
Submitted:
19 December 2023
Posted:
20 December 2023
You are already at the latest version
Abstract
Staphylococcus spp. are among the main pathogens responsible for important infections worldwide, both in humans and in domestic or wild animals. Chiropterans, popularly known as bats, inhabit natural environments and human constructions, and have been considered potential disseminators of several pathogens. Studies have shown the expression of the mecA gene in these animals, being a significant threat to One Health. The objective of this study was to determine antibiotic resistance and the presence of the mecA gene in Staphylococcus spp. isolates from bats from forest areas in the municipalities of Paraná, Brazil. A total of 87 frugivorous chiropterans belonging to the family Phyllostomidae were captured. Of these, 87 rectal and 87 oral swabs samples were collected, re-sulting in 32 Staphylococcus spp. isolates. Twenty-four isolates were characterized as coagu-lase-negative, and 8 coagulase-positive Staphylococcus spp. (STACP), which showed notable re-sistance to oxacillin (78.12%), penicillin G (65.62%) and to other antimicrobials as well. One STACP isolate was mecA-positive. Therefore, further studies are needed to elucidate the mechanisms of contamination and dissemination of these pathogens, emphasizing that anthropic actions in na-tural environments, especially those related to the use of antimicrobials, pesticides may be the main reasons for the presence of resistant bacteria in bats.
Keywords:
Antibiotics
; Wild animals
; Resistance genes
; Bats
; One Health
; Chiropterans
1. Introduction
Staphylococcus spp. are among the main pathogens responsible for important infections worldwide in both humans and animals [1, 2], and one of the most concerning mechanisms of virulence is their ability to acquire resistance to antibiotics [3].
MRSA (methicillin-resistant Staphylococcus aureus) was first identified shortly after the introduction of methicillin in 1960 and is considered one of the most serious threats to global health [4]. In addition, in recent years, other methicillin-resistant Staphylococcus spp., such as S. cohnii, S. hominis, S. lentus, S. sciuri, S. xylosus, S. epidermidis, S. lugdunensis and S. haemolyticus, known as MRS, gained importance as they were found to be responsible for a variety of infections in human populations, particularly in immunocompromised patients and in domestic and wild animals [5, 6].
Chiropterans, popularly known as bats, are wild animals belonging to the order Chiroptera. They inhabit natural environments such as forests and human constructions [7]. Because they are animals that fly long distances and because they inhabit the ceilings of homes and other types of buildings, they become important to human, animal and environmental health, since some authors have already reported that they are important reservoirs of different microorganisms, including Staphylococcus spp. and have also been considered potential disseminators of various pathogens [8, 9].
Studies have already demonstrated the presence of coagulase-positive MRS in bats [5, 10], and resistance is caused by the expression of the mecA gene, localized on the staphylococcal cassette chromosome mec (SCCmec). Resistance to methicillin and other beta-lactam antibiotics occurs due to a penicillin-binding protein (PBP2a) encoded by the mecA gene, that has low affinity for beta-lactams [11]. Usually, the mecA gene is found in hospital-associated MRSA (HA-MRSA), however, it may also normally be colonizing the nasal region of people in the community [12, 13].
Therefore, the presence of mecA in natural environments promotes global antimicrobial resistance, which is a significant concern for world health [14]. Accordingly, the aim of this study was to determine antibiotic resistance and the presence of the mecA gene in Staphylococcus spp. isolates of bats from forest areas in the municipalities of Paraná, Brazil.
2. Materials and Methods
2.1. Ethical Aspects, target population and sampling
A total of 87 bats were captured in forested areas in the municipalities of Cianorte, Japurá and São Tomé in the state of Paraná, from October 2017 to June 2019, under license from the Chico Mendes Institute for Biodiversity Conservation (ICMBio No. 60061-1) for the collection of biological material and with the approval of the Research Ethics Committee on Animal Experimentation (CEPEEA) of Universidade Paranaense under Protocol No. 34772/2018.
2.2. Capture of bats
The captures were made monthly, during eight nights, starting at sunset. The capture techniques were adapted from Greenhall and Paradiso [15], using ten nylon nets with a size of 14 meters in length and three meters in height. The nets were set up 0.5 to 3.0 meters above the ground, positioned across the trails in forest areas.
After capture, the animals were managed with leather gloves to avoid bites. Biometric data were collected, as a measurement of the forearm and weighing of the specimens, and identification was performed using a dichotomous key, following the criteria of Vieira [16], Vizotto and Taddei [17] and Jones and Carter [18].
The samples were collected using the Sterile swab GS-03 – Metal Rod. The first swab was introduced into the oral cavity of each chiropteran where circular and rotational movements were performed on the gums and tongue region. A second swab was collected from the rectal region, compressing it with rotational movements. All samples were stored and sent under refrigeration to the Laboratory of Preventive Veterinary Medicine and Public Health of the Postgraduate Program in Animal Science with an Emphasis on Bioactive Products at the University of Paraná (UNIPAR).
All handling with the bats was carried out by a team trained for this purpose and, immediately after data collection, the animals were released into the forest again.
2.3. Isolation and characterization of samples
In the laboratory, each swab was inserted into Brain Heart Infusion (BHI) Broth, and incubated at 37°C for 24 hours, and then, each sample was plated on mannitol medium and again incubated at 37°C for up to 48 hours for isolation of Staphylococcus spp. Each colony was subjected to analysis for macro and microscopic characteristics and biochemical tests for classification into coagulase-positive Staphylococcus (STACP) and coagulase-negative Staphylococcus (STACN) [19].
