
Submitted:
20 December 2023
Posted:
21 December 2023
You are already at the latest version
Abstract
Understanding the impact of drought stress on Arabica coffee physiology and metabolism is essential in the pursuit of developing drought-resistant varieties. In this study, we explored the physiological and metabolite changes in coffee genotypes exhibiting varying degrees of tolerance to drought—namely, the relatively tolerant Ca74110 and Ca74112, and the sensitive Ca754 and CaJ-19—under well-watered conditions and during terminal drought stress periods over two-time intervals (0 and 60 days following the onset of stress). The Metabolites profiling conducted uncovered significant associations between growth and the physiological characteristics of coffee genotypes with distinct drought tolerance behaviors. Initially, no marked differences were observed among genotypes or treatments. However, at the 60-day post-drought onset, notable increases in shoot growth, biomass, CO2 assimilation, pigments, and various physiological parameters were evident, particularly in the relatively tolerant genotypes. Metabolite profiling revealed elevations in glucose, maltose, amino acids, organic acids, and decreases in other metabolites, more pronounced in drought-tolerant genotypes. These alterations were more pronounced in the drought-tolerant genotypes, indicating a correlation with enhanced compatible solutes and energy-associated metabolites crucial for drought tolerance mechanisms. This research introduces GC-MS based metabolome profiling to Ethiopian coffee, shedding light on the intricate responses to drought stress and paving the way for the potential development of drought-resistant coffee seedlings in intensified agro-ecological zones.
Keywords:
drought stress
; coffea arabica
; growth
; gas exchanges
; metabolites
; network analysis
1. Introduction
Coffee stands as a vital global agricultural commodity, trailing only oil in importance. Its production in tropical and subtropical regions sustains millions of livelihoods [1,2]. Coffea arabica L. accounts for over 70% of the world’s coffee and is famed for its excellence [3]. Brazil leading Arabica coffee production at 44%, with Ethiopia contributing 5% [4]. Ethiopia, a top Arabica coffee producer in Africa, exports about 471,000 tons yearly, yielding 0.71 tons per hectare [5,6,7]. The looming specter of global climate change, however, threatens C. arabica cultivation, with water scarcity and drought. This poses significant challenges to coffee cultivation, disrupting suitable regions, yield, and quality, and inviting pests and diseases, causing economic losses [8,9].
Plants instinctively adjust catabolic and anabolic systems during drought, altering metabolic pathways to protect against damage [10,11]. This adaptation’s mechanism hinges on species, genotype, and stress intensity [12,13]. Metabolic adjustment involves accumulating compatible solutes, influencing pathways like sugar synthesis, photosynthesis, and more [14]. Certain metabolites increase during drought stress, like proline, serine, valine, and betaine, fostering tolerance [15,16], while others decrease, like Myo-inositol and glutamate [17]. Metabolite accumulation helps cell turgor maintenance, osmotic potential reduction, and oxidative damage protection [18]. Stress-resilient plants often sustain higher stress-related metabolite levels even under normal conditions [13,19]. Despite the importance of metabolomic components in drought tolerance, insights into C. arabica’s metabolomics under drought remain limited [20]. Understanding drought stress adaptation necessitates studying metabolomic responses in drought-tolerant and sensitive genotypes [21].
Metabolomic analysis, utilizing techniques like gas/liquid chromatography–mass spectrometry and nuclear magnetic resonance, reveals the level of small-molecule metabolites precisely [22,23,24]. Nevertheless, understanding coffee’s intricate genetic and molecular aspects remains a significant hurdle [2]. Recent focus on coffee has illuminated metabolite responses to drought [2], temperature [19], elevated CO2 [21], and water logging [25]. However, previous research in Ethiopia has predominantly approached drought stress in coffee from an agronomic and yield perspective, with limited comprehensive metabolomic profiling. Given metabolites’ role in osmoregulation during drought stress, bolstering coffee drought research with metabolomic investigations is crucial. Thus, this study aims to characterize and profile metabolites’ response to drought in tolerant and sensitive C. arabica genotypes, enhancing our grasp of drought adaptation and possibly guiding robust coffee varieties. By understanding these mechanisms, we may pave the way for more climate-resilient coffee varieties.
2. Materials and Methods
2.1. Plant Material
In this study, we utilized four C. arabica L. (Ca) genotypes sourced from Jimma Agricultural Research Center (JARC). The selection of these genotypes was based on their drought tolerance, comprising both relatively tolerant (Ca74112 and Ca74110) and sensitive (Ca754 and CaJ-19) varieties, as previously reported by Chekol et al. [7] and Tesfaye [26]. Our experimental focus was to investigate the interplay between growth, physiological performance, and metabolite responses within adult coffee genotypes. Adhering to the guidelines from WCR [27], we transplanted germinated coffee plants once they exhibited the first leaf pair, and were disease-free, had 3-5 cm tall stems (hypocotyls), and 2-3 cm secondary roots. These germinants were transplanted in 5 L plastic pots, with the side of the pot covered with aluminum foil to prevent excessive heat buildup. The pots were filled with 4 L of potting media (topsoil, compost, and sand in a 2:1:1 ratio, pH 5.4-6.8) with drainage holes. To address specific nutritional requirements and align with distinct coffee growth stages, we added 2.0 g of NPK/DAP fertilizers 5-7 cm below the seedlings. Subsequently, uniform-looking seedlings of each genotype were placed within a greenhouse environment and received consistent watering prior to the initiation of the drought stress treatments (Figurea S1–S3).
2.2. Growth Condition and Experiment Design
The research was conducted within a controlled greenhouse condition where the relative humidity ranges between 50-70%, with an average temperature of 24.5°C and a photon flux density of 850 ± 13 μmol m−2s−1. Coffee genotypes (240 days aged), with 7-8 leaf pairs and free from disease or nutrient deficiencies, were used for the study, and subjected to two conditions: well-watered (ww) and drought-stressed (ws). Under ww, soil moisture of 60–80% field water capacity, seedlings were irrigated every 3-4 days. In contrast, for the ws conditions, seedlings were initially fully irrigated to the same field water capacity and subsequently subjected to drought conditions by withholding water until the experiment’s end (around 300 days of coffee age). The study used a completely randomized block design (CRBD) with four genotypes and two water regimes, each replicated ten times, totaling 80 coffee plants.
To evaluate the coffee growth and physiological performance in response to drought stress, at 10-day intervals until the end of the experiment (for around 60 days), measurements such as shoot height, leaf relative water content, stomatal conductance, and net carbon assimilation rate were measured. Besides, evaluations of pigments of the coffee genotypes were conducted at the beginning of the experiment (0 timelines) and end of the experiment (60 days after drought started). For the metabolite analysis, we sampled the third matured leaf from upper new flesh growth at two distinct time points during the study: at the start of the experiment (0 days) and end of the experiment (after 60 days of the drought implementation period). These fresh biomass leaf samples were snap frozen in liquid nitrogen and stored at -80°C until further analysis.
2.3. Leaf Relative Water Content
Based on the works of Barrs and Weatherley [28], at 10-day intervals until the end of the experiment (for around 60 days), relative water content (RWC) from representative leaves of the coffee genotypes was determined following the parameters:
where FW is leaf fresh weight, DW is leaf dry weight, and TW is leaf turgid (re-saturated) weight.
The fresh weight of the leaves of the genotypes was measured, and for the determination of turgid weight, samples were soaked in distilled water for about 2 h at room temperature (20–22°C) and weighed. Furthermore, for the determination of dry weight, the samples were dried to a constant weight at 70°C. Sample weights were measured using balance to an accuracy of 0.0001 g (Sartorius, Karnataka, India).
2.4. Gas Exchange Measurements
Instantaneous gas exchange measurements were periodically measured out at 10 day intervals until the end of the experiment (for around 60 days). The rate of stomatal conductance (Gs, mol H2O m−2s−1) and net carbon assimilation (Anet, µmol CO2 m−2s−1) were collected using an open gas exchange system LI-6400 (LI-COR, Lincoln, Nebraska, USA) adjusted at 1000 μmol m−2s−1 photosynthetic photon flux density, 400 μmol CO2 mol−1 air reference CO2 concentration, and 500 μmol s−1 flow rates. The measurements were conducted between 9:00 and 11:00 a.m., on a young and fully expanded leaf.
2.5. Content of Photosynthetic Pigments
Healthy and fully expanded leaf discs from the genotypes, the same leaves used for gas exchange measurements, were collected, at 0 and 60 days after the start of drought treatment and analyzed for the concentration of pigments (chlorophylls) from the genotypes based on the protocols of Lichtenthaler [29] using a UV-VIS spectrophotometer (Model 3092, Maharashtra, India). The contents of chlorophyll a (chl-a), and chlorophyll b (chl-b), were measured following the calculation:
where Chl-a is the content of chlorophyll-a (mg g−1 tissue), and Chl-b is the content chlorophyll-b (mg g−1 tissue).
Chl-a (mg/g tissue) = 12.25 A663.2 – 2.79 A646.8
Chl-b (mg/g tissue) = 21.50 A646.8 – 5.10 A663.2
2.6. Vegetative Growth Measurements
To evaluate the growth performance in response to drought stress, shoot height (SH, cm, using meter scale) of the coffee genotypes were conducted at 10 day intervals until the end of the experiment (for around 60 days). Following sample harvesting (plants at around 300 days of age), shoot fresh biomass (g) and dry biomass (g, oven-dry biomass at 70°C for 24 h) were measured using balance to an accuracy of 0.0001 g (Sartorius, Karnataka, India).
2.7. Metabolite Analysis
To analyze metabolites, the dried leaf biomass samples, frozen using liquid nitrogen and stored at -80°C, of the well-watered and drought stressed conditions from 0TL and 60TL, were ground into constant weight of fine powder under liquid nitrogen using Mortar and pestles. The powder was oven-dried to a constant weight at 60°C for 24 h. Approximately 100 mg powder was weighed, and were extracted in a 1 ml methanol:chloroform:water extraction solution (2.5:1:1 v/v) [30]. The mixture was thoroughly vortexed (MX-S, Yuzhong, China) and kept in an orbital shaker (OS-20Pro, Zhejiang, China) for duration of 15 minutes. Following this initial preparation, the samples underwent centrifugation (MSLZL19-Neuvar, CA, USA) for 10 minutes at 12000 revolutions per minute (rpm) and placed at 4°C. The resulting supernatant was then carefully transferred to 2 mL screw-top tubes, mixed with 300 μl of chloroform and 300 μl of Mass Spec (MS) grade water and then centrifuged at 20,000× g for 2 min. Subsequently, 100 μl of the polar phase (water-methanol phase) was dried in a vacuum concentrator (Vacufuge Plus, Eppendorf, Hamburg, Germany) at 30°C for a period of 3 hours, and stored at -80°C.
The dried polar extracts were derivatized with 40 µL of 20 mg mL−1methoxyamine hydrochloride, followed by 70 µL of N-methyl-N-trimethylsilyltrifluoroacetamide (TMS derivatization) and 20 µL mL−1 of a mixture of fatty acid methyl esters (FAMES). For metabolite analysis, the study utilized Gas Chromatography-Mass Spectrometry (GC-MS), employing an Agilent 7890 system coupled with a DB-5MS capillary column coated with a 5% diphenyl and 95% dimethylpolysiloxane mixture. During injection, an aliquot of the analyte (1 μL) was injected in a splitless mode. Helium served as the carrier gas, with a specific temperature program ranging from 90°C to 285°C. Peaks were manually annotated, and ion intensity was determined and metabolites were identified through systematic comparison with established reference library derived from the Golm Metabolome Database [31], based on retention time and indices, and mass spectrum, that enabling us to gain insights into the intricate metabolic profiles of the coffee genotypes under study. In order to understand the alteration of the metabolites, the resulting ion intensity of relative concentration were transformed and normalized for the removal of measurement bias, and set for further statistical analysis.
