
Peach | Spain | AJ575108 Candidatus Phytoplasma prunorum (16SrX-B) |
Ludvíková et al., 2011 |
| 6. |
Prunus domestica (Rosaceae) |
Plum | Jordan | MH085227 Candidatus Phytoplasma solani (16SrXII) |
Salem et al., 2019 |
| 7. |
Prunus avium, and Prunus cerasus (Rosaceae) |
Sweet cherry, and Sour cherry |
Lithuania |
FJ231729 Candidatus Phytoplasma asteris’-related strains (16SrI) |
Valiunas et al., 2009 |
| 8. |
Santalum spp. (Santalaceae) |
Sandal | India | EF050071 ‘Candidatus Phytoplasma asteris’-related strains (16SrI-B) |
Khan et al., 2008 |
| 9. |
Eucalyptus spp. (Myrtaceae) |
Eucalyptus | Brazil | KM597065 CandidatusPhytoplasma aurantifolia (16SrII-C) |
Neves de Souza et al., 2015 |
| 10. |
Morus spp. (Moraceae) |
Mulberry | Japan and Korea | MT075612 Candidatus Phytoplasma asteris (16SrI) |
Ji et al. 2009 |
| 11. |
Paulownia tomentosa (Paulowniaceae) |
Paulownia | China | DQ851169 Candidatus Phytoplasma asteris (16SrI-D) |
Yue et al., 2008 |
| 12. |
Populus spp. (Salicaceae) |
Poplar | Bulgaria Colombia France |
AF503568 Candidatus Phytoplasma asteris (16SrI-P) |
Šeruga et al., 2003 Perilla-Henao et al. 2012 |
| 13. |
Ulmus sp. (U. americana, U. rubra, U. minor) Rubus fruticosus |
American elm Slippery elm Field Elm Blackberry |
Ohio, New York, West Virginia and Italy |
AF122910, AF122911, AY197649, AY197656 Elm yellows group (16SrV) |
Griffiths et al., 1999; Lee et al., 2004Swingle , 1938 |
Figure 1.
Phylogenetic tree shows relationship of different groups of phytoplasma associated with trees. Acholeplasma laidlawii is used as outgroup species to root up the tree.
Figure 1.
Phylogenetic tree shows relationship of different groups of phytoplasma associated with trees. Acholeplasma laidlawii is used as outgroup species to root up the tree.
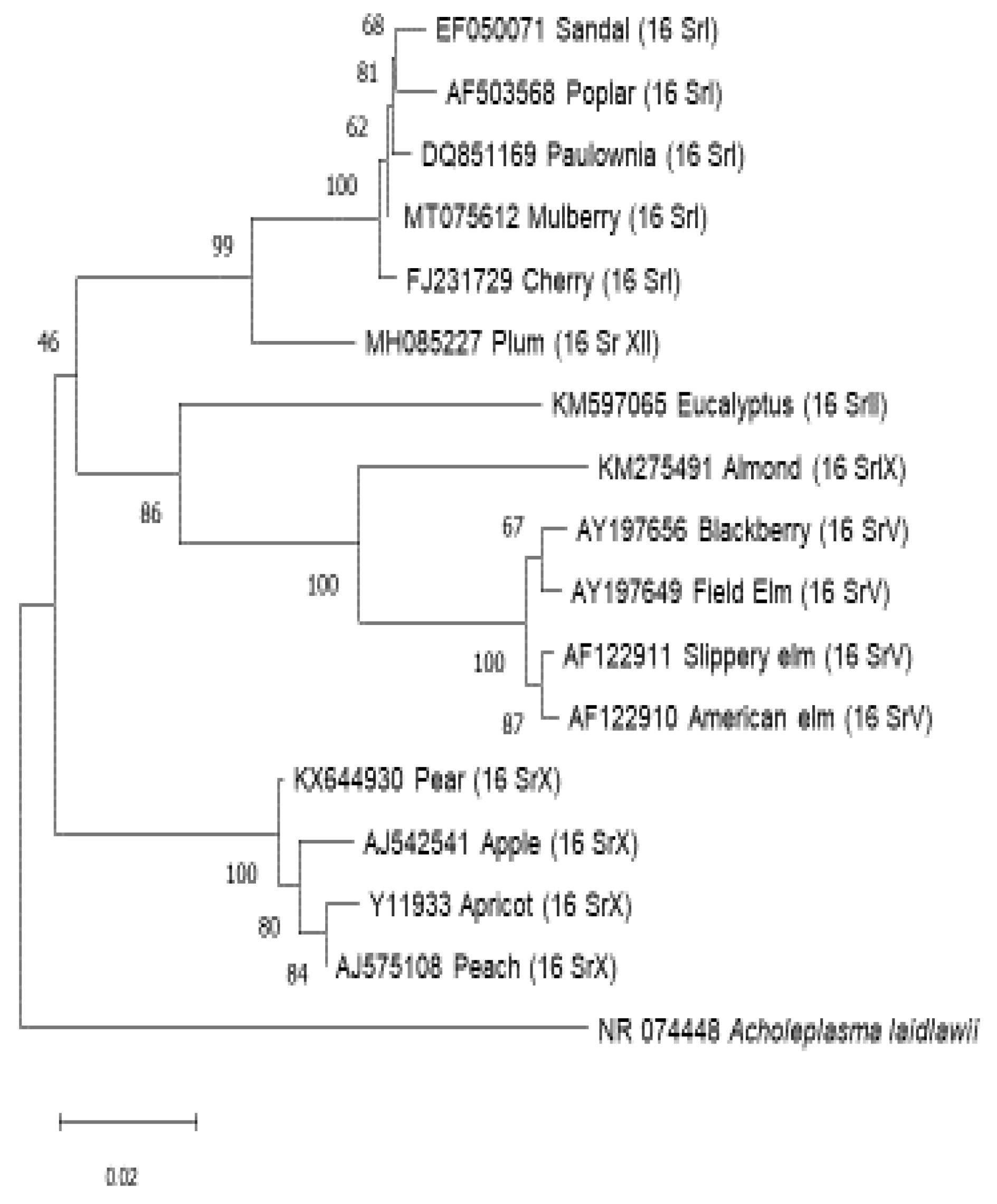
C. Discussion and Conclusion
Phytoplasma detection on trees are made detectable with the development of new technologies in the field of molecular assays. Some of the important diseases of phytoplasma on trees specially Apple proloferations , caused by Ca. P. mali, Pear decline caused by Ca.P.pyri and elm yellows caused by Ca. P. ulmi are very devastating diseases of forest and fruit trees worldwide causes upto 40-60% yield losses. Research on the role of phytoplasmas as the cause of diseases of forest trees has recently been intensified due to the availability of sensitive and specific detection methods, primarily based on PCR technology. These methods have allowed for the detection of phytoplasma infections in several trees for the first time, indicating that phytoplasma diseases may have a similar impact on forest trees as they do on fruit trees. As a result, there has been a significant increase in the number of phytoplasma diseases affecting trees and the associated phytoplasmas [156]. Many of these diseases are associated with taxonomically distinct phytoplasmas, which induce identical symptoms in a given plant and are found in the same areas or different countries. Furthermore, a single plant host may be infected with multiple phytoplasmas, a phenomenon that appears to be common in woody plants that are frequently visited by insect vectors over time.Malus domestica infected with 16SrX group of phytoplasma recorded from Germany, Pyrus communisinfected with 16SrX group phytoplasma recorded from Chile, Prunus dulcisinfected with 16SrIX group phytoplasma recorded from Lebanon, Prunus armeniacainfected with 16SrX group phytoplasma recorded from Czech Republic , Prunus persicainfected with 16SrX group phytoplasma recorded from Spain, Prunus domestica infected with 16SrXII group phytoplasma recorded from Jordan, Prunus avium infected with 16SrI group phytoplasma recorded from Lithuania, Santalum spp.infected with 16SrI group phytoplasma recorded from India, Eucalyptus spp.infected with 16SrII group phytoplasma recorded from Brazil, Morus spp. infected with 16SrI group phytoplasma recorded from Japan and korea, Paulownia tomentosainfected with 16SrI group phytoplasma recorded from China, Populus spp. infected with 16SrI group phytoplasma recorded from Bulgariaand Ulmus sp. infected with 16SrV group phytoplasma recorded from Italy, New York [24,40,72,88,98,111,115,124,130,136,147,150,152].