2.4. Phenotypic antimicrobial sensitivity tests
Antibiotic susceptibility tests were performed according to the Clinical and Laboratory Standards Institute (CLSI) [20]. Each Staphylococcus sp. was submitted to the antibiogram to determine the resistance profile, identifying the sample as to potential risk to public health. The samples were tested with a commercial kit (Laborclin, Pinhais, Paraná - BR), against drugs that have different mechanisms of action such as: cefepime (30 µg), ciprofloxacin (5 µg), chloramphenicol (30 µg), clindamycin (5 µg), erythromycin (15 µg), gentamicin (10 µg), oxacillin (1 µg), penicillin G (10 U), rifampicin (5 µg), sulfazotrim (25 µg), tetracycline (30 µg), and vancomycin (30 µg).
2.5. Bacterial multiresistance index
To calculate the multidrug resistance index, the formula described by Krumperman [21], a/(bc), was used, where a is the total number of antibiotics to which the samples showed resistance, b is the number of antibiotics tested, and c is the number of samples. The index to define whether the samples are high or low risk was 0.200, where ≤0.199 is considered low risk and ≥0.200 considered high risk.
To evaluate the multidrug resistance index of each sample, another formula was used, also described by Krumperman [21], a/b, where a is the number of antibiotics against which the isolate was resistant, and b the number of antibiotics tested.
2.6. Polymerase chain reaction (PCR): Staphylococcus aureus study
DNA of the STACP samples was extracted with the Purelink Genomic DNA Kit (Invitrogen, Carlsbad, CA, USA) according to the manufacturer's instructions, and PCR assays were performed using the primers Sa442-1 (5'-AAT CTT TGT CGG TAC ACGATA TTC TTC ACG-3 ') and Sa442-2 (5'-CGT AAT GAG ATT TCA GTA GAT AAT ACA ACA-3') according to Martineau et al. [22]. DNA amplification was done with Applied Biosystems model Veriti™ 96-Well Thermal Cycler.
The amplification products were visualized by electrophoresis on a 2% agarose gel stained with GelRed (Uniscience, Osasco, São Paulo, BR) using a 100-bp molecular marker (Invitrogen, Carlsbad, CA, USA), and the products were visualized as a single band of 241 bp. The standard strain ATCC 25923 of S. aureus was used as the positive control.
2.7. MecA gene determination
DNA of Staphylococcus spp. classified as resistant and with intermediate resistance to oxacillin was extracted using the Purelink Genomic DNA Kit (Invitrogen, Carlsbad, CA, USA) according to the manufacturer's instructions, and PCR assays were performed using the mecA1 primer (AAAATCGATGGTAAAGGTTGG) and mecA2 primer (AGTTCTGCAGTACCGGATTTG) following the method of Murakami et al. [23]. DNA amplification was performed with the Applied Biosystems model Veriti™ 96-Well Thermal Cycler.
The amplification products were visualized by electrophoresis on a 2% agarose gel stained with GelRed (Uniscience, Osasco, São Paulo, BR), along with a 100-bp molecular marker (Invitrogen, Carlsbad, California, USA), and the products were visualized as a single band of 533 bp. The standard strain ATCC 43300 of S. aureus served as the positive control and the standard strain ATCC 25923 of S. aureus as the negative control.
3. Results
A total of 87 chiropterans were captured, all belonging to the family Phyllostomidae. Of these, 28 (32.18%) were of the species Sturnira lilium, 22 (25.29%) of Carollia perspicillata, 20 (22.99%) of Platyrrhinus lineatus and 17 (19.54%) of Artibeus lituratus. Regarding the size and weight of the animals, there was a variation of 6 to 10 cm, and 12 to 76 g, respectively (Figure 1).
Of the 87 captured chiropterans, 32 Staphylococcus spp. isolates were obtained, 17 (53.12%) rectal and 15 (46.88%) oral. The isolates were characterized as STACN (24.75%) and STACP (8.25%).
Staphylococcus spp. showed marked differences in antibiotic resistance. Twenty-five isolates (78.12%) were resistant to oxacillin; 21 (65.62%) to penicillin G, 12 (37.50%) to erythromycin, besides other antibiotics (Table 1).
The resistance profile and the multi-resistance index of the Staphylococcus spp. isolates are shown in Table 2. The multi-resistance index of the isolates varied between 0.000 and 0.500, and results of 17 samples were greater than the established limit, which according to Krumperman [21], indicates that they are of high risk to public health, reaffirming the existence of multidrug-resistant bacteria present in the oral and rectal microbiota of bats.
In the molecular tests, no S. aureus was found; however, one STACP was positive for the mecA gene (Figure 2).
4. Discussion
In view of the results found in this work, the importance of this animal species for public health is evident, considering that human, animal and environmental health are closely related to each other.
The animals captured during this study were identified as belonging to four different species, namely Artibeus lituratus, Carollia perspicillata, Platyrrhinus lineatus and Sturnira lilium, all belonging to the family Phyllostomidae, order Chiroptera. Phyllostomids represent the richest family in species diversity in the Neotropical region, showing a varied range of eating habits; however, with a predominance of frugivorous species [24].
Those four species were found in all collection sites, showing that they are prevalent in the municipalities of Cianorte, Japurá and São Tomé, which are part of the microregion of Cianorte, Paraná, Brazil, as described by Ortêncio-Filho et al. [25].
The forested areas located in the municipalities where the collections took place, form interconnections through the presence of riparian forests bordering the rivers and streams of the region [26]. This characteristic shows the possibility of the movement of the chiropterans through those areas in a brief period of time, which allows the dissemination of different pathogens.
