Submitted:
23 December 2023
Posted:
25 December 2023
You are already at the latest version
Abstract
Abstract: Antimicrobial peptides (AMPs) are short peptides, usually made up of less than 100 amino acid residues. They are found in many living organisms and are an important factor in their innate immune systems. AMPs can be extracted from various living sources including bacteria, plants, animals and even humans. They are usually cationic peptides with an amphiphilic structure, which allows them to easily bind and interact with the cellular membranes of viruses, bacteria, fungi and other pathogens. They can act against both Gram-negative and Gram-positive pathogens and have various modes of action against them. Some attack the pathogens membrane, while others target their intracellular organelles as well as their nucleic acids, proteins and metabolic pathways. A crucial area of use of AMPs is in their ability to help with emerging antibiotic resistance: some AMPs are active against resistant strains and are susceptible to peptide engineering. This review considers AMPs from three key sources: plants, animals and humans, as well as their modes of action and some AMPs sequences.
Keywords:
Plant AMPs
; animal MPs
; human AMPs
; mechnism of action
; AMP sequnces
1. Introduction
Antimicrobial peptides are peptides, although varying in size, not usually longer than 100 amino acid residues [1]. They are an important part of the defense and the innate immune system of many living organisms such as bacteria, fungi, plants, fish, invertebrates, amphibians, crustaceans, insects, reptiles, mammals, humans etc. [1]. The review will focus on three in particular: plants, animals and humans and their respective modes of action.
AMPs are positively charged and have both hydrophilic and lipophilic properties, characterizing them as amphiphilic [2]. Their amphiphilic structure and cationic charge allow them to bind to a negatively-charged pathogen cell surface and insert themselves into the membrane, forming pores and channels, which eventually leads to cell death [2]. AMPs can be by their biosynthetic origin ribosomal or enzymatic (NRPS pathway). Ribosomally synthesized AMPs are divided into unmodified ribosomally synthesized peptides and ribosomally synthesized and posttranslationally modified peptides (RiPPs). RiPPs are being considered as potential alternatives of conventionally used antibiotics. Among these RiPPs are sactipeptides, which are a small subfamily of peptides. Sactipeptides (Sulfur-to-alpha carbon thioether cross-linked peptides) show various biological activities, such as antibacterial and hemolytic properties [3]. Nonribosomal peptide synthetases (NRPS) are large multimodular enzymes that synthesize a diverse variety of peptides. Many of these are currently used as pharmaceuticals, thanks to their antimicrobial activities (penicillin, vancomycin, daptomycin, echinocandin), immunosuppressant (cyclosporin) and anticancer compounds (bleomycin) [4].
AMPs use several modes of action when acting on pathogens, some of which target the membrane and others that focus on intracellular targets such as nucleic acids, protein synthesis, and repair pathways [5]. The antimicrobial peptides have a broad spectrum of action and are active against both Gram-positive and Gram-negative bacteria, viruses, fungi, and other pathogens [6]. They have been shown to be an effective substitute for antibiotics as they can act against antibiotic resistant strains and are naturally found in many living organisms, thus easy to access. In Kumar et al.’s review it is discussed how antimicrobial peptides can be chemically modified in order to improve their stability against proteolytic digestion while also retaining their strong antibacterial activity [7].
To date there are multiple antimicrobial peptides isolated from nature and because of their diversity they can also be classified in multiple categories based on their size, structure, modes of action etc. [7]. AMPs also differ in their activities; some can be active against bacteria – Gram-negative, Gram-positive or both – while others can be active against fungi or viruses. That said, AMPs can also possess some combination of antibacterial, antifungal and antiviral activities, or all of these activities at once. [7]. Based on their structure, AMPs can be divided into three subclasses [7,8], (Figure 1). The first one is AMPs with an alpha helical structure, mainly found in animals, specifically frogs and insects [7,8]. Some of the members of this subclass are cathelicidins, including LL-37 (also found in humans), and also magainins and aurein peptides. The second subclass are β-sheet AMPs, which include some members of the cathelicidin class, such as protegrins, and other classes like defensins and tachyplesins [7,8]. The third subclass of AMPs are known as having an extended coil structure The members of this subclass consist of mostly cathelicidins (indolicidin) and histatins [7,8]. Of note then, cathelicidins can be found across all three of these subclasses.
Cryptic peptides (cryptides) are small bioactive molecules obtained via the degradation of functionally active proteins [9,10,11]. Fesenko et al. describe that the winter moss Phycomitrella patens produces over 4 000 intracellular proteins, about 500 of them being secreted [10]. The moss was grown in stress conditions derived from adding the stress hormone methyl jasmonate. As a result, specific proteolysis was induced leading to the release of peptides with antimicrobial activities. Especially, the enzymes are considered as a reservoir of such cryptic peptides [12]. In their review, the authors describe host defense peptides (HDPs) in all canonical classes of enzymes and corresponding hydrolases. A promising anti-biofilm and immunomodulatory activity of human HDPGVF27 on two clinically relevant strains Burkholderia multivorans and Burkholderia cenocepacia were reported by Bosso et al. [13]. Burkholderia cenocepacia ia an opportunistic pathogen that commonly infect immunocompromised patients. Another study by Ciociola et al. [14] discussed the antifungal activity in vitro and in vivo of peptide derived from the C-terminus of albumin. This 13-residue peptide can penetrate and accumulate in Candida albicans cells causing gross morphological alterations in the fungus cellular structure. Novel cryptic peptides from PD-L1/2, a type 1 ribosome-inactivating protein (RIP) were reported in the study of Pizzo et al. [15]. The bioactive peptides were found in Phytolacca dioica L. The authors describe biofilm biomass, thickness, and structural components reduction in Gram-negative bacteria.
In this review we describe in details three key sources of antimicrobial peptides. We discuss the sources of AMPs in plants, their structures and activity as they are a crucial source of defense in plants against various pathogens, including viruses, bacteria and fungi [7,8]. Antimicrobial peptides are of particular importance in plants, as mentioned in Kumar et al.’s review, one of the reasons being their lack of adaptive immunity, meaning B-cell and T-cell mediated immunity [7].
The animal kingdom is also an important source of AMPs, and over the course of the review we cover the main classes of AMPs found in the different groups of animals. Vertebrate animals, contrary to plants, have at their disposal both an innate and adaptive immune system [7]. Nevertheless, some AMPs have been shown to have a critical role in their immune modulation and reduction of inflammation [7,16].
Although humans are mammals, the human body doesn’t produce the same amount of AMPs as other animals. This is discussed in the review by Li et al.[1], where it is mentioned that the only AMP from the cathelicidin family present in the human body is LL-37, as well as the inability of the human organism to express θ-defensins [1]. In this review we focus on human antimicrobial peptides, where they are produced and their activity against pathogens.
The last section of the review is dedicated to the different modes of action that antimicrobial peptides use to target and eliminate viruses, bacteria and fungi, the main ones being membrane targeting methods or focusing on intracellular targets.
2. AMPs from Plants
Antimicrobial peptides are very common sources of protection and defense in plants against pathogens, and are seen in all major organs of the plant, including seeds and flowers [1,17]. They are largely made up of cysteine and multiple disulfide bridges, which makes them compact and durable against high temperatures and chemicals [6,17]. Plant AMPs are divided into groups, based on their structures, which include thionins, defensins, hevein-like peptides, knottin-type peptides, α-hairpinins, lipid transfer proteins, snakins and non-cysteine rich peptides. These groups differ mainly in their cysteine content and disulfide bridge arrangement [6,18].
2.1. Groups of Plant AMPs.
Thionins are cationic peptides, found also in plants, with a length of 45 to 48 amino acids and contain 3 to 4 disulfide bridges [19]. Their activities are primarily antibacterial and antifungal and have an effect against both Gram-positive and Gram-negative bacteria [6,19].
Plant defensins are also cationic peptides with a similar length of 45 to 54 amino acids and contain 4 to 5 disulfide bridges [6,20], (Figure 2). They are non-toxic to plant and mammalian cells and thus are of interest for their potential use as medicinal agents. Similar to thionins, they have both antibacterial and antifungal properties and some have even been discovered to have anticancer activities [20].
Hevein-like peptides are cationic peptides and can contain from 29 to 45 amino acid residues [6,22]. They contain 3 to 5 disulfide bridges and are rich in glycine and aromatic residues [6,22]. Their antifungal activities are due to a chitin-binding domain which can damage the fungal cell wall, thus they have a wide range of antifungal activities [18,22].
The next class of plant AMPs, knottins, contain 30 residues, including cysteine residues and disulfide bonds [6,22]. Knottins possess a wide range of functions, some of which being hormone-like functions, inhibiting enzyme activity, cytotoxicity and anti-HIV [22]. They also have both antiviral and antibacterial activity but are toxic to humans due to their unselective contact with cell membranes [6,18,22].
α-Hairpinins (stable-like peptides) are rich in lysine and arginine class and form a helix-loop-helix structure [6,18,23]. They have both antibacterial and antifungal activities, they bind to DNA and inhibit RNA and protein synthesis by inhibiting trypsin and inactivating ribosomes activity [18,23].
Lipid transfer proteins (LTPs) are small, cationic, cysteine-rich proteins, made up of around 100 residues [6,18,22]. As their name suggests, they are able to transfer lipids between membranes and thus form pores and cause bacterial and fungal cell death [7]. LTPs are non-specific and can bind to a wide range of lipids such as phospholipids, fatty acids, acyl-coenzyme A etc. [22].
Snakins are described as small, cysteine-rich, cationic peptides, similar in structure to thionins and α-hairpinins [22,24]. They have antibacterial (against both Gram-negative and Gram-positive) and antifungal activities, although their modes of action are not fully understood yet [18,22,24].
The last class of AMPs are the non-cysteine rich peptides. They have 0 to 1 cysteine residues and are very structurally flexible, their activities are antibacterial (against Gram-negative and –positive bacteria) and antifungal but can also exhibit immune-stimulatory activities [6,22].
