
Submitted:
20 December 2023
Posted:
21 December 2023
You are already at the latest version
Abstract
Zidovudine (AZT) has significantly reduced the mortality and morbidity rates among AIDS patients, but it has been associated with hepatotoxicity. AZT treatment has been linked to disrupted lipid and glucose metabolism, and the enhancement of pro-inflammatory chemokines and other mediators, which can lead to liver steatosis. In contrast, the administration of adenosine (ADO) has proven to be an effective hepatoprotector against both acute and chronic liver damage. Therefore, present study was aimed to investigate the harmful effects of chronic AZT administration and the beneficial effects of the co-administered ADO. Chronic oral AZT administration did not induce hyperglycemia or dyslipidemia, but serum liver marker enzyme activities were increased, as well as evidence of liver steatosis and inflammation. These findings were associated with low insulin and glucagon levels, elevated serum levels of pro-inflammatory cytokines and disruptions in cell redox states in these animals. ADO co-administration attenuated the deleterious effects induced by AZT: blocking the production of inflammatory molecules, increasing serum insulin and glucagon levels, and restoring liver cellular redox state. Then, blood insulin levels seemed to be greatly influenced by the cellular redox state, which was altered by AZT administration. This alteration is likely to be connected to mitochondrial integrity and metabolism.
Keywords:
Liver redox state
; Fatty liver
; Hepatotoxicity
; Insulin
; Glucagon
; Lactic acidosis
; Cetosis
1. Introduction
Highly active anti-retroviral therapy (HAART) has significantly reduced mortality and morbidity rates in AIDS patients [1]. Zidovudine (3′-Azido-3′-deoxythymidine or AZT) is a nucleoside-analog reverse transcriptase inhibitor (NRTI), that remains as the cornerstone of treatment for a substantial portion of HAART recipients in developing countries [2]. However, it is important to note that all HIV drugs used in HAART are associated with varying degrees of adverse events [3]. Notably, HIV infection and antiretroviral drugs commonly impact organ systems such as the liver, kidneys, and heart [4]. The antiretroviral activity of AZT relies on the generation of AZT 5′-triphosphate [5]. Subsequent to this DNA damage, repair mechanisms come into play, primarily through the Nucleotide Excision Repair (NER) pathways [6], which also induce cell cycle arrest [7].
The entry of AZT into mitochondria and its subsequent conversion into AZT-5'-triphosphate within this organelle have been associated with detrimental effects on mitochondrial function, particularly when it becomes integrated into mitochondrial DNA (mtDNA) [8]. This interference alters mitochondrial bioenergetics and increases H2O2 production which impacts on respiratory complex-I function [9]. The elevated H2O2 levels rise reactive oxygen species (ROS) and peroxynitrite production, triggering single-strand DNA breaks, lipid peroxidation, protein oxidation/nitration, and mtDNA damage [10].
Prolonged exposure to antiretroviral drugs is associated with various long-term side effects, including impaired replication of mtDNA, which is manifested as microvesicular steatosis, steatohepatitis, or organ failure [11]. Additionally, long-term use of HAART has been linked to lactic acidosis, which is considered another manifestation of dysfunction in the mitochondrial respiratory chain [12]. Notably, the mitochondrial alterations induced by AZT appear to contribute to the association between HIV, long-term HAART use, and conditions such as dyslipidemia and diabetes mellitus (DM) [13], both of which are components of metabolic syndrome. It is worth emphasizing that these effects of the antiretroviral agent AZT are concentration-dependent (in the range of 25 to 100 mg ∙ kg-1) and time of exposure-dependent (ranging from 1 week to up to 12 weeks of treatment).
Administration of zidovudine alone or in combined HAART, have been associated with lipodystrophy and disturbed lipid and glucose metabolism, leading to the release of pro-inflammatory chemokines and mediators that increases the risk of DM and the onset of liver fibrosis and/or steatosis [14]. Furthermore, recent research has demonstrated that at a standard clinical dose, AZT reduces liver DNA synthesis and mitosis in rats subjected to 70% partial hepatectomy (PH), since AZT hinders liver regeneration altering PH-triggering oxidative events that play a role in the proliferation process [15].
Prior research conducted in our laboratory has demonstrated that the administration of adenosine has been tested as a hepatoprotective agent following acute ethanol intoxication [16], effectively preventing the accumulation of fat in the liver. Furthermore, adenosine delays the onset of fatty liver, hepatic necrosis, and the early oxidative stress and lipid peroxidation induced by carbon tetrachloride administration [17]. Notably, the administration of adenosine has been found to substantially reverse experimental cirrhosis and restore liver function, which are accompanied by an increased collagenolytic activity, due to decreased TIMPs levels, inhibition of cirrhosis-related oxidative stress and the restoration of the liver's capacity to proliferate effectively [18].
To investigate further, we conducted a prolonged oral administration of AZT at a slightly elevated dose, surpassing the therapeutic range (15 mg ● kg-1), to Wistar rats over a treatment period ranging from 1 to 8 weeks. During this study, we assessed the development of fatty liver, indicators of inflammation, hepatic cell redox status, and alterations in serum levels of pancreatic hormones. Additionally, we explored the potential of co-administering adenosine to mitigate the adverse effects of AZT.
2. Material and Methods
2.1. Reagents and Diagnostic Kits
Zidovudine (AZT) and adenosine (ADO) were procured from Biosynth (Carbosynth Ltd., China). Enzymes, coenzymes, and various other reagents were acquired from Sigma-Aldrich Chemical Co. (Mexico). Clinical metabolites were assessed using diagnostic kits sourced from Spinreact (Barcelona, Spain).
2.2. Animal Treatments
Male Wistar rats (n = 64) with an initial weight ranging from 190 to 210 g (2 months old) were individually housed, forming four experimental groups of four animals. They have free access to lab chow and water, within a controlled environment maintaining a 12:12-hour light/dark cycle. The amount of consumed water was meticulously recorded to determine the appropriate volume for dissolving AZT and ADO in the drinking water. This study was constituted for the following groups: A) Control group receiving only water, B) Control group consuming ADO at a dose of 30 mg ∙ kg-1/day, C) AZT alone group consuming 15 mg ∙ kg-1/day of AZT, and D) Animals in the combined treatment group, receiving 15 mg ∙ kg-1/day of AZT and 30 mg ∙ kg-1/day of ADO.
The treatment duration spanned from one to eight weeks, with a consistent intake of the nucleosides. Afterwards, rats underwent an overnight fast, anesthesia, and euthanasia. Whole blood was collected from the neck post-decapitation and serum was obtained through centrifugation. The liver and spleen were weighed, and the organ-to-body weight ratio was calculated. Liver samples were extracted for histological analysis, and the remainder of the liver was stored at -20 °C. All animal procedures adhered to the Biomedical-Ethics protocol of UNAM, in accordance with Federal Regulations for Animal Care and Experimentation (Ministry of Agriculture, SAGARPA, NOM-062-ZOO-1999).
2.3. Determination of “clinical” serum parameters
We quantified the serum levels of glucose, triglycerides (TG), cholesterol, albumin, total bilirubin, direct bilirubin, urea, and creatinine using diagnostic kits from Spinreact (Barcelona, Spain).
2.4. Determination of serum “marker” enzyme activities
In the sera collected from our experimental animals, we assessed the activities of γ-glutamyl transferase (γ-GT), amylase, and lipase using diagnostic kits from Spinreact (Barcelona, Spain). For other enzyme activities, including lactate dehydrogenase (LDH), alanine aminotransferase (ALT), aspartate aminotransferase (AST), and ornithine carbamoyl transferase (OCT), we employed previously established methods [19]. Additionally, we quantified the serum activity of arginase following the procedure outlined by Colombo and Konarska [20].
2.5. Measurement of Insulin, Glucagon, and Interleukin levels
Serum insulin and glucagon levels were quantified with ELISA kits sourced from RayBiotech (USA), while the serum concentrations of Interleukin-1β (IL-1β), Interleukin-4 (IL-4), Interleukin-6 (IL-6), and Interleukin-10 (IL-10) were determined using ELISA kits obtained from Sigma-Aldrich Chemical Co. (Mexico).
2.6. Histological assessment
Liver tissue samples were preserved in a 20% formaldehyde solution (pH 7.0 with PBS), followed by paraffin embedding, and then sectioned into 5 µm slices. These sections were subsequently stained with hematoxylin & eosin (H&E) using standard protocols. For histological analysis, a minimum of 4 biological replicates were utilized for each treatment, and five sections were systematically captured from each histological sample using a zig-zag approach. The primary histological observations encompassed findings of fatty liver, inflammatory infiltrate, and erythrocyte extravasation, which were graded as follows: 0 = absent, + = light, ++ = moderate, +++ = severe.
2.7. Estimating NAD/NADH Redox Potential via Pyruvate/Lactate and Acetoacetate/β-OH Butyrate Redox Pairs.
Acid extracts obtained from serum and liver samples were neutralized with 4 M K2CO3, and metabolites as lactate, pyruvate, acetoacetate and β-OH butyrate, were enzymatically determined as described elsewhere [21].
2.8. Calculations and statistics
Cytoplasmic and mitochondrial NAD/NADH ratios were calculated using the following formula: NAD/NADH = [oxidized substrate] / [reduced substrate] ∙ 1/Keq, while considering equilibrium constants for lactate and β-hydroxybutyrate dehydrogenases [22]. The results are presented as mean ± SE, and statistical significance of differences was evaluated through a two-way ANOVA assuming normal data distribution. Subsequently, an ad hoc Newman Keul’s test was applied, with significance defined as p < 0.01.
3. Results
3.1. Morphometric analysis and “clinical” parameters measured in sera from rats treated with AZT and ADO.
Neither zidovudine (AZT) nor ADO treatments significantly impacted the weight gain during the experimental procedure (data not shown). In addition, neither AZT nor ADO induced significant hepato- or splenomegaly (data not shown).