Based on this review, it has been determined that the Candidatus Phytoplasma asteris, 16SrI and Candidatus Phytoplasma prunorum, 16SrX group are the most widespread among the different phytoplasma groups affecting tree populations. Interestingly, the majority of trees affected by this group belong to the family Rosaceae, which might be because of the presence of potential vectors. Phytoplasma diseases have significant economic and ecological importance globally, particularly in forest trees. Over the years, various phytoplasma taxa have been identified in trees that are singly, doubly, or multiply infected, causing yellows and decline diseases. However, despite extensive research on well-studied phytoplasma diseases like EY and AshY, there is still a lack of information on insect vectors, plant host range, strain virulence, pathogenicity, and host tolerance and resistance for most of these diseases. To effectively manage these diseases, it is crucial to have a comprehensive understanding of these aspects. Further research is needed to investigate the role of phytoplasmas in asymptomatic trees and their relationship to the absence of symptoms. Additionally, the etiological role of various 'non-specific' phytoplasma taxa found in forest trees without pathological data requires further investigation. Moreover, there is still a lack of information on several aspects, including insect vectors, relationships between phytoplasmas and insect vectors, interactions between phytoplasmas and host plants, strain virulence, strain interference, host tolerance, host plant range, and the impact of phytoplasma infections on the growth and yield of affected plants.
Author Contributions
Conceptualization, S.M.; sequence data analysis, S.M. and R.K.; manuscript preparation, S.M. R.K. and C.M.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Beech, E.; Rivers, M.; Oldfield, S.; Smith, P. P. GlobalTreeSearch: The first complete global database of tree species and country distributions. Journal of Sustainable Forestry, 2017 36(5), 454-489. [CrossRef]
- Olson, D. M.; Dinerstein, E.; Wikramanayake, E. D.; Burgess, N. D.; Powell, G. V.,Underwood, E. C. ;Kassem, K. R. Terrestrial Ecoregions of the World: A New Map of Life on Earth: A new global map of terrestrial ecoregions provides an innovative tool for conserving biodiversity. BioScience,2001 , 51(11), 933-938. [CrossRef]
- Crowther, T. W.; Glick, H. B.; Covey, K. R.;Bettigole, C.; Maynard, D. S.; Thomas, S. M.; Bradford, M. A. Mapping tree density at a global scale. Nature, 2015 ,525(7568), 201-205.
- Bar-On, Y. M., Phillips, R., & Milo, R. (2018). The biomass distribution on Earth. Proceedings of the National Academy of Sciences, 115(25), 6506-6511. [CrossRef]
- Fazan, L.; Song, Y. G.; Kozlowski, G. The woody planet: From past triumph to manmade decline. Plants, 2020, 9(11), 1593. [CrossRef]
- Davies, N. G.; Abbott, S.; Barnard, R. C.; Jarvis, C. I.; Kucharski, A. J.; Munday, J. D.; Edmunds, W. J. Estimated transmissibility and impact of SARS-CoV-2 lineage B. 1.1. 7 in England. Science, 2021, 372(6538), eabg3055. [CrossRef]
- Wang, Y.; Wang, X.; Wang, K.; Chevallier, F.; Zhu, D.; Lian, J.; Canadell, J. G. The size of the land carbon sink in China. Nature, 2022, 603(7901), E7-E9. [CrossRef]
- Rao, G. P. Our understanding about phytoplasma research scenario in India. Indian phytopathology, 2021, 74(2), 371-401. [CrossRef]
- Oshima, K. Molecular biological study on the survival strategy of phytoplasma. Journal of General Plant Pathology, 2021, 87(6), 403-407. [CrossRef]
- Wei, W.; Zhao, Y. Phytoplasma taxonomy: nomenclature, classification, and identification. Biology, 2022, 11(8), 1119. [CrossRef]
- Cardwell, G.; Bornman, J. F.; James, A. P.; Black, L. J. A review of mushrooms as a potential source of dietary vitamin D. Nutrients, 2018, 10(10), 1498. [CrossRef]
- Hogenhout, S. A.; Van der Hoorn, R. A.; Terauchi, R.; Kamoun, S. Emerging concepts in effector biology of plant-associated organisms. Molecular plant-microbe interactions,2009, 22(2), 115-122. [CrossRef]
- Lorenz, D. H.; Eichhorn, K. W.; Bleiholder, H.; Klose, R.; Meier, U.; Weber, EGrowth Stages of the Grapevine: Phenological growth stages of the grapevine (Vitis vinifera L. ssp. vinifera)—Codes and descriptions according to the extended BBCH scale. Australian Journal of Grape and Wine Research, 1995, 1(2), 100-103. [CrossRef]
- Lee, I. M.; Gundersen-Rindal, D. E.;Bertaccini, APhytoplasma: ecology and genomic diversity. Phytopathology, 1998, 88(12), 1359-1366. [CrossRef]
- Jarausch, W.; Lansac, M.; Saillard, C.;Broquaire, J. M.;Dosba, F. PCR assay for specific detection of European stone fruit yellows phytoplasmas and its use for epidemiological studies in France. European Journal of Plant Pathology, 1998, 104, 17-27. [CrossRef]
- Martini, M.; Loi, N.; Ermacora, P.; Carraro, L.; Pastore, M. A real-time PCR method for detection and quantification ofCandidatus Phytoplasma prunorum'in its natural hosts. Bulletin of Insectology, 2007, 60(2), 251.