Although the captured species were exclusively frugivorous and have a predilection for forming their colonies in regions farther from human presence, direct exposure to humans may still exist, since they feed around fruit trees that are used for afforestation in cities or in orchards in the areas [27]. Another important aspect is the great flight capacity of these mammals, allowing them to travel long distances in a single night [28]. This behavior ends up narrowing the relationship of this animal species with humans, allowing their exposure to antibiotics that can cause the selection of microorganisms with resistance to different drugs and exposure to different species of resistant bacteria as well [29, 30].
Among the chiropterans samples, there were 32 Staphylococcus isolates, 17 (53.12%) from the rectal region and 15 (46.88%) from the oral region. Studies indicate that prevalence of Staphylococcus species in animals is common [31], and they can be found in the skin, mucosa, and intestinal microbiota of a wide variety of mammals [32] and birds [33]. Thus, once they are associated with the gastrointestinal tract, they are eliminated with the feces of colonized animals and become potential contaminants in the environment [34].
Coagulase-negative Staphylococcus spp. (75%) were the most frequent found in the samples of this study. However, it was not possible to identify isolated species. To the best of our knowledge, there are no studies in the literature relating coagulase-negative staphylococci to these animals, and therefore, studies on the identification of these bacterial species are crucial when it comes to public health.
In the last few decades, the increase in the number of STACN infections and their frequent resistance to methicillin, which indicates cross-resistance with all types of beta-lactams, has become an additional difficulty in the problem of infection control by this agent [35]. Another important aspect that must be considered is the resistance in other bacterial species, mainly Staphylococcus aureus, because resistance mechanisms and related genes as well show differences in functioning [36].
For a long time, STACN were considered saprophytes and rarely pathogenic, but they are currently recognized as etiologic agents of a series of infectious processes, being commonly isolated in human and animal clinical samples [37, 38]. In animals, STACN are mainly implicated in bovine mastitis [39, 40]. In humans, they are involved in nosocomial infections [38, 41]. An important factor is that the presence of these microorganisms colonizing wild animals, favor their maintenance in the environment, enabling the selection of new genetic forms.
In this work, the results were significant in terms of antimicrobial resistance. An alarming aspect is the increased resistance to erythromycin. According to Bouchiat et al. [42], the presence of isolates resistant to this antibiotic draws attention, since macrolides are used in case of intolerance to beta-lactams or resistance to methicillin and exert beneficial effects in infection treatments in these cases.
Krumperman [21] pointed out that results between 0.200 and 0.250 are arbitrary; however, they require attention. It is noteworthy that above this limit, the indices are considered, in fact, a high risk for public health and can contaminate the environment that the animals inhabit, thus reaching the human population.
Different results were found by Akobi et al. [5], in which no drug resistance was detected except for penicillin (3%). The results demonstrate that both the captured animals and the colonizing bacterial species were not exposed to the antimicrobials detected in this work, which indicates that the environment was possibly not contaminated by them and that possibly each location or country should have its specificity regarding antimicrobial resistance since this resistance is intrinsically related to the most used drugs in each location.
It is believed that the presence of resistance is associated with constant exposure or even remnants of antimicrobials possibly through contaminated food, hospital waste, groundwater, rivers and sewage drainages [30] and the successive selective pressure causing resistance gene transfer between isolates. Thus, this result indicates that the chiropterans in this study were exposed to different drugs [43, 44], increasing the possibility of selection and spread of antibiotic resistance [45].
Although the majority of Staphylococcus spp. shows resistance to beta-lactams, these drugs offer advantages due to their low cost, and therefore, they are recommended for the treatment of sensitive isolates [29, 32].
Therefore, the increased frequency of cross-resistance indicators with all classes of beta-lactams including even other classes of antimicrobials, found in bacteria colonizing wild animals is extremely worrisome [29, 32, 44]. This fact suggests the need for care in the use of these substances; since incorrect use along with the natural selection of microorganisms, probably resulted in the phenomenon of resistance, in both veterinary and human medicine, reaching the environment [43].
Antibiotic resistance in this study was 10.35% among the 32 samples analyzed, where only one of them was positive for the mecA gene in a STACP identified only at the genus level of (Staphylococcus sp.) in a specimen of a bat of the species Platyrrhinus lineatus. Animals of this species are frugivorous, and they usually seek human shelters, where, in fact, they eat the fruits in order to avoid predation [24]. This behavior makes direct contact between these animals and humans, increasing the risk of spreading the agent.
Accordingly, studies carried out with the feces of chiropterans in settlements in an urbanized environment in Africa have shown the proximity between bats and humans. S. aureus was identified, although the presence of the mecA gene has not been detected [4; 10].
Often resistance to oxacillin in Staphylococcus spp. is mainly the result of the mecA gene, along with the low affinity of PBP2a for this drug [11]. The assay of this gene serves as a diagnosis and helps in choosing the best antibiotic therapy to be instituted [46, 47]. Considering the occurrence of strains of Staphylococcus spp. resistant to oxacillin in veterinary medicine, and knowing that it is the drug of choice in the treatment of severe staphylococcal infections in humans, the possibility of zoonotic transmission of Staphylococcus spp. resistant to oxacillin indicates the need to monitor the isolate profiles and susceptibility to antibiotics in veterinary practice and also to monitor the consequences that this interaction can have at the public health level [48].
STACP can be considered reservoirs of these genes in hospital environment and can therefore transfer these factors to other bacteria [48]. This fact implies either the presence of antibiotics in the environment or exposure to other sources of contamination [49]. Therefore, when bats are in their natural habitat, they end up interacting directly or indirectly with other species of animals or even humans [50].