The administration of ADO alone significantly reduced serum glucose levels at 4 and 8 weeks compared to controls (Table 1). Administration of AZT led to significant hypoglycemia at 2 and 8 weeks post-treatment, which was rectified by combined AZT and ADO treatment. Indeed, co-administration of AZT and ADO resulted in elevated serum glucose levels at the beginning of the experiment (1 to 2 week; Table 1). ADO alone led to a decrease in serum TG levels compared to controls, starting at week 2, similar to that observed after AZT treatment (Table 1). Co-administration of AZT and ADO did not alter the reduced levels of circulating TG induced by either AZT or ADO alone. Conversely, serum cholesterol levels were significantly reduced only with the concurrent administration of the nucleosides at 2 and 8 weeks, in comparison to the control group (Table 1).
As indicators of liver function, we assessed serum albumin and bilirubin levels in rats treated with AZT and ADO. Animals receiving ADO alone or AZT + ADO did not exhibit significant changes in blood albumin levels (Table 1). Regarding total and direct bilirubin, a significant increase was observed in total bilirubin after 2 weeks of AZT administration, which promptly returned to control values (Table 1). Changes in total bilirubin appeared to be primarily associated with its indirect form, as the serum levels of direct bilirubin were only elevated at the outset of the experiment with combined treatment (Table 1). When assessing nitrogen metabolites as reflective of kidney function, minor but statistically significant changes were observed in animals administered with AZT and/or ADO (Table 1). Nonetheless, these results suggest that neither liver nor kidney functions were significantly affected by our experimental treatments.
3.2. Effects of AZT and ADO on Serum Enzyme Markers for Liver and Pancreas Integrity
When evaluating enzyme activities as markers of liver integrity, including transaminases, LDH, and γ-GT, we observed that ADO alone did not alter the enzyme profiles of these markers. However, the administration of AZT significantly increased serum activities of ALT and AST after 8 weeks, an effect that was completely mitigated by concurrent ADO treatment (Figure 1A and B). We observed an early rise in LDH activity when AZT was administered, which persisted throughout the experiment; once again, co-administration of ADO normalized LDH serum activity (Figure 1C). In contrast, AZT administration led to a reduction in γ-GT activity compared to the control group, and this effect remained unaltered by ADO administration (Figure 1D). Notably, the combined administration of AZT and ADO resulted in the lowest serum activities for all of these enzymes (Figure 1A to D).
The serum OTC activity increased after 4 weeks of AZT treatment, but the combined treatment led to a progressive and higher enhancement of this enzyme activity (Figure 1E). Conversely, ADO alone or in combination with AZT induced a substantial reduction in serum arginase activity, while the AZT-alone group exhibited a pattern for arginase activity similar to that of LDH (Figure 1F). We also assessed serum enzyme activities as markers of pancreatic integrity, specifically amylase and lipase. Here, only lipase activity showed a significant increase at 8 weeks after AZT treatment, which was effectively mitigated by co-administering ADO (Figure 1H). Unlike the metabolites described in Table 1, enzyme variations were evident depending on the treatment tested (Figure 1).
3.3. Effects of AZT and ADO on Serum Insulin and Glucagon Levels
We sought to determine whether fluctuations in insulin and/or glucagon played a role in the metabolic effects induced by the nucleosides in fasted animals. Oral ADO administration alone led to an initial increase in insulin levels after 2 weeks, which normalized thereafter. While AZT treatment alone did not result in significant changes, the combined treatment with ADO triggered a substantial and sustained increase in insulin compared to both the control and AZT groups, which disappears after 8 weeks of treatment (Figure 2A). In the same context, treatment with ADO alone or in combination with AZT also elevated serum glucagon concentrations (Figure 2B), maintaining the insulin/glucagon ratio in animals receiving the combined treatment when compared to the control group, except for AZT at the latest experimental time, where augmented this ratio (Figure 2C).
3.4. Histological assessment of the liver structure and serum levels of some cytokines involved in the inflammatory process
Liver tissue slices were subjected to histological examination to assess the presence of the following: 1) fatty change, 2) cell ballooning, 3) inflammatory infiltrates (PMN and macrophage cells), 4) hepatocellular disorganization, and 5) interstitial fibrosis. Figure 3 displays micrographs of liver tissue specimens observed with a 40X objective. In the control group, only a slight accumulation of fatty droplets was observed, primarily in relation to age. Conversely, animals treated with AZT alone exhibited a gradual increase in fat droplets, becoming more pronounced from 1 to 4 weeks and reaching peak accumulation at 8 weeks, indicative of liver steatosis (Figure 3). Co-administration with ADO also resulted in a progressive fat accumulation, but by the end of the experiment, the extent of fatty liver was significantly reduced (Figure 3).
Another cellular histological parameter we assessed was the presence of inflammatory infiltrate and erythrocyte extravasation. These features were observed with the AZT treatment, peaking at 4 weeks post-treatment, and subsequently subsided (Figure 3). Surprisingly, concurrent administration of ADO with AZT delayed this inflammatory response, leading to its persistence beyond that observed in the AZT-alone group (Figure 3).
To corroborate the observed fatty changes, we enzymatically quantified liver triglyceride (TG) levels. At the end of the experiment, control animals exhibited 8.4 ± 0.7 mg ∙ g-1 of liver compared to 17.8 ± 2.1 mg • g-1 of liver in the AZT group (p < 0.01 compared to controls). In contrast, animals receiving the combined treatment had liver TG levels of 11.2 ± 1.1 mg ∙ g-1 of liver (p < 0.01 compared to the AZT group). Notably, events such as hepatocellular disorganization, ballooning, or fibrosis were absent.
To confirm the presence of liver inflammation induced by AZT administration, we quantified serum levels of specific cytokines. Treatment with ADO alone or in combination with AZT significantly decreased the inflammatory cytokine IL-1β, while AZT alone markedly increased its serum levels after 8 weeks of treatment. Notably, this AZT-induced increase in IL-1β was completely attenuated by the concurrent administration of ADO (Figure 4A). A similar pattern was observed for IL-6, another pro-inflammatory cytokine. ADO alone significantly reduced IL-6 levels at 2 weeks post-treatment, while AZT alone induced a significant increase in IL-6 after 4 weeks. Once again, co-administration with ADO effectively mitigated the AZT-induced rise in IL-6 (Figure 4B).
The IL-4, a cytokine with various immune system effects associated with an anti-inflammatory response, exhibited an intriguing pattern. Treatment with ADO or AZT alone elevated serum IL-4 levels at 1 to 2 weeks, while co-administration of the nucleosides also increased IL-4 compared to controls (Figure 4C). At 4 weeks, the AZT group reached the highest serum IL-4 levels, which were partially reduced by the combined treatment. Interestingly, at the latest time point studied (8 weeks), we observed an inverse relationship in serum IL-4 levels between the AZT-alone group and the group co-administered with ADO (Figure 4C). The IL-10, a cytokine recognized for promoting a generalized anti-inflammatory and regulatory response, exhibited a robust induction by ADO, while AZT alone showed only a modest peak at 2 weeks of treatment (Figure 4D). Thereafter, neither group displayed significant variations in IL-10 levels (Figure 4D).
3.5. Effects of AZT and ADO on Serum, Liver NAD/NADH Redox State (Cytoplasmic and Mitochondrial), and levels of Ketone Bodies
our group has recently revealed a strong correlation between blood glucose variations, cellular redox state, and the insulin/glucagon ratio [23]. In light of these observations, we quantified the concentrations of pyruvate/lactate and acetoacetate/β-OH-butyrate redox pairs to estimate the NAD/NADH redox state in both the cytoplasm and mitochondria, respectively, and explored their potential reflection in serum levels in our experimental rat model.
In the serum, lactate levels were significantly reduced in the group of animals receiving the combined treatment of AZT plus ADO at all experimental time points, in comparison to controls and/or AZT alone (Figure 5, panels A). Conversely, AZT induced a significant increase in serum lactate at 8 weeks of treatment, an effect completely mitigated by co-administration with ADO (Figure 5, panels A).
Regarding serum pyruvate levels, ADO alone or in combination led to a decrease, whereas AZT alone resulted in a significant peak of serum pyruvate after 4 weeks, with a non-significant increase at 8 weeks of treatment (Figure 5, panels B). Consequently, serum NAD/NADH ratios were markedly increased by AZT administration. When ADO was co-administered with AZT, this ratio was also elevated at 1 week of treatment but subsequently declined, returning to the control range (Figure 5, panels C).
In contrast to serum observations, AZT alone reduced liver lactate levels, while co-administration with ADO restored lactate levels, and ADO alone only diminished lactate during the first 2 weeks in the liver (Figure 5, panels A). Liver pyruvate levels exhibited contrasting trends: ADO alone or in combination led to an increase, while AZT alone reduced this monocarboxylic acid only at the beginning of the treatment (Figure 5, panels B). Notably, chronic ADO administration induced a more oxidized liver cytoplasmic redox state, whereas AZT had a similar effect at 2 weeks, and the combination of nucleosides resulted in a higher NAD/NADH ratio at 4 weeks of treatment (Figure 5, panels C).
Regarding the acetoacetate/β-OH-butyrate redox pair, AZT administration significantly increased β-OH-butyrate levels, an effect that was reversed or normalized by co-administration with ADO (Figure 6, panels A). In terms of serum acetoacetate levels, AZT initially decreased them, a change that was normalized by co-administration with ADO (Figure 6, panels B). As a consequence, AZT alone consistently led to a lower NAD/NADH ratio at all experimental time points, an effect corrected in the AZT + ADO group, which exhibited the highest NAD/NADH ratio at 1 week of treatment (Figure 6, panels C).
Regarding the mitochondrial NAD/NADH redox state in the liver, AZT administration significantly decreased β-OH-butyrate levels, which were corrected by the combined treatment at 4 weeks, with no further significant changes observed thereafter (Figure 6, panels A). Conversely, liver acetoacetate levels were significantly reduced by treatments with ADO alone, AZT alone, or even in co-administration at earlier time points. However, simultaneous administration of AZT and ADO induced a peak in acetoacetate levels at 4 weeks of treatment (Figure 6, panels B). In animals treated with ADO or AZT alone, a reduced mitochondrial state was observed, particularly in those receiving AZT, but this was normalized by co-administration with ADO at later time points.
ADO alone decreased the amount of ketone bodies in the serum (at 8 weeks) and in the liver (at 4 weeks) (Table 2). AZT treatment resulted in increased serum ketone body levels (ketosis) at 2 and 4 weeks, while the opposite trend was seen in the liver, suggesting an elevated release of these metabolites (Table 2). Co-administration with ADO prevented AZT-induced ketosis and tended to decrease liver ketone body levels, possibly due to an anti-lipogenic effect of ADO (Table 2).
