
Submitted:
21 December 2023
Posted:
21 December 2023
You are already at the latest version
Abstract
Nutrition is one of the most influential environmental factor in both taxonomical shifts in gut microbiota as well as development of Type 2 Diabetes Mellitus (T2DM). Emerging evidence has shown that the effects of nutrition on both these parameters is not mutually exclusive and that changes in gut microbiota and related metabolites such as short chain fatty acids (SCFA) and branched chain amino acids (BCAA) may influence systemic inflammation and signaling pathways that contribute to pathophysiological processes associated with T2DM. With this background, our review highlights the effects of macronutrients, carbohydrates, proteins and lipids, as well as micronutrients, vitamins, and minerals, on T2DM, specifically through their alterations in gut microbiota and the metabolites they produce. Additionally, we describe the influences of common food groups, which incorporate varying combinations of these macronutrients and micronutrients, on both microbiota and metabolic parameters in the context of diabetes mellitus. Overall, nutrition is one of the first line modifiable therapies in the management of T2DM and better understanding of the mechanisms by which gut microbiota influence its pathophysiology provides opportunities for optimizing dietary interventions.
Keywords:
gut bacteria
; macronutrients
; micronutrients
; food groups
; insulin resistance
1. Introduction
The human gastrointestinal (GI) tract harbors trillions of gut microbiota, comprising about 500-1000 different bacterial species, which collectively weigh approximately 1-2 kilograms [1,2]. Analysis of the human microbial composition have shown that the gut microbiota of adults contains six phyla, with 90% of these bacterial species belonging to the phyla Bacteroidetes and Firmicutes, while the phyla Actinobacteria, Proteobacteria, Fusobacteria and Verrucomicrobia make up the rest [3]. The balance of the gut microbiota, namely eubiosis, is important in maintaining health and preventing diseases [4]. It has been well documented that the profile of the gut microbiota is unique to each individual host, and its composition is influenced by a variety of factors, resulting in taxonomical shifts in microbial species throughout a person’s lifetime [5,6]. This includes both extrinsic factors such as lifestyle, stress, medication, diet, disease status, as well as intrinsic factors, such as genetics, immune or metabolic [5,7] with extrinsic factors having the greatest effect on gut microbiota [7]. The influence of gut microbiota on homeostatic processes in the human body are multifaceted, with important roles in modulating metabolic processes [8], regulating immune responses [9], and maintaining overall host health [5]. More specifically, the effects of gut microbiota have been linked to a myriad of non-communicable diseases including type 2 diabetes mellitus (T2DM), dyslipidemia, obesity, Parkinson’s disease [10,11,12,13], to name a few. In particular, T2DM, a multifactorial chronic metabolic condition that is characterized by hyperglycemia, lipid imbalance and insulin resistance [14], remains a major global health threat affecting approximately 6% of the world’s population [15] and contributing significantly to the worldwide socio-economic burden [10]. The prevalence of T2DM continue to increase, with an estimated 530 million individuals being affected by the disease of which 22% are above the age of 70 [15]. Therefore, gaining a deeper understanding of the factors and mechanisms controlling hyperglycemia and insulin resistance is critical in the prevention, management, and effective therapeutic interventions of diabetes.
Gut microbiota has been long recognized as a key component in regulating host health and specific bacteria have been causally linked with the onset and progression of diseases, including diabetes. The extent of the effects of gut bacteria on T2DM has been attributed to both taxonomical shifts in gut microbiota as well as differential production of important gut metabolites including short chain fatty acids (SCFA), bile acids (BA) and amino acids (AA), which are shown to contribute to or protect against hyperglycemia and insulin resistance [10]. For example, bacterial species belonging to the genera Bifidobacterium and Lactobacillus, which are significant producers of SCFA, have been correlated with reduction of HbA1c serum levels [16]. Similarly, altered absorption of SCFA and BA have been observed in patients with T2DM as a result of increased gut barrier permeability induced by dysbiosis of gut flora [17] . Importantly, nutrition has been shown to be the primary modifiable factor of gut microbiota remodeling and in the development of T2DM, with various diets, food groups, macronutrients and micronutrients exerting different effects on its composition [18,19]. For example, beneficial diets such as Mediterranean diet, primarily composed of plant-based products, are inversely related with HgbA1c levels, waist circumference and insulin resistance [20], while animal-based product diets promote opposite effects [21]. Further, the type of food as well as the macronutrient and micronutrient composition of the diet exert distinct effects on gut microbiota and related metabolites, with major consequences on mechanisms regulating hyperglycemia and insulin resistance [21,22,23]. This review describes the key role of nutrients at the intersection between gut microbial eubiosis and the development of T2DM. It presents changes in gut microbiota composition profile of individuals with T2DM and how specific gut bacteria and related metabolites contribute to, or safeguard against, diabetes. Further, the effects of various macronutrients and micronutrients on microbiota-T2DM relationship and the impact of the common food groups on the gut microbial composition and T2DM are discussed.
2. Influence of Gut Microbiota on Hyperglycemia, Insulin Resistance and T2DM
Over the past several years, numerous studies have linked gut microbiota and T2DM, with factors such as systemic inflammation through production of lipopolysaccharides (LPS) [24], changes in gut membrane permeability and bile acid metabolism [25], all playing significant roles in the degree of insulin resistance in the host [1,10]. Gut bacteria has been associated with glucose intolerance as germ free mice show differential resistance against high fat diet-induced insulin resistance and adiposity [26,27,28]. More specifically, studies evaluating conventional gut microbial composition in T2DM, showed important trends in taxonomical shifts in gut bacteria that may have strong associations with the pathogenesis of condition. For example, an increase in Firmicutes/Bacteroidetes ratio has been linked to conditions associated with low-grade inflammation such as obesity and T2DM [29,30]. Further, the dysbiosis in T2DM has been characterized by a decrease in butyrate-producing bacterial species, mainly Roseburia intestinalis, Bifidobacterium spp., Akkermansia spp., and Faecalibacterium prausnitzii, and an increase in the abundance of unfavorable bacteria such as Clostridium clostridioforme, Clostridium hathewayi, Clostridium ramosum, Clostridium symbiosum, Bacteroides caccae, Escherichia Coli, Eggerthella spp., Fusobacterium and mucin-degrading bacterial genera, Ruminococcus [31,32,33,34]. Studies linking Lactobacillus with type 2 diabetes have been inconsistent [31].
The collective taxonomical shifts in gut microbiota composition are associated with increased gut and systemic inflammation, further contributing to the pathogenesis of T2DM [35] . For example, increase in the pathogenic gram-negative bacteria, leads to release of lipopolysaccharides (LPS), which is known to activate toll-like receptor 4 (TLR4) in adipocytes, promoting inflammatory signaling and cytokine expression [36]. Previous data has shown that LPS binding to TLR4 is associated with insulin resistance, since mice lacking the TLR4 are protected from suppressed insulin signaling and insulin-mediated changes in glucose metabolism [37]. Further studies have shown that adipocytes expressing TLR4 promote induction of pro-inflammatory cytokines, particularly IL-6 and IL-8 which decreased insulin-induced glucose uptake through downregulation of insulin-receptor substrate 1 (IRS-1) and glucose transporter 4 (GLUT4) [38]. Serine kinases, such as c-Jun-N-terminal Kinase (JNK) and inhibitor of nuclear factor kappa-B kinase subunit beta (IKKβ) contribute to insulin resistance through phosphorylation of IRS-1 [39,40]. Additionally, the mechanism behind increased insulin resistance, adiposity and lipid abnormalities is also thought to be due to LPS-induced increase in gut permeability by reduced expression of tight junction proteins, primarily zonula occludens-1 and minimally to claudin and occluden [41]. The impaired gut barrier integrity leads to translocation of LPS into the bloodstream and development of metabolic endotoxemia [42]. This, in turn, contributes to a sustained low-grade inflammation, via central insulin resistance and activation of the hypothalamic c-Jun N-terminal Kinase (JNK) cascade [43]. Taken together, these findings provide strong evidence for the influence of bacteria and its byproducts in altered insulin sensitivity (Figure 1).
The gut microbiota byproducts have been shown to exert generally protective effects on hyperglycemia and insulin resistance. Among them, SCFA, the enzymatically degraded end-product of anaerobic fermentation reactions of indigestible complex carbohydrates [44] such as butyrate, propionate, and acetate are the most metabolically important [45]. For example, oral supplementation of butyrate in a diabetic rodent model significantly decreased serum hemoglobin A1c, LPS and pro-inflammatory cytokines levels, while concomitantly improving gut integrity through measurement of intracellular adhesion molecules [46]. Importantly, these findings were accompanied by increased Firmicutes/Bacteroidetes ratio, which correlates with previous data showing the interplay between SCFA production and increases in gut microbial diversity [47]. Similar findings have been demonstrated with administration of a butyric acid derivative attenuating LPS-induced inflammation and insulin resistance with decreased phosphorylated IRS-1 measured in mouse adipocytes [48]. This involves the activated protein kinase (AMPK) dependent signaling, with beneficial effects including decreased inflammation, survival of β-cells, inhibition of insulin resistance and promotion of glucose metabolism and uptake [49]. Of note, acetate is also shown to have similar benefits, with increased AMPK signaling activity in the liver leading to hypoglycemic effects [49] . Similarly, SCFA have been shown to mitigate inflammatory processes, specifically through reprogramming metabolic activity of T lymphocytes [50]. For example, propionate enhanced expression of T regulatory cells, particularly Th17 helper cells and interleukin 10 (IL-10), ameliorating the negative effects of high-fat diet feeding [51]. T regulatory cells in adipocytes are shown to reduce adipose tissue inflammation and improve insulin resistance, further supporting the anti-inflammatory effects of SCFA on metabolic disease [52]. In addition to anti-inflammatory effects and improvement of hyperglycemia, SCFA also exert antidiabetic effects through insulin secretion in glucose-dependent manner by stimulating secretion of glucagon-like peptide 1 (GLP-1) via the free fatty acid receptors, FFAR2 and FFAR3 located on enteroendocrine cells [53]. Probiotic administration increased levels of both SCFA and SCFA-producing bacterial species, while decreasing pathogenic Escherichia coli and LPS [54]. Interestingly, it has also been shown that metformin, one of the first-line treatments for T2DM, promotes abundance of SCFA-producing gut microbiota, which is correlated to the secretion of GLP-1 [55]. Therefore, SCFA play an integral role in ameliorating T2DM through improving inflammation, activating important signaling pathways and modulation of gut peptides (Figure 1).
In addition, crosstalk between bile acids and gut microbiota play significant roles in the development or protection against hyperglycemia and insulin resistance [25]. Studies have shown that gut microbiota are involved in enzymatically converting primary bile acids into secondary bile acids [55], through expression of bile salt hydrolase activity [25]. Secondary bile acid binding to farsenoid X receptor (FXR) and Takeda G-protein coupled receptor 5 (TGR5) are shown to restructure gut microbiota and influence markers of T2DM [56]. For example, TGR5 activation can enhance pancreatic and liver function, leading to enteroendocrine L-cell mediated GLP-1 release and improved insulin resistance [57]. FXR agonists improve insulin resistance in diabetic animal models [58], however other studies showed that FXR deficiency has a similar effect [59,60]. Further, bile acids have been shown to increase insulin sensitivity through the fibroblast growth factor (FGF) activity, with FGF21 signaling found to have beneficial effects both in the liver and in adipose tissue, through decreased activation of the mammalian target of rapamycin complex 1 (Mtorc1) pathway [61] and increased activation of peroxisome proliferator-activated receptor γ (PPARγ) [62], respectively. Specifically, signaling via the Mtorc1 pathway promotes serine phosphorylation of IRS-1 (p-IRS-1), a known marker of insulin resistance [61]. Conversely, activation of PPARγ improves insulin mediated skeletal muscle glucose uptake and hepatic glucose production to enhance insulin sensitivity [63]. Overall, it is evident that these receptors and pathways influenced by enterohepatic circulation of bile acids and gut microbiota, are heavily implicated in glucose homeostasis and insulin sensitivity (Figure 1).
3. Influence of Nutrition in Modulating Gut Microbiota and Markers of T2DM
Nutrition plays a critical role in the intricate relationships between gut microbiota and pathophysiology of T2DM, and it is the key common factor when considering microbiota altering interventions to improve hyperglycemia and insulin resistance [64]. Nutrition shapes the gut microbiota, and it accounts for over 20% of the inter-individual microbiome variability in humans and 50% in mouse models [65,66]. Therefore, identification of different diets, macronutrients, micronutrients and food groups and their related effects on gut microbiota and T2DM is an important approach to prevent and control diabetes. Although not all the effects of the interactions between food components and T2DM are completely known, it is clear that diets rich in fruits and vegetables have beneficial effects on glucose metabolism [67]. For example, foods with a low glycemic index have beneficial effects on blood sugar, HbA1c, total cholesterol, LDL cholesterol and the inflammatory response in patients with diabetes and in the prevention of T2DM development across populations [68,69,70,71,72]. A recent study in obese women who followed a low-glycemic diet consisting mainly of whole grains, fish, vegetables, algae and perilla oil or a control diet consisting mainly of refined rice, bread, noodles, meat and processed foods showed a higher level of Gemminger formicilis, Collinsella aerofaciens, Escherichia coli and Bifidobacterium longum and a lower serum butyric acid level in those receiving the control diet compared to the low glycemic diet [73]. Gut dysbiosis and increases in abundance of pathogenic bacteria, especially Bacteroides have been reported in the presence of a carbohydrate rich diet [74]. However, not all studies showed significant differences in glycemic control of lipid profile in people who followed a low glycemic index diet compared to other types of diet [71,75,76].
Multiple studies have demonstrated associations between different dietary patterns and the risk for T2DM, however these associations are quite complex, because people do not consume individual foods, but mixtures of foods [77] causing corresponding changes in microbial composition. Further, it has been shown that long-term diet leads to the establishment of major enterotypes Prevotella, Bacteroides, and Ruminococcus [73,78] since diet is the main modulator of gut microbiota. Bifidobacterium spp., Lactobacillus spp., Bacteroides spp., Alistipes spp., Bilophila spp., Clostridium spp., Roseburia spp., Eubacterium spp., Enterococcus spp., Faecalibacterium prausnitzii, Akkermansia muciniphila, Escherichia coli, Helicobacter pylori, Streptococcus spp. [79] are among the many bacteria influenced by the diet. For example, Prevotella enterotype was associated with a high intake of carbohydrates, especially sugar, while the Bacteroides enterotype was associated with a high intake of meat [73]. In addition, specific diets such as the Western Diet (WD) and the Mediterranean diet (MD) have been shown to exert differential changes in the gut microbiota composition and ensuing metabolic functions. As such, an hypercaloric diet high in fats and animal proteins, characteristic of WD is associated with microbial dysbiosis [80] The WD is shown to increase unfavorable species such as Escherichia coli and Ruminococcus torques [81], which in turn, promotes increased gut permeability and metabolic endotoxemia through increased abundances in these gram-negative LPS-producing bacterial genera [82], contributing to insulin resistance. At the same time, it has been shown that the WD reduces SCFA-producing bacterial genera, such as Eubacterium and Roseburia [83]. These pro-inflammatory changes in gut permeability via the WD can be attributed, to some extent, to mTOR hyperactivation, which was improved after antibiotic introduction [84]. Further, the effects of similar diets are not limited to peripheral changes but are also associated with central insulin resistance as evidenced through increased serine phosphorylation of IRS-1 and inflammatory responses through nuclear factor kappa beta (NFKβ) and JNK activity [85]. In addition to phosphorylating IRS-1, JNK contributes to insulin resistance through promoting metabolic inflammation and negatively regulating interactions between PPARα-FGF21 as well as contributing to adiposity through dysregulation of the thyroid stimulating hormone (TSH) axis [86]. Studies also have shown that WD in patients with T2DM, promotes more C-peptide post-prandially, which is an endogenous marker of insulin secretion [87]. Increased insulin secretion was also reported after consumption of a Westernized diet, which preceded peripheral insulin resistance [88].
On the other hand, the MD, characterized by a high intake of dietary fiber, nuts, whole grains, and omega-3 poly-unsaturated fatty acids has been associated with favorable effects on gut microbiota composition and hyperglycemia [89]. Interestingly, the effects on gut microbiota are largely opposite from those observed after WD adherence, with MD consumption promoting increased relative abundance in main SCFA-producing genera like Lactobacillus, Bifidobacterium, Eubacterium and Faecalibacterium while reducing concentrations of Bacteroides and Prevotella spp. [90,91] , which collectively contribute to better glucose homeostasis. One large scale observational study of over 22,000 human participants who have adhered to the MD for 6 months showed that these individuals had lower risk of new onset T2DM [92]. The mechanisms behind improvements in glucose homeostasis include reduction of inflammatory processes [93], modulation of gut hormones [94,95] and altered production of microbial metabolites [96]. For example, 12-week adherence to the MD reduced pro-inflammatory cytokine interleukin-6 (IL-6) by 49% in T2DM patients [93]. Similarly, markers of inflammation such as C-reactive protein (CRP) and intracellular adhesion molecule-1 (ICAM-1) showed significant reduction post-MD [97]. ICAM-1 is heavily intertwined in T-cell mediated processes indicating that the adaptive immune response is also affected by this dietary intervention [98]. In addition, the MD exerts antioxidant and anti-hyperglycemic effects by augmenting GLP-1 activity in endothelial cells. These findings are also supported by two recent human studies showing that MD adherence over 210 days or 24 weeks, respectively, was associated with lower serum glucose, elevated fasting GLP-1 level, improved insulin resistance and HgbA1c [94,95]. Importantly, some of the bacterial genera changes associated with the MD, such as decreases in Prevotella and Bacteroides may influence markers of insulin resistance [96]. These genera have been shown to aggravate insulin resistance and cause glucose intolerance by elevating circulating levels of branched chain amino acids (BCAA) [96]. Studies have linked increased concentrations of BCAA to activation of mTORC1, a cell growth regulator, which causes dysregulation of insulin signaling [99]. In parallel, these BCAA are shown to be associated with pancreatic β-cell mitochondrial dysfunction and apoptosis, further contributing to insulin resistance [100]. Taken together, these findings support the role of dietary patterns in the remodeling of gut microbiota and resulting alterations in biomarkers associated with inflammation, hyperglycemia, and insulin signaling dysregulation.
4. Effects of Macronutrients on T2DM and Gut Microbiota
Various macronutrients exert distinct effects on gut microbiota and T2DM. In the following subsections, we explore the role of carbohydrates, dietary fibers and starches, as well as of proteins and lipids in mediating these effects.
4.1. Carbohydrates
Digestible carbohydrates are enzymatically degraded in the small intestine and are represented by starch and sugars, such as glucose, fructose, sucrose, and lactose [101]. Breakdown of these compounds stimulate insulin response by releasing glucose into the bloodstream thereby influencing insulin signaling [102]. Over the years, the relationship between carbohydrate intake, diabetes and gut microbiota has been studied, with differences shown in the post-prandial glycemic response determined both by the amount as well as by the type of carbohydrate consumed [103,104]. Low carbohydrate diets (LCD) have long been part of the main nutritional therapy regimen in the management of type 2 diabetes [105]. For example, a LCD, characterized by under 40% carbohydrates of total energy intake, had a beneficial effect on HgbA1c as compared to both very-low carbohydrate content or moderate carbohydrate content (40-64% of total energy intake) [106]. ADA guidelines include the importance of diets with low carbohydrate content in reducing HbA1c levels [107]. Numerous studies have supported the hypoglycemic effect of LCD through decreased blood sugar and increased insulin sensitivity, leading to lowering oral antidiabetic medications, while also improving lipid parameters such as increased HDL-cholesterol and decreased triglycerides [108,109,110,111]. Adherence to LCD reduced the risk for T2DM in children and adolescents [112] while a diet high in carbohydrates increased the risk of T2DM [113], clearly demonstrating the importance of this macronutrient in glucose homeostasis. Further a LCD is shown to modulate gut hormones such as GLP-1, while concomitantly promoting beneficial changes in gut microbiota and diabetic markers [22,114]. For example, LCD consumption for three months was associated with enhanced GLP-1 secretion in humans, reduced HgbA1c and increased abundance of SCFA-producing species, Roseburia, Ruminococcus and Eubacterium [22]. These SCFAs resulting from colonic carbohydrate fermentation act on free fatty acid receptors, FFAR2 (GPR43) and FFAR3 (GPR41), to stimulate GLP-1 release through the mitogen-activated protein kinase (MAPK) / extracellular regulated protein kinase (ERK) pathway [115]. For example, acetate increased GLP-1 secretion up to three-fold and butyrate by two-fold [115,116] in response to administration of Bifidobacterium, Lactobacillus and Enterococcus spp. Therefore, these metabolic signaling pathways involving both the host and bacteria pathways, play a significant role in metabolic health in response to carbohydrate intake (Figure 2).
On the other hand, high carbohydrate intake, including diets rich in glucose and fructose, promotes both metabolic disorders and intestinal dysbiosis [117] and have been associated with increase in the abundance of pathogenic bacteria, especially Bacteroides [74]. Additionally, metagenomic sequencing data of gut microbiota in animal models consuming carbohydrate-dense diets have shown increases in the Firmicutes to Bacteroidetes ratio as well as in pro-inflammatory Desulfovibrio vulgaris and mucin-degrading Akkermansia muciniphila [118]. These findings were accompanied by increased glucose intolerance, elevated serum glucose and a two and half fold increase in gut permeability [118,119]. The decrease in gut permeability may be attributed to the role of Akkermansia muciniphila, Bacteroides and Desulfovibrio spp. in inflammatory processes. In general, healthy amounts of Akkermansia have favorable effects on gut barrier integrity and inflammation [120], however, when in excess, an opposite effect is shown, with Akkermansia over-degrading the mucin layer leading to increased gut permeability and secretion of inflammatory cytokines [121]. Desulfovibrio, a gram-negative bacterium genera, known to produce hydrogen sulfide gas, increases T cell activity and systemic inflammation, known to cause important sequelae in both cognition and metabolic syndrome [122]. Desulfovibrio is also positively correlated with increases in fasting insulin, which can lead to insulin resistance [123]. Similarly, Bacteroides have been shown to degrade the mucin-layer when abundant and exhibit virulence factors that fuel their growth in conditions that cause low bacteria diversity [124]. Importantly, Bacteroides spp. increase biosynthesis of BCAA, an important marker for increased insulin resistance [96]. Taken together, these changes caused by high carbohydrate intake can contribute to metabolic endotoxemia, insulin resistance and hyperglycemia (Figure 2).
4.1.1. Dietary Fibers, Gut Microbiota and T2DM
Dietary fibers are plant components that are characterized by resistance to digestion and absorption in the small intestine [125]. Unlike digestible carbohydrates, dietary fiber is not enzymatically degraded in the small intestine but is fermented by microorganism resident in the large intestine [126]. Fibers are classified according to their physico-chemical characteristics, such as: fermentability, solubility and viscosity [127]. High intake of dietary fiber supports gut health [127] and promotes glycemic control, with recommendations of 25-50g/day of dietary fibers in diabetic patients [128]. There is compelling evidence demonstrating overall health benefits of diets rich in fiber and daily consumption of whole grains and bran was associated with decreased mortality due to cardiovascular causes, in patients with diabetes [106,129]. Moreover, patients with metabolic syndrome had a lower fiber intake than those without metabolic syndrome [130]. Similarly, high glycemic index diets and low fiber content are shown to induce metabolic syndrome in individuals with T2DM [131]. The effects of dietary fibers depend on their origin, noting that cereal fibers were more strongly associated with a decrease in the risk of diabetes, compared to fruit fibers, which had a weaker association [132]. However, dietary fibers derived from cereals and fruits have beneficial effects in controlling T2DM through improvements in inflammatory processes as measured through CRP and tumor necrosis factor alpha (TNF-α) levels [133]. that Dietary fibers also stimulate increases in circulating adiponectin, which serves as a marker of insulin sensitivity [134]. Overall, a high fiber intake was associated with a lower risk of type 2 diabetes [135,136] (Figure 2).
The effects of fiber on glycemic control is also influenced by the fiber viscosity [137,138]. For example, administration of psyllium, a soluble fiber, improved lipid profile and glycemic control [139]. In general, the soluble fiber has a stronger beneficial effect on T2DM compared to insoluble or non-viscous fiber. This may be due to the action of gut bacteria and its byproducts on the fiber substrate and their metabolic functions [140]. In addition, compared to insoluble fibers that are poorly fermentable, and have an important role in increasing the rate of intestinal transit, soluble fibers are highly fermentable, and are efficiently used by the gut microbiota [141]. Specifically, soluble dietary fibers have been shown to promote diversity of gut microbiota and serve as one of the most important substrates for gut microbiota [140]. Following fermentation, different metabolites are generated, including short-chain fatty acids [142], with the highest proportion (60%) being acetate, followed by propionate (25%) and butyrate (15%). Other generating byproducts also include gases such as methane and carbon dioxide [5]. As previously mentioned, SCFA are used as substrates for the metabolism of lipids, glucose and cholesterol and have a significant role in maintaining tissue barrier function and regulating gene expression and immunoregulation. They also provide energy support for colonocytes and regulate homeostasis of the colon, by maintaining the integrity of the intestinal mucosa and reducing inflammation, but also by promoting epithelial cell proliferation and differentiation and water absorption [5]. For example, when compared to cellulose, an insoluble fiber, inulin, which is a soluble fiber, provided significant protection against high fat diet-induced metabolic syndrome [143]. These protective mechanisms included enterocyte proliferation, anti-microbial gene expression and increased IL-22 expression, which improved low grade inflammation and prevented proliferation of unfavorable microbiota [143]. Therefore, dietary fiber represents an ideal source of carbohydrates accessible to the gut microbiota, that can be used to provide the host with energy and carbon sources [79]. Dietary fibers are also called prebiotics and they selectively stimulate the growth or activity of certain microorganisms [144]. The best-known sources of prebiotics are unrefined barley and oats, soy, inulins, but also non-digestible oligosaccharides, such as fructans, polydextrose, fructooligosaccharides, galactooligosaccharides, xylooligosaccharides and arabinooligosaccharides [145]. The high intake of dietary fiber is associated with the increase in the diversity of the gut microbiota, characterized mainly by the growth of Bacteroidetes and Prevotella spp., but also with the improvement of insulin resistance, the decrease of susceptibility to infections and malignant processes [146]. On the other hand, the lack of dietary fiber has contrasting effects, promoting decreased microbiota diversity [147], while also decreasing the production of butyrate, worsening insulin resistance, and increasing susceptibility to infections [148,149]. Importantly, the gut microbiota whose abundance is increased by dietary fibers ameliorate T2DM [145,150]. For example, Bifidobacterium spp. and other SCFA-related genera were shown to be increased while also enhancing GLP-1 secretion and improving HgbA1c levels [150] while harmful bacterial metabolites such as hydrogen sulfide and indole were reduced. Further, a positive correlation was observed between an increased amount of Roseburia, Lachnospira and Prevotella, and an increased level of short-chain fatty acids with a high intake of dietary fiber, with negative correlations with Ruminococcus and Streptococcus [151]. Similarly, a direct association between dietary fiber intake and gut microbiota diversity has also been observed in overweight pregnant women in which dietary fiber intake also decreased abundance of Bacteroides [152]. These findings are consistent with other studies showing that date consumption, which contains high amounts of dietary fibers, increases the abundance of Bifidobacterium, while having the opposite effect on Bacteroides spp. [153]. At the phylum level, the overall abundance of Bacteroidetes is increased thereby improving the Firmicutes to Bacteroidetes ratio [146]. Taken together, there is strong evidence for the role of dietary fibers in promoting beneficial effects of gut microbiota and T2DM.