2.8. Statistical Analysis
The collected data were subjected to statistical analysis using an Analysis of Variance test. Post hoc multiple comparisons were performed using Tukey’s honest significant difference test (p<0.05) to identify significant differences among the experimental groups. The dataset was transformed in Past version 4.0.3 [32]. Pearson correlation analysis, between all metabolite pairs and among metabolite, growth, and physiological traits, was performed after checking the assumptions of normality using the Shapiro–Wilk test. To reconstruct a metabolite network that would capture the coordinated changes in the metabolic profiles, threshold values were determined. Network visualization of metabolites was performed using Cytoscape version 3.10.1 [33], and the number of nodes, number of edges, edge to node ratio, network density, average node degree, characteristics path length, clustering coefficient, network heterogeneity, network diameter, network radius, and network centralization were investigated. To construct correlation-based networks of significant correlations, r>0.8 threshold values were applied. Principal component analyses were performed on the transformed (ws/ww), using RStudio (version 4.2.1). All statistical analyses were performed using SigmaPlot version 13 (Systat Software Inc., San Jose, CA, US).
3. Results
3.1. Shoot Growth and Biomass Were Affected by Drought Stress Treatments
Distinct variations in growth were evident among the four coffee genotypes following a 60-day period of drought-induced stress. When compared to well-watered coffee genotypes, those exposed to drought stress exhibited notably lower shoot height, shoot fresh and dry biomass at the end of the stress period, displaying statistically significant differences (p<0.05). Within the conditions of drought stress, markedly higher shoot height (Figure 1), shoot fresh biomass (Figure 2A), and shoot dry biomass (Figure 2B) were observed in the relatively tolerant genotypes Ca74112 (18.14±0.04 cm, 9.3 g, and 2.71 g) and Ca74110 (15.35±0.09 cm, 7.11 g, and 2.01 g), whereas lower values were recorded in the sensitive genotypes of Ca754 (11.38±0.3 cm, 4.43 g, and 1.17 g) and CaJ-19 (13.25±0.19 cm, 5.73 g, and 1.57 g), respectively. In comparison to the well-watered genotypes, at the conclusion of the experiment under drought stress conditions, the relatively tolerant genotypes Ca74112 (38.33%) and Ca74110 (43.59%) displayed the least reduction in shoot growth, while the sensitive genotypes Ca754 (50.84%) and CaJ-19 (47.17%) experienced higher reductions in shoot heights (Table S1). Similarly, at the end of the experiment, the least reduction in both shoot fresh and dry weight was observed in the relatively tolerant genotypes Ca74112 (31.01%, 30.87%) and Ca74110 (41.34%, 41.23%), compared to the sensitive genotypes Ca754 (62.14%, 62.13%) and CaJ-19 (60.21%, 60.25%) (Table S2).
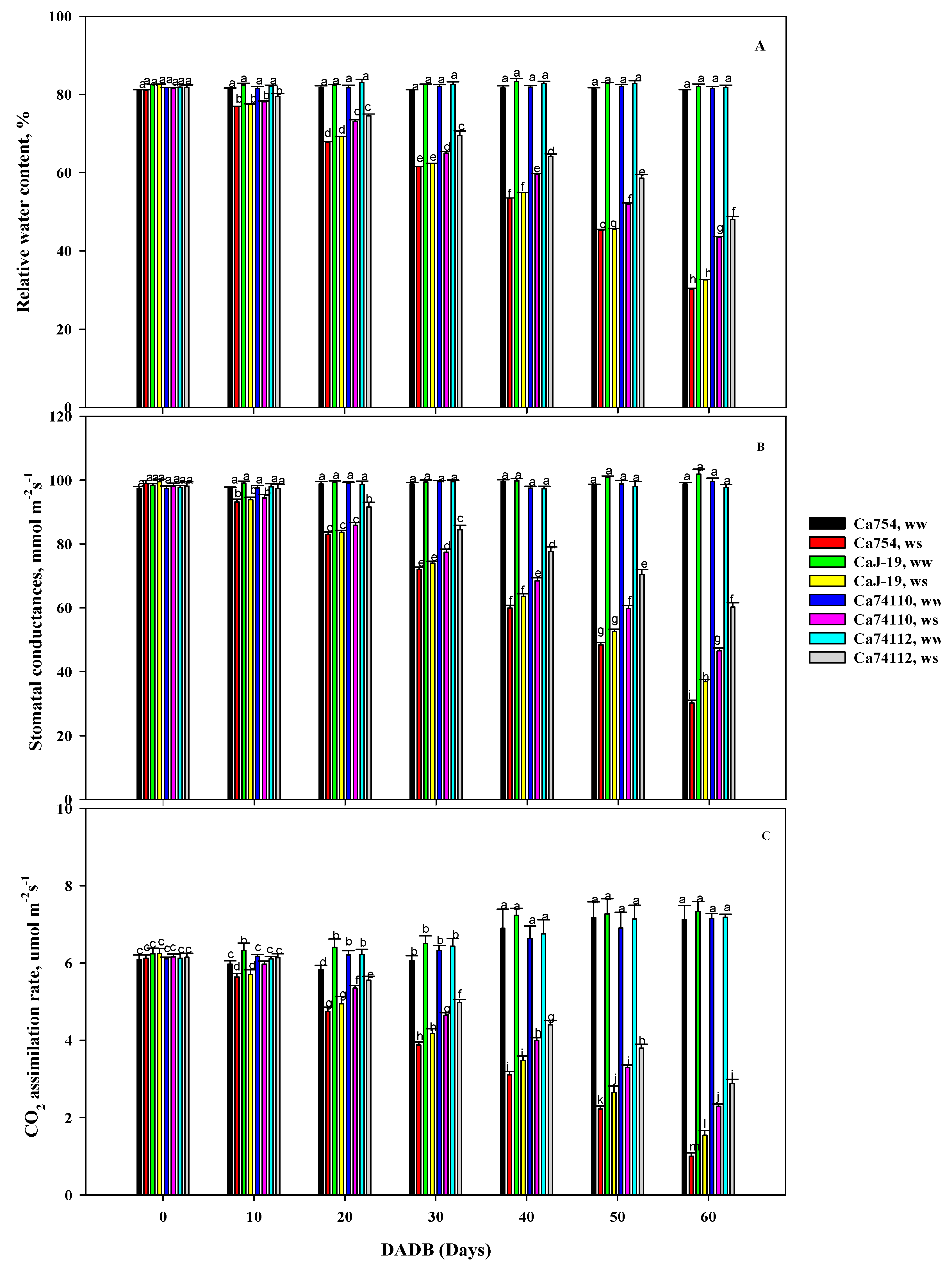
3.2. Difference of Relative Water Content among Coffee Genotypes under Drought Stress
In well-watered conditions, there were no significant (p<0.05) differences among the genotypes where the relative water content value ranges between 81.16% - 82.76% (Figure 3A). However, under drought-stressed conditions, at the end of the experiment, the mean relative water content was significantly (p<0.05) lower than those in well-watered conditions and the value of RWC was different among coffee genotypes, and a higher RWC were identified in the relatively tolerant genotypes of Ca74112 (48.09±0.8%, with 41.89% reduction than well-watered (ww) conditions) and Ca74110 (43.40±0.29%, with 46.74% reduction than well-watered (ww) conditions), and a lower RWC were recorded in the sensitive genotypes of Ca754 (30.24±0.21%, with 62.74% reduction than ww conditions) and CaJ-19 (32.57±0.13%, with 60.32% reduction than ww conditions) (Table S3).
3.3. Influence of Drought Stress in Stomatal Conductance among the Coffee Genotypes
At the initial stage, no significant differences in stomatal conductance (Gs, mmol m−2s−1) were observed between plants grown under drought stress and those under well-watered conditions. However, by the end of the experiments, noticeable variations in Gs were evident across genotypes. Among the drought-stressed plants, the highest Gs was recorded in the relatively tolerant genotypes Ca74112 (60.25 ± 1.38 mmol m−2s−1) and Ca74110 (46.51 ± 0.89 mmol m−2s−1), while the minimum Gs was recorded in the sensitive genotypes CaJ-19 (36.84 ± 0.71 mmol m−2s−1) and Ca754 (30.28 ± 0.86 mmol m−2s−1) (Figure 3B). Because of the imposed drought stress, there were reductions in Gs by 38.32%, 53.31%, 63.85%, and 69.34% in the genotypes Ca74112, Ca74110, CaJ-19, and Ca754, respectively (Table S4).
3.4. Drought Stress Associated Variation in Carbon Assimilation among Coffee Genotypes
The impact of drought stress on net carbon assimilation rate (Anet, µmol CO2 m−2s−1), stomatal conductance (Gs, mmol m−2s−1) and transpiration rate (E, mmol m−2s−1) were examined. There were no significant differences in the CO2 assimilation rate among the genotypes grown under control conditions. However, when subjected to drought stress conditions, all genotypes displayed notably distinct reductions in CO2 assimilation rate, exhibiting a gradual decline throughout the experiment (Figure 3C). The relatively tolerant genotypes of Ca74112 (2.89±0.11 µmol m−2s−1) and Ca74110 (2.29±0.06 µmol m−2s−1) recorded higher Anet values, while lower values were observed in the sensitive genotypes CaJ-19 (1.55±0.13 µmol m−2s−1) and Ca754 (1.00±0.09 µmol m−2s−1). In assessing the impact of drought stress on the reduction of CO2 assimilation rate, the sensitive genotype Ca754 experienced the highest reduction rate (85.96%), significantly greater than the lowest reduction observed in the tolerant genotype Ca74112 (59.85%) (Table S5).
Figure 3.
Effects of drought stress on (A) relative water content, (B) stomatal conductance, and (C) net assimilation rate of the four adult coffee genotypes, Ca754, CaJ-19, Ca74110, and Ca74112, under well water (ww) and drought stress conditions (ws), at different DADB. Bars indicate means±SD (n=10 replicates per genotype). Bars with the same letter indicate no significant difference at p<0.05 between samples. DADB indicates the number of days after drought stress begins.
Figure 3.
Effects of drought stress on (A) relative water content, (B) stomatal conductance, and (C) net assimilation rate of the four adult coffee genotypes, Ca754, CaJ-19, Ca74110, and Ca74112, under well water (ww) and drought stress conditions (ws), at different DADB. Bars indicate means±SD (n=10 replicates per genotype). Bars with the same letter indicate no significant difference at p<0.05 between samples. DADB indicates the number of days after drought stress begins.
3.5. Variations of Photosynthetic Pigments under Drought Stress among Coffee Genotypes
Plants grown under well-watered conditions consistently exhibited significantly higher pigment content compared to those experiencing drought stresses. Across all tested coffee genotypes, drought stress led to a notable decline in Chl-a (Figure 4A) and Chl-b (Figure 4B) content, while in well-watered plants, chlorophyll levels remained relatively stable throughout the experiment. By the end of the drought stress period, genotype Ca74112 recorded the significantly highest and lowest Chl-a and Chl-b values (1.09 mg g−1fw and 0.21 mg g−1fw, respectively), while Ca754 had the lowest values (0.63 mg g−1fw for Chl-a and 0.14 mg g−1fw for Chl-b) (p<0.05). Comparing the reduction rates of Chl-a and Chl-b due to drought stress, the sensitive genotype Ca754 displayed the highest reduction rates (56.96% for Chl-a and 57.14% for Chl-b), whereas the tolerant genotype Ca74112 exhibited lower reduction rates (28.56% for Chl-a and 38.72% for Chl-b) (Table S6).
