
Submitted:
14 December 2023
Posted:
21 December 2023
You are already at the latest version
Abstract
Brown leaf spot disease caused by Alternaria spp. is among the most common diseases in potato crops. Typical brown spot symptoms were observed in commercial potato-cultivated areas of northern Korea during June 2021–2022. In total, 77 isolates were collected, and based on sequence analysis of the internal transcribed spacer (ITS) region, the collected isolates were identified as Alternaria spp. (83.1%). Phylogenetic analysis revealed that a majority of these isolates clustered within a clade that included A. alternata. Additionally, the ITS region and rpb2 yielded the most informative sequences for the identification of A. alternata. Pathogenicity tests confirmed that the collected pathogens elicited identical symptoms in the field. In the pathogenicity test performed on seven commercial cultivars, the pathogens exhibited strong virulence in both wound and non-wound inoculations. Among the cultivars tested, Arirang-1ho, Arirang-2ho, and Golden Ball were resistant to the pathogens. Furthermore, when testing effective fungicides in vitro, mancozeb and difenoconazole were found to be effective agents for inhibiting mycelial growth. In summary, our findings suggest that A. alternata plays a critical role in leaf disease in potato crop regions and emphasise the necessity for continuous monitoring and management to protect against this disease in South Korea.
Keywords:
Alternaria alternata
; brown spot disease
; pathogenicity test
; phylogenetic analysis
; Solanum tuberosum L.
1. Introduction
Potatoes (Solanum tuberosum L.) are a staple non-cereal food crop and the fourth most productive crop after maize, wheat, and rice worldwide [1]. Various pathogenic infections threaten the potato crop and can lead to poor quality and reduced yield. These include late blight caused by Phytophthora infestans, early blight caused by Alternaria solani, and brown leaf spot disease caused by Alternaria alternata [2]. In a previous study conducted in Korea, more than 60% of potato leaves exhibited brown leaf spots caused by A. alternata infection [3].
Unlike early blight, potato brown leaf spot disease caused by A. alternata manifests as small, irregular, circular spots, with sizes ranging from pinpoint to 12 mm, on the lower leaves that turn into dark brown spots without concentric rings [4]. This disease occurs throughout the potato-growing season when environmental conditions are favourable for the pathogen. As the disease progresses, the entire leaf becomes chlorotic and turns brown, and the leaf edges curl up, similar to early blight. Eventually, the infected leaves wither and die, leaving the wilted leaves hanging from the potato plant. Dorby et al. (1984) reported that yields could be reduced by as much as 18% under conditions of high relative humidity and temperature, high pathogen density, and susceptible hosts [5].
This disease has been relatively underresearched in comparison to the investigations on early and late blight in potato. However, a brown spot disease caused by A. alternata has been recently reported in several countries [1,3,6,7,8]. Previous studies have reported that A. arborescens, A. tenuissima, A. tomatophila, A. grandis, A. solani, and A. alternata are the causal pathogens of brown leaf blight symptoms in potatoes [1,9,10,11].
The genus Alternaria is difficult to identify based solely on morphology, and employing the commonly used internal transcribed spacer (ITS) region for sequence-based identification remains challenging. Phylogenetic analysis using concatenated sequences offers a potential solution for resolving fungi species classification within the genus [11,12]. Woudenberg et al. (2015) generated sequences from seven loci: the ITS region, glyceraldehyde-3-phosphate dehydrogenase (gapdh), translation elongation factor 1-alpha (tef1), RNA polymerase second-largest subunit (rpb2), Alternaria major allergen gene (Alt a 1), endopolygalacturonase (endoPG), and OPA10-1 [11]. Therefore, employing methods beyond morphological and phylogenetic analyses, focusing on these seven loci, would aid in the definitive identification of Alternaria species.
Identifying the disease pathogen is the first step towards its control. In a previous study, we reported the causal pathogen, A. alternata, responsible for brown leaf spot symptoms observed during the harvest season in the potato cultivation areas of Yeoncheon, Gyeonggi Province, Korea [3]. In this study, we employed a comprehensive set of sequences involving seven loci, namely, ITS, gapdh, tef1, rpb2, Alt a1, endoPG, and OPA10-1, and generated a phylogenetic tree, consequently identifying the causal pathogen as A. alternata [3]. Developing resistant potato cultivars stands as the most efficient strategy for effective control. However, similar to potato early blight, no resistant cultivars against potato brown leaf spot have been reported to date [13]. Therefore, implementing efficient fungicide application will help mitigate the reduction in potato yield caused by this pathogen [14].
Consequently, the objectives of this study were: (i) to identify the predominant causal pathogen responsible for leaf blight symptoms, including leaf spots, in leaves collected from Yeoncheon, Baengnyeongdo, and Goseong between 2020 and 2021 via morphological and molecular analyses and, if dominant pathogens were determined; (ii) to analyse the differences in the representatives of the dominant species via phylogenetic analysis; (iii) to examine their pathogenicity on different potato cultivars; and (iv) to identify effective fungicides. In this study, we identified A. alternata as the primary pathogen responsible for leaf spot and blight symptoms in samples collected from the three regions between 2020 and 2021. Furthermore, we performed a phylogenetic analysis using seven loci and found that the ITS and RPB2 loci served as the primary informative sequences for distinguishing A. alternata.
2. Materials and Methods
2.1. Isolation of Fungal Isolates
Potato leaves that developed symptoms were collected from three different regions from June to August 2020–2021. Infected tissues (5 ×5 mm) from the diseased leaf samples were immersed in 70% ethanol for 1 min, rinsed three times in sterilised water, dried, placed on water agar amended with 100 ppm of streptomycin, and then incubated in the dark at 25 °C for 3–7 days. After hyphae emerged from the tissues, the fungal isolates were transferred onto potato dextrose agar (PDA; Difco Laboratories, Detroit, MI, USA) or V8-Juice agar medium (8% V8-Juice, 1.5% agar, pH 7). All fungal strains were stored at 4°C in sterile distilled water or at -80°C in 15% glycerol in agar blocks on which the fungi were grown for long-term storage until the experiment.
2.2. Fungal Cultures and DNA Extraction
All the collected isolates were subjected to DNA extraction. Fungal isolates were grown in 5 mL of potato dextrose broth (Difco Laboratories) at 25°C for 7 days. Genomic DNA was isolated as described previously [15] or purified using the NucleoSpin Plant kit (Macherey-Nagel, Dűren, Germany) according to the manufacturer’s instructions. The DNA concentration was estimated using a NanoDrop ND-1000 spectrophotometer (NanoDrop Technologies, Inc. Wilmington, USA). DNA concentration was adjusted to 12.5 ng/μL for each isolate and subjected to PCR amplification.
2.3. PCR and Sequencing
PCR was performed using an ABI 2720 Thermal Cycler (Applied Biosystems, Foster City, CA, USA). PCR amplification was performed with 25 ng of genomic DNA and 2 pmol/L of each primer (Table 1) using the i-StarMAX II PCR master mix (iNtRON Biotechnology Inc., Seongnam, Korea). The amplification conditions were as follows: (a) initial denaturation at 96°C for 1 min; (b) 2 cycles of denaturation at 94°C for 1 min, annealing at 52 °C for 1 min, and elongation at 72°C for 1 min; (c) 28 cycles of denaturation at 94°C for 30 sec, annealing at 55°C for 30 sec, and elongation at 72°C for 1 min; and (d) elongation at 72°C for 3 min.
PCR products were resolved via 0.8% agarose gel electrophoresis and bidirectionally sequenced by the Bioneer sequencing service (Bioneer Inc., Daejeon, Korea) on both strands with the same primers used for PCR amplification. Sequence assembly was performed using the SeqMan program DNAStar (Madison, WI, USA) and CodonCode Aligner V3.5.4. software (CodonCode Co., USA). The aligned sequences were subjected to a BLASTn search in the GenBank database (http://www.ncbi.nlm.nih.gov/BLAST/). All the generated sequences were deposited in GenBank (Supplementary Table S1).