4. Discussion
Since the introduction of HAART in the mid-1990s, the lifespan of individuals living with HIV has substantially increased. However, the major causes of morbidity and mortality in these patients have shifted towards liver, renal, and cardiovascular diseases [24]. In the context of liver health, AZT has been associated with peroxide production and oxidative stress in hepatocytes [25], possibly linked to AZT-induced glutathione depletion [26].
Oxidative stress can lead to elevated levels of liver enzymes, often considered specific markers of liver toxicity. Treatment with AZT has been shown to significantly increase liver enzyme levels, including alkaline phosphatase, ALT, and AST [27]. Early initiation of HAART is often associated with marked elevations in AST, ALT, and BUN, with some resolution expected as treatment progresses [28]. Nevertheless, there is evidence that a significantly increased number of HIV-infected individuals treated with AZT have died due to hepatotoxicity resulting from lipid dysregulation, steatosis, steatohepatitis, hepatomegaly, and abnormal liver function [29].
Furthermore, there have been reports of coexisting dyslipidemia and hyperglycemia in individuals starting HAART that includes AZT, significantly increasing the risk factors for the development of both diseases [30]. In contrast to what is observed in patients, the administration of AZT in our experimental model led to significant hypoglycemia at 2 and 8 weeks after treatment, a phenomenon corrected by the combined treatment of AZT plus ADO, which, in fact, induced a transient hyperglycemia (Table 1). Additionally, AZT decreased serum TG levels without significantly affecting cholesterol levels (Table 1). Therefore, we did not observe chronic hyperglycemia or dyslipidemia resulting from AZT administration in our study.
Conversely, AZT treatment significantly altered marker enzymes of liver integrity (Figure 1). The nucleoside increased ALT, AST, and LDH levels, primarily after 8 weeks of administration, while decreasing γ-GT activity. Co-administration with ADO normalized these parameters (Figure 1). Furthermore, AZT increased the serum activity of arginase, while that of OCT was clearly diminished (Figure 1). Both type-I arginase and OCT are used to assess hepatocellular damage, including acute hepatitis, cirrhosis, or cancer [31,32].
In accordance with the serum enzymatic patterns induced by AZT, we observed liver fat accumulation in rats treated with AZT alone, reaching its peak at 8 weeks, a condition considered as liver steatosis (Figure 3). This phenomenon was partially prevented or delayed by simultaneous administration with ADO (Figure 3).
Liver inflammation, as assessed by polymorphonuclear (PMN), macrophages, and erythrocyte infiltration, was observed in AZT-treated animals primarily after 4 weeks of treatment, but appeared to resolve thereafter. In this context, ADO administration clearly delayed the liver's inflammatory histological pattern induced by AZT (Figure 3). To explain the AZT effects observed in this study, it has been reported that AZT adversely affects the oxidation of free fatty acids within hepatic mitochondria, which can be associated to NRTIs-induced insulin resistance and dyslipidemia, leading to liver TG accumulation and subsequent hepatic steatosis [24].
In mice, AZT exposure reduces mtDNA copy number in the liver, which can also affect pancreatic islet size. Administration of AZT reduced the mean insulin-positive cell area/islet when compared with control mice. Therefore, reduced mtDNA copy number (caused by AZT treatment) could impair insulin supply and sensing [33].
In our study, administration of ADO alone induced an enhancement of insulin after 2 weeks, which normalized thereafter. While treatment with AZT alone did not promote significant changes, combined treatment with ADO elicited a drastic increase in insulin compared to either the control or AZT groups (Figure 2A). Additionally, treatment with ADO alone or simultaneously with AZT elevated the serum concentration of glucagon (Figure 2B), maintaining the insulin/glucagon ratio in animals receiving the combined treatment, compared to controls (Figure 2C). Our results indicate that AZT did not induce insulin resistance in our rats at any time during the treatment, and hyperinsulinemia was only present in animals receiving AZT + ADO (Figure 2). Although it is challenging to explain how ADO combined with AZT can elevate serum glucose levels in overnight fasted rats, it is possible that the ADO-induced increase in serum glucagon may partially explain this effect, by affecting the liver gluconeogenesis. During fasting, increased glucagon levels play a key role in stimulating fatty acid oxidation and activating PPAR-α through its effects on PKA, suggesting a potential crosstalk between these signaling pathways [34].
AZT treatment in female C57BL/6J mice significantly increased hepatic TG levels, inflammation, and oxidative stress. Nitration, glycation, and lipid peroxidation were also significantly higher in the AZT-treated mice compared to vehicle controls [35]. During inflammation, cytokines released by immune cells, including TNF-α and IL-1β, directly affect the endothelium, increasing the expression of cell adhesion molecules (ICAM-1 and VCAM-1) and E-selectin [36].
Administration of ADO, either alone or with AZT, induced a significant decrease in the inflammatory cytokines IL-1β and IL-6, which were elevated by AZT alone (4 to 8 weeks post-treatment). As for the anti-inflammatory cytokines, treatment with ADO or AZT alone led to an increase in the serum concentration of IL-4 at 1 to 2 weeks after administration (Figure 4). Indeed, it has been reported that the IFC-305 compound (an aspartic-salt derivative of ADO) showed an antioxidant and anti-inflammatory activities, probably by IL-10 and arginase activity, highlighting that the IFC-305 prevents or even reverses an experimental model of carbon tetrachloride-induced cirrhosis in rats [37].
Previous studies have demonstrated that adverse effects of nucleoside analogs used in HAART are directly linked to mitochondrial injury [38]. Mitochondrial damage manifests as abnormal morphology, depletion of mitochondrial-encoded enzymes, and reduced numbers of mitochondrial genes [39]. From here, mitochondrial disruption results in energy loss, electron leakage from the electron transport system, increased levels of reactive oxygen species (ROS), oxidative damage, and significant imbalances in cellular redox states (i.e., an increased NADH/NAD ratio), which shift the pyruvate/lactate balance in favor of increased lactate production [40]. Liver and skeletal muscle tissues are recognized as important targets for mitochondrial injury induced by nucleoside analogs [10,41]. Additionally, AZT renders mitochondrial membranes more susceptible to damage by free fatty acids, induces morphological changes, and activates nuclear factor κBp65 in cardiac tissue in mice [9]. AZT also promotes ROS-mediated oxidative damage, triggering ADP-ribosylation reactions and accelerated NAD+ catabolism, all contributing to the development of liver toxicity in animal models [27]. Hence, we explored the potential role of changes in cellular redox states and their deleterious effects on the liver in animals treated with AZT (Figs. 5 and 6, Table 2).
Under our experimental conditions, AZT induced lactic acidosis, but this lactate does not appear to be produced by the liver, as observed in Figure 5. Additionally, the levels of pyruvate showed fewer changes induced by either AZT or ADO. Therefore, the serum NAD/NADH ratio was more oxidized in animals receiving AZT alone, and the liver cytoplasmic NAD/NADH ratio was only transiently oxidized by AZT alone. Co-administration with ADO normalized the serum NAD/NADH ratio and induced a more oxidized liver cytoplasmic NAD/NADH ratio of animals administered with AZT (Figure 5).
The serum amount of β-OH-butyrate was augmented by AZT treatment and inversely correlated with the liver level of β-OH-butyrate, while the serum acetoacetate showed minimal changes. Thus, AZT clearly induced a more reduced serum NAD/NADH ratio of the ketone bodies redox-pair, which reflects that of liver mitochondria (Figure 6). The alterations in the fluctuations of cytoplasmic and mitochondrial redox states, with the former being more oxidized and the latter showing a reduced condition, especially after 2 and 4 weeks of treatment, could be related to progressive mitochondrial dysfunction induced by AZT, similar to well-known hepatotoxins [21].
5. Conclusions
Our long-lasting oral administration of AZT to rats for 1 up to 8 weeks did not result in hyperglycemia or dyslipidemia attributable to the nucleoside treatment. We did not observe alterations in parameters indicative of liver function. However, we did record elevations in marker enzymes in liver tissue, despite the absence of necrotic images in liver histological samples. Fatty infiltration indicated the onset of liver steatosis and inflammation. These findings were associated with hypoglycemia, low serum insulin and glucagon levels, and elevated serum levels of pro-inflammatory cytokines in animals treated with AZT. Co-administration of adenosine largely attenuated the deleterious effects induced by AZT, including the production of inflammatory molecules. Furthermore, it increased serum levels of insulin and glucagon, and restored the altered cellular redox state in the liver caused by AZT. Thus, blood insulin levels were significantly influenced by the cellular redox state, which was altered by chronic AZT administration, likely linked to mitochondrial integrity and metabolism.
Authors Contribution: Conceptualization: MLCZ, ABO, RHM; Methodology: MLCZ, LSS, RHM, RPD; Investigation; MLCZ, ABO, LSS, RHM; Visualization: MLCZ, LSS, RHM, ABO, RPD; Funding acquisition: MLCZ, RHM; Project administration: MLCZ, RHM; Supervision: MLCZ, RHM; Writing (Original draft): MLCZ, ABO, RHM; Writing (Review & editing): MLCZ, LSS, RHM, ABO, RPD.
Funding
Authors acknowledge the partial funding support to the present investigation from PAPIIT grants # IT200420 and # IN210623 of the Dirección General de Asuntos del Personal Académico (DGAPA).
Data Availability Statement
There is availability to share data and materials.
Acknowledgments
We thank the expert aid from Biols. Gerardo Coello, Ana María Escalante, Eng. Juan Manuel Barbosa, and Ivette Rosas, for designing figures, as well as to Eng. Aurey Galván and Manuel Ortínez, for maintenance and supervision of the laboratory equipment. The valuable aid of medical Vets Claudia Rivera and Hector Malagón is also acknowledged.
Conflicts of Interest
Authors declare that they do not have any kind of conflict of interest.
