
Submitted:
19 December 2023
Posted:
20 December 2023
You are already at the latest version
Abstract
Sechium compositum (Cucurbitaceae) is a wild species that is distributed in the Soconusco region, Chiapas, Mexico, and the border with Guatemala. This species has an intangible biochemical value resulting from the pharmacological relevance of its secondary metabolites. However, as a consequence of the lack of knowledge about its importance, it is being displaced from its habitat at an accelerated rate, incurring the risk of genetic loss. Therefore, an in vitro culture protocol with two experimental phases was evaluated to propagate, conserve, and regenerate this species. The first phases considered the multiplication of seedlings, adding seven concentrations (0.1, 0.2, 0.4, 0.6, 0.8, 1.0, 1.2 mg mL-1) of 6-benzylaminopurine (BA) and thidiazuron (TDZ) and evaluating the number of buds, shoots, and seedling height. The best multiplication response was recorded with 0.1, 0.2, 0.4, and 1.0 mg L-1 of BA and 0.1 mg L-1 of TDZ, as well as the MS base culture medium. With 0.1 mg L-1 of BA, 52-mm height, 1.36 shoots, and 9.22 buds were obtained, surpassing the MS control (MS culture medium alone); likewise, twice more roots (80%) and a 14% reduction in the bud structure were packed in 50 repetitions. The second phase consisted of inducing callus formation from stem and leaf explants through the addition of 0.5, 1.0, and 2.0 mg L-1 of TDZ and 2,4-Dichlorophenoxyacetic acid (2,4-D) to the medium. The results demonstrated that the induction of callogenesis in S. compositum has a better start from the stem in a medium with 2.0 mg L-1 of 2,4-D, whose value was 4.89 —equivalent to 97.8% callus formation around the explant. The addition of 500 mg L-1 of polyvinylpyrrolidone (PVP) is also suggested to reduce oxidation. This protocol represents a significant advance in the conservation, multiplication, and callogenesis of S. compositum and contributes to its rescue and revaluation in the face of the danger of extinction.
Keywords:
agrobiodiversity
; in vitro multiplication
; Cucurbitaceae
; underutilized species
; Sechium (P. Br.)
1. Introduction
Agrobiodiversity includes local or native varieties, tolerated, encouraged, cultivated, in process of domestication ecotypes, and their wild relatives [1,2]. Phytogenetic resources for food and agriculture, implicit in agrobiodiversity, play a fundamental role in human development, providing significant benefits [3] that contribute to the scientific, technological, socioeconomic, and cultural progress of megadiverse countries [4].
Some of these local varieties and their wild relatives have not yet been fully characterized, which creates a major limitation for the identification of new uses or improvement of known ones. In many cases, the commercial success of a genotype can indirectly displace local varieties whose use or consumption are less popular —especially wild relatives, which have been rendered fragile, placed in conditions of erosion, or lost by the lack of research about their potential uses [5,6,7].
The conservation of these phytogenetic resources requires the application of in situ and ex situ strategies [8]. The first focuses on the maintenance of plant species and traditional agricultural systems in the habitats of origin, where they have developed specific phenotypic and genetic characteristics. This effort entails the protection of natural areas, wild ecosystems, and other environments within their original context [8]. In this process, the lore of rural communities plays a crucial role, since their populations have been the guardians of these plants and their uses for generations [5,9,10].
Ex situ conservation is mainly carried out in germplasm banks or scientific collections [10,11]and consists of the preservation of orthodox and recalcitrant seeds [12,13] outside natural habitats. Both approaches play a fundamental role in the protection and conservation of the genetic diversity of plant species, contributing to food security and global biodiversity.
In this context, chayote (Sechium P.Br.) (Cucurbitaceae) emerges as an important phytogenetic resource for Mexico (its center of origin and domestication). The genus includes ten species, of which only S. edule and S. tacaco are cultivated as food [14]. The other eight (S. chinantlense, S. compositum, S. hintonii, S. talamancense, S. panamense, S. pittieri, S. venosum, and S. vilosum) are wild species [15].
S. compositum is distributed in the Soconusco region, Chiapas, Mexico, and the border with Guatemala [14]. In 2011, the proximity of rural communities placed five S. compositum populations in the status of very high fragility in Chiapas [16]. This limited geographical distribution and lack of knowledge of any use among rural inhabitants has promoted its displacement by economically profitable crops, such as coffee, corn, and forage species [16].
According to recent studies by [17], both the biological variants of S. edule and two identified morphotypes of S. compositum have a high content of secondary metabolites with pharmacological activity-specifically tetracyclic triterpenes, phenols, and flavonoids [18,19] with antileukemic [20] and antifungal [21] potential, which opens a window of opportunity for its revaluation and therefore contributes to its conservation.
The ex situ conservation of S. compositum faces significant challenges, given its climbing nature and recalcitrant seed, which hinders its preservation through traditional methods [22]. In response to these challenges, a viable alternative is its establishment under in vitro conditions. This approach not only allows the preservation of the species, but also its regeneration and possible reintegration into its original habitat [23,24]. Furthermore, in vitro preservation is a source of tissues that are valuable for bioprospecting research [18], as well as the induction of mutagenesis for future applications. This biotechnological technique has several applications, including the study of physiological aspects [25], clonal propagation [26], production of secondary metabolites [27] plant regeneration [28,29] and obtaining disease-free varieties [23,30,31].
A significant number of research works have focused on the in vitro establishment, conservation, and regeneration of S. edule [22,32,33,34,35,36] in addition to clonal propagation, rooting, and acclimatization protocols [22,37,38]. However, unlike the case of other species, explant-based callogenesis protocols have not been studied [39]. Callogenesis is the basis for massive in vitro propagation through indirect organogenesis or indirect somatic embryogenesis [40,41]; however, no such protocol has been developed for wild species.
The objective was to develop an in vitro multiplication protocol from bud explants, as well as a callogenesis protocol for Sechium compositum (Donn. Sm.) C. Jeffrey, with the purpose of obtaining tissue for the regeneration of the species, consequently contributing to its conservation and the possibility of studies for new uses.
2. Materials and Methods
The research was divided into two experimental phases. The first considered the multiplication of seedlings through the addition of 0.1, 0.2, 0.4, 0.6, 0.8, 1.0, 1.2 mg mL-1 concentrations of the 6-benzylaminopurine (BA, PhytoTechnology Laboratories® B800, Shawnee Mission, KS, USA) and thidiazuron (TDZ, Sigma-Aldrich® P6186, San Luis, MO, USA) growth regulators, in order to observe differences in number of buds, shoots, and seedling height, with the MS plant culture medium [42]. The second phase consisted of the induction of callus formation from stem and leaf explants, with the addition of 0.5, 1.0, and 2.0 mg L-1 concentrations of the TDZ and 2,4-D growth regulators to the medium.
2.1. Biological material
Vegetative material of S. compositum from accession 321-05 of the Sechium P. Br. Germplasm Bank (19° 08’ 48” N and 97° 57’ 00” W), was used. The original growth conditions of the accession are high evergreen forest. However, since its collection (2005), it has been acclimatized to mountain cloud forest conditions: 1,340 m.a.s.l., 19 °C average annual temperature, 85% relative humidity, and 2,250 mm annual precipitation. The soils are nutrient-rich vitric luvisols with moderate fertility, coarse texture, and fragments of volcanic glass, slightly acidic to acidic pH (4.3-6.5), abundance of organic matter, low levels of calcium, and high levels of iron, manganese, and zinc [43].
2.2. Experimental phase 1: disinfection procedure for the establishment of biological material
Twenty-centimeter-long tips of plagiotropic stems (vine) were collected up to the apical bud. The axillary buds were cut from the vine in the laboratory; the following disinfection procedure was then applied: they were washed with soap (Axion®) and water, placed in 70% (v/v) alcohol for 1.0 min, and disinfected with sodium hypochlorite bleach (Cloralex®) at 20% (v/v) for 10 min while stirring. They were immediately washed with sterile distilled water in a laminar flow hood. Finally, they were established in a MS medium supplemented with 1.5 mgL-1 Plant Preservation Mixture (PPMTM, Plant Cell Technology, Inc.). The responsive buds were multiplied and used for the development of biotrials for the establishment, multiplication, and induction of calluses.
2.2.1. In vitro base culture medium and general maintenance conditions
The base culture medium for the in vitro establishment, maintenance, and callogenesis was the MS [42] medium with vitamins (PhytoTechnology Laboratories® M519, Shawnee Mission, KS, USA) under standard conditions. They were gelled with 9 g L-1 of agar (PhytoTechnology Laboratories® A111, Shawnee Mission, KS, USA) supplemented with 30 g L-1 of D-sucrose (PhytoTechnology Laboratories® S391, Shawnee Mission, KS, USA); the result was sterilized in an automatic autoclave at 120 °C, at 0.1 MPa pressure for 20 min. Seven mL of medium were placed in 15-cm glass tubes with plastic lids; meanwhile, 25 mL of medium were placed in glass bottles with plastic lids. The pH was adjusted from 5.7 to 5.8 with 1.0 N sodium hydroxide (BAKER ANALYZED®) or 1.0 N hydrochloric acid (MERCK®). The incubation conditions for all tests were 25 ±1 °C and a 16-h light photoperiod with 3,000 lux intensity.
2.2.2. In vitro multiplication
Two growth regulators were added to the MS base culture medium to induce seedling growth and in vitro shoot formation from buds of S. compositum: 1) 6-benzylaminopurine (BA PhytoTechnology Laboratories® B800, Shawnee Mission, KS, USA), at 0.1, 0.2, 0.4, 0.6, 0.8, 1.0, 1.2 mg mL-1 concentrations; and 2) thidiazuron (TDZ, Sigma-Aldrich® P6186, San Luis, MO, USA) in the same concentrations. The control consisted only of a MS culture medium (MS control). The 14 treatments plus the control were distributed in a completely randomized arrangements with ten repetitions. They were incubated at 25 ± 1 °C with a 16-h light photoperiod. The evaluation was carried out after four months, recording stem length (mm), number of shoots, number of buds, presence of roots, and formation of structures (e.g., calli). To check the effect of the minimum BA concentration, buds were sown on a MS medium supplemented with 0.1 mg L-1 of BA and compared with the MS base culture medium. The sample size was n=50 repetitions per completely randomized treatment.
2.3. Experimental phase 2: callogenesis induction
Stem and leaf explants were taken to obtain callus in vitro. Two growth regulators were added to the MS base culture medium: 2,4-D (2,4-Dichlorophenoxyacetic acid; Phytotechnology Laboratories® D295, Shawnee Mission, KS, USA) and TDZ (Sigma-Aldrich® P6186, San Luis, MO, USA), both with 0.5, 1.0, 2.0 mg L-1 concentrations and 18 repetitions. The control was MS culture medium by itself (MS control), with 10 repetitions per explant. A completely randomized design was applied (n=236). They were incubated at 25 ±1 °C, with a 16-h light photoperiod, and 3,000 lux intensity. The evaluation was carried out 30 d after establishment, recording fresh weight (g) and level of callus formation (%) according to the scale described in Table 1.
The estimated volume of callus was calculated using the regression model, based on non-intrusive measurements reported by Ramos-Parra et al. [44]. with the following equation:
Estimated volume of callus = 1.019 + 0.044(d12h) + 0.106(d22h)
where: ß’s= numbers that indicate the parameters of the models (1.019, 0.044, 0.106).
d1= diameter 1, linear dimension parallel to the medium and largest horizon, considering that the container is in front of the observer.
d2= diameter 2, dimension parallel to the medium and perpendicular to d1.
h= height (h), dimension perpendicular to d1, which were substituted in the model to obtain the estimated callus volume.