2.4. Phylogenetic Analysis
Phylogenetic analysis was performed on A. alternata isolates, and the dominant group was identified using ITS region sequencing. Multiple sequence alignments of the concatenated sequences were generated using ClustalX [24] and manually adjusted. Sequence divergence was estimated using the MEGA computer package version 11 [25] and the Tamura-Nei model of evolution [26]. Phylogenetic analyses of the sequence data consisted of a maximum likelihood analysis of both the individual data partitions and the combined dataset.
2.5. Pathogenicity Test
To test for pathogenicity, 1-month-old potato plants (S. tuberosum cultivar (cv.) Superior) grown in a 25°C growth chamber were sprayed with a conidial suspension (1×106 conidia/mL) containing 250 ppm Tween 20 prepared from 7 to 14-day-old cultures of the selected Alternaria spp. isolates. Sterile, distilled water was used as the control. The inoculated plants were placed in a plastic box (50 × 40 × 45 cm) to maintain high humidity and incubated in the dark at 25°C for 1 day. The box was transferred to a growth chamber, and the plants were grown under a 16-h photoperiod, fluorescent lighting, and maintained at a temperature of 25°C and >70% humidity. Disease severity was measured 7 days after inoculation. The assay was performed in triplicate.
2.6. Virulence Test on Commercial Cultivars
Seven commercial potato cultivars, Arirang-1ho, Arirang-2ho, Golden Ball, Daekwang, Daeji, Superior, and Chubaek, were obtained from the Highland Agriculture Research Institute, National Institute of Crop Science, Rural Development Administration, Korea. The cultivars were grown in a greenhouse (23–30°C). For large-scale screening, a detached leaf assay was performed using leaves from 45-day-old plants from all seven potato cultivars.
Healthy leaves were placed in a plastic box and maintained in a watered state using cotton. Then, a single leaf was inflicted with wounds 10 times at each of the three inoculation sites using a micro pin. The inoculum was prepared with 6-mm agar plugs from the 7-day-old A. alternata culture in a V8 juice agar medium. For inoculation, the mycelial agar plugs were placed upside down onto the detached leaves.
Following 1 day-post inoculation (dpi) in the dark at 25°C, the inoculum was removed from the infection sites, and the containers were incubated in a growth chamber (16 h light with >70% humidity and at 25°C). The symptoms were observed at 7 dpi. An uninoculated V8-juice agar plug was used as a control. All the experiments were performed twice. Disease leaf area (DLA) was calculated by measuring the area of observed symptoms relative to the total observed area using the ImageJ software 1.48 version [27]. The formula used was as follows:
Disease leaf area (%)= (symptom-developed area within the total observed area/total observed area) × 100.
2.7. In Vitro Screening of Fungicide Sensitivity
The ability of the fungicides to inhibit the radial growth of the A. alternata isolates was assayed. To select a suitable fungicide for the control of A. alternata isolates, different fungicides with varied mechanisms of action were selected: mancozeb and chlorothalonil (exhibiting multi-site contact activity); difenoconazole (inhibiting 14α-sterol demethylase, categorised as DMI-fungicides); boscalid and pydiflumetofen (succinate dehydrogenase inhibitors, SDHI fungicide); kresoxim-methyl (a quinone outside inhibitor, categorised as a QoI fungicide); and thiophanate-methyl (inhibiting tubulin polymerisation, classified as an MBC fungicide) (Table 2).
After the A. alternata isolates were selected, the ability of the fungicides to inhibit the radial growth of these isolates was evaluated using the agar dilution method [28]. The selected fungicide was added to the PDA medium before solidification, and the mycelial plug from the edge of the hyphae was cultured for 7 days in the PDA medium using a 6-mm cork borer. The mycelial plugs were inoculated into the fungicide medium.
After 7 days of incubation at 25°C, the radial growth in both the control (C, PDA) and treated (T, PDA amended with fungicide) sets was measured. The percentages of radial growth inhibition (I) and corrected inhibition (IC) were calculated as previously described [29]. In brief, two formulae were used:
where C is the diameter of the fungal colony from the selected isolates on the PDA plate, T is the diameter of the colony on the treated plates, and C0 is the diameter of the primary fungal mycelial disc (6 mm).
I (%) = [(C-T)/C] × 100
IC (%) = [(C-T)/(C-C0)] × 100
2.8. Statistical Analysis
The results of the study were presented as the mean ± standard deviation. Statistical analysis was performed using one-way analysis of variance in SPSS (Ver. 20.0, SPSS Inc.). Duncan’s multiple range test was used to determine significance at the 95% probability level.
3. Results
3.1. Collection of Fungal Isolates from Potatoes with Brown Leaf Spot Disease
We investigated the occurrence of diseases affecting potatoes cultivated in three northern regions (Yeoncheon, Goseong, and Baengnyeongdo) of South Korea from June and July 2020 to 2021 (Table 3). Brown leaf spot disease was observed in 2020 and 2021, with a particularly severe outbreak in Yeoncheon in 2021 (Figure 1a). The molecular identification of the isolates was performed by analysing their morphological characteristics and ITS region sequences, cross-referencing the obtained ITS sequences with the results of the NCBI BLAST search (Table 3).
During the first round of sampling (2020), we collected 33 isolates (84.6%) of Alternaria spp. and six isolates (15.4%) of Fusarium spp. from the entire collection area (Figure 1d, left). In the second round (in 2021), we collected 31 isolates (81.6%) of Alternaria spp., 2 isolates (5.3%) of Fusarium spp., 2 isolates (5.3%) of Boeremia spp., 2 isolates (5.3%) of Stagonosporopsis spp., and 1 isolate (2.6%) of Colletotrichum sp. from the same collection area (Figure 1d, centre). Altogether, the 77 isolates collected across the 2-year period comprised Alternaria spp. (64 isolates), Fusarium spp. (8 isolates), Boeremia spp. (2 isolates), Stagonosporopsis spp. (2 isolates), and Colletotrichum sp. (1 isolate) (Figure 1d, right).
3.2. Phylogenetic Analysis of Alternaria spp. Isolates Using Seven Barcoding Genes
Given that members of the Alternaria genus were the predominant fungal species throughout the study period, we obtained nucleotide sequence information for seven barcoding genes: ITS, gapdh, tef1, rpb2, Alt a 1, endoPG, and OPA10-2, for species-level identification of the 55 selected Alternaria spp. isolates. Single gene sequences (ITS (Supplementary Figure S1), gapdh (Supplementary Figure S2), tef1 (Supplementary Figure S3), rpb2 (Supplementary Figure S4), Alt a 1 (Supplementary Figure S5), endoPG (Supplementary Figure S6), and OPA10-2 (Supplementary Figure S7)). Additionally, a concatenated multigene phylogeny encompassing seven genes was generated (Figure 2). The multigene phylogenetic tree revealed that 51 isolates corresponded to A. alternata, while the remaining four isolates comprised two strains of A. arborescens and two strains of A. solani (Figure 2). Notably, the phylogenetic tree generated from rpb2 sequences showed similar clustering (Supplemental Figure S4) to the concatenated seven-gene phylogenetic tree; two isolates (SYP-F0352 and SYP-F035) clustered with the type strain A. solani CBS109157, and two isolates (SYP-F0713 and SYP-F0714) were associated with A. arborescens CBS 102605 (Supplemental Figure S4). However, in the remaining single-gene tree, isolates of A. alternata, A. arborescens, and A. solani could not be clearly distinguished.