- Yvon, M.; Thébaud, G.; Alary, R.; Labonne, G.Specific detection and quantification of the phytopathogenic agent ‘Candidatus Phytoplasma prunorum’. Molecular and Cellular Probes, 2009, 23(5), 227-234. [CrossRef]
- Pignatta, D.; PoggiPollinI, C.; Giunchedi, L.; Ratti, C.; Reggiani, N.; Forno, F.; Pelato, E. A real-time PCR assay for the detection of European Stone Fruit Yellows Phytoplasma (ESFYP) in plant propagation material. In Acta horticulturae-Proceedings of the 20th International symposium on virus and virus-like diseases of temperate fruit crops-Fruit virus diseases 2008 (Vol. 781, pp. 499-503).
- Thébaud, G.; Yvon, M..; Alary, R.,; Sauvion, N.; Labonne, G. Efficient transmission of ‘Candidatus Phytoplasma prunorum’is delayed by eight months due to a long latency in its host-alternating vector. Phytopathology, 2009, 99(3), 265-273. [CrossRef]
- Nikolic, G.; Zlatkovic, S.; Cakic, M.; Cakic, S.; Lacnjevac, C.; Rajic, Z. Fast fourier transform IR characterization of epoxy GY systems crosslinked with aliphatic and cycloaliphatic EH polyamine adducts. Sensors, 2010, 10(1), 684-696. [CrossRef]
- Torres, M. A.; Dangl, J. L. Functions of the respiratory burst oxidase in biotic interactions, abiotic stress and development. Current opinion in plant biology, 2005, 8(4), 397-403. [CrossRef]
- De Jonghe, K.; De Roo, I.; Maes, M. Fast and sensitive on-site isothermal assay (LAMP) for diagnosis and detection of three fruit tree phytoplasmas. European journal of plant pathology, 2017, 147, 749-759. [CrossRef]
- Tan, C. M.; Lin, Y. C.; Li, J. R.; Chien, Y. Y.; Wang, C. J.; Chou, L.; Yang, J. Y. Accelerating complete phytoplasma genome assembly by immunoprecipitation-based enrichment and MinION-based DNA sequencing for comparative analyses. Frontiers in Microbiology, 2021, 12, 766221. [CrossRef]
- Seemüller, E.; Schneider, B. ‘Candidatus Phytoplasma mali’,‘Candidatus Phytoplasma pyri’and ‘Candidatus Phytoplasma prunorum’, the causal agents of apple proliferation, pear decline and European stone fruit yellows, respectively. International Journal of Systematic and Evolutionary Microbiology, 2004, 54(4), 1217-1226. [CrossRef]
- Lee, I. M.; Gundersen-Rindal, D. E.; Bertaccini, A. Phytoplasma: ecology and genomic diversity. Phytopathology, 1998, 88(12), 1359-1366. [CrossRef]
- Fránová, J.;Špak, J.; Šimková, M. First report of a 16SrIII-B subgroup phytoplasma associated with leaf reddening, virescence and phyllody of purple coneflower. European Journal of Plant Pathology, 2013, 136, 7-12.
- Cieślińska, M.; Hennig, E.; Kruczyńska, D.;Bertaccini, A. Genetic diversity of'Candidatos Phytoplasma mali'strains in Poland. PhytopathologiaMediterranea, 2015, 477-487. [CrossRef]
- Seemüller, E.; Sule, S.; Kube, M.; Jelkmann, W.; Schneider, B. The AAA+ ATPases and HflB/FtsH proteases of ‘Candidatus Phytoplasma mali’: phylogenetic diversity, membrane topology, and relationship to strain virulence. Molecular Plant-Microbe Interactions, 2013, 26(3), 367-376. [CrossRef]
- Janik, K.; Panassiti, B.; Kerschbamer, C.; Burmeister, J.; Trivellone, V. Phylogenetic Triage and Risk Assessment: How to Predict Emerging Phytoplasma Diseases. Biology, 2023, 12(5), 732. [CrossRef]
- Kingston-Smith, A.; Wilkins, P. W.; Humphreys, M. O. Leaves of high yielding perennial ryegrass contain less aggregated Rubisco than S23. Molecular breeding for the genetic improvement of forage crops and turf, 2005, 227, 113. [CrossRef]
- Barthel, D.; Cullinan, C.; Mejia-Aguilar, A.; Chuprikova, E.; McLeod, B. A.; Kerschbamer, C.; Janik, K. Identification of spectral ranges that contribute to phytoplasma detection in apple trees–A step towards an on-site method. Spectrochimica Acta Part A: Molecular and Biomolecular Spectroscopy, 2023, 303, 123246. [CrossRef]
- Kamińska, M.; Śliwa, H. First report of phytoplasma belonging to apple proliferation group in roses in Poland. Plant Disease, 2004, 88(11), 1283-1283. [CrossRef]
- Mehle, N.; Ravnikar, M.; Seljak, G.; Knapic, V.; Dermastia, M. The most widespread phytoplasmas, vectors and measures for disease control in Slovenia. Phytopathogenic mollicutes, 2011, 1(2), 65-76. [CrossRef]
- Musetti, R. Management and ecology of phytoplasma diseases of grapevine and fruit crops. In Integrated management of diseases caused by fungi, phytoplasma and bacteria 2008(pp. 43-60). Dordrecht: Springer Netherlands. [CrossRef]
- Tedeschi, R.; Alma, A. Candidatus Phytoplasma mali': the current situation of insect vectors in northwestern Italy. Bulletin of Insectology, 2007, 60(2), 187.