4.1.2. Starch, Gut Microbiota and T2DM
Starch provides approximately 20 to 40% of the energy requirements for most people and is classified according to the degree of enzymatic hydrolysis [154]. Some rapid digestible starch are hydrolyzed in less than 20 minutes of enzymatic digestion [154] while slow digestible starch are absorbed in the small intestine after approximately 100 minutes of enzymatic digestion. The resistant starch is not hydrolyzed even after 120 minutes of enzymatic incubation [155]. Resistant starch is considered dietary fiber found both naturally in cereals, fruits and vegetables, but also may be added into processed foods [156]. Certain resistant starches are considered prebiotics as well, with positive effects in the prevention or even improvement of metabolic diseases, including metabolic syndrome and T2DM [157]. There are currently five types of known resistant starch: type 1 resistant starch, found in whole grain or coarsely ground bread and durum wheat pasta; type 2 found in negated potato starch, green banana starch, gingko starch and corn starch; type 3. amylose and retrograded starch; type 4, a chemically modified starch; and type 5, an amylose-lipid complex [157]. Foods such as potatoes, rice, pasta, and breakfast cereals contain less than 2.5% resistant starch. On the other hand, certain foods such as boiled legumes and peas, but also other cooked and cooled starchy foods contain a higher amount of resistant starch (5-15%) [157]. The western diet contains mostly foods with a low content of resistant starch.
A growing number of studies suggest the importance of resistant starch in reducing the risk of type 2 diabetes [158,159,160]. Reductions in postprandial blood glucose have been observed when carbohydrates from a meal were replaced with resistant starch [161]. Furthermore, the results of a meta-analysis reported that resistant starch supplementation is associated with improvement in blood glucose, insulinemia, insulin sensitivity and resistance, especially in patients with diabetes and overweight or obesity [160]. More specifically, Type 1 and type 2 resistant starches have been associated with improved postprandial blood glucose, and in addition, type 2 resistant starch has been associated with improved postprandial insulin response and fasting blood glucose [162]. Use of type 3 resistant starch for 3 to 11 weeks resulted in reduction in fasting blood glucose, triglycerides, and total cholesterol in a diabetic mice model [163]. Similarly, type 4 resistant starch introduction significantly reduced post-prandial glucose by 33% [164]. Importantly, the glycemic response following starch intake varied, with lower glycemic and insulinemia responses after raw starch intake, compared to cooked starch [165].
Furthermore, it is well-documented that resistant starch, similar to dietary fibers, is also important in supporting the gut microbiota, through fermentation reactions mediated by resident bacteria which may explain its effects on improving insulin resistance, reduced glucose absorption and glucose homeostasis [166]. For example, consumption of resistant starch type 4 led to an increase in the abundance of Actinobacteria and Bacteroidetes and a decrease in the abundance of Firmicutes [167]. Further, consumption of type 4 resistant starch increased Bifidobacterium adolescentis and Parabacteroides distasonis, while type 2 resistant starch led to an increase in the abundance of Ruminococcus bromii and Eubacterium rectale [167]. Similarly, a study carried out in overweight men showed an increase in the abundance of the same species, Ruminococcus bromii and Eubacterium rectale, in men who consumed diets high in resistant starch [168]. Overall, the data support the exceptional ability of Ruminococcus bromii to degrade resistant starch, due to its carbohydrate active enzyme activity, and in turn, starch serves as a nutrient to increase its abundance [169,170]. These gut microbial changes caused by the resistant starch along with increased in Akkermansia were associated with concomitant benefits in metabolic parameters, including decreased LDL, increased GLP-1 secretion, acetate, and early phase insulin secretion [171].
4.2. Proteins, Gut Microbiota and T2DM
The major functional and structural component of body cells is protein [172]. The current recommendation for protein intake for healthy individuals is 0.8 grams/kg body weight per day or 10-35% of the total energy intake [173] with no difference for patients with diabetes. However, in diabetes complications such as diabetic nephropathy, the recommendation is to reduce protein intake [172,174]. Dietary proteins have received considerable attention for their role in the control of body weight given their demonstrated effects on enhanced satiety and maintenance of lean body mass during weight loss. However, the exact role of proteins in control of diabetes is not as well defined as it is for other macronutrients [104]. Notwithstanding, increase in protein intake has been shown to improve insulin sensitivity by maintaining muscle mass during weight loss in elderly patients with prediabetes or type 2 diabetes [173]. Likewise, a 30% calorie protein diet was associated with improvement of some cardiovascular risk factors though HgbA1c levels were not affected [106]. However, an improvement in insulin sensitivity was observed in a group of obese women who followed a hypocaloric and high protein diet compared to those who followed a hypocaloric and hyperglycemic diet [175], although no changes in blood glucose were observed. Further, an association has been observed between improvement of insulin release and the maintenance of low blood sugar and milk proteins, casein and whey [173]. Moreover, whey proteins are potent stimuli of insulin and incretin secretion such as GLP-1 and GIP that are known to lower blood sugar through stimulation of insulin and inhibition of glucagon secretion, resulting in the inhibition of hepatic glucose production, as well as inhibition of gastric emptying [176,177]. However, other studies did not find a positive association between increase protein intake and lowering the risk of type 2 diabetes, compared to a low protein intake [113], therefore there is a need for more studies to examine the influence of protein on T2DM.
It is important to note that different type of proteins have differing effects on insulin signaling. For example, animal proteins promote insulin resistance and are associated with increased risk for diabetes [178,179,180,181]. On the other hand, plant proteins promote insulin sensitivity [182] and improved glycemic control in patients with type 2 diabetes [183]. Also, increasing the intake of vegetable proteins was associated with a lower probability of relapse of type 2 diabetes [184], and probability of developing type 2 diabetes and its comorbidities [185]. Further, vegetable proteins are the main components of the beneficial Mediterranean diets, while animal proteins, such as red and processed meats are characteristic of the Western diets, which have opposite effects on both gut microbiota and T2DM as described in earlier sections. Therefore, there is strong evidence showing the importance of the type of proteins as it relates to derangement of metabolic parameters.
Similar to other macronutrients, proteins are metabolized by gut microbes into metabolites such as short-chain fatty acids, but also neurotransmitters, amino acid substrates, and organic acids that have physiological effects both locally and systemically [186]. Amino acids are fermented by gut bacteria in the distal colon, and protein fermentation leads to less production of short-chain fatty acids (SCFA) and more production of branched-chain amino acids (BCAA) and potentially toxic substrates, such as ammonia, when compared to carbohydrate fermentation [5]. BCAA include leucine, isoleucine and valine, and are not naturally synthesized in humans, making them nutritionally essential, with their most common food source being proteins. Their role in insulin resistance is thought to be 3-fold via activation of mTORc1 uncoupling of IRS-1, mitochondrial dysfunction through toxic accumulation of BCAA and altered expression of the following genes in humans (BCKDHA, PPM1K, IVD and KLF15) contributing to altered insulin signaling and therefore resistance [100]. Importantly, it has been well documented that animal proteins are metabolized into a higher content of BCAA than plant proteins, which can explain the differences in insulin resistance between the two protein types [187]. In turn, dietary proteins contribute to changes in microbial composition. For example, a high protein diet was associated with a reduction in the abundance of propionate and butyrate-producing bacteria such as Akkermansia, Faecalibacterium, Roseburia, Eubacterium, while increasing abundance of Escherichia, Shigella, Enterococcus, and Streptococcus [188,189] in a rodent model. These findings are consistent with data showing decrease in fecal butyrate, but not propionate or acetate in response to high protein intake in humans [190] but see also [191] where no significant results were seen although the study populations differed between the studies (overweight vs. endurance athletes).
Additionally, increased abundance of Clostridium, unnamed Clostridiales and Allobaculum and a decreased relative concentrations of Eubacterium, Akkermansia, Mucispirillum, Ruminococcus, Johnsonella, Alistipes, Butyrivibrio, and Blautia were also observed after high protein intake [192]. Similar increase in the abundance of Bacteroidaceae was observed at a high protein intake, given that nitrogen from dietary proteins promotes increase in Bacteroidaceae [193]. Increased nitrogen production from gut microbiota and resulting reactive nitrogen species generally relates to growth of unfavorable microbial genera, as SCFA, particularly butyrate, have been shown to limit its production [194]. Further, a high-protein hypoglycemic diet decreased amounts of Roseburia and Eubacterium rectale which correlated with a decrease in fecal butyrate [195]. In addition to decreased SCFA, the increase in trimethylamine N-oxide (TMAO), a bacterial byproduct with proatherogenic and pro-diabetic effects, that is positively correlated with increased concentrations of anaerobic bacteria such as Bacteroides, Alistipes and Bilophila as well as animal products, including red meat sources of protein [79,196]. An increase in the abundance of Bacteroides and Clostridia and a decrease in the abundance of Bifidobacterium adolescentis have also been seen in individuals consuming a diet rich in beef compared to those who did not consume meat [197]. Similar changes in the microbiota composition following protein consumption have been reported at the phylum and class level. For example, high abundance in Firmicutes, especially Clostridia and Bacilli, was observed in rats fed beef, pork or fish protein. An increase in the abundance of Bacteroidetes was seen in rats fed soy protein, while a decrease in Bacteroidetes abundance was noticed in rats fed fish protein [198]. Compared to animal proteins, plant proteins increase Bifidobacterium and Lactobacillus but also decrease the amount of Bacteroides fragilis and Clostridium perfringens [5]. For example, soy proteins have been associated with promoting Bifidobacterium, Lactobacillus, Butyricicoccus, Parabacteroides, Lachnospiraceae and Akkermansia muciniphila [199]. Collectively, these studies demonstrate the intricate relationship between protein intake, changes in the gut microbiota composition and how these taxonomical shifts may influence hyperglycemia and T2DM related parameters.
4.3. Lipids, Gut Microbiota and T2DM
Lipids are considered naturally occurring compounds composed of fatty acids or related derivates, that are soluble in organic solvents, but insoluble in water [200]. Lipid intake has somewhat controversial direct effects on blood glucose [104], though it may influence insulin sensitivity [201]. The current nutritional recommendations are to decrease consumption of saturated lipids and trans fatty acids and increase monounsaturated and polyunsaturated fatty acids [202]. It has been consistently shown that diets high in fats increase the risk of type 2 diabetes through impaired glucose tolerance and binding of insulin to its receptors, resulting in altered glucose transport and accumulation of triglycerides in skeletal muscles [203]. By contrast, a hypolipidemic diet was had no effect on the incidence of diabetes after 8 years, compared to a control diet [204], even when compared to low carbohydrate diets [205]. Therefore, the type and quality of lipids consumed are more important in the risk of developing T2DM [203].
Similar to proteins, lipids from plant sources provide better health benefits than lipids from animal sources [206], and a high intake of plant-based lipids has been associated with a significantly lower risk for the occurrence of T2DM [113]. Results from a meta-analysis report inverse correlation between the incidence of T2DM and high intake of vegetable-based lipids, especially plant-derived α-linolenic acid and for polyunsaturated fatty acids [202]. Conversely, the intake of trans fatty acids has been associated with all-cause mortality, T2DM and ischemic heart disease [207]. Similarly, saturated fats cause unfavorable changes in energy balance, insulin resistance, and fat-cell differentiation [208], though all-cause mortality was not shown to be increased the same way it was after trans-fat consumption [207]. Interestingly, replacing dietary saturated fat with omega-6 polyunsaturated fatty acids resulted in a lower risk of diabetes and related sequelae [209]. A higher intake of omega-6 polyunsaturated fatty acids was associated with a lower risk for diabetes [210]. More specifically, replacing saturated fatty acids with polyunsaturated fatty acids (PUFA) was associated with a 35% decrease in the risk of developing T2DM, and replacing trans fatty acids with PUFA correlated with a 40% reduction in the same parameter [211]. Therefore, the most beneficial lipids in promoting better insulin sensitivity are PUFA, with omega-6 and omega-3 PUFA, being extensively studied [212]. A recent meta-analysis of 67 studies showed that increased supplementation of omega-3 PUFA decreased the risk of developing T2DM [212]. Importantly, the mechanisms of the beneficial effects of PUFA involves gut microbiota and related decreases in production of inflammatory mediators [213]. Specifically, studies have demonstrated increased Bifidobacterium, Bacteroidetes to Firmicutes ratio, fecal SCFA, concomitantly with attenuation of high fat-diet induced insulin resistance and liver inflammation following omega-3 PUFA introduction [214]. Inflammatory markers such as IL-1β, TNF-α, IL-8, IL-6 and interferon-γ were significantly reduced following omega-3 supplementation which also improved fat accumulation and metabolic parameters [215].
Conversely, increased consumption of dietary lipids, particularly saturated fatty acids and trans fats influences the composition of the gut microbiota negatively, decreasing bacterial diversity [216] and increasing Firmicutes/Bacteroidetes ratio [216,217]. These effects also contribute to the development of obesity through leptin resistance and promotion of low-grade systemic inflammation through the LPS/TLR4 pathway, that are characteristics of dysbiosis [218,219]. A decrease in bacterial diversity and an increase in the abundance of Faecalibacterium prausnitzii was also observed following increased consumption of saturated fats [152,220,221]. Also, increased intake of fats is also associated with an increase in the abundance of Rikenellaceae and Bacteroides and other anaerobic genera [79,222]. Similarly, a high intake of trans fatty acids was associated with a decrease in the abundance of Bacteroidetes and an increase in the abundance of Proteobacteria and Desulfovibrionaceae [223]. The opposite is also true, with low intake of these unfavorable lipids resulted in relatively increased abundance of beneficial bacteria such as Bifidobacterium, but also with improvements in glycemia and total cholesterol [220]. For example, an increase in the abundance of Bifidobacterium and Adlercreutzia, Lactobacillus and Streptococcus and Akkermansia muciniphila was shown in mice given fish oil and an increase in the abundance of Bacteroides and Bilophila in mice given lard, which aggravates white adipose tissue inflammation [224]. Overall, dietary lipids serve as an important macronutrient in modulating gut microbiota composition and metabolic parameters underlying T2DM.
5. Effects of Micronutrients on T2DM and Gut Microbiota
Micronutrients, including vitamins and minerals, have also been heavily implicated in gut microbial remodeling and glucose homeostasis. In the following subsections, we describe the role of various vitamins and minerals in modulating these processes.
5.1. Vitamins, Gut Microbiota and T2DM
Vitamins are organic substances classified as either fat soluble (vitamin A, D, E, K) or water soluble (vitamin B complex, C) with antioxidant vitamins such as A, C, D and E shown to be decreased in individuals with diabetes, while vitamin D deficiency is associated with the development of diabetes and its sequelae [225,226]. For example, 6-month vitamin D supplementation improved HgbA1c, with decreased production of oxidation products and oxygen free radicals [227]. Similarly, high dose cholecalciferol, the active form of Vitamin D, improved clinical manifestations of cutaneous microcirculation, inflammatory markers and peripheral neuropathy [228]. Supplementation also decreases inflammatory gene expression, particularly of IL-6, IL-10, IFN-γ, which serve potential benefit in protecting against T2DM development and disease progression through their roles in influencing platelet mediated inflammation [229,230]. Further, cholecalciferol decreases insulin resistance through increased activity of insulin receptors and enhanced expression of PPAR-γ [231]. This is also demonstrated in individuals with obesity or overweight where supplementation reduced fasting blood glucose, decreased truncal subcutaneous fat and reversed to normoglycemia [232]. Gut microbiota, vitamin D and the development of metabolic diseases, including T2DM [233], has been described as a three-way axis with vitamin D anti-inflammatory actions playing a central role. Particularly, vitamin D binding to the Vitamin D receptor (VDR) influences gut microbial composition, with findings showing increased Bifidobacterium and Akkermansia species, which exert anti-inflammatory effects and improve insulin resistance [234]. The VDR can degrade lithocholic acid, a secondary bile acid, as well as regulate enzymes that mediate cholesterol conversion into bile acids [235]. Interestingly, animal studies have shown that Vitamin D upregulates tight junction proteins to improve intestinal barrier integrity, reduced LPS production and suppressed hyperinsulinemia, hyperglycemia and neuroinflammation [236]. Vitamin A supplementation has also been shown to exert beneficial effects in modulating microbiota, enhancing pancreatic β cell activity, and reducing inflammatory responses [237], though the literature is not as robust as compared to that on vitamin D.
Vitamin K is another fat-soluble vitamin whose metabolism has been intricately related with gut microbiota, with recent meta-analyses demonstrating its benefits in T2DM risk [238], specifically through improvement in fasting blood glucose and insulin resistance scores. Recent studies have shown the importance of Vitamin K2 supplementation in improving glycemic homeostasis and insulin sensitivity in T2DM via gut microbiota [239]. Specifically, after 6 months of vitamin K2 supplementation there were significant reductions in fasting serum glucose, insulin and HbA1c levels in patients with T2DM and significant glucose tolerance improvement in diet-induced obesity mice. In addition, increased concentrations of secondary bile acids (lithocholic and taurodeoxycholic acid) and short-chain fatty acids (acetic acid, butyric acid, and valeric acid) were found in human and mouse feces that were accompanied by an increased abundance of the genera responsible for the biosynthesis of these metabolites. Further fecal microbiota transplant of these microbiota into high-fat diet induced obesity rat model showed significant improvements in glucose tolerance through activation of colonic bile acid receptors, increased GLP-1, and anti-inflammatory effects [239] (Figure 3).
Water-soluble vitamins have also been implicated in various facets related to T2DM [240]. For example, patients with prediabetes and T2DM have a higher vitamin C [240] requirement than those without, thus dietary approaches may help with treatment of the condition. Further, treatment with metformin, currently the first-line treatment of T2DM, has been shown to cause vitamin B12 deficiency [241]. Recent study showed that metformin can assist gut microbiota in contributing to this deficiency [242]. Using functional and genomic analyses via high-throughput screens in E. coli and C. elegans it was shown that metformin helps bacteria gather B12 from the environment by increasing the expressions of B12 transporter genes thus reducing the B12 supply to T2D patients taking the drug over time [242]. It is important to note that Vitamin B12 deficiency can further exacerbate sequelae of T2DM such as peripheral neuropathy and even macrovascular complications in patients with the condition [243]. Therefore, B12 supplementation is important in patients on metformin treatment. Collectively, there is strong evidence showing the impact of vitamins on gut microbiota and associated T2DM metabolic abnormalities (Figure 3).
5.1. Minerals, Gut Microbiota and T2DM
Minerals, including zinc, calcium, selenium, potassium, magnesium, copper and iron are found in various food groups that serve an essential role in health, as well as glucose metabolism [244]. For example, zinc has been shown to have important effects in activating cell signaling pathway that can prolong the action of insulin and modulate insulin receptors [245]. Specifically, zinc is highly involved in insulin processing, storage, and secretion in pancreatic β-cells, with two zinc molecules required to coordinate these processes [246]. Further, zinc serves as an important antioxidant that improves markers of diabetes mellitus through reduction of reactive oxygen species (ROS) [247]. Some of these processes may also be mediated by gut microbiota, as zinc deficiency has been shown to promote unfavorable effects on microbial composition and inflammatory markers [248]. Commensal bacterial species work to increase bioavailability of zinc and iron, while pathogenic species promote the opposite [249], therefore dysbiosis in T2DM that increases unfavorable gut microbial composition may, to a certain extent, be attributed to zinc and iron deficiency [250].
Iron is the most abundant trace metal in the human body and has also been intricately related with glucose metabolism [251]. Pancreatic β-cells are heavily involved in controlling iron homeostasis through release of hepcidin, which binds transferrin, the molecule that transports iron in the blood [252]. Further, iron deficiency is correlated with impaired insulin release [253]. The Fenton reaction, which generates radical species from iron, can amplify glucose-induced insulin secretion [254], however, when ROS species accumulate in states of iron overload, these redox reactions can lead to insulin resistance and pancreatic β-cells apoptosis [255]. As such, iron supplementation has been shown to have negative effects on gut microbial composition, with observed decreases in Lactobacillus and Bifidobacterium, with relative increases in pro-inflammatory Escherichia coli [256]. Therefore, low-iron diets can protect from the development of metabolic disease through improved TGF-β signaling [257]. Similarly, trace elements such as copper, when in excess, have similar negative effects through production of reactive oxygen species, promoting insulin resistance [245], though the effects of copper on gut microbiota are not well understood.
Calcium is the most abundant mineral in humans with calcium signaling influencing insulin secretion and resistance through its systemic importance and associations with other vitamin and mineral deficiencies such as hypomagnesia and hypovitaminosis D [258]. Study findings have shown that hypocalcemia and hypomagnesia is correlated with increased plasma blood glucose and HgbA1c in humans [259]. A favorable gut microbiome is essential for optimal calcium absorption, with SCFA production found to lower pH in the colon, making calcium more soluble and therefore absorbable [244]. The interrelationship between gut microbiota and magnesium is similar, with magnesium supplementation promoting Lactobacillus spp. production and reciprocally, resulting SCFA production can improve mineral absorption (Figure 3). Therefore, states of dysbiosis, such as T2DM, contribute to mineral deficiencies through impairing mineral metabolism, absorption, and other important processes [260].
6. Food Groups and Related Effects on Gut Microbiota and T2DM
Considering the effects of macro and micronutrients on gut microbiota and T2DM discussed above, food groups and dietary patterns are major determinants of the gut microbiota-metabolic disorder axis. Therefore, in the following sections we present the interrelations between food groups (cereals, whole grains, fruits, vegetables, dairy products, meat products and oil seeds) gut microbiota and T2DM and how food patterns and adherence to certain diets modulate the relative abundance of several gut microbiota taxa and their impact on physiologic, metabolic and hormonal functions that impinge on the prevention, development and management of T2DM (Table 1).
6.1. Cereals and Cereal Products
Cereals are composed of whole grains, wheat, oats, rye and barley, which have shown to have beneficial effects on metabolic health and gut microbiota [261,262]. Over the years, increasing number of studies have elucidated the effects of cereals products on T2DM, largely showing that the components within cereals decrease risk of developing the disease and related sequelae [263,264,265,266]. Dietary recommendations for cereals in metabolic disease include increasing the intake of cereals with whole grains and limiting the intake of refined grains or cereals with processed sugars and artificial sweeteners [267]. Both whole wheat intake and intake of barley, oat and rye have been associated with improved blood glucose levels [268,269] and increased insulin sensitivity [270]. It should be noted that the beneficial effects of cereals appear when the intake is high, at least 4 grams of β-glucans daily [269,271]. For example, a supplement of up to 50 grams of whole grains per day was associated with a 25% decrease in the risk of T2DM [272]. Similarly, the consumption of two servings of whole grains per day was associated with a 21% decrease in the risk of T2DM [273] while refined grain intake of 200-400 grams per day was associated with a 6-14% increase in the risk of T2DM [272]. The composition of the whole grains such as magnesium and phytochemicals, isoflavins and lignins were also associated with beneficial effects in T2DM [274]. Adding cereal fiber to meals, reduced post-prandial insulin release, indicating the important roles of fiber in improving insulin sensitivity [266]. Taken together, this data provides strong evidence for cereal products in reducing the risk of T2DM development.
Further, cereal based dietary approaches are shown to affect multiple metabolic parameters in individuals already diagnosed with T2DM, some of which may be related to changes in gut microbiota. For example, after 3-month adherence to high dietary fiber-based cereal there were favorable trends in lipids, HgbA1, body mass index (BMI), adipose distribution and fasting C-peptide levels [263]. Similarly, high fiber rye, a component of healthy cereal, is shown to improve similar parameters when compared to refined wheats [264]. In addition to improving metabolic parameters, the high fiber rye diet produced important changes in gut microbiota, including elevated SCFA- producing Agathobacter and decreased Ruminococcus torques, with associated increases in plasma butyrate concentrations [264]. When compared to refined grains, the whole grain, has immunomodulatory effects that were associated with microbial composition alterations [275]. For example, introduction of wheat grains after a 2 week Western-style diet, improved SCFA, increased SCFA-producing Lachnospira and reduced pro-inflammatory Enterobacteriaceae family which correlated with positive changes in effector memory T cell activity and acute innate immune response [275]. Other immunomodulatory effects of cereals have also been described in the literature, with reduced activity of pro-inflammatory cytokines, TNF-α and IL-6, being observed after consumption [276,277]. In rodent studies, the effects of wheat also improved GPR41/43 receptor expression and enhanced GLP-1 secretion with concomitant increases in SCFA-producing bacteria, providing further insights into the multitude effects that cereals have on metabolic disease [278].
In addition to the changes described above, it seems that a general increase in Bifidobacterium and Lactobacillus spp. are common after cereal consumption, an effect consistent with other recent studies [265]. Previous studies have also showed that diets rich in whole wheat compared to refined wheats exhibit abundance of Bifidobacterium and relative decreases in Bacteroides after 12-week intervention [279]. The type of cereal consumed is also important in determining microbiota shifts. For example, an increase in the abundance of Bifidobacterium and Lactobacillus was seen in the gut microbiota of people who consumed whole grain cereals for breakfast, compared to the microbiota of people who consumed cereals based on wheat bran [280]. A corn-based cereal diet increased abundance of fecal Bifidobacteirum after a 3-week intervention, as compared to a refined-corn based cereal [281]. These changes in the composition of the gut microbiota could be observed even at a low intake of whole corn (29.6% of the recommended total of 48 g [281]). However, the opposite is also true, with sugar additives and processing seen in refined cereals, shown to have negative effects on both gut microbial composition and related metabolites [282]. Therefore, it is evident that eating cereals with naturally occurring fibers can be beneficial in preventing or treating metabolic derangements in T2DM, while avoiding refined cereals and cereals with additives is also important.