References
- Palella, F.J., Jr; Delaney, K.M.; Moorman, A.C.; Loveless, M.O.; Fuhrer, J.; Satten, G.A.; Aschman, D.J.; Holmberg, S.D. Declining morbidity and mortality among patients with advanced human immunodeficiency virus infection. HIV Outpatient Study Investigators. N. Engl. J. Med. 1998, 338, 853–860. [Google Scholar] [CrossRef]
- Chadwick, D.; Stanley, A.; Sarfo, S.; Appiah, L.; Ankcorn, M.; Foster, G.; Schwab, U.; Phillips, R.; Geretti, A.M. Response to antiretroviral therapy in occult hepatitis B and HIV co-infection in West Africa. AIDS. 2013, 27, 139–141. [Google Scholar] [CrossRef] [PubMed]
- UNAIDS. UNAIDS Data 2017. 2017. Available online: http://www.unaids. org/sites/default/files/media_asset/20170720_Data_book_ 2017_en.pdf.
- Smith, C.J.; Ryom, L.; Weber, R.; Morlat, P.; Pradier, C.; Reiss, P.; Kowalska, J.D.; de Wit, S.; Law, M.; el Sadr, W.; et al. D:A:D Study Group. Trends in underlying causes of death in people with HIV from 1999 to 2011 (D:A:D): A multicohort collaboration. Lancet. 2014, 384, 241–248. [Google Scholar] [CrossRef] [PubMed]
- Fang, J.L.; Han, T.; Wu, Q.; Land, F.A.; Chang, C.W.; Guo, L.; Fuscoe, J.C. Differential gene expression in human hepatocyte cell lines exposed to the antiretroviral agent zidovudine. Arch. Toxicol. 2014, 88, 609–623. [Google Scholar] [CrossRef] [PubMed]
- Wu, Q.; Beland, F.A.; Chang, C.W.; Fang, J.L. Role of DNA repair pathways in response to zidovudine-induced DNA damage in immortalized human liver THLE2 cells. Int. J. Biomed. Sci. 2013, 9, 18–25. [Google Scholar] [PubMed]
- Fang, J.L.; Beland, F.A. Long-term exposure to zidovudine delays cell cycle progression, induces apoptosis, and decreases telomerase activity in human hepatocytes. Toxicol. Sci. 2009, 111, 120–130. [Google Scholar] [CrossRef]
- Govindarajan, R.; Leung, G.P.; Zhou, M.; Tse, C.M.; Wang, J.; Unadkat, J.D. Facilitated mitochondrial import of antiviral and anticancer nucleoside drugs by human equilibrative nucleoside transporter-3. Am. J. Physiol. Gastrointest. Liver Physiol. 2009, 296, G910–G922. [Google Scholar] [CrossRef] [PubMed]
- Pupure, J.; Fernandes, M.A.; Santos, M.S.; Moreno, A.J.; Kalvinsh, I.; Klusa, V.; Oliveira, C.R. Mitochondria as the target for mildronate's protective effects in azidothymidine (AZT)-induced toxicity of isolated rat liver mitochondria. Cell Biochem. Funct. 2008, 26, 620–631. [Google Scholar] [CrossRef] [PubMed]
- de la Asunción, J.G.; del Olmo, M.L.; Sastre, J.; Pallardó, F.V.; Viña, J. Zidovudine (AZT) causes an oxidation of mitochondrial DNA in mouse liver. Hepatology 1999, 29, 985–987. [Google Scholar] [CrossRef] [PubMed]
- Morris, A.A.; Taanman, J.W.; Blake, J.; Cooper, J.M.; Lake, B.D.; Malone, M.; Love, S.; Clayton, P.T.; Leonard, J.V.; Schapira, A.H. Liver failure associated with mitochondrial DNA depletion. J. Hepatol. 1998, 28, 556–563. [Google Scholar] [CrossRef] [PubMed]
- Chariot, P.; Drogou, I.; de Lacroix-Szmania, I.; Eliezer-Vanerot, M.C.; Chazaud, B.; Lombès, A.; Schaeffer, A.; Zafrani, E.S. Zidovudine-induced mitochondrial disorder with massive liver steatosis, myopathy, lactic acidosis, and mitochondrial DNA depletion. J. Hepatol. 1999, 30, 156–160. [Google Scholar] [CrossRef]
- Abebe, M.; Kinde, S.; Belay, G.; Gebreegziabxier, A.; Challa, F.; Gebeyehu, T.; Nigussie, P.; Tegbaru, B. Antiretroviral treatment associated hyperglycemia and dyslipidemia among HIV infected patients at Burayu Health Center, Addis Ababa, Ethiopia: A cross-sectional comparative study. BMC Res. Notes 2014, 7, 380. [Google Scholar]
- Han, W.M.; Apornpong, T.; Lwin, H.M.S.; Thammapiwan, S.; Boonrungsirisap, J.; Gatechompol, S.; Ubolyam, S.; Tangkijvanich, P.; Kerr, S.J.; Avihingsanon, A. Non-alcoholic fatty liver disease and non-alcoholic steatohepatitis with liver fibrosis as predictors of new-onset Diabetes Mellitus in people living with HIV: A longitudinal cohort study. Clin. Infect. Dis. 2023, 21, ciad433. [Google Scholar]
- Butanda-Ochoa, A.; Hernández-Espinosa, D.R.; Olguín-Martínez, M.; Sánchez-Sevilla, L.; Rodríguez, M.R.; Chávez-Rentería, B.; Aranda-Fraustro, A.; Hernández-Muñoz, R. A single zidovudine (AZT) administration delays hepatic cell proliferation by altering oxidative state in the regenerating rat liver. Oxid. Med. Cell. Longev. 2017, 2017, 8356175. [Google Scholar] [CrossRef]
- Hernández-Muñoz, R.; Santamaría, A.; García-Sáinz, J.A.; Piña, E.; Chagoya de Sámchez, V. On the mechanism of ethanol-induced fatty liver and its reversibility by adenosine. Arch. Biochem Biophys. 1978, 190, 155–162. [Google Scholar] [CrossRef] [PubMed]
- Hernández-Muñoz, R.; Glender, W.; Díaz-Muñoz, M.; García-Sáinz, J.A.; Chagoya de Sánchez, V. Effects of adenosine on liver cell damage induced by carbon tetrachloride. Biochem. Pharmacol. 1984, 33, 2599–2604. [Google Scholar] [CrossRef]
- Hernández-Muñoz, R.; Díaz-Muñoz, M.; Suárez-Cuenca, J.A.; Trejo-Solís, C.; López, V.; Sánchez-Sevilla, L.; Yáñez, L.; De Sánchez, V.C. Adenosine reverses a preestablished CCl4-induced micronodular cirrhosis through enhancing collagenolytic activity and stimulating hepatocyte cell proliferation in rats. Hepatology 2001, 34, 677–687. [Google Scholar] [CrossRef]
- Morales-González, J.A.; Gutiérrez-Salinas, J.; Yañez, L.; Villagómez-Rico, C.; Badillo-Romero, J.; Hernández-Muñoz, R. Morphological and biochemical effects of a low ethanol dose on rat liver regeneration. Dig. Dis. Sci. 1999, 44, 1963–1974. [Google Scholar] [CrossRef] [PubMed]
- Colombo, J.P.; Konarska, L. Arginase. In Methods of Enzymatic Analysis; Bergmeyer, H.U., Bergmeyer, J., Grassl, M., Eds.; Verlag Chemie: Deerfield Beach, FL, USA, 1984; Volume VII, pp. 285–294. [Google Scholar]
- Hernández-Muñoz, R.; Díaz-Muñoz, M.; de Sánchez, V.C. Possible role of cell redox state on collagen metabolism in carbon tetrachloride-induced cirrhosis as evidenced by adenosine administration to rats. Biochim. Biophys. Acta. 1994, 1200, 93–99. [Google Scholar] [CrossRef] [PubMed]
- Hernández-Muñoz, R.; Contreras-Zentella, M.L. Involvement of cell oxidant status and redox state in the increased non-enzymatic ethanol oxidation by the regenerating rat liver. Biochem. Pharmacol. 2019, 161, 63–72. [Google Scholar] [CrossRef] [PubMed]
- Martiñón-Gutiérrez, G.; Luna-Castro, M.; Hernández-Muñoz, R. Role of insulin/glucagon ratio and cell redox state in the hyperglycemia induced by exposure to a 60-Hz magnetic field in rats. Sci. Rep. 2021, 11, 11666. [Google Scholar] [CrossRef] [PubMed]
- Bakasis, A.D.; Androutsakos, T. Liver fibrosis during antiretroviral treatment in HIV-infected individuals. Truth or tale? Cells 2021, 10, 1212. [Google Scholar] [CrossRef] [PubMed]
- Majid, S.; Khanduja, K.L.; Gandhi, R.K.; Kapur, S.; Sharma, R.R. Influence of ellagic acid on antioxidant defense system and lipid peroxidation in mice. Biochem. Pharmacol. 1991, 42, 1441–1445. [Google Scholar] [CrossRef]