- Jarausch, B.; Fuchs, A.; Schwind, N.; Krczal, G.; Jarausch, W. Cacopsyllapicta as most important vector for ‘Candidatus Phytoplasma mali’in Germany and neighbouring regions. Bulletin of Insectology, 2007, (2), 189-190.
- Mayer, D. M.; Kuenzi, M.; Greenbaum, R.; Bardes, M.; Salvador, R. B. How low does ethical leadership flow? Test of a trickle-down model. Organizational behavior and human decision processes, 2009, 108(1), 1-13. [CrossRef]
- Carraro, L.; Loi, N.; Ermacora, P. Transmission characteristics of the European stone fruit yellows phytoplasma and its vector Cacopsyllapruni. European Journal of Plant Pathology, 2001, 107, 695-700.
- Blomquist, C. L.; Kirkpatrick, B. C. Frequency and seasonal distribution of pear psylla infected with the pear decline phytoplasma in California pear orchards. Phytopathology, 2002, 92(11), 1218-1226. [CrossRef]
- Facundo, R.; Quiroga, N.; Méndez, P.; Zamorano, A.; Fiore, N. First report of ‘Candidatus Phytoplasma pyri’on pear in Chile. Plant Disease, 2017, 101(5), 830-830. [CrossRef]
- Hashemi-Tameh, M.; Bahar, M.;Zirak, L. ‘Candidatus Phytoplasma asteris’ and ‘Candidatus Phytoplasma aurantifolia’, new phytoplasma species infecting apple trees in Iran. Journal of Phytopathology, 2014, 162(7-8), 472-480. [CrossRef]
- Seemüller, E.; Marcone, C.; Lauer, U.; Ragozzino, A.; Göschl, M. Current status of molecular classification of the phytoplasmas. Journal of plant pathology,1998, 3-26.
- Etropolska, A;Jarausch, W;Jarausch, B;Trenchev, G. Detection of European fruit tree phytoplasmas and their insect vectors in important fruit-growing regions in Bulgaria. Bulgarian Journal of Agricultural Science. 2015, 21(6):1248-53.
- Seemüller,E; Kampmann, M; Kiss , E; Schneider, B. HflB gene-based phytopathogenic classification of ‘Candidatus Phytoplasma mali’strains and evidence that strain composition determines virulence in multiply infected apple trees. Molecular plant-microbe interactions. 2011 , 24(10):1258-66. [CrossRef]
- Koch, K. Sucrose metabolism: regulatory mechanisms and pivotal roles in sugar sensing and plant development. Current opinion in plant biology. 2004 , 1;7(3):235-46. [CrossRef]
- Kaviani , Rad, A.; Balasundram, S.K; Azizi , S.; Afsharyzad, Y.; Zarei, M.; Etesami, H.; Shamshiri, R. R. An overview of antibiotic resistance and abiotic stresses affecting antimicrobial resistance in agricultural soils. International Journal of Environmental Research and Public Health. 2022 , 12;19(8):4666. [CrossRef]
- Zirak, L.; Bahar, M.; Ahoonmanesh, A. Characterization of phytoplasmas related to ‘Candidatus Phytoplasma asteris’ and peanut WB group associated with sweet cherry diseases in Iran. Journal of Phytopathology. 2010 , 158(1):63-5. [CrossRef]
- Chin, C.H.; Chen , S.H.; Wu , H. H.; Ho, C.W.; Ko , M.T.; Lin, C. Y. CytoHubba: identifying hub objects and sub-networks from complex interactome. BMC systems biology. 2014, 8(4):1-7. [CrossRef]
- Vyas ,S. R.; Chakravorty, S.C. Wines from Indian grapes. Haryana Agricultural University; 1971.
- Joshi ,V.K.; Sandhu, D.K.; Jaiswal, S. Effect of addition of SO sub (2) on solid-state fermentation of apple pomace. Current science. Bangalore. 1995;69(3):263-4.
- Giunchedi, L.; Marani, F.; Credi ,R. Mycoplasma-like bodies associated with Plum decline (leptonecrosis)/Corpiriferibili a micoplasmiassociait al deperimento del Susino (leptonecrosi). Phytopathologiamediterranea. 1978, 1:205-9.
- Morvan, G. Apricot chlorotic leaf roll. EPPO Bulletin. 1977;7(1):37-55. [CrossRef]
- Pollini , C.P.; Giunchedi, L; Gambin, E. Presence of mycoplasma-like organisms in peach trees in Northern-Central Italy. PhytopathologiaMediterranea. 1993 ,1:188-92. [CrossRef]
- Lederer, W.; Seemüller, E. Demonstration of mycoplasmas in Prunus species in Germany. Journal of Phytopathology. 1992 ,134(2):89-96. [CrossRef]
- Lorenz ,M,G.; Wackernagel, W. Bacterial gene transfer by natural genetic transformation in the environment. Microbiological reviews. 1994, ;58(3):563-602. [CrossRef]
- Navrátil , M.; Šturdík, E.; Gemeiner, P. Batch and continuous mead production with pectate immobilised, ethanol-tolerant yeast. Biotechnology Letters. 2001, 23:977-82. [CrossRef]
- Álvarez-Fernández ,A.; García-Laviña, P.; Fidalgo ,C.; Abadía, J,.; Abadía, A. Foliar fertilization to control iron chlorosis in pear (Pyrus communis L.) trees. Plant and Soil. 2004 , 263:5-15.
- Carraro ,L.; Ferrini, F.; Labonne, G.; Ermacora, P.; Loi , N. Seasonal infectivity of Cacopsyllapruni, vector of European stone fruit yellows phytoplasma. Annals of Applied Biology. 2004, 144(2):191-5. [CrossRef]
- Jarausch,W.; Jarausch-Wehrheim, B.; Danet, J.L.; Broquaire, J.M.; Dosba, F.; Saillard,C; Garnier , M. Detection and indentification of European stone fruit yellows and other phytoplasmas in wild plants in the surroundings of apricot chlorotic leaf roll-affected orchards in southern France. European Journal of Plant Pathology. 2001 ,107:209-17. [CrossRef]
- Vargas-Asencio ,J.; Wojciechowska, K.; Baskerville , M.; Gomez, A. L.; Perry , K. L.; Thompson, J.R. The complete nucleotide sequence and genomic characterization of grapevine asteroid mosaic associated virus. Virus Research. 2017 , 2;227:82-7. [CrossRef]
- Duduk ,B.; Botti ,S.; Ivanović , M.; Krstić, B.; Dukić ,N.; Bertaccini,A. Identification of phytoplasmas associated with grapevine yellows in Serbia. Journal of Phytopathology. 2004 , 152(10):575-9. [CrossRef]
- Fialova, R.; Navratil, M.; Lauterer, P.; Navrkalova, V. Candidatus Phytoplasma prunorum': the phytoplasma infection of Cacopsyllapruni from apricot orchards and from overwintering habitats in Moravia (Czech Republic). Bulletin of Insectology. 2007 , 1;60(2):183.