Prevention of oxidation in calli
Independent tests were carried out with activated carbon and polyvinylpyrrolidone (PVP) to reduce the effect of oxidation on calli, to obtain calli that maintained normal growth characteristics, to increase the areas of potentially active calli, and to decrease brown-yellow areas.
A) Test with activated carbon
One hundred twenty stems were sown in a medium with 2.0 mg L-1 of 2,4-D and 2.0 mg L-1 of activated carbon (PhytoTechnology Laboratories® C325, Shawnee Mission, KS, USA). Callus formation was evaluated during 30 d.
B) Test with polyvinylpyrrolidone (PVP) as antioxidant agent
In this trial, stems were sown in the callus formation medium (2.0 mg L-1 of 2,4-D) with 250 mg L-1 and 500 mg L-1 concentrations of polyvinylpyrrolidone (PVP; Sigma-Aldrich® P2307, San Luis, MO, USA), and they were compared with the control medium without PVP. The development of each callus (%) was evaluated taking into consideration the characteristics described in Table 1, under a completely randomized design with ten repetitions. In a parallel experiment, level 5 calli (Table 1) formed from stem were resown in a medium with 2.0 mg L-1 of 2,4-D, to which 250 mg L-1 and 500 mg L-1 of polyvinylpyrrolidone were added. The percentage of callus formation was evaluated considering characteristics such as growth of the mass, decrease in brown areas (sign of oxidation), and increase in light areas (potential active callus) (Table 2). Both experiments were established under a completely randomized design, with ten repetitions, and were evaluated at 20 and 30 d. The results were analyzed using the Kruskal-Wallis One Way Analysis of Variance on Ranks test.
2.4. Feasibility of the callogenesis protocol
The repeatability of the callogenesis protocol was checked by sowing 120 stems in a MS medium supplemented with 2.0 mg L-1 of the 2,4-D regulator. Callus formation, diameter 1, diameter 2, height, and estimated volume were evaluated at 25 d, using the equation proposed by [44]. Subsequently, 60 calli were divided into two fragments and resown in a MS medium supplemented with a 2.0 mg L-1 concentration of the 2,4-D regulator and 500 mg L-1 of PVP. At 30 d, callus formation, diameter 1, diameter 2, height, and estimated volume were evaluated following the equation proposed by [44].
2.5. Statistical analysis
All treatments had a completely randomized experimental design. When the assumptions of normality and homogeneity were not met, the nonparametric Kruskal-Wallis test was applied. For the bud multiplication test, the following was applied: the Wilcoxon analysis of multiple range comparison of paired sides by the Dwass Method [45], Steel [46], Critchlow-Fligner [47] (DSCF method) was used to determine the following variables: seedling height, number of buds, and number of shoots. For the callus and root formation variables, it was performed using the Mann-Whitney U Test (P = <0.001). The callogenesis experiment was analyzed through the Wilcoxon analysis of multiple range comparison of paired sides by the Dwass Method [45], Steel [46], Critchlow-Fligner [47] (DSCF method). In addition, a Mann-Whitney U Test comparative analysis (P = <0.001) was carried out separately for calli inducted from the stem explant and from the leaf explant. The efficiency of adding PVP to the medium to reduce oxidation in callus was determined by the Mann-Whitney U Test (P = <0.001). The data were analyzed in the SAS® 2022 On Demand for Academics statistical software, online, and using the SigmaPlot [14.0] software.
3. Results
3.1. Experimental phase 1: in vitro multiplication
Under the comparison of ranges by the DSCF method (P < 0.05), treatment 2 (1.0 mg L-1 of BA) recorded the tallest seedling heights (64.46 mm), which doubled the value of the control and other treatments (Table 3).
Treatment 9, which consisted of the application of a 0.1 mg L-1 concentration of TDZ demonstrated a significant increase in growth, with a 30.32 mm average value. Regarding the number of buds, outstanding results were observed with 0.1, 0.2, 0.4, and 1.0 mg L-1 concentrations of BA (treatments 2, 3, 4, and 7), as well as with 0.1 mg L-1 of TDZ (treatment 9) and the control group; all of them exhibited optimal responses, with a range of 10-16 buds (Table 3).
Figure 1 shows that treatments 6, 7, 8, and 10 had a different callus and root formation response than the control (P= <0.001). This could be the starting point for the development of trials that promote greater root development or callus formation. The factors that promote callus formation were the type and the concentration of the growth regulator.
Optimization treatments aimed at the in vitro growth of S. compositum must address several essential traits, including seedling size, number of buds, and induction of a greater number of shoots. These considerations are essential to ensure adequate multiplication and root development, even in situations where callus formation is limited. In this study, treatments that can meet such conditions include 0.1 mg L-1 and 1.0 mg L-1 of BA and 0.1 mg L-1 of TDZ (Figure 1).
To validate the results of the first trial, 0.1 mg L-1 of BA —one of the concentrations that optimize the process— was compared with the MS control. According to the analysis of variance (P=<0.001), there were differences in all the variables evaluated. The following average values were recorded for the 50 repetitions evaluated for each treatment with 0.1 mg L-1 of BA: a height of 52 mm, 1.36 shoots, and 9.22 buds. These figures were higher than the MS control (Figure 2).
Remarkably, root formation was observed in 50 representative repetitions: 0.1 mg L-1 of BA generated twice the number of root than the MS control (80%); even callus formation was observed (16%) without limiting its growth (Figure 2). In this regard, the cytokinins can induce high proliferation and cell division —i.e., they increase the multiplication rate of plants, in addition to inducing root elongation and an increase in shoot production.
When the five reseedings of S. compositum are exceeded, a structure of packed buds (“rosettes”) is usually formed (Figure 3). Even when it has a significant number of buds, this structure is difficult to divide for their multiplication. This situation may be caused by a loss of morphogenic competence in the mature material (morphological, biochemical, and molecular differences); consequently, the ontogenic age of the plant material causes the absence of elongation, since younger plant explants led to a greater elongation rate of the axillary buds.
Figure 2 indicates a significant decrease in the formation of the rosette-shaped structure (Figure 3C) when 0.1 mg L-1 of BA was added to the base culture medium, reducing it threefold compared to the MS control, in which a 42 % rosette formation was observed. These results suggests that BA acts as a rejuvenating agent for the material. Both cytokinins and BA promote the elimination of the dormancy of axillary buds, as well as the formation of lateral shoots.
3.2. Experimental phase 2: callogenesis induction
In the separate analysis of the explants (leaf and stem), compared with the control, the treatments with 2,4-D resulted in differences for all the variables evaluated using the Mann-Whitney U Test (P = <0.001). Furthermore, the concentration of this regulator can vary within a range of 0.5 to 2.0 mg L-1, highlighting that the highest concentration resulted in a notable improvement in callus development (Figure 4).
The callus formed was a compact mass with a white and cottony periphery, as well as some light yellow-green middle areas, especially in levels 3 and 4 (Figure 5D,E,I,J). The explants of level 5 calli are fully covered with a white layer (Figure 6A–C). After 40 d in the medium, an oxidation process began, during which the calli changed from light white yellow to brown-yellow (Figure 7D). This oxidation indicator lasted about 60 d and then the tissue died.
Specific assays for callogenesis of S. compositum showed that the establishment of stem and leaf fragments in a MS medium supplemented with 0.5, 1.0, and 2.0 mg L-1 of TDZ is hardly the best option for callus formation, especially with leaf explants, because callus formation does not surpass level 2, i.e., less than 50% (Table 4, Figure 4). In addition, the tissue mass was green in the center and brown in the periphery (Figure 5A–C,F–H). On the contrary, when the medium was supplemented with three concentrations of 2,4-D (0.5, 1.0, and 2.0 mg L-1), an average ~level 5 callus formation was induced —i.e., 100% callus formation at 30 d from the stem explant. The highest concentration was statistically different in callus formation (4.89 ± 0.08).
In this sense, the success of callogenesis of S. compositum depends on the type of growth regulator and the explant. The concentration did not show differences in callus fresh weight, diameter, height, volume, and root formation. Similar results were obtained by Soto-Contreras et al. [39] who evaluated the effect of 0.5, 1.0, and 1.5 mg L-1 concentrations at of 2,4-Dichlorophenoxyacetic acid on the induction of callus formation from different vitroplant explants of S. edule. Their results show that, with the highest concentration, the nodal segments recorded 100% formation of white calli and a fresh weight of 1.74 g, while the calli of the leaf segments were greenish and had a fresh weight of 0.35 g.
Control of callus oxidation
a) Activated carbon
The inclusion of an antioxidant in the medium is an alternative to reduce oxidation in in vitro culture. In this case, 120 repetitions of an experiment in which callus formation was induced from the stem in the callogensis medium (2.0 mg L-1 of 2,4-D) with 2.0 mg L-1 of activated carbon were evaluated. Under these conditions, the stems showed induction of callus formation at 7 d, with swelling of the explant at the ends where the cut was made (Figure 6A). After 15 days (Figure 6B), the callus development around the stem was more noticeable. However, after 30 d of evaluation (Figure 6C), 100% stopped their development with level 3 calli (Table 1) and generated roots; only one explant generated shoots and leaves (Figure 6D). Therefore, activated carbon in a callus formation initiation process is not suitable for S. compositum.
There are reports that mention that the addition of activated charcoal to the best callus formation treatments generates a negative effect (for example, slow cell proliferation of Brosimum alicastrum callus).
b) Application of polyvinylpyrrolidone (PVP)
According to Figure 8, the level of development of calli with desirable characteristics (Figure 7A,C) at 20 and 35 d (Table 1) show that the calli in the formation medium (control) reached level 5 (100%) at 35 d. Meanwhile, when 250 mg L-1 of PVP were added to the medium, the formation level reached 2.2 ± 0.13 (30%); for its part, the application of 500 mg L-1 of PVP to the medium resulted in a formation level of 3.9 ± 0.38 (72.5%). Therefore, adding PVP from the initial stage of callus formation should decrease callus induction from the stem by 27.5%. Both the control and the 500 mg L-1 concentration at of PVP showed differences at 35 d, according to the Mann-Whitney U Test (P = <0.001).
Figure 9 indicates that the resowing of callus of average level (4.5) from stem explant, in a medium with 2.0 mg L-1 of 2,4-D, to which two concentrations of polyvinylpyrrolidone (PVP: 250 and 500 mg L-1) were added, did not register differences (P = >0.05) at 20 and 35 d. However, with 500 mg L-1, 100% calli with desired characteristics were obtained in a resown callus (Figure 7C) —i.e., they presented mostly active zones, increase in mass, root formation, and transparent medium (Table 2). In that sense, adding PVP to the medium for resown calli helps to reduce the oxidation process.
3.3. Validation of the callogenesis protocol
As a result of the validation of the callogenesis protocol, the 120 calli sown from stem in a medium with a of 2.0 mg L-1 concentration of the 2,4-D regulator recorded the following results at 25 d: average formation values of up to 70% ±29.6%; diameter 1 of 7.74 ± 2.5 mm; diameter 2 of 8.41 ± 2.4 mm; height of 7.35 ±2.2 mm; and a volume of 27.89 ±19.5 mm3. Half of these calli were divided into two groups and resown for new callogenesis, on a medium that included 500 mg L-1 of PVP. A 30-day evaluation reported 100% callus development with active zones, increase in mass, root formation, and transparent medium.
4. Discussion
4.1. Experimental phase 1: in vitro multiplication
Previous work with S. edule shows similar responses to the application of different concentrations of growth regulators (e.g., BA and TDZ). Authors such as [36] recorded a more efficient propagation of S. edule plants by adding 0.1 mg L-1 of BA, and 0.1 mg L-1 of GA3 to the MS base culture medium. Mora also identified that concentrations greater than 0.5 mg L-1 of BA induce callus formation. Other authors such as [48] observed that the association of different growth regulators with BA had significant advantages (e.g., greater shoot length) over KN and 2iP, after four weeks of culture. They also mention the synergistic effect of BA and IAA when the induction of shoots is similar to BA. The maximum number of nodes and shoot length was observed with 2.0 μM of BA; this number increased when they were subcultured in a culture medium with the same concentration.