In Table 1. are presented some of best studied plant antimicrobial peptides, their source and application [25,26,27,28,29,30,31].

Table 1.
Natural AMPs expressed in plant hosts and their application and activities against pathogens [8].
Table 1.
Natural AMPs expressed in plant hosts and their application and activities against pathogens [8].
| AMP | Plant Species | Application and activity | Reference |
|---|---|---|---|
| Thi2.1 (Thionin) | Tomato (Lycopersicon esculentum) | Crop protection | [17,25] |
| Mj-AMP2 (Knottin) | Rice (Oryza sativa) | Resistance to fungal pathogens | [17,26] |
| Lipid Transfer Proteins (LTPs) | Tobacco (Nicotina tabacum) | Resistance to pathogens | [17,27] |
| Petunia Floral defensins | Banana (Musa spp.) | Effective resistance against pathogenic fungal Fusarium oxysporum | [17,28] |
| PmAMP1 (cysteine-rich protein) | Canola (Brassica napus) | Resistance against fungal pathogens (Leptosphaeria maculans) | [17,29] |
| SN-1 (Snakin) | Wheat (Triticum aestivum) | Antifungal activity in vitro and enhanced resistance to fungus (Gaeumannomyces graminis) | [17,30] |
| Pro-SmAMP2 (Heiven-like peptide) | Potato (Solanum tuberosum) | Crop protection from Alternaria sp. and Fusarium sp. | [17,31] |
As seen from the Table 1 the daily foods consumed worldwide contains antimicrobial peptides.
2.2. AMPs Present in Vegetables
In recent years there have been a lot of experiments proving the existence of AMPs in vegetables we consume daily, such as tomatoes, onions, garlic, peppers, chili peppers, which are going to be covered in the next few paragraphs. Some other examples include, LTPs extracted from leaves of spinach, barley, maize and sugar beet and from seeds of cowpea, and radish have been demonstrated to inhibit phytopathogens, such as Pseudomonas solanacearum, Clavibacter michiganensis, Fusarium solani, Rhizoctonia solani, Trichoderma viride and Cercospora beticola [32].
2.2.1. AMPs in Tomato
In Herbel et. al’s study, it is mentioned that tomato plants express two members of the snakin peptide family, named snakin-1, SN1 and snakin-2, SN2 [33]. These peptides have been described as small in molecular weight, having a cationic net charge, and six disulfide bonds [33]. Snakins are thought to act through pore formation in the biomembrane of pathogenic cells, but the exact mode of action is not yet fully understood. There is evidence that SN2 can target the phospholipid membrane of both bacteria and fungi non-specifically and causes agglomeration of the cells [33]. In Herbel et. al’s study, an untreated adult fruiting tomato plant was used to determine SN2 expression levels in a healthy plant. Results of their experiment show that the highest expression of SN2 is found in leaves and flowers of the tomato plant [33]. They also tested ripe tomato fruits and divided them into skin, pulp, and seeds. The pulp and skin showed moderate expression, whereas in the seeds, the gene expression level was not detectably higher than in the stem. These results show that every plant organ expresses a distinct amount of SN2 [33]. The authors found a higher quantity of SN2 in leaves and flowers of the tomato plant, as compared to in the shoot, seeds, or roots [33]. Importantly, what the study also conducted is that SN2 is expressed constitutively in tomato plants, even if they do not undergo a defense response [33]. In Slezina et. al’s study is described that cysteine rich peptides (CRPs) are also present in tomatoes [34]. The authors describe them to be active against plant pathogen Clavibacter michiganensis - Gram-positive bacteria, as well as the fungus Cryptococcus neoformans responsible for fungal meningitis and encephalitis in humans [34].
2.2.2. AMPs in Onion
In Taggar et. al’s study, an AMP, named as peptide-Ba49, was isolated from Bacillus subtilis subsp. spizizenii strain cultivated from Allium cepa, also known as the common onion, which exhibited strong antibacterial activity against S. aureus ATCC 25923 [35,36]. The mode of action of this peptide on S. aureus was elucidated by the authors to be through change in membrane potential and by triggering the production of reactive oxygen species (ROS) [35,36]. Furthermore, the peptide-Ba49 prevented the formation of S. aureus biofilm at low concentration and showed its potential to degrade the mature biofilm of S. aureus [35,36]. The peptide-Ba49 also exhibited intracellular killing potential against S. aureus in the macrophage cells, and was found to bolster the fibroblast cell migration in the scratch assay at low concentration, exhibiting a wound healing efficacy of this peptide [35,36].
2.2.3. AMPs in Garlic
In Ezeorba et. al’s review, three peptides were isolated from Laba garlic named F3-3-a, F3-3-b, and F3-3-c peptides with molecular weights of 693.72 Da, 737.80 Da, and 629.79 Da, respectively [37,38]. F3-3-a was identified as a pentapeptide Tyr-Asn-His-Asn-Phe (YNHNF), F3-3- b was a hexapeptide Trp-Pro-Thr-Ser-Phe-Thr (WPTSFT), and F3-3-c a hexapeptide Ala-Val-Asp-Arg-Ala-Val (AVDRAV) [37,38]. The antimicrobial activity of the three peptides against Escherichia coli, Staphylococcus aureus, Salmonella enteritidis, and Bacillus subtilis showed that F3-3-b and F3-3-c had a significant inhibitory activity on the growth of the four bacteria, especially the latter peptide [37,38]. According to the authors the difference in amino acid composition and conformation may account for the observed difference in the antimicrobial activity of the peptides [37]. The composition of hydrophobic amino acids in the peptides was 20%, 50%, and 67 % for F3-3-a, F3-3-b, and F3-3-c peptides, respectively [37]. The presence of hydrophobic amino acids, Val and Ala, and basic amino acid Arg in F3-3-c, which had the highest antimicrobial activity, corroborates with previous findings that Val/Arg residues enhance the antibacterial activity of peptides [37]. The bactericidal action of F3-3-c was demonstrated to be by causing physical damage to the bacteria cell membrane, thereby initiating leakage of cellular content [37].
The same review mentions a new AMP isolated from garlic, AsR416, with a molecular weight of 3799.52 Da, which was found to contain cysteine disulfide bonds, α-helix and β-sheet structures, 1-aspartine, L-histidine, n-acetyl-D-glucosamine 6 phosphates, N1-acetyl spermidine, analine, and L-arogenate [38]. AsR416 demonstrated antibacterial activity against Gram-negative bacteria, including phytopatogens Agrobacterium tumefaciens, Xanthomonas campestris pv. oryzicola, Ralstonia solanacearum, and Gram-positive bacteria such as human pathogens Bacillus anthrax, Bacillus cereus, Bacillus subtilis, and plant pathogens Clavibacter fangii, Clavibacter michiganensis [38].
Li et. al reported, a novel antifungal peptide named NpRS with nine amino acids (RSLNLLMFR) i.e., (Arg, Ser, Leu, Asn, Leu, Met, Phe and Arg) was obtained in garlic [39]. The peptide was found to significantly inhibit the growth of Candida albicans. According to the authors the mode of action of this peptide is through membrane destruction and interference of ribosome-related pathways and protein synthesis [39]. The resistance gene CDR1 for azole was found to be down-regulated and the drug resistance was hardly developed in 21 days by the serial passage study. Garlic extracts and garlic oil were also extensively verified with antifungal activity against Candida spp. Garlic oil was constituted by a number of linear sulfur-containing volatile compounds where diallyl disulfide (DDS) and diallyl trisulfide (DTS) were the most abundant volatile compounds. The authors assessed the antifungal activity of NpRS against C. albicans through minimum inhibition concentration (MIC) and the time-kill kinetics assay [39]. The MIC of NpRS against C. albicans was found to be 0.27 mM. The time-kill kinetics assay was performed with fungal strains to determine the mode of action of NpRS on the growth of C. albicans. At certain time intervals, aliquots were taken and determined by OD600. According to the results, NpRS exhibited significant fungicide activity (p < 0.05) and extended the lag phase of C. albicans at the MIC concentration [39].
2.2.4. AMPs in Chili Pepper
The Capsicum genus is known to produce at least 10 known antimicrobial peptides, including ɣ-Thionin, found in C. chinense [40]. AMPs such as LTPs and defensins were found in seeds of C. annuum [41]. Results indicated that three protein fractions of chili pepper seeds display antifungal activities against different fungi. An inhibitory effect of F1, F2 and F3 fractions on the growth of all fungi tested was noticed at concentrations of 70 and 150 µg mL-1. A notable inhibitory effect, mainly of the F3 fraction, was also observed on the growth of S. cerevisiae yeast at concentrations of 70 and 150 µg mL-1, demonstrating 70% and 100% of inhibition, respectively [41]. The authors tested the F3 fraction against the yeasts S. cerevisiae, C. guilliermondii, C. parapsilosis, K. marxiannus, P. membranifaciens, C. tropicalis and C. albicans. The IC50 value for C. albicans, C. guilliermondii, K. marxiannus and P. membranifaciens for example, can be observed at a concentration of <16 µg mL-1 [31]. A growth inhibition of S. cerevisiae was observed at a concentration of <32 µg mL-1 and for yeasts C. parapsilosis and C. tropicalis at <64 µg mL-1 [41].
In Santos et. al’s study after protein extraction from the fruit of Capsicum chinense different fractions were obtained, named F1 to F10 [42]. Peptides in the F4 and F5 fractions were sequenced and revealed similarity with the plant antimicrobial peptides like non-specific lipid transfer proteins and defensin-like peptide. The F4 and F5 fractions presented strong antimycotic activity against the phytopathogenic fungi Fusarium solani and Fusarium oxysporum, causing toxic effects on them, leading to membrane permeabilization, endogenous reactive oxygen species increase, activation of metacaspase and loss of mitochondrial function [42].
Table 2 contains different types of plant AMPs and their mode of action against human- and food-borne pathogens.
Table 2.