- Gupta ,S; Handa, A.; Brakta,A.; Negi , G.; Tiwari, R.K.; Lal ,M. K.; Kumar , R. First report of ‘Candidatus Phytoplasma asteris’ associated with yellowing, scorching and decline of almond trees in India. PeerJ.2023 , 28;11:e15926.
- Verdin , E.; Salar , P.; Danet ,J.L.; Choueiri, E.; Jreijiri, F.; El Zammar , S.; Gelie, B.; Bove, J.M.; Garnier, M. ‘Candidatus Phytoplasma phoenicium’sp. nov., a novel phytoplasma associated with an emerging lethal disease of almond trees in Lebanon and Iran. International Journal of Systematic and Evolutionary Microbiology. 2003 , 53(3):833-8. [CrossRef]
- Salehi ,M.; Izadpanah, K.; Almond Brooming. InProceedings of the 12th Iranian Plant Protection Congress 2 7 September 1995 Karadj 1995.
- Abou-Jawdah ,Y.; Dakhil, H.; El-Mehtar, S.; Lee, I.M. Almond witches'-broom phytoplasma: a potential threat to almond, peach, and nectarine. Canadian Journal of Plant Pathology. 2003 ,1;25(1):28-32. [CrossRef]
- Lova, M.M.; Quaglino, F.; Abou-Jawdah, Y.; Choueiri ,E.; Sobh, H.; Casati ,P.; Tedeschi, R.; Alma , A.; Bianco, P.A. Identification of new 16SrIX subgroups,-F and-G, among'Candidatus Phytoplasma phoenicium'strains infecting almond, peach and nectarine in Lebanon. Phytopathologiamediterranea. 2011, 1;50(2):273-82.
- Choueiri , E.; Verdin ,E.; Danet ,J. L.; Jreijiri,F.; El Zammar, S.; Salard , P.; Bové, J.M.; Garnier ,M. A Phytoplasma Disease Of Almond In Lebanon. Options Méditerranéennes: Série B. Etudes et Recherches. 2003(45):123-5.
- Salehi, M.; Izadpanah, K.; Heydarnejad, J. Characterization of a new almond witches’ broom phytoplasma in Iran. Journal of Phytopathology. 2006 ;154(7-8):386-91. [CrossRef]
- Abou-Jawdah, Y.; Sobh, H.; Akkary,M. First report of Almond witches’ broom phytoplasma 1 (‘Candidatus Phytoplasma phoenicium’) causing a severe disease on nectarine and peach trees in Lebanon. EPPO bulletin. 2009 ,39(1):94-8. [CrossRef]
- Salehi, M.; Salehi, E.; Abbasian, M.; Izadpanah, K. Wild almond (Prunus scoparia), a potential source of almond witches’ broom phytoplasma in Iran. Journal of Plant Pathology. 2015 1;97(2):377-81.
- Quaglino, F; Kube, M.; Jawhari.; M.; Abou-Jawdah, Y.; Siewert , C.; Choueiri, E.; Sobh, H.; Casati, P.; Tedeschi.; R.; Lova ,M. M.; Alma, A. ‘Candidatus Phytoplasma phoenicium’associated with almond witches’-broom disease: from draft genome to genetic diversity among strain populations. BMC microbiology. 2015 15:1-5. [CrossRef]
- Casati ,P.; Quaglino, F.; Abou-Jawdah ,Y.; Picciau, L.; Cominetti, A.; Tedeschi , R.; Jawhari, M.; Choueiri , E.; Sobh , H.; Lova , M. M.; Beyrouthy, M. Wild plants could play a role in the spread of diseases associated with phytoplasmas of pigeon pea witches'-broom group (16SrIX). Journal of Plant Pathology. 2016 , 1:71-81.
- Torres, E.; Martin, M. P.; Paltrinieri , S.; Vila, A.; Masalles, R.; Bertaccini, A. Spreading of ESFY phytoplasmas in stone fruit in Catalonia (Spain). Journal of Phytopathology. 2004, 152(7):432-7. [CrossRef]
- Sertkaya ,G.; Martini , M.; Ermacora , P.; Musetti, R.; Osler , R. Detection and characterization of phytoplasmas in diseased stone fruits and pear by PCR-RFLP analysis in Turkey. Phytoparasitica. 2005 , 33:380-90. [CrossRef]
- Mehle, N.; Ravnikar, M.; Seljak, G.; Knapic, V.; Dermastia, M. The most widespread phytoplasmas, vectors and measures for disease control in Slovenia. Phytopathogenic mollicutes. 2011;1(2):65-76. [CrossRef]
- Tarca ,A.L.; Bhatti ,G.; Romero, R. A comparison of gene set analysis methods in terms of sensitivity, prioritization and specificity. PloS one. 2013 ,15;8(11):e79217. [CrossRef]
- Žežlina, I.; Rot, M.; Kač, M.; Trdan, S. Causal agents of stone fruit diseases in Slovenia and the potential for diminishing their economic impact–a review. Plant Protection Science. 2016 , 28;52(3):149-57. [CrossRef]
- Gentit, P.; Delbos, R. P.; Candresse, T.; Dunez, J. Characterization of a new nepovirus infecting apricot in Southeastern France: apricot latent ringspot virus. European journal of plant pathology.2001 , 107:485-94. [CrossRef]
- Kison , H.; Seemüller , E. Differences in strain virulence of the European stone fruit yellows phytoplasma and susceptibility of stone fruit trees on various rootstocks to this pathogen. Journal of Phytopathology. 2001 , 4;149(9):533-41. [CrossRef]
- Pérez-Pastor, A.; Domingo , R.; Torrecillas , A.; Ruiz-Sánchez , M. C. Response of apricot trees to deficit irrigation strategies. Irrigation Science. 2009, 27:231-42. [CrossRef]
- Genini, M.; Ramel , M. E. Distribution of European stone fruit yellows phytoplasma in apricot trees in Western Switzerland. InXIX International Symposium on Virus and Virus-like Diseases of Temperate Fruit Crops-Fruit Tree Diseases 657 2003 , 21 (pp. 455-458).