The best medium for the induction of longer roots contained 10 mΜ of indole-3-butyric acid. With lower concentrations of TDZ, multiple adventitious buds were induced, but shoots did not regenerate as was the case of an inoculation of different concentrations of BA.
BA evidently promotes the generation of shoots, growth, and stimulation of root formation, in both S. edule and S. compositum, enhancing micropropagation. Authors such as [22] induced the proliferation of axillary shoots from axenic nodal segments obtained from seedlings germinated in vitro. Nodal segments were cultured in MS medium supplemented with 0.1 mg L-1 of 6-benzylaminopurine (BA), producing 8.0 ± 0.4 shoots per explant, with a 92% regeneration frequency. Furthermore, the regeneration of the seedlings was optimized through the addition of 0.1 mg L-1 of BA and 0.05 mg L-1 of gibberellic acid (GA3) to the medium, obtaining 5.3 ± 1.9 shoots per average explant.
Regarding the number of shoots, treatments 2, 7, and 9 stood out with an average of 2.1, 2.0, and 2.1 shoots, respectively. These data are considered indicative of the multiplication rate for each treatment (Table 3). The results highlight the significantly positive influence of the treatments in shoot growth and multiplication. Overall, this difference in responses is since BA significantly stimulates the formation of axillary buds, while the TDZ response is associated with the formation of adventitious buds [49].
TDZ favored the formation of basal calli from 0.2 mg L-1 to 1.2 mg L-1 (Treatments 10, 11, 12, 13, 14, and 15). In the case of BA, treatments 5, 6, and 8 (0.6 mg L-1, 0.8 mg L-1, and 1.2 mg L-1, respectively) formed more calli than treatments 2, 3, 4, and 7, which had the lowest concentrations (0.1, 0.2, 0.4, and 1.0 mg L-1, respectively). For example, in their research on the in vitro application of different concentrations of cytokinins in Castanea sativa Mill [49] demonstrated that two of their TDZ and BA treatments recorded a higher (80%) formation of basal callus, which improved the establishment of the seedlings. They also mention that the basal callus accumulates substances and hormones necessary for the in vitro response, which, according to [50], suggesting a relationship between the callus formation and the establishment of seedlings.
In our study, we observed that BA increases the formation of shoots and number of buds. Important differences were recorded comparing the MS control (MS culture medium by itself) and the MS base culture medium (MS medium supplemented with 0.1 mg L-1 of 6-benzylaminopurine) that improved the micropropagation of S. compositum from buds. BA (6-benzylaminopurine) has been the growth regulator par excellence for the optimization of the induction, multiplication, or regeneration stages by direct organogenesis in other species and has recorded more shoots and longer seedlings than MS control. Likewise, depending on the species or variety, and the concentrations or combinations with other regulators, media supplemented with BA are significantly more potent [51,52,53].
4.2. Experimental phase 2: Callogenesis induction
Theoretically, high concentrations of auxins promote root formation, while high concentrations of cytokinins promote shoot regeneration. A balance between auxins and cytokinins leads to callus development [54]. However, multiple research works have reported that the use of 2,4-Dichlorophenoxyacetic acid (2,4-D) induces in vitro callus formation from various explants, positioning it as one of the regulators that better promotes callogenesis, whether, or not it is combined with cytokinins [55,56,57,58].
Other authors such as [23] have reported that cytokinins (BA and TDZ) promote callus formation. Using in vitro stem segments of S. edule, they obtained 70% callus formation with 1.0 mg L-1 of the BA growth regulator, and 76% with 0.4 mg L-1 of TDZ, both at 30 days of evaluation. Compared with the first stage of this research, high concentrations of TDZ (0.2-1.2 mg L-1) and BA (0.5, 0.6, and 1.2 mg L-1) promoted the formation of callus on the development of the seedling.
Table 4 and Figure 4 show that the best callus formation treatment was based on stem explant in a medium with 2.0 mg L-1 of 2,4-D (highest concentration), recording an average value of 4.89 (equivalent to 97.8% callus formation around the explant). The weight and volume variables indicated that the best treatments started from the stem explant and the medium enriched with 2,4-D, without differences between concentrations. The presence of a root is an indication that calli can become embryogenic —i.e., they can be the point of origin for the promotion of the regeneration of the seedling (indirect organogenesis) [59,60]; therefore, the best treatments started from the stem in a medium enriched with 2,4-D, in the three concentrations evaluated.
Likewise, [61] and other authors evaluated the effect of adding PVP to the culture medium as an antioxidant and observed that the treatment with 500 mg L-1 decreased the loss of explants of the guava variety CHRG (Psidium guajava L.), recording a higher frequency of explants and a lower level of necrosis by oxidation (7%). Other authors such as [62] observed that the PVP treatment doubled the size of callus of Taxus globosa Schlecht: the callus showed friability and 100% feasibility using fluorescein diacetate (FdA). They also observed that, despite having an antioxidant, the calli of Taxus globosa induced with this medium slightly darkened in the subculture, due to the presence of phenolic compounds.
5. Conclusions
The results show the feasibility of using growth regulators, such as BA, TDZ, and 2,4-D, for the in vitro multiplication and callogenesis of S. compositum. These results do not rule out the possibility that the MS medium may continue to be the base culture medium for the maintenance and propagation of seedlings; however, supplementing it with 0.1 mg L-1 of BA will doubtlessly optimize the multiplication and regeneration of seedlings. For callogenesis from stem explants, the best callus-inducing growth regulator was 2.0 mg L-1 of 2,4-D. The protocol for the induction, maintenance, and multiplication of calli was validated through the induction of calli from the stem in a MS medium supplemented with 2.0 mg L-1 of 2,4-D and resowing after 20 d in a medium with 2.0 mg L-1 of 2,4-D at + 500 mg L-1 of PVP. The success of callus maintenance consists of observing the beginning of oxidation (even in a medium with PVP) and carry out frequent subcultures of the most active parts of the callus, approximately 20 days after sowing or resowing.
Author Contributions
Conceptualization, J.C.-I. and M.d.l.L.R.-F.; methodology, J.C.-I. and M.d.L.A.-G.; validation, L.d.M.R.-P., M.S.H. and I.C.-J.; investigation, M.d.l.L.R.-F., J.C.-I. and C.R.C.M.; resources, L.d.M.R.-P.; writing—original draft preparation, J.C.-I., I.C.-J., and M.d.L.A.-G.; writing—review and editing, J.C.-I. and L.d.M.R.-P. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Data Availability Statement
We encourage all authors of articles published in MDPI journals to share their research data. In this section, please provide details regarding where data supporting reported results can be found, including links to publicly archived datasets analyzed or generated during the study. Where no new data were created, or where data is unavailable due to privacy or ethical restrictions, a statement is still required. Suggested Data Availability Statements are available in section “MDPI Research Data Policies” at https://www.mdpi.com/ethics.
Acknowledgments
To the Sechium edule germplasm bank in Mexico, from Interdisciplinary Research Group on Sechium edule in Mexico, A.C. and Biotechnology Laboratory, CENID-COMEF of INIFAP, Mexico
Conflicts of Interest
The authors declare no conflict of interest.
References
- Casas, A., & Vallejo, M. Agroecología y agrobiodiversidad. Crisis ambiental en México, 2019; 103.
- CDB. Convenio de Diversidad Biológica. Available online: https://www.cbd.int/convention/text/default.shtml.
- Lobo, M., Medina, C.I. Conservación de recursos genéticos de la agrobiodiversidad como apoyo al desarrollo de sistemas de producción sostenibles. Revista Corpoica. Ciencia y Tecnología Agropecuaria, 2009; 10(1): 33-42.
- Cadena-Íñiguez, J., Trejo-Téllez, B. I., Morales-Flores, F. J., & Ruíz-Vera, V. M. Análisis de tratados internacionales relacionados con recursos genéticos y su congruencia con el marco jurídico mexicano. In Proceedings from the 22th International Congress on Project Engineering. Comunicaciones presentadas al XXII Congreso Internacional de Ingeniería de Proyectos, celebrado en Madrid del 11 al 13 de julio de 2018. (p. 97). Asociación española de ingeniería de proyectos (AEIPRO).
- Cadena-Iñiguez, J. El chayote (Sechium edule (Jacq.) Sw., importante recurso fitogenético mesoamericano. Agro Productividad, 2010; 3(2).
- Rodas, Y. C. R., Galarza, M. L. A., Hernández, M. S., Íñiguez, J. C., & Solano, V. M. C. Gestión de un recurso fitogenético para producción de metabolitos secundarios en proyectos de diversificación económica rural. In Comunicaciones presentadas al XXV Congreso Internacional de Ingeniería de Proyectos: celebrado en Alcoy del 6 al 9 de julio de 2021 (p. 101). Asociación española de ingeniería de proyectos (AEIPRO).
- Salas, S. M. P., Aguilar-Galván, F., & Sandoval, L. H. Plantas silvestres comestibles de la Barreta, Querétaro, México y su papel en la cultura alimentaria local. Revista Etnobiología. 2021; Vol, 19(1), 41-62.
- Organización de las Naciones Unidas para la Alimentación y la Agricultura. AGP- Conservación de los Recursos Fitogenéticos. Available online: https://www.fao.org/agriculture/crops/core-themes/theme/seeds-pgr/conservation/en (accessed on 2023).
- Cadena Iñiguez, J., González Santos, R., Cuevas Sánchez, J., Riviello Flores, M. D. L. L., & Ruiz Posadas, L. D. M. La Conservación in situ de la Biodiversidad Agrícola, y la generación de proyectos en ejidos y comunidades de México. In Proceedings from the 22th International Congress on Project Engineering. Comunicaciones presentadas al XXIV Congreso Internacional de Ingeniería de Proyectos, celebrado en Madrid del 07 al 09 de julio de 2020. (p. x). Asociación española de ingeniería de proyectos (AEIPRO).
- González-Santos, R., Cadena-Iñiguez, J., Morales-Flores, F. J., Ruiz-Vera, V. M., Pimentel-López, J., & Peña-Lomelí, A. Model for the conservation and sustainable use of plant genetic resources in México. Wulfenia J, 2015; 22, 333-353.
- Beeching JR, Marmey P, Hughes MA, Charrier A. Evaluation of molecular approaches for determining genetic diversity in Cassava germplasm. Proc. 2nd Internat. Scient. Meet. The Cassava Biotechnology Network. Bogor, Indonesia. 1994; pp. 22-26.
- Organización de las Naciones Unidas para la Alimentación y la Agricultura. El segundo informe sobre el estado de los recursos fitogenéticos para la alimentación y la agricultura en el mundo. Available online: https://www.fao.org/documents/card/es?details=22fabd61-4b41-5bb3-b6be-d38e1516dccc/ (accessed on 23 november 2023 ).
- Vargas-Bermúdez J. D. Conservación In situ de plantas nativas herbáceas, medicinales o bioplaguicidas, como reservorio de diversidad genética y cultural, mariscal sucre-guayas. Doctoral Dissertation, Universidad Agraria Del Ecuador, 2020.
- Barrera-Guzmán, L. A., Cadena-Iñiguez, J., Legaria-Solano, J. P., & Sahagún-Castellanos, J. Phylogenetics of the genus Sechium P. Brown: A review. Spanish Journal of Agricultural Research, 2021; 19(1), e07R01-e07R01.