- de la Asunción, J.G.; Del Olmo, M.L.; Gómez-Cambronero, G.; Sastre, J.; Pallardó, F.V.; Viña, J. AZT induces oxidative damage to cardiac mitochondria: Protective effect of vitamins C and E. Life Sci. 2004, 76, 47–56. [Google Scholar] [CrossRef] [PubMed]
- Tikoom, K.; Tamta, A.; Ali, I.Y.; Gupta, J.; Gaikwad, A.B. Tannic acid prevents azidothymidine (AZT) induced hepatotoxicity and genotoxicity along with change in expression of PARG and histone H3 acetylation. Toxicol. Lett. 2008, 177, 90–96. [Google Scholar] [CrossRef] [PubMed]
- Tadesse, B.T.; Foster, B.A.; Kabeta, A.; Ayalew, F.; H/Meskel, G.; Jerene, D.; Makonnen, E.; Aklillu, E. Hepatic and renal toxicity and associated factors among HIV-infected children on antiretroviral therapy: A prospective cohort study. HIV Med. 2019, 20, 147–156. [Google Scholar] [CrossRef]
- Acosta, B.S.; Grimsley, E.W. Zidovudine-associated type B lactic acidosis and hepatic steatosis in an HIV-infected patient. South Med. J. 1999, 92, 421–423. [Google Scholar] [CrossRef]
- Schulte-Hermann, K.; Schalk, H.; Haider, B.; Hutterer, J.; Gmeinhart, B.; Pichler, K.; Brath, H.; Dorner, T.E. Impaired lipid profile and insulin resistance in a cohort of Austrian HIV patients. J. Infect. Chemother. 2016, 22, 248–253. [Google Scholar] [CrossRef]
- Ikemoto, M.; Tsunekawa, S.; Toda, Y.; Totani, M. Liver-type arginase is a highly sensitive marker for hepatocellular damage in rats. Clin. Chem. 2001, 47, 946–948. [Google Scholar] [CrossRef] [PubMed]
- Díaz-Muñoz, M.; Hernández-Muñoz, R. Molecular and biochemical features of the mitochondrial enzyme ornithine transcarbamylase: A possible new role as a signaling factor. Curr. Med. Chem. 2010, 17, 2253–2260. [Google Scholar] [CrossRef] [PubMed]
- Silva, J.P.; Köhler, M.; Graff, C.; Oldfors, A.; Magnuson, M.A.; Berggren, P.O.; Larsson, N.G. Impaired insulin secretion and beta-cell loss in tissuespecific knockout mice with mitochondrial diabetes. Nat. Genet. 2000, 26, 336–340. [Google Scholar] [CrossRef]
- Begriche, K.; Igoudjil, A.; Pessayre, D.; Fromenty, B. Mitochondrial dysfunction in NASH: Causes, consequences and possible means to prevent it. Mitochondrion 2006, 6, 1–28. [Google Scholar] [CrossRef] [PubMed]
- Aukrust, P.; Müller, F.; Svardal, A.M.; Ueland, T.; Berge, R.K.; Frøland, S.S. Disturbed glutathione metabolism and decreased levels in human immunodeficiency virus-infected patients during highly active antiretroviral therapy—potential immuno-modulatory effects of antioxidants. J. Infect. Dis. 2003, 188, 232–238. [Google Scholar] [CrossRef] [PubMed]
- Mondal, D.; Pradhan, L.; Ali, M.; Agrawal, K.C. HAART drugs induce oxidative stress in human endothelial cells and increase endothelial recruitment of mononuclear cells: Exacerbation by inflammatory cytokines and amelioration by antioxidants. Cardiovasc. Toxicol. 2004, 4, 287–302. [Google Scholar] [CrossRef]
- Pérez-Cabeza de Vaca, R.; Domínguez-López, M.; Guerrero-Celis, N.; Rodríguez-Aguilera, J.R.; Chagoya de Sánchez, V. Inflammation is regulated by the adenosine derivative molecule, IFC-305, during reversion of cirrhosis in a CCl4 rat model. Int. Immunopharmacol. 2018, 5, 12–23. [Google Scholar] [CrossRef] [PubMed]
- Miller, K.D.; Cameron, M.; Wood, L.V.; Dalakas, M.C.; Kovacs, J.A. Lactic acidosis and hepatic steatosis associated with use of stavudine: Report of four cases. Ann. Intern. Med. 2000, 1333, 192–196. [Google Scholar] [CrossRef]
- Casademont, J.; Barrientos, A.; Grau, J.M.; Pedrol, E.; Estivill, X.; Urbano-Márquez, A.; Nunes, V. The effect of zidovudine on skeletal muscle mtDNA in HIV-1 infected patients with mild or no muscle dysfunction. Brain. 1996, 119, 1357–1364. [Google Scholar] [CrossRef]
- Vickers, A.E.; Bentley, P.; Fisher, R.L. Consequences of mitochondrial injury induced by pharmaceutical fatty acid oxidation inhibitors is characterized in human and rat liver slices. Toxicol. In Vitro. 2006, 20, 1173–1182. [Google Scholar] [CrossRef]
- Pan-Zhou, X.R.; Cui, L.; Zhou, X.J.; Sommadossi, J.P.; Darley-Usmar, V.M. Differential effects of antiretroviral nucleoside analogs on mitochondrial function in HepG2 cells. Antimicrob. Agents Chemother. 2000, 44, 496–503. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Effects of AZT and ADO on serum activities of marker enzymes for liver and pancreas integrity. Results represent the mean ± SEM of four individual observations per experimental group for serum activities of alanine aminotransferase (ALT; panel A), aspartate aminotransferase (AST; panel B), lactate dehydrogenase (LDH; panel C), γ-glutamyl transferase (γ-GT; panel D), ornithine transcarbamylase (OCT; panel E), and arginase (panel F), all expressed as IU • L-1. Additionally, the pancreatic enzymes amylase (panel G) and lipase (panel H) are expressed in the same units. Control values are represented by the horizontal gray bar. The symbols and colors represent the experimental groups at the top of panel A. Statistical significance: *p < 0.01 compared to the control group, and **p < 0.01 vs. the group receiving AZT alone.
Figure 1.
Effects of AZT and ADO on serum activities of marker enzymes for liver and pancreas integrity. Results represent the mean ± SEM of four individual observations per experimental group for serum activities of alanine aminotransferase (ALT; panel A), aspartate aminotransferase (AST; panel B), lactate dehydrogenase (LDH; panel C), γ-glutamyl transferase (γ-GT; panel D), ornithine transcarbamylase (OCT; panel E), and arginase (panel F), all expressed as IU • L-1. Additionally, the pancreatic enzymes amylase (panel G) and lipase (panel H) are expressed in the same units. Control values are represented by the horizontal gray bar. The symbols and colors represent the experimental groups at the top of panel A. Statistical significance: *p < 0.01 compared to the control group, and **p < 0.01 vs. the group receiving AZT alone.
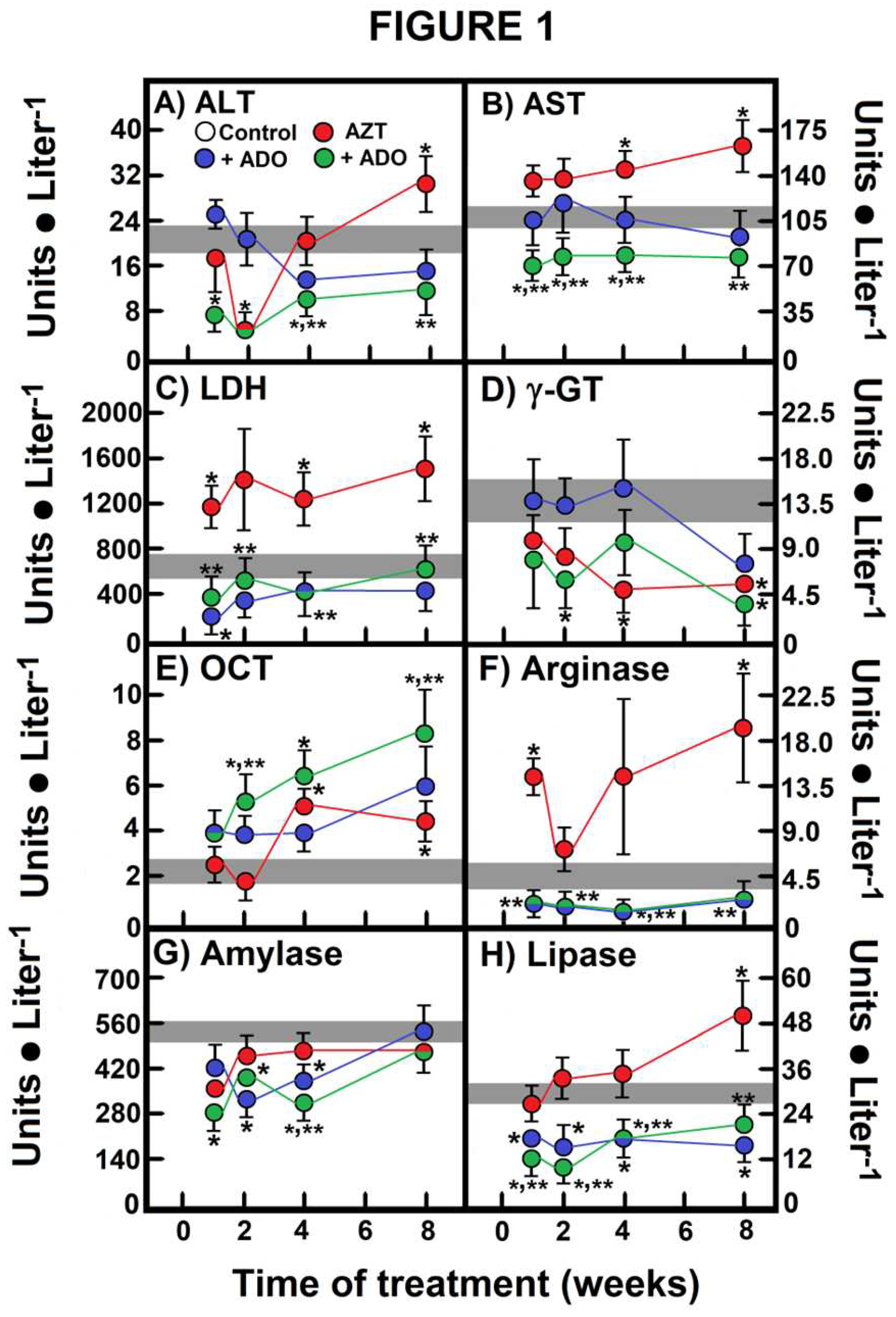
Figure 2.
Effects of AZT and ADO on serum activities of marker enzymes for liver and pancreas integrity. Results represent the mean ± SEM of four individual observations per experimental group for insulin (panel A) and glucagon (panel B), expressed as pmoles • ml-1. In panel C, the insulin/glucagon ratio is depicted. Control values are represented by the horizontal gray bar.The symbols and colors indicate the experimental groups at the top of panel A. Statistical significance: *p < 0.01 compared to the control group, and **p < 0.01 vs. the group receiving AZT alone.
Figure 2.
Effects of AZT and ADO on serum activities of marker enzymes for liver and pancreas integrity. Results represent the mean ± SEM of four individual observations per experimental group for insulin (panel A) and glucagon (panel B), expressed as pmoles • ml-1. In panel C, the insulin/glucagon ratio is depicted. Control values are represented by the horizontal gray bar.The symbols and colors indicate the experimental groups at the top of panel A. Statistical significance: *p < 0.01 compared to the control group, and **p < 0.01 vs. the group receiving AZT alone.