Summary of AMPs expressed in vegetables, their modes of action and activity against bacteria and fungi.
Table 2.
Summary of AMPs expressed in vegetables, their modes of action and activity against bacteria and fungi.
| Vegetable AMPs | Mode of action | Active against |
|---|---|---|
| Tomato (snakin SN2) | Pore formation, agglomeration of the cells | S. cerevisiae |
| Onion (Ba-49) | Disruption of the cell membrane, triggering the production of ROS, preventing the formation of biofilm and degrading the formation of mature biofilm | S. aureus |
| Garlic (F3-3-a, F3-3-b, F3-3-c) | Disruption of the cell membrane | E. coli, S. aureus, Salmonella enteritidis, B. subtilis |
| Chili pepper (F3 fraction) | Membrane permeabilization, production of ROS | S. cerevisiae, C. guilliermondii, C. parapsilosis, K. marxiannus, P. membranifaciens, C. tropicalis, C. albicans |
As seen from Table 2, plant peptides are active against pronounced pathogens like E. coli, S. aureus, S. enteridis, C. albicans, C. troppicals and C. parapsilosis. Besides pathogens’ cell membrane disruption, membrane pore formation, agglomeration, ROS production is worthy to highlight also the blocking of pathogen’s biofilm formation or its dysfunction by AMPs.
Other than plant AMPs, another plant-derived compounds are polyphenolic extracts which also possess antimicrobial properties such as antibiofilm activity against C. jejuni, reported by Elgamoudi et. al [43]. Polyphenol-rich cranberry and other berry extracts have been found to have a strong antibiofilm effect on dual-species Streptococcus mutans-Candida albicans biofilms and sole Streptococcus mutans biofilms [43]. Polyphenols have high ability to bind to proline-rich peptides, however the interaction between these compounds, present in plants is still not fully understood [44]. Pharmacokinetics, bioavailability, absorption and metabolism are thought to be affected [44]. The authors concluded that there is a synergism between polyphenols and proline-rich peptides (PRP), by controlling PRP’s gene expression.
Antibacterial activity of different plants against a Gr (+) B. subtilis and Gr (-) negative E. coli shown in Table 3 is due to the presence of AMPs or polyphenols in plants demonstrating strong activities against various pathogens. Highest antimicrobial action on B. subtilis was demonstrated by orange skin onion (27 mm), red skin onion (25 mm), cayenne pepper (24 mm) and on E. coli by garlic (30 mm) and cayenne pepper (25 mm).
Table 3.
Antibacterial activity of AMP/polyphenol containing vegetables (inhibition zones d in mm) against B. subtilis and E. coli type strains [45].
Table 3.
Antibacterial activity of AMP/polyphenol containing vegetables (inhibition zones d in mm) against B. subtilis and E. coli type strains [45].
| Vegetable/plant vegetative organ | Inhibition zone d on B. subtilis NIBMCC 8752 | Inhibition zone d on E. coli NIBMCC 8751 |
|---|---|---|
| Parsley (leaves) | 2 | 0 |
| Tomato (seeds) | 5 | 0 |
| Cayenne pepper (tissue discs) | 24 | 25 |
| Cayenne pepper (seeds) | 7 | 11 |
| Onion orange skin (mature bulbs) | 27 | 3 |
| Onion red skin (mature bulbs) | 25 | 3 |
| Onion young (fresh bulbs) | 0 | 0 |
| Garlic (mature bulbs) | 7 | 30 |
| Garlic young (fresh bulbs) | 2 | 0 |
3. AMPs from Animals
AMPs are widely observed in animals, with the defensin and cathelicidin families being the most common. Cathelicidins are positively charged, amphipathic AMPs, which vary in size and structure[1,46]. Defensins are proven to be an important part of animal’s defense system [1,47,48]. They are cationic and made up of around 29 to 42 amino acids and three pairs of intramolecular disulfide bonds [18,47,48]. Defensins are divided into three groups based on the position of the disulfide bonds, those being α-defensins, β-defensins and θ-defensins [1,47,48].
AMPs in invertebrates are extremely important as these organisms lack an adaptive immune response [6]. Some of the main classes of AMPs found in invertebrates are: in insects (defensins and cecropins), molluscs and nematodes (defensins), horseshoe crabs (big defensins), invertebrate β-defensins and crustaceans (crustins) [6]. There has been a recent discovery of a novel AMP found in marine worms, inhabiting contrasted habitats. The identified AMPs are alvinellacin (ALV), arenicin (ARE) and polaricin (POL – the novel AMP) All three of the AMPs show bactericidal activity against the bacteria typical of the habitat [49].
Vertebrate AMPs largely vary in size (15-200 residues) and are present in fish, amphibians, birds and mammals [6]. AMPs found in fish include β-defensins, cathelicidins, hepicidins, histone-derived peptides and piscidins [18]. Cathelicidins are secreted by secretory granules of immune cells and activate when cleaved [6,46]. After being activated they permeabilize lipid membranes and can act against Gram-positive and negative bacteria [6,46,50]. Fish β-defensins have antibacterial activity and act also against fish-specific viruses. Fish hepcidins are described as cysteine-rich hormones that regulate iron homeostasis and demonstrate antibacterial activity against both Gram-negative and positive bacteria [6].
The main source of AMPs in amphibians are frogs and toads [1]. The most common class that has been observed in them are the magainins, which have an antibacterial activity against both Gram-negative and -positive bacteria and even some fungi [1], (Figure 3).
Insects have also been described to have a large amount of AMPs such as cecropins, which again have been found to have a broad activity against Gram-negative and positive bacteria and some fungi. Other classes of insect AMPs include defensins, proline-rich peptides and attacins [1,52].
Reptile and avian AMPs have been described to be cathelicidins and defensins [6]. A cathelicidin named OH-CATH is a peptide secreted from a king cobra and is active against bacteria such as Pseudomonas aeruginosa and Enterobacter aerogenes. Avian cathelicidins in chicken are fowlicidines which are active against both Gram-negative and positive bacteria [6]. Besides in chicken, cathelicidins or cathelicidin-like peptides are described in duck, turkey, pheasant and quail. Reptile β-defensins were first found in the leukocytes of a European pond turtle TBD-1. Crotamine, pelovaterin and turtle egg-white protein are other AMPs found in reptiles. Whereas avian β-defensins are discovered in chicken-AvBD-1; ostrich-ostricacins and mallard duck-AvBD2, AvBD9 [6].
Mammalian-derived AMPs are mostly from the cathelicidin and defensin classes but others include platelet antimicrobial proteins, hepcidins and dermicidins [6]. As mentioned, LL-37 from the cathelicidin family is the most studied and understood cathelicidin [6,46]. It is active against various Gram-negative and positive bacteria and, if applied on wounds, promotes healing [6,46]. Another member from the cathelicidin family present in mammals is cathelicidin 4, or indolicidin, which is a tryptophan and proline-rich peptide, secreted from bovine neutrophils, which can act against both Gram-negative and positive bacteria [6,46], (Figure 4).
It forms pores in the cell membrane and inhibits DNA synthesis [6,46]. Protegrins (PG) are also from the cathelicidin family, which are secreted from porcine white blood cells and act by increasing the membrane permeabilization and inhibiting RNA synthesis [6,46,54]. Mammalian α-defensins are secreted by promyelocytes, neutrophil precursor cells, and Paneth cells [6,48]. α-defensins isolated from a guinea pig’s neutrophils can be used against S. aureus and E. coli, and those isolated from rabbits have a broad activity against Gram-negative and positive bacteria. Mammalian β-defensins were first isolated from bovine mucosal epithelial cells and have been identified to be active against Gram-positive and negative pathogens [6,48]. Θ-defensins structure is different to α- and β-defensins and are active against B. anthrax, S. aureus and C. albicans [6].
Table 4.
Different classes of AMPs found in animal hosts. Optimized after [18] .
Table 4.
Different classes of AMPs found in animal hosts. Optimized after [18] .
| Animals | AMPs | Reference |
|---|---|---|
| Mammalians | Cathelicidins
Defensins Platelet antimicrobial proteins Dermicidins Hepcidins |
[6,18,46] |
| Reptiles | Defensins Cathelicidins |
[6,18] |
| Fish | β-defensins Cathelicidins Hepicidins (HAMP1 and HAMP2) Histone-derived peptides Piscidins (1-7) |
[6,18] |
| Amphibians | Magainins Cancrins |
[1,6,18] |
| Crustaceans | Crustins | [6,18] |
4. AMPs from Humans
Humans, similarly to animals, can produce AMPs, with LL-37 being the only member from the cathelicidin family that is present in humans [1,46]. LL-37 is produced by epithelial cells and neutrophils and has many activities, such as antibacterial activity, regulation of inflammation, modulation of cell death and has further potential in treating drug-resistant bacterial infections. Although LL-37 is primarily found on newborns’ skin, human AMPs can be found in various parts of the body including eyes, ears, skin, the respiratory tract and lungs, intestines and urethra [1,46]. Some AMPs are constantly being produced in the body, whereas others only when there is an infection or inflammation present [1].
Another class of human AMPs are the defensins. Human α-defensins HNP1-4 are found to be secreted by neutrophils and HD5-6 in the intestinal tract by Paneth cells [6,55]. The most studied and understood defensin is the HNP1, which has antibacterial activity against E. coli, S. aureus, S. epidermis by inhibiting DNA and protein synthesis. Human β-defensins (HBD1-4) have been found to be expressed in many parts of the body including the respiratory, gastrointestinal and urinary tracts, testis and keratinocytes [6,55]. Human β-defensin 2 (HBD-2) is primarily found in elders [1,55], but HBD1-3 has antibacterial activity against Gram-negative bacteria including P. aeruginosa and E. coli and yeasts such as C. albicans. HBD-3 also expresses antibacterial activities against Gram-positive bacteria such as S. pyogenes and S. aureus [6,55]. θ-defensins are not expressed in humans or primates [6].