- Avendaño-Arrazate, C. H., Cadena Iiguez, J., Arevalo Galarza, M. L. C., Campos Rojas, E., Cisneros Solano, V. M., Aguirre Medina, J.F. Las variedades del chayote mexicano, recurso ancestral con potencial de comercialización (No. Libro 635.62 A8). Grupo Interdisciplinario de Investigación en Sechium edule en México (GISeM), 2010.
- GISeM. Rescatando y aprovechando los recursos fitogenéticos de Mesoamérica. Volumen 2: Chayote. Grupo Interdisciplinario de Investigación en Sechium edule en México. Colegio de Postgraduados. Texcoco, Estado de México, 2011; 24p.
- Iñiguez-Luna, M. I., Cadena-Iñiguez, J., Soto-Hernández, R. M., Morales-Flores, F. J., Cortes-Cruz, M., & Watanabe, K. N. Natural bioactive compounds of Sechium spp. for therapeutic and nutraceutical supplements. Frontiers in Plant Science, 2021;12, 772389.
- Iñiguez-Luna, M. I., Cadena-Iñiguez, J., Soto-Hernández, R. M., Morales-Flores, F. J., Cortes-Cruz, M., Watanabe, K. N., ... & Cadena-Zamudio, J. D. Bioprospecting of Sechium spp. varieties for the selection of characters with pharmacological activity. Scientific Reports, 2021; 11(1), 1-12.
- Rivera-Ponce, E. A., Cadena-Iñiguez, J., Cisneros-Solano, V. M., Soto-Hernández, R. M., San Miguel-Chávez, R., García-Osorio, C., & Arévalo-Galarza, M. D. L. Composición fitoquímica y uso potencial del jugo de Sechium compositum (Donn. Sm.) C. Jeffrey. Agro-Divulgación, 2022; 2(4).
- Aguiñiga Sánchez, I. Potencial antileucémico in vitro de extractos de cuatro genotipos de Sechium spp (Cucurbitaceae) (Master's thesis), 2013.
- Gordillo Salinas, L. S. Actividad antifúngica de Sechium compositum contra Botrytis cinerea y Colletotrichum gloeosporioides en condiciones in vitro (Master's thesis), 2019.
- Cruz-Martínez, V., Castellanos-Hernández, O. A., Acevedo-Hernández, G. J., Torres-Morán, M. I., Gutiérrez-Lomelí, M., Ruvalcaba-Ruiz, D., ... & Rodríguez-Sahagún, A. Genetic fidelity assessment in plants of Sechium edule regenerated via organogenesis. South African Journal of Botany, 2017; 112, 118-122.
- Castillo-Martínez C, Cisneros-Solano VM, Hernández-Marini R, Cadena-Iñiguez J, Avendaño-Arrazate CH. Conservación y multiplicación de una colección de Sechium spp. Colegio de Postgraduados y Grupo Interdisciplinario de Investigación Sechium edule en México A.C, México, 2013.
- Cruz-Martínez, V. O., Acevedo-Hernández, G. J., Castellanos-Hernández, O. A., & Rodríguez-Sahagún, A. Regeneración de Sechium edule mediante organogénesis y uso de marcadores ramp para evaluar la fidelidad genética. Biotecnología y Sustentabilidad, 2016; 1(1).
- Wei, Y., Li, Z., Lv, L. et al. Overexpression of MbICE3 increased the tolerance to cold and drought in lettuce (Lactuca sativa L.). In Vitro Cell.Dev.Biol.-Plant. 2023. [CrossRef]
- Romo-Paz, F., Orozco-Flores, J.D., Delgado-Aceves, L. et al. Micropropagation of Physalis angulata L. and P. chenopodifolia Lam. (Solanaceae) via indirect organogenesis. In Vitro Cell.Dev.Biol.-Plant, 2023; 59, 497–506. [CrossRef]
- Alihodzic, A., Davis, J., Roberts, C. et al. Production of secondary metabolites in regenerated Southern wormwood (Artemisia abrotanum L.) under various experimental conditions. In Vitro Cell.Dev.Biol.-Plant, 2023. [CrossRef]
- Behera, S., Kar, S.K., Monalisa, K. et al. Assessment of genetic, biochemical fidelity, and therapeutic activity of in vitro regenerated Hedychium coronarium. In vitro Cell.Dev.Biol.-Plant, 2023. [CrossRef]
- Zhang, W., Dai, W. In vitro plant regeneration of ‘Prelude’ red raspberry (Rubus idaeus L.). In Vitro Cell.Dev.Biol.-Plant, 2023; 59, 461–466. [CrossRef]
- Gaspar, T., Kevers, C., Penel, C., Greppin, H., Reid, D. M., & Thorpe, T. A. Plant hormones and plant growth regulators in plant tissue culture. In vitro Cellular & Developmental Biology-Plant, 1996; 32, 272-289.
- Kaur, K., Dolker, D., Behera, S., & Pati, P. K. Critical factors influencing in vitro propagation and modulation of important secondary metabolites in Withania somnifera (L.) dunal. Plant Cell, Tissue and Organ Culture (PCTOC), 2022; 149(1-2), 41-60.
- Alvarenga, S., & Morera, J. In vitro micropropagation of the chayote (Sechium edule Jacq. Sw.). Tecnologia en Marcha (Costa Rica), 1992; vol. 11, no 3.
- Abdelnour, A., Ramírez, C., & Engelmann, F. Micropropagación de chayote (Sechium edule Jacq. SW.) a partir de brotes vegetativos. Agronomía mesoamericana, 2002; 13(2), 147-151.
- Abdelnour-Esquivel, A., Bermudez, L. C., Rivera, C., & Alvarenga-Venutolo, S. Cultivo de meristemas, termo y quimioterapia en chayote (Sechium edule Jacq. Sw.) para la erradicación del virus del mosaico del chayote (ChMV). Manejo Integrado de Plagas y Agroecología Número 77 (Abril 2006), 2006.
- Del Ángel, O., Vela, G., Rodríguez, A., Gómez, M., & García, H. (2014). Desinfección y regeneración eficiente de chayote in vitro (Sechium edule Jacq. Sw.). In Ciencias Agropecuarias Handbook T-II: Congreso Interdisciplinario de Cuerpos Académicos, ECORFAN, 2014; p. 23.
- Mora, D. F., Esquivel, A. A., & Venutolo, S. A. Micropropagación de fenotipos seleccionados de chayote. Tecnología en marcha, 1999; 13(3), 9-15.
- Abdelnour-Esquivel, A., Brenes-Madriz, J., & Alvarenga-Venutolo, S. Guía Técnica Semilla de Chayote. 2015; [PDF] Available online: chrome-extension://efaidnbmnnnibpcajpcglclefindmkaj. Available online: https://repositoriotec.tec.ac.cr/bitstream/handle/2238/6768/gu%C3%ADa-tecnica-semillachayote-14%20SET-15.pdf?sequence=1.
- García, J. G., Alvarado, E. S., & Bolaños, J. A. Efecto del AIA y el AIB sobre el enraizamiento in vitro de brotes de Sechium edule (Jacq.) Sw. Biotecnología Vegetal, 2015; 15(1).
- Soto-Contreras, A., Núñez-Pastrana, R., Rodríguez-Deméneghi, M. V., Aguilar-Rivera, N., Galindo-Tovar, M. E., & Ramírez-Mosqueda, M. A. Indirect organogenesis of Sechium edule (Jacq.) Swartz. In Vitro Cellular & Developmental Biology-Plant, 2022; 58(6), 903-910.
- Da Silva, R. F. Sistema de regeneración in vitro por embriogénesis somática indirecta en variedades venezolanas de arroz (Oryza sativa L.). Revista Científica UDO Agrícola, 2012; 12(1), 55-68.
- Riviello-Cogco, E., Robledo-Paz, A., Gutiérrez-Espinosa, M. A., Suárez-Espinosa, J., & Mascorro-Gallardo, J. O. Maduración y germinación de embriones somáticos de Coffea arabica cv. colombia. Revista Fitotecnia Mexicana, 2021; 44(2), 161-161.
- Classic Murashige, T., & Skoog, F. A revised medium for rapid growth and bioassays with tobacco tissue cultures. Physiol. Plant, 1962; 15, 473-497.
- Cadena Iñiguez, J., Soto Hernández, M., Arévalo Galarza, M., Avendaño Arrazate, C. H., Aguirre Medina, J. F., & Ruiz Posadas, L. D. M. Caracterización bioquímica de variedades domesticadas de chayote Sechium edule (Jacq.) Sw. comparadas con parientes silvestres. Revista Chapingo. Serie Horticultura, 2011; 17(SPE2), 45-55.
- Ramos-Parra, M., Ulín-Montejo, F., Aguilar-Nieto, J. A., Solis-Trapala, I. L., & Fierro-Carbajal, J. B. Modelación y estimación del volumen de tejido vegetal in vitro de Strombocactus disciformis basada en mediciones no intrusivas. Universidad y ciencia, 2010; 26(2), 195-203.
- Dwass, C. Some k-sample rank-order test. Contributions to probability and Statistics. 1960; Pp 198-202.
- Steel, R. A rank sum test for comparing all pairs of treatments. Technometrics, 1960; Vol. 2, No. 2 pp 197-207.
- Critchlow, D. y Fligner, M. On distribution-free multiple comparisons in the one-way analysis of variance. Commun. Statist.- Theory meth., 1991; Vol. 20, No. 1 pp 127-139.
- Thilagam, D., Kumudini, B. S., & Manohar, S. H. Regeneration of Sechium edule (jacq) sw. From sterile in vitro nodal explants and assessment of clonalfidelity using issr and rapd markers. International Journal of Agricultural, 2016; Vol. 6, pag. 285-292.
- Larson, C. G., Hasbún, R., Jofré, M. P., Sánchez-Olate, M., & Ríos, D. Efecto del genotipo y fuente de citoquinina en la etapa de iniciación de cultivo in vitro de tejido adulto de Castanea sativa Mill. Gayana. Botánica, 2017; 74(1), 30-40.
- Meier-Dinkel, A., Becker, B., Duckstein, D. Micropropagation and ex vitro rooting of several clones of late-flushing Quercus robur L. Annales des Sciences Forestières, 1993; 50(1): 319-322.
- Castillo-Martínez, C. R., Gutiérrez-Espinosa, M., Buenrostro-Nava, M. T., Cetina Alcalá, V. M., & Cadena Iñiguez, J. Regeneración de plantas de Paulownia elongata Steud: Por organogénesis directa. Revista mexicana de ciencias forestales, 2012; 3(10), 41-49.
- Gil C, A. I., Ariza C, C. A., Castillo T, L. M., Salgado D, L. E., Banda Sánchez, L., & Vanegas M, L. E. Induction of organogenesis in vitro with 6-benzylaminopurine in Cattleya trianae Linden & Rchb. f. Revista UDCA Actualidad & Divulgación Científica, 2019; 22(2).
- Mora-Cruz, Y., López-Peralta, M. C. G., Hernández-Meneses, E., & Cruz-Huerta, N. Regeneración in vitro de plantas de prosthechea vitellina (lindley) we higging por organogenesis directa. Revista Fitotecnia Mexicana, 2023; 46(1), 33-33.
- Hussain, A., Ahmed Qarshi, I., Nazir, H. & Ullah, I. Plant tissue culture: current status and opportunities. Recent advances in plant in vitro culture, 2012, vol. 6, no 10, p. 1-28. [CrossRef]
- Chen, X., Ye, C., Yang, H., Ji, W., Xu, Z., Ye, S., ... & Zhu, X. Callogenesis and plant regeneration in peony (Paeonia× suffruticosa) using flower petal explants. Horticulturae, 2022; 8(5), 357.
- Hernández-Amasifuen, A. D., Cortez-Lázaro, A. A., Argüelles-Curaca, A., & Díaz-Pillasca, H. B. Callogénesis in vitro de durazno (Prunus persica L.) var. Huayco rojo a partir de explantes foliares. Ciencia y Tecnología Agropecuaria, 2022; 23(1), e2032-e2032.