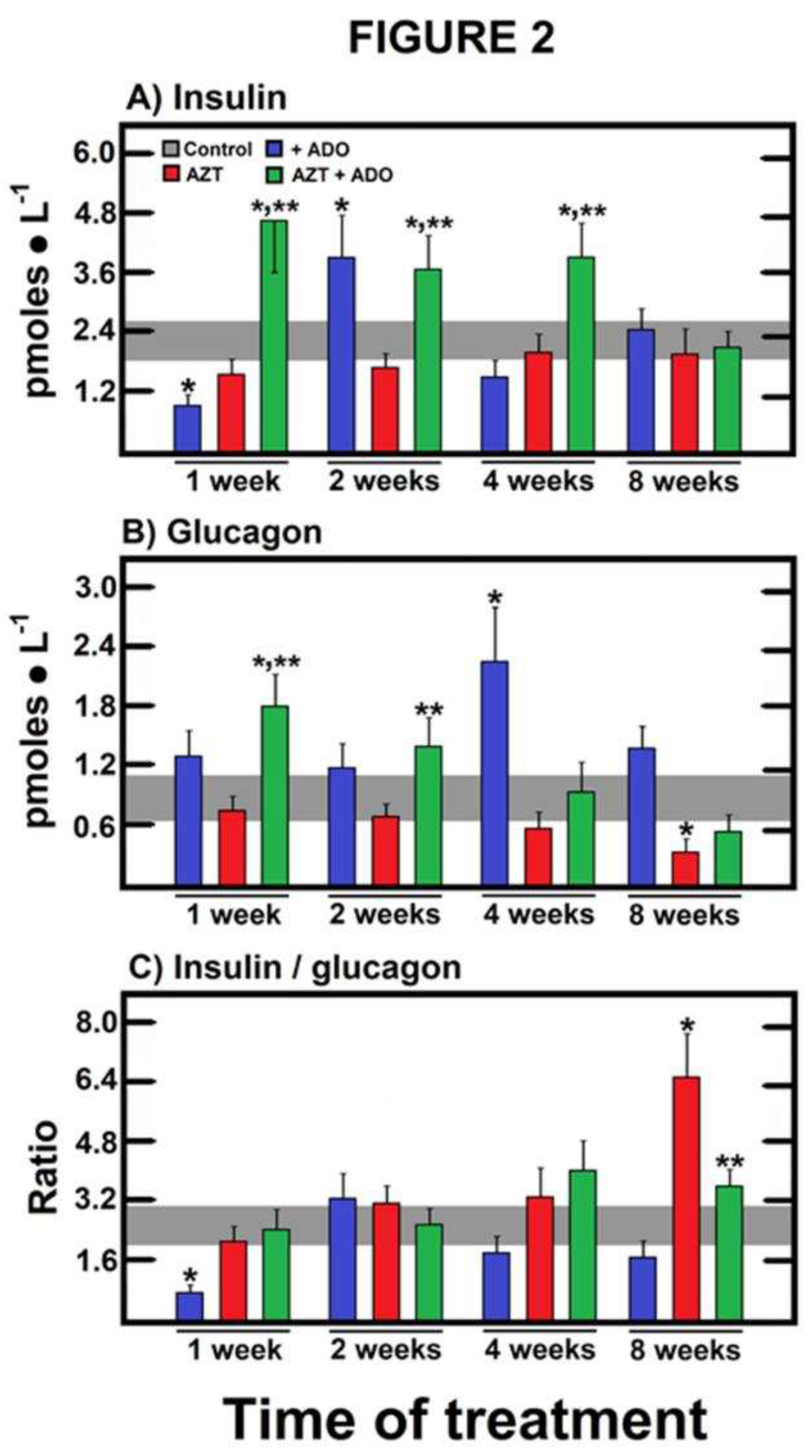
Figure 3.
Histological survey of livers from rats treated with zidovudine (AZT) and adenosine (ADO). Representative micrographs of the four experimental groups: A) control, B) ADO, C) AZT, and D) AZT + ADO. For histological analysis, four biological samples were used per treatment, and five sections or fields were examined using a 40X objective and photographically registered from each histological sample employing the zig-zag methodology. The table at the bottom displays the average of the histological findings, graded according to the scoring system described at the bottom of the table.
Figure 3.
Histological survey of livers from rats treated with zidovudine (AZT) and adenosine (ADO). Representative micrographs of the four experimental groups: A) control, B) ADO, C) AZT, and D) AZT + ADO. For histological analysis, four biological samples were used per treatment, and five sections or fields were examined using a 40X objective and photographically registered from each histological sample employing the zig-zag methodology. The table at the bottom displays the average of the histological findings, graded according to the scoring system described at the bottom of the table.
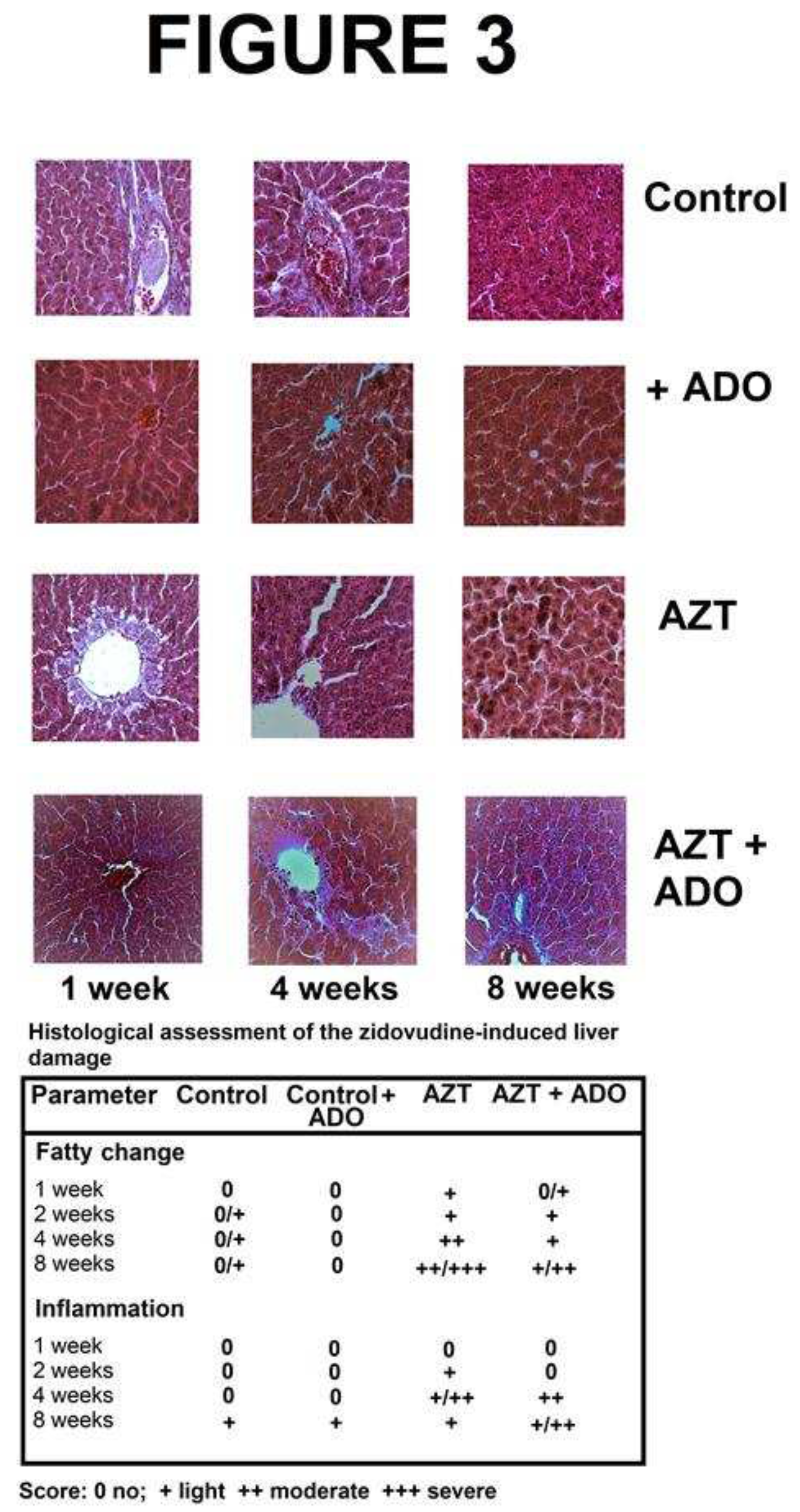
Figure 4.
Effects of zidovudine (AZT) and adenosine (ADO) on serum concentration of cytokines involved in the inflammatory process. Results represent the mean ± SEM of four individual observations per experimental group for interleukin-1β (IL-1β, panel A), interleukin-6 (IL-6, panel B), interleukin-4 (IL-4, panel C), and interleukin-10 (IL-10, panel D). Control values are represented by the horizontal gray bar.Symbols and colors for the experimental groups are indicated at the top of panel A. Statistical significance: *p < 0.01 against the control group, and **p < 0.01 vs. the group of AZT alone.
Figure 4.
Effects of zidovudine (AZT) and adenosine (ADO) on serum concentration of cytokines involved in the inflammatory process. Results represent the mean ± SEM of four individual observations per experimental group for interleukin-1β (IL-1β, panel A), interleukin-6 (IL-6, panel B), interleukin-4 (IL-4, panel C), and interleukin-10 (IL-10, panel D). Control values are represented by the horizontal gray bar.Symbols and colors for the experimental groups are indicated at the top of panel A. Statistical significance: *p < 0.01 against the control group, and **p < 0.01 vs. the group of AZT alone.