6.2. Fruits and Vegetables
In general, some of the healthiest foods are considered vegetables and fruits, due to their content in dietary fiber, vitamins, minerals, and flavonoids [283]. Multiple studies have demonstrated the inverse associations between consumption of green leafy vegetables and the risk of developing T2DM [67,284,285], the consumption of fruits and T2DM [286], as well as the intake of mixed fruits and vegetables and T2DM [287]. Specifically, an intake of 0.2 servings per day of green leafy vegetables reduced the risk for type 2 diabetes by 13% [284], with similar findings in another meta-analysis showing risk reduction of 14% [285]. Changes in microbial shifts after consumption of fruits and vegetables have also been described with study findings showing decreased abundance of the Lachnospiraceae family, including Ruminococcus, and increased concentrations of Faecalibacterium and Lactobacillus [288]. Further metagenomic sequencing studies combining two large human cohorts have shown changes that include increased abundance of Faecalibacterium prausnitzii, Akkermansia muciniphila, Ruminococcaceae, Clostridiales, and Acidaminococcus and with decrease in the abundance of Fusobacterium [289].
Vegetables and fruits are sources of antioxidants that have been associated with augmenting glucose metabolism, by improving oxidative stress [290], particularly given their high content of flavonoids and polyphenols [291]. Interestingly, flavonoids are shown to modulate gut microbiota-related metabolic processes, particularly through suppression of lipogenesis and up-regulation of lipolysis, via the FXR pathway in bile acid metabolism [292]. These effects of flavonoids were corelated with increased Akkermansia, and reductions in Lachnoclostridium, Desulfovibrio, Colidextribacter and Blautia, all of which are strongly associated with metabolic parameters [292]. Further, flavonoid based dietary interventions alleviated inflammation as measured through LPS/TLR-4, TNF-α, IL-6, and IL-10, while also improving insulin resistance, HgbA1c and oral glucose tolerance [293]. Interestingly, GLP-1 release was also enhanced following flavonoid introduction. The beneficial effects of fruit and vegetable flavonoids also are shown by improvement of intestinal barrier integrity, as well as promoting islet cell proliferation and suppression of islet cell apoptosis [294]. Flavonoids also modulate glucose metabolism by up-regulation of IRS/AKT signaling pathway to increase GLUT4 translocation and synthesis of glycogen, while concomitantly improving the Firmicutes to Bacteroidetes ratio [295]. As such, flavonoids, a major component of fruits and vegetables, exerts a multitude of metabolic benefits at the intersection between gut microbiota and glucose homeostasis.
In addition to flavonoids, fruits and vegetables are comprised of other beneficial bioactive phytochemical-based nutrients, including vitamin C and carotenoids, which contribute to insulin sensitivity [296,297]. Also, green leafy vegetables contain magnesium which is inversely associated with increased risk for type 2 diabetes [298]. The association between fruit and vegetable intake and reduced risk of type 2 diabetes may be due to their dietary fiber content [299], and subsequent effects on weight loss in overweight individuals [300]. Fruit and vegetable juices, depending on their content, may have differing outcomes on both gut microbiota and T2DM [301,302,303,304]. Fruit juices that are altered by added sugar or artificial sweeteners pose harmful risks to the gut metabolic profile [301]. For example, artificial sweetening of fruit beverages results in modest change in gut microbiota, particularly in the ratio of Firmicutes to Bacteroidetes [301]. However, introduction of natural fruit or vegetable extracts or juice generally have favorable effects [302,303,304]. In a pre-diabetic rodent model, blueberry juice improved microbiota composition as well as metabolic parameters including insulin signaling, inflammation, ketogenesis and fatty acid oxidation [303]. Similarly, pomegranate juice can reduce post-prandial glycemic response after eating a high carbohydrate meal, primarily breads [305]. Overall, fruits and vegetables are an important food group in maintaining a healthy microbiota profile since diets high in fruits, vegetables, legumes, and whole grains are accompanied by optimal body weight, reduced inflammation, and lower insulin resistance.
6.3. Milk and Dairy Products
Dairy products are rich in protein, B vitamins, and minerals, such as calcium, magnesium, potassium, phosphorus, and zinc, all of which having important effects on gut microbiota composition [306]. Dairy proteins, especially whey proteins, are associated with improved insulin sensitivity, and a reduced risk of type 2 diabetes [307]. Interestingly, high quantities of dairy consumption (2 servings per day) in adolescence were associated with a 38% decreased risk of developing T2DM in middle-aged women [308]. Further, an inverse correlation was observed between the intake of skimmed or semi-skimmed dairy products and the risk of type 2 diabetes [309]. This decreased risk was seen with 200 grams (g) of skimmed dairy product intake, with improvement of risk up to 6% with every additional 200g, up to a daily total of 600g [272]. Another study has shown that 1 serving of dairy per day has beneficial effects on T2DM risk reduction of 9% in men and 4% in women [310,311]. Dairy consumption produces specific compositional changes in gut microbiota. For example, introduction of dairy products or intake of yogurt for three weeks led to decreased Bacteroides fragilis [306] and an abundant growth of Lactobacillus and Bifidobacterium [312]. Similarly, consumption of kefir, a yogurt-based drink over the next 4 weeks, increased abundance of Lactobacillus [313,314], with associated elevated levels in fecal SCFA [314]. Interestingly, in studies on murine models, yogurt-derived Lactobacillus plantarum has been shown to ameliorate the reduction in pancreatic β-cell mass with notable improvements in insulin resistance [315]. Taken together, these studies show that dairy consumption prompt significant changes in the composition of gut microbiota that are beneficial to the host in mitigating the deleterious effects of T2DM.
6.3. Meat and Meat Products
The recommendations for patients with type 2 diabetes regarding the intake of meat and meat products are similar to the recommendations for healthy individuals, i.e., one portion/day or the equivalent of 100-150 grams of lean meat per day [316]. Lean meat and meat products are sources of protein with high biological value, but also important sources of iron and vitamin B12 [317]. However, red meats are shown to exert negative effects on both contributing to T2DM development and worsening of the condition [316]. Several positive associations have been reported between the intake of processed red meats and increased blood glucose concentrations, insulin levels, and risk for obesity [318,319]. Moreover, the risk for type 2 diabetes was associated with the intake of red meat up to 100 grams per day [272], but also with the intake of up to 50 grams per day of processed meat products [272,320]. These effects have been attributed to the content of heterocyclic amines and nitrates affecting glucose metabolism [321,322]. These metabolites contribute to insulin resistance through adverse effects on pancreatic β cell function and insulin-like growth factor (IGF-1) [323]. Further, these inorganic nitrates, present in processed meats, promote DNA damage through conversion to cytotoxic agents such as peroxy-nitrite as well as reactive oxygen species, which increase pro-inflammatory cytokine production and hamper glucose homeostasis [324]. Red meats also enhance the presence of dietary advanced glycosylated end products (dAGE), the result of the Maillard reaction that occurs between amino acids and reducing sugars [325]. These dAGE products are shown to increase insulin resistance, while restricted intake of dietary glycoxidation products improve insulin sensitivity in diabetic mice [326]. Additionally, it has been shown that hyperglycemia further enhances the glycation process, thus worsening complications of uncontrolled diabetes. Therefore, red meats are a source of inorganic nitrates and substrates for generation of dAGE, which may contribute to development of insulin resistance and complicate pre-existing diabetes.
Red meat may also be detrimental to gut microbial composition. It has been shown that red meat decreases Lactobacillus, Paralactobacillus, Prevotella, while also decreasing SCFA in animal models [327]. Further, administration of beef, red meat derivate, in mouse and rat study models led to an increase in the amount of Clostridium and Blautia and a decrease in the amount of Bifidobacterium and Akkermansia [328]. Addition of butyrate containing starch was shown to reverse the negative effects of red meat diet adherence through increased abundances of Clostridium coccoides, Clostridium leptum, Lactobacillus spp, Parabacteroides distasonis, Ruminococcus bromii, but also to a decrease in the amount of Ruminococcus torques, Ruminococcus gnavus, Escherichia Coli [329]. However, the effects on gut microbial composition are dependent of the type of meat and proteins they contain [330]. For example, a study evaluating the gut microbiota of individuals consuming chicken meat is characterized by the highest proportion of Prevotella 9 (22.45%), followed by Dialister, Faecalibacterium, Megamonas, Prevotella, Roseburia, Alloprevotella, Ruminococcaceae, Eubacterium and Succinivibrio, while the gut microbiota of individuals consuming pork is characterized by the highest proportion of Bacteroides (17.3%), followed by Faecalibacterium, Roseburia, Dialister, Ruminococcus, Blautia, Megamonas, Agathobacter, Subdoligranulum and Eubacterium [331]. On the other hand, pork intake, decreased the amount of Blautia, Bifidobacterium and Alistipes, and increased the amount of Akkermansia muciniphila and Ruminococcaceae [332]. Collectively, intake of pork meat induced low-grade inflammation, induced oxidative stress and upregulation of lipid metabolism genes such as PPAR-α and PPAR-γ [332]. Further, an increase in the abundance of Lactobacillus and a decrease in SCFA levels and SCFA-producing bacterial species such as Fusobacterium, Bacteroides and Prevotella have been reported in laboratory mice fed beef, pork or fish proteins, compared to mice that were given protein from sources other than meat, such as soy or casein [333]. Similarly, laboratory rats fed chicken meat had the highest abundance of Lactobacillus, compared to laboratory rats fed soy and had the highest abundance of Ruminococcus and the lowest abundance of Lactobacillus [334]. The results of a systematic review showed that the administration of beef in mouse and rat study models led to an increase in the amount of Clostridium and Blautia and a decrease in the amount of Bifidobacterium and Akkermansia [328]. Collectively, these changes indicate that meats derived from chicken have more favorable effects on gut microbiota and insulin resistance as compared to pork and red meats.
6.5. Nuts, Oils and Oilseeds
Tree nuts have been shown to exert favorable effects on gut microbiota and metabolic parameters [335,336]. For example, replacing starchy foods with peanuts or almonds in patients with type 2 diabetes led to improvements in blood glucose, HgbA1c and inflammatory markers [335]. In addition, daily intake of raw or roasted almonds for 4 weeks promoted Bifidobacterium spp. and Lactobacillus spp. and inhibited the growth of Enterococcus spp. Interestingly, administration of raw almonds had a greater Bifidobacteria-promoting effect than roasted almonds, with both roasted and raw almonds having a potential prebiotic effect, including regulating gut bacteria and improving metabolic activities [336]. Similarly, nut intake promotes an increase in the abundance of Faecalibacterium, Clostridium, Dialister and Roseburia and a decrease in the abundance of Ruminococcus, Dorea, Oscillopira and Bifidobacterium [337]. Pistachio consumption led to an increase in the abundance of potentially beneficial, butyrate-producing bacteria [338] while eating whole, roasted or chopped almonds is associated with an increase in the abundance of Lachnospira, Roseburia [339]. These alterations in gut microbiota were associated with concomitant decrease in pro-inflammatory secondary bile acid production and LDL cholesterol, two interrelated parameters in the development of hyperglycemia and insulin resistance [337]. Also, a diet enriched with 20% peanut protein was effective in increasing the amount of Bifidobacterium and reducing the amount of Enterobacteria and Clostridium perfringens in rats [340].
Oilseeds are important sources of polyunsaturated and monounsaturated fatty acids [341,342] and their consumption has been associated with decreased risk for type 2 diabetes [343]. For example, dietary flaxseed oil, given their rich composition of omega-3 was associated with decreased Firmicutes and pro-inflammatory markers such as IL-1β, TNF-α, IL-6, increased Bacteroidetes and Alistipes that negatively correlate with LPS production [344]. Further, direct markers of hyperglycemia showed significant improvement, particularly in fasting blood glucose and glycated hemoglobin. Interestingly, superoxide dismutase (SOD) activity was increased as well, with previous studies showing that SOD activity can improve diabetes-induced mitochondrial electron transport dysfunction and diabetes complications such as retinopathy [345]. Meta-analyses of human studies confirm these beneficial anti-inflammatory effects of oilseeds, with decreased CRP and IL-6 activity leading to improved endothelial function and metabolic activity [346]. Oil seeds cause significant changes in the gut composition profile, such as increased Lactobacillus spp. and SCFA, with reduced production of harmful metabolites such as TMAO [347]. As such, oilseeds and nuts serve as a healthy food source that are intricate components of the Mediterranean diet and modulate important metabolic processes associated with T2DM.
7. Conclusions and Perspective
Over the years, substantial evidence has accumulated supporting the influence of dietary changes in modulating gut microbiota in ways that safeguard against or contribute to the development of T2DM. In this review, we identified the roles of key macronutrients, micronutrients, and various food groups in these processes. Particularly, we showed the effects of important microbiota metabolites including SCFA, BCAA, TMAO, secondary bile acids, gut hormones and inflammatory signaling and how nutrients that are associated with the development of T2DM have distinct metabolite profiles that mechanistically lead to or protect against insulin resistance. In general, SCFA-producing species such as Lactobacillus, Faecalibacterium, Akkermansia, Faecalibacterium Eubacterium, are induced by favorable nutrients such as cereals, nuts, oilseeds, fruits, and vegetables, while inflammatory species such as Escherichia coli, Ruminococcus torques and Bacteroides spp. are associated with red meats, fats, and sugary foods. The various food groups discussed are components of common diets such as the MD and WD, which are also shown to have contrasting effects on gut microbiota and development of T2DM. Though important trends, mechanisms and associations have been described regarding this topic, the relationship between nutrients, gut microbiota, hyperglycemia, insulin resistance and the host is complex and new insights are constantly changing our understanding of these processes. Limitations within the studies presented also exist. Increasing a number of human studies have elucidated the mechanisms described within this review, though animal studies serve as important models in demonstrating the associations with gut microbiota, their metabolites and insulin resistance given that 90% of the gut microbiota between mice and humans are deemed to be similar [348]. Still, these murine studies may not translate fully in humans therefore the interpretation should be taken with caution when attempting to generalize these findings. Nevertheless, there is significant data showing that gut microbiota is heavily involved at the intersection of nutrition and T2DM.

Table 1.
Food Groups and resulting effects on T2DM.
| Food Group | Study Period | Outcome measured | Results/Implications | Subject type | Reference |
|---|---|---|---|---|---|
| Cereals and Cereal Products | Meta-Analysis | Diabetes risk | 2 servings of whole grains decreased risk of developing T2DM by 21% Refined grain intake increased risk of developing T2DM by 6-14% |
Humans | [272] |
| 3 months | Metabolic Parameters | Improvements in lipid quality, HgbA1c, BMI, adipose distribution, and fasting C-peptide levels | Humans | [263] | |
| 6 weeks | Gut Microbiota and Inflammatory Markers | Whole grains improved effector memory T cell activity and acute innate immune response Increased quantity of SCFA and SCFA-producing genera including Lachnospira Decreased relative abundances of pro-inflammatory bacterial family Enterobacteriaceae |
Humans | [275] | |
| 9 weeks | Gut Microbiota and Gut Hormones | Increased SCFA-producing species Increased GLP-1 secretion |
Mice | [278] | |
| 12 weeks | Gut Microbiota | Increased Bifidobacterium and decreased Bacteroides | Humans | [279] | |
| Fruits and Vegetables | Meta-Analysis | Diabetes risk | Intake of 0.2 servings per day of green leafy vegetables reduced the risk for type 2 diabetes by 13% | Humans | [284] |
| Gut Microbiota and Metabolic Parameters | Increased Akkermansia Reduced Lachnoclostridium, Desulfovibrio, Colidextribacter and Blautia Up-regulation of lipolysis through the FXR, bile acid metabolism pathway |
Mice | [292] | ||
| Inflammatory Markers and Metabolic Parameters | Reduced LPS/TLR-4 activity, TNF-α, and IL-6 Improved IL-10 Improved insulin resistance and HgbA1c Increased GLP-1 secretion |
Mice | [293] | ||
| Glucose Metabolism and Gut Microbiota | Up-regulation of the IRS/AKT signaling pathway to increase GLUT4 translocation and synthesis of glycogen Improved Firmicutes/Bacteroidetes ratio |
Mice | [295] | ||
| Milk and Dairy Products | Diabetes risk | 1 serving of dairy per day has beneficial effects on T2DM risk reduction of 9% in men and 4% in women | Humans | [310,311] | |
| 3 weeks | Gut Microbiota | Increased Bifidobacterium and Lactobacillus spp. Increased serum IgA Decreased Bacteroides fragilis |
Humans | [306,312] | |
| Gut Microbiota and Pancreatic Function |
Lactobacillus isolated from yogurt increased SCFA levels and SCFA receptors, GPR41/43 Increased SCFA-producing genera Inhibited reduction of β-cell mass |
Mice | [315] | ||
| Meat and Meat Products | Meta-Analysis | Diabetes Risk | Risk for T2DM is increased with intake of 100 grams of red meat per day Risk for T2DM is increased with intake of 50 grams of processed meat per day |
Humans | [272,320] |
| Gut Microbiota | Red meat decreases Lactobacillus, Paralactobacillus, Prevotella, while also decreasing SCFA | Dogs | [327] | ||
| 1-4 weeks | Gut Microbiota | Increased Clostridium and Blautia Decreased Bifidobacterium and Akkermansia |
Mice | [328] | |
| 3 months | Gut Microbiota, Inflammatory and Metabolic parameters | Pork meat decreased Blautia, Bifidobacterium, Alistipes Induced low-grade inflammation Induced oxidative stress Up-regulated lipid metabolism genes including PPAR-α and PPAR-γ |
Mice | [332] | |
| Nuts, Oils and Oilseeds | 3 months | Parameters of T2DM | Peanuts or almonds in patients with T2DM improved blood glucose, HgbA1c and inflammatory markers like IL-6 expression | Humans | [335] |
| 6 weeks | Gut Microbiota and Metabolic Parameters | Nut intake increase in the abundance of Faecalibacterium, Clostridium, Dialister and Roseburia and a decrease in the abundance of Ruminococcus, Dorea, Oscillopira and Bifidobacterium Decreased pro-inflammatory bile acid production and LDL cholesterol |
Humans | [337] | |
| 5 weeks | Gut Microbiota and Inflammatory Parameters |
Dietary flaxseed oil decreased severity of T2DM Improved Firmicutes to Bacteroidetes ratio, while increasing Alistipes Reduction in as IL-1β, TNF-α, IL-6, and LPS production |
Rats | [344] |
Abbreviations: T2DM, Type 2 Diabetes Mellitus, HgbA1c, Hemoglobin A1c; BMI, body mass index; SCFA, short chain fatty acid; GLP-1, glucagon-like peptide 1; FXR, Farsenoid X Receptor; LPS, lipopolysaccharides; TLR-4, Toll-like receptor 4; TNF-α, Tumor Necrosis Factor alpha; IL, interleukin; GLUT4, Glucose Transporter 4; IRS, insulin receptor substrate; IgA, Immunoglobulin A; PPAR, peroxisome proliferator-activated receptor; LDL, low density lipoprotein.
Author Contributions
Conceptualization, S.H., O.C.I., and M.C.; methodology, S.H., O.C.I., and M.C.; investigation, S.H., O.C.I.; writing—original draft preparation, S.H., O.C.I.; writing—review and editing, M.C.; visualization, S.H.; supervision, M.C.; project administration, M.C.; funding acquisition, M.C. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the project: The analysis of interrelationship between gut microbiota and the host with applications in the prevention and control of type 2 diabetes”, co-financed by the European Regional Development Fund through the Competitiveness Operational Program under the contract number 120/16.09.2016.