3.6. Alteration of Metabolites under Drought Stress Conditions
To investigate the molecular changes associated with drought tolerance, we examined metabolic responses at two distinct time points: 0 and 60 days into the drought implementation periods. A profiling analysis revealed 50 identified metabolites spanning sugars, amino acids, and intermediates from pathways such as the tricarboxylic acid cycle (TCA), glycolysis, γ-aminobutyric acid (GABA) shunt, and shikimic pathways. The relative concentrations of each metabolite are represented as fold changes (ws/ww) (see Figure 5, Figures S4 and S5, Table S7). Initially, no significant differences in metabolite alterations were observed between relatively tolerant and sensitive coffee genotypes in both treatment groups. However, after 60 days of drought stress initiation (DADB), a noteworthy shift occurred. The relatively tolerant genotypes, Ca74112 (with 46 altered metabolites - 30 up and 16 down) and Ca74110 (with 46 altered metabolites - 29 up and 17 down), displayed significantly higher alterations compared to the sensitive genotypes, CaJ-19 (42 altered metabolites - 23 up and 19 down) and Ca754 (40 altered metabolites - 22 up and 18 down). At the outset of the study (0TL), no significant metabolite alterations were evident between the well-watered/control (ww) and drought-stressed (ws) treatments. However, by 60 DADB, substantial alterations in metabolite accumulation were observed compared to the control (p<0.01). At 60 DADB, the relatively tolerant genotypes exhibited markedly higher alterations and increased metabolite accumulation compared to the sensitive genotypes, with a notably greater magnitude. Specifically, in the relatively tolerant genotypes (Ca74110 and Ca74112), the most substantial up-accumulation was observed in glucose (68.67 and 93-fold), tryptophan (67 and 80-fold), maltose (42.4 and 42.27-fold), galactose (25.8 and 37.7-fold), L-Cysteine (2 and 48.44-fold), lactose (27.12 and 32.87-fold), lysine (14.18 and 22.71-fold), methionine (20.4 and 18.15-fold), leucine (9.66 and 14.45-fold), pyruvic acid (13.06 and 12.83-fold), ribose (8.82 and 9.59-fold), and other amino acids and organic acids. Conversely, in Ca754, CaJ-19, Ca74110 and Ca74112 respectively, significant reductions (p<0.05) were observed in glutamate (0.13, 0.16, 0.15 and 0.15-fold), aconitic acid (0.65, 0.56, 0.37 and 0.28-fold), glucose-6-phosphate (0.53, 0.68, 0.12 and 0.33-fold), γ-aminobutyric acid (0.49, 0.39, 0.36 and 0.48-fold), fructose-6-phosphate (0.74, 0.81, 0.29 and 0.41-fold), asparagine (0.38, 0.29, 0.51 and 0.57-fold), glutamine (0.23, 0.22, 0.19 and 0.59-fold), aspartate (0.66, 0.47, 0.56 and 0.63-fold), alanine (0.82, 0.98, 0.74 and 0.64-fold), myo-inositol (0.69, 0.68, 0.67 and 0.68-fold), and citric acid (0.55, 0.27, 0.63 and 0.86-fold) among the four genotypes (see Figures S6–S9, Table S8).
3.7. Correlation of Specific Metabolites with Growth And Physiology
An interesting aspect of this study was to combine knowledge from growth and physiological response of coffee genotypes with metabolic data. For this purpose, a correlation analysis of the growth and physiological parameters with metabolic response, at 60 DADB, was performed. Among the highly increased sugar metabolites, a strong positive correlation of glucose, maltose, and galactose, with shoot fresh biomass (r=0.89; r=0.837; r=0.927, respectively), RWC (r=0.939; r=0.942; r=0.993, respectively), Anet (r=0.946; r=0.905; r=0.964, respectively), and Gs (r=0.884; r=0.842; r=0.936, respectively) were observed. Besides, a strong positive correlation of tryptophan, L-Cysteine, and proline with shoot fresh biomass (r=0.867; r=0.886; r=0.908, respectively), RWC (r=0.969; r=0.771; r=0.987, respectively), Anet (r=0.917; r=0.805; r=0.948, respectively), and Gs (r=0.88; r=0.891; r=0.918, respectively) were observed. From the TCA cycle and glycolysis pathway, a strong positive correlation for malic and pyruvic acid with RWC (r=0.922; r=0.968, respectively), Anet (r=0.841; r=0.923, respectively), and Gs (r=0.808; r=0.878, respectively) were observed. A strong negative correlation of myo-inositol with shoot fresh biomass (r=-0.613), RWC (r=-0.695), Anet (r=-0.713), Gs (r=-0.595), and E (r=-0.61) were observed. Glutamate was weakly correlated with shoot fresh biomass (r=0.539), RWC (r=0.426), Anet (r=0.562), and Gs (r=0.496). α-ketoglutaric acid and glucose-6-phosphate were negatively correlated with shoot fresh biomass (r=-0.966; r=-0.573, respectively), RWC (r=-0.933; r=-0.758, respectively), Anet (r=-0.933; r=-0.682, respectively), and Gs (r=-0.976; r=-0.584, respectively). Data for all significant correlations among the growth, physiology and metabolites are available in Table S9.
3.8. Correlation-Based Network Analysis to Identify Coordinated Stress Induced Metabolic Perturbation
The 60 days of drought stress caused an increase in the number of edges, network densities, average node degree and other key values of metabolites network analysis, and the increased value was more characterized in the relatively tolerant than the sensitive coffee genotypes. Thus, at r>0.8, the mean values of network analysis in the relatively drought tolerant genotypes showed that, they had 87 nodes, 1250 number of edges, 14.4 to 1 edge to node ratio, 0.359 network density, 29.762 average node degree, 4.268 characteristics path length, 0.924 clustering coefficient, 0.385 network heterogeneity, 10 network diameter, 5 network radius, and 0.126 network centralization, while the sensitive genotypes had 80 nodes, 1411 number of edges, 17.6 to 1 edge to node ratio, 0.907 network density, 36.293 average node degree, 1.094 characteristics path length, 0.991 clustering coefficient, 0.211 network heterogeneity, 3 network diameter, 2 network radius, and 0.071 network centralization (Figure 6 and Figure 7, Table S10).
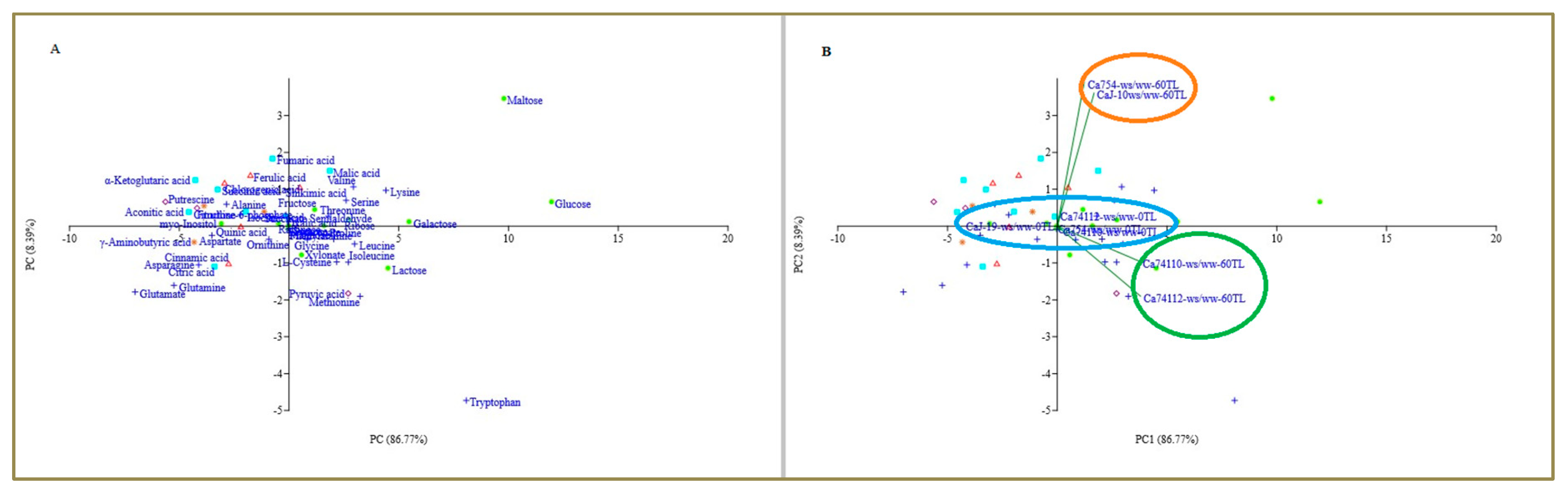
3.9. PCA Analysis
PCA results demonstrate an obvious metabolite accumulation and genotype category during the drought stress exposition. The first principal component (PC1) and second principal component (PC2) represent 86.77% and 8.39% of the PCA, respectively. The contribution of metabolites for PC1 was dominated by glucose, maltose, tryptophan, galactose, lactose, lysine, methionine, leucine, valine, ribose, isoleucine, pyruvic acid, and serine; whereas, maltose, fumaric acid, malic acid, ferulic acid, α-Ketoglutaric acid, chlorogenic acid, valine, shikimic acid, and succinic acid were major contributors for separation along PC2 (Figure 8, Tables S11 and S12).
In order to understand the drought induced responsive metabolites, the metabolite’s up- and down-accumulation score values were considered for the analysis in each genotype, and the score value of PC1 (86.77%) was taken as the weight. As a result of this, significantly (p<0.001) the most responsive metabolite PC1 score values were identified in glucose (11.96 PCA score value), maltose (9.79 PCA score value), tryptophan (8.08 PCA score value), galactose (5.47 PCA score value), lactose (4.50 PCA score value), lysine (4.42 PCA score value), methionine (3.23 PCA score value), leucine (2.99 PCA score value), valine (2.94 PCA score value),ribose (2.71 PCA score value), isoleucine (2.70 PCA score value), pyruvic acid (2.70 PCA score value), and serine (2.60 PCA score value) (Table S12).
Similarly, following the same protocol, to briefly assess the drought tolerance capacity of the genotypes, the genotypes-against the respective water treatments- score values were taken into consideration. The result showed that the PC score value of the genotypes under drought stressed conditions was significantly (p<0.05) higher than those under well-watered conditions. However, among the coffee genotypes under drought stressed conditions, significantly the highest PC1 score values were identified by the relatively tolerant genotypes of Ca74112 (0.972 PCA score value) and Ca74110 (0.977 PCA score value) which were followed by the sensitive genotypes of CaJ-19 (0.803 PCA score value) and Ca754 (0.679 PCA score value) (Figures S10–S12, Table S13).
Figure 9.
Principle component analysis (PCA) plot (x – first component, y – second component) plot indicating (A) the row labels of each metabolite and (B) the biplot of the four coffee genotypes, Ca754, CaJ-19, Ca74110, and Ca74112 based on GC/MS analysis, at 0 and 60 days after drought begins. Sugars (orange, dot), amino acids (blue, plus), TCA (aqua blue, square), glycolysis (pink, diamond), GABA shut (brown, star), and shikimik pathway (red, triangle).
Figure 9.
Principle component analysis (PCA) plot (x – first component, y – second component) plot indicating (A) the row labels of each metabolite and (B) the biplot of the four coffee genotypes, Ca754, CaJ-19, Ca74110, and Ca74112 based on GC/MS analysis, at 0 and 60 days after drought begins. Sugars (orange, dot), amino acids (blue, plus), TCA (aqua blue, square), glycolysis (pink, diamond), GABA shut (brown, star), and shikimik pathway (red, triangle).