According to Peterson et al. [51], the presence of these bacteria in wild animals is extremely alarming, where they point out the contamination of the environment where bats live and configure a high dispersion potential, where the forms of dissemination are not restricted to physical contact only, but dispersal also through the air. According to Banskar et al. [52], this information is important not only for a better understanding of this interaction between bats and bacteria, but also, in the screening of possible pathogens present in bats, which generally serve as a reservoir for these organisms.
Fessler et al. [53] highlights that epidemiological studies on antimicrobial resistance in wild animals should be popularized, since resistant bacteria are possibly selected in human and veterinary hospitals, where antimicrobials are used in clinical therapy or as growth promoters [54].
From an epidemiological point of view, it is important to determine the organisms that act as a reservoir of resistant bacteria, and therefore, the identification of species and the genes involved with resistance is a prerequisite for monitoring the spread of contamination from animal and human populations and the environment itself.
5. Conclusions
In Staphylococcus spp. from rectal and oral samples of bats captured in forested areas in the municipalities of Cianorte, Japurá and São Tome in Paraná, Brazil, the presence of STACN and STACP was found, with predominance of STACN. These isolates showed high resistance to oxacillin, penicillin G and erythromycin.
The identification of high percentages of multidrug resistance found in Staphylococcus isolates from bat is very important, considering the ability of these animals to fly and the possibility of spreading these agents.
The presence of the mecA gene was also observed in one of the STACP isolates, which is an alarming factor since fruit bats colonized by Staphylococcus spp. with the mecA gene, can easily spread these microorganisms far with flying.
In view of the results found, it is important to emphasize that anthropic actions exerted in the natural environment, especially those directly related to the use of antimicrobials in clinical treatments, in the disinfection of hospital environments and in agricultural products, may be the main reasons for the presence of bacteria resistant in bats, since there is no effective control of the use of antimicrobials, thus exempting this animal species from being responsible for this phenomenon.
Thus, it is essential that more studies are carried out to accurately identify the Staphylococcus species associated with bats, as well as the identification of other genes associated with resistance in these organisms, emphasizing the characteristics of bats that make them possible reservoirs and enable the spread of these bacteria in the environment.
Author Contributions
Writing of the research project and the scientific article, M.A.M.; Collection of biological samples, I.C.S; Animal capture, M.H.C and G.M.; Molecular laboratory analysis and tabulation of results, A.C.C.A.B; Phenotypic laboratory analysis and tabulation of results, F.G.C.B; Scientific writing review and tabulation of results, L.A.M; Molecular laboratory analysis, K.S; Phenotypic laboratory analysis, J.H.; Collection of biological samples, M.D.; English revision, M.A.; Scientific writing revision, L.N.B.; Oversight of all project stages and scientific writing revision, D.D.G. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by Universidade Paranaense and the Araucária Foundation (CP 09/2016 Institutional Program for Basic and Applied Research - Protocol: 47661.491.48325.12122016 and CP 10/2018 Pro-Equipment Program - Protocol: 51176.543.48325.29102018) and Coordenação de Aperfeiçoamento de Pessoal de Nível Superior (CAPES) grant PROSUP scholarships.
Institutional Review Board Statement
The experiment was approved by the Research Ethics Committee on Animal Experimentation of Universidade Paranaense (CEPEEA—protocol 34772/2018). (Annex 1 and 2).
Data Availability Statement
The data presented in this study are available on request from the corresponding author. The data are not publicly available due to ethical and privacy issues.
Acknowledgments
The authors thank the Universidade Paranaense; Fundação Araucária; and Coordenação de Aperfeiçoamento de Pessoal de Nível Superior (CAPES).
Conflicts of Interest
The authors declare no conflict of interest.
References
- Pourramezan, N.; Ohadian Moghadam, S.; Pourmand, M. R. Methicillin-resistant Staphylococcus aureus tracking spread among health-care workers and hospitalized patients in critical wards at a university hospital, Tehran, Iran. New Microbes New Infect 2019, 27, 29–35. [CrossRef]
- Van Duijkeren, E.; Box, A.T.A.; Heck, M.E.O.C.; Wannet, W.J.B.; Fluit, A.C. Methicillin-resistant staphylococci isolated from animals. Vet. Microbiol. 2004, 103, 91–97. [CrossRef]
- Mrochen, D.M.; Grumann, D.; Schulz, D.; Gumz, J.; Trübe, P.; Pritchett-Corning, K.; Johnson, S.; Nicklas, W.; Kirsch, P.; Martelet, K. van den Brandt, J.; Berg, S.; Bröker, B.M.; Wiles, S.; Holtfreter, S. Global spread of mouse-adapted Staphylococcus aureus lineages CC1, CC15 , and CC88 among mouse breeding facilities. Int. J. Med. Microbiol. 2018, 308, 598–606. [CrossRef]
- Blodkamp, S.; Kadlec, K.; Gutsmann, T.; Naim, H.Y.; von Köckritz-Blickwede, M.; Schwarz, S. In vitro activity of human and animal cathelicidins against livestock-associated methicillin-resistant Staphylococcus aureus. Vet. Microbiol. 2015, 194, 107-111. [CrossRef]
- Akobi, B.; Aboderin, O.; Sasaki, T.; Shittu, A. Characterization of Staphylococcus aureus isolates from faecal samples of the straw-coloured fruit bat (Eidolon helvum) in Obafemi Awolowo University (OAU), Nigeria. BMC Microbiol. 2012, 12, 279. [CrossRef]
- Walther, B.; Wieler, L.H.; Friedrich, A.W.; Hanssen, A.M.; Kohn, B.; Brunnberg, L.; Lübke-Becker, A. Methicillin-resistant Staphylococcus aureus (MRSA) isolated from small and exotic animals at a university hospital during routine microbiological examinations. Vet. Microbiol. 2008, 127, 171-178. [CrossRef]
- Russo, D.; Ancillotto, L. Sensitivity of bats to urbanization: A review. Mamm. Biol. 2015, 80, 205-212Buzón-Durán, L.; Capita, R.; Alonso-Calleja, C. Antibiotic susceptibility of methicillin-resistant staphylococci (MRS) of food origin: A comparison of agar disc diffusion method and a commercially available miniaturized test. Food Microbiol. 2018, 72, 220–224.