Conflicts of Interest
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
References
- Salgaço, M.K.; Oliveira, L.G.S.; Costa, G.N.; Bianchi, F.; Sivieri, K. Relationship between gut microbiota, probiotics, and type 2 diabetes mellitus. Appl Microbiol Biotechnol. 2019, 103, 9229–9238. [Google Scholar] [CrossRef] [PubMed]
- Sender, R.; Fuchs, S.; Milo, R. Revised Estimates for the Number of Human and Bacteria Cells in the Body. PLOS Biol. 2016, 14, e1002533. [Google Scholar] [CrossRef]
- Banaszak, M.; Górna, I.; Woźniak, D.; Przysławski, J.; Drzymała-Czyż, S. Association between Gut Dysbiosis and the Occurrence of SIBO, LIBO, SIFO and IMO. Microorganisms 2023, 11, 573. [Google Scholar] [CrossRef] [PubMed]
- Gomaa, E.Z. Human gut microbiota/microbiome in health and diseases: A review. Antonie van Leeuwenhoek 2020, 113, 2019–2040. [Google Scholar] [CrossRef] [PubMed]
- Rinninella, E.; Cintoni, M.; Raoul, P.; Lopetuso, L.R.; Scaldaferri, F.; Pulcini, G.; Miggiano, G.A.D.; Gasbarrini, A.; Mele, M.C. Food Components and Dietary Habits: Keys for a Healthy Gut Microbiota Composition. Nutrients 2019, 11, 2393. [Google Scholar] [CrossRef] [PubMed]
- Rinninella, E.; Raoul, P.; Cintoni, M.; Franceschi, F.; Miggiano, G.A.D.; Gasbarrini, A.; Mele, M.C. What Is the Healthy Gut Microbiota Composition? A Changing Ecosystem across Age, Environment, Diet, and Diseases. Microorganisms 2019, 7, 14. [Google Scholar] [CrossRef] [PubMed]
- Leeming, E.R.; Johnson, A.J.; Spector, T.D.; Le Roy, C.I. Effect of Diet on the Gut Microbiota: Rethinking Intervention Duration. Nutrients 2019, 11, 2862. [Google Scholar] [CrossRef] [PubMed]
- Chambers, E.S.; Byrne, C.S.; Morrison, D.J.; Murphy, K.G.; Preston, T.; Tedford, C.; Garcia-Perez, I.; Fountana, S.; Serrano-Contreras, J.I.; Holmes, E.; et al. Dietary supplementation with inulin-propionate ester or inulin improves insulin sensitivity in adults with overweight and obesity with distinct effects on the gut microbiota, plasma metabolome and systemic inflammatory responses: A randomised cross-over trial. Gut 2019, 68, 1430–1438. [Google Scholar] [CrossRef] [PubMed]
- Pickard, J.M.; Zeng, M.Y.; Caruso, R.; Núñez, G. Gut microbiota: Role in pathogen colonization, immune responses, and inflammatory disease. Immunol. Rev. 2017, 279, 70–89. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Zhang, J.; Cheng, Y.; Zhu, M.; Xiao, Z.; Ruan, G.; Wei, Y. Gut microbiota: A new target for T2DM prevention and treatment. Front. Endocrinol. 2022, 13, 958218. [Google Scholar] [CrossRef] [PubMed]
- Hamamah, S.; Hajnal, A.; Covasa, M. Impact of Nutrition, Microbiota Transplant and Weight Loss Surgery on Dopaminergic Alterations in Parkinson's Disease and Obesity. Int. J. Mol. Sci. 2022, 23, 7503. [Google Scholar] [CrossRef] [PubMed]
- Hamamah, S.; Aghazarian, A.; Nazaryan, A.; Hajnal, A.; Covasa, M. Role of Microbiota-Gut-Brain Axis in Regulating Dopaminergic Signaling. Biomedicines 2022, 10, 436. [Google Scholar] [CrossRef]
- Flaig, B.; Garza, R.; Singh, B.; Hamamah, S.; Covasa, M. Treatment of Dyslipidemia through Targeted Therapy of Gut Microbiota. Nutrients 2023, 15, 228. [Google Scholar] [CrossRef]
- Wu, Y.; Ding, Y.; Tanaka, Y.; Zhang, W. Risk Factors Contributing to Type 2 Diabetes and Recent Advances in the Treatment and Prevention. Int. J. Med Sci. 2014, 11, 1185–1200. [Google Scholar] [CrossRef]
- Khan, M.A.B.; Hashim, M.J.; King, J.K.; Govender, R.D.; Mustafa, H.; Al Kaabi, J. Epidemiology of Type 2 Diabetes–Global Burden of Disease and Forecasted Trends. J. Epidemiol. Glob. Health 2020, 10, 107–111. [Google Scholar] [CrossRef] [PubMed]
- Hsieh, M.-C.; Tsai, W.-H.; Jheng, Y.-P.; Su, S.-L.; Wang, S.-Y.; Lin, C.-C.; Chen, Y.-H.; Chang, W.-W. The beneficial effects of Lactobacillus reuteri ADR-1 or ADR-3 consumption on type 2 diabetes mellitus: A randomized, double-blinded, placebo-controlled trial. Sci. Rep. 2018, 8, 16791. [Google Scholar] [CrossRef] [PubMed]
- Zhao, L.; Lou, H.; Peng, Y.; Chen, S.; Fan, L.; Li, X. Elevated levels of circulating short-chain fatty acids and bile acids in type 2 diabetes are linked to gut barrier disruption and disordered gut microbiota. Diabetes Res. Clin. Pr. 2020, 169, 108418. [Google Scholar] [CrossRef]
- Beam, A.; Clinger, E.; Hao, L. Effect of Diet and Dietary Components on the Composition of the Gut Microbiota. Nutrients 2021, 13, 2795. [Google Scholar] [CrossRef] [PubMed]
- Zmora, N.; Suez, J.; Elinav, E. You are what you eat: Diet, health and the gut microbiota. Nat. Rev. Gastroenterol. Hepatol. 2019, 16, 35–56. [Google Scholar] [CrossRef] [PubMed]
- Skantze, V.; Hjorth, T.; Wallman, M.; Brunius, C.; Dicksved, J.; Pelve, E.A.; Esberg, A.; Vitale, M.; Giacco, R.; Costabile, G.; et al. Differential Responders to a Mixed Meal Tolerance Test Associated with Type 2 Diabetes Risk Factors and Gut Microbiota—Data from the MEDGI-Carb Randomized Controlled Trial. Nutrients 2023, 15, 4369. [Google Scholar] [CrossRef] [PubMed]
- Maestri, M.; Santopaolo, F.; Pompili, M.; Gasbarrini, A.; Ponziani, F.R. Gut microbiota modulation in patients with non-alcoholic fatty liver disease: Effects of current treatments and future strategies. Front. Nutr. 2023, 10, 1110536. [Google Scholar] [CrossRef] [PubMed]
- Ren, M.; Zhang, H.; Qi, J.; Hu, A.; Jiang, Q.; Hou, Y.; Feng, Q.; Ojo, O.; Wang, X. An Almond-Based Low Carbohydrate Diet Improves Depression and Glycometabolism in Patients with Type 2 Diabetes through Modulating Gut Microbiota and GLP-1: A Randomized Controlled Trial. Nutrients 2020, 12, 3036. [Google Scholar] [CrossRef] [PubMed]
- Kumar, M.; Pal, N.; Sharma, P.; Kumawat, M.; Sarma, D.K.; Nabi, B.; Verma, V.; Tiwari, R.R.; Shubham, S.; Arjmandi, B.; et al. Omega-3 Fatty Acids and Their Interaction with the Gut Microbiome in the Prevention and Amelioration of Type-2 Diabetes. Nutrients 2022, 14, 1723. [Google Scholar] [CrossRef]
- Carranza-Naval, M.J.; Vargas-Soria, M.; Hierro-Bujalance, C.; Baena-Nieto, G.; Garcia-Alloza, M.; Infante-Garcia, C.; del Marco, A. Alzheimer’s Disease and Diabetes: Role of Diet, Microbiota and Inflammation in Preclinical Models. Biomolecules 2021, 11, 262. [Google Scholar] [CrossRef] [PubMed]
- Gao, R.; Meng, X.; Xue, Y.; Mao, M.; Liu, Y.; Tian, X.; Sui, B.; Li, X.; Zhang, P. Bile acids-gut microbiota crosstalk contributes to the improvement of type 2 diabetes mellitus. Front. Pharmacol. 2022, 13, 1027212. [Google Scholar] [CrossRef] [PubMed]
- Rabot, S.; Membrez, M.; Bruneau, A.; Gérard, P.; Harach, T.; Moser, M.; Raymond, F.; Mansourian, R.; Chou, C.J.; Janssen, A.W.F.; et al. Germ-free C57BL/6J mice are resistant to high-fat-diet-induced insulin resistance and have altered cholesterol metabolism. FASEB J. 2010, 24, 4948–4959. [Google Scholar] [CrossRef]
- Hamamah, S.; Covasa, M. Gut Microbiota Restores Central Neuropeptide Deficits in Germ-Free Mice. Int. J. Mol. Sci. 2022, 23, 11756. [Google Scholar] [CrossRef] [PubMed]
- Kimura, I.; Ozawa, K.; Inoue, D.; Imamura, T.; Kimura, K.; Maeda, T.; Terasawa, K.; Kashihara, D.; Hirano, K.; Tani, T.; et al. The gut microbiota suppresses insulin-mediated fat accumulation via the short-chain fatty acid receptor GPR43. Nat. Commun. 2013, 4, 1829. [Google Scholar] [CrossRef] [PubMed]
- Ley, R.E.; Bäckhed, F.; Turnbaugh, P.; Lozupone, C.A.; Knight, R.D.; Gordon, J.I. Obesity alters gut microbial ecology. Proc. Natl. Acad. Sci. USA 2005, 102, 11070–11075. [Google Scholar] [CrossRef] [PubMed]
- Turnbaugh, P.J.; Ley, R.E.; Mahowald, M.A.; Magrini, V.; Mardis, E.R.; Gordon, J.I. An Obesity-Associated Gut Microbiome with Increased Capacity for Energy Harvest. Nature 2006, 444, 1027–1031. [Google Scholar] [CrossRef] [PubMed]
- Gurung, M.; Li, Z.; You, H.; Rodrigues, R.; Jump, D.B.; Morgun, A.; Shulzhenko, N. Role of gut microbiota in type 2 diabetes pathophysiology. EBioMedicine 2020, 51, 102590. [Google Scholar] [CrossRef] [PubMed]
- Sabatino, A.; Regolisti, G.; Cosola, C.; Gesualdo, L.; Fiaccadori, E. Intestinal Microbiota in Type 2 Diabetes and Chronic Kidney Disease. Curr. Diabetes Rep. 2017, 17, 16. [Google Scholar] [CrossRef] [PubMed]
- Larsen, N.; Vogensen, F.K.; Van Den Berg, F.W.J.; Nielsen, D.S.; Andreasen, A.S.; Pedersen, B.K.; Al-Soud, W.A.; Sørensen, S.J.; Hansen, L.H.; Jakobsen, M. Gut Microbiota in Human Adults with Type 2 Diabetes Differs from Non-Diabetic Adults. PLoS ONE 2010, 5, e9085. [Google Scholar] [CrossRef] [PubMed]
- Karlsson, F.; Tremaroli, V.; Nielsen, J.; Bäckhed, F. Assessing the Human Gut Microbiota in Metabolic Diseases. Diabetes 2013, 62, 3341–3349. [Google Scholar] [CrossRef] [PubMed]
- Saad, M.J.A.; Santos, A.; Prada, P.O. Linking Gut Microbiota and Inflammation to Obesity and Insulin Resistance. Physiology 2016, 31, 283–293. [Google Scholar] [CrossRef]
- Lu, Y.-C.; Yeh, W.-C.; Ohashi, P.S. LPS/TLR4 signal transduction pathway. Cytokine 2008, 42, 145–151. [Google Scholar] [CrossRef]
- Shi, H.; Kokoeva, M.V.; Inouye, K.; Tzameli, I.; Yin, H.; Flier, J.S. TLR4 links innate immunity and fatty acid–induced insulin resistance. J. Clin. Investig. 2006, 116, 3015–3025. [Google Scholar] [CrossRef] [PubMed]
- Franchini, M.; Monnais, E.; Seboek, D.; Radimerski, T.; Zini, E.; Kaufmann, K.; Lutz, T.; Reusch, C.; Ackermann, M.; Muller, B.; et al. Insulin Resistance and Increased Lipolysis in Bone Marrow Derived Adipocytes Stimulated with Agonists of Toll-like Receptors. Horm. Metab. Res. 2010, 42, 703–709. [Google Scholar] [CrossRef]
- Samuel, V.T.; Shulman, G.I. Mechanisms for Insulin Resistance: Common Threads and Missing Links. Cell 2012, 148, 852–871. [Google Scholar] [CrossRef] [PubMed]
- Jayaraman, S.; Devarajan, N.; Rajagopal, P.; Babu, S.; Ganesan, S.K.; Veeraraghavan, V.P.; Palanisamy, C.P.; Cui, B.; Periyasamy, V.; Chandrasekar, K. β-Sitosterol Circumvents Obesity Induced Inflammation and Insulin Resistance by down-Regulating IKKβ/NF-κB and JNK Signaling Pathway in Adipocytes of Type 2 Diabetic Rats. Molecules 2021, 26, 2101. [Google Scholar] [CrossRef]
- Zou, P.; Yang, F.; Ding, Y.; Zhang, D.; Liu, Y.; Zhang, J.; Wu, D.; Wang, Y. Lipopolysaccharide downregulates the expression of ZO-1 protein through the Akt pathway. BMC Infect. Dis. 2022, 22, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Lebrun, L.J.; Pallot, G.; Nguyen, M.; Tavernier, A.; Dusuel, A.; Pilot, T.; Deckert, V.; Dugail, I.; Le Guern, N.; De Barros, J.-P.P.; et al. Increased Weight Gain and Insulin Resistance in HF-Fed PLTP Deficient Mice Is Related to Altered Inflammatory Response and Plasma Transport of Gut-Derived LPS. Int. J. Mol. Sci. 2022, 23, 13226. [Google Scholar] [CrossRef] [PubMed]
- Rorato, R.; Borges, B.d.C.; Uchoa, E.T.; Antunes-Rodrigues, J.; Elias, C.F.; Elias, L.L.K. LPS-Induced Low-Grade Inflammation Increases Hypothalamic JNK Expression and Causes Central Insulin Resistance Irrespective of Body Weight Changes. Int. J. Mol. Sci. 2017, 18, 1431. [Google Scholar] [CrossRef] [PubMed]
- Puertollano, E.; Kolida, S.; Yaqoob, P. Biological significance of short-chain fatty acid metabolism by the intestinal microbiome. Curr. Opin. Clin. Nutr. Metab. Care 2014, 17, 139–144. [Google Scholar] [CrossRef] [PubMed]
- Morrison, D.J.; Preston, T. Formation of short chain fatty acids by the gut microbiota and their impact on human metabolism. Gut Microbes 2016, 7, 189–200. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.-H.; Gao, C.-L.; Guo, H.-L.; Zhang, W.-Q.; Huang, W.; Tang, S.-S.; Gan, W.-J.; Xu, Y.; Zhou, H.; Zhu, Q. Sodium butyrate supplementation ameliorates diabetic inflammation in db/db mice. J. Endocrinol. 2018, 238, 231–244. [Google Scholar] [CrossRef] [PubMed]
- Akhtar, M.; Chen, Y.; Ma, Z.; Zhang, X.; Shi, D.; Khan, J.A.; Liu, H. Gut microbiota-derived short chain fatty acids are potential mediators in gut inflammation. Anim. Nutr. 2021, 8, 350–360. [Google Scholar] [CrossRef] [PubMed]
- Jung, T.W.; Park, H.S.; Choi, G.H.; Kim, D.; Lee, T. β-aminoisobutyric acid attenuates LPS-induced inflammation and insulin resistance in adipocytes through AMPK-mediated pathway. J. Biomed. Sci. 2018, 25, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Entezari, M.; Hashemi, D.; Taheriazam, A.; Zabolian, A.; Mohammadi, S.; Fakhri, F.; Hashemi, M.; Hushmandi, K.; Ashrafizadeh, M.; Zarrabi, A.; et al. AMPK signaling in diabetes mellitus, insulin resistance and diabetic complications: A pre-clinical and clinical investigation. Biomed. Pharmacother. 2021, 146, 112563. [Google Scholar] [CrossRef]
- Hu, M.; Alhamwe, B.A.; Santner-Nanan, B.; Miethe, S.; Harb, H.; Renz, H.; Potaczek, D.P.; Nanan, R.K. Short-Chain Fatty Acids Augment Differentiation and Function of Human Induced Regulatory T Cells. Int. J. Mol. Sci. 2022, 23, 5740. [Google Scholar] [CrossRef] [PubMed]
- Haase, S.; Mäurer, J.; Duscha, A.; Lee, D.-H.; Balogh, A.; Gold, R.; Müller, D.N.; Haghikia, A.; Linker, R.A. Propionic Acid Rescues High-Fat Diet Enhanced Immunopathology in Autoimmunity via Effects on Th17 Responses. Front. Immunol. 2021, 12. [Google Scholar] [CrossRef] [PubMed]
- Ilan, Y.; Maron, R.; Tukpah, A.-M.; Maioli, T.U.; Murugaiyan, G.; Yang, K.; Wu, H.Y.; Weiner, H.L. Induction of regulatory T cells decreases adipose inflammation and alleviates insulin resistance in ob/ob mice. Proc. Natl. Acad. Sci. 2010, 107, 9765–9770. [Google Scholar] [CrossRef]
- Rameshrad, M.; Razavi, B.M.; Lalau, J.-D.; E De Broe, M.; Hosseinzadeh, H. An overview of glucagon-like peptide-1 receptor agonists for the treatment of metabolic syndrome: A drug repositionin. Iran J Basic Med Sci . 2020, 23, 568. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Dilidaxi, D.; Wu, Y.; Sailike, J.; Sun, X.; Nabi, X.-H. Composite probiotics alleviate type 2 diabetes by regulating intestinal microbiota and inducing GLP-1 secretion in db/db mice. Biomed. Pharmacother. 2020, 125, 109914. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.; Lou, X.; Jiang, C.; Ji, X.; Tao, X.; Sun, J.; Bao, Z. Gut microbiota is correlated with gastrointestinal adverse events of metformin in patients with type 2 diabetes. Front. Endocrinol. 2022, 13, 1044030. [Google Scholar] [CrossRef] [PubMed]
- Molinero, N.; Ruiz, L.; Sánchez, B.; Margolles, A.; Delgado, S. Intestinal Bacteria Interplay With Bile and Cholesterol Metabolism: Implications on Host Physiology. Front. Physiol. 2019, 10, 185. [Google Scholar] [CrossRef]
- Thomas, C.; Gioiello, A.; Noriega, L.; Strehle, A.; Oury, J.; Rizzo, G.; Macchiarulo, A.; Yamamoto, H.; Mataki, C.; Pruzanski, M.; et al. TGR5-Mediated Bile Acid Sensing Controls Glucose Homeostasis. Cell Metab. 2009, 10, 167–177. [Google Scholar] [CrossRef]
- Cariou, B.; van Harmelen, K.; Duran-Sandoval, D.; van Dijk, T.H.; Grefhorst, A.; Abdelkarim, M.; Caron, S.; Torpier, G.; Fruchart, J.-C.; Gonzalez, F.J.; et al. The Farnesoid X Receptor Modulates Adiposity and Peripheral Insulin Sensitivity in Mice. J. Biol. Chem. 2006, 281, 11039–11049. [Google Scholar] [CrossRef]
- Bjursell, M.; Wedin, M.; Admyre, T.; Hermansson, M.; Böttcher, G.; Göransson, M.; Lindén, D.; Bamberg, K.; Oscarsson, J.; Bohlooly-Y, M. Ageing Fxr Deficient Mice Develop Increased Energy Expenditure, Improved Glucose Control and Liver Damage Resembling NASH. PLoS ONE 2013, 8, e64721. [Google Scholar] [CrossRef]
- Prawitt, J.; Abdelkarim, M.; Stroeve, J.H.M.; Popescu, I.; Duez, H.; Velagapudi, V.R.; Dumont, J.; Bouchaert, E.; van Dijk, T.H.; Lucas, A.; et al. Farnesoid X Receptor Deficiency Improves Glucose Homeostasis in Mouse Models of Obesity. Diabetes 2011, 60, 1861–1871. [Google Scholar] [CrossRef] [PubMed]
- Gong, Q.; Hu, Z.; Zhang, F.; Cui, A.; Chen, X.; Jiang, H.; Gao, J.; Chen, X.; Han, Y.; Liang, Q.; et al. Fibroblast growth factor 21 improves hepatic insulin sensitivity by inhibiting mammalian target of rapamycin complex 1 in mice. J. Hepatol. 2016, 64, 425–438. [Google Scholar] [CrossRef] [PubMed]
- Wu, L.; Mo, W.; Feng, J.; Li, J.; Yu, Q.; Li, S.; Zhang, J.; Chen, K.; Ji, J.; Dai, W.; et al. Astaxanthin attenuates hepatic damage and mitochondrial dysfunction in non-alcoholic fatty liver disease by up-regulating the FGF21/PGC-1α pathway. Br J Pharmacol. 2020, 177, 3760–3777. [Google Scholar] [CrossRef] [PubMed]
- Abaj, F.; Rafiee, M.; Koohdani, F. A personalised diet approach study: Interaction between PPAR-γ Pro12Ala and dietary insulin indices on metabolic markers in diabetic patients. J. Hum. Nutr. Diet. 2022, 35, 663–674. [Google Scholar] [CrossRef] [PubMed]
- Tilg, H.; Moschen, A.R. Food, immunity, and the microbiome. Gastroenterology 2015, 148, 1107–1119. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Zhang, M.; Wang, S.; Han, R.; Cao, Y.; Hua, W.; Mao, Y.; Zhang, X.; Pang, X.; Wei, C.; et al. Interactions between gut microbiota, host genetics and diet relevant to development of metabolic syndromes in mice. ISME J. 2009, 4, 232–241. [Google Scholar] [CrossRef] [PubMed]
- Rothschild, D.; Weissbrod, O.; Barkan, E.; Kurilshikov, A.; Korem, T.; Zeevi, D.; Costea, P.I.; Godneva, A.; Kalka, I.N.; Bar, N.; et al. Environment dominates over host genetics in shaping human gut microbiota. Nature 2018, 555, 210–215. [Google Scholar] [CrossRef] [PubMed]
- Cooper, A.J.; Forouhi, N.G.; Ye, Z.; Buijsse, B.; Arriola, L.; Balkau, B.; Barricarte, A.; Beulens, J.W.; Boeing, H.; Büchner, F.L.; et al. Fruit and vegetable intake and type 2 diabetes: EPIC-InterAct prospective study and meta-analysis. Eur J Clin Nutr. 2012, 66, 1082–1092. [Google Scholar] [CrossRef] [PubMed]
- Villegas, R.; Liu, S.; Gao, Y.-T.; Yang, G.; Li, H.; Zheng, W.; Shu, X.O. Prospective Study of Dietary Carbohydrates, Glycemic Index, Glycemic Load, and Incidence of Type 2 Diabetes Mellitus in Middle-aged Chinese Women. Arch. Intern. Med. 2007, 167, 2310–2316. [Google Scholar] [CrossRef] [PubMed]
- Jenkins, D.J.; Kendall, C.W.; McKeown-Eyssen, G.; Josse, R.G.; Silverberg, J.; Booth, G.L.; Vidgen, E.; Josse, A.R.; Nguyen, T.H.; Corrigan, S.; et al. Effect of a low-glycemic index or a high-cereal fiber diet on type 2 diabetes: A randomized trial. Jama 2008, 300, 2742–2753. [Google Scholar] [CrossRef] [PubMed]
- Ojo, O.; Ojo, O.O.; Adebowale, F.; Wang, X.H. The Effect of Dietary Glycaemic Index on Glycaemia in Patients with Type 2 Diabetes: A Systematic Review and Meta-Analysis of Randomized Controlled Trials. Nutrients 2018, 10, 373. [Google Scholar] [CrossRef]
- Kabir, M.; Oppert, J.-M.; Vidal, H.; Bruzzo, F.; Fiquet, C.; Wursch, P.; Slama, G.; Rizkalla, S.W. Four-week low-glycemic index breakfast with a modest amount of soluble fibers in type 2 diabetic men. Metabolism 2002, 51, 819–826. [Google Scholar] [CrossRef] [PubMed]
- Jimenez-Cruz, A.; Bacardi-Gascon, M.; Turnbull, W.H.; Rosales-Garay, P.; Severino-Lugo, I. A Flexible, Low-Glycemic Index Mexican-Style Diet in Overweight and Obese Subjects With Type 2 Diabetes Improves Metabolic Parameters During a 6-Week Treatment Period. Diabetes Care 2003, 26, 1967–1970. [Google Scholar] [CrossRef]
- Hur, H.J.; Wu, X.; Yang, H.J.; Kim, M.J.; Lee, K.-H.; Hong, M.; Park, S.; Kim, M.-S. Beneficial Effects of a Low-Glycemic Diet on Serum Metabolites and Gut Microbiota in Obese Women With Prevotella and Bacteriodes Enterotypes: A Randomized Clinical Trial. Front. Nutr. 2022, 9, 861880. [Google Scholar] [CrossRef] [PubMed]
- Bhute, S.S.; Mefferd, C.C.; Phan, J.R.; Ahmed, M.; Fox-King, A.E.; Alarcia, S.; Villarama, J.V.; Abel-Santos, E.; Hedlund, B.P. A High-Carbohydrate Diet Prolongs Dysbiosis and Clostridioides difficile Carriage and Increases Delayed Mortality in a Hamster Model of Infection. Microbiol. Spectr. 2022, 10, e0180421. [Google Scholar] [CrossRef] [PubMed]
- Heilbronn, L.K.; Clifton, P.M. C-reactive protein and coronary artery disease: Influence of obesity, caloric restriction and weight loss. J. Nutr. Biochem. 2002, 13, 316–321. [Google Scholar] [CrossRef] [PubMed]
- Wolever, T.M.; Brand-Miller, J.C.; Abernethy, J.; Astrup, A.; Atkinson, F.; Axelsen, M.; Björck, I.; Brighenti, F.; Brown, R.; Brynes, A.; et al. Measuring the glycemic index of foods: Interlaboratory study. Am. J. Clin. Nutr. 2008, 87, 247S–257S. [Google Scholar] [CrossRef]
- Schwedhelm, C.; Boeing, H.; Hoffmann, G.; Aleksandrova, K.; Schwingshackl, L. Effect of diet on mortality and cancer recurrence among cancer survivors: A systematic review and meta-analysis of cohort studies. Nutr. Rev. 2016, 74, 737–748. [Google Scholar] [CrossRef] [PubMed]
- Hur, H.J.; Yang, H.J.; Kim, M.J.; Lee, K.-H.; Kim, M.-S.; Park, S. Association of Polygenic Variants with Type 2 Diabetes Risk and Their Interaction with Lifestyles in Asians. Nutrients 2022, 14, 3222. [Google Scholar] [CrossRef] [PubMed]
- Singh, R.K.; Chang, H.-W.; Yan, D.; Lee, K.M.; Ucmak, D.; Wong, K.; Abrouk, M.; Farahnik, B.; Nakamura, M.; Zhu, T.H.; et al. Influence of diet on the gut microbiome and implications for human health. J. Transl. Med. 2017, 15, 73. [Google Scholar] [CrossRef] [PubMed]
- Bibbò, S.; Ianiro, G.; Giorgio, V.; Scaldaferri, F.; Masucci, L.; Gasbarrini, A.; Cammarota, G. The role of diet on gut microbiota composition. Eur. Rev. Med. Pharmacol. Sci. 2016, 20, 4742–4749. [Google Scholar] [PubMed]
- Zhou, X.; Han, D.; Xu, R.; Li, S.; Wu, H.; Qu, C.; Wang, F.; Wang, X.; Zhao, Y. A Model of Metabolic Syndrome and Related Diseases with Intestinal Endotoxemia in Rats Fed a High Fat and High Sucrose Diet. PLoS ONE 2014, 9, e115148. [Google Scholar] [CrossRef] [PubMed]
- Tomas, J.; Mulet, C.; Saffarian, A.; Cavin, J.-B.; Ducroc, R.; Regnault, B.; Tan, C.K.; Duszka, K.; Burcelin, R.; Wahli, W.; et al. High-fat diet modifies the PPAR-γ pathway leading to disruption of microbial and physiological ecosystem in murine small intestine. Proc. Natl. Acad. Sci. 2016, 113, E5934–E5943. [Google Scholar] [CrossRef]
- David, L.A.; Maurice, C.F.; Carmody, R.N.; Gootenberg, D.B.; Button, J.E.; Wolfe, B.E.; Ling, A.V.; Devlin, A.S.; Varma, Y.; Fischbach, M.A.; et al. Diet rapidly and reproducibly alters the human gut microbiome. Nature 2014, 505, 559–563. [Google Scholar] [CrossRef] [PubMed]
- Xie, Y.; Zhao, Y.; Shi, L.; Li, W.; Chen, K.; Li, M.; Chen, X.; Zhang, H.; Li, T.; Matsuzawa-Ishimoto, Y.; et al. Gut epithelial TSC1/mTOR controls RIPK3-dependent necroptosis in intestinal inflammation and cancer. J Clin Invest. 2020, 130, 2111–2128. [Google Scholar] [CrossRef]
- Kothari, V.; Luo, Y.; Tornabene, T.; O'Neill, A.M.; Greene, M.W.; Geetha, T.; Babu, J.R. High fat diet induces brain insulin resistance and cognitive impairment in mice. Biochim. et Biophys. Acta (BBA) - Mol. Basis Dis. 2017, 1863, 499–508. [Google Scholar] [CrossRef] [PubMed]
- Solinas, G.; Becattini, B. JNK at the crossroad of obesity, insulin resistance, and cell stress response. Mol. Metab. 2016, 6, 174–184. [Google Scholar] [CrossRef]
- Chen, Y.; Wang, X.; Zhang, M.-F.; Li, Y.-X.; Li, Y.; Gu, T.; Xia, F.-Z.; Yu, J.; Lu, Y.-L. Short reaction of C-peptide, glucagon-like peptide-1, ghrelin and endomorphin-1 for different style diet in type 2 diabetic patients. Chin. Med. J. 2011, 124, 3485–3489. [Google Scholar] [CrossRef] [PubMed]