3.3. Pathogenicity Test
Infections caused by Alternaria spp. involve the direct invasion of host plants through the stomata and/or wounds [30]. To investigate the possibility of infection through stomata, we inoculated conidia onto the entire unwounded surface of the Superior potato cultivar. Based on the results of the phylogenetic analysis, the isolates were clustered into three distinct groups, and we selected isolates from each of these the three groups for virulence testing (Figure 2): six isolates (SYP-F0939, SYP-F0942, SYP-F0934, SYP-F0941, SYP-F0944, and SYP-F0946) from Group I, one isolate (SYP-F0936) from Group II, and four isolates (SYP-F0935, SYP-F0940, SYP-F0943, and SYP-F0945) from Group III.
Disease symptoms were first observed 3 days post-inoculation (dpi) in most isolates, and severe necrotic brown spot symptoms were observed after 7 dpi. Necrosis progressed from the outer edges to the inner regions of the leaves, causing them to turn black and wilt (Figure 3). These symptoms are similar to those observed in the field (Figure 1). In particular, two strains classified in Group I, SYP-F0939 and SYP-F0942, caused severe disease, leading to the death of all potato seedlings. No symptoms were observed in the control seedlings. The isolates retrieved from all diseased leaves were confirmed to be A. alternata based on their sequences at the rpb2 locus, establishing A. alternata as the causative agent of this disease.
3.4. Virulence Test on Commercial Cultivars
Typical symptoms of Alternaria spp. infection were observed in both wounded and non-wounded leaves within 7 days of the detached leaf assay. These symptoms were similar to those observed in the field. The results of the virulence test for all seven cultivars indicated that the disease incidence depended on both potato cultivars and A. alternata isolates in wounded and non-wounded inoculations. The control showed no symptoms on either wounded or non-wounded leaves (Figure 4a).
Potato cv. Chubaek: Typical or acute progressive symptoms included brownish-black lesions, leaf discoloration, and purple coloration in response to some isolates (Figure 4a). Among the seven cultivars tested, the abundance of Chubaek was relatively high. The DLA rates for all isolates (12, 2, and 12 isolates selected from A. alternata groups I, II, and III, respectively, see Figure 2) were 33.2% and 52.2% with non-wounded (Figure 4b) and wounded (Figure 4c) inoculations, respectively. These results indicate the heightened susceptibility of the cv. Chubaek to the A. alternata isolates obtained from the potato fields, among the seven cultivars.
The potato cvs. Daekwang, Daeji, and Superior exhibited typical brownish-black lesions and halo formations (Figure 4a). The DLA rates for all isolates were 16.4%, 17.9%, and 19.3% for non-wounded inoculations and 19.6%, 31.0%, and 33.0%, respectively (Figure 4b).
The incidence of disease was the lowest in potato cvs. Arirang-1ho, Arirang-2ho, and Golden Ball when inoculated with the entire set of isolates (Figure 4). The DLA rates of cvs. Arirang-1ho, Arirang-2ho, and Golden Ball were 1.2%, 2.2%, and 3.0%, respectively, for non-wounded inoculations (Figure 4a,b). For wounded inoculations (Figure 4c), the values were 11.5%, 12.3%, and 9.1%, respectively, indicating resistance to the disease. Consistent re-isolation of the pathogen from the symptoms in most cultivars confirmed A. alternata as the causative agent responsible for the observed symptoms.
3.5. Selection of Appropriate Fungicides for Potato Brown Spot Disease in Korea
In total, 10 isolates were used for the fungicide selection experiment, with two selected from each of the three groups (I, II, and III) of A. alternata and two isolates each from A. arborescens and A. solani (Figure 2). The fungicides mancozeb and difenoconazole effectively controlled the mycelial growth of all isolates, with control rates ranging from 100% to 73.1% (Figure 5, Table 4). Pydiflumetofen and boscalid exhibited lower, yet significant, inhibitory effects on mycelial growth than mancozeb and difenoconazole. In contrast, chlorothalonil, kresoxim-methyl, and thiophanate-methyl had limited effectiveness, controlling mycelial growth rates ranging from a maximum of 56% to a minimum of 3.2% (Figure 5; Table 4).
4. Discussion
This study aimed to determine the causal pathogen responsible for potato brown leaf spot in the three northern regions of South Korea between June and July in 2020–2021, identify resistant potato cultivars against this pathogen, and select the most effective commercially available fungicides for controlling this issue in Korean potato crops.
Our results strongly suggest that A. alternata is the major pathogen responsible for brown leaf spot disease in these regions. Although A. alternata has long been reported as a major pathogen in potato cultivation areas in Europe [31,32], the United States [6,10,13], China [1], Israel [5], South Africa [33], and Russia [34], it was only first reported in Korea in 2023 [3]. However, subsequent research on the distribution, the presence of resistant varieties, and the selection of effective control agents against A. alternata in South Korea is lacking.
The phylogenetic analysis revealed genetic disparities, showing three distinct clusters—groups I, II, and III—among A. alternata isolates, primarily distinguished based on the ITS region and the rpb2 gene. In addition, the rpb2 gene sequences were found to be highly effective in identifying strains of A. arborescens, A. solani, and A. alternata. Although previous studies have highlighted the applicability of employing a combination of various gene sequences in phylogenetic analysis for distinguishing Alternaria spp. [1,2,10,13,14,32], our study accurately identified A. alternata using a range of gene sequences including the ITS region, gapdh, tef1, rpb2, Alt a1, endoPG, and OPA 10-1. We believe that more extensive genetic information can offer deeper insights. However, despite the genotype-based clustering into groups I, II, and III, these differences did not translate to notable differences in phenotype. Significant differences were not noted in pathogenicity, virulence across the seven cultivars, or sensitivity to fungicides among these genotype-based groups. This suggests that genetic distinctions might not hold significant influence.
Notably, all 10 selected isolates exhibited resistance to the QoI fungicide, kresoxim-methyl (Figure 5; Table 4). Therefore, testing for resistance to other QoI fungicides, such as azoxystrobin, is warranted, owing to the widespread use of QoIs in commercial potato fields in Korea, which indicates a high probability of resistance development in Alternaria spp. Dube et al. (2014) reported the presence of azoxystrobin-resistant A. alternata isolates, which are commonly used to control early blight [33]. These isolates harboured a single-point mutation in the cyt b gene of the mitochondrial genome, resulting in an amino acid substitution from glycine to alanine at position 143 (G143A) in A. alternata [33], which highlights the feasibility of confirming resistance to QoI-class pesticides through PCR amplification and sequencing. This emphasises the importance of continuous monitoring and management of resistant strains of Alternaria spp. to implement effective fungicide application strategies.
5. Conclusions
This study identified A. alternata, A. arborescens, and A. solani as the causative agents of brown leaf spot disease in potatoes in South Korea through morphological and phylogenetic analyses. Among these, A. alternata was found to be the major pathogen. Pathogenicity tests confirmed that all the selected isolates exhibited the same symptoms as those observed in the field. When inoculated with and without wounds, the commercial cultivars Arirang-1ho, Arirang-2ho, and Golden Ball exhibited resistance, Daekwang, Daeji, and Superior demonstrated moderate resistance, and Chubaek was found to be susceptible. In vitro screening identified mancozeb and difenoconazole as the most effective fungicides for inhibiting fungal growth, making them the most suitable fungicide options. Our study revealed that brown spot disease caused by A. alternata, which has been previously reported in various countries, is a potential major disease affecting potato production in Korea. Further research is required to investigate the genetic diversity and environmental factors influencing the occurrence of this disease.
Supplementary Materials
The following supporting information can be downloaded at: www.mdpi.com/xxx/s1, Phylogenetic tree constructed based on the ITS region (Figure S1), gapdh (Figure S2), tef1 (Figure S3), rpb2 (Figure S4), Alt a1 (Figure S5), endoPG (Figure S6), OPA 10-2 (Figure S7) sequence of the Alternaria spp.; Table S1: Isolates used in this study and their GenBank accession numbers.