- Sidek, N., Nulit, R., Kong, Y. C., Yien, C. Y. S., Sekeli, R., & EL-Barghathi, M. F. (2022). Callogenesis and somatic embryogenesis of Oryza sativa L.(cv. MARDI Siraj 297) under the influence of 2, 4-dichlorophenoxyacetic acid and kinetin. AIMS Agriculture and Food, 2022.
- Zayova, E., Nedev, T., & Stancheva, I. Callus via shoot organogenesis and plant regeneration of Stevia rebaudiana Bertoni. In Proceedings of the Bulgarian Academy of Sciences, 2022; Vol. 75, No. 4, pp. 620-628.
- Da Silva, R. F. Sistema de regeneración in vitro por embriogénesis somática indirecta en variedades venezolanas de arroz (Oryza sativa L.). Revista Científica UDO Agrícola, 2012; 12(1), 55-68.
- Phillips, G. C., & Garda, M. Plant tissue culture media and practices: an overview. In Vitro Cellular & Developmental Biology-Plant, 2019; 55, 242-257.
- Domínguez-Perales, L. A., Domínguez-Álvarez, J. L., Cruz-Izquierdo, S., Santacruz-Varela, A., Barrientos-Priego, A., Padilla-Ramírez, J. S., & Gutiérrez-Espinosa, M. A. Propagación in vitro de selecciones de guayabo (Psidium guajava L.). Revista fitotecnia mexicana, 2016; 39(3), 285-295.
- Cureño, H. B., Hernadez, R. S., valdivia, A. R., Télle, L. T., Martínez, M., Vazquez, M. E., ... & Uptón, J. L. Extracción y cuantificación de taxoides por HPLC en hojas in situ y en callos inducidos in vitro de Taxus globosa Schlecht. III Jornadas Internacionales sobre el Tejo (Taxus baccata L.), 2011.
Figure 1.
Effect of six concentrations of thidiazuron (TDZ) and 6-benzylaminopurine (BA), compared to the MS control, in which the presence of callus and root in seedlings of Sechium compositum was evaluated. Means ± SE (standard error). Where the treatments are: 1: MS control, 2: MS+BA (0.1 mg L-1), 3: MS+BA (0.2 mg L-1), 4: MS+BA (0.4 mg L-1), 5: MS+BA (0.6 mg L-1), 6: MS+BA (0.8 mg L-1), 7: MS+BA (1.0 mg L-1), 8: MS+BA (1.2 mg L-1), 9: MS+TDZ (0.1mg L-1), 10: MS+TDZ (0.2mg L-1), 11: MS+TDZ (0.4 mg L-1), 12: MS+TDZ (0.6mg L-1), 13: MS+TDZ (0.8mg L-1), 14: MS+TDZ (1.0 mg L-1), 15: MS+TDZ (1.2 mg L-1). Kruskal-Wallis One Way Analysis of Variance on Ranks (P = <0.001); Mann-Whitney U Test (P= <0.001). * Differences between control and treatments (P < 0.05).
Figure 1.
Effect of six concentrations of thidiazuron (TDZ) and 6-benzylaminopurine (BA), compared to the MS control, in which the presence of callus and root in seedlings of Sechium compositum was evaluated. Means ± SE (standard error). Where the treatments are: 1: MS control, 2: MS+BA (0.1 mg L-1), 3: MS+BA (0.2 mg L-1), 4: MS+BA (0.4 mg L-1), 5: MS+BA (0.6 mg L-1), 6: MS+BA (0.8 mg L-1), 7: MS+BA (1.0 mg L-1), 8: MS+BA (1.2 mg L-1), 9: MS+TDZ (0.1mg L-1), 10: MS+TDZ (0.2mg L-1), 11: MS+TDZ (0.4 mg L-1), 12: MS+TDZ (0.6mg L-1), 13: MS+TDZ (0.8mg L-1), 14: MS+TDZ (1.0 mg L-1), 15: MS+TDZ (1.2 mg L-1). Kruskal-Wallis One Way Analysis of Variance on Ranks (P = <0.001); Mann-Whitney U Test (P= <0.001). * Differences between control and treatments (P < 0.05).
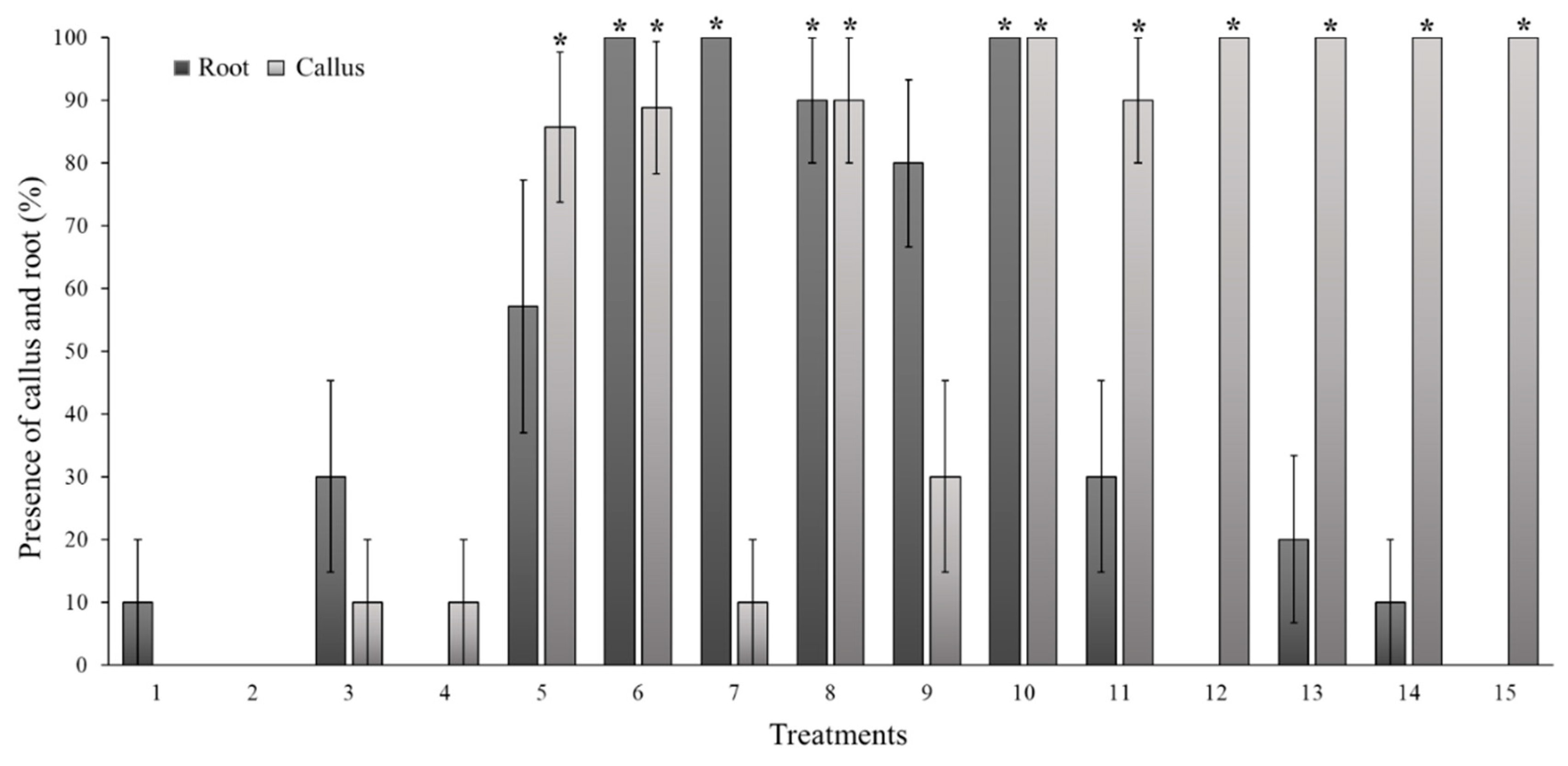
Figure 2.
Effect of 0.1 mg L-1 of BA, compared to MS control. Average seedling height, average number of shoots, buds, percentage of root, callus, and rosette structure of the in vitro seedlings of Sechium compositum. Means ± SE (Standard Error) of n=50 repetitions per treatment. Kruskal-Wallis One Way Analysis of Variance on Ranks (P = <0.001); Mann-Whitney U Test (P= <0.001). * Differences between control and treatments (P < 0.05).
Figure 2.
Effect of 0.1 mg L-1 of BA, compared to MS control. Average seedling height, average number of shoots, buds, percentage of root, callus, and rosette structure of the in vitro seedlings of Sechium compositum. Means ± SE (Standard Error) of n=50 repetitions per treatment. Kruskal-Wallis One Way Analysis of Variance on Ranks (P = <0.001); Mann-Whitney U Test (P= <0.001). * Differences between control and treatments (P < 0.05).
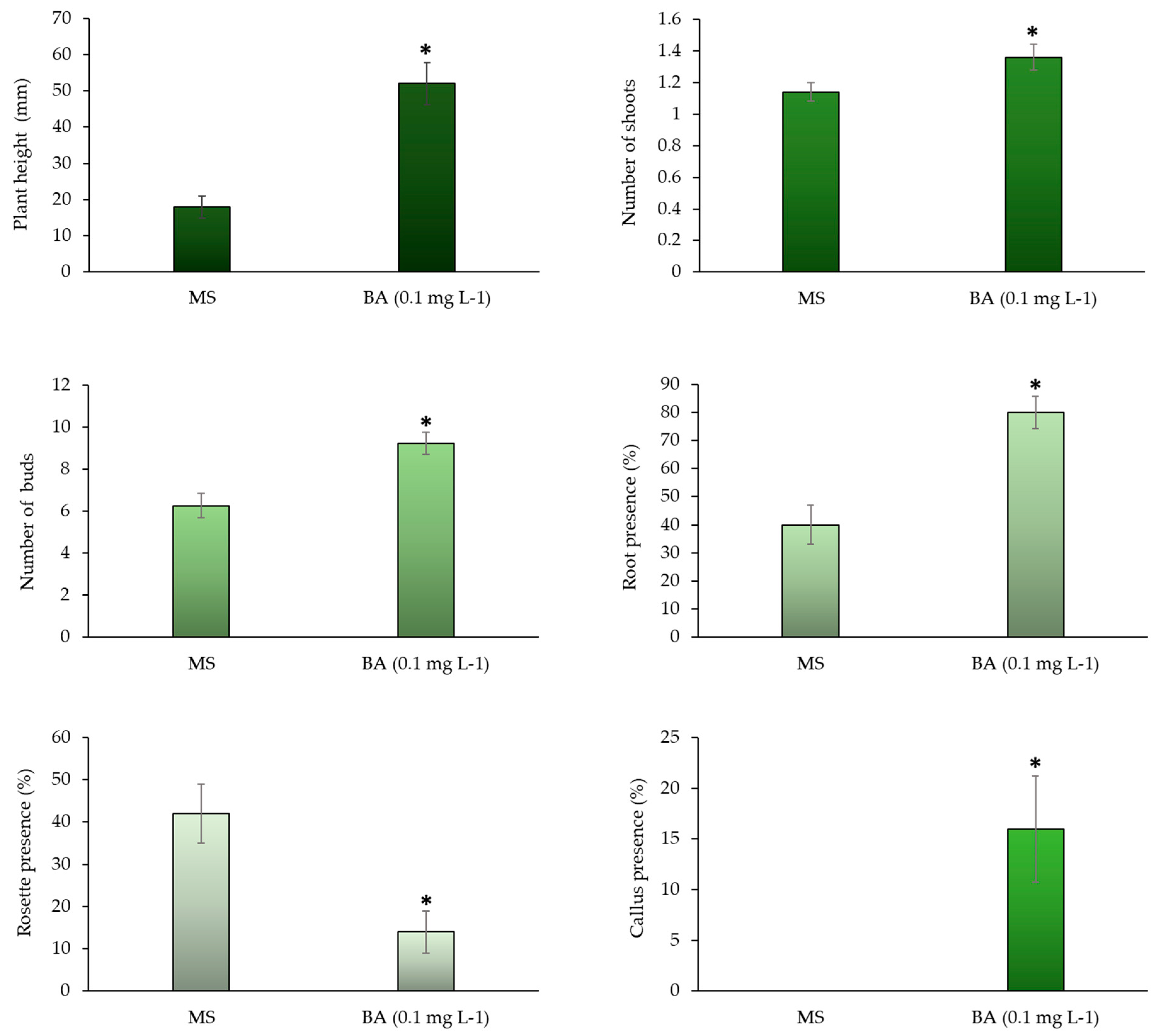
Figure 3.