4. Discussion
4.1. Genotypic Variability and Physiological Responses
The physiological impact of drought stress on plants is often manifested in restricted growth and developmental limitations due to the scarcity of water [34]. Coffee plants, both at the seedling or mature stages, exhibit high sensitivity to soil moisture levels, profoundly affecting their subsequent growth and development [7,35]. Consistent with the findings of Silva et al. [9], our study noted a significant decline in the growth performance of coffee plants under drought stress, reflected in reduced shoot length and lower fresh and dry biomass. Notably, after a 60-day drought stress period, relatively higher shoot length and increased fresh and dry biomass were observed in the relatively tolerant genotypes Ca74110 and Ca74112 compared to the sensitive genotypes Ca754 and CaJ-19. Under drought stress, any decrease in turgor pressure and water potential can impede cell division, expansion, and elongation, leading to reduced leaf area, smaller leaf size, and ultimately lower photosynthetic rates by limiting CO2 assimilation [7,9,34]. Studies by DaMatta et al. [36] and Wei et al. [37] have highlighted the impact of drought stress on diminishing shoot and root growth in coffee and Lycium barbarum plants, respectively. However, previous research by Chekol et al. [7] and Dias et al. [38] suggest that in tolerant genotypes, enhanced growth responses are linked to water conservation mechanisms that enable coffee plants to sustain cell division and elongation processes. Studies by Caine et al. [25] and Xiong et al. [39] on rice and oak plants, respectively, also support the notion that limited water availability triggers metabolic responses favoring cellular division. This, in turn, favoring the development of dermal tissue, ground tissue, and vascular tissue, essential components contributing to the plant’s adaptation to drought stress. Shoot growth and development serve as key indicators of a plant’s response to drought stress and are often considered key parameters in assessing a plant’s drought tolerance [40]. Similar with the observations of Mirian et al. [41], our results indicate that the relatively tolerant genotypes Ca74110 and Ca74112 maintain higher growth metrics—such as shoot length, fresh and dry biomass, and specific leaf area—more effectively than the sensitive genotypes Ca754 and CaJ-19 under drought stress conditions.
4.2. Relative Water Contents, Gas Exchange and Pigments Variations Among Coffee Genotypes Towards Drought Stress
In the current study, drought stress significantly impacted various physiological parameters of coffee genotypes, notably reducing leaf relative water content, net assimilation rate, stomatal exchange, transpiration, and chlorophyll pigments compared to well-watered conditions. Similar reductions in these parameters under drought stress were reported in other studies on coffee [7,9], cowpea [42], and other tolerant crops [43]. The tolerant coffee genotypes (Ca74110 and Ca74112), displayed higher relative water content even under drought stress, aligning with findings in potato genotypes reported by Soltys-Kalina et al. [44].
Drought stress often leads to decreased photosynthesis assimilation rates, stomatal conductance, and transpiration rates, affecting gaseous exchange parameters [34,39]. Similarly, tolerant coffee genotypes exhibited better physiological performances in these parameters compared to sensitive genotypes under drought stress [7,38]. The reduction in photosynthesis rate under drought is usually associated with stomatal closure and decreased internal CO2 concentrations, impacting CO2 fixation and pigment synthesis [9,45,46]. This decline in photosynthesis rate, along with reduced stomatal conductance and transpiration rates, often leads to diminished pigment synthesis [47]. Tolerant genotypes (Ca74110 and Ca74112) maintained higher pigment contents even under drought stress compared to sensitive genotypes (Ca754 and CaJ-19). Drought stress often affects the structure and function of photosynthetic pigments by damaging thylakoid membranes and reducing the activity of essential enzymes like RUBISCO [48]. This stress-associated decline in chlorophyll content is observed in various crops, indicating damage to light-harvesting complex proteins, impacting photon absorption and electron transport [49]. Perhaps, relatively drought-tolerant genotypes (Ca74110 and Ca74112) might possess better protective mechanisms against chlorophyll degradation enzymes than sensitive genotypes (Ca754 and CaJ-19).
4.3. Drought Stress Causes Variability in Metabolite Alteration among Coffee Genotypes
Drought stress triggers significant changes in the biosynthesis and transport of primary and secondary metabolites, orchestrating adjustments in plants’ physiological and biochemical processes [17]. Differentiating between tolerant and sensitive behaviors, plants respond diversely to shifting soil moisture regime [9]. Tolerant plants usually sustain metabolic processes and defense responses, whereas sensitive ones operate in the opposite manner [50]. In our study, we focused on sugars, amino acids, and organic acids synthesis in different pathways like glycolysis, GABA shut, TCA cycle, and shikimic path. After a 60-day drought stress period, relatively tolerant genotypes (28 in Ca74110 and 29 in Ca74112) up-accumulated more metabolites compared to sensitive genotypes (25 in Ca754 and 21 in CaJ-19). Conversely, sensitive genotypes (18 in Ca754 and 22 in CaJ-19) down-accumulated more metabolites than relatively tolerant ones (17 in Ca74110 and 16 in Ca74112). This reflects significantly higher metabolite up-accumulation in the relatively tolerant Ca74112 and Ca74110 genotypes compared to sensitive Ca754 and CaJ-19 genotypes (p<0.01). According to Fabregas and Fernie [20], organic biomolecules like sugars, amino acids, and others play pivotal roles in osmotic adjustment during drought stress by regulating vacuolar osmotic potential. Kapoor et al. [51] also noted that these metabolite responses to drought vary not only between species but also among genotypes and different parts of the plant. The PCA analysis further revealed distinct separation and clustering between the relatively tolerant and sensitive coffee genotypes, indicating diverse mechanisms of metabolite alteration in response to drought stress. Previous findings by Rodrigues et al. [2] on coffee plants and Hochberg et al. [13] on grapevines suggest that prolonged drought stress triggers adjustments in various metabolites, enhancing the plants’ resilience to drought. Similarly, Xiong et al. [39] on Quercus species indicate that during drought stress, metabolite alterations are more pronounced in tolerant genotypes, providing enhanced resistance to manage plant growth and development.
Sugars are highly sensitive metabolite classes to drought stress and extensively studied [16]. Profiling 12 sugars showed increased levels across all genotypes under drought stress condition, notably higher in relatively tolerant Ca74110 and Ca74112, particularly in glucose, maltose, galactose, lactose, and ribose, compared to sensitive Ca754 and CaJ-19. However, in the current study myo-inositol concentration decreased universally, indicating its drought sensitivity. These sugars play key roles in osmotic adjustments, membrane stability, and maintaining leaf water content during drought stress [20]. Previous studies by Urano et al. [52], Krasensky and Jonak [53], Fabregas et al. [54], Ogbaga et al. [55], and Pires et al. [56] align with our findings, demonstrating increased fructose, glucose, raffinose, and other sugar levels during drought stress in various plants. Additionally, similar to our observations, Urano et al. [52] also noted a reduction in myo-inositol content under drought conditions.
The study identified 19 amino acids, showing up-accumulation in certain amino acids across all genotypes, particularly pronounced in relatively tolerant Ca74110 and Ca74112, including tryptophan, L-Cysteine, lysine, methionine, valine, leucine, isoleucine, serine, and proline compared to sensitive Ca754 and CaJ-19. However, alanine, aspartate, glutamate, glutamine, and asparagine decreased universally. During drought stress, amino acids usually act as osmolytes and scavengers of reactive oxygen species, influencing cellular functions [57]. Proline accumulation correlates with drought tolerance [58,59], as demonstrated by various studies including Konieczna et al. [34], Zhang et al. [60], and Joshi et al. [61]. Decreases in certain amino acids during drought stress might be due to redirect metabolic activities towards proline biosynthesis [62]. Enhanced protein degradation or inhibition of biosynthesis presumably contributes to amino acid increases during prolonged drought [56]. Amino acids linked to pyruvate metabolism increase might be due to their involvement in gluconeogenesis to alleviate transamination products [63]. The correlation-based network analysis also demonstrated heightened coordinated metabolic activities in relatively tolerant coffee genotypes, showcasing their resilience to drought stress. Hochberg et al.’s [13] finding in grapevines research supported this, indicating that prolonged drought stress can boost metabolic network density. Likewise, Sanchez et al. [64] observed increased network connectivity in Lotus genotypes facing salt stress, aligning with these findings.
Among the identified 8 tricarboxylic acid (TCA) cycle intermediates, mostly marked reduction levels across all genotypes were apparent, particularly higher in relatively tolerant Ca74110 and Ca74112 (malic acid, oxalic acid) compared to sensitive Ca754 and CaJ-19. TCA cycle metabolites’ responses in drought stress remain less pronounced than sugars and amino acids. Araujo et al. [65] also noted limited alterations in the TCA cycle during drought stress, similar to this study. Fabregas et al. [54] demonstrated analogous changes in Arabidopsis, linking increased malic acid and oxalic acid levels to suppressed malate dehydrogenase, aiding nutrient uptake and intracellular ionic regulation under drought conditions. The increase in malic acid level and oxalic acid is associated with decreased sink tissue utilization due to malate dehydrogenase suppression [66]. Yang et al. [67] also reported declines in citrate, succinate, α-ketoglutarate, and fumarate in drought-stressed maize kernels, aligning with this study’s findings.
The study examined 11 metabolites from glycolysis, GABA, and shikimic biosynthetic pathways, observing increased levels universally in all genotypes, notably more pronounced in relatively tolerant Ca74110 and Ca74112 (pyruvic acid, shikimic acid) compared to sensitive Ca754 and CaJ-19. However, fructose-6-phosphate, glucose-6-phosphate, γ-aminobutyric acid, succinate semialdehyde, and putrescine exhibited reduced levels across all genotypes. Consistent with this study, Rabara et al. [17] also reported decreased glucose-6-phosphate and fructose-6-phosphate in tobacco and soybean leaves under drought stress. Guo et al. [15] observed a declining trend in GABA shunt metabolites in wheat under drought. As per Kinnersley’s review [68], γ-aminobutyric acid (GABA) levels increased in response to drought stress in various plant species.