Author Contributions
Conceptualization, J.P., J.C., Y.K. (Yangseon Kim) and S.-Y.P.; methodology, J.P., S.K. and S.-Y.P.; validation, J.P., S.K. and S.-Y.P.; formal analysis, J.P., S.K., M.J., S.A., M.-H.J., and S.-Y.P.; investigation, J.P., S.K., M.J., S.A., Y.K. (Youngjun Kim), J.Y., M.-H.J.; resources, J.P., and S.-Y.P.; data curation, J.P., S.K., M.J., S.A., M.-H.J., and S.-Y.P.; writing—original draft preparation, J.P. and S.-Y.P.; writing—review and editing, S.-Y.P.; visualization, J.P., M.-H.J., and S.-Y.P.; supervision, S.-Y.P.; project administration, E.Y.K., and S.-Y.P.; funding acquisition, E.Y.K.
Funding
Please add: This work was supported by a grant from the Rural Development Administration (PJ0152782023).
Institutional Review Board Statement
Not applicable.
Data Availability Statement
The data that support the findings of this study are available in the Supplementary Materials of this article.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Zheng:, H.H.; Zhao, J.; Wang, T.Y.; Wu, X.H. Characterization of Alternaria species associated with potato foliar disease in China. Plant Pathol. 2016, 64, 425–433. [Google Scholar] [CrossRef]
- van der Waals, J.E.; Korsten, L.; Slippers, B. Genetic diversity among Alternaria solani isolated from potatoes in South Africa. Plant Dis. 2004, 88, 959–964. [Google Scholar] [CrossRef] [PubMed]
- Choi, J.; Jeung, M.-H.; Choi, E.D.; Park, J.; Park, S.-Y. First report of brown spot caused by Alternaria alternata on potato (Solanum tuberosum) in Korea. Plant Dis. 2023, 107, 2253. [Google Scholar] [CrossRef] [PubMed]
- Stevenson, W.R.; Loria, R.; Franc, G.D.; Weingartner, D.P. Compendium of potato diseases, Second Edition; The American Phytopathological Society: St, Paul, MN, USA, 2001. [Google Scholar]
- Dorby, S.; Dinoor, A.; Prusky, D.; Barkaigolan, R. Pathogenicity of Alternaria alternata on potato in Israel. Phytopathology 1984, 74, 537–542. [Google Scholar] [CrossRef]
- Fairchild, K.L.; Miles, T.D.; Wharton, P.S. Assessing fungicide resistance in population of Alternaria in Idaho potato fields. Crop Prot. 2013, 49, 31–39. [Google Scholar] [CrossRef]
- Soleimani, M.J.; Kirk, W. Enhance resistance to Alternaria alternata causing pototo brown leaf spot disease by using some plant defense inducers. J. Plant Prot. Res. 2012, 52, 83–90. [Google Scholar] [CrossRef]
- Thomma, B.P. Alternaria spp.: From general saprophyte to specific parasite. Mol. Plant Pathol. 2003, 4, 225–236. [Google Scholar] [CrossRef]
- Rodrigues, T.T.M.S.; Berbee, M.L.; Simmons, E.G.; Cardoso, C.R.; Reis, A.; Maffia, L.A.; Mizubuti, E.S.G. First report of Alternaria tomatophila and A. grandis causing early blight on tomato and potato in Brazil. New Des. Rep. 2010, 22, 28. [Google Scholar] [CrossRef]
- Tymon, L.S.; Cummings, T.F.; Johnson, D.A. Identification and aggressiveness of three Alternaria spp. on potato foliage in the US Northwest. Plant Dis. 2016, 100. [Google Scholar] [CrossRef]
- Woudenberg, J.H.; Seidl, M.F.; Groenewald, J.Z.; de Vries, M.; Stielow, J.B.; Thomma, B.P.; Crous, P.W. Alternaria section Alternaria: Species, formae speciales or pathotypes? Stud. Mycol. 2015, 82, 1–21. [Google Scholar] [CrossRef]
- Woudenberg, J.H.; Truter, M.; Groenewald, J.Z.; Crous, P.W. Large-spored Alternaria pathogens in section Porri disentangled. Stud. Mycol. 2014, 79, 1–47. [Google Scholar] [CrossRef] [PubMed]
- Ding, S.; Meinholz, K.; Cleveland, K.; Jordan, S.A.; Gevens, A.J. Diversity and virulence of Alternaria spp. causing potato early blight and brown spot in Wisconsin. Phytopathology 2018, 109, 436–445. [Google Scholar] [CrossRef] [PubMed]
- Meng, J.W.; Zhu, W.; He, M.H.; Wu, E.J.; Duan, G.H.; Xie, Y.K.; Jin, Y.J.; Yang, L.N.; Shang, L.P.; Zhan, J. Population genetic analysis reveals cryptic sex in the phytopathogenic fungus Alternaria alternata. Sci. Rep. 2015, 5, 18250. [Google Scholar] [CrossRef] [PubMed]
- Chi, M.-H.; Park, S.-Y.; Lee, Y.-H. A quick and safe method for fungal DNA extraction. Plant Pathol. J. 2009, 25, 108–111. [Google Scholar] [CrossRef]
- de Hoog, G.S.; Gerrits van den Ende, A.H. Molecular diagnostics of clinical strains of filamentous Basidiomycetes. Mycoses. 1998, 41, 183–189. [Google Scholar] [CrossRef] [PubMed]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics; Academic Press: New York, 1990; Volume 18, pp. 315–322. [Google Scholar]
- Berbee, M.L.; Pirseyedi, M.; Hubbard, S. Cochliobolus phylogenetics and the origin of known, highly virulent pathogens, inferred from ITS and glyceraldehyde-3-phosphate dehydrogenase gene sequences. Mycologia 1999, 91, 964–977. [Google Scholar] [CrossRef]
- Carbone, I.; Kohn, L.M. A method for designing primer sets for speciation studies in filamentous ascomycetes. Mycologia 1999, 91, 553–556. [Google Scholar] [CrossRef]
- O'Donnell, K.; Kistler, H.C.; Cigelnik, E.; Ploetz, R.C. Multiple evolutionary origins of the fungus causing Panama disease of banana: Concordant evidence from nuclear and mitochondrial gene genealogies. Proc. Natl. Acad. Sci. U S A 1998, 95, 2044–2049. [Google Scholar] [CrossRef] [PubMed]
- Sung, G.H.; Sung, J.M.; Hywel-Jones, N.L.; Spatafora, J.W. A multi-gene phylogeny of Clavicipitaceae (Ascomycota, Fungi): Identification of localized incongruence using a combinational bootstrap approach. Mol. Phylogenet. Evol. 2007, 44, 1204–1223. [Google Scholar] [CrossRef]