- Brøns, C.; Jensen, C.B.; Storgaard, H.; Hiscock, N.J.; White, A.; Appel, J.S.; Jacobsen, S.; Nilsson, E.; Larsen, C.M.; Astrup, A.; et al. Impact of short-term high-fat feeding on glucose and insulin metabolism in young healthy men. J. Physiol. 2009, 587, 2387–2397. [Google Scholar] [CrossRef]
- Riccio, P.; Rossano, R. Diet, Gut Microbiota, and Vitamins D + A in Multiple Sclerosis. Neurotherapeutics. 2018, 15, 75–91. [Google Scholar] [CrossRef] [PubMed]
- Meslier, V.; Laiola, M.; Roager, H.M.; De Filippis, F.; Roume, H.; Quinquis, B.; Giacco, R.; Mennella, I.; Ferracane, R.; Pons, N.; et al. Mediterranean diet intervention in overweight and obese subjects lowers plasma cholesterol and causes changes in the gut microbiome and metabolome independently of energy intake. Gut 2020, 69, 1258–1268. [Google Scholar] [CrossRef]
- Newman, T.M.; Shively, C.A.; Register, T.C.; Appt, S.E.; Yadav, H.; Colwell, R.R.; Fanelli, B.; Dadlani, M.; Graubics, K.; Nguyen, U.T.; et al. Diet, obesity, and the gut microbiome as determinants modulating metabolic outcomes in a non-human primate model. Microbiome 2021, 9, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Sobiecki, J.G.; Imamura, F.; Davis, C.R.; Sharp, S.J.; Koulman, A.; Hodgson, J.M.; Guevara, M.; Schulze, M.B.; Zheng, J.S.; Agnoli, C.; et al. A nutritional biomarker score of the Mediterranean diet and incident type 2 diabetes: Integrated analysis of data from the MedLey randomised controlled trial and the EPIC-InterAct case-cohort study. PLoS Med. 2023, 20, e1004221. [Google Scholar] [CrossRef]
- Al-Aubaidy, H.A.; Dayan, A.; Deseo, M.A.; Itsiopoulos, C.; Jamil, D.; Hadi, N.R.; Thomas, C.J. Twelve-Week Mediterranean Diet Intervention Increases Citrus Bioflavonoid Levels and Reduces Inflammation in People with Type 2 Diabetes Mellitus. Nutrients 2021, 13, 1133. [Google Scholar] [CrossRef] [PubMed]
- Di Mauro, A.; Tuccinardi, D.; Watanabe, M.; Del Toro, R.; Monte, L.; Giorgino, R.; Rampa, L.; Rossini, G.; Kyanvash, S.; Soare, A.; et al. The Mediterranean diet increases glucagon-like peptide 1 and oxyntomodulin compared with a vegetarian diet in patients with type 2 diabetes: A randomized controlled cross-over trial. Diabetes/Metabolism Res. Rev. 2020, 37, e3406. [Google Scholar] [CrossRef] [PubMed]
- Papamichou, D.; Panagiotakos, D.B.; Holmes, E.; Koutsakis, P.; Katsoulotos, H.; Loo, R.L.; Itsiopoulos, C. The rationale and design of a Mediterranean diet accompanied by time restricted feeding to optimise the management of type 2 diabetes: The MedDietFast randomised controlled trial. Nutr. Metab. Cardiovasc. Dis. 2021, 32, 220–230. [Google Scholar] [CrossRef] [PubMed]
- Pedersen, H.K.; Gudmundsdottir, V.; Nielsen, H.B.; Hyotylainen, T.; Nielsen, T.; Jensen, B.A.H.; Forslund, K.; Hildebrand, F.; Prifti, E.; Falony, G.; et al. Human gut microbes impact host serum metabolome and insulin sensitivity. Nature 2016, 535, 376–381. [Google Scholar] [CrossRef] [PubMed]
- Gudban, N.; Yehuda, I.; Nasir, W.; Soboh, S.; Tamir, S.; Blum, A. Effect of Telemedicine Dietary Intervention on Endothelial Function in Patients with Type 2 Diabetes Mellitus on Mediterranean Diet. 2021, 23, 89–93. Isr Med Assoc J . 2021, 23, 89–93. [Google Scholar] [PubMed]
- van de Stolpe, A.; van der Saag, P.T. Intercellular adhesion molecule-1. J Mol Med 1996, 74, 13–33. [Google Scholar] [CrossRef] [PubMed]
- Zhou, M.; Shao, J.; Wu, C.-Y.; Shu, L.; Dong, W.; Liu, Y.; Chen, M.; Wynn, R.M.; Wang, J.; Wang, J.; et al. Targeting BCAA Catabolism to Treat Obesity-Associated Insulin Resistance. Diabetes 2019, 68, 1730–1746. [Google Scholar] [CrossRef]
- Lynch, C.J.; Adams, S.H. Branched-chain amino acids in metabolic signalling and insulin resistance. Nat. Rev. Endocrinol. 2014, 10, 723–736. [Google Scholar] [CrossRef] [PubMed]
- Chandel, N.S. Carbohydrate Metabolism. Cold Spring Harb Perspect Biol. 2021, 13. [Google Scholar] [CrossRef] [PubMed]
- Zhao, L.; Hutchison, A.T.; Heilbronn, L.K. Carbohydrate intake and circadian synchronicity in the regulation of glucose homeostasis. Curr. Opin. Clin. Nutr. Metab. Care 2021, 24, 342–348. [Google Scholar] [CrossRef] [PubMed]
- Lütjens, A.; Verleur, H.; Plooij, M. Glucose and insulin levels on loading with different carbohydrates. Clin. Chim. Acta 1975, 62, 239–243. [Google Scholar] [CrossRef] [PubMed]
- Koloverou, E.; Panagiotakos, D.B. Macronutrient Composition and Management of Non-Insulin-Dependent Diabetes Mellitus (NIDDM): A New Paradigm for Individualized Nutritional Therapy in Diabetes Patients. Rev. Diabet. Stud. 2016, 13, 6–16. [Google Scholar] [CrossRef] [PubMed]
- Reynolds, A.; Mann, J.; Cummings, J.; Winter, N.; Mete, E.; Te Morenga, L. Carbohydrate quality and human health: A series of systematic reviews and meta-analyses. Lancet 2019, 393, 434–445. [Google Scholar] [CrossRef] [PubMed]
- Wheeler, M.L.; Dunbar, S.A.; Jaacks, L.M.; Karmally, W.; Mayer-Davis, E.J.; Wylie-Rosett, J.; Yancy, W.S. Response to Comment on: Wheeler et al. Macronutrients, Food Groups, and Eating Patterns in the Management of Diabetes: A Systematic Review of the Literature, 2010. Diabetes Care 2012, 35, 434–445. [Google Scholar] [CrossRef] [PubMed]
- Classification and Diagnosis of Diabetes: Standards of Medical Care in Diabetes-2021. Diabetes Care 2021, 44 (Suppl S1), S15–S33. [CrossRef]
- Boden, G.; Sargrad, K.; Homko, C.; Mozzoli, M.; Stein, T.P. Effect of a Low-Carbohydrate Diet on Appetite, Blood Glucose Levels, and Insulin Resistance in Obese Patients with Type 2 Diabetes. Ann. Intern. Med. 2005, 142, 403–411. [Google Scholar] [CrossRef]
- Davis, N.J.; Tomuta, N.; Schechter, C.; Isasi, C.R.; Segal-Isaacson, C.; Stein, D.; Zonszein, J.; Wylie-Rosett, J. Comparative Study of the Effects of a 1-Year Dietary Intervention of a Low-Carbohydrate Diet Versus a Low-Fat Diet on Weight and Glycemic Control in Type 2 Diabetes. Diabetes Care 2009, 32, 1147–1152. [Google Scholar] [CrossRef]
- Yancy, W.S.; Foy, M.; Chalecki, A.M.; Vernon, M.C.; Westman, E.C. A low-carbohydrate, ketogenic diet to treat type 2 diabetes. Nutr. Metab. 2005, 2, 34. [Google Scholar] [CrossRef]
- Kokubo, E.; Morita, S.; Nagashima, H.; Oshio, K.; Iwamoto, H.; Miyaji, K. Blood Glucose Response of a Low-Carbohydrate Oral Nutritional Supplement with Isomaltulose and Soluble Dietary Fiber in Individuals with Prediabetes: A Randomized, Single-Blind Crossover Trial. Nutrients 2022, 14, 2386. [Google Scholar] [CrossRef] [PubMed]
- Gow, M.L.; Garnett, S.P.; Baur, L.A.; Lister, N.B. The Effectiveness of Different Diet Strategies to Reduce Type 2 Diabetes Risk in Youth. Nutrients 2016, 8, 486. [Google Scholar] [CrossRef]
- Alhazmi, A.; Stojanovski, E.; McEvoy, M.; Garg, M.L. Macronutrient Intakes and Development of Type 2 Diabetes: A Systematic Review and Meta-Analysis of Cohort Studies. J. Am. Coll. Nutr. 2012, 31, 243–258. [Google Scholar] [CrossRef] [PubMed]
- Seino, Y.; Maekawa, R.; Ogata, H.; Hayashi, Y. Carbohydrate-induced secretion of glucose-dependent insulinotropic polypeptide and glucagon-like peptide-1. J Diabetes Investig. 2016, 1 (Suppl 1), 27–32. [Google Scholar] [CrossRef]
- Zhang, J.-M.; Sun, Y.-S.; Zhao, L.-Q.; Chen, T.-T.; Fan, M.-N.; Jiao, H.-C.; Zhao, J.-P.; Wang, X.-J.; Li, F.-C.; Li, H.-F.; et al. SCFAs-Induced GLP-1 Secretion Links the Regulation of Gut Microbiome on Hepatic Lipogenesis in Chickens. Front. Microbiol. 2019, 10, 2176. [Google Scholar] [CrossRef]
- Hernández, M.A.G.; Canfora, E.E.; Jocken, J.W.E.; Blaak, E.E. The Short-Chain Fatty Acid Acetate in Body Weight Control and Insulin Sensitivity. Nutrients 2019, 11, 1943. [Google Scholar] [CrossRef] [PubMed]
- Takeuchi, T.; Kubota, T.; Nakanishi, Y.; Tsugawa, H.; Suda, W.; Kwon, A.T.-J.; Yazaki, J.; Ikeda, K.; Nemoto, S.; Mochizuki, Y.; et al. Gut microbial carbohydrate metabolism contributes to insulin resistance. Nature 2023, 621, 389–395. [Google Scholar] [CrossRef]
- Do, M.H.; Lee, E.; Oh, M.-J.; Kim, Y.; Park, H.-Y. High-Glucose or -Fructose Diet Cause Changes of the Gut Microbiota and Metabolic Disorders in Mice without Body Weight Change. Nutrients 2018, 10, 761. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Liang, X.-F.; He, S.; Chen, X.; Wang, J.; Li, J.; Zhu, Q.; Zhang, Z.; Li, L.; Alam, M.S. Effects of High Carbohydrate Diet-Modulated Microbiota on Gut Health in Chinese Perch. Front. Microbiol. 2020, 11. [Google Scholar] [CrossRef]
- Derrien, M.; Belzer, C.; de Vos, W.M. Akkermansia muciniphila and its role in regulating host functions. Microb. Pathog. 2017, 106, 171–181. [Google Scholar] [CrossRef]
- Chiantera, V.; Laganà, A.S.; Basciani, S.; Nordio, M.; Bizzarri, M. A Critical Perspective on the Supplementation of Akkermansia muciniphila: Benefits and Harms. Life 2023, 13, 1247. [Google Scholar] [CrossRef] [PubMed]
- Singh, S.B.; Carroll-Portillo, A.; Lin, H.C. Desulfovibrio in the Gut: The Enemy within? Microorganisms 2023, 11, 1772. [Google Scholar] [CrossRef] [PubMed]
- Gu, X.-M.; Lu, C.-Y.; Pan, J.; Ye, J.-Z.; Zhu, Q.-H. Alteration of intestinal microbiota is associated with diabetic retinopathy and its severity: Samples collected from southeast coast Chinese. World J. Diabetes 2023, 14, 862–882. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Wang, J.; Yi, J.; Liu, Y.; Yu, Z.; Chen, S.; Liu, X. Increased mucin-degrading bacteria by high protein diet leads to thinner mucus layer and aggravates experimental colitis. J. Gastroenterol. Hepatol. 2021, 36, 2864–2874. [Google Scholar] [CrossRef]
- DeVries, J.W. On defining dietary fibre. Proc. Nutr. Soc. 2003, 62, 37–43. [Google Scholar] [CrossRef] [PubMed]
- Makki, K.; Deehan, E.C.; Walter, J.; Bäckhed, F. The Impact of Dietary Fiber on Gut Microbiota in Host Health and Disease. Cell Host Microbe 2018, 23, 705–715. [Google Scholar] [CrossRef] [PubMed]
- Gill, S.K.; Rossi, M.; Bajka, B.; Whelan, K. Dietary fibre in gastrointestinal health and disease. Nat. Rev. Gastroenterol. Hepatol. 2021, 18, 101–116. [Google Scholar] [CrossRef] [PubMed]
- Anderson, J.W.; Randles, K.M.; Kendall, C.W.C.; Jenkins, D.J.A. Carbohydrate and Fiber Recommendations for Individuals with Diabetes: A Quantitative Assessment and Meta-Analysis of the Evidence. J. Am. Coll. Nutr. 2004, 23, 5–17. [Google Scholar] [CrossRef] [PubMed]
- Veronese, N.; Solmi, M.; Caruso, M.G.; Giannelli, G.; Osella, A.R.; Evangelou, E.; Maggi, S.; Fontana, L.; Stubbs, B.; Tzoulaki, I. Dietary fiber and health outcomes: An umbrella review of systematic reviews and meta-analyses. Am. J. Clin. Nutr. 2018, 107, 436–444. [Google Scholar] [CrossRef] [PubMed]
- Steemburgo, T.; Dall'Alba, V.; Almeida, J.C.; Zelmanovitz, T.; Gross, J.L.; de Azevedo, M.J. Intake of soluble fibers has a protective role for the presence of metabolic syndrome in patients with type 2 diabetes. Eur. J. Clin. Nutr. 2007, 63, 127–133. [Google Scholar] [CrossRef]
- Silva, F.M.; Steemburgo, T.; de Mello, V.D.; Tonding, S.F.; Gross, J.L.; Azevedo, M.J. High dietary glycemic index and low fiber content are associated with metabolic syndrome in patients with type 2 diabetes. J. Am. Coll. Nutr. 2011, 30, 141–148. [Google Scholar] [CrossRef]
- Schulze, M.B.; Schulz, M.; Heidemann, C.; Schienkiewitz, A.; Hoffmann, K.; Boeing, H. Fiber and magnesium intake and incidence of type 2 diabetes: A prospective study and meta-analysis. Arch Intern Med. 2007, 167, 956–965. [Google Scholar] [CrossRef] [PubMed]
- Qi, L.; van Dam, R.M.; Liu, S.; Franz, M.; Mantzoros, C.; Hu, F.B. Whole-Grain, Bran, and Cereal Fiber Intakes and Markers of Systemic Inflammation in Diabetic Women. Diabetes Care 2006, 29, 207–211. [Google Scholar] [CrossRef] [PubMed]
- Qi, L.; Meigs, J.B.; Liu, S.; Manson, J.E.; Mantzoros, C.; Hu, F.B. Dietary Fibers and Glycemic Load, Obesity, and Plasma Adiponectin Levels in Women With Type 2 Diabetes. Diabetes Care 2006, 29, 1501–1505. [Google Scholar] [CrossRef] [PubMed]
- AlEssa, H.B.; Bhupathiraju, S.N.; Malik, V.S.; Wedick, N.M.; Campos, H.; Rosner, B.; Willett, W.C.; Hu, F.B. Carbohydrate quality and quantity and risk of type 2 diabetes in US women. Am. J. Clin. Nutr. 2015, 102, 1543–1553. [Google Scholar] [CrossRef] [PubMed]
- Schulze, M.B.; Liu, S.; Rimm, E.B.; E Manson, J.; Willett, W.C.; Hu, F.B. Glycemic index, glycemic load, and dietary fiber intake and incidence of type 2 diabetes in younger and middle-aged women. Am. J. Clin. Nutr. 2004, 80, 348–356. [Google Scholar] [CrossRef]
- McRorie, J.W., Jr. Evidence-Based Approach to Fiber Supplements and Clinically Meaningful Health Benefits, Part 1: What to Look for and How to Recommend an Effective Fiber Therapy. Nutr. Today 2015, 50, 82–89. [Google Scholar] [CrossRef] [PubMed]
- McRorie, J.W., Jr.; McKeown, N.M. Understanding the Physics of Functional Fibers in the Gastrointestinal Tract: An Evidence-Based Approach to Resolving Enduring Misconceptions about Insoluble and Soluble Fiber. J. Acad. Nutr. Diet. 2017, 117, 251–264. [Google Scholar] [CrossRef]
- Ziai, S.A.; Larijani, B.; Akhoondzadeh, S.; Fakhrzadeh, H.; Dastpak, A.; Bandarian, F.; Rezai, A.; Badi, H.N.; Emami, T. Psyllium decreased serum glucose and glycosylated hemoglobin significantly in diabetic outpatients. J. Ethnopharmacol. 2005, 102, 202–207. [Google Scholar] [CrossRef]
- Guan, Z.-W.; Yu, E.-Z.; Feng, Q. Soluble Dietary Fiber, One of the Most Important Nutrients for the Gut Microbiota. Molecules 2021, 26, 6802. [Google Scholar] [CrossRef] [PubMed]
- Holscher, H.D. Dietary fiber and prebiotics and the gastrointestinal microbiota. Gut Microbes 2017, 8, 172–184. [Google Scholar] [CrossRef] [PubMed]
- Danneskiold-Samsøe, N.B.; Barros, H.D.d.F.Q.; Santos, R.; Bicas, J.L.; Cazarin, C.B.B.; Madsen, L.; Kristiansen, K.; Pastore, G.M.; Brix, S.; Júnior, M.R.M. Interplay between food and gut microbiota in health and disease. Food Res. Int. 2018, 115, 23–31. [Google Scholar] [CrossRef] [PubMed]
- Zou, J.; Chassaing, B.; Singh, V.; Pellizzon, M.; Ricci, M.; Fythe, M.D.; Kumar, M.V.; Gewirtz, A.T. Fiber-Mediated Nourishment of Gut Microbiota Protects against Diet-Induced Obesity by Restoring IL-22-Mediated Colonic Health. Cell Host Microbe 2017, 23, 41–53. [Google Scholar] [CrossRef] [PubMed]
- Singh, S.P.; Jadaun, J.S.; Narnoliya, L.K.; Pandey, A. Prebiotic Oligosaccharides: Special Focus on Fructooligosaccharides, Its Biosynthesis and Bioactivity. Appl. Biochem. Biotechnol. 2017, 183, 613–635. [Google Scholar] [CrossRef] [PubMed]
- Pandey, K.R.; Naik, S.R.; Vakil, B.V. Probiotics, prebiotics and synbiotics- a review. J Food Sci Technol. 2015, 52, 7577–7587. [Google Scholar] [CrossRef]
- Leshem, A.; Segal, E.; Elinav, E. The Gut Microbiome and Individual-Specific Responses to Diet. mSystems 2020, 5. [Google Scholar] [CrossRef] [PubMed]
- Halmos, E.P.; Christophersen, C.T.; Bird, A.R.; Shepherd, S.J.; Gibson, P.R.; Muir, J.G. Diets that differ in their FODMAP content alter the colonic luminal microenvironment. Gut 2015, 64, 93–100. [Google Scholar] [CrossRef] [PubMed]
- Leshem, A.; Liwinski, T.; Elinav, E. Immune-Microbiota Interplay and Colonization Resistance in Infection. Mol. Cell 2020, 78, 597–613. [Google Scholar] [CrossRef]
- Liu, D.; Wen, B.; Zhu, K.; Luo, Y.; Li, J.; Li, Y.; Lin, H.; Huang, J.; Liu, Z. Antibiotics-induced perturbations in gut microbial diversity influence metabolic phenotypes in a murine model of high-fat diet-induced obesity. Appl. Microbiol. Biotechnol. 2019, 103, 5269–5283. [Google Scholar] [CrossRef] [PubMed]
- Zhao, L.; Zhang, F.; Ding, X.; Wu, G.; Lam, Y.Y.; Wang, X.; Fu, H.; Xue, X.; Lu, C.; Ma, J.; et al. Gut bacteria selectively promoted by dietary fibers alleviate type 2 diabetes. Science 2018, 359, 1151–1156. [Google Scholar] [CrossRef] [PubMed]
- De Filippis, F.; Pellegrini, N.; Vannini, L.; Jeffery, I.B.; La Storia, A.; Laghi, L.; Serrazanetti, D.I.; Di Cagno, R.; Ferrocino, I.; Lazzi, C.; et al. High-level adherence to a Mediterranean diet beneficially impacts the gut microbiota and associated metabolome. Gut Microbiota 2016, 65, 1812–1821. [Google Scholar] [CrossRef] [PubMed]
- Röytiö, H.; Mokkala, K.; Vahlberg, T.; Laitinen, K. Dietary intake of fat and fibre according to reference values relates to higher gut microbiota richness in overweight pregnant women. Br. J. Nutr. 2017, 118, 343–352. [Google Scholar] [CrossRef] [PubMed]
- Eid, N.; Enani, S.; Walton, G.; Corona, G.; Costabile, A.; Gibson, G.; Rowland, I.; Spencer, J.P. The impact of date palm fruits and their component polyphenols, on gut microbial ecology, bacterial metabolites and colon cancer cell proliferation. J. Nutr. Sci. 2014, 3, e46. [Google Scholar] [CrossRef] [PubMed]
- Sajilata, M.G.; Singhal, R.S.; Kulkarni, P.R. Resistant Starch-A Review. Compr Rev Food Sci Food Saf. 2006, 5, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Mora-Flores, L.P.; Casildo, R.M.-T.; Fuentes-Cabrera, J.; Pérez-Vicente, H.A.; de Anda-Jáuregui, G.; Neri-Torres, E.E. The Role of Carbohydrate Intake on the Gut Microbiome: A Weight of Evidence Systematic Review. Microorganisms 2023, 11, 1728. [Google Scholar] [CrossRef] [PubMed]
- Brumer, H. Sticking to starch. J Biol Chem. 2022, 298, 102049. [Google Scholar] [CrossRef] [PubMed]
- Birt, D.F.; Boylston, T.; Hendrich, S.; Jane, J.-L.; Hollis, J.; Li, L.; McClelland, J.; Moore, S.; Phillips, G.J.; Rowling, M.; et al. Resistant Starch: Promise for Improving Human Health. Adv. Nutr. Int. Rev. J. 2013, 4, 587–601. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Y.; Zhang, N.; Zhou, Y.; Zhou, Z.; Bai, Y.; Strappe, P.; Blanchard, C. Manipulations of glucose/lipid metabolism and gut microbiota of resistant starch encapsulated Ganoderma lucidum spores in T2DM rats. Food Sci. Biotechnol. 2021, 30, 755–764. [Google Scholar] [CrossRef]
- Zhang, C.; Ma, S.; Wu, J.; Luo, L.; Qiao, S.; Li, R.; Xu, W.; Wang, N.; Zhao, B.; Wang, X.; et al. A specific gut microbiota and metabolomic profiles shifts related to antidiabetic action: The similar and complementary antidiabetic properties of type 3 resistant starch from Canna edulis and metformin. Pharmacol. Res. 2020, 159, 104985. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Chen, J.; Song, Y.-H.; Zhao, R.; Xia, L.; Chen, Y.; Cui, Y.-P.; Rao, Z.-Y.; Zhou, Y.; Zhuang, W.; et al. Effects of the resistant starch on glucose, insulin, insulin resistance, and lipid parameters in overweight or obese adults: A systematic review and meta-analysis. Nutr. Diabetes 2019, 9, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Robertson, M.D. Dietary-resistant starch and glucose metabolism. Curr. Opin. Clin. Nutr. Metab. Care 2012, 15, 362–367. [Google Scholar] [CrossRef] [PubMed]
- Pugh, J.E.; Cai, M.; Altieri, N.; Frost, G. A comparison of the effects of resistant starch types on glycemic response in individuals with type 2 diabetes or prediabetes: A systematic review and meta-analysis. Front. Nutr. 2023, 10, 1118229. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Dong, L.; Wu, J.; Qiao, S.; Xu, W.; Ma, S.; Zhao, B.; Wang, X. Intervention of resistant starch 3 on type 2 diabetes mellitus and its mechanism based on urine metabonomics by liquid chromatography-tandem mass spectrometry. Biomed. Pharmacother. 2020, 128, 110350. [Google Scholar] [CrossRef] [PubMed]
- Stewart, M.L.; Zimmer, J.P. Postprandial glucose and insulin response to a high-fiber muffin top containing resistant starch type 4 in healthy adults: A double-blind, randomized, controlled trial. Nutrition 2018, 53, 59–63. [Google Scholar] [CrossRef]
- Collings, P.; Williams, C.; MacDonald, I. Effects of cooking on serum glucose and insulin responses to starch. BMJ 1981, 282, 1032. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Zhang, M.; Ma, Q.; Tian, B.; Nie, C.; Chen, Z.; Li, J. Health beneficial effects of resistant starch on diabetes and obesity via regulation of gut microbiota: A review. Food Funct. 2020, 11, 5749–5767. [Google Scholar] [CrossRef] [PubMed]
- Martínez, I.; Kim, J.; Duffy, P.R.; Schlegel, V.L.; Walter, J. Resistant Starches Types 2 and 4 Have Differential Effects on the Composition of the Fecal Microbiota in Human Subjects. PLoS ONE 2010, 5, e15046. [Google Scholar] [CrossRef] [PubMed]
- Walker, A.W.; Ince, J.; Duncan, S.H.; Webster, L.M.; Holtrop, G.; Ze, X.; Brown, D.; Stares, M.D.; Scott, P.; Bergerat, A.; et al. Dominant and diet-responsive groups of bacteria within the human colonic microbiota. ISME J. 2010, 5, 220–230. [Google Scholar] [CrossRef] [PubMed]
- Mukhopadhya, I.; Moraïs, S.; Laverde-Gomez, J.; Sheridan, P.O.; Walker, A.W.; Kelly, W.; Klieve, A.V.; Ouwerkerk, D.; Duncan, S.H.; Louis, P.; et al. Sporulation capability and amylosome conservation among diverse human colonic and rumen isolates of the keystone starch-degrader Ruminococcus bromii. Environ. Microbiol. 2017, 20, 324–336. [Google Scholar] [CrossRef] [PubMed]
- Ze, X.; Duncan, S.H.; Louis, P.; Flint, H.J. Ruminococcus bromii is a keystone species for the degradation of resistant starch in the human colon. ISME J. 2012, 6, 1535–1543. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Ouyang, Y.; Li, H.; Shen, L.; Ni, Y.; Fang, Q.; Wu, G.; Qian, L.; Xiao, Y.; Zhang, J.; et al. Metabolic phenotypes and the gut microbiota in response to dietary resistant starch type 2 in normal-weight subjects: A randomized crossover trial. Sci. Rep. 2019, 9, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Beasley, J.M.; Wylie-Rosett, J. The Role of Dietary Proteins Among Persons with Diabetes. Curr. Atheroscler. Rep. 2013, 15, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Beaudry, K.M.; Devries, M.C. Nutritional Strategies to Combat Type 2 Diabetes in Aging Adults: The Importance of Protein. Front. Nutr. 2019, 6, 138. [Google Scholar] [CrossRef] [PubMed]
- Devries, M.C.; Sithamparapillai, A.; Brimble, K.S.; Banfield, L.; Morton, R.W.; Phillips, S.M. 'Changes in Kidney Function Do Not Differ between Healthy Adults Consuming Higher- Compared with Lower- or Normal-Protein Diets: A Systematic Review and Meta-Analysis. J. Nutr. 2018, 148, 1760–1775. [Google Scholar] [CrossRef] [PubMed]
- Piatti, P.; Monti, L.; Magni, F.; Fermo, I.; Baruffaldi, L.; Nasser, R.; Santambrogio, G.; Librenti, M.; Galli-Kienle, M.; Pontiroli, A.; et al. Hypocaloric high-protein diet improves glucose oxidation and spares lean body mass: Comparison to hypocaloric high-carbohydrate diet. Metabolism 1994, 43, 1481–1487. [Google Scholar] [CrossRef] [PubMed]
- Cho, Y.M.; Fujita, Y.; Kieffer, T.J. Glucagon-Like Peptide-1: Glucose Homeostasis and Beyond. Annu. Rev. Physiol. 2014, 76, 535–559. [Google Scholar] [CrossRef] [PubMed]
- Ma, J.; Stevens, J.E.; Cukier, K.; Maddox, A.F.; Wishart, J.M.; Jones, K.L.; Clifton, P.M.; Horowitz, M.; Rayner, C.K. Effects of a Protein Preload on Gastric Emptying, Glycemia, and Gut Hormones After a Carbohydrate Meal in Diet-Controlled Type 2 Diabetes. Diabetes Care 2009, 32, 1600–1602. [Google Scholar] [CrossRef] [PubMed]
- Song, Y.; Manson, J.E.; Buring, J.E.; Liu, S. A prospective study of red meat consumption and type 2 diabetes in middle-aged and elderly women: The women's health study. Diabetes Care. 2004, 27, 2108–2115. [Google Scholar] [CrossRef] [PubMed]