5. Conclusions
Under drought stress, coffee plants accumulate diverse metabolites including sugars, amino acids, TCA, GABA shut, glycolysis, and shikimic pathways, vital in regulating osmotic potential, ion transport, turgor pressure, and cell membrane stability. These metabolic changes reflect adaptive responses for drought tolerance. Comparing metabolic profiles and physiological responses, relative tolerant genotypes (Ca74110 and Ca74112) exhibit higher compatible solutes, active metabolite synthesis, and faster growth than sensitive group under drought stress. To address coffee demand amidst ongoing drought, enhancing drought tolerance in Coffee arabica genotypes is essential. In this regard, adjusting metabolic gene expression via genetic engineering stands as a potential approach. Implementing metabolic engineering in drought tolerance development programs could significantly enhance sustainable coffee bean yields and productivity to meet the growing demand for coffee consumption.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org, Figure S1. Seeds of the four C. arabica genotypes used in this study; Figure S2. Representative example of C. arabica sand germination process of the nine genotype seeds: (A) early stage (maximum 26.0±2.31 days), (B) matchstick stage (max. 32.0±2.09 days), (C) butterfly stage (max. 46.0±2.23 days), and (D) transplanting stage (max. 53.2±3.86 days); Figure S3. Transplanting the C. arabica genotypes (A) from the sand media, (B) pulling up the genotypes without root damage, (C) the initial seedling, (D) initial seedling after transplanting, (E) seedlings at the age of 6 leaf pairs (6-month-old), (F) at the start of the experiment when genotypes developed 7-8 leaf pairs (240-270 days, 8-9 month old); Figure S4. The stacked bar chart representation of the metabolites response, in percentage, of the coffee genotypes growing under ww and ws conditions, at 0 and 60 days after drought stress begins. ww and ws refers to well-water and drought stress, respectively; Figure S5. Box and jitter plot representation of the coffee genotypes growing under ww and ws conditions, at 0 and 60 days after drought stress begins. ww and ws refers to well-water and drought stress, respectively; Figure S6. Fold changes (drought stress/well-watered) value of sugars accumulation in the four coffee genotypes, Ca754, CaJ-19, Ca74110, and Ca74112, at 0 and 60 days after drought stress begins. ww and ws refers to well-watered/control and drought stressed, respectively. Fold changes were calculated using the formula (value of ws per ww); Figure S7. Fold changes (drought stress/well-watered) value of amino acids accumulation in the four coffee genotypes, Ca754, CaJ-19, Ca74110, and Ca74112, at 0 and 60 days after drought stress begins. ww and ws refers to well-watered/control and drought stressed, respectively. Fold changes were calculated using the formula (value of ws per ww); Figure S8. Fold changes (drought stress/well-watered) value of intermediate metabolites from the tricarboxylic acid pathway in the four coffee genotypes, Ca754, CaJ-19, Ca74110, and Ca74112, at 0 and 60 days after drought stress begins. ww and ws refers to well-watered/control and drought stressed, respectively. Fold changes were calculated using the formula (value of ws per ww); Figure S9. Fold changes (drought stress/well-watered) value of intermediate metabolites from the glycolysis, GABA shut and shikimic pathway in the four coffee genotypes, Ca754, CaJ-19, Ca74110, and Ca74112, at 0 and 60 days after drought stress begins. ww and ws refers to well-watered/control and drought stressed, respectively. Fold changes were calculated using the formula (value of ws per ww); Figure S10. Eigenvalue and variance of the principal component analysis of the coffee genotypes growing under ww and ws conditions, at 0 and 60 days after drought stress begins. ww and ws refers to well-water and drought stress, respectively; Figure S11. PC1 loading value of the coffee genotypes growing under ww and ws conditions, at 0 and 60 days after drought stress begins. ww and ws refers to well-water and drought stress, respectively; Figure S12. PC2 loading value of the coffee genotypes growing under ww and ws conditions, at 0 and 60 days after drought stress begins. ww and ws refers to well-water and drought stress, respectively. Table S1. Mean values and SD of stem height (cm); Table S2. Mean values and SD of fresh and dry weight (g); Table S3. Mean values and SD of relative water content (%); Table S4. Mean values and SD of stomatal conductance (Gs, mmol m−2s−1); TableS5. Mean values and SD of net carbon assimilation (Anet, µmol CO2 m−2s−1); Table S6. Mean values and SD of photosynthetic Chl-a and Chl-b (mg g−1fw); Table S7. Row data of the relative concentration of metabolites of the coffee genotypes growing under ww and ws conditions, at 0 and 60 days after drought stress begins. ww and ws refers to well-water and drought stress, respectively; TableS8. Transformed fold change (Log2) of metabolites of the coffee genotypes growing under ww and ws conditions, at 0 and 60 days after drought stress begins. ww and ws refers to well-water and drought stress, respectively; Table S9. Pearson correlation analysis and heat-map of growth, physiology, and metabolites of coffee genotypes of the four coffee genotypes, Ca754, CaJ-19, Ca74110, and Ca74112, under ww and ws conditions, both at 0-TL and 60-TL periods; Table S10. Metabolite network analysis of coffee genotypes of the four coffee genotypes, Ca754, CaJ-19, Ca74110, and Ca74112, under ww and ws conditions, both at 0-TL and 60-TL periods; Table S11. Eigenvalue and variance of the principal component analysis of the coffee genotypes growing under ww and ws conditions, at 0 and 60 days after drought stress begins. ww and ws refers to well-water and drought stress, respectively; TableS12. Heat-map and PCA score value representing the total metabolite’s up- and down-accumulation of the four coffee genotypes, Ca754, CaJ-19, Ca74110, and Ca74112, after 60 days after drought begins. Red, the lowest value; green, highest value; and, in between colors (yellow, orange, light green) represent the medium value; Table S13. Heat-map and PCA score value representing the drought tolerance capacity of the four coffee genotypes, Ca754, CaJ-19, Ca74110, and Ca74112, after 60 days after drought begins. Red, the lowest value; green, highest value; and, in between colors (yellow, orange, light green) represent the medium value.
Author Contributions
Conceptualization, A.D., B.W., and H.C.; methodology, A.D., T.S., and H.C.; investigation, H.C.; writing-original draft, H.C.; writing-review and editing of the manuscript, H.C., A.D., G.B.D., and A.M.-A.; visualization, A.D., H.C., and A.M.-A.; funding acquisition, A.D., B.W., and G.B.D.; resources, A.D., B.W., and H.C.; supervision, A.D., T.S., B.W., G.B.D., and A.M.-A. All authors have read and agreed to the published version of the manuscript.
Funding
We thank Addis Ababa University for providing the research fund (LT/PY-038/2019) and the Department of Plant Biology and Biodiversity Management for facilitating the financial support.
Institutional Review Board Statement
Not applicable.
Data Availability Statement
Relevant data applicable to this research are within the paper and are also available on request from the corresponding author.
Acknowledgments
We are grateful to the Jimma Agricultural Research Center for providing the seeds of coffee (C. arabica) genotypes, and Addis Ababa University for supporting the coffee metabolite’s analysis work.
Conflicts of Interest
The authors declare that they have no competing interests.
References
- Kufa, T. Biomass production and distribution in seedlings of Coffea arabica genotypes under contrasting nursery environments in south-western Ethiopia. Agri. Sci. 2012, 3, 835–843. [Google Scholar] [CrossRef]
- Rodrigues, A.M.; Jorge, T.; Osorio, S.; Pott, D.M.; Lidon, F.C.; DaMatta, F.M.; Marques, I.; Ribeiro- Barros, A.I.; Ramalho, J.C.; Antonio, C. Primary metabolite profile changes in coffea spp. promoted by single and combined exposure to drought and elevated CO2 concentration. Metabolites 2021, 11, 427. [Google Scholar] [CrossRef] [PubMed]
- Elmar, R.; Jean-François, M. The major evolutionary lineages of the coffee family (Rubiaceae, Angiosperms). Combined analysis (nDNA and cpDNA) to infer the position of Coptosapelta and Luculia, and Supertree construction based on rbcL, rps16, trnL-trnF and atpB-rbcL Data. A new classification in two subfamilies, Cinchonoideae and Rubioideae. Sys. Geog. Plants 2006, 76, 85–145. [Google Scholar]
- FAOSTAT. The United Nations Food and Agriculture Organisation of the United Nations (FAO). Crop Statistics: New York, USA, 2021; pp 1-1357.
- CSA (Central Statistical Agency). Agricultural Sample Survey: Area and Production of Crops; Central Statistical Agency of Ethiopia: Addis Ababa, Ethiopia, 2019; pp. 1–450. [Google Scholar]
- ECX (Ethiopian Commodity Exchange). The Ethiopian Annual Export Products, Annual Reports; ECX publishing: Addis Ababa, Ethiopia, 2019; pp. 1–130. [Google Scholar]
- Chekol, H.; Bezuayehu, Y.; Warkineh, B.; Shimber, T.; Mierek-Adamska, A.; Dąbrowska, G.B.; Degu, A. Unraveling Drought Tolerance and Sensitivity in Coffee Genotypes: Insights from Seed Traits, Germination, and Growth-Physiological Responses. Agriculture 2023, 13, 1754. [Google Scholar] [CrossRef]
- Leon-Burgos, A.F.; Unigarro, C.; Balaguera-Lopez, H.E. Can prolonged conditions of water deficit alter photosynthetic performance and water relations of coffee plants in central-west Colombian? S. Afr. J. Bot. 2022, 149, 366–375. [Google Scholar] [CrossRef]
- da Silva, P.C.; Junior, W.Q.R.; Ramos, M.L.G.; Rocha, O.C.; Veiga, A.D.; Silva, N.H.; Brasileiro, L.d.O.; Santana, C.C.; Soares, G.F.; Malaquias, J.V.; et al. Physiological Changes of Arabica Coffee under Different Intensities and Durations of Water Stress in the Brazilian Cerrado. Plants 2022, 11, 2198. [Google Scholar] [CrossRef] [PubMed]
- Arbona, V.; Manzi, M.; De Ollas, C.; Gómez-Cadenas, A. Metabolomics as a Tool to Investigate Abiotic Stress Tolerance in Plants. Int. J. Mol. Sci. 2013, 14, 4885–4911. [Google Scholar] [CrossRef] [PubMed]
- Kumar, M.; Patel, M.K.; Kumar, N.; Bajpai, A.B.; Siddique, K.H.M. Metabolomics and Molecular Approaches Reveal Drought Stress Tolerance in Plants. Int. J. Mol. Sci. 2021, 22, 9108. [Google Scholar] [CrossRef] [PubMed]
- Witt, S.; Luis, G.; Jan, L.; Jill, C.; Axel, T.; Jose, L.; Natalia, P.; Alisdair, R. Metabolic and phenotypic responses of greenhouse-grown maize hybrids to experimentally controlled drought stress. Mol. Plant 2012, 5, 401–417. [Google Scholar] [CrossRef]
- Hochberg, U.; Degu, A.; Toubiana, D.; Gendler, T.; Nikoloski, Z.; Rachmilevitch, S.; Fait, A. Metabolite profiling and network analysis reveal coordinated changes in grapevine water stress response. BMC Plant Biol. 2013, 13, 184. [Google Scholar] [CrossRef]
- Fiehn, O.; Kopka, J.; Dörmann, P.; Altmann, T.; Trethewey, R.N.; Willmitzer, L. Metabolite profiling for plant functional genomics. Nat. Biotechnol. 2000, 18, 1157–1161. [Google Scholar] [CrossRef] [PubMed]
- Guo, R.; Shi, L.; Jiao, Y.; Li, M.; Zhong, X.; Gu, F.; Liu, Q.; Xia, X.; Li, H. Metabolic responses to drought stress in the tissues of drought-tolerant and drought-sensitive wheat genotype seedlings. AoB Plants 2018, 10, ply016. [Google Scholar] [CrossRef] [PubMed]
- Lozano-Elena, F.; Fàbregas, N.; Coleto-Alcudia, V.; Caño-Delgado, A.I. Analysis of metabolic dynamics during drought stress in Arabidopsis plants. Sci. Data 2022, 9, 90. [Google Scholar] [CrossRef] [PubMed]