- Liu, Y.J.; Whelen, S.; Hall, B.D. Phylogenetic relationships among ascomycetes: Evidence from an RNA polymerse II subunit. Mol. Biol. Evol. 1999, 16, 1799–1808. [Google Scholar] [CrossRef]
- Hong, S.G.; Cramer, R.A.; Lawrence, C.B.; Pryor, B.M. Alt a 1 allergen homologs from Alternaria and related taxa: Analysis of phylogenetic content and secondary structure. Fungal Genet. Biol. 2005, 42, 119–129. [Google Scholar] [CrossRef] [PubMed]
- Andrew, M.; Peever, T.L.; Pryor, B.M. An expanded multilocus phylogeny does not resolve morphological species within the small-spored Altemrnaria species complex. Mycologia 2009, 101, 95–109. [Google Scholar] [CrossRef] [PubMed]
- Thompson, J.D.; Gibson, T.J.; Plewniak, F.; Jeanmougin, F.; Higgins, D.G. The CLUSTAL_X windows interface: Flexible strategies for multiple sequence alignment aided by quality analysis tools. Nucleic Acids Res. 1997, 25, 4876–4882. [Google Scholar] [CrossRef] [PubMed]
- Tamura, K.; Nei, M. Estimation of the number of nucleotide substitutions in the control region of mitochondrial DNA in humans and chimpanzees. Mol. Biol. Evol. 1993, 10, 512–526. [Google Scholar] [CrossRef] [PubMed]
- Schneider, C.A.; Rasband, W.S.; Eliceiri, K.W. NIH Image to ImageJ: 25 years of image analysis. Nat. Methods 2012, 9, 671–675. [Google Scholar] [CrossRef] [PubMed]
- Hanlon, A.; Taylor, M.; Dick, J. Agar dilution susceptibility testing; CRC Press: The journal CRC Press eBooks, 2007. [Google Scholar]
- Gorai, P.S.; Ghosh, R.; Konra, S.; Mandal, N.C. Biological control of early blight disease of potato caused by Alternaria alternata EBP3 by an endophytic bacterial strain Bacillus velenzensis SEB1. Biol. Control 2021, 156, 104551. [Google Scholar] [CrossRef]
- Agrios, G.N. Plant Patholgy 5th edition; Elsevier Academic Press: Amsterdam, 2005. [Google Scholar]
- Edin, E.; Liljeroth, E.; Andersson, B. Long term field sampling in Sweden reveal a shift in occurrence of cytochrome b genotype and amino acid substitution F129L in Alternaria solani, together with a high incidence of the G143A substitution in Alternaria alternata. Eur. J. Plant Pathol. 2019, 155, 627–641. [Google Scholar] [CrossRef]
- Vandecasteelea, M.; Landschoota, S.; Carrettea, J.; Verwaerena, J.; Hofte, M.; Audenaerta, K.; Haesaert, G. Species prevalence and disease progression studies demonstrate a seasonal shift in the Alternaria population composition on potato. Plant Pathol. 2018, 67, 327–336. [Google Scholar] [CrossRef]
- Dube, J.P.; Truter, M.; van der Waals, J.E. First report of resistance to QoI fungicides in Alternaria alternata isolates from potato in South Africa. Plant Dis. 2014, 98, 1431. [Google Scholar] [CrossRef]
- Kokaeva, L.Y.; Belosokhov, A.F.; Doeva, L.Y.; Skolotneva, E.S.; Elansky, S.N. Distribution of Alternaria species on blighted potato and tomato leaves in Russia. J. Plant Dis. Prot. 2018, 125, 205–212. [Google Scholar] [CrossRef]
Figure 1.
Naturally occurring leaf brown spot on potatoes, representatives of collected isolates, and percentage distribution of fungal isolates during 2020–2021 in Korea. (a) Brown spot symptoms on potato leaves collected from Yeoncheon (June 2021). Symptoms on the front of the leaves (left), back of the leaves (centre), and stem (right); (b) 14-day-old cultures on PDA from five representative fungal species, including Alternaria spp. (far left): Fusarium spp. (second from the left), Boeremia spp. (centre), Stagonosporopsis spp. (right of centre), and Colletotrichum sp. (far right); (c) Spores of each fungus; and (d) Percentage distribution of fungal species from 2020 (left), 2021 (centre), and the combined collection from 2020 to 2021 (right).
Figure 1.
Naturally occurring leaf brown spot on potatoes, representatives of collected isolates, and percentage distribution of fungal isolates during 2020–2021 in Korea. (a) Brown spot symptoms on potato leaves collected from Yeoncheon (June 2021). Symptoms on the front of the leaves (left), back of the leaves (centre), and stem (right); (b) 14-day-old cultures on PDA from five representative fungal species, including Alternaria spp. (far left): Fusarium spp. (second from the left), Boeremia spp. (centre), Stagonosporopsis spp. (right of centre), and Colletotrichum sp. (far right); (c) Spores of each fungus; and (d) Percentage distribution of fungal species from 2020 (left), 2021 (centre), and the combined collection from 2020 to 2021 (right).

Figure 2.
Phylogenetic analysis. Phylogenetic tree constructed based on concatenated sequences of ITS, gapdh, tef1, rpb2, Alt a1, endoPG, and OPA10-2 from 23 strains of Alternaria spp. Reference sequences were retrieved from GenBank (accession numbers shown in supplementary Table 1). the tree was constructed using the maximum likelihood method, and bootstrap values (1,000 replications) are shown in front of each node. MEGA X was used for the analysis.
Figure 2.
Phylogenetic analysis. Phylogenetic tree constructed based on concatenated sequences of ITS, gapdh, tef1, rpb2, Alt a1, endoPG, and OPA10-2 from 23 strains of Alternaria spp. Reference sequences were retrieved from GenBank (accession numbers shown in supplementary Table 1). the tree was constructed using the maximum likelihood method, and bootstrap values (1,000 replications) are shown in front of each node. MEGA X was used for the analysis.
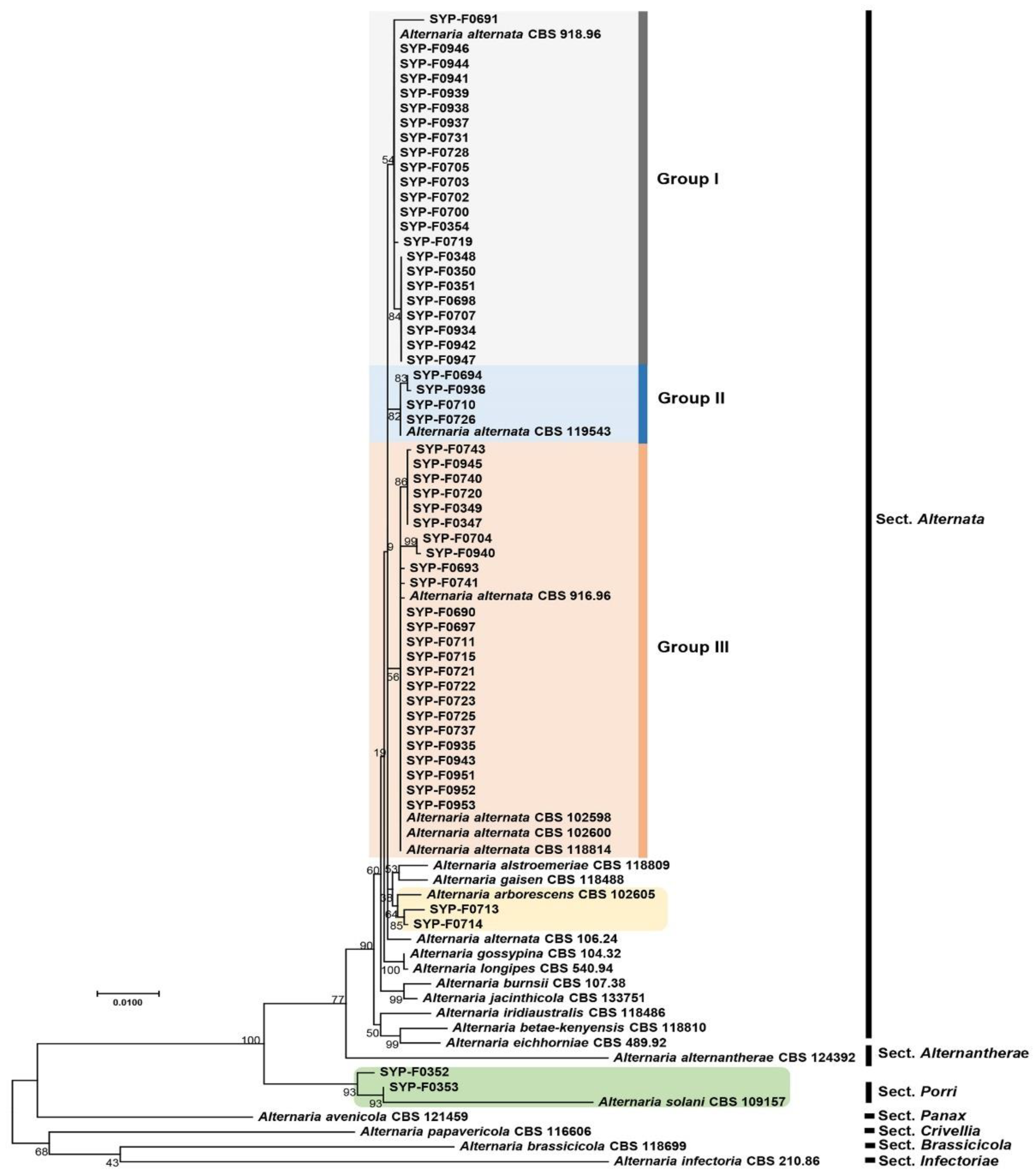
Figure 3.