Sechium compositum established in vitro: A) +2-months old seedling. B) +2-months old seedling with two shoots and formation of basal callus. C) Seedling with packed buds (rosette structure).
Figure 3.
Sechium compositum established in vitro: A) +2-months old seedling. B) +2-months old seedling with two shoots and formation of basal callus. C) Seedling with packed buds (rosette structure).

Figure 4.
Effect of three concentrations of thidiazuron (TDZ) and 2,4-dichlorophenoxyacetic acid (2,4-D) on callus generation, from leaf (A) and stem (B) explants of Sechium compositum. The following treatments were used: 1.- MS control-stem, 2.- Stem MS+2,4-D (0.5 mg L-1), 3.- Stem MS+2,4-D (1.0 mg L-1), 4.- Stem MS+2,4-D (2.0 mg L-1), 5.- Stem MS+TDZ (0.5 mg L-1), 6.- Stem MS+TDZ (1.0 mg L-1), 7.- Stem MS+TDZ )(2.0 mg L-1, 8.- MS control-leaf, 9.- Leaf MS+2,4-D (0.5 mg L-1), 10.- Leaf MS+2,4-D (1.0 mg L-1), 11.- Leaf MS+2,4-D (2.0 mg L-1), 12.- Leaf MS+TDZ (0.5 mg L-1), 13.- Leaf MS+TDZ (1.0 mg L-1), 14.- Leaf MS+TDZ (2.0 mg L-1). Means ± Standard Error (SE). Comparative analysis Mann-Whitney U Test (P = <0.001). *Differences between the control and the treatments.
Figure 4.
Effect of three concentrations of thidiazuron (TDZ) and 2,4-dichlorophenoxyacetic acid (2,4-D) on callus generation, from leaf (A) and stem (B) explants of Sechium compositum. The following treatments were used: 1.- MS control-stem, 2.- Stem MS+2,4-D (0.5 mg L-1), 3.- Stem MS+2,4-D (1.0 mg L-1), 4.- Stem MS+2,4-D (2.0 mg L-1), 5.- Stem MS+TDZ (0.5 mg L-1), 6.- Stem MS+TDZ (1.0 mg L-1), 7.- Stem MS+TDZ )(2.0 mg L-1, 8.- MS control-leaf, 9.- Leaf MS+2,4-D (0.5 mg L-1), 10.- Leaf MS+2,4-D (1.0 mg L-1), 11.- Leaf MS+2,4-D (2.0 mg L-1), 12.- Leaf MS+TDZ (0.5 mg L-1), 13.- Leaf MS+TDZ (1.0 mg L-1), 14.- Leaf MS+TDZ (2.0 mg L-1). Means ± Standard Error (SE). Comparative analysis Mann-Whitney U Test (P = <0.001). *Differences between the control and the treatments.
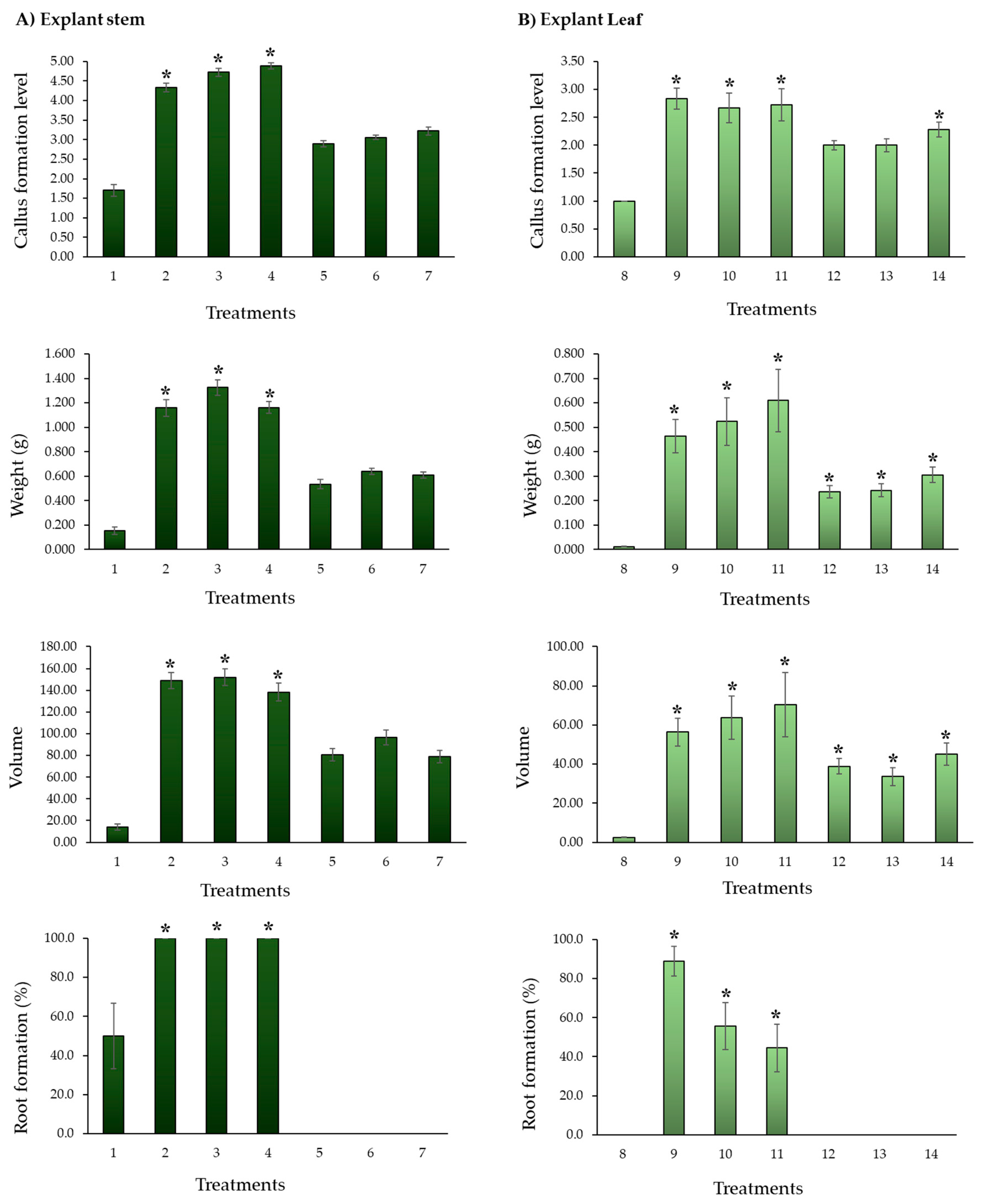
Figure 5.
Callogenesis in Sechium compositum. Oxidized callus obtained from leaf explant: A, B, C, F, G, H. Callus obtained from stem explant: D, E, I, J. All observed through optical microscope.
Figure 5.
Callogenesis in Sechium compositum. Oxidized callus obtained from leaf explant: A, B, C, F, G, H. Callus obtained from stem explant: D, E, I, J. All observed through optical microscope.
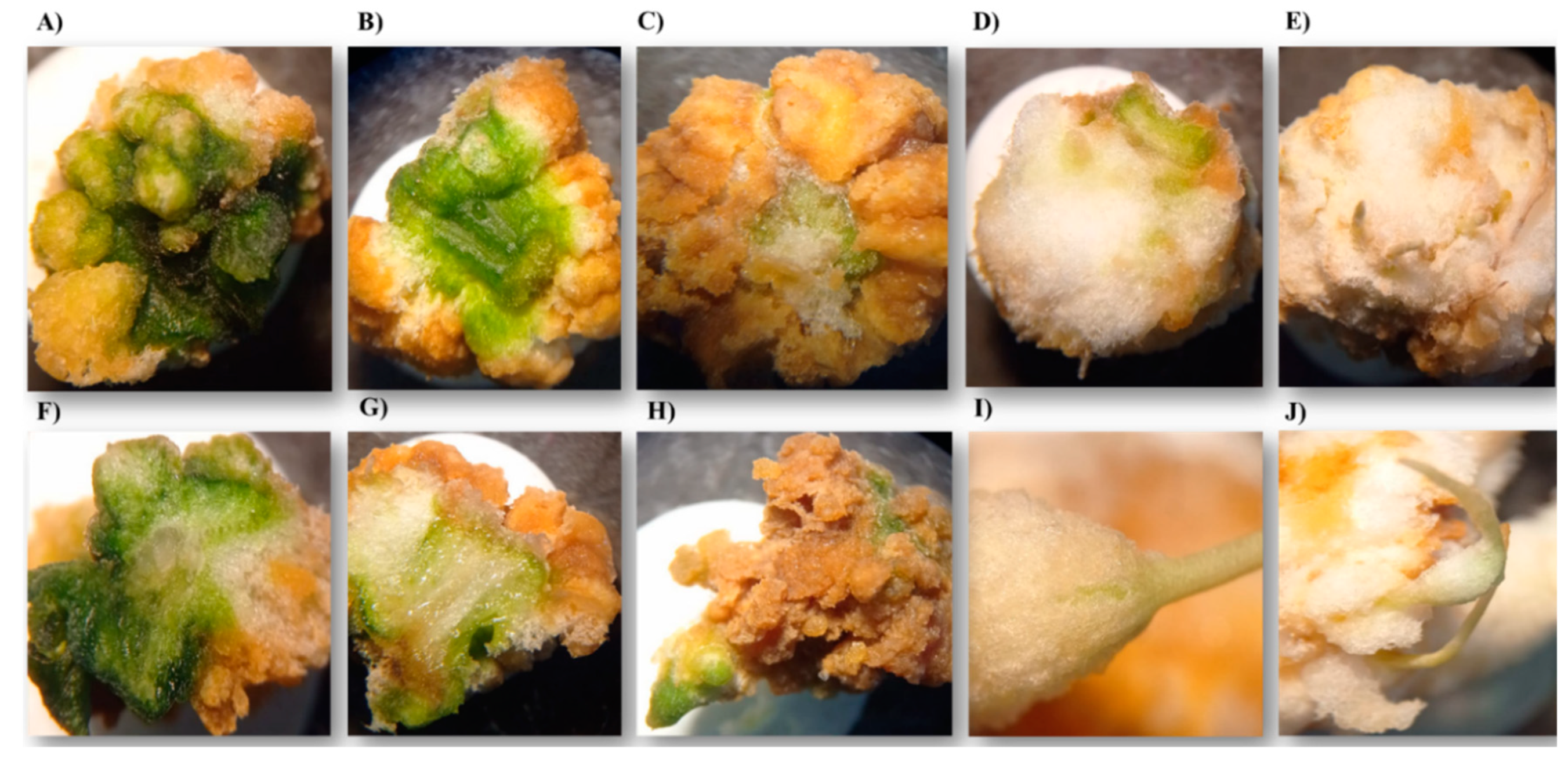
Figure 6.