- Vandžurová, A.; Bačkor, P.; Javorský, P.; Pristaš, P. Staphylococcus nepalensis in the guano of bats (Mammalia). Vet. Microbiol. 2013, 164, 116-121. [CrossRef]
- Sens-Junior, H.; Trindade, W.A.; Oliveira, A.F.; Zaniolo, M.M.; Serenini, G.F.; Araujo-Ceranto, J.B.; Gonçalves, D.D.; Germano, R.M. Bacterial resistance in bats from the Phyllostomidae family and its relationship with unique health. Pesqui Vet Bras 2018, 38, 1207–1216. [CrossRef]
- Olatimehin, A.; Shittu, A.O.; Onwugamba, F.C.; Mellmann, A.; Becker, K.; Schaumburg, F. Staphylococcus aureus Complex in the Straw-Colored Fruit Bat ( Eidolon helvum ) in Nigeria. Front Microbiol 2018, 9, 1–7. [CrossRef]
- Liao, X.; Cullen, P.J.; Liu, D.; Muhammad, A.I.; Chen, S.; Ye, X.; Wang, J.; Ding, T. Combating Staphylococcus aureus and its methicillin resistance gene (mecA) with cold plasma. Science of the Total Environment 2018, 645, 1287–1295. [CrossRef]
- Alrabiah, K.; Al Alola, S.; Al Banyan, E.; Al Shaalan, M.; Al Johani, S. Characteristics and risk factors of hospital acquired–methicillin-resistant Staphylococcus aureus (HA-MRSA) infection of pediatric patients in a tertiary care hospital in Riyadh, Saudi Arabia. Int J Pediatr Adolesc Med 2016, 3, 71-77. [CrossRef]
- Shahbazian, J. H., Hahn, P. D., Ludwig, S., Ferguson, J., Baron, P., Christ, A., ... & Davis, M. F. (2017). Multidrug and mupirocin resistance in environmental methicillin-resistant Staphylococcus aureus (MRSA) isolates from homes of people diagnosed with community-onset MRSA infection. Applied and environmental microbiology, 83(22), e01369-17. [CrossRef]
- Kesharwani, A.K.; Mishra, J. Detection of β-lactamase and antibiotic susceptibility of clinical isolates of Staphylococcus aureus. Biocatal Agric Biotechnol 2019, 17, 720–725. [CrossRef]
- Greenhall, A. M.; Paradiso, J. L. Bats and bat banding. Bureau of Sport Fisheries and Wildlife Resource Publication 1968, 72, 47.
- Vieira, C. O. C. Ensaio monográfico sobre os quirópteros do Brasil. Arquivo de Zoologia do Estado de São Paulo 1942, 3, 219-471.
- Vizoto, L. D.; Taddei, V. A. Chave para determinação de quirópteros brasileiros. Boletim de Ciências 1973, 1, 1-72.
- Jones, J. K.; Carter, D. C. Annotated checklist, with keys to subfamilies and genera. In: Biology of bats of the new world family Phyllostomidae, part I. Spec. Publ. Mus. Tex. Tech Univ 1976, 107-38.
- Quinn, P. J.; Carter, M. E.; Markey, B.; Carter, G. R. Clinical Veterinary Microbiology, 2 nd ed.; Wolfe publishing : London, England, 1994.
- CLINICAL AND LABORATORY STANDARDS INSTITUTE (CLSI). 2018. M100–S127. Performance Standards for Antimicrobial Susceptibility Testing: 28th Informational Supplement. Clinical and Laboratory Standards Institute, Wayne, PA.
- Krumperman, P.H. Multiple Antibiotic Resistance Indexing of Escherichia coli to Identify High-Risk Sources of Fecal Contamination of Foodst. Appl. Environ. Microbiol 1983, 46, 165–170.
- Martineau, F.; Picard, F. J.; Roy, P. H.; Ouellette, M.; Bergeron, M. G. Species-specific and ubiquitous-DNA-based assays for rapid identification of Staphylococcus aureus. J. Clin. Microbiol 1998, 36, 618-623. [CrossRef]
- Murakami, K.; Minamide, W.; Wada, K.; Nakamura, E.; Teraoka, H.; Watanabe, S. Identification of methicillin-resistant strains of staphylococci by polymerase chain reaction. J. Clin. Microbiol 1991, 29, 2240-2244. [CrossRef]
- Reis, N.R.; Peracchi, A.L.; Pedro, W.A.; Lima, I.P. Mamíferos do Brasil, 2nd ed.; Universidade Estadual de Londrina: Londrina, Brazil, 2011; pp. 437.
- Ortêncio-Filho, H.; Reis, N.R.; Pinto, D.; Anderson, R.; Testa, D.A.; Marques, M.A. Levantamento dos morcegos (Chiroptera, Mammalia) do parque municipal do Cinturão Verde de cianorte. Chiroptera Neotropical 2005 , 11, 211–215.