5. Modes of Action and Mechanisms
5.1. Antiviral AMPs
AMPs that are able to target viruses usually have mechanisms that target both RNA and DNA viruses but depending on the way they act, they can be divided into three groups [6,50,54]. The first one consists of AMPs that target the viral envelope. They act by embedding themselves in the envelope of the virus and thus causing instability and disrupting the mode of action of the virus, an example of which is AMP LL-37 [1,6,50,54]. The second mode of action that AMPs can have against viruses is to prevent them from binding to specific receptors on the target cells by binding to them themselves [6,50,54]. For example, defensins have the ability to link with herpes simplex virus glycopeptides and thus prevent the virus from binding to the target cells receptors [6]. The third group of antiviral AMPs are ones that target internal components by either damaging or blocking certain viral proteins, interfering with viral transcription, targeting the viral nucleocapsid or inhibiting the virus from leaving the host cell [6,50,54].
5.2. Antibacterial AMPs
Antibacterial AMPs have been found to be mostly cationic and amphipathic, which act by interacting with the negatively charged bacterial membrane and causing instability and disruption [6,56]. They can act primarily against Gram-negative bacteria, but some AMPs like daptomycin have a broad spectrum of action against Gram-positive bacteria as well. Glycopeptides act by binding to cell wall precursors and inhibiting the formation of the peptidoglycan [6,56,57]. AMPs have also been suggested as a treatment strategy against H. pylori [58]. Antibacterial AMPs have two major modes of action: either targeting the bacterial membrane or non-membrane targeting, although AMPs can use both mechanisms [6,56,57].
5.3. Antifungal AMPs
Fungi, compared to bacteria, have a cellular wall which mostly consists of chitin. Antifungal AMPs have a similar mode of action to antibacterial AMPs: barrel-stave, toroidal pore and carpet-like models [6,59,60]. These will be further described in the following section. Some examples of antifungal AMPs which use these methods of penetrating the fungal cell wall are LL-37 and dermaseptin. Other than these methods, echinocandins, indolicidin and buforins can inhibit the 1,3-β-glucan and chitin biosynthesis and can also act on other intracellular targets such as nucleic acids and repair mechanisms [6,59,60].
5.4. Membrane-Targeting AMPs
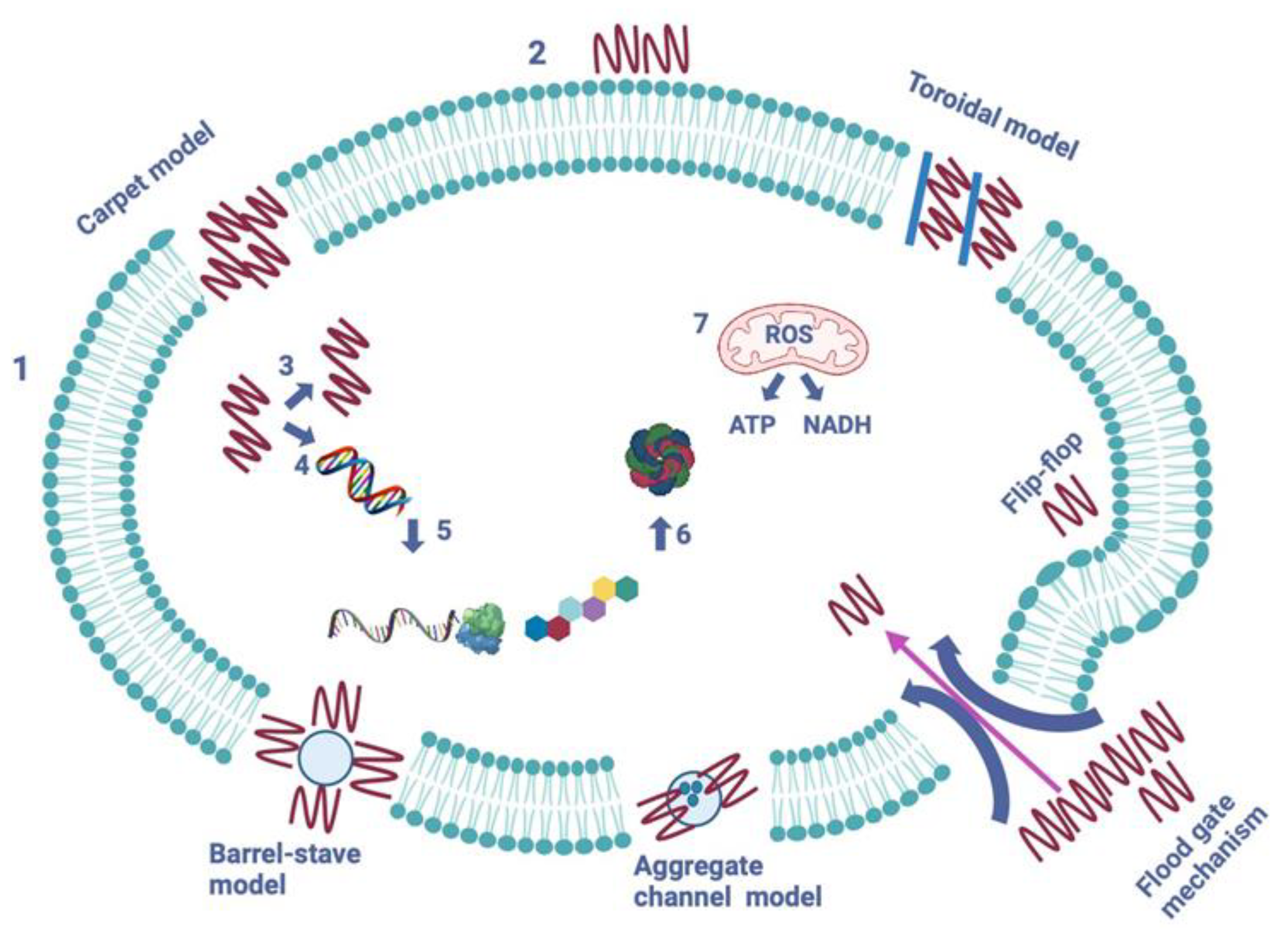
As previously noted, various antibacterial AMPs use a cell membrane targeting mechanism, which consists of the positively charged and amphipathic AMP binding to a negatively charged and hydrophobic cell membrane phospholipids, leading to the formation of pores and channels in the cell wall [6,38,50,56]. Thus far five models of cell membrane modes of action have been described, which are barrel-stave, toroidal-pore, carpet, aggregate channel and flood gate mechanism models [1,56]. The five models can be seen in Figure 5.
The pores formed in the barrel-stave model lead to a cytoplasmic outflow, membrane collapse, permeability and eventually cell death [6,56], (Figure 5). AMPs that use this mode of action include, but are not limited to, ceratotoxins, protegrins and alamethicin [1,56].
The toroidal-pore model suggests that AMPs can cross the lipid membrane by interacting with the lipid head groups, which causes the lipid bilayer to bend and for the peptides to insert themselves into the membrane bilayer and form pores and channels through it. Magainins, protegrins, actinoporins and melittin are some the AMPs which use this mode of action [1,6,56].
The next model, described as the carpet model, AMPs cover the cell membrane’s surface like a carpet and interact with the membrane’s phospholipid head groups [1,6,56]. This eventually leads to high peptide concentrations, damaging and permeation of the phospholipid bilayer. Some of the AMPs that use this mode of action are cecropins, magainin and indolicidin [1,6,56].
In the aggregate channel model, AMPs are described to spontaneously form unstructured peptide aggregates, which surround the pathogen’s membrane. That causes the formation of channels and leakage of cytoplasmic fluid [57].
The last model is a recently proposed mechanism, called floodgate mechanism. During the early stage of the attack, α-helical AMPs form transient toroidal gaps in the pathogen’s cell membrane. It is proposed that AMPs first stress the hydrophobic and electrostatic membrane and after the initial attack they recruit nearby unbound peptides [57].
5.5. Non-Membrane Targeting
Some AMPs can directly penetrate the cell membrane through endocytosis [1,50]. This way they directly act on important bacterial organelles, intracellular proteins or can target RNA, DNA or protein synthesis [1,6,50]. The way AMPs target nucleic acids or protein synthesis is by binding to them and destroying their conformation. Histone-derived AMPs are the most common to use this mechanism, and examples include buforin II as well as indolicidin, the latter of which having activity against both Gram-negative and -positive pathogens. Another way of targeting and inhibiting nucleic acid and protein synthesis is by targeting enzymes and protein in certain metabolic pathways [1,50]. For example, indolicidin can act on type I DNA topoisomerase and thus inhibit the relaxation of double-stranded DNA. Other AMPs can act on RNA polymerase, DNA gyrase and other DNA replication-associated proteins [1,6,50]. Targets for AMPs are also the nucleic acid damage repair pathways; they can disrupt damage response pathways and signaling and instead promote cell death and apoptosis. Some peptides can also target ribosomes to inhibit protein synthesis by acting on the translation pathway, usually caused by proline-rich AMPs. PrAMPs can also act on the folding and assembly of the proteins by inhibiting the bacterial heat shock protein [1,50].
In Table 5 are presented the amino acid sequences of some AMPs varying in length and content, most of which have been taken from The National Center for Biotechnology’s (NCBI) database [61].
Table 5.