- Rabara, R.C.; Tripathi, P.; Rushton, P.J. Comparative Metabolome Profile between Tobacco and Soybean Grown under Water-Stressed Conditions. BioMed Res. Int. 2017, 2017, 3065251. [Google Scholar] [CrossRef] [PubMed]
- Drapal, M.; Amah, D.; Schöny, H.; Brown, A.; Swennen, R.; Fraser, P.D. Assessment of metabolic variability and diversity present in leaf, peel and pulp tissue of diploid and triploid Musa spp. Phytochemistry 2020, 176, 112388. [Google Scholar] [CrossRef] [PubMed]
- Obata, T.; Witt, S.; Jan, L.; Natalia, P.R.; Igor, F.; Salima, Y.; Jose, L.; Jill, E.C.; Alisdair, R. Metabolite profiles of maize leaves in drought, heat, and combined stress field trials reveal the relationship between metabolism and grain yield. Plant Physio. 2015, 169, 2665–2683. [Google Scholar] [CrossRef]
- Fàbregas, N.; Fernie, A.R. The metabolic response to drought. J. Exp. Bot. 2019, 70, 1077–1085. [Google Scholar] [CrossRef]
- Impa, S.M.; John, S.; Inga, K.; Raju, B.; Toshihiro, O.; Krishna, J. Carbon balance and source-sink metabolic changes in winter wheat exposed to high night-time temperature. Plant Cell Env. 2019, 42, 1233–1246. [Google Scholar] [CrossRef] [PubMed]
- Roessner, U.; Beckles, D.M. Metabolite Measurements; Springer: New York, NY, USA, 2009; pp. 39–69. [Google Scholar]
- Schwender, J. Plant Metabolic Networks; Junker, B.H., Ed.; Springer: Dordrecht, The Netherlands, 2009. [Google Scholar]
- Lu, Y.; Stegemann, S.; Agrawal, S.; Karcher, D.; Ruf, S.; Bock, R. Horizontal Transfer of a Synthetic Metabolic Pathway between Plant Species. Curr. Biol. 2017, 27, 3034–3041. [Google Scholar] [CrossRef]
- Caine, R.S.; Yin, X.; Sloan, J.; Harrison, E.L.; Mohammed, U.; Fulton, T.; Biswal, A.K.; Dionora, J.; Chater, C.C.; Coe, R.A.; et al. Rice with reduced stomatal density conserves water and has improved drought tolerance under future climate conditions. New Phytol. 2019, 221, 371–384. [Google Scholar] [CrossRef]
- Tesfaye, S. Variations among coffee (Coffea arabica L.) genotypes for sensitivity to drought induced by soil drying at early stages of growth in Ethiopia. J. Plant Breed. Crop Sci. 2018, 5, 453–462. [Google Scholar]
- WCR (World Coffee Research). Good Practice Guide Coffee Nursery Management; WCR publishing: Portland, OR, USA, 2019; pp. 1–80. [Google Scholar]
- Barrs, H.D.; Weatherley, P.E. A Re-Examination of the Relative Turgidity Technique for Estimating Water Deficits in Leaves. Aust. J. Biol. Sci. 1962, 15, 413–428. [Google Scholar] [CrossRef]
- Lichtenthaler, H.K. Chlorophylls and carotenoids, the pigments of photosynthetic biomembranes. Methods Enzy. 1987, 148, 350–382. [Google Scholar] [CrossRef]
- Lisec, J.; Schauer, N.; Kopka, J.; Willmitzer, L.; Fernie, A.R. Gas chromatography mass spectrometry–based metabolite profiling in plants. Nat. Protoc. 2006, 1, 387–396. [Google Scholar] [CrossRef] [PubMed]
- Kopka, J.; Schauer, N.; Krueger, S.; Birkemeyer, C.; Usadel, B.; Bergmuller, E.; Dormann, P.; Weckwerth, W.; Gibon, Y.; Stitt, M.; et al. GMD@CSB.DB: the Golm Metabolome Database. Bioinformatics 2005, 21, 1635–1638. [Google Scholar] [CrossRef]
- Hammer, Ø.; David, A.; Harper, T.; Paul, D.R. PAST: paleontological statistics software package for education and data analysis. Palaeo. Electro. 2001, 4, 9. [Google Scholar]
- Shannon, P.; Markiel, A.; Ozier, O.; Baliga, N.S.; Wang, J.T.; Ramage, D.; Amin, N.; Schwikowski, B.; Ideker, T. Cytoscape: A software environment for integrated models of Biomolecular Interaction Networks. Genome Res. 2003, 13, 2498–2504. [Google Scholar] [CrossRef]
- Konieczna, W.; Warchoł, M.; Mierek-Adamska, A.; Skrzypek, E.; Waligórski, P.; Piernik, A.; Dąbrowska, G.B. Changes in physio-biochemical parameters and expression of metallothioneins in Avena sativa L. in response to drought. Sci. Rep. 2023, 13, 1–16. [Google Scholar] [CrossRef]
- Gugliuzza, G.; Talluto, G.; Martinelli, F.; Farina, V.; Bianco, R.L. Water Deficit Affects the Growth and Leaf Metabolite Composition of Young Loquat Plants. Plants 2020, 9, 274. [Google Scholar] [CrossRef]
- DaMatta, F.M.; Ronchi, C.P.; Maestri, M.; Barros, R.S. Ecophysiology of coffee growth and production. Braz. J. Plant Physiol. 2007, 19, 485–510. [Google Scholar] [CrossRef]
- Wei, Y.; Xu, X.; Tao, H.; Wang, P. Growth performance and physiological response in the halophyte Lyciumbarbarum grown at salt-affected soil. An. Applied Biol. 2006, 149, 263–269. [Google Scholar] [CrossRef]
- Dias, P.C.; Araujo, W.L.; Moraes, G.A.; Barros, R.S.; DaMatta, F.M. Morphological and physiological responses of two coffee progenies to soil water availability. J. Plant Physiol. 2007, 164, 1639–1647. [Google Scholar] [CrossRef]
- Xiong, S.; Wang, Y.; Chen, Y.; Gao, M.; Zhao, Y.; Wu, L. Effects of Drought Stress and Rehydration on Physiological and Biochemical Properties of Four Oak Species in China. Plants 2022, 11, 679. [Google Scholar] [CrossRef] [PubMed]
- Cai, C.T.; Cai, Z.Q.; Yao, T.Q.; Qi, X. Vegetative growth and photosynthesis in coffee plants under different watering and fertilization managements in Yunnan, SW China. Photosynthetica 2007, 45, 455–461. [Google Scholar] [CrossRef]
- Eira, M.T.S.; da Silva, E.A.A.; De Castro, R.D.; Dussert, S.; Walters, C.; Bewley, J.D.; Hilhorst, H.W.M. Coffee seed physiology. Braz. J. Plant Physiol. 2006, 18, 149–163. [Google Scholar] [CrossRef]
- Hayatu, M.; Muhammad, S.Y.; Habibu, U.A. Effect of water stress on the leaf relative water content and yield of some cowpea (Vigna unguiculata (L) Walp.) genotype. Int. J. Sci. Technol. Res. 2014, 3, 148–152. [Google Scholar]
- Pirzad, A.; Shakiba, M.R.; Zehtab-Salmasi, S.; Mohammadi, S.A. Effect of water stress on leaf relative water content, chlorophyll, proline and soluble carbohydrates in Matricaria chamomilla L. J. Medic. Plants Res. 2011, 5, 2483–2488. [Google Scholar]
- Soltys-Kalina, D.; Plich, J.; Strzelczyk-Żyta, D.; Śliwka, J.; Marczewski, W. The effect of drought stress on the leaf relative water content and tuber yield of a half-sib family of ‘Katahdin’-derived potato cultivars. Breed. Sci. 2016, 66, 328–331. [Google Scholar] [CrossRef]
- Wang, Z.; Li, G.; Sun, H.; Ma, L.; Guo, Y.; Zhao, Z.; Gao, H.; Mei, L. Effects of drought stress on photosynthesis and photosynthetic electron transport chain in young apple tree leaves. Biol. Open 2018, 7, bio035279. [Google Scholar] [CrossRef]
- Zhang, A.; Liu, M.; Gu, W. Effect of drought on photosynthesis, total antioxidant capacity, bioactive component accumulation, and the transcriptome of Atractylodeslancea. BMC Plant Biol. 2021, 21, 293. [Google Scholar] [CrossRef]
- Taiz, L.; Zeiger, E. Plant Physiology; Sinauer Associates: Sunderland, MA, USA, 2010; pp. 1–782. [Google Scholar]
- Mafakheri, A.; Siosemardeh, A.F.; Bahramnejad, B.; Struik, P.C.; Sohrabi, Y. Effect of drought stress on yield, proline and chlorophyll contents in three chickpea cultivars. Aus. J. Crop Sci. 2010, 4, 580–585. [Google Scholar] [CrossRef]
- Nikolaeva, M.K.; Maevskaya, S.N.; Shugaev, A.G.; Bukhov, N.G. Effect of drought on chlorophyll content and antioxidant enzyme activities in leaves of three wheat cultivars varying in productivity. Russ. J. Plant Physiol. 2010, 57, 87–95. [Google Scholar] [CrossRef]
- Isah, T. Stress and defense responses in plant secondary metabolites production. Biol. Res. 2019, 52, 39. [Google Scholar] [CrossRef] [PubMed]
- Kapoor, D.; Bhardwaj, S.; Landi, M.; Sharma, A.; Ramakrishnan, M.; Sharma, A. The Impact of Drought in Plant Metabolism: How to Exploit Tolerance Mechanisms to Increase Crop Production. Appl. Sci. 2020, 10, 5692. [Google Scholar] [CrossRef]
- Urano, K.; Maruyama, K.; Ogata, Y.; Morishita, Y.; Takeda, M.; Sakurai, N.; Suzuki, H.; Saito, K.; Shibata, D.; Kobayashi, M.; et al. Characterization of the ABA-regulated global responses to dehydration in Arabidopsis by metabolomics. Plant J. 2009, 57, 1065–1078. [Google Scholar] [CrossRef] [PubMed]
- Krasensky, J.; Jonak, C. Drought, salt, and temperature stress-induced metabolic rearrangements and regulatory networks. J. Exp. Bot. 2012, 63, 1593–1608. [Google Scholar] [CrossRef] [PubMed]
- Fabregas, N.; Lozano-Elena, F.; Blasco-Escamez, D. Over-expression of the vascular brassinosteroid receptor BRL3 confers drought resistance without penalizing plant growth. Nat. Com. 2018, 9, 4680. [Google Scholar] [CrossRef] [PubMed]
- Ogbaga, C.C.; Stepien, P.; Dyson, B.C.; Rattray, N.J.W.; Ellis, D.I.; Goodacre, R.; Johnson, G.N. Biochemical Analyses of Sorghum Varieties Reveal Differential Responses to Drought. PLoS ONE 2016, 11, e0154423. [Google Scholar] [CrossRef] [PubMed]
- Pires, M.V.; Pereira Júnior, A.A.; Medeiros, D.B.; Daloso, D.M.; Pham, P.A.; Barros, K.A.; Engqvist, M.K.; Florian, A.; Krahnert, I.; Maurino, V.G.; et al. The influence of alternative pathways of respiration that utilize branched-chain amino acids following water shortage in Arabidopsis. Plant Cell Environ. 2016, 39, 1304–1319. [Google Scholar] [CrossRef]
- Ahanger, M.A.; Ashraf, M.; Bajguz, A.; Ahmad, P. Brassinosteroids Regulate Growth in Plants Under Stressful Environments and Crosstalk with Other Potential Phytohormones. J. Plant Growth Regul. 2018, 37, 1007–1024. [Google Scholar] [CrossRef]
- Singh, T.N.; Aspinall, D.; Paleg, L.G. Proline Accumulation and Varietal Adaptability to Drought in Barley: a Potential Metabolic Measure of Drought Resistance. Nat. New Biol. 1972, 236, 188–190. [Google Scholar] [CrossRef] [PubMed]
- Szabados, L.; Savouré, A. Proline: a multifunctional amino acid. Trends Plant Sci. 2010, 15, 89–97. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Zhang, Y.; Du, Y.; Chen, S.; Tang, H. Dynamic metabolomic responses of tobacco (Nicotiana tabacum) plants to salt stress. J. Proteome Res. 2011, 10, 1904–1914. [Google Scholar] [CrossRef] [PubMed]
- Joshi, V.; Joung, J.-G.; Fei, Z.; Jander, G. Interdependence of threonine, methionine and isoleucine metabolism in plants: accumulation and transcriptional regulation under abiotic stress. Amino Acids 2010, 39, 933–947. [Google Scholar] [CrossRef] [PubMed]
- Lehmann, S.; Funck, D.; Szabados, L.; Rentsch, D. Proline metabolism and transport in plant development. Amino Acids 2010, 39, 949–962. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Mao, C.; Shi, Z.; Kou, X. The Amino Acid Metabolic and Carbohydrate Metabolic Pathway Play Important Roles during Salt-Stress Response in Tomato. Front. Plant Sci. 2017, 8, 1231. [Google Scholar] [CrossRef] [PubMed]
- Sanchez, D.H.; Pieckenstain, F.L.; Szymanski, J.; Erban, A.; Bromke, M.; Hannah, M.A.; Kraemer, U.; Kopka, J.; Udvardi, M.K. Comparative Functional Genomics of Salt Stress in Related Model and Cultivated Plants Identifies and Overcomes Limitations to Translational Genomics. PLoS ONE 2011, 6, e17094. [Google Scholar] [CrossRef] [PubMed]
- Araujo, W.L.; Nunes-Nesi, A.; Nikoloski, Z.; Sweetlove, L.J.; Fernie, A.R. Metabolic control and regulation of the tricarboxylic acid cycle in pho- tosynthetic and heterotrophic plant tissues. Plant Cell and Envi. 2012, 35, 1–21. [Google Scholar] [CrossRef] [PubMed]
- Rzepka, A.; Rut, G.; Krupa, J. Effect of abiotic stress factors on fluctuations in contents of malate and citrate and on malic enzyme activity in moss gametophores. Photosynthetica 2009, 47, 141–145. [Google Scholar] [CrossRef]
- Yang, L.; Fountain, J.C.; Ji, P.; Ni, X.; Chen, S.; Lee, R.D.; Kemerait, R.C.; Guo, B. Deciphering drought-induced metabolic responses and regulation in developing maize kernels. Plant Biotechnol. J. 2018, 16, 1616–1628. [Google Scholar] [CrossRef]
- Kinnersley, A.M. Gamma aminobutyric acid (GABA) and plant responses to stress. Crit. Revi. Plant Sci. 2000, 19, 479–509. [Google Scholar] [CrossRef]
Figure 1.