- Leandro, N.; Angelis, D.E.; Domingo, B.L. Parque Municipal Cinturão Verde De Cianorte – Módulo Mandhuy E Os Principais Impactos Da Área De Entorno. Rev GEOMAE 2011, 2, 51–70.
- Haddock, J.K.; Threlfall, C.G.; Law, B.; Hochuli, D.F. Light pollution at the urban forest edge negatively impacts insectivorous bats. Biological Conservation 2019, 236, 17–28. [CrossRef]
- Chaverri, G.; Kunz, T. H. Ecological determinants of social systems: perspectives on the functional role of roosting ecology in the social behavior of tent-roosting bats. Advances in the Study of Behavior 2010, 275-318.
- Witte, W.; Cuny, C.; Klare, I.; Nübel, U.; Strommenger, B.; Werner, G. Emergence and spread of antibiotic-resistant Gram-positive bacterial pathogens. Int J Med Microbiol 2008, 298, 365–377. [CrossRef]
- Gomes, G. P. L. A.; Souza, A. C. S.; Leão-Vasconcelos, L. N.O; Melo Costa, D.; Alves, S. B.; Neves, H. C. C.; Pereira, M. S. Manual resuscitators in successive use in the same patient: reservoir of multi- and extensively resistant bacteria. J. Hosp. Infect 2017, 95, 87–90. [CrossRef]
- Pristas, P.; Vandz, A. Staphylococcus nepalensis in the guano of bats (Mammalia). Vet. Microbiol 2013, 164, 116-121.
- Nowakiewicz, A.; Ziółkowska, G.; Zięba, P.; Gnat, S.; Wojtanowicz-Markiewicz, K.; Trościańczyk, A. Comparative Immunology , Microbiology and Infectious Diseases Coagulase-positive Staphylococcus isolated from wildlife : Identification , molecular characterization and evaluation of resistance profiles with focus on a methicillin-resistant strain. Comp. Immunol. Microbiol. Infect. Dis 2016, 44, 21–28.
- Ben Yahia, H.; Chairat, S.; Hamdi, N.; Gharsa, H.; Sallem, R.B.; Ceballos, S.; Torres, C.; Slama, K. B.Antimicrobial resistance and genetic lineages of faecal enterococci of wild birds: Emergence of vanA and vanB2 harbouring Enterococcus faecalis. Innov Food Sci Emerg Technol 2018, 52, 936–941. [CrossRef]
- Garrido, A. M.; Gálvez, A.; Pulido, R. P. Antimicrobial resistance in enterococci. Journal of Infectious Diseases and Therapy 2014, 2.
- Kmeť, V.; Čuvalová, A.; Stanko, M. Small mammals as sentinels of antimicrobial-resistant staphylococci. Folia Microbiol 2018, 63, 665–668. [CrossRef]
- Watanabe, S.; Ito, T.; Sasaki, T.; Li, S.; Uchiyama, I.; Kishii, K.; Kikuchi, K.; Skov R.L.; Hiramatsu, K. Genetic diversity of staphylocoagulase genes (coa): insight into the evolution of variable chromosomal virulence factors in Staphylococcus aureus. PLoS One 2009, 4, 5714. [CrossRef]
- Becker, K.; Heilmann, C.; Peters, G. Coagulase-negative staphylococci. Clinical microbiology reviews 2014, 27, 870-926.
- Van Alen, S.; Ballhausen, B.; Peters, G.; Friedrich, A. W.; Mellmann, A.; Köck, R.; Becker, K. In the centre of an epidemic: Fifteen years of LA-MRSA CC398 at the University Hospital Münster. Vet. Microbiol 2017, 200, 19–24. [CrossRef]
- Bonsaglia, E. C. R.; Silva, N. C. C.; Rossi, B. F.; Camargo, C. H.; Dantas, S. T. A.; Langoni, H.;Guimarães, F.F.; Lima, F.S.; Fitzgerald, J.R.; Júnior, F.; Rall, V. L. M. Molecular epidemiology of methicillin-susceptible Staphylococcus aureus (MSSA) isolated from milk of cows with subclinical mastitis. Microbial Pathogenesis 2018, 124, 130–135. [CrossRef]
- Yang, F.; Liu, L.; Wang, L.; Wang, X.; Li, X.; Luo, J.; Zhang, Z.; Zhang, S.; Yan, Z.; Li, H. Penicillin-resistant characterization of Staphylococcus aureus isolated from bovine mastitis in Gansu, China. J Integr Agric 2017, 16, 1874–1878. [CrossRef]
- Miller, W. R.; Munita, J. M.; Arias, C. A. Mechanisms of antibiotic resistance in enterococci. Expert Rev Anti Infect Ther 2014, 12, 1221–1236. [CrossRef]
- Bouchiat, C.; El-Zeenni, N.; Chakrakodi, B.; Nagaraj, S.; Arakere, G.; Etienne, J. Epidemiology of Staphylococcus aureus in Bangalore, India: Emergence of the ST217 clone and high rate of resistance to erythromycin and ciprofloxacin in the community. New Microbes New Infect 2015, 7, 15–20. [CrossRef]
- Dadashi, M.; Hajikhani, B.; Darban-Sarokhalil, D.; van Belkum, A.; Goudarzi, M. Mupirocin Resistance in Staphylococcus aureus: A Systematic Review and Meta-Analysis. J Glob Antimicrob Resist 2019. [CrossRef]
- von Wintersdorff, C. J.; Penders, J.; Stobberingh, E. E.; Lashof, A. M. O.; Hoebe, C. J.; Savelkoul, P. H.; Wolffs, P. F. High rates of antimicrobial drug resistance gene acquisition after international travel, the Netherlands. Emerging Infectious Diseases 2014, 20, 649–657.