- Sluijs, I.; Beulens, J.W.; van der A, D.L.; Spijkerman, A.M.; Grobbee, D.E.; van der Schouw, Y.T. Dietary Intake of Total, Animal, and Vegetable Protein and Risk of Type 2 Diabetes in the European Prospective Investigation into Cancer and Nutrition (EPIC)-NL Study. Diabetes Care 2009, 33, 43–48. [Google Scholar] [CrossRef] [PubMed]
- Shang, X.; Scott, D.; Hodge, A.M.; English, D.R.; Giles, G.G.; Ebeling, P.R.; Sanders, K.M. Dietary protein intake and risk of type 2 diabetes: Results from the Melbourne Collaborative Cohort Study and a meta-analysis of prospective studies. Am. J. Clin. Nutr. 2016, 104, 1352–1365. [Google Scholar] [CrossRef] [PubMed]
- van Nielen, M.; Feskens, E.J.; Mensink, M.; Sluijs, I.; Molina, E.; Amiano, P.; Ardanaz, E.; Balkau, B.; Beulens, J.W.; Boeing, H.; et al. Dietary protein intake and incidence of type 2 diabetes in Europe: The EPIC-InterAct Case-Cohort Study. Diabetes Care. 2014, 37, 1854–1862. [Google Scholar] [CrossRef] [PubMed]
- Adeva-Andany, M.M.; Fernández-Fernández, C.; Carneiro-Freire, N.; Vila-Altesor, M.; Ameneiros-Rodríguez, E. The differential effect of animal versus vegetable dietary protein on the clinical manifestations of diabetic kidney disease in humans. Clin. Nutr. ESPEN 2022, 48, 21–35. [Google Scholar] [CrossRef] [PubMed]
- Viguiliouk, E.; Stewart, S.E.; Jayalath, V.H.; Ng, A.P.; Mirrahimi, A.; De Souza, R.J.; Hanley, A.J.; Bazinet, R.P.; Mejia, S.B.; Leiter, L.A.; et al. Effect of Replacing Animal Protein with Plant Protein on Glycemic Control in Diabetes: A Systematic Review and Meta-Analysis of Randomized Controlled Trials. Nutrients 2015, 7, 9804–9824. [Google Scholar] [CrossRef] [PubMed]
- Gutierrez-Mariscal, F.M.; Alcalá-Diaz, J.F.; Quintana-Navarro, G.M.; de la Cruz-Ares, S.; Torres-Peña, J.D.; Cardelo, M.P.; Arenas-Larriva, A.P.; Malagón, M.M.; Romero-Cabrera, J.L.; Ordovás, J.M.; et al. Changes in quantity plant-based protein intake on type 2 diabetes remission in coronary heart disease patients: From the CORDIOPREV study. Eur. J. Nutr. 2023, 62, 1903–1913. [Google Scholar] [CrossRef] [PubMed]
- de la Cruz-Ares, S.; Gutiérrez-Mariscal, F.M.; Alcalá-Díaz, J.F.; Quintana-Navarro, G.M.; Podadera-Herreros, A.; Cardelo, M.P.; Torres-Peña, J.D.; Larriva, A.P.A.-D.; Pérez-Martínez, P.; Delgado-Lista, J.; et al. Quality and Quantity of Protein Intake Influence Incidence of Type 2 Diabetes Mellitus in Coronary Heart Disease Patients: From the CORDIOPREV Study. Nutrients 2021, 13, 1217. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.; Zhang, X.; Liu, H.; Brown, M.A.; Qiao, S. Dietary Protein and Gut Microbiota Composition and Function. Curr. Protein Pept. Sci. 2018, 20, 145–154. [Google Scholar] [CrossRef] [PubMed]
- Jäger, R.; Zaragoza, J.; Purpura, M.; Iametti, S.; Marengo, M.; Tinsley, G.M.; Anzalone, A.J.; Oliver, J.M.; Fiore, W.; Biffi, A.; et al. Probiotic Administration Increases Amino Acid Absorption from Plant Protein: A Placebo-Controlled, Randomized, Double-Blind, Multicenter, Crossover Study. Probiotics Antimicrob. Proteins 2020, 12, 1330–1339. [Google Scholar] [CrossRef] [PubMed]
- Mu, C.; Yang, Y.; Luo, Z.; Guan, L.; Zhu, W. The Colonic Microbiome and Epithelial Transcriptome Are Altered in Rats Fed a High-Protein Diet Compared with a Normal-Protein Diet. J. Nutr. 2016, 146, 474–483. [Google Scholar] [CrossRef] [PubMed]
- Mu, C.; Yang, Y.; Luo, Z.; Zhu, W. Temporal microbiota changes of high-protein diet intake in a rat model. Anaerobe 2017, 47, 218–225. [Google Scholar] [CrossRef]
- Beaumont, M.; Portune, K.J.; Steuer, N.; Lan, A.; Cerrudo, V.; Audebert, M.; Dumont, F.; Mancano, G.; Khodorova, N.; Andriamihaja, M.; et al. Quantity and source of dietary protein influence metabolite production by gut microbiota and rectal mucosa gene expression: A randomized, parallel, double-blind trial in overweight humans. Am. J. Clin. Nutr. 2017, 106, 1005–1019. [Google Scholar] [CrossRef] [PubMed]
- Moreno-Pérez, D.; Bressa, C.; Bailén, M.; Hamed-Bousdar, S.; Naclerio, F.; Carmona, M.; Pérez, M.; González-Soltero, R.; Montalvo-Lominchar, M.G.; Carabaña, C.; et al. Effect of a Protein Supplement on the Gut Microbiota of Endurance Athletes: A Randomized, Controlled, Double-Blind Pilot Study. Nutrients 2018, 10, 337. [Google Scholar] [CrossRef] [PubMed]
- Holmes, A.J.; Chew, Y.V.; Colakoglu, F.; Cliff, J.B.; Klaassens, E.; Read, M.N.; Solon-Biet, S.M.; McMahon, A.C.; Cogger, V.C.; Ruohonen, K.; et al. Diet-Microbiome Interactions in Health Are Controlled by Intestinal Nitrogen Source Constraints. Cell Metab. 2016, 25, 140–151. [Google Scholar] [CrossRef] [PubMed]
- Reese, A.T.; Pereira, F.C.; Schintlmeister, A.; Berry, D.; Wagner, M.; Hale, L.P.; Wu, A.; Jiang, S.; Durand, H.K.; Zhou, X.; et al. Microbial nitrogen limitation in the mammalian large intestine. Nat. Microbiol. 2018, 3, 1441–1450. [Google Scholar] [CrossRef] [PubMed]
- Agarwal, U.; Hu, Q.; Baldwin, R.L.; Bequette, B.J. Role of rumen butyrate in regulation of nitrogen utilization and urea nitrogen kinetics in growing sheep1. J. Anim. Sci. 2015, 93, 2382–2390. [Google Scholar] [CrossRef] [PubMed]
- Russell, W.R.; Gratz, S.W.; Duncan, S.H.; Holtrop, G.; Ince, J.; Scobbie, L.; Duncan, G.; Johnstone, A.M.; Lobley, G.E.; Wallace, R.J.; et al. High-protein, reduced-carbohydrate weight-loss diets promote metabolite profiles likely to be detrimental to colonic health. Am. J. Clin. Nutr. 2011, 93, 1062–1072. [Google Scholar] [CrossRef] [PubMed]
- Jia, J.; Dou, P.; Gao, M.; Kong, X.; Li, C.; Liu, Z.; Huang, T. Assessment of Causal Direction Between Gut Microbiota–Dependent Metabolites and Cardiometabolic Health: A Bidirectional Mendelian Randomization Analysis. Diabetes 2019, 68, 1747–1755. [Google Scholar] [CrossRef] [PubMed]
- Hentges, D.J.; Maier, B.R.; Burton, G.C.; A Flynn, M.; Tsutakawa, R.K. Effect of a high-beef diet on the fecal bacterial flora of humans. Cancer Res. 1977, 37, 568–571. [Google Scholar] [PubMed]
- Wu, S.; Bhat, Z.F.; Gounder, R.S.; Ahmed, I.A.M.; Al-Juhaimi, F.Y.; Ding, Y.; Bekhit, A.E.-. .-D.A. Effect of Dietary Protein and Processing on Gut Microbiota—A Systematic Review. Nutrients 2022, 14, 453. [Google Scholar] [CrossRef] [PubMed]
- Han, W.; Zhuang, X.; Liu, Q.; Sun, B.; Miao, H.; Zhang, X. Fermented soy whey induced changes on intestinal microbiota and metabolic influence in mice. Food Sci. Hum. Wellness 2021, 11, 41–48. [Google Scholar] [CrossRef]
- von Frankenberg, A.D.; Silva, F.M.; de Almeida, J.C.; Piccoli, V.; Nascimento, F.V.D.; Sost, M.M.; Leitão, C.B.; Remonti, L.L.R.; Umpierre, D.; Reis, A.F.; et al. Effect of dietary lipids on circulating adiponectin: A systematic review with meta-analysis of randomised controlled trials. Br. J. Nutr. 2014, 112, 1235–1250. [Google Scholar] [CrossRef] [PubMed]
- Gao, H.; Geng, T.; Huang, T.; Zhao, Q. Fish oil supplementation and insulin sensitivity: A systematic review and meta-analysis. Lipids Heal. Dis. 2017, 16, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Neuenschwander, M.; Barbaresko, J.; Pischke, C.R.; Iser, N.; Beckhaus, J.; Schwingshackl, L.; Schlesinger, S. Intake of dietary fats and fatty acids and the incidence of type 2 diabetes: A systematic review and dose-response meta-analysis of prospective observational studies. PLOS Med. 2020, 17, e1003347. [Google Scholar] [CrossRef] [PubMed]
- Steyn, N.; Mann, J.; Bennett, P.; Temple, N.; Zimmet, P.; Tuomilehto, J.; Lindström, J.; Louheranta, A. Diet, nutrition and the prevention of type 2 diabetes. PhotoniX 2004, 7, 147–165. [Google Scholar] [CrossRef] [PubMed]
- Tinker LF, Bonds DE, Margolis KL, Manson JE, Howard BV, Larson J, Perri MG, Beresford SA, Robinson JG, Rodríguez B; et al. Low-fat dietary pattern and risk of treated diabetes mellitus in postmenopausal women: The Women's Health Initiative randomized controlled dietary modification trial. Arch Intern Med. 2008, 168, 1500–1511. [CrossRef]
- Wang, L.-L.; Wang, Q.; Hong, Y.; Ojo, O.; Jiang, Q.; Hou, Y.-Y.; Huang, Y.-H.; Wang, X.-H. The Effect of Low-Carbohydrate Diet on Glycemic Control in Patients with Type 2 Diabetes Mellitus. Nutrients 2018, 10, 661. [Google Scholar] [CrossRef] [PubMed]
- Hu, F.B.; Manson, J.E.; Stampfer, M.J.; Colditz, G.; Liu, S.; Solomon, C.G.; Willett, W.C. Diet, Lifestyle, and the Risk of Type 2 Diabetes Mellitus in Women. New Engl. J. Med. 2001, 345, 790–797. [Google Scholar] [CrossRef] [PubMed]
- de Souza, R.J.; Mente, A.; Maroleanu, A.; I Cozma, A.; Ha, V.; Kishibe, T.; Uleryk, E.; Budylowski, P.; Schünemann, H.; Beyene, J.; et al. Intake of saturated and trans unsaturated fatty acids and risk of all cause mortality, cardiovascular disease, and type 2 diabetes: Systematic review and meta-analysis of observational studies. BMJ 2015, 351, h3978. [Google Scholar] [CrossRef] [PubMed]
- Rosqvist, F.; Iggman, D.; Kullberg, J.; Cedernaes, J.; Johansson, H.-E.; Larsson, A.; Johansson, L.; Ahlström, H.; Arner, P.; Dahlman, I.; et al. Overfeeding Polyunsaturated and Saturated Fat Causes Distinct Effects on Liver and Visceral Fat Accumulation in Humans. Diabetes 2014, 63, 2356–2368. [Google Scholar] [CrossRef] [PubMed]
- Hu, F.B. Do functional foods have a role in the prevention of cardiovascular disease? Circulation 2011, 124, 538–540. [Google Scholar] [CrossRef]
- Brown, T.J.; Brainard, J.; Song, F.; Wang, X.; Abdelhamid, A.; Hooper, L. Omega-3, omega-6, and total dietary polyunsaturated fat for prevention and treatment of type 2 diabetes mellitus: Systematic review and meta-analysis of randomised controlled trials. BMJ 2019, 366, l4697. [Google Scholar] [CrossRef] [PubMed]
- Salmerón, J.; Hu, F.B.; E Manson, J.; Stampfer, M.J.; A Colditz, G.; Rimm, E.B.; Willett, W.C. Dietary fat intake and risk of type 2 diabetes in women. Am. J. Clin. Nutr. 2001, 73, 1019–1026. [Google Scholar] [CrossRef] [PubMed]
- Jiang, H.; Wang, L.; Wang, D.; Yan, N.; Li, C.; Wu, M.; Wang, F.; Mi, B.; Chen, F.; Jia, W.; et al. Omega-3 polyunsaturated fatty acid biomarkers and risk of type 2 diabetes, cardiovascular disease, cancer, and mortality. Clin. Nutr. 2022, 41, 1798–1807. [Google Scholar] [CrossRef] [PubMed]
- Tortosa-Caparrós, E.; Navas-Carrillo, D.; Marín, F.; Orenes-Piñero, E. Anti-inflammatory effects of omega 3 and omega 6 polyunsaturated fatty acids in cardiovascular disease and metabolic syndrome. Crit. Rev. Food Sci. Nutr. 2017, 57, 3421–3429. [Google Scholar] [CrossRef] [PubMed]
- Hereu, M.; Ramos-Romero, S.; Busquets, C.; Atienza, L.; Amézqueta, S.; Miralles-Pérez, B.; Nogués, M.R.; Méndez, L.; Medina, I.; Torres, J.L. Effects of combined d-fagomine and omega-3 PUFAs on gut microbiota subpopulations and diabetes risk factors in rats fed a high-fat diet. Sci. Rep. 2019, 9, 16628. [Google Scholar] [CrossRef] [PubMed]
- Kobyliak, N.; Abenavoli, L.; Falalyeyeva, T.; Mykhalchyshyn, G.; Boccuto, L.; Kononenko, L.; Kyriienko, D.; Komisarenko, I.; Dynnyk, O. Beneficial effects of probiotic combination with omega-3 fatty acids in NAFLD: A randomized clinical study. Minerva Medica 2018, 109, 418–428. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Zhang, M.; Pang, X.; Zhao, Y.; Wang, L.; Zhao, L. Structural resilience of the gut microbiota in adult mice under high-fat dietary perturbations. ISME J. 2012, 6, 1848–1857. [Google Scholar] [CrossRef] [PubMed]
- Okazaki, Y.; Sekita, A.; Chiji, H.; Kato, N. Consumption of lily bulb modulates fecal ratios of firmicutes and bacteroidetes phyla in rats fed a high-fat diet. Food Sci. Biotechnol. 2016, 25, 153–156. [Google Scholar] [CrossRef]
- de Lartigue, G.; Barbier de la Serre, C.; Espero, E.; Lee, J.; Raybould, H.E. Diet-induced obesity leads to the development of leptin resistance in vagal afferent neurons. Am J Physiol Endocrinol Metab. 2011, 301, E187–E195. [Google Scholar] [CrossRef] [PubMed]
- Kim, K.-A.; Gu, W.; Lee, I.-A.; Joh, E.-H.; Kim, D.-H. High Fat Diet-Induced Gut Microbiota Exacerbates Inflammation and Obesity in Mice via the TLR4 Signaling Pathway. PLoS ONE 2012, 7, e47713. [Google Scholar] [CrossRef] [PubMed]
- Fava, F.; Gitau, R.; Griffin, B.A.; Gibson, G.R.; Tuohy, K.M.; Lovegrove, J.A. The type and quantity of dietary fat and carbohydrate alter faecal microbiome and short-chain fatty acid excretion in a metabolic syndrome ‘at-risk’ population. Int. J. Obes. 2013, 37, 216–223. [Google Scholar] [CrossRef]
- Hildebrandt, M.A.; Hoffmann, C.; Sherrill–Mix, S.A.; Keilbaugh, S.A.; Hamady, M.; Chen, Y.-Y.; Knight, R.; Ahima, R.S.; Bushman, F.; Wu, G.D. High-Fat Diet Determines the Composition of the Murine Gut Microbiome Independently of Obesity. Gastroenterology 2009, 137, 1716–1724. [Google Scholar] [CrossRef] [PubMed]
- Daniel, H.; Gholami, A.M.; Berry, D.; Desmarchelier, C.; Hahne, H.; Loh, G.; Mondot, S.; Lepage, P.; Rothballer, M.; Walker, A.; et al. High-fat diet alters gut microbiota physiology in mice. ISME J. 2013, 8, 295–308. [Google Scholar] [CrossRef] [PubMed]
- Ge, Y.; Liu, W.; Tao, H.; Zhang, Y.; Liu, L.; Liu, Z.; Qiu, B.; Xu, T. Effect of industrial trans-fatty acids-enriched diet on gut microbiota of C57BL/6 mice. Eur. J. Nutr. 2018, 58, 2625–2638. [Google Scholar] [CrossRef] [PubMed]
- Caesar, R.; Tremaroli, V.; Kovatcheva-Datchary, P.; Cani, P.D.; Bäckhed, F. Crosstalk between Gut Microbiota and Dietary Lipids Aggravates WAT Inflammation through TLR Signaling. Cell Metab. 2015, 22, 658–668. [Google Scholar] [CrossRef]
- Valdés-Ramos, R.; Guadarrama-López, A.L.; Martínez-Carrillo, B.E.; Benítez-Arciniega, A.D. Vitamins and type 2 diabetes mellitus. Endocr Metab Immune Disord Drug Targets. 2015, 15, 54–63. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Tan, H.; Tang, J.; Li, J.; Chong, W.; Hai, Y.; Feng, Y.; Lunsford, L.D.; Xu, P.; Jia, D.; et al. Effects of Vitamin D Supplementation on Prevention of Type 2 Diabetes in Patients With Prediabetes: A Systematic Review and Meta-analysis. Diabetes Care 2020, 43, 1650–1658. [Google Scholar] [CrossRef]
- Cojic, M.; Kocic, R.; Klisic, A.; Kocic, G. The Effects of Vitamin D Supplementation on Metabolic and Oxidative Stress Markers in Patients With Type 2 Diabetes: A 6-Month Follow Up Randomized Controlled Study. Front. Endocrinol. 2021, 12. [Google Scholar] [CrossRef] [PubMed]
- Karonova, T.; Stepanova, A.; Bystrova, A.; Jude, E.B. High-Dose Vitamin D Supplementation Improves Microcirculation and Reduces Inflammation in Diabetic Neuropathy Patients. Nutrients 2020, 12, 2518. [Google Scholar] [CrossRef] [PubMed]
- Hoseini, R.; Rahim, H.A.; Ahmed, J.K. Decreased inflammatory gene expression accompanies the improvement of liver enzyme and lipid profile following aerobic training and vitamin D supplementation in T2DM patients. BMC Endocr. Disord. 2022, 22, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Johny, E.; Jala, A.; Nath, B.; Alam, J.; Kuladhipati, I.; Das, R.; Borkar, R.M.; Adela, R. Vitamin D Supplementation Modulates Platelet-Mediated Inflammation in Subjects With Type 2 Diabetes: A Randomized, Double-Blind, Placebo-Controlled Trial. Front. Immunol. 2022, 13, 869591. [Google Scholar] [CrossRef] [PubMed]
- Dunlop, T.W.; Väisänen, S.; Frank, C.; Molnár, F.; Sinkkonen, L.; Carlberg, C. The human peroxisome proliferator-activated receptor delta gene is a primary target of 1alpha,25-dihydroxyvitamin D3 and its nuclear receptor. J Mol Biol. 2005, 349, 248–260. [Google Scholar] [CrossRef]
- Bhatt, S.P.; Misra, A.; Pandey, R.M.; Upadhyay, A.D.; Gulati, S.; Singh, N. Vitamin D Supplementation in Overweight/obese Asian Indian Women with Prediabetes Reduces Glycemic Measures and Truncal Subcutaneous Fat: A 78 Weeks Randomized Placebo-Controlled Trial (PREVENT-WIN Trial). Sci Rep. 2020, 10, 220. [Google Scholar] [CrossRef] [PubMed]
- Sukik, A.; Alalwani, J.; Ganji, V. Vitamin D, Gut Microbiota, and Cardiometabolic Diseases-A Possible Three-Way Axis. Int J Mol Sci. 2023, 24, 940. [Google Scholar] [CrossRef] [PubMed]
- Singh, P.; Rawat, A.; Alwakeel, M.; Sharif, E.; Al Khodor, S. The potential role of vitamin D supplementation as a gut microbiota modifier in healthy individuals. Sci. Rep. 2020, 10, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Bakke, D.; Chatterjee, I.; Agrawal, A.; Dai, Y.; Sun, J. Regulation of Microbiota by Vitamin D Receptor: A Nuclear Weapon in Metabolic Diseases. Nucl. Recept. Res. 2018, 5. [Google Scholar] [CrossRef] [PubMed]
- Hussein, H.M.; Elyamany, M.F.; Rashed, L.A.; Sallam, N.A. Vitamin D mitigates diabetes-associated metabolic and cognitive dysfunction by modulating gut microbiota and colonic cannabinoid receptor 1. Eur. J. Pharm. Sci. 2021, 170, 106105. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Zhou, J.; Zhang, Y.; Tang, J.; Sun, B.; Xu, W.; Wang, X.; Chen, Y.; Sun, Z. Changes in Intestinal Microbiota Are Associated with Islet Function in a Mouse Model of Dietary Vitamin A Deficiency. J. Diabetes Res. 2020, 2020, 1–10. [Google Scholar] [CrossRef]
- Qu, B.; Yan, S.; Ao, Y.; Chen, X.; Zheng, X.; Cui, W. The relationship between vitamin K and T2DM: A systematic review and meta-analysis. Food Funct. 2023, 14, 8951–8963. [Google Scholar] [CrossRef]
- Zhang, Y.; Liu, L.; Wei, C.; Wang, X.; Li, R.; Xu, X.; Zhang, Y.; Geng, G.; Dang, K.; Ming, Z.; et al. Vitamin K2 supplementation improves impaired glycemic homeostasis and insulin sensitivity for type 2 diabetes through gut microbiome and fecal metabolites. BMC Med. 2023, 21, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Wilson, R.; Willis, J.; Gearry, R.; Skidmore, P.; Fleming, E.; Frampton, C.; Carr, A. Inadequate Vitamin C Status in Prediabetes and Type 2 Diabetes Mellitus: Associations with Glycaemic Control, Obesity, and Smoking. Nutrients 2017, 9, 997. [Google Scholar] [CrossRef]
- Infante, M.; Leoni, M.; Caprio, M.; Fabbri, A. Long-term metformin therapy and vitamin B12 deficiency: An association to bear in mind. World J. Diabetes 2021, 12, 916–931. [Google Scholar] [CrossRef] [PubMed]
- Yao, L.; Wang, Y.; Qin, S.; Zhu, S.; Wu, L. The antidiabetic drug metformin aids bacteria in hijacking vitamin B12 from the environment through RcdA. Commun. Biol. 2023, 6, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Kakarlapudi, Y.; Kondabolu, S.K.; Tehseen, Z.; Khemani, V.; J, S.K.; Nousherwani, M.D.; Saleem, F.; Abdelhameed, A.N. Effect of Metformin on Vitamin B12 Deficiency in Patients With Type 2 Diabetes Mellitus and Factors Associated With It: A Meta-Analysis. Cureus 2022, 14, e32277. [Google Scholar] [CrossRef] [PubMed]
- Bielik, V.; Kolisek, M. Bioaccessibility and Bioavailability of Minerals in Relation to a Healthy Gut Microbiome. Int. J. Mol. Sci. 2021, 22, 6803. [Google Scholar] [CrossRef] [PubMed]
- Bjørklund, G.; Dadar, M.; Pivina, L.; Doşa, M.D.; Semenova, Y.; Aaseth, J. The Role of Zinc and Copper in Insulin Resistance and Diabetes Mellitus. Curr. Med. Chem. 2020, 27, 6643–6657. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.V. Zinc and insulin in pancreatic beta-cells. Endocrine 2013, 45, 178–189. [Google Scholar] [CrossRef] [PubMed]
- Kloubert, V.; Rink, L. Zinc as a micronutrient and its preventive role of oxidative damage in cells. Food Funct. 2015, 6, 3195–3204. [Google Scholar] [CrossRef] [PubMed]
- Sauer, A.K.; Grabrucker, A.M. Zinc Deficiency During Pregnancy Leads to Altered Microbiome and Elevated Inflammatory Markers in Mice. Front. Neurosci. 2019, 13, 1295. [Google Scholar] [CrossRef] [PubMed]
- Celis, A.I.; Relman, D.A. Competitors versus Collaborators: Micronutrient Processing by Pathogenic and Commensal Human-Associated Gut Bacteria. Mol. Cell 2020, 78, 570–576. [Google Scholar] [CrossRef] [PubMed]
- Fukunaka, A.; Fujitani, Y. Role of Zinc Homeostasis in the Pathogenesis of Diabetes and Obesity. Int. J. Mol. Sci. 2018, 19, 476. [Google Scholar] [CrossRef] [PubMed]
- Sha, W.; Hu, F.; Xi, Y.; Chu, Y.; Bu, S. Mechanism of Ferroptosis and Its Role in Type 2 Diabetes Mellitus. J. Diabetes Res. 2021, 2021, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Kulaksiz, H.; Fein, E.; Redecker, P.; Stremmel, W.; Adler, G.; Cetin, Y. Pancreatic beta-cells express hepcidin, an iron-uptake regulatory peptide. J Endocrinol. 2008, 197, 241–249. [Google Scholar] [CrossRef] [PubMed]
- Berthault, C.; Staels, W.; Scharfmann, R. Purification of pancreatic endocrine subsets reveals increased iron metabolism in beta cells. Mol Metab. 2020, 42, 101060. [Google Scholar] [CrossRef] [PubMed]
- Leloup, C.; Tourrel-Cuzin, C.; Magnan, C.; Karaca, M.; Castel, J.; Carneiro, L.; Colombani, A.-L.; Ktorza, A.; Casteilla, L.; Pénicaud, L. Mitochondrial Reactive Oxygen Species Are Obligatory Signals for Glucose-Induced Insulin Secretion. Diabetes 2009, 58, 673–681. [Google Scholar] [CrossRef] [PubMed]
- Cooksey, R.C.; Jouihan, H.A.; Ajioka, R.S.; Hazel, M.W.; Jones, D.L.; Kushner, J.P.; McClain, D.A. Oxidative Stress, β-Cell Apoptosis, and Decreased Insulin Secretory Capacity in Mouse Models of Hemochromatosis. Endocrinology 2004, 145, 5305–5312. [Google Scholar] [CrossRef]
- Paganini, D.; Zimmermann, M.B. The effects of iron fortification and supplementation on the gut microbiome and diarrhea in infants and children: A review. Am. J. Clin. Nutr. 2017, 106, 1688S–1693S. [Google Scholar] [CrossRef] [PubMed]
- Salaye, L.; Bychkova, I.; Sink, S.; Kovalic, A.J.; Bharadwaj, M.S.; Lorenzo, F.; Jain, S.; Harrison, A.V.; Davis, A.T.; Turnbull, K.; et al. A Low Iron Diet Protects from Steatohepatitis in a Mouse Model. Nutrients 2019, 11, 2172. [Google Scholar] [CrossRef] [PubMed]
- Ozcan, L.; Tabas, I. Calcium signalling and ER stress in insulin resistance and atherosclerosis. J. Intern. Med. 2016, 280, 457–464. [Google Scholar] [CrossRef]
- Faysal, M.R.; Akter, T.; Hossain, M.S.; Begum, S.; Banu, M.; Tasnim, J.; Sultana, I.; Krishna, S.P.; Alam, S.; Akter, T.; et al. Study of Serum Calcium and Magnesium Levels in Type 2 Diabetes Mellitus Patients. Mymensingh Med. J. MMJ 2023, 32, 54–60. [Google Scholar] [PubMed]
- Bergillos-Meca, T.; Cabrera-Vique, C.; Artacho, R.; Moreno-Montoro, M.; Navarro-Alarcón, M.; Olalla, M.; Giménez, R.; Seiquer, I.; Ruiz-López, M.D. Does Lactobacillus plantarum or ultrafiltration process improve Ca, Mg, Zn and P bioavailability from fermented goats’ milk? Food Chem. 2015, 187, 314–321. [Google Scholar] [CrossRef]
- Kulathunga, J.; Simsek, S. A Review: Cereals on Modulating the Microbiota/Metabolome for Metabolic Health. Curr. Nutr. Rep. 2022, 11, 371–385. [Google Scholar] [CrossRef] [PubMed]
- van Trijp, M.P.H.; Schutte, S.; Esser, D.; Wopereis, S.; Hoevenaars, F.P.M.; Hooiveld, G.J.E.J.; A Afman, L. Minor Changes in the Composition and Function of the Gut Microbiota During a 12-Week Whole Grain Wheat or Refined Wheat Intervention Correlate with Liver Fat in Overweight and Obese Adults. J. Nutr. 2020, 151, 491–502. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Shi, Y.; Wei, D.; Ni, W.; Zhu, N.; Yan, X. Impact of a high dietary fiber cereal meal intervention on body weight, adipose distribution, and cardiovascular risk among individuals with type 2 diabetes. Front. Endocrinol. 2023, 14, 1283626. [Google Scholar] [CrossRef] [PubMed]