Pathogenicity test. One-month-old potato cultivar Superior plants were inoculated by spraying with conidial suspension (1 × 105 conidia/mL). The photo captured 7 days post-inoculation.
Figure 3.
Pathogenicity test. One-month-old potato cultivar Superior plants were inoculated by spraying with conidial suspension (1 × 105 conidia/mL). The photo captured 7 days post-inoculation.

Figure 4.
Detached leaf assay without/with wound and box plot analysis. (a) Detached leaf assay. Sterilised distilled water was used as the control. Isolates for the virulence test were selected from groups I, II, and III on the left, which are identical to the groups shown in Figure 2. Box plot analysis using data from the disease leaf area (b) without wound, and (c) with wound of virulence test. Bars indicate the standard error of the means (n = 25).
Figure 4.
Detached leaf assay without/with wound and box plot analysis. (a) Detached leaf assay. Sterilised distilled water was used as the control. Isolates for the virulence test were selected from groups I, II, and III on the left, which are identical to the groups shown in Figure 2. Box plot analysis using data from the disease leaf area (b) without wound, and (c) with wound of virulence test. Bars indicate the standard error of the means (n = 25).
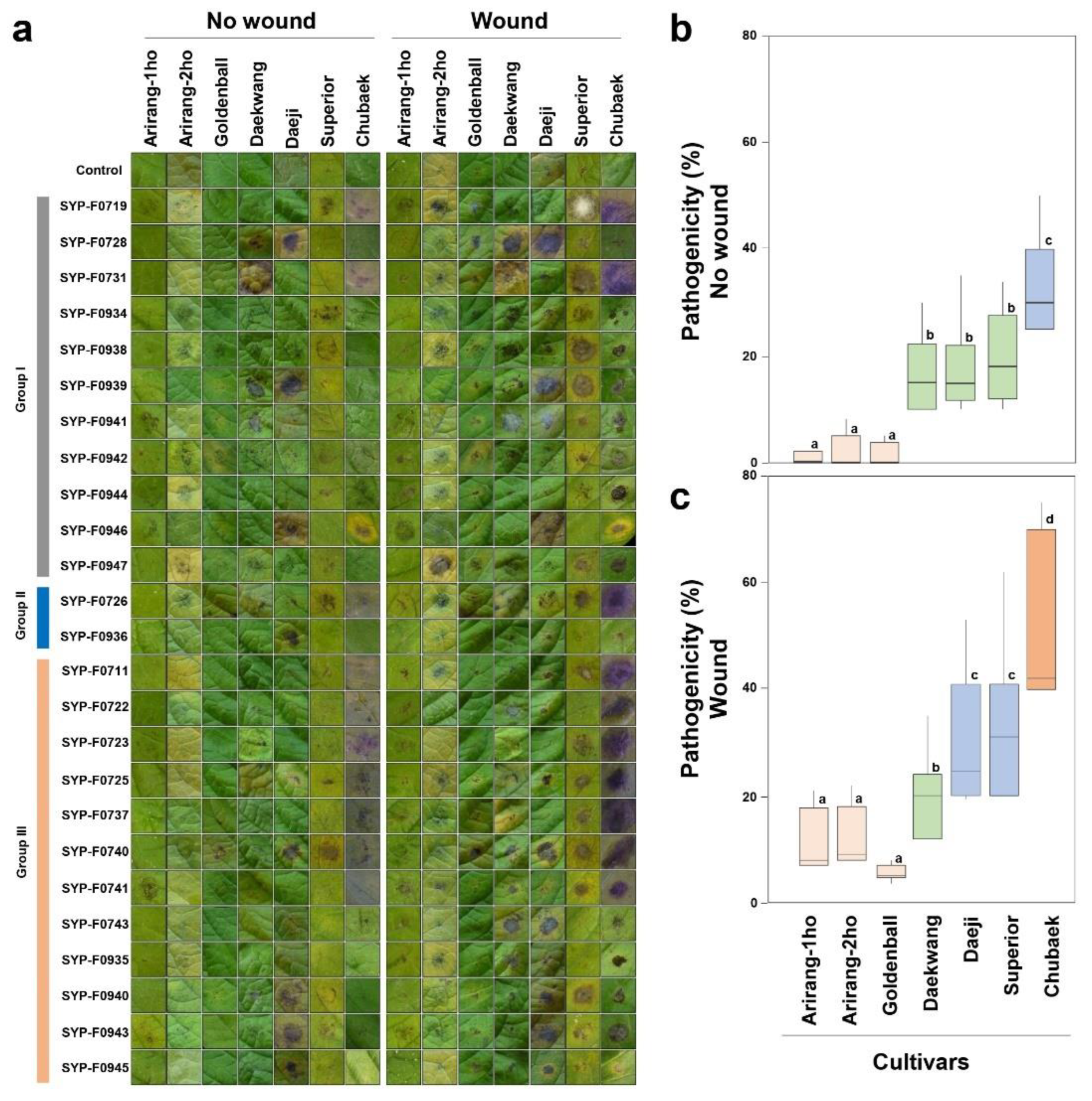
Figure 5.
Effect of seven fungicides on growth rates of selected Alternaria spp. isolates causing potato brown leaf spot. (a) Selected Alternaria spp. isolates grown for 7 days on PDA containing mancozeb (1,500 μg/mL), chlorothalonil (1,253 μg/mL), difenoconazole (34 µg/mL), pydiflumetofen (46 μg/mL), boscalid (328 μg/mL), kresoxim-methyl (148 μg/mL), or thiophanate-methyl (700 μg/mL).
Figure 5.
Effect of seven fungicides on growth rates of selected Alternaria spp. isolates causing potato brown leaf spot. (a) Selected Alternaria spp. isolates grown for 7 days on PDA containing mancozeb (1,500 μg/mL), chlorothalonil (1,253 μg/mL), difenoconazole (34 µg/mL), pydiflumetofen (46 μg/mL), boscalid (328 μg/mL), kresoxim-methyl (148 μg/mL), or thiophanate-methyl (700 μg/mL).


Table 1.