Effects of drought stress on shoot length, of the four adult coffee genotypes, Ca754, CaJ-19, Ca74110, and Ca74112, under well water (ww) and drought stress conditions (ws), at different DADB. Bars indicate means±SD (n=10 replicates per genotype). Bars with the same letter indicate no significant difference at p<0.05 between samples. DADB indicates the number of days after drought stress begins.
Figure 1.
Effects of drought stress on shoot length, of the four adult coffee genotypes, Ca754, CaJ-19, Ca74110, and Ca74112, under well water (ww) and drought stress conditions (ws), at different DADB. Bars indicate means±SD (n=10 replicates per genotype). Bars with the same letter indicate no significant difference at p<0.05 between samples. DADB indicates the number of days after drought stress begins.
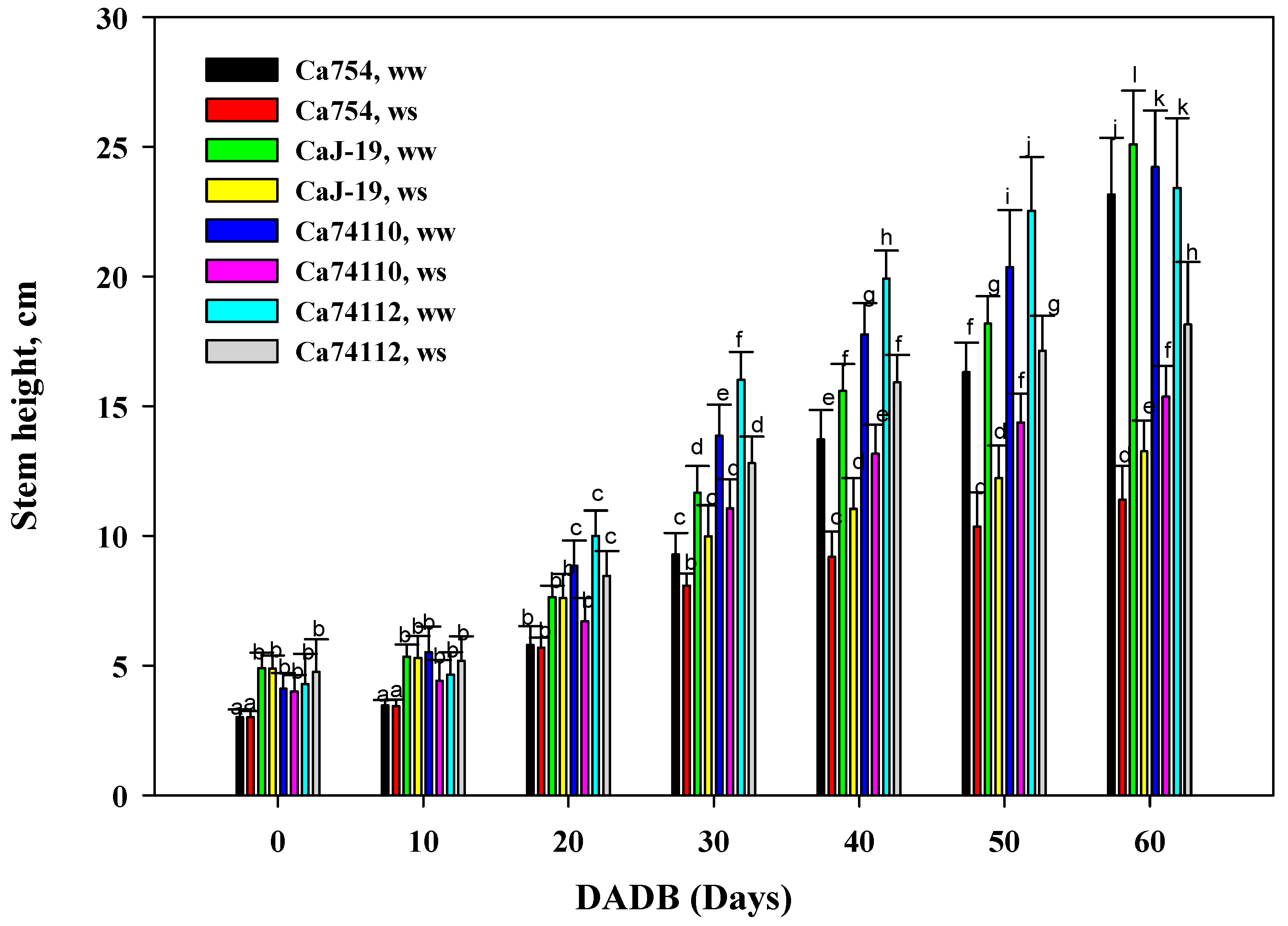
Figure 2.
Effects of drought stress on shoot (A) fresh biomass, and (B) dry biomass of the four adult coffee genotypes, Ca754, CaJ-19, Ca74110, and Ca74112, under well water (ww) and drought stress conditions (ws), 60 days after drought stress begins. Bars indicate means±SD (n=10 replicates per genotype). Bars with the same letter indicate no significant difference at p<0.05 between samples.
Figure 2.
Effects of drought stress on shoot (A) fresh biomass, and (B) dry biomass of the four adult coffee genotypes, Ca754, CaJ-19, Ca74110, and Ca74112, under well water (ww) and drought stress conditions (ws), 60 days after drought stress begins. Bars indicate means±SD (n=10 replicates per genotype). Bars with the same letter indicate no significant difference at p<0.05 between samples.
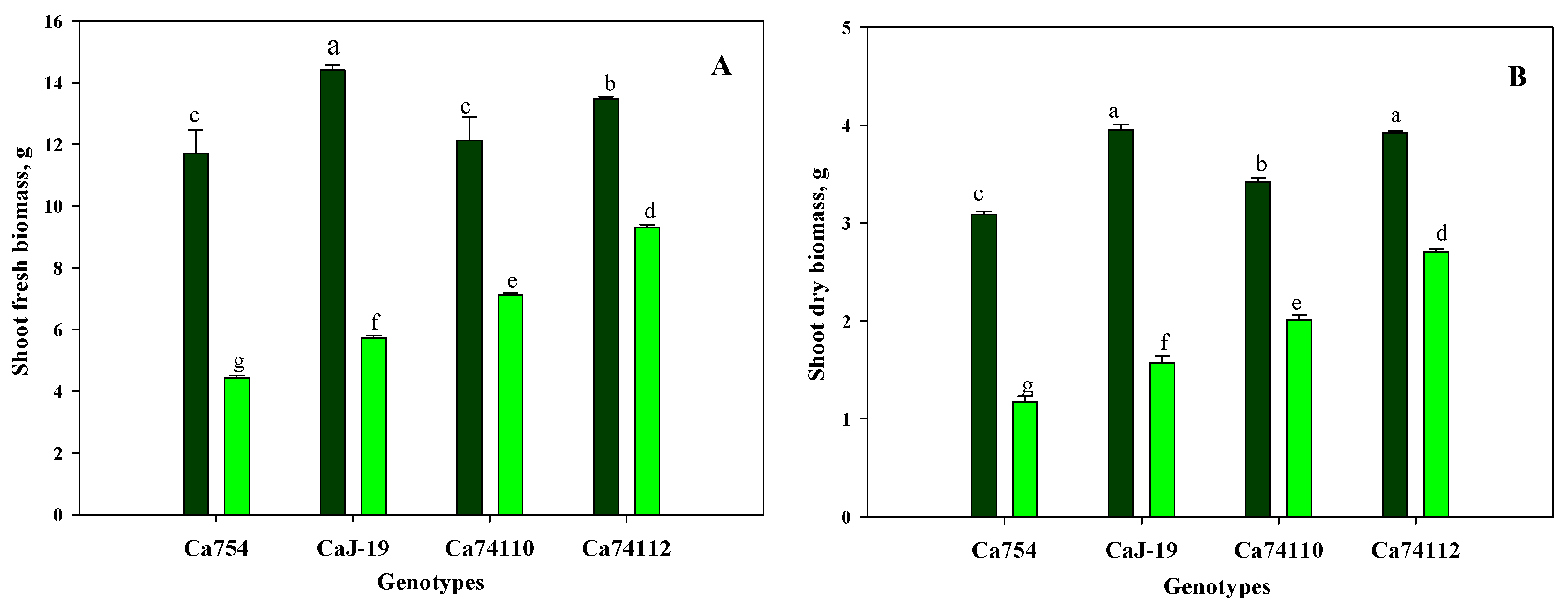
Figure 4.
Effects of drought stress on (A) chlorophyll-a, and (B) chlorophyll-b of the four adult coffee genotypes, Ca754, CaJ-19, Ca74110, and Ca74112, under well water (ww) and drought stress conditions (ws), at 0 and 60 days after drought stress begins. Bars indicate means±SD (n=10 replicates per genotype). Bars with the same letter indicate no significant difference at p<0.05 between samples. DADB indicates the number of days after drought stress begins.
Figure 4.
Effects of drought stress on (A) chlorophyll-a, and (B) chlorophyll-b of the four adult coffee genotypes, Ca754, CaJ-19, Ca74110, and Ca74112, under well water (ww) and drought stress conditions (ws), at 0 and 60 days after drought stress begins. Bars indicate means±SD (n=10 replicates per genotype). Bars with the same letter indicate no significant difference at p<0.05 between samples. DADB indicates the number of days after drought stress begins.