- Koncz , L.S.; Petróczy, M.; Pénzes, B.; Ladányi, M.; Palkovics , L.; Gyócsi, P.; Nagy , G.; Ágoston, J.; Fail , J. Detection of ‘CandidatusPhythoplasmaprunorum’in Apricot Trees and its Associated Psyllid Samples. Agronomy. 2023 , 9;13(1):199. [CrossRef]
- Lorenz, K.H.; Dosba, F.; Poggi-Pollini, C.; Llacer, G.; Seemüller, E. Phytoplasma diseases of Prunus species in Europe are caused by genetically similar organisms/Phytoplasma-Krankheit von Prunus-Arten in Europa werdendurchgenetischeinheitlicheOrganismenhervorgerufen. Zeitschrift für Pflanzenkrankheiten und Pflanzenschutz/Journal of Plant Diseases and Protection. 1994 ,1:567-75.
- Delic ,D.; Martini ,M.; Ermacora , P.; Myrta , A.; Carraro, L. Identification of fruit tree phytoplasmas and their vectors in Bosnia and Herzegovina. EPPO bulletin. 2007 , 37(2):444-8. [CrossRef]
- Ambrožič Turk, B.; Stopar , M.; Fajt , N. Blossom thinning of'Redhaven'peach in Slovenia. InXXVIII International Horticultural Congress on Science and Horticulture for People (IHC2010): International Symposium on Plant 932 2010 , 22 (pp. 251-254).
- Cieślińska, M.; Morgaś, H. Detection and identification of ‘Candidatus Phytoplasma prunorum’,‘Candidatus Phytoplasma mali’and ‘Candidatus Phytoplasma pyri’in stone fruit trees in Poland. Journal of Phytopathology. 2011,159(4):217-22. [CrossRef]
- Ludvikova,H.; Franova, J.; Sucha , J. Phytoplasmas in apricot, peach and sour cherry orchards in East Bohemia, Czech Republic. Bulletin of Insectology. 2011, 64(Supplement):S67-8. [CrossRef]
- Mehle ,N.; Ravnikar, M.; Seljak, G.; Knapic, V, Dermastia M. The most widespread phytoplasmas, vectors and measures for disease control in Slovenia. Phytopathogenic mollicutes. 2011;1(2):65-76.
- Bodnár, D.; Csüllög, K.; Tarcali,G. Review of the biology of plant psyllid (Cacopsyllapruni, Scopoli 1763), and its role in the spreading of European stone fruit yellows, ESFY-phytoplasma with Hungarian data. Acta AgrariaDebreceniensis. 2018, 30(74):25-33. [CrossRef]
- Etropolska,A.; Lefort , F. First report of candidatus phytoplasma prunorum, the european stone fruit yellows phytoplasma on peach trees on the territory of canton of Geneva, Switzerland. International Journal of Phytopathology. 2019 , 30;8(2):63-7. [CrossRef]
- Ermacora, P.; Loi, N.; Ferrini , F, Loschi, A, Martini M, Osler R, Carraro L. Hypo-and hyper-virulence in apricot trees infected by European stone fruit yellows. Julius-Kühn-Archiv (427). 2010 , 29:197-200.
- Marcone,C; Guerra, L.J.; Uyemoto, J. K. Phytoplasmal diseases of peach and associated phytoplasma taxa. Journal of Plant Pathology. 2014 , 1;96(1).
- Ragozzino ,A. CHAPTER 46: Peach Rosette Phytoplasma. InVirus and Virus-Like Diseases of Pome and Stone Fruits 2011 (pp. 251-253). The American Phytopathological Society.
- Salehi, M.; Hosseini , S. A.; Salehi , E.; Quaglino, F.; Bianco, P.A. Peach witches’-broom, an emerging disease associated with ‘Candidatus Phytoplasma phoenicium’and ‘Candidatus Phytoplasma aurantifolia’in Iran. Crop protection. 2020 ,1;127:104946. [CrossRef]
- Dosba, F.; Orliac , S.; Dutrannoy, F.; Maison , P.; Massonià , G.; Audergon, J. M. Evaluation of resistance to plum pox virus in apricot trees. InXVInternational Symposium on Fruit Tree Diseases 309 1991 , 8 (pp. 211-220). [CrossRef]
- Landi ,F.; Prandini , A.; Paltrinieri , S.; Missere,D.; Bertaccini, A. Susceptibility to European stone fruit yellows phytoplasma of new and old plum varieties. InCurrent status and perspectives of phytoplasma disease research and management. 2010 (pp. 84-84).
- Pollini, C.P.; Bissani, R.; Giunchedi,L.; Vindimian, E. Occurrence of phytoplasma infection in European plums (Prunus domestica). Journal of Phytopathology. 1995 , 143(11-12):701-3. [CrossRef]
- Sauvion, N.; Lachenaud, O.; Genson , G.; Rasplus J, Labonne, G. Are there several biotypes of Cacopsyllapruni?.Bulletin of Insectology. 2007 , 1;60(2):185.
- Peccoud, J.; Labonne .G.; Sauvion,N. Molecular test to assign individuals within the Cacopsyllapruni complex. PLoS One. 2013 , 19;8(8):e72454. [CrossRef]
- Paltrinieri, S.; Bertaccini, A.; Lugli , A.; Monari , W. Three years of molecular monitoring of phytoplasma spreading in a plum growing area in Italy. InXIX International Symposium on Virus and Virus-like Diseases of Temperate Fruit Crops-Fruit Tree Diseases 657 2003 , 21 (pp. 501-506). [CrossRef]
- Salem ,A. E.; Naweto, M.; Mostafa ,M, Authority AE. Combined effect of gamma irradiation and chitosan coating on physical and chemical properties of plum fruits. Journal of Nuclear Technology in Applied Science. 2016;4:91-102.
- Bertaccini,A.; Duduk, B.; Paltrinieri ,S.; Contaldo, N. Phytoplasmas and phytoplasma diseases: a severe threat to agriculture. American Journal of Plant Sciences. 2014 , 27;2014. [CrossRef]
- Bernhard ,R.; Marenaud, C.; Eymet, J.; Sechet, J.; Fos, A.; Moutous, G. A complex disease of certain Prunus:'Molieres decline'. ComptesRendus des Seances de l'Academied'Agriculture de France. 1977;3(3):178-88.