Amino acid sequences of some AMPs.
| AMP | Source | Sequence | Reference |
|---|---|---|---|
| LL-37 | Human (Homo sapiens) | LLGDFFRKSKEKIGKEFKRIVQRIKDFLRNLVPRTES | [62] |
| Indolicidin | Cattle (Bos taurus) | ILPWKWPWWPWRR-amide | |
| Crotamine | South American rattlesnake (Crotalus durissus) | YKQCHKKGGHCFPKEKICLPPSSDFGKMDCRWRWKCCKKGSG, Cys4-Cys36, Cys11-Cys30, Cys18-Cys37 | [62] |
| Cancrin | Crab-eating frog (Rana cancrivora) | GSAQPYKQLHKVVNWDPYG | [63] |
| Melittin | Honey bee (Apis mellifera) | GIGAVLKVLTTGLPALISWIKRKRQQ-NH2 | [64] |
| Buforin II | Asian toad (Duttaphrynus melanostictus) | TRSSRAGLQFPVGRVHRLLRK | [65] |
| HNP-1 | Human (Homo sapiens) | ACYCRIPACIAGERRYGTCIpYQGRLWAFCC | [62] |
| HBD-2 | Human (Homo sapiens) | GIGDPVTCLKSGAICHPVFCPRRYKQIGTCGLPGTKCCKKP | [62] |
| Protegrin | Pig (Sus scrofa) | RGGRLCYCRRRFCVCVGR-amide | [62] |
| Magainin 2 | African clawed frog (Xenopus laevis) | GIGKFLHSAKKFGKAFVGEIMNS | [62] |
| Cecropin A | Cecropia moth (Hyalophora cecropia) | KWKLFKKIEKVGQNIRDGIIKAGPAVAVVGQATQIAK-amide | [62] |
| NpRS | Garlic (Allium sativum) | RSLNLLMFR | [39] |
| SN1 | Potato (Solanum tuberosum) | MKLFLLTLLLVTLVITPSLIQTTMAGSNFCDSKCKLRCSKAGLADRCLKYCGICCEECKCVPSGTYGNKHECPCYRDKKNSKGKSKCP | [61] |
| SN2 | Tomato (Solanum lycopersicum) | MAISKALFASLLLSLLLLEQVQSIQTDQVSSNAISEGADSYKKIDCGGACAARCRLSSRPRLCHRACGTCCARCNCVPPGTSGNTETCPCYASLTTHGNKRKCP | [61] |
| CC-AMP1 | Ghost Pepper (Capsicum chinense × frutescens) | ZETLDPICMAKCVLKCGKKAWCLTKCIAGCVL | [40] |
| γ-Purothionin | Wheat (Triticum turgidum) | KICRRRSAGF KGPCMSNKNCAQVCQQEGWG GGNCDGPFRRCKCIRQC | [61,66,67] |
| α-1-Purothionin (precursor) | Bread wheat (Triticum aestivum) | MGSKGLKGVMVCLLILGLVL EQVQVEGKSCCRTTLGRNCYNLCRSRGAQK LCSTVCRCKLTSGLSCPKGFPKLALESNSDEPDTIEYCNLGCRSSVCDYMVNAAADDEEM KLYVENCGDACVNFCNGDAGLTSLDA | [61] |
| Thi2.1 | Thale cress (Arabidopsis thaliana) | MKGRILILSLLIMSLVMAQVQVEAKICCPSNQARNGYSVCRIRFSKGRCMQVSGCQNSDTCPRGWVNAILENSADATNEHCKLGCETSVCGAMNTLQNSDASEIVNGASEQCAKGCSIFCTKSYVVPPGPPKLL | [61] |
| Mj-AMP2 | Garden four-o’clock (Mirabilis jalapa) | MAKVPIAFLKFVIVLILFIAMSGMIEACIGNG GRCNENVGPPYCCSGFCLRQPNQGYGVCRNR | [61] |
| Lipid transfer protein | Common tobacco (Nicotiana tabacum) | MEMVGKIA CFVVLCMVVVAPHAEALSCGQVQSGLAPCLPYLQGRGPLGSCCGGVKGLLGAAKSLSDRKTACTCLKSAANAIKGIDMGKAAGLPGACGVNIPYKISPSTDCSKVQ | [61] |
| Flower -derived plant defensin 2 | Petunia x hybrida | MARSICFFAVATLALMLFAAYEAEAATCKAECPTWDGICINKGPCVKCCKAQPEKFTDGHCSKVLRRCLCTKPCATEEATATLANEVKTMAEALVEEDMME | [61] |
| PmAMP1 | Western white pine (Pinus monticola) | METKHLAYVMFVLVSLFLAMAQPSQASYFSAWVGPGCNNHNARYNKCGCSNISHNVHGGYEFVYQGQAPTAYNTNNCKGVAQTRFSSNVNQACSNFAWKSVFIQC | [61] |
| SmAMP2 | Chickweed (Stellaria media) | MLNMKSFALLMLFATLVGVTIAYDPNGKCGRQYGKCRAGQCCSQYGYCGSGSKYCAHNTPLSEIEPTAAGQCYRGRCSGGLCCSKYGYCGSGPAYCGLGMCQGSCLPDMPNHPAQIQARTEAAQAEAQAEAYNQANEAAQVEAYYQAQTQAQPQVEPAVTKAP | [61] |
| Crustin | Red swamp crayfish (Procambarus clarkii) | MLRVLVLSMLVVAALGHLPRPKPPQPGCNYYCTKPEGPNKGAKYCCGPQFLPLIREEKHNGFCPPPLKDCTRILPPQVCPHDGHCPINQKCCFDTCLDLHTCKPAHFYIN | [61] |
| Hepicidin HAMP2.3 | Gilthead seabream (Sparus aurata) | MKTFSVAVAVAIVLTFICLQESSAVSFTEVQELEEPMSNDGPIAAYKEMPEDSWKMGYGSRRWKCRFCCRCCPRMRGCGLCCRF | [61] |
| Histone -derived, partial | Catla (Labeo catla) | MSGRGKTGGKARAKAKTRSSRAGLQFPVGRVHRLLRKGNYAERVGAGAPVYLAAVLEYLTAEILELAGNAARDNKKTRIIP | [61] |
| Piscidin 2 | Schlegel’s black rockfish (Sebastes schlegelii) | MRFIMLFLVLSMVVLMAEPGEAFIHHIFGAIKRIFGDKQRDMADQQELDQRAFDRERAFN | [61] |
| Proline-rich AMP | Green mud crab (Scylla paramamosain) | MRLLWLLVALAAVVPAAMPASAGYFPGRPPFPRPFPRPPSRPFPRPPFPGPFPRPYPWR |
[61] |
| Attacin | House fly (Musca domestica) | MFTKSIAIIVFLATLAVVNAQFGGSITSNS RGGADVFARLGHQFGDNKRNFGGGVFAAGNTLGGPVTRGAFLSGNADRFGGSLSHSRTDNFGSTFSQKLNANLFQNDKHKLDANAFHSRTNLDNGFKFNTVGGGLDYNHANGHGASVTASRIPQLNMNTVDVTGKANLWK SADRATSLDLTGGVSKNFGG PLDGQTNKHI GVGLSHDF |
[61] |
| Oh-Cath | King cobra (Ophiophagus hannah) | MEGFFWKTLLVVGALAIGGTSSLPHKPLTY EEAVDLAVSIYNSKSGEDSLYRLLEAVPPPEWDPLSESNQELNFTIKETVCLVAEERSLEECDFQEDGAI MGCTGYYFFGESPPVLVLTCKPVGEEEEQK QEEGNEEEKEVEKEEKEEDEKDQPRRVKRF KKFFKKLKNSVKKRAKKFFKKPRVIGVSIPF |
[61] |
| TBD-1 | European pond turtle (Emys orbicularis) | YDLSKNCRLRGGICYIGKCPRRFFRSGSCS RGNVCCLRFG | [61] |
| Pelovaterin | Chinese soft-shelled turtle (Pelodiscus sinensis) | DDTPSSRCGSGGWGPCLPIVDLLCIVHVTV GCSGGFGCCRIG | [61] |
| avian β-defensin 1 | Japanese quail (Coturnix japonica) | MKIVYLLFPFILLLAHGAAGSSRDLGKREQ CYRQKGFCAFLKCPSLTIISGKCSRFHVCCKNIWG | [61] |
| α-defensin 5, Paneth cell-specific | Human (Homo sapiens) | MRTIAILAAI LLVALQAQAESLQERADEAT TQKQSGEDNQDLAISFAGNGLSALRTSGSQARATCYCRTG RCATRESLSGVCEISGRLYRLCCR | [61] |
| θ-defensin-1 | Rhesus monkey (Macaca mulatta) | RCICTRGFCRCLCRRGVC | [61] |
6. Discussion on the benefits and limitations of AMPs
Antibiotic-resistant pathogens have become an urgent contemporary problem as many resistant strains continue to emerge [68]. Antimicrobial peptides can be considered as an alternative therapeutic agent and may therefore be crucial for the fight against antibiotic resistance [6,69]. As AMPs have many mechanisms of actions against pathogens, additional to clinical, they also have potential benefits for agricultural biotechnology [17,69]. There is potential with natural AMPs, such as plant-based AMPs, in terms of large scale production yield, particularly as biotechnological applications in their cultivation [8]. Their clinical efficacy has already been shown [6]. AMPs also have potential as natural food preservatives for food and packaging, as well as a range of health benefits when included in food [19,70]. Interestingly, defatted egg yolk proteins, have been found to be a great source for bioactive peptides, which can potentially find application as natural preservatives [71].
Along with their advantages, natural AMPs have shown to face some limitations as well, such as poor absorption, metabolism, short half-life and low permeability, which have all proven to be a challenge in developing drug-alternative AMPs [6,72]. According to multiple articles, one of the main limitations are their poor stability and susceptibly to proteolytic degradation, which can lead to a reduced half-life and limit their possible route of administration in the pathogen [18,73]. Environmental stress can also be a contributing factor, also leading to degradation of the peptides [74]. While there have been some strategies to address these challenges, there is still perhaps more work to be done to ensure the frequent future use of natural AMPs [6,72]. Another disadvantage is that large-scale production and high cost of production limit their use [18]. Furthermore, as of yet, only a few substances have been approved by regulatory bodies. Those approved include glycopeptides such as vancomycin and teicoplanin, daptomycin and polymyxin B, but many more are currently being investigated and tested for their use [6]. So far many strategies have been developed to increase the stability of AMPs, but that has shown to decrease their antimicrobial activity [75].