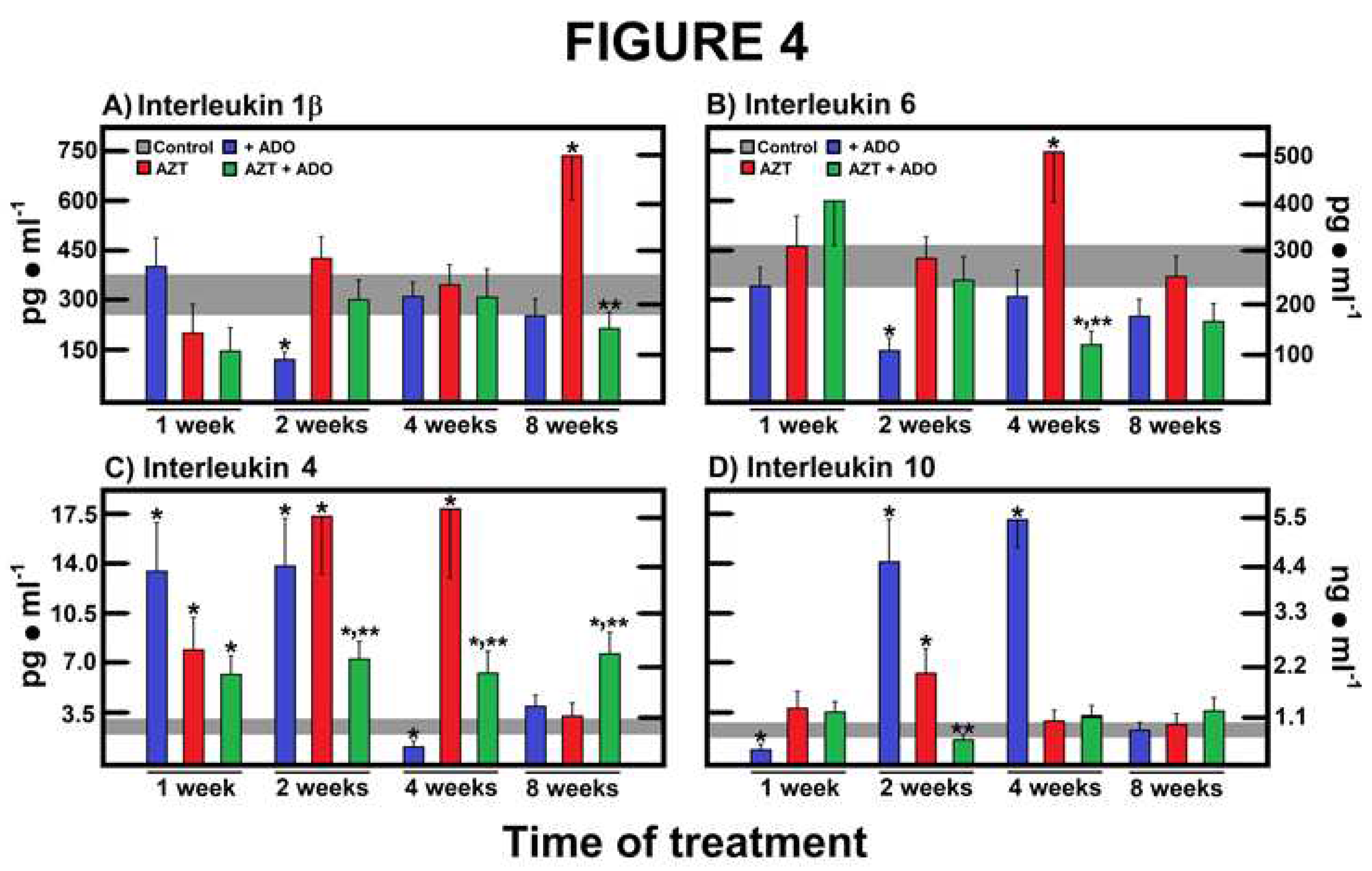
Figure 5.
Effects of zidovudine (AZT) and adenosine (ADO) on the cytoplasmic liver NAD/NADH redox state and their possible reflection in serum. Results are the mean ± SEM of four individual observations per experimental group for serum and liver lactate levels (panels A), serum and liver pyruvate levels (panels B), and serum and liver NAD/NADHcyto ratio (panels C). Control values are represented by the horizontal gray bar.Symbols and colors for the experimental group are indicated at the top of panel A. Statistical significance: *p < 0.01 against the control group, and **p < 0.01 vs. the group treated with AZT alone.
Figure 5.
Effects of zidovudine (AZT) and adenosine (ADO) on the cytoplasmic liver NAD/NADH redox state and their possible reflection in serum. Results are the mean ± SEM of four individual observations per experimental group for serum and liver lactate levels (panels A), serum and liver pyruvate levels (panels B), and serum and liver NAD/NADHcyto ratio (panels C). Control values are represented by the horizontal gray bar.Symbols and colors for the experimental group are indicated at the top of panel A. Statistical significance: *p < 0.01 against the control group, and **p < 0.01 vs. the group treated with AZT alone.
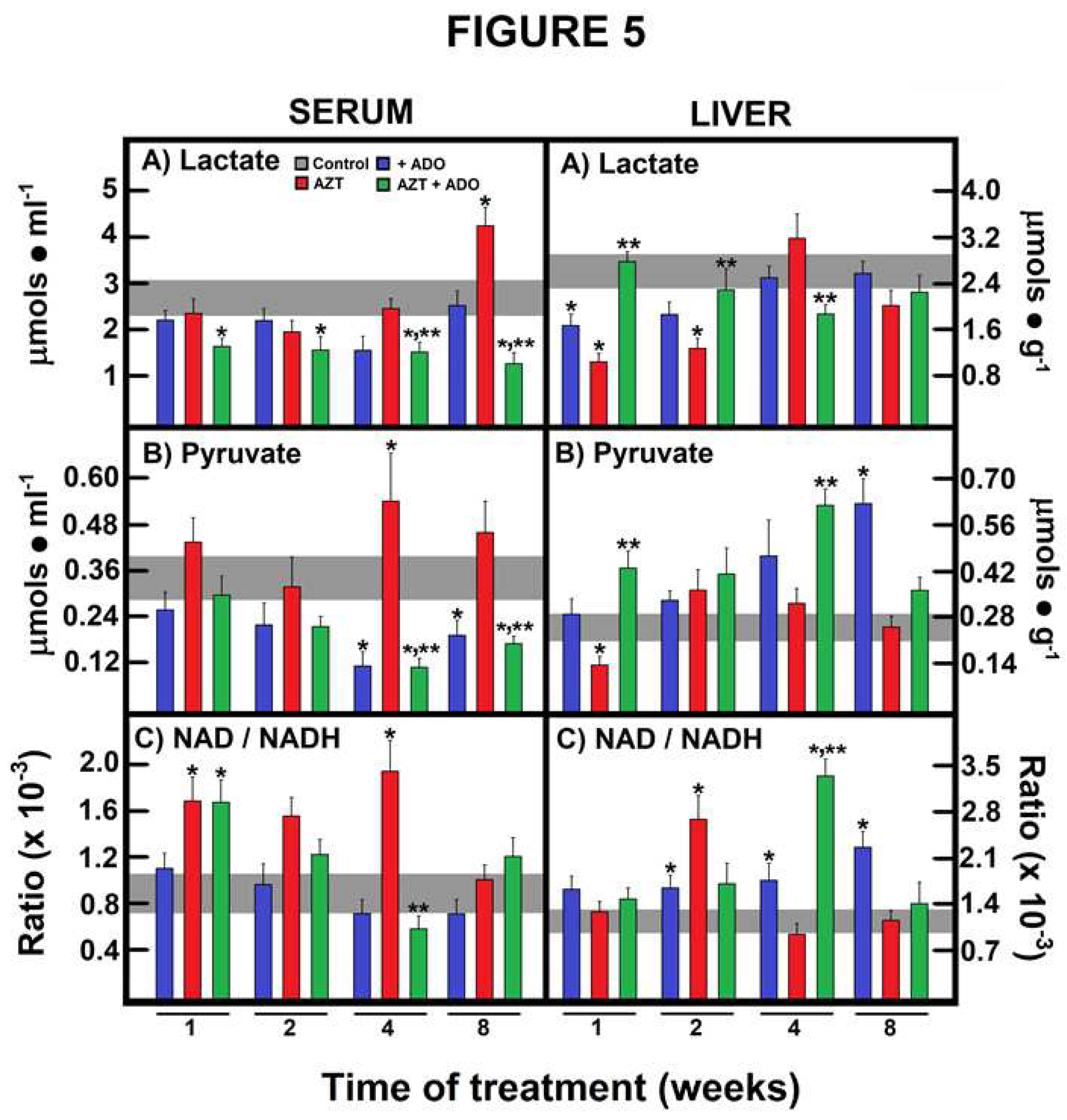
Figure 6.
Effects of zidovudine (AZT) and adenosine (ADO) on mitochondrial liver NAD/NADH redox state and its possible reflection in serum. Results are the mean ± SEM of four individual observations per experimental group for serum and liver β-OH butyrate levels (panels A), serum and liver acetoacetate levels (panels B), and serum and liver NAD/NADHmito ratio (panels C). Control values are represented by the horizontal gray bar.Symbols and colors for the experimental group are indicated at the top of panel A. Statistical significance: *p < 0.01 against the control group, and **p < 0.01 vs. the group treated with AZT alone.
Figure 6.
Effects of zidovudine (AZT) and adenosine (ADO) on mitochondrial liver NAD/NADH redox state and its possible reflection in serum. Results are the mean ± SEM of four individual observations per experimental group for serum and liver β-OH butyrate levels (panels A), serum and liver acetoacetate levels (panels B), and serum and liver NAD/NADHmito ratio (panels C). Control values are represented by the horizontal gray bar.Symbols and colors for the experimental group are indicated at the top of panel A. Statistical significance: *p < 0.01 against the control group, and **p < 0.01 vs. the group treated with AZT alone.