- Davis , R.E.; Dally , E.L.; Zhao, Y.; Lee, I.M.; Wei, W.; Wolf, T. K.; Beanland , L.; LeDoux , D.G.; Johnson, D.A.; Fiola, J.A.; Walter-Peterson , H. Unraveling the Etiology of North American Grapevine Yellows (NAGY): Novel NAGY Phytoplasma Sequevars Related to ‘Candidatu s Phytoplasma pruni’. Plant Disease. 2015 5;99(8):1087-97. [CrossRef]
- Uyemoto,J.K.; Kirkpatrick, B.C. CHAPTER 44: X-Disease Phytoplasma. InVirus and Virus-Like Diseases of Pome and Stone Fruits 2011 (pp. 243-245). The American Phytopathological Society.
- Kirkpatrick, M.; Lofsvold, D.; Bulmer ,M. Analysis of the inheritance, selection and evolution of growth trajectories. Genetics.1990 , 1;124(4):979-93. [CrossRef]
- Arocha Rosete, Y.; To, H.; Evans, M.; White , K.; Saleh , M.; Trueman , C.; Tomecek , J.; Van Dyk , D.; Summerbell , R.C.; Scott , J.A. Assessing the use of DNA detection platforms combined with passive wind-powered spore traps for early surveillance of potato and tomato late blight in Canada. Plant Disease. 2021 30;105(11):3610-22. [CrossRef]
- Risto ,A.; Abdalla , M.; Myrelid, P. Staging Pouch Surgery in Ulcerative Colitis in the Biological Era. Clinics in Colon and Rectal Surgery. 2022 , 17;35(01):058-65. [CrossRef]
- Fernández, L.; Méndez, J.; Guijarro,J. A. Molecular virulence mechanisms of the fish pathogen Yersinia ruckeri. Veterinary microbiology. 2007 ,15;125(1-2):1-0. [CrossRef]
- Griffiths, H.M.; Sinclair, W.A.; Boudon-Padieu, E.; Daire, X.; Lee, I.M.; Sfalanga, A.; Bertaccini, A. Phytoplasmas associated with elm yellows: molecular variability and differentiation from related organisms. Plant Disease. 1999;83(12):1101-4. [CrossRef]
- Marcone, C.; Valiunas, D.; Mondal, S.; Sundararaj, R. On some significant phytoplasma diseases of forest trees: an update. Forests. 2021;12(4):408. [CrossRef]
- Thomas, S.; Balasundaran, M. Detection of sandal spike phytoplasma by polymerase chain reaction. Current science. 1999;76(12):1574-6.
- Khan, J.A.; Srivastava, P.; Singh, S.K. Identification of a ‘Candidatus Phytoplasma asteris’-related strain associated with spike disease of sandal (Santalum album) in India. Plant Pathology. 2006;55(4):572. [CrossRef]
- Khan, J.A.; Singh, S.K.; Ahmad, J. Characterisation and phylogeny of a phytoplasma inducing sandal spike disease in sandal (Santalum album). Annuals of Applied Biology. 2008;153(3):365-72. [CrossRef]
- Rao, M.S.; Muniyappa, V. Epidemiology of sandal spike-disease. Tree Mycoplasmas and Mycoplasma Diseases; Hiruki, C., Ed.; University of Alberta Press: Edmonton, AB, Canada. 1988:57-8.
- Marcone, C.; Lee, I.M.; Davis, R.E.; Ragozzino, A.; Seemüller, E. Classification of aster yellows-group phytoplasmas based on combined analyses of rRNA and tuf gene sequences. International Journal of Systematic and Evolutionary Microbiology. 2000;50(5):1703-13. [CrossRef]
- Lee, I.M.; Martini, M.; Marcone, C.; Zhu, S.F. Classification of phytoplasma strains in the elm yellows group (16SrV) and proposal of ‘Candidatus Phytoplasma ulmi’for the phytoplasma associated with elm yellows. International Journal of Systematic and Evolutionary Microbiology. 2004;54(2):337-47. [CrossRef]
- Weintraub, P.G.; Beanland, L. Insect vectors of phytoplasmas. Annu. Rev. Entomol.2006;51:91-111. [CrossRef]
- Tiwarekar, B.; Kirdat, K.; Sundararaj, R.; Yadav, A. Updates on phytoplasma diseases associated with sandalwood in Asia. In: Phytoplasma Diseases of Major Crops, Trees, and Weeds. Elsevier; 2023.309–20. [CrossRef]
- Sastry, K.S.; Thakur, R.N.; Gupta, J.H.; Pandotra, V.R. Three virus diseases of Eucalyptus citriodora. Indian Phytopathology. 1971.
- Marcone, C. Current status of phytoplasma diseases of forest and landscape trees and shrubs. Journal of Plant Pathology. 2015;1.9-36.
- Marcone, C.; Ragozzino, A.; Seemuller, E. Detection of an elm yellows-related phytoplasma in eucalyptus trees affected by little-leaf disease in Italy. Plant Disease. 1996, 80, 669–673.
- Neves de Souza, A.; Leão de Carvalho, S.; Nascimento da Silva, F.; Alfenas, A.C.; Zauza, E.A.; Carvalho. C.M. First report of phytoplasma associated with Eucalyptus urophylla showing witches’ broom in Brazil. Phytopathogenic Mollicutes. 2015;5(1s):S83-4. [CrossRef]
- Salehi, M.; Hosseini, S.E.; Salehi, E. First report of a ‘Candidatus Phytoplasma asteris’ related phytoplasma associated with eucalyptus little leaf disease in Iran. Journal of Plant Pathology. 2016;98(1):175.
- Azimi, M.; Farokhi-Nejad, R.; Mehrabi-Koushki, M. First report of a ‘Candidatus Phytoplasma aurantifolia’-related strain associated with leaf roll symptoms on eucalyptus in Iran. New Disease Reports. 2017;35(4):3. [CrossRef]
- Baghaee-Ravari, S.; Jamshidi, E.; Falahati-Rastegar, M. “Candidatus Phytoplasma solani” associated with Eucalyptus witches’ broom in Iran. Forest Pathology. 2018;48(1):e12394. [CrossRef]
- de Souza, B. R.; Freitas, I. A. S.; de Araujo Lopes, V.; do Rosario Rosa, V.; Matos, F. S. Growth of Eucalyptus plants irrigated with saline water. African Journal of Agricultural Research. 2015;10(10), 1091-1096.