Multiple strategies are currently in development to counter the above AMP limitations and so far various design strategies have been used to improve proteolytic stability such as sequence modification, cyclization, peptidomimetics, and nanotechnology [6,17,54,71,73]. Many of the plant-based limitations in particular have already been addressed in recent years [17]. Chemical modification has been shown to be the most frequent and easiest way to improve AMPs activity and biocompatibility [7,72]. For example, modifying CLEC3A – derived AMPs, has shown an increase in their activity, especially against drug-resistant bacteria [76]. Strategies have been developed to reduce the proteolytic degradation of AMPs, one of which is the development of fully functional mimics of CAMPs (cationic AMPs), which are able to evade proteases [77]. In Cafaro et. al’s study a peptoid, named P13#1 is designed to mimic cathelicidins. The authors report strong biological activities of the peptoid, similar to those of human cathelicidin LL-37 and antimicrobial and anti-inflammatory activities comparable to ampicillin and gentamicin, without showing toxicity [77]. A different strategy implemented by Tortorella et. al to overcome the easy degradation of AMPs was to synthesize the N-glycosilated form of LL-III AMP [74]. The authors concluded that glycosylation did not affect the peptide’s mode of action and biological activity, and it in fact made it more resistant to proteolytic degradation [74]. Another way to tackle the disadvantage of reduced antimicrobial activity, when increasing the stability of AMPs was described by He et. al [75]. In their research the authors introduced hydrophobic group modifications at the N-terminus of proteolysis-resistant AMPs D1 [75]. By end-tagging an Nal, the peptide N1 showed strong antimicrobial activity by damaging bacterial cell membranes and inhibiting the bacterial energy metabolism and retained its stability [R11]. Potential approaches to boost the synthesis of next-generation AMPs to be used as antimicrobial drugs are (i) modification of the amide bond, (ii) encapsulation of the peptide in a suitable matrix, (iii) modification of the amino acid composition, and (iv) insertion of diastereomers, combined with de novo design strategies [57]. With regards to regulation and approval, for plant-derived AMPs, regulatory bodies have been providing frameworks in recent years, which for example, has led to the approval of the therapeutic enzyme Elelyso by the FDA [17,78]. Hopefully in future years more plant-derived products will be made available for use [17,78].
The overall future use of AMPs is promising [79,80,81]. Recent advancements have brought us closer to successful implementation [6,78,79]. As studies continue, so too does our ability to increase bioavailability and efficacy, as well as improve the efficiency of production and lower production costs, therefore allowing for greater quantities to be produced [82]. Over the coming decade, we are likely to see further advances in capability, more successful case studies on a trial level, greater capacity to produce and distribute, as well as widespread regulatory approval.
Acknowledgments
This work was conducted in UPIZ-Biology-MF-NBU and financially supported by Grant NBU-GFSR-27/2023, New Bulgarian University, Sofia, Bulgaria.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Li, X.; Zuo, S.; Wang, B.; Zhang, K.; Wang, Y. Antimicrobial Mechanisms and Clinical Application Prospects of Antimicrobial Peptides. Molecules 2022, 27, 2675. [Google Scholar] [CrossRef]
- Chrom, C.L.; Renn, L.M.; Caputo, G.A. Characterization and Antimicrobial Activity of Amphiphilic Peptide AP3 and Derivative Sequences. Antibiotics 2019, 8, 20. [Google Scholar] [CrossRef]
- Chen, Y.; Wang, J.; Li, G.; Yang, Y.; Ding, W. Current Advancements in Sactipeptide Natural Products. Frontiers in Chemistry 2021, 9. [Google Scholar] [CrossRef]
- Iacovelli, R.; Bovenberg, R.A.L.; Driessen, A.J.M. Nonribosomal Peptide Synthetases and Their Biotechnological Potential in Penicillium Rubens. Journal of Industrial Microbiology and Biotechnology 2021, 48. (7-8). [Google Scholar] [CrossRef]
- Mansour, S.C.; Pena, O.M.; Hancock, R.E. Host defense peptides: front-line immunomodulators. Trends Immunol. 2014, 35, 443–450. [Google Scholar] [CrossRef]
- Bin Hafeez, A.; Jiang, X.; Bergen, P.J.; Zhu, Y. Antimicrobial Peptides: An Update on Classifications and Databases. Int. J. Mol. Sci. 2021, 22, 11691. [Google Scholar] [CrossRef]
- Kumar, P.; Kizhakkedathu, J.N.; Straus, S.K. Antimicrobial Peptides: Diversity, Mechanism of Action and Strategies to Improve the Activity and Biocompatibility In vivo. Biomolecules 2018, 8, 4. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Hu, S.; Jian, W.; Xie, C.; Yang, X. Plant antimicrobial peptides: Structures, functions, and applications. Bot. Stud. 2021, 62, 5. [CrossRef]
- Pizzo, E.; Cafaro, V.; Donato, A.D.; Notomista, E.Cryptic Antimicrobial Peptides: Identification Methods and Current Knowledge of Their Immunomodulatory Properties. Current Pharmaceutical Design 24 (10), 1054–1066. [CrossRef]
- Fesenko, I.; Azarkina, R.; Kirov, I.; Kniazev, A.; Filippova, A.; Grafskaia, E.; Lazarev, V.; Zgoda, V.; Butenko, I.; Bukato, O.; Lyapina, I.; Nazarenko, D.; Elansky, S.; Mamaeva, A.; Ivanov, V.; Govorun, V. Phytohormone Treatment Induces Generation of Cryptic Peptides with Antimicrobial Activity in the Moss Physcomitrella Patens. BMC Plant Biology 2019, 19((1)). [Google Scholar] [CrossRef] [PubMed]
- de Oliveira Costa, B.; Franco, O.L. Cryptic Host Defense Peptides: Multifaceted Activity and Prospects for Medicinal Chemistry. Current Topics in Medicinal Chemistry 2020, 20((14)), 1274–1290. [Google Scholar] [CrossRef] [PubMed]
- Bosso, A.; Maro, A.D.; Cafaro, V.; Donato, A.D.; Notomista, E.; Pizzo, E. Enzymes as a Reservoir of Host Defence Peptides. Current Topics in Medicinal Chemistry 2020, 20((14)), 1310–1323. [Google Scholar] [CrossRef]
- Bosso, A.; Gaglione, R.; Di Girolamo, R.; Veldhuizen, E.J.A.; García-Vello, P.; Fusco, S.; Cafaro, V.; Monticelli, M.; Culurciello, R.; Notomista, E.; Arciello, A.; Pizzo, E. Human Cryptic Host Defence Peptide GVF27 Exhibits Anti-Infective Properties against Biofilm Forming Members of the Burkholderia Cepacia Complex. Pharmaceuticals 2022, 15((2)), 260. [Google Scholar] [CrossRef]
- Ciociola, T.; Zanello, P.P.; D’Adda, T.; Galati, S.; Conti, S.; Magliani, W.; Giovati, L. A Peptide Found in Human Serum, Derived from the C-Terminus of Albumin, Shows Antifungal Activity in Vitro and in Vivo. Microorganisms 2020, 8((10)), 1627. [Google Scholar] [CrossRef]
- Pizzo, E.; Pane, K.; Bosso, A.; Landi, N.; Ragucci, S.; Russo, R.; Gaglione, R.; Torres, M.D.T.; de la Fuente-Nunez, C.; Arciello, A.; Di Donato, A.; Notomista, E.; Di Maro, A. Novel Bioactive Peptides from PD-L1/2, a Type 1 Ribosome Inactivating Protein from Phytolacca Dioica L. Evaluation of Their Antimicrobial Properties and Anti-Biofilm Activities. Biochimica et Biophysica Acta (BBA) - Biomembranes 2018, 1860((7)), 1425–1435. [Google Scholar] [CrossRef]
- Luo, Y.; Song, Y. Mechanism of Antimicrobial Peptides: Antimicrobial, Anti-Inflammatory and Antibiofilm Activities. Int. J. Mol. Sci. 2021, 22, 11401. [Google Scholar] [CrossRef] [PubMed]
- Shanmugaraj, B.; Bulaon, C.J.I.; Malla, A.; Phoolcharoen, W. Biotechnological Insights on the Expression and Production of Antimicrobial Peptides in Plants. Molecules 2021, 26, 4032. [Google Scholar] [CrossRef] [PubMed]
- Dini, I.; De Biasi, M.-G.; Mancusi, A. An Overview of the Potentialities of Antimicrobial Peptides Derived from Natural Sources. Antibiotics 2022, 11, 1483. [Google Scholar] [CrossRef] [PubMed]
- Höng, K.; Austerlitz, T.; Bohlmann, T.; Bohlmann, H. The Thionin Family of Antimicrobial Peptides. PLOS ONE 2021, 16((7)), e0254549. [Google Scholar] [CrossRef]
- Kovaleva, V.; Bukhteeva, I.; Kit, O.Y.; Nesmelova, I.V. Plant Defensins from a Structural Perspective. Int. J. Mol. Sci. 2020, 21, 5307. [Google Scholar] [CrossRef]
- PDB ID 6NOM Pinheiro-Aguiar, R.; do Amaral, V.S.G.; Pereira, I.B.; Kurtenbach, E.; Almeida, F.C.L. Nuclear Magnetic Resonance Solution Structure of Pisum Sativum Defensin 2 Provides Evidence for the Presence of Hydrophobic Surface-Clusters. Proteins 2020, 88((1)), 242–246. [Google Scholar] [CrossRef]
- Tam, J.P.; Wang, S.; Wong, K.H.; Tan, W.L. Antimicrobial Peptides from Plants. Pharmaceuticals 2015, 8, 711–757. [Google Scholar] [CrossRef]
- Slavokhotova, A.A.; Rogozhin, E.A. Defense Peptides from the α-Hairpinin Family Are Components of Plant Innate Immunity. Frontiers in Plant Science 2020, 11. [Google Scholar] [CrossRef] [PubMed]
- Su, T.; Han, M.; Cao, D.; Xu, M. Molecular and Biological Properties of Snakins: The Foremost Cysteine-Rich Plant Host Defense Peptides. J. Fungi 2020, 6, 220. [Google Scholar] [CrossRef] [PubMed]
- Chan, Y.-L.; Prasad, V.; Sanjaya; Kuei Hung Chen; Po Chang Liu; Chan, M.-T.; Cheng, C.-P. Transgenic Tomato Plants Expressing an Arabidopsis Thionin (Thi2.1) Driven by Fruit-Inactive Promoter Battle against Phytopathogenic Attack. 2005, 221 (3), 386–393. [CrossRef]