- Iversen, K.N.; Dicksved, J.; Zoki, C.; Fristedt, R.; Pelve, E.A.; Langton, M.; Landberg, R. The Effects of High Fiber Rye, Compared to Refined Wheat, on Gut Microbiota Composition, Plasma Short Chain Fatty Acids, and Implications for Weight Loss and Metabolic Risk Factors (the RyeWeight Study). Nutrients 2022, 14, 1669. [Google Scholar] [CrossRef] [PubMed]
- Fava, F.; Ulaszewska, M.M.; Scholz, M.; Stanstrup, J.; Nissen, L.; Mattivi, F.; Vermeiren, J.; Bosscher, D.; Pedrolli, C.; Tuohy, K.M. Impact of wheat aleurone on biomarkers of cardiovascular disease, gut microbiota and metabolites in adults with high body mass index: A double-blind, placebo-controlled, randomized clinical trial. Eur. J. Nutr. 2022, 61, 2651–2671. [Google Scholar] [CrossRef] [PubMed]
- Gaesser, G.A.; Rodriguez, J.; Patrie, J.T.; Whisner, C.M.; Angadi, S.S. Effects of Glycemic Index and Cereal Fiber on Postprandial Endothelial Function, Glycemia, and Insulinemia in Healthy Adults. Nutrients 2019, 11, 2387. [Google Scholar] [CrossRef]
- Williams, P.G. The Benefits of Breakfast Cereal Consumption: A Systematic Review of the Evidence Base. Adv. Nutr. Int. Rev. J. 2014, 5, 636S–673S. [Google Scholar] [CrossRef] [PubMed]
- Harris Jackson, K.; West, S.G.; Vanden Heuvel, J.P.; Jonnalagadda, S.S.; Ross, A.B.; Hill, A.M.; Grieger, J.A.; Lemieux, S.K.; Kris-Etherton, P.M. Effects of whole and refined grains in a weight-loss diet on markers of metabolic syndrome in individuals with increased waist circumference: A randomized controlled-feeding trial. Am J Clin Nutr. 2014, 100, 577–586. [Google Scholar] [CrossRef]
- Tosh, S.M. Review of human studies investigating the post-prandial blood-glucose lowering ability of oat and barley food products. Eur. J. Clin. Nutr. 2013, 67, 310–317. [Google Scholar] [CrossRef] [PubMed]
- Pereira, M.A.; Jacobs, D.R., Jr.; Pins, J.J.; Raatz, S.K.; Gross, M.D.; Slavin, J.L.; Seaquist, E.R. Effect of whole grains on insulin sensitivity in overweight hyperinsulinemic adults. Am. J. Clin. Nutr. 2002, 75, 848–855. [Google Scholar] [CrossRef]
- Nilsson, A.C.; Oöstman, E.M.; Granfeldt, Y.; Björck, I.M. Effect of cereal test breakfasts differing in glycemic index and content of indigestible carbohydrates on daylong glucose tolerance in healthy subjects. Am. J. Clin. Nutr. 2008, 87, 645–654. [Google Scholar] [CrossRef] [PubMed]
- Schwingshackl, L.; Hoffmann, G.; Lampousi, A.-M.; Knüppel, S.; Iqbal, K.; Schwedhelm, C.; Bechthold, A.; Schlesinger, S.; Boeing, H. Food groups and risk of type 2 diabetes mellitus: A systematic review and meta-analysis of prospective studies. Eur. J. Epidemiology 2017, 32, 363–375. [Google Scholar] [CrossRef] [PubMed]
- Seo, Y.S.; Lee, H.-B.; Kim, Y.; Park, H.-Y. Dietary Carbohydrate Constituents Related to Gut Dysbiosis and Health. Microorganisms 2020, 8, 427. [Google Scholar] [CrossRef] [PubMed]
- Davison, K.M.; Temple, N.J. Cereal fiber, fruit fiber, and type 2 diabetes: Explaining the paradox. J. Diabetes its Complicat. 2018, 32, 240–245. [Google Scholar] [CrossRef] [PubMed]
- Vanegas, S.M.; Meydani, M.; Barnett, J.B.; Goldin, B.; Kane, A.; Rasmussen, H.; Brown, C.; Vangay, P.; Knights, D.; Jonnalagadda, S.; et al. Substituting whole grains for refined grains in a 6-wk randomized trial has a modest effect on gut microbiota and immune and inflammatory markers of healthy adults. Am. J. Clin. Nutr. 2017, 105, 635–650. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Wang, X.; Sun, S.; Liu, S.; Huan, Y.; Li, R.; Liu, Q.; Cao, H.; Zhou, T.; Lei, L.; et al. Effects of a ready-to-eat cereal formula powder on glucose metabolism, inflammation, and gut microbiota in diabetic db/db mice. Food Sci Nutr. 2020, 8, 4523–4533. [Google Scholar] [CrossRef] [PubMed]
- Roager, H.M.; Vogt, J.K.; Kristensen, M.; Hansen, L.B.S.; Ibrügger, S.; Mærkedahl, R.B.; Bahl, M.I.; Lind, M.V.; Nielsen, R.L.; Frøkiær, H.; et al. Whole grain-rich diet reduces body weight and systemic low-grade inflammation without inducing major changes of the gut microbiome: A randomised cross-over trial. Gut 2017, 68, 83–93. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Zhang, D.; Yang, Z.; Hao, Y.; Wang, Z.; Wang, J.; Wang, Z. Wheat Alkylresorcinols Modulate Glucose Homeostasis through Improving GLP-1 Secretion in High-Fat-Diet-Induced Obese Mice. J. Agric. Food Chem. 2023, 71, 16125–16136. [Google Scholar] [CrossRef] [PubMed]
- Christensen, E.G.; Licht, T.R.; Kristensen, M.; I Bahl, M. Bifidogenic effect of whole-grain wheat during a 12-week energy-restricted dietary intervention in postmenopausal women. Eur. J. Clin. Nutr. 2013, 67, 1316–1321. [Google Scholar] [CrossRef] [PubMed]
- Costabile, A.; Klinder, A.; Fava, F.; Napolitano, A.; Fogliano, V.; Leonard, C.; Gibson, G.R.; Tuohy, K.M. Whole-grain wheat breakfast cereal has a prebiotic effect on the human gut microbiota: A double-blind, placebo-controlled, crossover study. Br. J. Nutr. 2007, 99, 110–120. [Google Scholar] [CrossRef] [PubMed]
- Carvalho-Wells, A.L.; Helmolz, K.; Nodet, C.; Molzer, C.; Leonard, C.; McKevith, B.; Thielecke, F.; Jackson, K.G.; Tuohy, K.M. Determination of the in vivo prebiotic potential of a maize-based whole grain breakfast cereal: A human feeding study. Br J Nutr. 2010, 104, 1353–1356. [Google Scholar] [CrossRef] [PubMed]
- Yu, D.; Zhu, L.; Gao, M.; Yin, Z.; Zhang, Z.; Zhu, L.; Zhan, X. A Comparative Study of the Effects of Whole Cereals and Refined Cereals on Intestinal Microbiota. Foods 2023, 12, 2847. [Google Scholar] [CrossRef] [PubMed]
- Basiak-Rasała, A.; Różańska, D.; Zatońska, K. Food groups in dietary prevention of type 2 diabetes. Rocz Panstw Zakl Hig . 2019, 70, 347–357. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Fan, Y.; Zhang, X.; Hou, W.; Tang, Z. Fruit and vegetable intake and risk of type 2 diabetes mellitus: Meta-analysis of prospective cohort studies. BMJ Open 2014, 4, e005497. [Google Scholar] [CrossRef] [PubMed]
- Carter, P.; Gray, L.J.; Troughton, J.; Khunti, K.; Davies, M.J. Fruit and vegetable intake and incidence of type 2 diabetes mellitus: Systematic review and meta-analysis. BMJ 2010, 341, c4229. [Google Scholar] [CrossRef] [PubMed]
- Mursu, J.; Virtanen, J.K.; Tuomainen, T.-P.; Nurmi, T.; Voutilainen, S. Intake of fruit, berries, and vegetables and risk of type 2 diabetes in Finnish men: The Kuopio Ischaemic Heart Disease Risk Factor Study. Am. J. Clin. Nutr. 2014, 99, 328–333. [Google Scholar] [CrossRef] [PubMed]
- Wang, P.Y.; Fang, J.C.; Gao, Z.H.; Zhang, C.; Xie, S.Y. Higher intake of fruits, vegetables or their fiber reduces the risk of type 2 diabetes: A meta-analysis. J Diabetes Investig. 2016, 7, 56–69. [Google Scholar] [CrossRef] [PubMed]
- Lakshmanan, A.P.; Mingione, A.; Pivari, F.; Dogliotti, E.; Brasacchio, C.; Murugesan, S.; Cusi, D.; Lazzaroni, M.; Soldati, L.; Terranegra, A. Modulation of gut microbiota: The effects of a fruits and vegetables supplement. Front. Nutr. 2022, 9, 930883. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Z.; Sun, T.-Y.; He, Y.; Gou, W.; Zuo, L.-S.; Fu, Y.; Miao, Z.; Shuai, M.; Xu, F.; Xiao, C.; et al. Dietary fruit and vegetable intake, gut microbiota, and type 2 diabetes: Results from two large human cohort studies. BMC Med. 2020, 18, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Sotoudeh, G.; Abshirini, M.; Bagheri, F.; Siassi, F.; Koohdani, F.; Aslany, Z. Higher dietary total antioxidant capacity is inversely related to prediabetes: A case-control study. Nutrition 2018, 46, 20–25. [Google Scholar] [CrossRef] [PubMed]
- Rienks, J.; Barbaresko, J.; Oluwagbemigun, K.; Schmid, M.; Nöthlings, U. Polyphenol exposure and risk of type 2 diabetes: Dose-response meta-analyses and systematic review of prospective cohort studies. Am. J. Clin. Nutr. 2018, 108, 49–61. [Google Scholar] [CrossRef] [PubMed]
- Duan, R.; Guan, X.; Huang, K.; Zhang, Y.; Li, S.; Xia, J.; Shen, M. Flavonoids from Whole-Grain Oat Alleviated High-Fat Diet-Induced Hyperlipidemia via Regulating Bile Acid Metabolism and Gut Microbiota in Mice. J. Agric. Food Chem. 2021, 69, 7629–7640. [Google Scholar] [CrossRef]
- Yang, T.; Zhou, W.; Xu, W.; Ran, L.; Yan, Y.; Lu, L.; Mi, J.; Zeng, X.; Cao, Y. Modulation of gut microbiota and hypoglycemic/hypolipidemic activity of flavonoids from the fruits of Lycium barbarum on high-fat diet/streptozotocin-induced type 2 diabetic mice. Food Funct. 2022, 13, 11169–11184. [Google Scholar] [CrossRef] [PubMed]
- Su, M.; Tang, T.; Tang, W.; Long, Y.; Wang, L.; Liu, M. Astragalus improves intestinal barrier function and immunity by acting on intestinal microbiota to treat T2DM: A research review. Front. Immunol. 2023, 14, 1243834. [Google Scholar] [CrossRef] [PubMed]
- Gong, X.; Xiong, L.; Bi, C.; Zhang, B. Diosmetin ameliorate type 2 diabetic mellitus by up-regulating Corynebacterium glutamicum to regulate IRS/PI3K/AKT-mediated glucose metabolism disorder in KK-Ay mice. Phytomedicine 2021, 87, 153582. [Google Scholar] [CrossRef]
- Tarwadi, K.; Agte, V. Potential of commonly consumed green leafy vegetables for their antioxidant capacity and its linkage with the micronutrient profile. Int. J. Food Sci. Nutr. 2003, 54, 417–425. [Google Scholar] [CrossRef]
- Harding, A.H.; Wareham, N.J.; Bingham, S.A.; Khaw, K.; Luben, R.; Welch, A.; Forouhi, N.G. Plasma vitamin C level, fruit and vegetable consumption, and the risk of new-onset type 2 diabetes mellitus: The European prospective investigation of cancer--Norfolk prospective study. Arch Intern Med. 2008, 168, 1493–1499. [Google Scholar] [CrossRef] [PubMed]
- Larsson, S.C.; Wolk, A. Magnesium intake and risk of type 2 diabetes: A meta-analysis. J. Intern. Med. 2007, 262, 208–214. [Google Scholar] [CrossRef] [PubMed]
- Liese, A.D.; Roach, A.K.; Sparks, K.C.; Marquart, L.; D'Agostino, R.B.; Mayer-Davis, E.J. Whole-grain intake and insulin sensitivity: The Insulin Resistance Atherosclerosis Study. Am. J. Clin. Nutr. 2003, 78, 965–971. [Google Scholar] [CrossRef]
- Schwingshackl, L.; Hoffmann, G.; Kalle-Uhlmann, T.; Arregui, M.; Buijsse, B.; Boeing, H. Fruit and Vegetable Consumption and Changes in Anthropometric Variables in Adult Populations: A Systematic Review and Meta-Analysis of Prospective Cohort Studies. PLoS ONE 2015, 10, e0140846. [Google Scholar] [CrossRef]
- Ramne, S.; Brunkwall, L.; Ericson, U.; Gray, N.; Kuhnle, G.G.C.; Nilsson, P.M.; Orho-Melander, M.; Sonestedt, E. Gut microbiota composition in relation to intake of added sugar, sugar-sweetened beverages and artificially sweetened beverages in the Malmö Offspring Study. Eur J Nutr. 2021, 60, 2087–2097. [Google Scholar] [CrossRef]
- Hu, R.; Zeng, F.; Wu, L.; Wan, X.; Chen, Y.; Zhang, J.; Liu, B. Fermented carrot juice attenuates type 2 diabetes by mediating gut microbiota in rats. Food Funct. 2019, 10, 2935–2946. [Google Scholar] [CrossRef] [PubMed]
- Nunes, S.; Viana, S.D.; Preguiça, I.; Alves, A.; Fernandes, R.; Teodoro, J.S.; Matos, P.; Figueirinha, A.; Salgueiro, L.; André, A.; et al. Blueberry Counteracts Prediabetes in a Hypercaloric Diet-Induced Rat Model and Rescues Hepatic Mitochondrial Bioenergetics. Nutrients 2021, 13, 4192. [Google Scholar] [CrossRef] [PubMed]
- Fidélix, M.; Milenkovic, D.; Sivieri, K.; Cesar, T. Microbiota modulation and effects on metabolic biomarkers by orange juice: A controlled clinical trial. Food Funct. 2020, 11, 1599–1610. [Google Scholar] [CrossRef] [PubMed]
- Kerimi, A.; Nyambe-Silavwe, H.; Gauer, J.S.; Tomás-Barberán, F.A.; Williamson, G. Pomegranate juice, but not an extract, confers a lower glycemic response on a high-glycemic index food: Randomized, crossover, controlled trials in healthy subjects. Am J Clin Nutr. 2017, 106, 1384–1393. [Google Scholar] [CrossRef] [PubMed]
- Aslam, H.; Marx, W.; Rocks, T.; Loughman, A.; Chandrasekaran, V.; Ruusunen, A.; Dawson, S.L.; West, M.; Mullarkey, E.; Pasco, J.A.; et al. The effects of dairy and dairy derivatives on the gut microbiota: A systematic literature review. Gut Microbes 2020, 12. [Google Scholar] [CrossRef] [PubMed]
- Pal, S.; Ellis, V.; Dhaliwal, S. Effects of whey protein isolate on body composition, lipids, insulin and glucose in overweight and obese individuals. Br. J. Nutr. 2010, 104, 716–723. [Google Scholar] [CrossRef] [PubMed]
- Malik, V.S.; Sun, Q.; van Dam, R.M.; Rimm, E.B.; Willett, W.C.; Rosner, B.; Hu, F.B. Adolescent dairy product consumption and risk of type 2 diabetes in middle-aged women. Am. J. Clin. Nutr. 2011, 94, 854–861. [Google Scholar] [CrossRef] [PubMed]
- Tong, X.; Dong, J.-Y.; Wu, Z.-W.; Li, W.; Qin, L.-Q. Dairy consumption and risk of type 2 diabetes mellitus: A meta-analysis of cohort studies. Eur. J. Clin. Nutr. 2011, 65, 1027–1031. [Google Scholar] [CrossRef]
- Choi, H.K.; Willett, W.C.; Stampfer, M.J.; Rimm, E.; Hu, F.B. Dairy consumption and risk of type 2 diabetes mellitus in men: A prospective study. Arch Intern Med. 2005, 165, 997–1003. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Choi, H.K.; Ford, E.; Song, Y.; Klevak, A.; Buring, J.E.; Manson, J.E. A Prospective Study of Dairy Intake and the Risk of Type 2 Diabetes in Women. Diabetes Care 2006, 29, 1579–1584. [Google Scholar] [CrossRef]
- Link-Amster, H.; Rochat, F.; Saudan, K.Y.; Mignot, O.; Aeschlimann, J.M. Modulation of a specific humoral immune response and changes in intestinal flora mediated through fermented milk intake. FEMS Immunol Med Microbiol. 1994, 10, 55–63. [Google Scholar] [CrossRef] [PubMed]
- Yilmaz, I.; Dolar, M.E.; Ozpinar, H. Effect of administering kefir on the changes in fecal microbiota and symptoms of inflammatory bowel disease: A randomized controlled trial. Turk. J. Gastroenterol. 2020, 30, 242–253. [Google Scholar] [CrossRef] [PubMed]
- Butler, M.I.; Bastiaanssen, T.F.S.; Long-Smith, C.; Berding, K.; Morkl, S.; Cusack, A.-M.; Strain, C.; Busca, K.; Porteous-Allen, P.; Claesson, M.J.; et al. Recipe for a Healthy Gut: Intake of Unpasteurised Milk Is Associated with Increased Lactobacillus Abundance in the Human Gut Microbiome. Nutrients 2020, 12, 1468. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.; Song, J.; Huang, Y.; Li, X.; Wang, H.; Zhang, Y.; Suo, H. Lactobacillus plantarum SHY130 isolated from yak yogurt attenuates hyperglycemia in C57BL/6J mice by regulating the enteroinsular axis. Food Funct. 2021, 13, 675–687. [Google Scholar] [CrossRef]
- Gray, A.; Threlkeld, R.J. Nutritional Recommendations for Individuals with Diabetes. In Feingold KR, Anawalt B, Blackman MR, Boyce A, Chrousos G, Corpas E; et al., editors. Endotext. South Dartmouth (MA): MDText.com, Inc. Copyright © 2000-2023, MDText.com, Inc.; 2000. [Google Scholar]
- Gille, D.; Schmid, A. Vitamin B12 in meat and dairy products. Nutr. Rev. 2015, 73, 106–115. [Google Scholar] [CrossRef] [PubMed]
- Fretts, A.M.; Follis, J.L.; A Nettleton, J.; Lemaitre, R.N.; Ngwa, J.S.; Wojczynski, M.K.; Kalafati, I.P.; Varga, T.V.; Frazier-Wood, A.C.; Houston, D.K.; et al. Consumption of meat is associated with higher fasting glucose and insulin concentrations regardless of glucose and insulin genetic risk scores: A meta-analysis of 50,345 Caucasians. Am. J. Clin. Nutr. 2015, 102, 1266–1278. [Google Scholar] [CrossRef] [PubMed]
- Rouhani, M.H.; Salehi-Abargouei, A.; Surkan, P.J.; Azadbakht, L. Is there a relationship between red or processed meat intake and obesity? A systematic review and meta-analysis of observational studies. Obes. Rev. 2014, 15, 740–748. [Google Scholar] [CrossRef] [PubMed]
- Pan, A.; Sun, Q.; Bernstein, A.M.; Schulze, M.B.; Manson, J.E.; Willett, W.C.; Hu, F.B. Red meat consumption and risk of type 2 diabetes: 3 cohorts of US adults and an updated meta-analysis. Am. J. Clin. Nutr. 2011, 94, 1088–1096. [Google Scholar] [CrossRef] [PubMed]
- de la Monte, S.M.; Tong, M.; Lawton, M.; Longato, L. Nitrosamine exposure exacerbates high fat diet-mediated type 2 diabetes mellitus, non-alcoholic steatohepatitis, and neurodegeneration with cognitive impairment. Mol. Neurodegener. 2009, 4, 54. [Google Scholar] [CrossRef]
- Micha, R.; Michas, G.; Mozaffarian, D. Unprocessed Red and Processed Meats and Risk of Coronary Artery Disease and Type 2 Diabetes – An Updated Review of the Evidence. Curr. Atheroscler. Rep. 2012, 14, 515–524. [Google Scholar] [CrossRef] [PubMed]
- Tong, M.; Neusner, A.; Longato, L.; Lawton, M.; Wands, J.R.; De La Monte, S.M. Nitrosamine exposure causes insulin resistance diseases: Relevance to type 2 diabetes mellitus, non-alcoholic steatohepatitis, and Alzheimer's disease. J. Alzheimer's Dis. 2009, 17, 827–844. [Google Scholar]
- Wolk, A. Potential health hazards of eating red meat. J. Intern. Med. 2016, 281, 106–122. [Google Scholar] [CrossRef] [PubMed]
- White, D.L.; Collinson, A. Red Meat, Dietary Heme Iron, and Risk of Type 2 Diabetes: The Involvement of Advanced Lipoxidation Endproducts. Adv. Nutr. Int. Rev. J. 2013, 4, 403–411. [Google Scholar] [CrossRef] [PubMed]
- Hofmann, S.M.; Dong, H.-J.; Li, Z.; Cai, W.; Altomonte, J.; Thung, S.N.; Zeng, F.; Fisher, E.A.; Vlassara, H. Improved Insulin Sensitivity Is Associated With Restricted Intake of Dietary Glycoxidation Products in the db/db Mouse. Diabetes 2002, 51, 2082–2089. [Google Scholar] [CrossRef] [PubMed]
- Sandri, M.; Dal Monego, S.; Conte, G.; Sgorlon, S.; Stefanon, B. Raw meat based diet influences faecal microbiome and end products of fermentation in healthy dogs. BMC Vet. Res. 2017, 13, 65. [Google Scholar] [CrossRef]
- Albracht-Schulte, K.; Islam, T.; Johnson, P.; Moustaid-Moussa, N. Systematic Review of Beef Protein Effects on Gut Microbiota: Implications for Health. Adv. Nutr. Int. Rev. J. 2020, 12, 102–114. [Google Scholar] [CrossRef] [PubMed]
- Le Leu, R.K.; Winter, J.M.; Christophersen, C.T.; Young, G.P.; Humphreys, K.J.; Hu, Y.; Gratz, S.W.; Miller, R.B.; Topping, D.L.; Bird, A.R.; et al. Butyrylated starch intake can prevent red meat-induced O6-methyl-2-deoxyguanosine adducts in human rectal tissue: A randomised clinical trial. Br. J. Nutr. 2015, 114, 220–230. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Li, Y.; Feng, S.; He, K.; Guo, L.; Chen, W.; Wang, M.; Zhong, L.; Wu, C.; Peng, X.; et al. Differential Effects of Dietary White Meat and Red Meat on NAFLD Progression by Modulating Gut Microbiota and Metabolites in Rats. Oxidative Med. Cell. Longev. 2022, 2022, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Shi, J.; Di Zhao, D.; Zhao, F.; Wang, C.; Zamaratskaia, G.; Li, C. Chicken-eaters and pork-eaters have different gut microbiota and tryptophan metabolites. Sci. Rep. 2021, 11, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Zou, X.; Zhao, D.; Zhao, F.; Li, C. Pork Meat Proteins Alter Gut Microbiota and Lipid Metabolism Genes in the Colon of Adaptive Immune-Deficient Mice. Mol Nutr Food Res. 2020, 64, e1901105. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.; Lin, X.; Li, H.; Li, Y.; Shi, X.; Zhao, F.; Xu, X.; Li, C.; Zhou, G. Intake of Meat Proteins Substantially Increased the Relative Abundance of Genus Lactobacillus in Rat Feces. PLoS ONE 2016, 11, e0152678. [Google Scholar] [CrossRef]
- Zhu, Y.; Shi, X.; Lin, X.; Ye, K.; Xu, X.; Li, C.; Zhou, G. Beef, Chicken, and Soy Proteins in Diets Induce Different Gut Microbiota and Metabolites in Rats. Front. Microbiol. 2017, 8, 1395. [Google Scholar] [CrossRef] [PubMed]
- Hou, Y.-Y.; Ojo, O.; Wang, L.-L.; Wang, Q.; Jiang, Q.; Shao, X.-Y.; Wang, X.-H. A Randomized Controlled Trial to Compare the Effect of Peanuts and Almonds on the Cardio-Metabolic and Inflammatory Parameters in Patients with Type 2 Diabetes Mellitus. Nutrients 2018, 10, 1565. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Wang, W.; Huang, G.; Zhang, W.; Ni, L. In vitro and in vivo evaluation of the prebiotic effect of raw and roasted almonds (Prunus amygdalus). J. Sci. Food Agric. 2016, 96, 1836–1843. [Google Scholar] [CrossRef]
- Holscher, H.D.; Guetterman, H.M.; Swanson, K.S.; An, R.; Matthan, N.R.; Lichtenstein, A.H.; A Novotny, J.; Baer, D.J. Walnut Consumption Alters the Gastrointestinal Microbiota, Microbially Derived Secondary Bile Acids, and Health Markers in Healthy Adults: A Randomized Controlled Trial. J. Nutr. 2018, 148, 861–867. [Google Scholar] [CrossRef] [PubMed]
- Ukhanova, M.; Wang, X.; Baer, D.J.; Novotny, J.A.; Fredborg, M.; Mai, V. Effects of almond and pistachio consumption on gut microbiota composition in a randomised cross-over human feeding study. Br. J. Nutr. 2014, 111, 2146–2152. [Google Scholar] [CrossRef] [PubMed]
- Holscher, H.D.; Taylor, A.M.; Swanson, K.S.; Novotny, J.A.; Baer, D.J. Almond Consumption and Processing Affects the Composition of the Gastrointestinal Microbiota of Healthy Adult Men and Women: A Randomized Controlled Trial. Nutrients 2018, 10, 126. [Google Scholar] [CrossRef] [PubMed]
- Peng, M.; Bitsko, E.; Biswas, D. Functional Properties of Peanut Fractions on the Growth of Probiotics and Foodborne Bacterial Pathogens. J. Food Sci. 2015, 80, M635–41. [Google Scholar] [CrossRef] [PubMed]
- Jiang, R.; Manson, J.E.; Stampfer, M.J.; Liu, S.; Willett, W.C.; Hu, F.B. Nut and Peanut Butter Consumption and Risk of Type 2 Diabetes in Women. JAMA 2002, 288, 2554–2560. [Google Scholar] [CrossRef] [PubMed]
- Pan, X.; Siloto, R.M.; Wickramarathna, A.D.; Mietkiewska, E.; Weselake, R.J. Identification of a Pair of Phospholipid:Diacylglycerol Acyltransferases from Developing Flax (Linum usitatissimum L.) Seed Catalyzing the Selective Production of Trilinolenin. J. Biol. Chem. 2013, 288, 24173–24188. [Google Scholar] [CrossRef]
- Viguiliouk, E.; Kendall, C.W.C.; Mejia, S.B.; Cozma, A.I.; Ha, V.; Mirrahimi, A.; Jayalath, V.H.; Augustin, L.S.A.; Chiavaroli, L.; Leiter, L.A.; et al. Effect of Tree Nuts on Glycemic Control in Diabetes: A Systematic Review and Meta-Analysis of Randomized Controlled Dietary Trials. PLoS ONE 2014, 9, e103376. [Google Scholar] [CrossRef] [PubMed]
- Zhu, L.; Sha, L.; Li, K.; Wang, Z.; Wang, T.; Li, Y.; Liu, P.; Dong, X.; Dong, Y.; Zhang, X.; et al. Dietary flaxseed oil rich in omega-3 suppresses severity of type 2 diabetes mellitus via anti-inflammation and modulating gut microbiota in rats. Lipids Heal. Dis. 2020, 19, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.-Y.; Kim, M.; Bin Oh, S.; Kim, H.-Y.; Kim, C.; Kim, T.-Y.; Park, Y.-H. Superoxide dismutase 3 prevents early stage diabetic retinopathy in streptozotocin-induced diabetic rat model. PLoS ONE 2022, 17, e0262396. [Google Scholar] [CrossRef] [PubMed]
- Schwingshackl, L.; Christoph, M.; Hoffmann, G. Effects of Olive Oil on Markers of Inflammation and Endothelial Function—A Systematic Review and Meta-Analysis. Nutrients 2015, 7, 7651–7675. [Google Scholar] [CrossRef] [PubMed]
- Sánchez-Quintero, M.J.; Delgado, J.; Medina-Vera, D.; Becerra-Muñoz, V.M.; Queipo-Ortuño, M.I.; Estévez, M.; Plaza-Andrades, I.; Rodríguez-Capitán, J.; Sánchez, P.L.; Crespo-Leiro, M.G.; et al. Beneficial Effects of Essential Oils from the Mediterranean Diet on Gut Microbiota and Their Metabolites in Ischemic Heart Disease and Type-2 Diabetes Mellitus. Nutrients 2022, 14, 4650. [Google Scholar] [CrossRef] [PubMed]
- Park, J.C.; Im, S.-H. Of men in mice: The development and application of a humanized gnotobiotic mouse model for microbiome therapeutics. Exp. Mol. Med. 2020, 52, 1383–1396. [Google Scholar] [CrossRef]
Figure 1.