- Dhanyalakshmi, K.H.; Nataraja, K.N. Mulberry (Morus spp.) has the features to treat as a potential perennial model system. Plant signaling&behavior. 2018;13(8):e1491267. [CrossRef]
- Ji, X.; Gai, Y.; Zheng, C.; Mu, Z. Comparative proteomic analysis provides new insights into mulberry dwarf responses in mulberry (Morus alba L.). Proteomics. 2009;9(23):5328-39. [CrossRef]
- Doi, Y.O.; Teranaka, M.; Yora, K.; Asuyama, H. Mycoplasma-or PLT group-like microorganisms found in the phloem elements of plants infected with mulberry dwarf, potato witches' broom, aster yellows, or paulownia witches' broom. Japanese Journal of Phytopathology. 1967;33(4):259-66. [CrossRef]
- Ishiie, T.; Doi, Y.; Yora, K.; Asuyama, H. Suppressive effects of antibiotics of tetracycline group on symptom development of mulberry dwarf disease. Japanese Journal of Phytopathology. 1967;33(4):267-75. [CrossRef]
- Kim, Y.H.; La, Y.J.; Kim, Y.T. Transmission and histochemical detection of mulberry dwarf mycoplasma in several herbaceous plants. The Plant Pathology Journal. 1985;1(3):184-9.
- Yuan-zhang, K.; Feng-ping, Z.; Zhi-song, X.; Pei-gen, C. Studies on the mechanism of mulberry varietal resistance to mulberry yellow dwarf disease. International Journal of Tropical Plant Diseases. 1999;17(1-2):103-14.
- Luo, L.; Zhang, X.; Meng, F.; Wang, Y.; Zhou, Y.; Liu, J. Draft genome sequences resources of mulberry dwarf phytoplasma strain MDGZ-01 associated with mulberry yellow dwarf (MYD) diseases. Plant Disease. 2022;106(8):2239-42. [CrossRef]
- Yue, H.N.; Wu, Y.F.; Shi, Y.Z.; Wu, K.K.; Li, Y.R. First report of paulownia witches'-broom phytoplasma in China. Plant disease. 2008;92(7):1134-. [CrossRef]
- Doi, Y.; Asuyama, H. Paulownia witches'broom disease. In Mycoplasma diseases of trees and shrubs. 1981 (pp. 135-145). Academic Press.
- Hiruki, C.Paulownia witches’-broom disease important in East Asia. Acta Horticulture. 1999;469: 63–68. [CrossRef]
- Weintraub, P. G.; Beanland, L. Insect vectors of phytoplasmas. Annu. Rev. Entomol., 2006 , 51, 91-111. [CrossRef]
- Deng, M.; Dong, Y.; Xu, S.; Huang, S.; Zhai, X.; Fan, G. Genome-Wide Identification and Expression of the Paulownia fortunei MADS-Box Gene Family in Response to Phytoplasma Infection. Genes. 2023;14(3):696. [CrossRef]
- Atanasoff, D. Stammhexenbesenbei Ulmen und anderenBäumen. Archives of Phytopathology & Plant Protection. 1973;9(4):241-3. [CrossRef]
- Van Der Meer, F.A. Witches' brooms in poplars?.Populier. 1980;17(2):42-3.
- Cousin, M.T.; Sharma, A.K. Mycoplasmalike Organisms (MLOs) Associated With the Witches' Broom Disease of Poplar. PhytopathologischeZeitschrift. 1986;117(4). [CrossRef]
- Seemüller, E.; Lederer, W. MLO-associated decline of Alnus glutinosa, Populus tremula and Crataegus monogyna. Journal of Phytopathology. 1988;121(1):33-9. [CrossRef]
- Cousin, M.T. Witches' broom: a phytoplasma disease of poplar.1996.
- Berges, R.; Cousin, M.T.; Roux, J.; Mäurer R.; Seemüller, E. Detection of phytoplasma infections in declining Populus nigra ‘Italica’trees and molecular differentiation of the aster yellows phytoplasmas identified in various Populus species. European journal of forest pathology. 1997;27(1):33-43. [CrossRef]
- Šeruga, M.; Škorić, D.; Botti, S.; Paltrinieri, S.; Juretić, N.; Bertaccini, A.F. Molecular characterization of a phytoplasma from the aster yellows (16SrI) group naturally infecting Populus nigra L.‘Italica’trees in Croatia. Forest pathology. 2003;33(2):113-25. [CrossRef]
- Mitrović, J.; Paltrinieri, S.; Contaldo, N.; Bertaccini, A.; Duduk, B. Occurrence of two ‘Candidatus Phytoplasma asteris’-related phytoplasmas in poplar trees in Serbia. Bulletin of Insectology. 2011;64(Supplement):S57-8.
- Cousin, M.T.; Roux, J.; Boudon-Padieu, E.; Berges, R.; Seemuller, E.; Hiruki, C. Use of heteroduplex mobility analysis (HMA) for differentiating phytoplasma isolates causing witches’ broom disease on Populus nigra cv italica and stolbur or big bud symptoms on tomato. Journal of Phytopathology. 1998;146(2-3):97-102. [CrossRef]
- Perilla-Henao, L.M.; Dickinson, M.; Franco-Lara, L. First report of ‘Candidatus Phytoplasma asteris’ affecting woody hosts (Fraxinus uhdei, Populus nigra, Pittosporum undulatum, and Croton spp.) in Colombia. Plant Disease. 2012;96(9):1372-. [CrossRef]
- Marcone, C. Elm yellows: A phytoplasma disease of concern in forest and landscape ecosystems. Forest Pathology. 2017;47(1):e12324. [CrossRef]
- Swingle, R.U. A phloem necrosis of Elm. Phytopathology. 1938;28(10).
- Jacobs, K.A.; Lee, I.M.; Griffiths, H.M.; MillerJr, F.D.; Bottner, K.D. A new member of the clover proliferation phytoplasma group (16SrVI) associated with elm yellows in Illinois. Plant Disease. 2003;87(3):241-6. [CrossRef]
- Matteoni, J.A.; Sinclair, W.A. Elm yellows and ash yellows. Tree mycoplasmas and mycoplasma diseases. 1988:19-31.
- Mittempergher, L. Elm yellows in Europe. InThe elms: Breeding, conservation, and disease management.2000:103-119. Boston, MA: Springer US.
- Marcone, C. Current status of phytoplasma diseases of peach. InVIII International Peach Symposium1084. 2013:569-578. [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