- Prasad, B.D.; Jha, S.; Chattoo, B.B. Transgenic Indica Rice Expressing Mirabilis Jalapa Antimicrobial Protein (Mj-AMP2) Shows Enhanced Resistance to the Rice Blast Fungus Magnaporthe Oryzae. Plant Science 2008, 175((3)), 364–371. [Google Scholar] [CrossRef]
- Sarowar, S.; Kim, Y.J.; Kim, K.D.; Hwang, B.K.; Ok, S.H.; Shin, J.S. Overexpression of Lipid Transfer Protein (LTP) Genes Enhances Resistance to Plant Pathogens and LTP Functions in Long-Distance Systemic Signaling in Tobacco. Plant Cell Reports 2008, 28((3)), 419–427. [Google Scholar] [CrossRef] [PubMed]
- Ghag, S.B.; Shekhawat, U.K.S.; Ganapathi, T.R. Petunia Floral Defensins with Unique Prodomains as Novel Candidates for Development of Fusarium Wilt Resistance in Transgenic Banana Plants. PLoS ONE 2012, 7((6)), e39557. [Google Scholar] [CrossRef] [PubMed]
- Verma, S.; Yajima, W.; Rahman, M.; Shah, S.; Liu, J.-J.; A.K.M. Ekramoddoullah; Nat. A Cysteine-Rich Antimicrobial Peptide from Pinus Monticola (PmAMP1) Confers Resistance to Multiple Fungal Pathogens in Canola (Brassica Napus). Plant Molecular Biology 2012, 79 (1-2), 61–74. [CrossRef]
- Rong, W.; Qi, L.; Wang, J.; Du, L.; Xu, H.; Wang, A.; Zhang, Z. Expression of a Potato Antimicrobial Peptide SN1 Increases Resistance to Take-All Pathogen Gaeumannomyces Graminis Var. Tritici in Transgenic Wheat. Functional & Integrative Genomics 2013, 13((3)), 403–409. [Google Scholar] [CrossRef]
- Vetchinkina, E.M.; Komakhina, V.V.; Vysotskii, D.A.; Zaitsev, D.V.; Smirnov, A.N.; Babakov, A.V.; Komakhin, R.A. [Expression of Plant Antimicrobial Peptide Pro-SmAMP2 Gene Increases Resistance of Transgenic Potato Plants to Alternaria and Fusarium Pathogens. Genetika 2016, 52((9)), 1055–1068. [Google Scholar] [CrossRef]
- Diz, M.S.S.; Carvalho, A.O.; Rodrigues, R.; Neves-Ferreira, A.G.C.; Da Cunha, M.; Alves, E.W.; Okorokova-Façanha, A.L.; Oliveira, M.A.; Perales, J.; Machado, O.L.T.; Gomes, V.M. Antimicrobial Peptides from Chili Pepper Seeds Causes Yeast Plasma Membrane Permeabilization and Inhibits the Acidification of the Medium by Yeast Cells. Biochimica Et Biophysica Acta 2006, 1760((9)), 1323–1332. [Google Scholar] [CrossRef]
- Herbel, V.; Sieber-Frank, J.; Wink, M. The Antimicrobial Peptide Snakin-2 Is Upregulated in the Defense Response of Tomatoes (Solanum Lycopersicum) as Part of the Jasmonate-Dependent Signaling Pathway. Journal of Plant Physiology 2017, 208, 1–6. [Google Scholar] [CrossRef]
- Slezina, M.P.; Istomina, E.A.; Kulakovskaya, E.V.; Abashina, T.N.; Odintsova, T.I. Synthetic Oligopeptides Mimicking γ-Core Regions of Cysteine-Rich Peptides of Solanum lycopersicum Possess Antimicrobial Activity against Human and Plant Pathogens. Curr. Issues Mol. Biol. 2021, 43, 1226–1242. [Google Scholar] [CrossRef]
- Taggar, R.; Singh, S.; Bhalla, V.; Bhattacharyya, M.S.; Sahoo, D.K. Deciphering the Antibacterial Role of Peptide from Bacillus Subtilis Subsp. Spizizenii Ba49 against Staphylococcus Aureus. Frontiers in Microbiology 2021, 12. [Google Scholar] [CrossRef]
- Ramita Taggar; Manoj Jangra; Dwivedi, A.; Bansal, K.; Patil, P.B.; Mani Shankar Bhattacharyya; Hemraj Nandanwar; Sahoo, D.K. Bacteriocin Isolated from the Natural Inhabitant of Allium Cepa against Staphylococcus Aureus. World Journal of Microbiology & Biotechnology 2021, 37 (2). [CrossRef]
- Chidike Ezeorba, T.P.; Ezugwu, A.L.; Chukwuma, I.F.; Anaduaka, E.G.; Udenigwe, C.C. Health-Promoting Properties of Bioactive Proteins and Peptides of Garlic (Allium Sativum). Food Chemistry 2024, 435, 137632. [Google Scholar] [CrossRef]
- Gao, X.; Chen, Y.; Chen, Z.; Xue, Z.; Jia, Y.; Guo, Q.; Ma, Q.; Zhang, M.; Chen, H. Identification and Antimicrobial Activity Evaluation of Three Peptides from Laba Garlic and the Related Mechanism. Food & Function 2019, 10((8)), 4486–4496. [Google Scholar] [CrossRef]
- Li, S.; Wang, Y.; Zhou, J.; Wang, J.; Zhang, M.; Chen, H. Structural Characterization, Cytotoxicity, and the Antifungal Mechanism of a Novel Peptide Extracted from Garlic (Allium sativa L.). Molecules 2023, 28, 3098. [Google Scholar] [CrossRef] [PubMed]
- Culver, K.D.; Allen, J.L.; Shaw, L.N.; Hicks, L.M. Too Hot to Handle: Antibacterial Peptides Identified in Ghost Pepper. Journal of natural products 2021, 84((8)), 2200–2208. [Google Scholar] [CrossRef] [PubMed]
- Ribeiro, S.F.F.; Carvalho, A.O.; Da Cunha, M.; Rodrigues, R.; Cruz, L.P.; Melo, V.M.M.; Vasconcelos, I.M.; Melo, E.J.T.; Gomes, V.M. Isolation and Characterization of Novel Peptides from Chilli Pepper Seeds: Antimicrobial Activities against Pathogenic Yeasts. Toxicon 2007, 50((5)), 600–611. [Google Scholar] [CrossRef]
- de Azevedo dos Santos, L.; Taveira, G.; da Silva, M.; da Silva Gebara, R.; da Silva Pereira, L.; Perales, J.; Teixeira-Ferreira, A.; de Oliveira Mello, É.; de Oliveira Carvalho, A.; Rodrigues, R.; Gomes, V. Antimicrobial Peptides from Capsicum Chinense Fruits: Agronomic Alternatives against Phytopathogenic Fungi. Bioscience Reports 2020, 40((8)). [Google Scholar] [CrossRef] [PubMed]
- Elgamoudi, B.A.; Korolik, V. Campylobacter Biofilms: Potential of Natural Compounds to Disrupt Campylobacter jejuni Transmission. Int. J. Mol. Sci. 2021, 22, 12159. [Google Scholar] [CrossRef]
- Pérez-Gregorio, R.; Soares, S.; Mateus, N.; de Freitas, V. Bioactive Peptides and Dietary Polyphenols: Two Sides of the Same Coin. Molecules 2020, 25, 3443. [Google Scholar] [CrossRef]
- Satchanska, G. Antibacterial Activity of Plant Polyphenols. Secondary Metabolites - Trends and Reviews 2022. IntechOpen. [CrossRef]
- Kościuczuk, E.M.; Lisowski, P.; Jarczak, J.; Strzałkowska, N.; Jóźwik, A.; Horbańczuk, J.; Krzyżewski, J.; Zwierzchowski, L.; Bagnicka, E. Cathelicidins: Family of Antimicrobial Peptides. A Review. Molecular Biology Reports 2012, 39((12)), 10957–10970. [Google Scholar] [CrossRef]
- Chang, T.L.; Vargas, J.; DelPortillo, A.; Klotman, M.E. Dual Role of α-Defensin-1 in Anti–HIV-1 Innate Immunity. Journal of Clinical Investigation 2005, 115((3)), 765–773. [Google Scholar] [CrossRef] [PubMed]
- Shafee, T.M.A.; Lay, F.T.; Phan, T.K.; Anderson, M.A.; Hulett, M.D. Convergent Evolution of Defensin Sequence, Structure and Function. Cellular and Molecular Life Sciences 2016, 74((4)), 663–682. [Google Scholar] [CrossRef] [PubMed]
- Bruno, R.; Boidin-Wichlacz, C.; Melnyk, O.; Zeppilli, D.; Landon, C.; Thomas, F.; Cambon, M.-A.; Lafond, M.; Mabrouk, K.; Massol, F.; Hourdez, S.; Maresca, M.; Jollivet, D.; Tasiemski, A. The Diversification of the Antimicrobial Peptides from Marine Worms Is Driven by Environmental Conditions. Science of The Total Environment 2023, 879, 162875. [Google Scholar] [CrossRef] [PubMed]
- Le, C.-F.; Fang, C.-M.; Sekaran, S.D. Intracellular Targeting Mechanisms by Antimicrobial Peptides. Antimicrobial Agents and Chemotherapy 2017, 61((4)). [Google Scholar] [CrossRef] [PubMed]
- PDB ID 2MAG Gesell, J.; Zasloff, M.; Opella, S.J. Journal of Biomolecular NMR 1997, 9 (2), 127–135. [CrossRef]
- Mylonakis, E.; Podsiadlowski, L.; Muhammed, M.; Vilcinskas, A. Diversity, Evolution and Medical Applications of Insect Antimicrobial Peptides. Philosophical Transactions of the Royal Society B: Biological Sciences 2016, 371((1695)), 20150290. [Google Scholar] [CrossRef] [PubMed]
- PDB ID 1G89 Rozek, A.; Friedrich, C.L.; Hancock, R.E. Structure of the Bovine Antimicrobial Peptide Indolicidin Bound to Dodecylphosphocholine and Sodium Dodecyl Sulfate Micelles. Biochemistry 2000, 39((51)), 15765–15774, PDB. [Google Scholar] [CrossRef]
- Kalle Pärn; Elo Eriste; Langel, Ü. The Antimicrobial and Antiviral Applications of Cell-Penetrating Peptides. Springer eBooks 2015, 223–245. [CrossRef]
- Ahmed, A.; Siman-Tov, G.; Hall, G.; Bhalla, N.; Narayanan, A. Human Antimicrobial Peptides as Therapeutics for Viral Infections. Viruses 2019, 11, 704. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.-Y.; Yan, Z.-B.; Meng, Y.-M.; Hong, X.-Y.; Shao, G.; Ma, J.-J.; Cheng, X.-R.; Liu, J.; Kang, J.; Fu, C.-Y. Antimicrobial Peptides: Mechanism of Action, Activity and Clinical Potential. Military Medical Research 2021, 8, 48. [Google Scholar] [CrossRef]
- Kapil, S.; Sharma, V. D-Amino Acids in Antimicrobial Peptides: A Potential Approach to Treat and Combat Antimicrobial Resistance. Canadian Journal of Microbiology 2020. [Google Scholar] [CrossRef]
- Hamza Olleik; Perrier, J.; Hijazi, A.; Baydoun, E.; Maresca, M. Antimicrobial Peptides and Peptidomimetics as Treatment Option for Helicobacter Pylori Infection. WORLD SCIENTIFIC eBooks 2023, 25–56. [CrossRef]
- Buda De Cesare, G.; Cristy, S.A.; Garsin, D.A.; Lorenz, M.C. Antimicrobial Peptides: A New Frontier in Antifungal Therapy. mBio 2020, 11((6)). [Google Scholar] [CrossRef]
- Fernández de Ullivarri, M.; Arbulu, S.; Garcia-Gutierrez, E.; Cotter, P.D. Antifungal Peptides as Therapeutic Agents. Frontiers in Cellular and Infection Microbiology 2020, 10. [Google Scholar] [CrossRef] [PubMed]
- NCBI. National Center for Biotechnology Information. Nih.gov. https://www.ncbi.nlm.nih.gov/.