Calli obtained from stem of Sechium compositum sown in medium (2.0 mg L-1 of 2,4-D) with 2.0 mg L-1 of activated carbon. Where: A) Stem explant for callus formation observed 7 days after sowing (callus formation begins at the ends of the stem where the cut was made); B) Callus observed 15 days after sowing (white callus begins to develop on the surface of the stem); C) Callus observed 30 days after sowing (most of the calli formed roots roots and their development stopped); D) Single seedling regenerated at 30 d, in the medium with activated carbon from stem explant.
Figure 6.
Calli obtained from stem of Sechium compositum sown in medium (2.0 mg L-1 of 2,4-D) with 2.0 mg L-1 of activated carbon. Where: A) Stem explant for callus formation observed 7 days after sowing (callus formation begins at the ends of the stem where the cut was made); B) Callus observed 15 days after sowing (white callus begins to develop on the surface of the stem); C) Callus observed 30 days after sowing (most of the calli formed roots roots and their development stopped); D) Single seedling regenerated at 30 d, in the medium with activated carbon from stem explant.

Figure 7.
Calli obtained from stem of Sechium compositum. Where: A) Callus with desired characteristics (white, with few yellow areas); B) Callus culture was divided for multiplication and resown in medium with 2.0 mg L-1 of 2,4-D; C) Calli resown in a medium with 2.0 mg L-1 of 2,4-D with PVP; D) Oxidized callus.
Figure 7.
Calli obtained from stem of Sechium compositum. Where: A) Callus with desired characteristics (white, with few yellow areas); B) Callus culture was divided for multiplication and resown in medium with 2.0 mg L-1 of 2,4-D; C) Calli resown in a medium with 2.0 mg L-1 of 2,4-D with PVP; D) Oxidized callus.
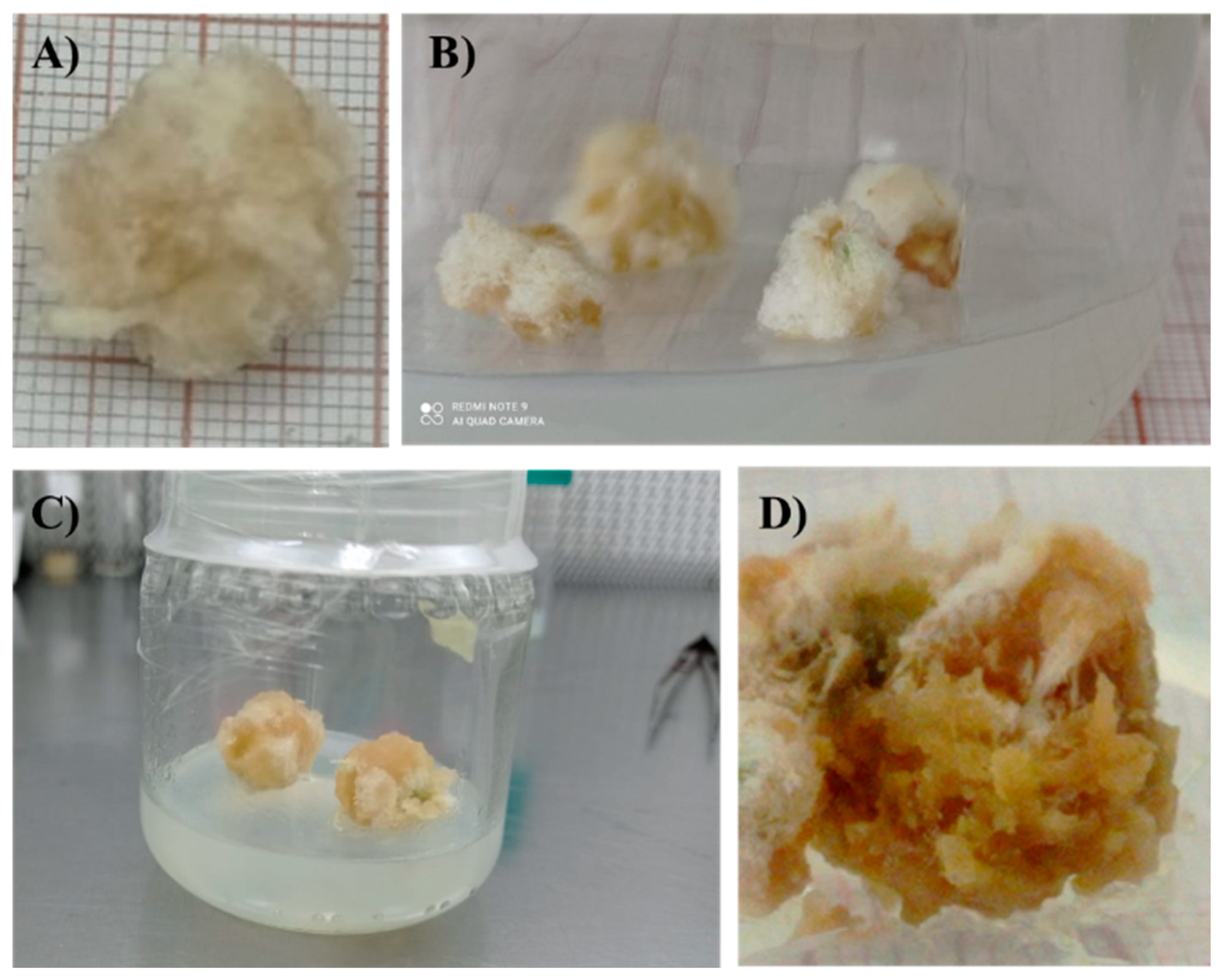
Figure 8.
Effect of adding two concentrations of polyvinylpyrrolidone (PVP) to the medium with 2.0 mg L-1 of 2,4-D for the induction of callus from stem of Sechium compositum. Callus development level at 20 and 30 days. Means ± Standard Error (SE). Comparative analysis with Mann-Whitney U Test (P = <0.001). *Differences between the control and the treatments.
Figure 8.
Effect of adding two concentrations of polyvinylpyrrolidone (PVP) to the medium with 2.0 mg L-1 of 2,4-D for the induction of callus from stem of Sechium compositum. Callus development level at 20 and 30 days. Means ± Standard Error (SE). Comparative analysis with Mann-Whitney U Test (P = <0.001). *Differences between the control and the treatments.
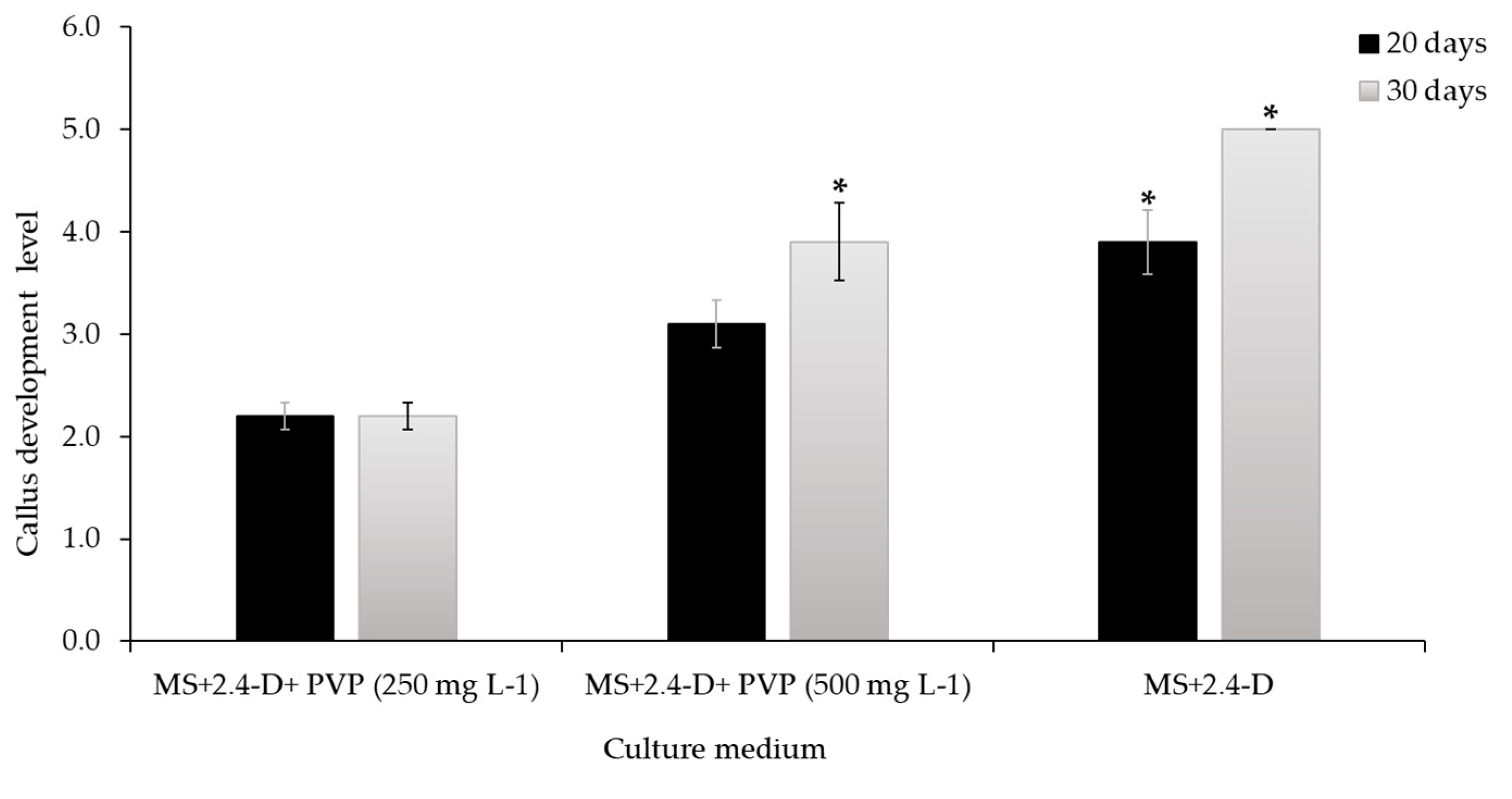
Figure 9.
Effect of the addition of two concentrations of polyvinylpyrrolidone (PVP) to the medium with 2.0 mg L-1 of 2,4-D on the growth of sub cultured calli with desirable characteristics in Sechium compositum (reduction in oxidation, survival, and mass growth); evaluated at 20 and 30 days. Means ± Standard Error (SE). Kruskal-Wallis One Way Analysis of Variance on Ranks (P = >0.05). There are no differences between the treatments.
Figure 9.
Effect of the addition of two concentrations of polyvinylpyrrolidone (PVP) to the medium with 2.0 mg L-1 of 2,4-D on the growth of sub cultured calli with desirable characteristics in Sechium compositum (reduction in oxidation, survival, and mass growth); evaluated at 20 and 30 days. Means ± Standard Error (SE). Kruskal-Wallis One Way Analysis of Variance on Ranks (P = >0.05). There are no differences between the treatments.