Primers used for PCR and sequencing.
| Locusa | Primer | Primer Sequence (5’-3’) | References |
|---|---|---|---|
| ITS | V9G | TTACGTCCCTGCCCTTTGTA | [16] |
| ITS4 | CCTCCGCTTATTGATATGC | [17] | |
| gapdh | gpd1 | CAACGGCTTCGGTCGCATTG | [18] |
| gpd2 | GCCAAGCAGTTGGTTGTGC | [18] | |
| tef1 | EF1-728F | CATCGAGAAGTTCGAGAAGG | [19] |
| EF1-986R | TAC TTG AAG GAA CCC TTA CC | [19] | |
| EF2 | GGARGTACCAGTSATCATGTT | [20] | |
| rpb2 | RPB2-5F2 | GGGGWGAYCAGAAGAAGGC | [21] |
| fRPB2-7cR | CCCATRGCTTGTYYRCCCAT | [22] | |
| Alt a 1 | Alt-For | ATGCAGTTCACCACCATCGC | [23] |
| Alt-Rev | ACGAGGGTGAYGTAGGCGTC | [23] | |
| endoPG | PG3 | TACCATGGTTCTTTCCGA | [24] |
| PG2b | GAGAATTCRCARTCRTCYTGRTT | [24] | |
| OPA10-2 | OPA 10-2R | GATTCGCAGCAGGGAAACTA | [24] |
| OPA 10-2L | TCGCAGTAAGACACA TTCTACG | [24] |
aITS: internal transcribed spacer regions 1 and 2 and interventing 5.8S nrDNA, gapdh: glyceraldehyde 3-phosphate dehydrogenase, tef1: translation elongation factor 1-alpha, rpb2: RNA polymerase second largest subunit, Alt a 1: Alternaria major allergen gene, endoPG: endopolygalacturonase, and OPA10-2: anonymous region.
Table 2.
Chemical names, group name, formulation and final concentration on the medium used in the Alternaria trials.
Table 2.
Chemical names, group name, formulation and final concentration on the medium used in the Alternaria trials.
| Chemical Name | Target Site | Group Name | Formulation (%) |
Final conc. of Fungicide on the Medium (μg/mL) |
|---|---|---|---|---|
| Mancozeb | Multi-site contact activity |
Dithiocarbamates | 75 | 1,500 |
| Chlorothalonil | Chloronitriles | 75 | 1,253 | |
| Difenoconazole | Inhibit sterol biosynthesis in membrane | C14-methylase in sterol biosynthesis | 10 | 34 |
| Pydiflumetofen | “Complex II” Succinate dehydrogenase |
Succinate-dehydrogenase inhibitor (SDHI) | 18.35 | 46 |
| Boscalid | 49.30 | 328 | ||
| Krexosim-methyl | Inhibit mitochondrial respiration | Quinone outside inhibitor (QoI) | 40.20 | 148 |
| Thiophanate-methyl | B1 tubulin polymerization |
Methyl benzimidazole carbamates (MBC) | 70 | 700 |
Table 3.
List of collected isolates, locality, species, and GenBank accession numbers for ITS region sequences.
Table 3.
List of collected isolates, locality, species, and GenBank accession numbers for ITS region sequences.
| Isolates | The Closest Matched GenBank Taxa | GenBank Accession nos. | Query Over |
Percent | Collected Regions | Date of Isolation |
|---|---|---|---|---|---|---|
| SYP-F0690 | Alternaria alternata | OR787445.1 | 100 | 100 | Goseong | June 26, 2020 |
| SYP-F0691 | Alternaria alternata | OR787445.1 | 100 | 100 | Goseong | June 26, 2020 |
| SYP-F0693 | Alternaria alternata | OR787445.1 | 100 | 100 | Goseong | June 26, 2020 |
| SYP-F0694 | Alternaria alternata | OR787445.1 | 100 | 100 | Goseong | June 26, 2020 |
| SYP-F0697 | Alternaria alternata | OR787445.1 | 100 | 100 | Goseong | June 26, 2020 |
| SYP-F0698 | Alternaria alternata | OR787445.1 | 100 | 100 | Goseong | June 26, 2020 |
| SYP-F0700 | Alternaria alternata | OR787445.1 | 100 | 100 | Goseong | June 26, 2020 |
| SYP-F0701 | Alternaria alternata | OR787445.1 | 100 | 100 | Goseong | June 26, 2020 |
| SYP-F0702 | Alternaria alternata | OR787445.1 | 100 | 100 | Goseong | June 26, 2020 |
| SYP-F0703 | Alternaria alternata | OR787445.1 | 100 | 100 | Goseong | June 26, 2020 |
| SYP-F0704 | Alternaria alternata | OR787445.1 | 100 | 100 | Goseong | June 26, 2020 |
| SYP-F0705 | Alternaria alternata | OR787445.1 | 100 | 100 | Goseong | June 26, 2020 |
| SYP-F0706 | Alternaria alternata | OR787445.1 | 100 | 100 | Goseong | June 26, 2020 |
| SYP-F0707 | Alternaria alternata | OR787445.1 | 100 | 100 | Goseong | June 26, 2020 |
| SYP-F0696 | Fusarium acuminatum | MT635295.1 | 100 | 100 | Goseong | June 26, 2020 |
| SYP-F0688 | Fusarium equiseti | MT560375.1 | 100 | 100 | Goseong | June 26, 2020 |
| SYP-F0689 | Fusarium equiseti | MT560375.1 | 100 | 100 | Goseong | June 26, 2020 |
| SYP-F0692 | Fusarium equiseti | MT560375.1 | 100 | 100 | Goseong | June 26, 2020 |
| SYP-F0708 | Fusarium equiseti | MT560375.1 | 100 | 100 | Goseong | June 26, 2020 |
| SYP-F0709 | Fusarium equiseti | MT560375.1 | 100 | 100 | Goseong | June 26, 2020 |
| SYP-F0347 | Alternaria alternata | MH992147.1 | 100 | 100 | Goseong | July 22, 2020 |
| SYP-F0348 | Alternaria alternata | OR787445.1 | 100 | 100 | Goseong | July 22, 2020 |
| SYP-F0349 | Alternaria alternata | KX816031.1 | 100 | 100 | Goseong | July 22, 2020 |
| SYP-F0350 | Alternaria alternata | OR787445.1 | 100 | 99.81 | Goseong | July 22, 2020 |
| SYP-F0351 | Alternaria alternata | OR787445.1 | 100 | 100 | Goseong | July 22, 2020 |
| SYP-F0354 | Alternaria alternata | OR787445.1 | 100 | 100 | Baengnyeongdo | July 22, 2020 |
| SYP-F0352 | Alternaria solani | MT498268.1 | 100 | 100 | Baengnyeongdo | July 22, 2020 |
| SYP-F0353 | Alternaria solani | OR787445.1 | 100 | 100 | Baengnyeongdo | July 22, 2020 |
| SYP-F0937 | Alternaria alternata | OR787445.1 | 100 | 100 | Yeoncheon | June 27, 2020 |
| SYP-F0938 | Alternaria alternata | OR787445.1 | 100 | 100 | Yeoncheon | June 27, 2020 |
| SYP-F0939 | Alternaria alternata | OR787445.1 | 100 | 100 | Yeoncheon | June 27, 2020 |
| SYP-F0940 | Alternaria alternata | OR787445.1 | 100 | 100 | Yeoncheon | June 27, 2020 |
| SYP-F0941 | Alternaria alternata | OR787445.1 | 100 | 100 | Yeoncheon | June 27, 2020 |
| SYP-F0942 | Alternaria alternata | OR787445.1 | 100 | 100 | Yeoncheon | June 27, 2020 |