Effects of Microbiota and Metabolites on Glucose Homeostasis and Insulin Resistance. A) Overgrowth of inflammatory microbial species, particularly gram-negative bacteria, promotes increased LPS and gut permeability. LPS/TLR-4 binding leads to IRS-1 phosphorylation, decrease GLUT-4 uptake, thereby increasing serum blood glucose and worsening peripheral insulin resistance. LPS/TLR-4 binding contributes to pro-inflammatory cytokine release and metabolic endotoxemia. Metabolic endotoxemia contributes to the hypothalamic JNK cascade and c-JUN activity. c-JUN interacts with IRS-1 to promote central insulin resistance. B) SCFA producing microbiota have the capability to enzymatically catalyze fermentation reactions to produce SCFA; butyrate, propionate, and acetate. SCFA enhance GLP-1 secretion, increase relative abundances of beneficial bacteria, improve gut barrier integrity, and reduce phosphorylation of IRS-1 to improve insulin resistance. C1) Primary bile acids are created in the liver, which are further converted by gut microbiota into secondary bile acids. Secondary bile acids bind to their receptors, FXR and TGR5 to enhance GLP-1 secretion and stimulate the FGF21 pathway to increase PPAR-γ, improving insulin resistance. At the same time, they promote activation of Mtorc1 pathway, which uncouples IRS-1 and promote insulin resistance. C2) Gut microbiota produce TMA, which are converted by FMO in the liver to TMA-O. TMA-O contributes to atherosclerosis, inflammatory processes, bile acid dysregulation and dysbiosis leading to increased insulin resistance. Abbreviations: LPS, Lipopolysaccharides; TLR-4, Toll-like receptor 4; IL-1, interleukin-1; TNF-α, Tumor necrosis factor alpha; IL-6, interleukin-6; IRS-1, Insulin receptor substrate 1; p-IRS-1, phosphorylated insulin receptor substrate 1; GLUT-4, glucose transport 4; JNK, Jun amino terminal kinase; C4, 4 carbon; C3, 3 carbon; C2, 2 carbon; GLP-1, Glucagon-like peptide 1; BA, bile acids; FGF21, Fibroblast Growth Factor 21; Mtorc1, Mammalian target of rapamycin complex 1; PPAR-γ, Peroxisome proliferator-activated receptor gamma; FMO, Flavin-containing monooxygenase; TMA, Trimethylamine N; TMA-O, Trimethylamine N-oxide.
Figure 1.
Effects of Microbiota and Metabolites on Glucose Homeostasis and Insulin Resistance. A) Overgrowth of inflammatory microbial species, particularly gram-negative bacteria, promotes increased LPS and gut permeability. LPS/TLR-4 binding leads to IRS-1 phosphorylation, decrease GLUT-4 uptake, thereby increasing serum blood glucose and worsening peripheral insulin resistance. LPS/TLR-4 binding contributes to pro-inflammatory cytokine release and metabolic endotoxemia. Metabolic endotoxemia contributes to the hypothalamic JNK cascade and c-JUN activity. c-JUN interacts with IRS-1 to promote central insulin resistance. B) SCFA producing microbiota have the capability to enzymatically catalyze fermentation reactions to produce SCFA; butyrate, propionate, and acetate. SCFA enhance GLP-1 secretion, increase relative abundances of beneficial bacteria, improve gut barrier integrity, and reduce phosphorylation of IRS-1 to improve insulin resistance. C1) Primary bile acids are created in the liver, which are further converted by gut microbiota into secondary bile acids. Secondary bile acids bind to their receptors, FXR and TGR5 to enhance GLP-1 secretion and stimulate the FGF21 pathway to increase PPAR-γ, improving insulin resistance. At the same time, they promote activation of Mtorc1 pathway, which uncouples IRS-1 and promote insulin resistance. C2) Gut microbiota produce TMA, which are converted by FMO in the liver to TMA-O. TMA-O contributes to atherosclerosis, inflammatory processes, bile acid dysregulation and dysbiosis leading to increased insulin resistance. Abbreviations: LPS, Lipopolysaccharides; TLR-4, Toll-like receptor 4; IL-1, interleukin-1; TNF-α, Tumor necrosis factor alpha; IL-6, interleukin-6; IRS-1, Insulin receptor substrate 1; p-IRS-1, phosphorylated insulin receptor substrate 1; GLUT-4, glucose transport 4; JNK, Jun amino terminal kinase; C4, 4 carbon; C3, 3 carbon; C2, 2 carbon; GLP-1, Glucagon-like peptide 1; BA, bile acids; FGF21, Fibroblast Growth Factor 21; Mtorc1, Mammalian target of rapamycin complex 1; PPAR-γ, Peroxisome proliferator-activated receptor gamma; FMO, Flavin-containing monooxygenase; TMA, Trimethylamine N; TMA-O, Trimethylamine N-oxide.
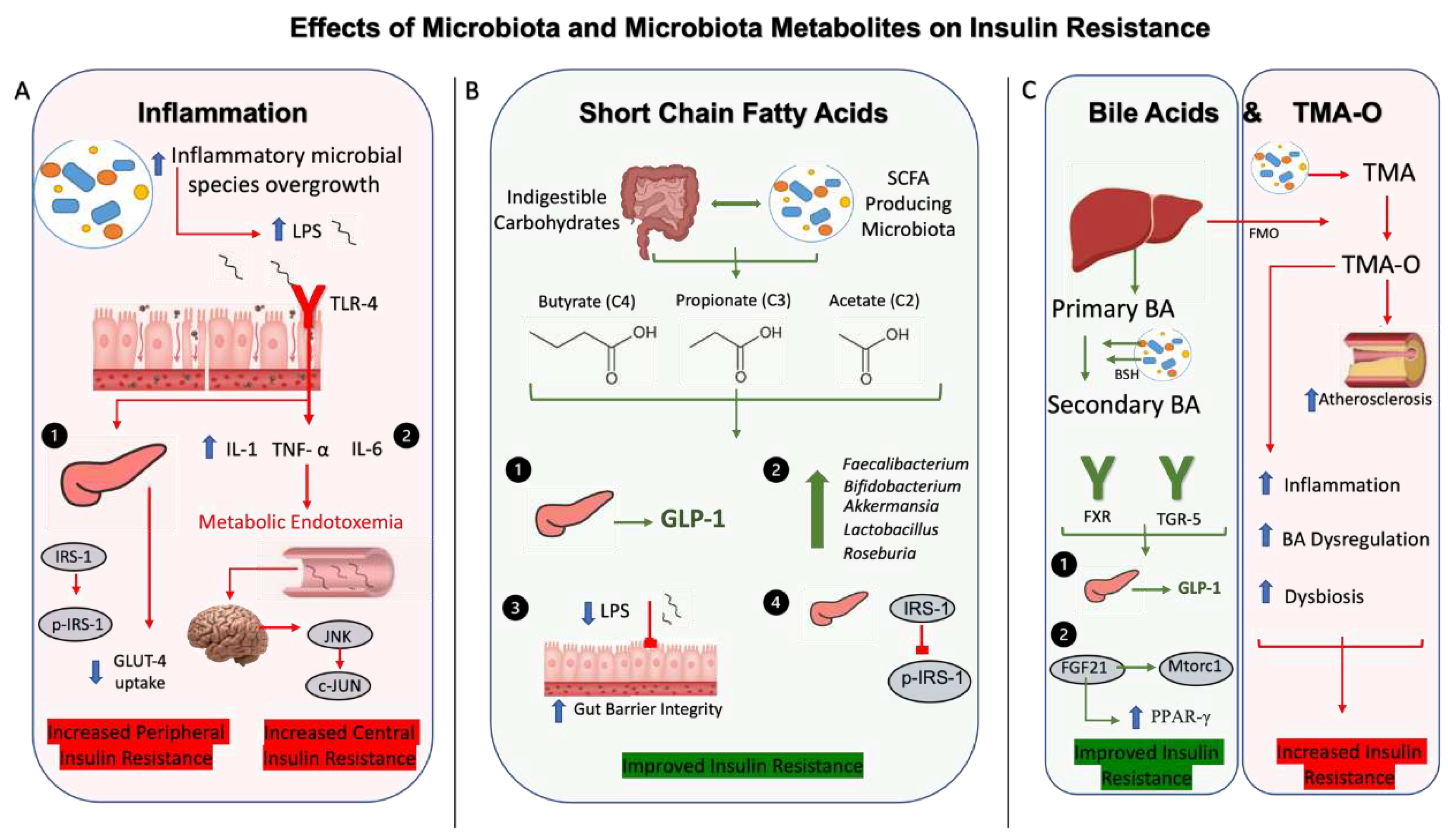
Figure 2.
Effects of Macronutrients; Carbohydrates, Proteins, Lipids on Gut Microbiota and Type 2 Diabetes Mellitus. Carbohydrates are comprised by fibers and starches. Fibers and starches promote beneficial gut microbial changes, which improve insulin resistance through increased SCFA concentrations, enhancing GLP-1 secretion, improving low grade inflammation and metabolic parameters including HgbA1c, post-prandial glucose secretion, LDL, and triglycerides. Lean meats promote beneficial gut microbial changes, but have neutral effects on HgbA1c, though they do reduce post-prandial glucose secretion. Red meats generally contribute to harmful changes in microbial composition, which promote elevations in negative microbiota metabolites including TMAO and BCAA, while decreasing SCFA, which increase risk of developing T2DM and worsening insulin sensitivity. Trans fats also promote harmful changes in microbial composition, which negatively affects leptin and insulin resistance, increases LPS/TLR4 binding and is associated with up to 40% increased risk of developing T2DM when compared to polyunsaturated fatty acids. Omega-3 polyunsaturated fatty acids, on the other hand, confer beneficial effects on gut microbiota, which decreases inflammatory markers, improves insulin sensitivity, and increases SCFA, to overall decrease the risk of developing T2DM. Abbreviations: T2DM, Type 2 Diabetes Mellitus; SCFA, Short Chain Fatty Acids; HgbA1c, Hemoglobin A1c; GLP-1, Glucagon like peptide 1; LDL, low density lipoprotein; TMAO, Trimethylamine N-oxide. BCAA, Branched chain amino acids; LPS/TLR4, Lipopolysaccharides / Toll-like receptor 4; PUFA, polyunsaturated fatty acids.
Figure 2.
Effects of Macronutrients; Carbohydrates, Proteins, Lipids on Gut Microbiota and Type 2 Diabetes Mellitus. Carbohydrates are comprised by fibers and starches. Fibers and starches promote beneficial gut microbial changes, which improve insulin resistance through increased SCFA concentrations, enhancing GLP-1 secretion, improving low grade inflammation and metabolic parameters including HgbA1c, post-prandial glucose secretion, LDL, and triglycerides. Lean meats promote beneficial gut microbial changes, but have neutral effects on HgbA1c, though they do reduce post-prandial glucose secretion. Red meats generally contribute to harmful changes in microbial composition, which promote elevations in negative microbiota metabolites including TMAO and BCAA, while decreasing SCFA, which increase risk of developing T2DM and worsening insulin sensitivity. Trans fats also promote harmful changes in microbial composition, which negatively affects leptin and insulin resistance, increases LPS/TLR4 binding and is associated with up to 40% increased risk of developing T2DM when compared to polyunsaturated fatty acids. Omega-3 polyunsaturated fatty acids, on the other hand, confer beneficial effects on gut microbiota, which decreases inflammatory markers, improves insulin sensitivity, and increases SCFA, to overall decrease the risk of developing T2DM. Abbreviations: T2DM, Type 2 Diabetes Mellitus; SCFA, Short Chain Fatty Acids; HgbA1c, Hemoglobin A1c; GLP-1, Glucagon like peptide 1; LDL, low density lipoprotein; TMAO, Trimethylamine N-oxide. BCAA, Branched chain amino acids; LPS/TLR4, Lipopolysaccharides / Toll-like receptor 4; PUFA, polyunsaturated fatty acids.
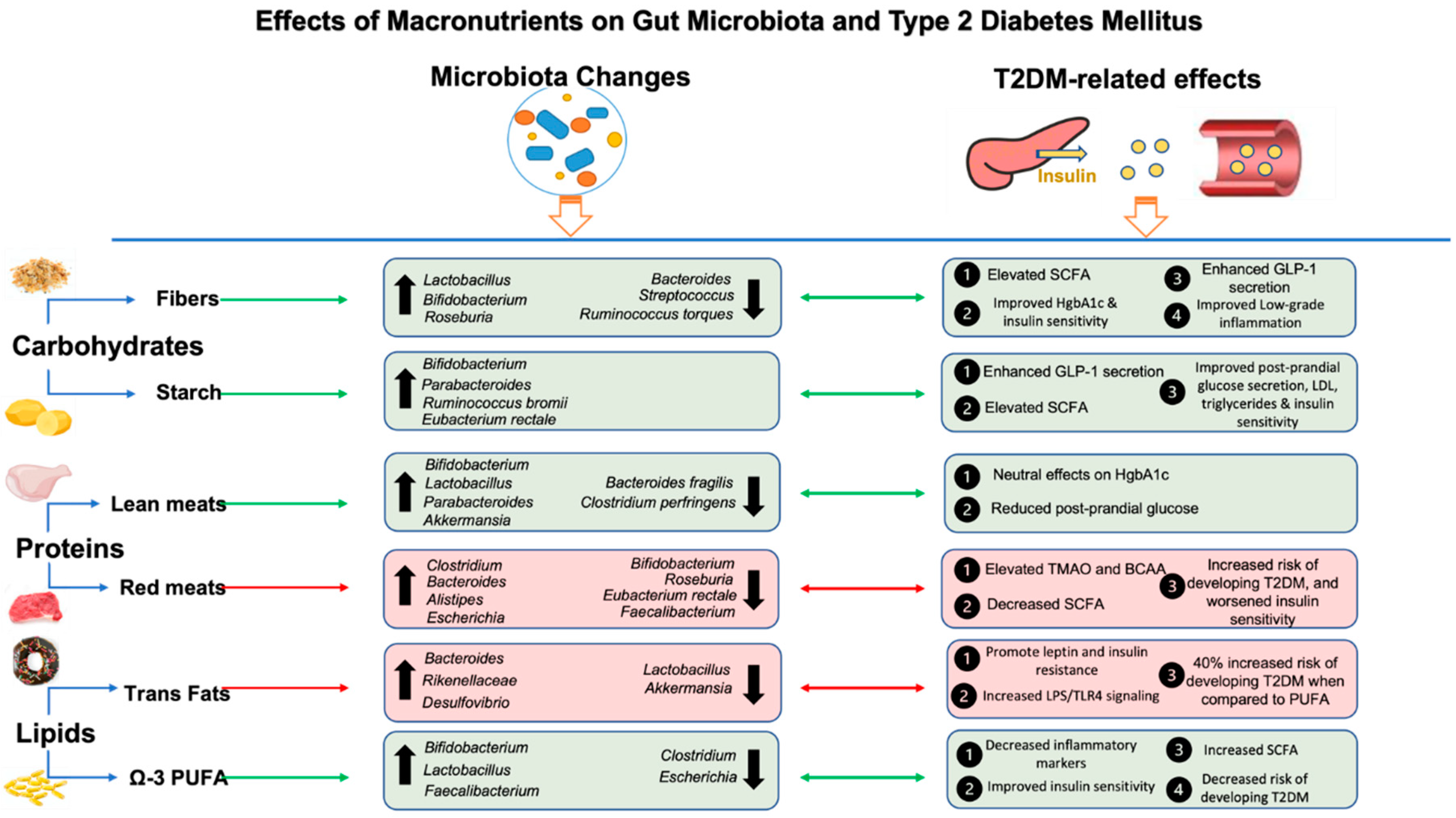
Figure 3.
Effects of Micronutrients; Vitamins and Minerals on Gut Microbiota and Type 2 Diabetes Mellitus. Vitamins generally promote beneficial changes in gut microbiota. Vitamin D has been shown to promote beneficial effects on HgbA1c and insulin resistance through decreasing reactive oxygen species, pro-inflammatory markers, and degradation of excess secondary bile acids. Water-soluble vitamins including B12 are affected by Metformin, the first-line treatment for T2DM, while Vitamin C and B12 requirements are increased, therefore supplementation is important in patients with the condition. Vitamin K improves bile acid profile, increases GLP-1 and increases SCFA to improve metabolic parameters. Minerals including Calcium and Zinc have beneficial effects on microbiota, while iron excess is associated with more negative changes in bacterial composition. Calcium improves blood glucose and HgbA1c, while increasing relative abundances of SCFA to increases mineral absorbtion through lowering colonic pH. Zinc decreases oxidative stress and is involvement in insulin storage and secretion in pancreatic β-cells. Iron, on the other hand, increases inflammatory species abundance, such as Escherichia, to promote insulin resistance, radical oxygen and nitrogen species and β-cell apoptosis. Abbreviations: T2DM, Type 2 Diabetes Mellitus; SCFA, Short Chain Fatty Acids; HgbA1c, Hemoglobin A1c; GLP-1, Glucagon like peptide 1; LPS, lipopolysaccharides; ROS, reactive oxygen species.
Figure 3.
Effects of Micronutrients; Vitamins and Minerals on Gut Microbiota and Type 2 Diabetes Mellitus. Vitamins generally promote beneficial changes in gut microbiota. Vitamin D has been shown to promote beneficial effects on HgbA1c and insulin resistance through decreasing reactive oxygen species, pro-inflammatory markers, and degradation of excess secondary bile acids. Water-soluble vitamins including B12 are affected by Metformin, the first-line treatment for T2DM, while Vitamin C and B12 requirements are increased, therefore supplementation is important in patients with the condition. Vitamin K improves bile acid profile, increases GLP-1 and increases SCFA to improve metabolic parameters. Minerals including Calcium and Zinc have beneficial effects on microbiota, while iron excess is associated with more negative changes in bacterial composition. Calcium improves blood glucose and HgbA1c, while increasing relative abundances of SCFA to increases mineral absorbtion through lowering colonic pH. Zinc decreases oxidative stress and is involvement in insulin storage and secretion in pancreatic β-cells. Iron, on the other hand, increases inflammatory species abundance, such as Escherichia, to promote insulin resistance, radical oxygen and nitrogen species and β-cell apoptosis. Abbreviations: T2DM, Type 2 Diabetes Mellitus; SCFA, Short Chain Fatty Acids; HgbA1c, Hemoglobin A1c; GLP-1, Glucagon like peptide 1; LPS, lipopolysaccharides; ROS, reactive oxygen species.
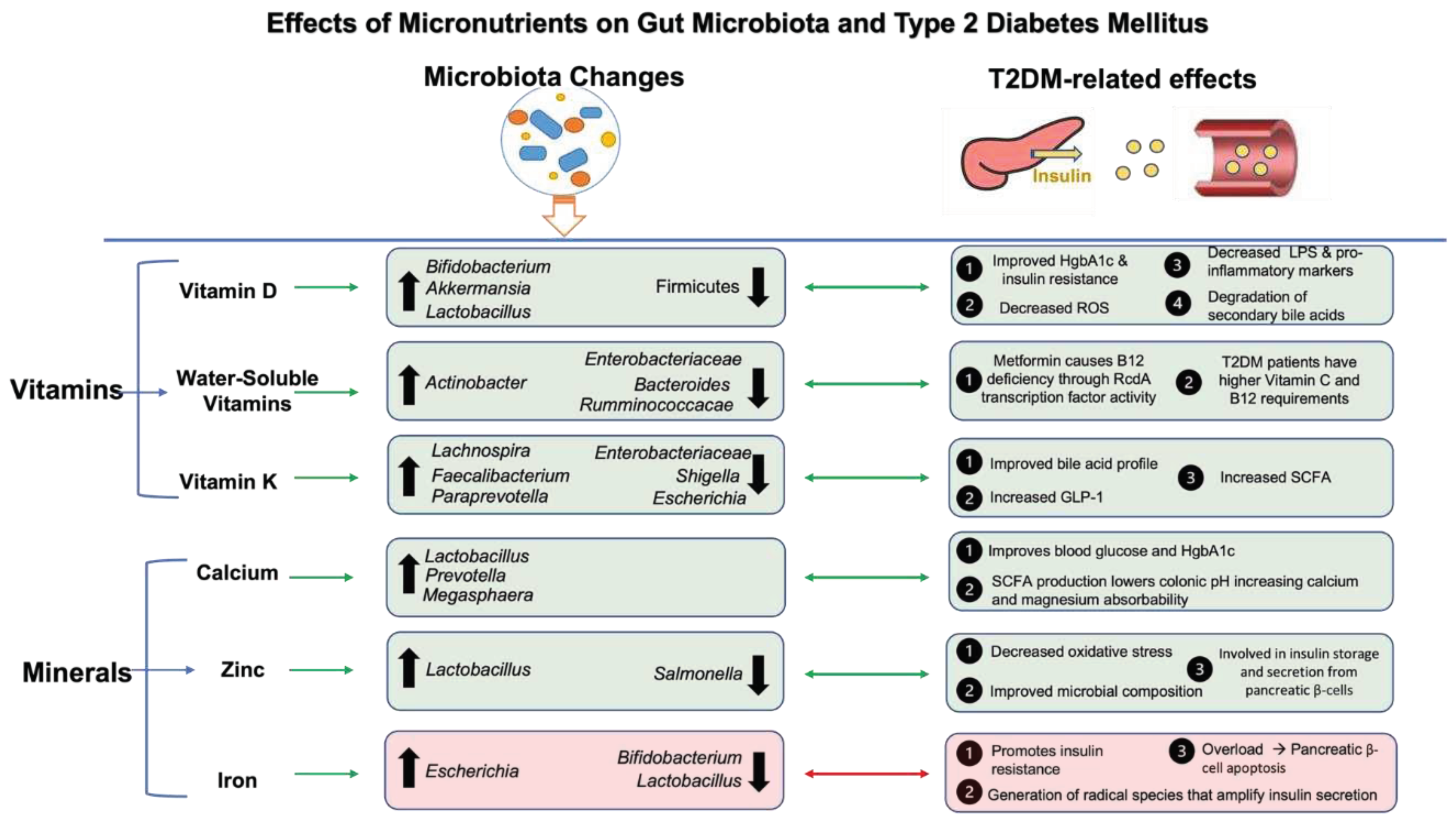
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