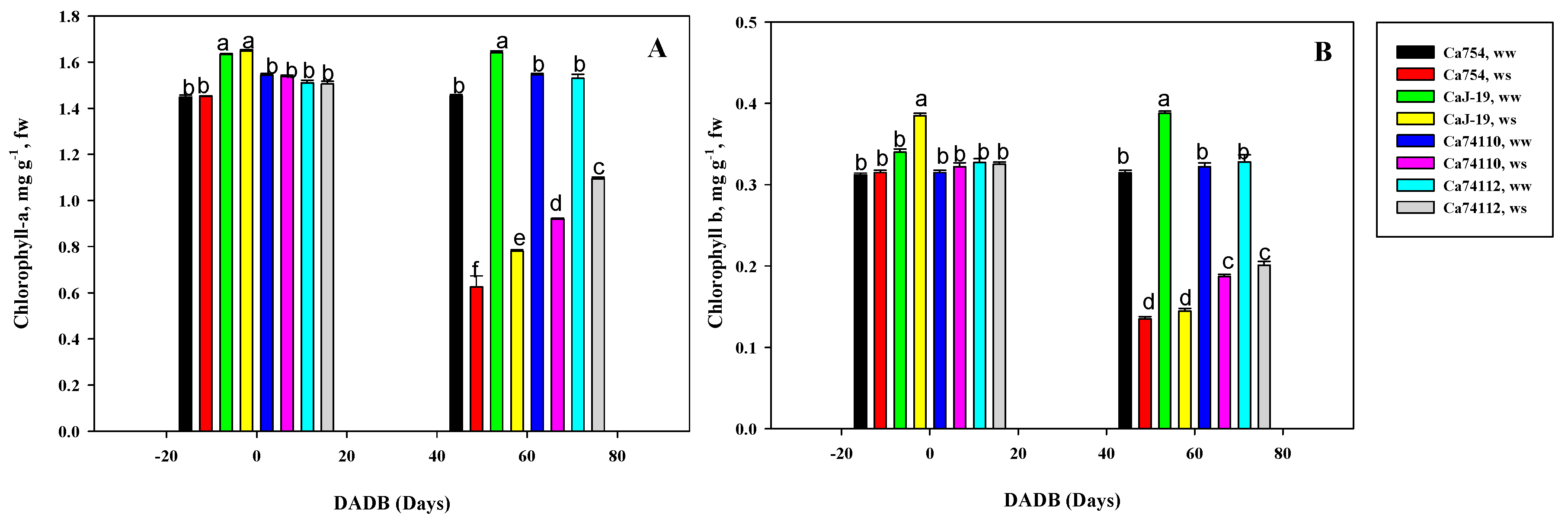
Figure 5.
Metabolic responses to drought stress in leaves of four coffee genotypes, Ca754, CaJ-19, Ca74110, and Ca74112. Values are the logarithmic (Log2) transformed fold changes (ws/ww, drought stressed/well-watered) of selected leaf metabolites at 0 and 60 days after drought stress begins. Bolded figures represent significant difference between ww and ws treatments as tested by the Tukey’s honest test (p<0.05). Green, red, and others represent an increase, decrease, and in between, in terms of metabolite alteration. TL refers time line. (-) indicates that the metabolite is not detected in the respective genotype and/or treatment.
Figure 5.
Metabolic responses to drought stress in leaves of four coffee genotypes, Ca754, CaJ-19, Ca74110, and Ca74112. Values are the logarithmic (Log2) transformed fold changes (ws/ww, drought stressed/well-watered) of selected leaf metabolites at 0 and 60 days after drought stress begins. Bolded figures represent significant difference between ww and ws treatments as tested by the Tukey’s honest test (p<0.05). Green, red, and others represent an increase, decrease, and in between, in terms of metabolite alteration. TL refers time line. (-) indicates that the metabolite is not detected in the respective genotype and/or treatment.
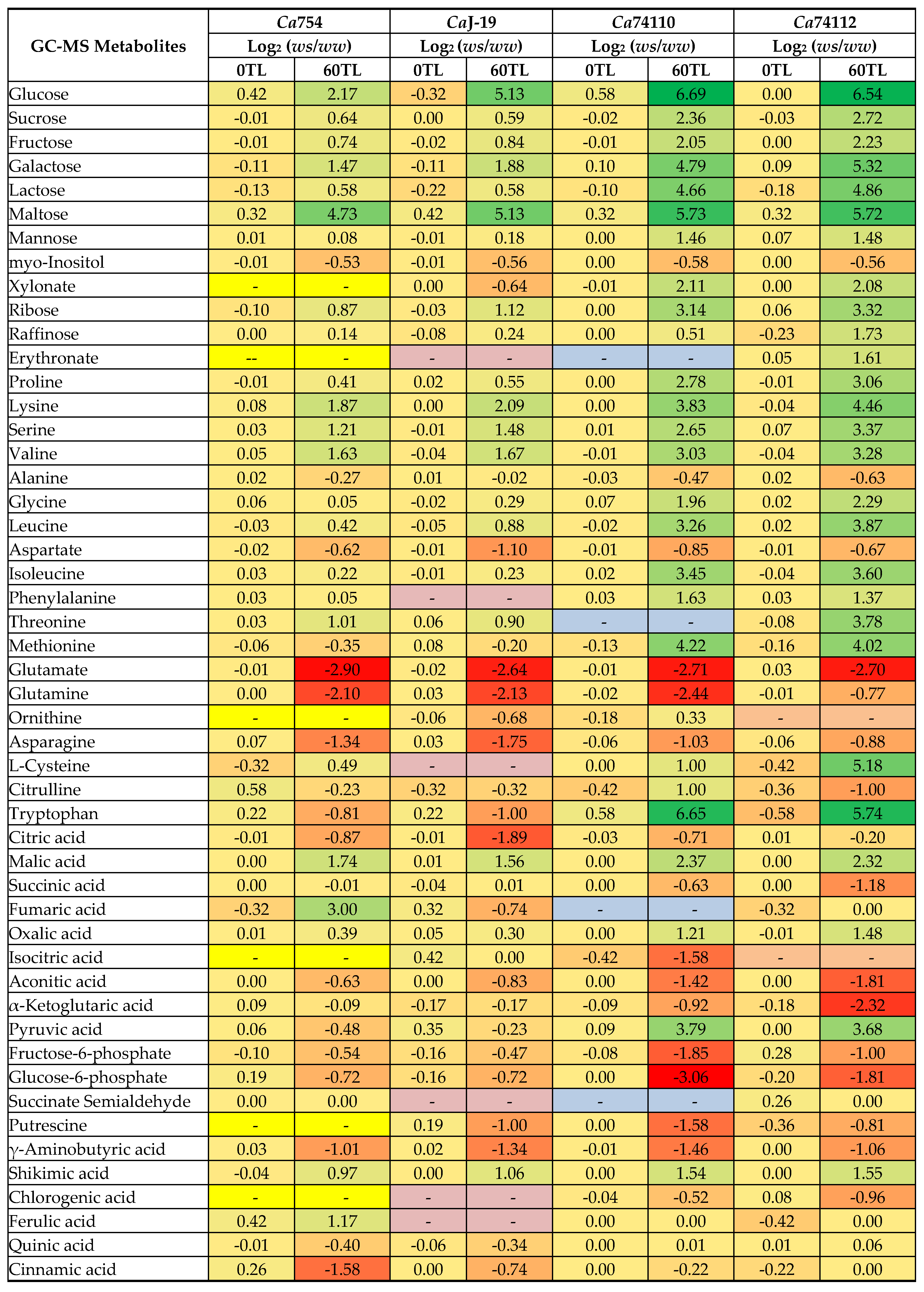
Figure 6.
Changes in Ca754 and CaJ-19 metabolite interactions as a result of drought stress conditions at 0TL and 60-TL. Nodes correspond to metabolites and colors correspond to compound classes as detailed in the figure legend. Edges between nodes represent Pearson correlations at r≥0.8. TL refers time line.
Figure 6.
Changes in Ca754 and CaJ-19 metabolite interactions as a result of drought stress conditions at 0TL and 60-TL. Nodes correspond to metabolites and colors correspond to compound classes as detailed in the figure legend. Edges between nodes represent Pearson correlations at r≥0.8. TL refers time line.
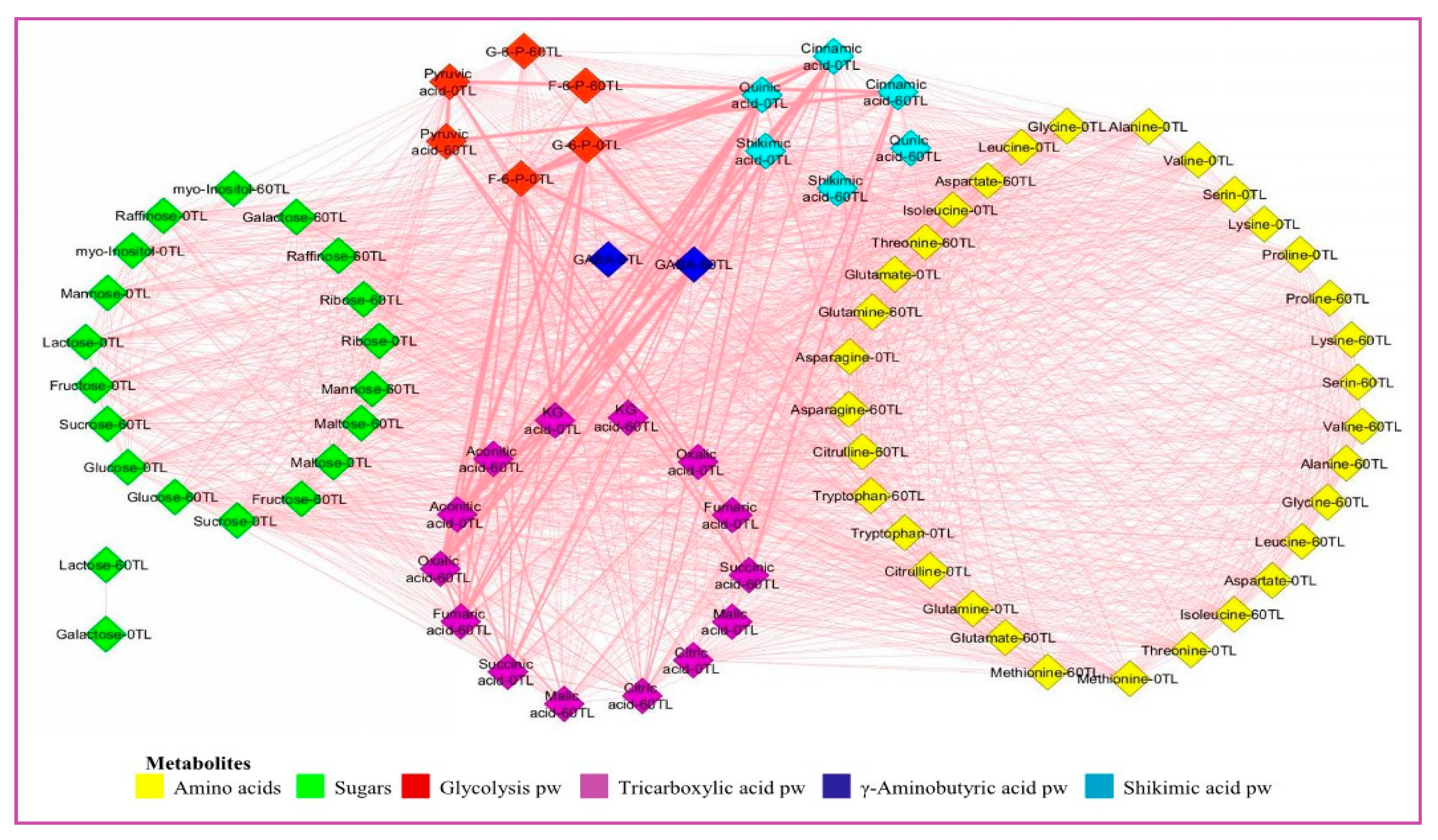
Figure 7.
Changes in Ca74110 and Ca74112 metabolite interactions because of drought stress conditions at 0TL and 60-TL. Nodes correspond to metabolites and colors correspond to compound classes as detailed in the figure legend. Edges between nodes represent Pearson correlations at r≥0.8. TL refers time line.
Figure 7.
Changes in Ca74110 and Ca74112 metabolite interactions because of drought stress conditions at 0TL and 60-TL. Nodes correspond to metabolites and colors correspond to compound classes as detailed in the figure legend. Edges between nodes represent Pearson correlations at r≥0.8. TL refers time line.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