| SYP-F0943 | Alternaria alternata | OR787445.1 | 100 | 100 | Yeoncheon | June 27, 2020 |
| SYP-F0944 | Alternaria alternata | OR787445.1 | 100 | 100 | Yeoncheon | June 27, 2020 |
| SYP-F0945 | Alternaria alternata | OR787445.1 | 100 | 100 | Yeoncheon | June 27, 2020 |
| SYP-F0946 | Alternaria alternata | OR787445.1 | 100 | 100 | Yeoncheon | June 27, 2020 |
| SYP-F0947 | Alternaria alternata | OR787445.1 | 100 | 100 | Yeoncheon | June 27, 2020 |
| SYP-F0710 | Alternaria alternata | OR787445.1 | 100 | 100 | Yeoncheon | June 22, 2021 |
| SYP-F0711 | Alternaria alternata | OR787445.1 | 100 | 100 | Yeoncheon | June 22, 2021 |
| SYP-F0712 | Alternaria alternata | OR787445.1 | 100 | 100 | Yeoncheon | June 22, 2021 |
| SYP-F0715 | Alternaria alternata | OR687203.1 | 100 | 99.61 | Yeoncheon | June 22, 2021 |
| SYP-F0716 | Alternaria alternata | OR787445.1 | 100 | 100 | Yeoncheon | June 22, 2021 |
| SYP-F0717 | Alternaria alternata | OR787445.1 | 100 | 100 | Yeoncheon | June 22, 2021 |
| SYP-F0718 | Alternaria alternata | MT524302.1 | 100 | 100 | Yeoncheon | June 22, 2021 |
| SYP-F0719 | Alternaria alternata | OR787445.1 | 100 | 100 | Yeoncheon | June 22, 2021 |
| SYP-F0720 | Alternaria alternata | OR787445.1 | 100 | 100 | Yeoncheon | June 22, 2021 |
| SYP-F0721 | Alternaria alternata | OR734592.1 | 100 | 100 | Yeoncheon | June 22, 2021 |
| SYP-F0722 | Alternaria alternata | OR787445.1 | 100 | 100 | Yeoncheon | June 22, 2021 |
| SYP-F0723 | Alternaria alternata | OR787445.1 | 100 | 100 | Yeoncheon | June 22, 2021 |
| SYP-F0725 | Alternaria alternata | OR787445.1 | 100 | 100 | Yeoncheon | June 22, 2021 |
| SYP-F0726 | Alternaria alternata | OR787445.1 | 100 | 100 | Yeoncheon | June 22, 2021 |
| SYP-F0728 | Alternaria alternata | OR787445.1 | 100 | 100 | Yeoncheon | June 22, 2021 |
| SYP-F0729 | Alternaria alternata | OK315470.1 | 100 | 100 | Yeoncheon | June 22, 2021 |
| SYP-F0731 | Alternaria alternata | OR787445.1 | 100 | 100 | Yeoncheon | June 22, 2021 |
| SYP-F0737 | Alternaria alternata | OR787445.1 | 100 | 100 | Yeoncheon | June 22, 2021 |
| SYP-F0740 | Alternaria alternata | OR787445.1 | 100 | 100 | Yeoncheon | June 22, 2021 |
| SYP-F0741 | Alternaria alternata | ON599295.1 | 100 | 98.33 | Yeoncheon | June 22, 2021 |
| SYP-F0742 | Alternaria alternata | OR787445.1 | 100 | 100 | Yeoncheon | June 22, 2021 |
| SYP-F0743 | Alternaria alternata | MT498268.1 | 100 | 100 | Yeoncheon | June 22, 2021 |
| SYP-F0934 | Alternaria alternata | OR787445.1 | 100 | 100 | Yeoncheon | June 22, 2021 |
| SYP-F0935 | Alternaria alternata | OR787445.1 | 100 | 100 | Yeoncheon | June 22, 2021 |
| SYP-F0936 | Alternaria alternata | OR787445.1 | 100 | 100 | Yeoncheon | June 22, 2021 |
| SYP-F0713 | Alternaria arborescens | MT212228.1 | 100 | 100 | Yeoncheon | June 22, 2021 |
| SYP-F0714 | Alternaria arborescens | OR787445.1 | 100 | 100 | Yeoncheon | June 22, 2021 |
| SYP-F0745 | Alternaria longipes | MT524302.1 | 100 | 100 | Yeoncheon | June 22, 2021 |
| SYP-F0736 | Boeremia exigua | KY555024.1 | 100 | 100 | Yeoncheon | June 22, 2021 |
| SYP-F0733 | Boeremia exigua | MT397284.1 | 100 | 100 | Yeoncheon | June 22, 2021 |
| SYP-F0730 | Colletotrichum nymphaeae | LC435466.1 | 100 | 100 | Yeoncheon | June 22, 2021 |
| SYP-F0732 | Fusarium equiseti | MK752407.1 | 100 | 100 | Yeoncheon | June 22, 2021 |
| SYP-F0734 | Fusarium graminearum | OR346117.1 | 100 | 100 | Yeoncheon | June 22, 2021 |
| SYP-F0724 | Stagonosporopsis dennisii | OQ158929.1 | 100 | 99.18 | Yeoncheon | June 22, 2021 |
| SYP-F0727 | Stagonosporopsis dennisii | OK315470.1 | 100 | 100 | Yeoncheon | June 22, 2021 |
| SYP-F0951 | Alternaria alternata | OR787445.1 | 100 | 100 | Baengnyeongdo | July 19, 2021 |
| SYP-F0952 | Alternaria alternata | OR787445.1 | 100 | 100 | Baengnyeongdo | August 1, 2021 |
| SYP-F0953 | Alternaria alternata | OR787445.1 | 100 | 100 | Baengnyeongdo | August 1, 2021 |
Table 4.
Results of inhibition rate of seven selected fungicides against Alternaria spp.
| Isolates | Control | Mancozeb | Defenoconazole | Pydiflumetofen | Boscalid | Chlorothalonil | Krexoxim-methyl | Thiophanate-methyl |
|---|---|---|---|---|---|---|---|---|
| SYP-F0934 | 0.0±0.0g,*,# | 100.0±0.0a | 90.0±1.6b | 77.3±4.2c | 77.3±1.6c | 46.4±1.6d | 38.2±3.1e | 14.6±3.1f |
| SYP-F0944 | 0.0±0.0g | 100.0±0.0a | 96.3±1.6b | 75.7±1.6c | 72.9±1.6c | 58.0±2.8d | 39.3±3.2e | 20.6±1.6f |
| SYP-F0945 | 0.0±0.0f | 100.0±0.0a | 90.5±1.6b | 84.8±3.3c | 81.0±1.6c | 41.9±1.6d | 44.8±4.4d | 25.7±2.9e |
| SYP-F0936 | 0.0±0.0g | 100.0±0.0a | 87.3±2.5b | 75.8±1.9c | 68.1±1.9d | 40.0±1.0e | 3.2±1.9g | 10.9±1.7f |
| SYP-F0935 | 0.0±0.0f | 100.0±0.0a | 83.7±2.7b | 77.3±1.6bc | 80.0±5.7c | 51.9±1.6d | 55.5±1.6d | 28.2±1.6e |
| SYP-F0943 | 0.0±0.0f | 100.0±0.0a | 92.6±1.6b | 82.4±4.2c | 80.6±2.8c | 56.5±3.2d | 50.0±2.8e | 45.4±4.2e |
| SYP-F0713 | 0.0±0.0h | 100.0±0.0a | 88.1±0.0c | 97.0±0.0b | 74.3±1.7d | 41.6±1.7e | 32.7±3.4f | 13.9±0.0g |
| SYP-F0714 | 0.0±0.0f | 100.0±0.0a | 90.0±8.7b | 76.0±3.0c | 80.0±1.7c | 44.9±1.7d | 43.9±1.7d | 14.9±1.7e |
| SYP-F0352 | 0.0±0.0g | 100.0±0.0a | 89.0±2.8b | 79.8±1.6c | 79.8±1.6c | 52.2±1.6d | 43.1±1.6e | 14.6±7.3f |
| SYP-F0353 | 0.0±0.0f | 98.7±2.2a | 73.1±20.0b | 74.4±2.2b | 66.7±2.2b | 37.2±2.2c | 25.6±2.2c | 5.1±2.2d |
Data were presented as mean ± SD from three replicates. # Duncan’s multiple test was used to determine significance at the 95% probability level. The same letters in a column showed no significant difference.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