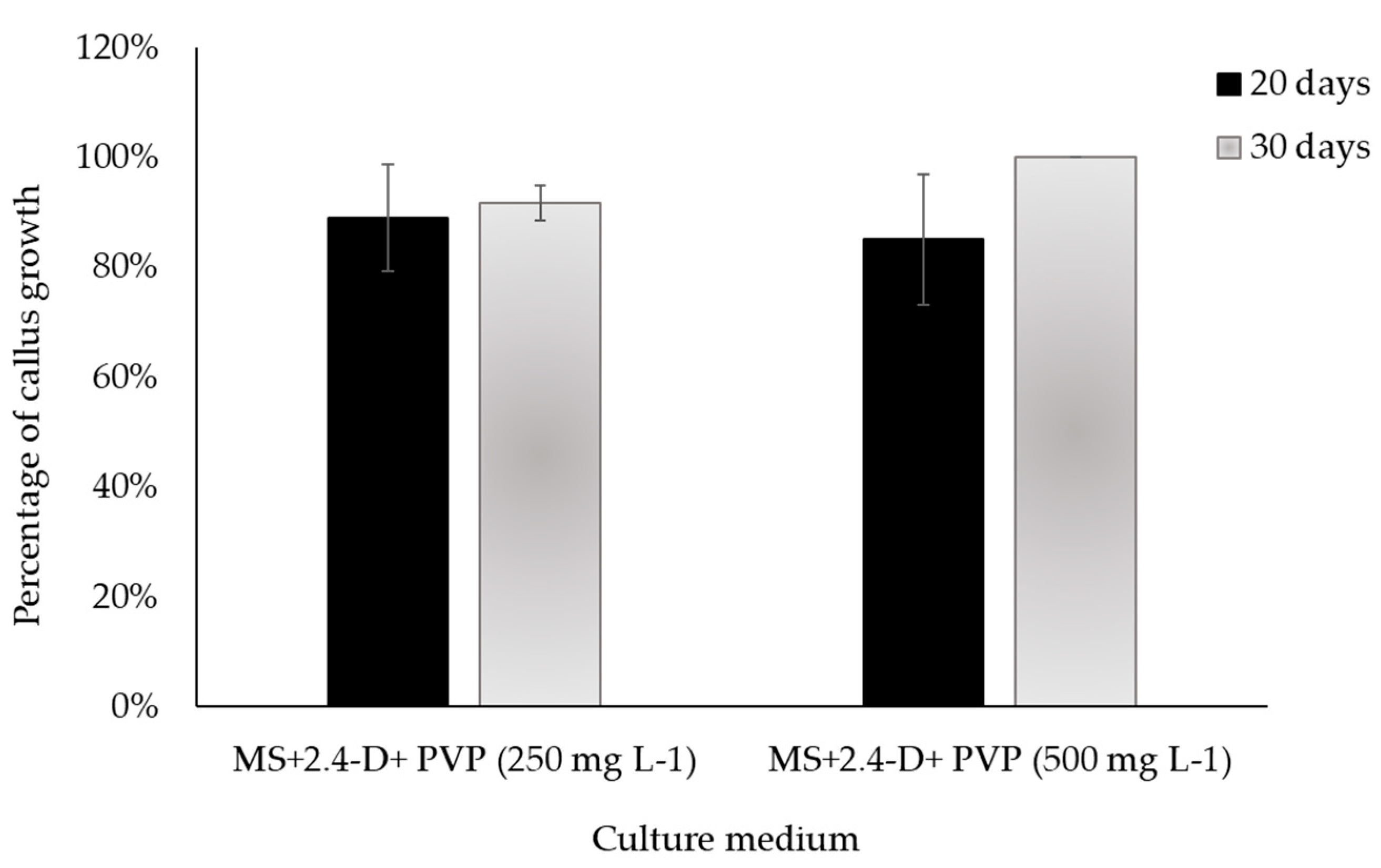

Table 1.
Five-level scale used to determine callus formation from stem and leaf explants of Sechium compositum.
Table 1.
Five-level scale used to determine callus formation from stem and leaf explants of Sechium compositum.
| Level | Callus formation scale (%) | Description |
| 1 | 0 | There is no tissue response. |
| 2 | 1-25 | The tissue swells (turgor) and begins to form a light-yellow callus at the ends. |
| 3 | 26-50 | The ends surrounding tissue areas show a greater amount of white callus. |
| 4 | 52-75 | A green tissue portion is observed at the top. The rest of the callus is white. |
| 5 | 76-100 | The callus has completely covered the tissue and there is an increase in the white mass, with a slight brown tone in small areas. |
Table 2.
Scale used to determine the percentage of callus for multiplication.
| Callus development (%) | Description |
| 0 | Brown callus and yellow medium are observed. |
| 25 | The callus maintains a greater number of brown areas and the mass does not increase. The medium looks slightly yellow. |
| 50 | A greater percentage of potentially active callus is observed, and the medium turns a light yellow. |
| 75 | A considerable decrease in brown areas is observed, along with has a greater number of active areas in the callus, and a transparent medium. |
| 100 | The callus presents mostly or all active zones, its mass increases, and root formation is observed. The medium is transparent. |
Table 3.
Effect of seven concentrations of thidiazuron (TDZ) and 6-benzylaminopurine (BA) on height and proliferation of buds and shoots of Sechium compositum.
Table 3.
Effect of seven concentrations of thidiazuron (TDZ) and 6-benzylaminopurine (BA) on height and proliferation of buds and shoots of Sechium compositum.
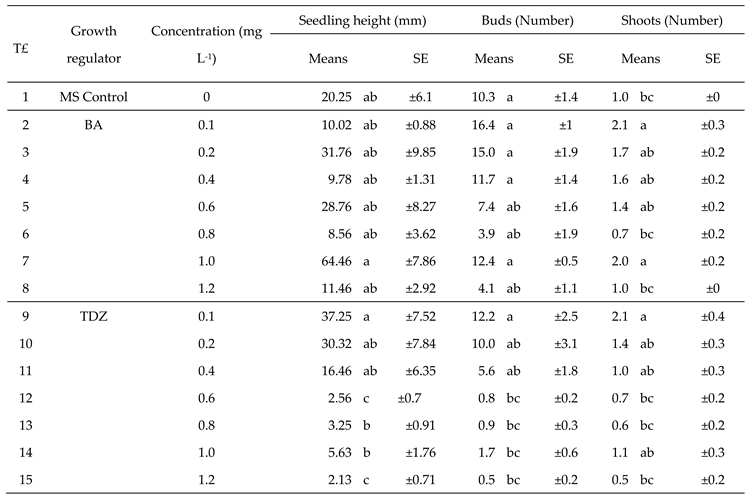 |
Means ± SE (standard error). Kruskal-Wallis Analysis Pr > Chisq < 0.0001. Means with the same letter are not statistically different according to the Wilcoxon matched pairs signed rank test versus the Steel-Dwass-Critchlow-Fligner method (P < 0.05). T£ = Treatments.
Table 4.
Effect of three concentrations of thidiazuron (TDZ) and 2,4-dichlorophenoxyacetic acid (2,4-D) on the generation of callus, from stem and leaf explants of Sechium compositum.
Table 4.
Effect of three concentrations of thidiazuron (TDZ) and 2,4-dichlorophenoxyacetic acid (2,4-D) on the generation of callus, from stem and leaf explants of Sechium compositum.
| T | E | GR |
[GRC] (mg L-1) |
Callus Formation level | Weight (g) | Ø 1 | Ø 2 | Height | Volume | Root formation (%) | ||||||||||||||
| X̅ | SE | X̅ | SE | X̅ | SE | X̅ | SE | X̅ | SE | X̅ | SE | X̅ | SE | |||||||||||
| 1 | Stem | MS | 1.70 | e | ±0.15 | 0.153 | c | ±0.029 | 7.15 | de | ±0.44 | 5.70 | cd | ±0.51 | 4.75 | c | ±0.51 | 14.06 | ce | ±2.75 | 50.0 | abc | ±16.7 | |
| 2 | 2,4-D | 0.5 | 4.33 | b | ±0.11 | 1.160 | a | ±0.068 | 16.26 | a | ±0.30 | 13.48 | a | ±0.51 | 11.73 | a | ±0.31 | 149.11 | a | ±7.31 | 100.0 | a | ±0.0 | |
| 3 | 1.0 | 4.72 | ab | ±0.11 | 1.326 | a | ±0.064 | 16.51 | a | ±0.30 | 12.95 | a | ±0.31 | 11.69 | a | ±0.28 | 152.05 | a | ±7.66 | 100.0 | a | ±0.0 | ||
| 4 | 2.0 | 4.89 | a | ±0.08 | 1.161 | a | ±0.048 | 16.08 | a | ±0.40 | 12.55 | a | ±0.25 | 11.12 | a | ±0.26 | 138.28 | a | ±8.14 | 100.0 | a | ±0.0 | ||
| 5 | TDZ | 0.5 | 2.89 | c | ±0.08 | 0.534 | b | ±0.039 | 13.07 | b | ±0.43 | 10.68 | b | ±0.40 | 9.64 | b | ±0.31 | 80.82 | b | ±5.68 | 0.0 | ce | ±0.0 | |
| 6 | 1.0 | 3.06 | c | ±0.06 | 0.638 | b | ±0.026 | 13.79 | b | ±0.30 | 10.08 | b | ±0.28 | 10.54 | b | ±0.37 | 96.43 | b | ±6.69 | 0.0 | ce | ±0.0 | ||
| 7 | 2.0 | 3.22 | c | ±0.10 | 0.607 | b | ±0.026 | 13.12 | b | ±0.36 | 10.40 | b | ±0.41 | 9.41 | b | ±0.31 | 79.01 | b | ±5.61 | 0.0 | ce | ±0.0 | ||
| 8 | Sheet | MS | 1.00 | e | ±0.00 | 0.012 | d | ±0.001 | 5.72 | e | ±0.05 | 4.61 | d | ±0.39 | 0.92 | d | ±0.11 | 2.45 | f | ±0.17 | 0.0 | cde | ±0.0 | |
| 9 | 2,4-D | 0.5 | 2.83 | cd | ±0.19 | 0.464 | bc | ±0.068 | 11.48 | bc | ±0.48 | 9.04 | b | ±0.51 | 8.12 | bc | ±0.55 | 56.37 | bcd | ±7.05 | 88.9 | ab | ±7.6 | |
| 10 | 1.0 | 2.67 | cde | ±0.27 | 0.523 | bc | ±0.098 | 12.07 | b | ±0.96 | 8.93 | b | ±0.74 | 6.78 | bc | ±0.78 | 63.73 | bc | ±11.11 | 55.6 | ab | ±12.1 | ||
| 11 | 2.0 | 2.72 | cde | ±0.29 | 0.610 | bc | ±0.128 | 11.61 | b | ±1.00 | 8.81 | b | ±0.91 | 7.52 | bc | ±1.02 | 70.47 | bc | ±16.39 | 44.4 | bcd | ±12.1 | ||
| 12 | TDZ | 0.5 | 2.00 | e | ±0.08 | 0.236 | c | ±0.024 | 10.17 | cd | ±0.51 | 7.83 | c | ±0.40 | 7.21 | c | ±0.37 | 38.82 | cde | ±3.93 | 0.0 | ce | ±0.0 | |
| 13 | 1.0 | 2.00 | de | ±0.11 | 0.241 | c | ±0.027 | 9.77 | cd | ±0.59 | 6.99 | c | ±0.39 | 6.42 | c | ±0.47 | 33.58 | cde | ±4.65 | 0.0 | ce | ±0.0 | ||
| 14 | 2.0 | 2.28 | de | ±0.14 | 0.305 | c | ±0.032 | 11.39 | c | ±0.54 | 8.06 | c | ±0.29 | 6.64 | c | ±0.45 | 45.15 | cd | ±5.73 | 0.0 | ce | ±0.0 | ||
Means (X̅) ± Standard Error (SE). Kruskal-Wallis Analysis Pr > Chisq <0.0001. Means with the same letter are not statistically different according to Wilcoxon matched pairs signed rank test versus the Steel-Dwass-Critchlow-Fligner Method (P < 0.05). Treatment (T), Explant (E), Growth Regulator (GR), Growth Regulator Concentration [GRC], Diameter (Ø).
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions, or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