- Mühldorfer, K. Bats and Bacterial Pathogens: A Review. Zoonoses Public Health 2013, 60, 93–103. [CrossRef]
- Ito, T.; Hiramatsu, K.; Tomasz, A.; De Lencastre, H.; Perreten, V.; Holden, M. T.; Coleman, D. C.; Goering, R.; Giffard, P. M.; Skov, R. L.; Zhang, K.; Westh, H.; O'Brien, F.; Tenover, F. C.; Oliveira, D. C.; Boyle-Vavra, S.; Laurent, F.; Kearns, A. M.; Kreiswirth, B.; Ko, K. S.; Grundmann, H.; Sollidu, J. E.; John Jr., J. F.; Daum, R.; Soderquist, B.; Buist, G. Guidelines for reporting novel mecA gene homologues. Antimicrob Agents Chemother 2012, 56, 4997-4999.
- Koukos, G.; Sakellari, D.; Arsenakis, M.; Tsalikis, L.; Slini, T.; Konstantinidis, A. Prevalence of Staphylococcus aureus and methicillin resistant Staphylococcus aureus ( MRSA ) in the oral cavity. Arch. Oral Biol 2015, 60, 1410–1415. [CrossRef]
- Wendlandt, S.; Feßler, A. T.; Monecke, S.; Ehricht, R.; Schwarz, S.; Kadlec, K. The diversity of antimicrobial resistance genes among staphylococci of animal origin. Int. J. Med. Microbiol, Jena 2013, 303, 338–349. [CrossRef]
- Resman, F.; Thegerström, J.; Månsson, F.; Ahl, J.; Tham, J.; Riesbeck, K. The prevalence, population structure and screening test specificity of penicillin-susceptible Staphylococcus aureus bacteremia isolates in Malmö, Sweden. Journal of Infection 2016, 73, 129–135.
- Cheatham, S.; Thapaliya, D.; Taha, M.; Milliken, K.; Dalman, M. R.; Kadariya, J.; Grenier, D.; Smith, T. C. Prevalence of Staphylococcus aureus and methicillin-resistant S aureus on environmental surfaces in Ohio nursing homes. American Journal of Infection Control 2019, 000, 1–5. [CrossRef]
- Peterson, L. R.; Boehm, S.; Beaumont, J. L.; Patel, P. A.; Schora, D. M.; Peterson, K. E.; Burdsall, D.; Hines, C.; Fausone, M.; Robicsek, A.; Smith, B. A. Reduction of methicillin-resistant Staphylococcus aureus infection in long-term care is possible while maintaining patient socialization: A prospective randomized clinical trial. American Journal of Infection Control 2016, 44, 1622-1627. [CrossRef]
- Banskar, S.; Mourya, D.T.; Shouche, Y.S. Bacterial diversity indicates dietary overlap among bats of different feeding habits. Microbiol Res 2016, 182, 99–108. [CrossRef]
- Feßler, A. T.; Schuenemann, R.; Kadlec, K.; Hensel, V.; Brombach, J.; Murugaiyan, J.; Oechtering, G.; Burgener, I.A.; Schwarz, S. Methicillin-resistant Staphylococcus aureus (MRSA) and methicillin-resistant Staphylococcus pseudintermedius (MRSP) among employees and in the environment of a small animal hospital. Vet. Microbiol 2018, 221, 153–158. [CrossRef]
- Baptista, I.; Rocha, S.M.; Cunha, Â.; Saraiva, J.A.; Almeida, A. Inactivation of Staphylococcus aureus by high pressure processing : An overview. Innov Food Sci Emerg Technol 2016, 36, 128–149. [CrossRef]
Figure 1.
Chiroptera species captured in the cities of Cianorte, Japurá and São Tomé in Paraná, Brazil, 2017-2019.
Figure 1.
Chiroptera species captured in the cities of Cianorte, Japurá and São Tomé in Paraná, Brazil, 2017-2019.

Figure 2.
PCR assay for the mecA gene in isolates of Staphylococcus spp. From rectal and oral swabs of bats captured in the cities of Cianorte, Japurá and São Tomé in Paraná, Brazil, 2017-2019. Legend: 2% agarose gel. Lane 1 – molecular marker; lane 5 – positive sample for the mecA gene; lane 2 – negative control (ATCC 43300); lane 3 – positive control (ATCC 25923).
Figure 2.
PCR assay for the mecA gene in isolates of Staphylococcus spp. From rectal and oral swabs of bats captured in the cities of Cianorte, Japurá and São Tomé in Paraná, Brazil, 2017-2019. Legend: 2% agarose gel. Lane 1 – molecular marker; lane 5 – positive sample for the mecA gene; lane 2 – negative control (ATCC 43300); lane 3 – positive control (ATCC 25923).