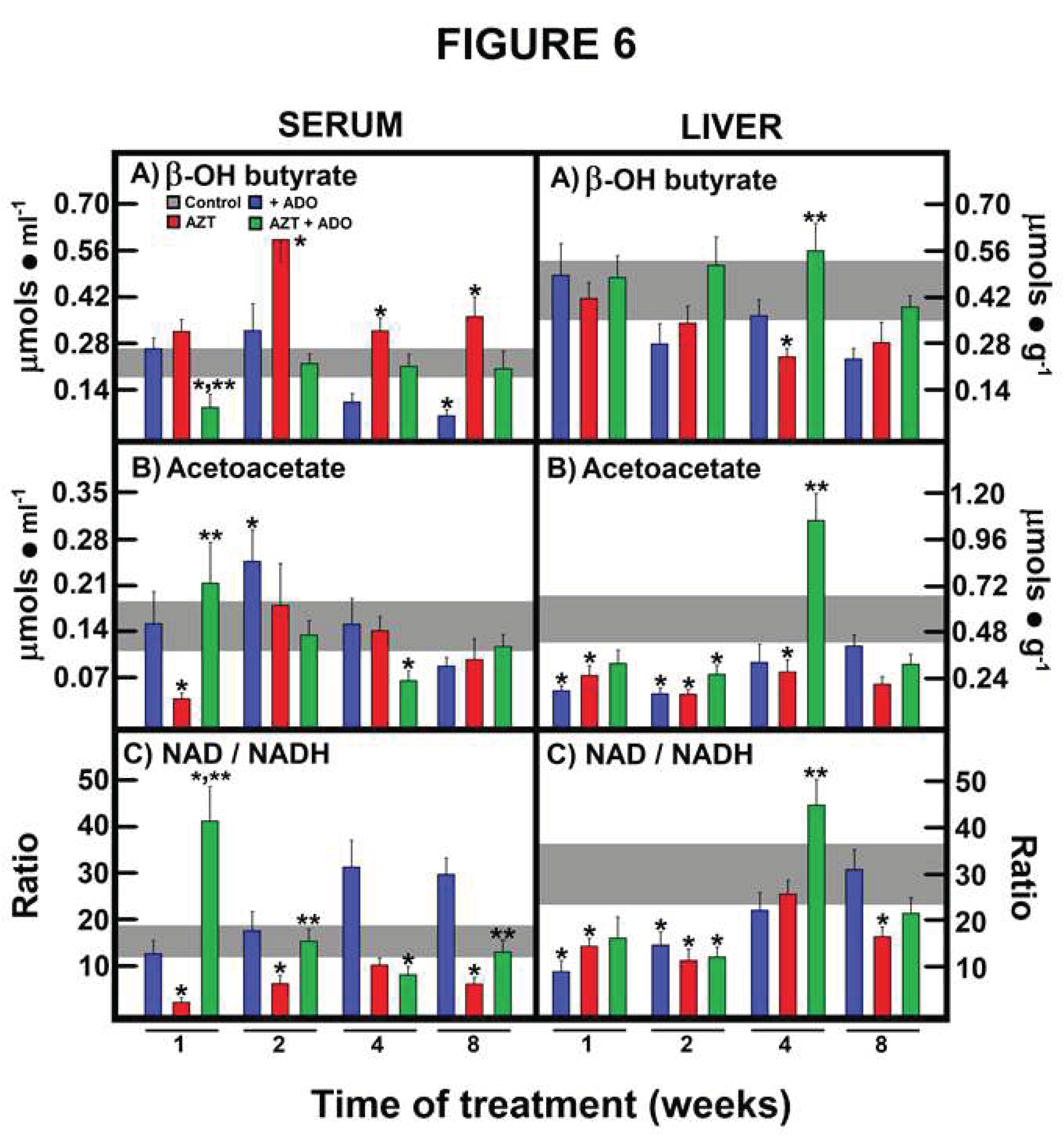

Table 1.
Clinical parameters determined in sera from rats treated with zidovudine (AZT) and adenosine (ADO).
Table 1.
Clinical parameters determined in sera from rats treated with zidovudine (AZT) and adenosine (ADO).
| Parameter (Time) | Controls | Controls ADO | AZT | AZT + ADO |
|---|---|---|---|---|
| Glucose | ||||
| 1 week | 88 ± 5 | 84 ± 7 | 80 ± 12 | 138 ± 10*,** |
| 2 weeks | 100 ± 6 | 90 ± 7 | 67 ± 6* | 137 ± 12*,** |
| 4 weeks | 107 ± 7 | 82 ± 6* | 101 ± 4 | 133 ± 11** |
| 8 weeks | 118 ± 9# | 90 ± 5* | 67 ± 3* | 125 ± 10** |
| TG | ||||
| 1 week | 57 ± 5 | 50 ± 8 | 50 ± 6 | 51 ± 5 |
| 2 weeks | 66 ± 6 | 38 ± 6* | 40 ± 7* | 40 ± 4* |
| 4 weeks | 50 ± 4 | 31 ± 3* | 34 ± 3* | 45 ± 7 |
| 8 weeks | 61 ± 7 | 35 ± 4* | 35 ± 6* | 33 ± 5* |
| Cholesterol | ||||
| 1 week | 58 ± 6 | 58 ± 9 | 47 ± 6 | 53 ± 5 |
| 2 weeks | 63 ± 12 | 47 ± 3 | 54 ± 4 | 42 ± 4* |
| 4 weeks | 60 ± 11 | 48 ± 3 | 48 ± 3 | 47 ± 7 |
| 8 weeks | 54 ± 5 | 49 ± 4 | 41 ± 6 | 35 ± 5* |
| Albumin | ||||
| 1 week | 3.1 ± 0.1 | 2.7 ± 0.2 | 2.7 ± 0.1* | 2.7 ± 0.2 |
| 2 weeks | 3.1 ± 0.2 | 2.7 ± 0.3 | 2.6 ± 0.1 | 2.7 ± 0.2 |
| 4 weeks | 3.7 ± 0.3 | 2.9 ± 0.4 | 2.9 ± 0.2 | 2.6 ± 0.1* |
| 8 weeks | 2.8 ± 0.3 | 2.8 ± 0.3 | 2.7 ± 0.1 | 3.3 ± 0.3 |
| Bilirubine (T) | ||||
| 1 week | 0.37± 0.11 | 0.38 ± 0.06 | 0.22 ± 0.04 | 0.22 ± 0.04 |
| 2 weeks | 0.19 ± 0.03 | 0.14 ± 0.02 | 0.66 ± 0.18* | 0.22 ± 0.04 |
| 4 weeks | 0.44 ± 0.08 | 0.20 ± 0.03* | 0.23 ± 0.04 | 0.31 ± 0.05 |
| 8 weeks | 0.29 ± 0.04 | 0.29 ± 0.05 | 0.14 ± 0.03* | 0.44 ± 0.07 |
| Bilirubine (D) | ||||
| 1 week | 0.05 ± 0.01 | 0.06 ± 0.06 | 0.04 ± 0.01 | 0.14 ± 0.03*,** |
| 2 weeks | 0.09 ± 0.02 | 0.02 ± 0.02* | 0.07 ± 0.01 | 0.07 ± 0.02** |
| 4 weeks | 0.10 ± 0.02 | 0.08 ± 0.03 | 0.05 ± 0.02 | 0.04 ± 0.01* |
| 8 weeks | 0.08 ± 0.02 | 0.05 ± 0.01 | 0.04 ± 0.01 | 0.03 ± 0.01 |
| Urea | ||||
| 1 week | 45 ± 3 | 42 ± 3 | 39 ± 5 | 42 ± 3 |
| 2 weeks | 47 ± 4 | 50 ± 5 | 44 ± 5 | 46 ± 4 |
| 4 weeks | 40 ± 4 | 44 ± 5 | 57 ± 5* | 49 ± 5 |
| 8 weeks | 54 ± 6 | 49 ± 6 | 45 ± 4 | 50 ± 6 |
| Creatinine | ||||
| 1 week | 0.55 ± 0.04 | 42 ± 3 | 0.53 ± 0.07 | 0.57 ± 0.5 |
| 2 weeks | 0.53 ± 05 | 50 ± 5 | 0.79 ± 0.09* | 0.66 ± 0.06 |
| 4 weeks | 0.64 ± 0.08 | 44 ± 5 | 0.86 ± 0.09 | 0.75 ± 0.08 |
| 8 weeks | 0.77 ± 0.11 | 49 ± 6 | 0.92 ± 0.07 | 0.85 ± 0.10 |
The 'clinical parameters' represent the mean ± SE of 4 independent observations per experimental point. All metabolites are expressed as mg • dL-1, except for albumin, which is expressed as g • dL-1. Abbreviations: (T) = Total and (D) = Direct. Statistical significance: *p < 0.01 compared to controls, **p < 0.01 compared to the AZT group, and #p < 0.05 compared to the control group (1 week of treatment).
Table 2.
Serum and liver levels of total ketone bodies in rats treated with zidovudine (AZT) and adenosine (ADO).
Table 2.
Serum and liver levels of total ketone bodies in rats treated with zidovudine (AZT) and adenosine (ADO).
| Parameter (Time) | Controls | Controls ADO | AZT | AZT + ADO |
|---|---|---|---|---|
| Ketone bodies | ||||
| Serum | ||||
| 1 week | 0.49 ± 0.07 | 0.42 ± 0.08 | 0.36 ± 0.06 | 0.33 ± 0.08 |
| 2 weeks | 0.34 ± 0.05 | 0.56 ± 0.12 | 0.77 ± 0.16* | 0.35 ± 0.05** |
| 4 weeks | 0.31 ± 0.04 | 0.26 ± 0.05 | 0.49 ± 0.05* | 0.30 ± 0.04** |
| 8 weeks | 0.35 ± 0.05 | 0.16 ± 0.02* | 0.46 ± 0.04 | 0.33 ± 0.05 |
| Liver | ||||
| 1 week | 0.91 ± 0.14 | 0.66 ± 0.08 | 0.68 ± 0.11 | 0.79 ± 0.17 |
| 2 weeks | 1.03 ± 0.15 | 0.45 ± 0.06* | 0.52 ± 0.10* | 0.77 ± 0.11 |
| 4 weeks | 1.24 ± 0.20 | 0.69 ± 0.08* | 0.52 ± 0.06* | 0.30 ± 0.04*,** |
| 8 weeks | 0.71 ± 0.15 | 0.56 ± 0.07 | 0.48 ± 0.04 | 0.60 ± 0.09 |
The total ketone bodies are the mean ± SE of 4 independent observations per experimental point as the sum of β-OH-butyrate and acetoacetate and expressed as nmoles ∙ ml-1 of serum or as nmoles ∙ g-1 of liver. Statistical significance: *p< 0.01 against controls, **p< 0.01 vs. the AZT group.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