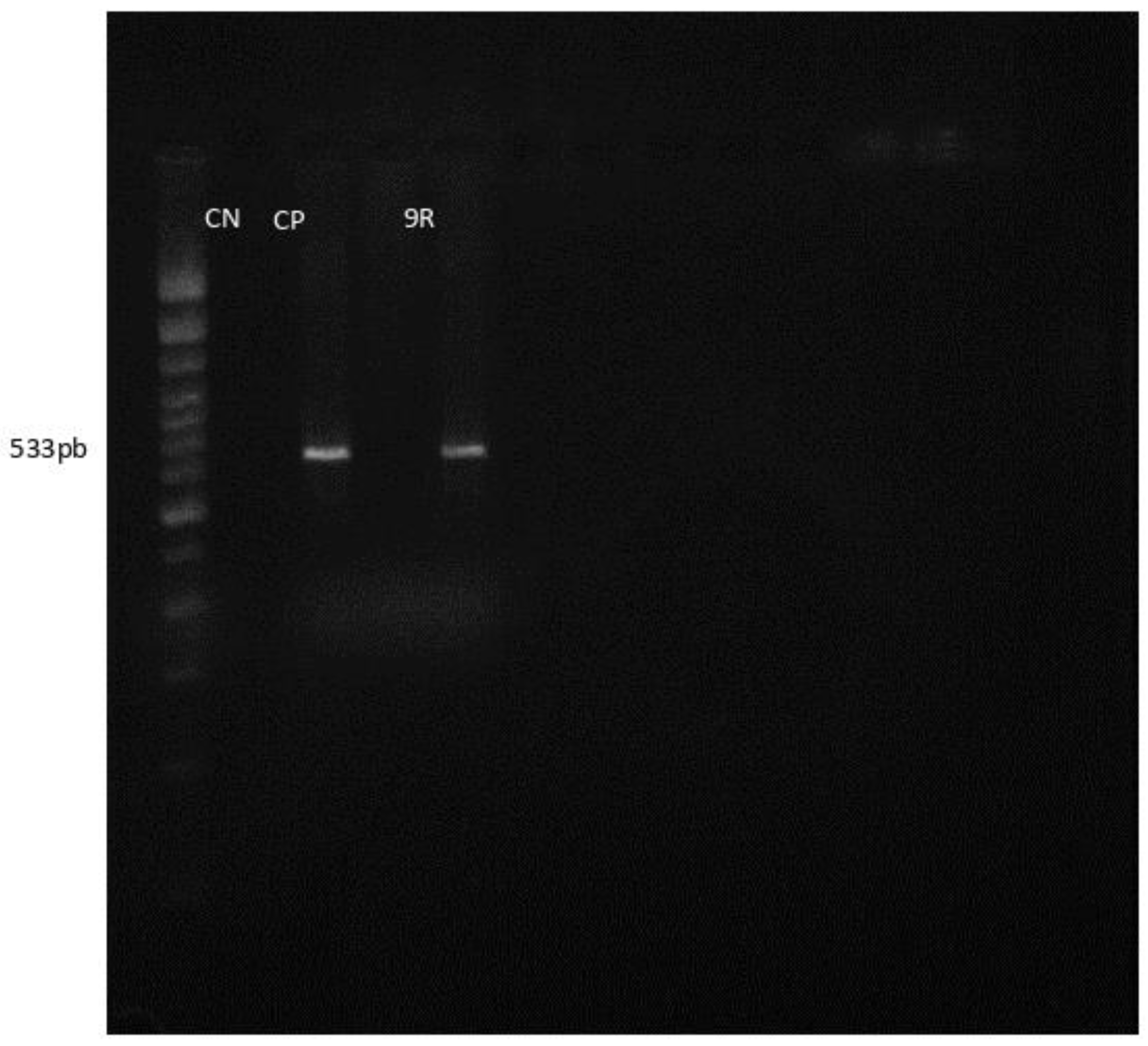

Table 1.
Resistance profiles to different antimicrobials of coagulase-negative Staphylococcus spp (CNSTA) isolates from bat guano in the cities of Cianorte, Japurá and São Tomé in northwest Paraná, Brazil, 2019.
Table 1.
Resistance profiles to different antimicrobials of coagulase-negative Staphylococcus spp (CNSTA) isolates from bat guano in the cities of Cianorte, Japurá and São Tomé in northwest Paraná, Brazil, 2019.
| Bacterial Resistance | |||||
|---|---|---|---|---|---|
| Antimicrobials | Feces | ||||
| R | Total | % | |||
| Ampicillin Cephalothin Oxacillin Amoxicillin+clavulanic acid Norfloxacin Enrofloxacin Gentamicin |
5 5 5 4 1 0 0 |
6 6 6 6 6 6 6 |
83.33 83.33 83.33 66.66 16.66 0% 0% |
||
| TOTAL | 20 | ||||
Legend: R = Resistant; % = Percentage.
Table 2.
Resistance profile and antimicrobial multidrug resistance indexes of isolates of coagulase-negative Staphylococcus spp. (CNSTA) from the guano of bats collected from roofs in the cities of Cianorte, Japurá, and São Tomé in northwestern Paraná, Brazil, 2019.
Table 2.
Resistance profile and antimicrobial multidrug resistance indexes of isolates of coagulase-negative Staphylococcus spp. (CNSTA) from the guano of bats collected from roofs in the cities of Cianorte, Japurá, and São Tomé in northwestern Paraná, Brazil, 2019.
| Sample | Antimicrobials and Resistance | Antimicrobial Resistance Index | |||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| AMC | AMP | CFL | ENO | GEN | NOR | OXA | |||||||||||
| 12 | S | S | S | I | S | R | S | 0.14 | |||||||||
| 13* | R | R | R | I | S | S | R | 0.57 | |||||||||
| 14 | S | R | R | S | S | S | R | 0.42 | |||||||||
| 15* | R | R | R | S | I | S | R | 0.57 | |||||||||
| 16* | R | R | R | S | S | S | R | 0.57 | |||||||||
| 19* | R | R | R | S | I | S | R | 0.57 | |||||||||
Legend: I — Intermediate, R — Resistant, S — Sensitive — AMC — Amoxicillin+clavulanic acid, AMP —Ampicillin, CFL — Cephalothin, ENO — Enrofloxacin, GEN — Gentamicin, NOR — Norfloxacin, OXA — Oxacillin.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