- Almeida, P.F.; Pokorny, A. 5.10 Interactions of Antimicrobial Peptides with Lipid Bilayers. ScienceDirect. https://www.sciencedirect.com/science/article/abs/pii/B9780123749208005154 (accessed 2023-11-06).
- Huan, Y.; Kong, Q.; Mou, H.; Yi, H. Antimicrobial Peptides: Classification, Design, Application and Research Progress in Multiple Fields. Frontiers in Microbiology 2020, 11. [Google Scholar] [CrossRef] [PubMed]
- Guha, S.; Ferrie, R.P.; Ghimire, J.; Ventura, C.R.; Wu, E.; Sun, L.; Kim, S.Y.; Wiedman, G.R.; Hristova, K.; Wimley, W.C. Applications and Evolution of Melittin, the Quintessential Membrane Active Peptide. Biochemical Pharmacology 2021, 193, 114769. [Google Scholar] [CrossRef] [PubMed]
- Park, C.B.; Yi, K.S.; Matsuzaki, K.; Kim, M.S.; Kim, S.C. Structure-Activity Analysis of Buforin II, a Histone H2A-Derived Antimicrobial Peptide: The Proline Hinge Is Responsible for the Cell-Penetrating Ability of Buforin II. Proceedings of the National Academy of Sciences of the United States of America 2000, 97((15)), 8245–8250. [Google Scholar] [CrossRef] [PubMed]
- Morris, C.F. The Antimicrobial Properties of the Puroindolines, a Review. World Journal of Microbiology & Biotechnology 2019, 35((6)). [Google Scholar] [CrossRef]
- Colilla, F.J.; Rocher, A.; Mendez, E. γ-Purothionins: Amino Acid Sequence of Two Polypeptides of a New Family of Thionins from Wheat Endosperm. FEBS Letters 1990, 270((1-2)), 191–194. [Google Scholar] [CrossRef] [PubMed]
- Mahlapuu, M.; Håkansson, J.; Ringstad, L.; Björn, C. Antimicrobial Peptides: An Emerging Category of Therapeutic Agents. Frontiers in Cellular and Infection Microbiology 2016, 6. [Google Scholar] [CrossRef] [PubMed]
- Moretta, A.; Scieuzo, C.; Petrone, A.M.; Salvia, R.; Manniello, M.D.; Franco, A.; Lucchetti, D.; Vassallo, A.; Vogel, H.; Sgambato, A.; Falabella, P. Antimicrobial Peptides: A New Hope in Biomedical and Pharmaceutical Fields. Frontiers in Cellular and Infection Microbiology 2021, 11. [Google Scholar] [CrossRef]
- Rai, M.; Pandit, R.; Gaikwad, S.; Kövics, G. Antimicrobial Peptides as Natural Bio-Preservative to Enhance the Shelf-Life of Food. Journal of Food Science and Technology 2016, 53((9)), 3381–3394. [Google Scholar] [CrossRef]
- Czelej, M.; Czernecki, T.; Garbacz, K.; Wawrzykowski, J.; Jamioł, M.; Michalak, K.; Walczak, N.; Wilk, A.; Waśko, A. Egg Yolk as a New Source of Peptides with Antioxidant and Antimicrobial Properties. Foods 2023, 12, 3394. [Google Scholar] [CrossRef]
- Mahlapuu, M.; Björn, C.; Ekblom, J. Antimicrobial Peptides as Therapeutic Agents: Opportunities and Challenges. Critical Reviews in Biotechnology 2020, 40((7)), 978–992. [Google Scholar] [CrossRef]
- Lai, Z.; Yuan, X.; Chen, H.; Zhu, Y.; Dong, N.; Shan, A. Strategies Employed in the Design of Antimicrobial Peptides with Enhanced Proteolytic Stability. Biotechnology Advances 2022, 59, 107962. [Google Scholar] [CrossRef] [PubMed]
- Tortorella, A.; Leone, L.; Lombardi, A.; Pizzo, E.; Bosso, A.; Winter, R.; Petraccone, L.; Del Vecchio, P.; Oliva, R. The Impact of N-Glycosylation on the Properties of the Antimicrobial Peptide LL-III. Scientific Reports 2023, 13((1)), 3733. [Google Scholar] [CrossRef]
- He, S.; Yang, Z.; Li, X.; Wu, H.; Zhang, L.; Shan, A.; Wang, J. Boosting Stability and Therapeutic Potential of Proteolysis-Resistant Antimicrobial Peptides by End-Tagging β-Naphthylalanine. Acta Biomaterialia 2023, 164, 175–194. [Google Scholar] [CrossRef]
- Meinberger, D.; Drexelius, M.G.; Grabeck, J.; Hermes, G.; Roth, A.; Elezagic, D.; Neundorf, I.; Streichert, T.; Klatt, A.R. Modified CLEC3A-Derived Antimicrobial Peptides Lead to Enhanced Antimicrobial Activity against Drug-Resistant Bacteria. Antibiotics 2023, 12, 1532. [Google Scholar] [CrossRef]
- Cafaro, V.; Bosso, A.; Di Nardo, I.; D’Amato, A.; Izzo, I.; De Riccardis, F.; Siepi, M.; Culurciello, R.; D’Urzo, N.; Chiarot, E.; et al. The Antimicrobial, Antibiofilm and Anti-Inflammatory Activities of P13#1, a Cathelicidin-like Achiral Peptoid. Pharmaceuticals 2023, 16, 1386. [Google Scholar] [CrossRef]
- Chen, C.H.; Lu, T.K. Development and Challenges of Antimicrobial Peptides for Therapeutic Applications. Antibiotics 2020, 9, 24. [Google Scholar] [CrossRef]
- Lei, J.; Sun, L.; Huang, S.; Zhu, C.; Li, P.; He, J.; Mackey, V.; Coy, D.H.; He, Q. The Antimicrobial Peptides and Their Potential Clinical Applications. American Journal of Translational Research 2019, 11((7)), 3919–3931. [Google Scholar]
- Malanovic, N.; Lohner, K. Antimicrobial Peptides Targeting Gram-Positive Bacteria. Pharmaceuticals 2016, 9, 59. [Google Scholar] [CrossRef]
- Shamseddine, L.; Roblin, C.; Veyrier, I.; Basset, C.; De Macedo, L.; Boyeldieu, A.; Maresca, M.; Nicoletti, C.; Brasseur, G.; Kieffer-Jaquinod, S.; Courvoisier-Dezord E.; Amouric, A.; Carpentier, P.; Campo, N.; Berge, M.; Polard, P.; Perrier, J.; Duarte, V.; Lafond, M. Mechanistic and functional aspectsof the Ruminococcin C sactipeptide isoforms. iScience 2023, 26, 107563, 1-20. [CrossRef]
- Toke, O. Antimicrobial Peptides: New Candidates in the Fight against Bacterial Infections. Biopolymers 2005, 80((6)), 717–735. [Google Scholar] [CrossRef]
Figure 1.
Simplified models of alpha helix, β-sheet and extended coil structures. Created in BioRender.com.
Figure 1.
Simplified models of alpha helix, β-sheet and extended coil structures. Created in BioRender.com.
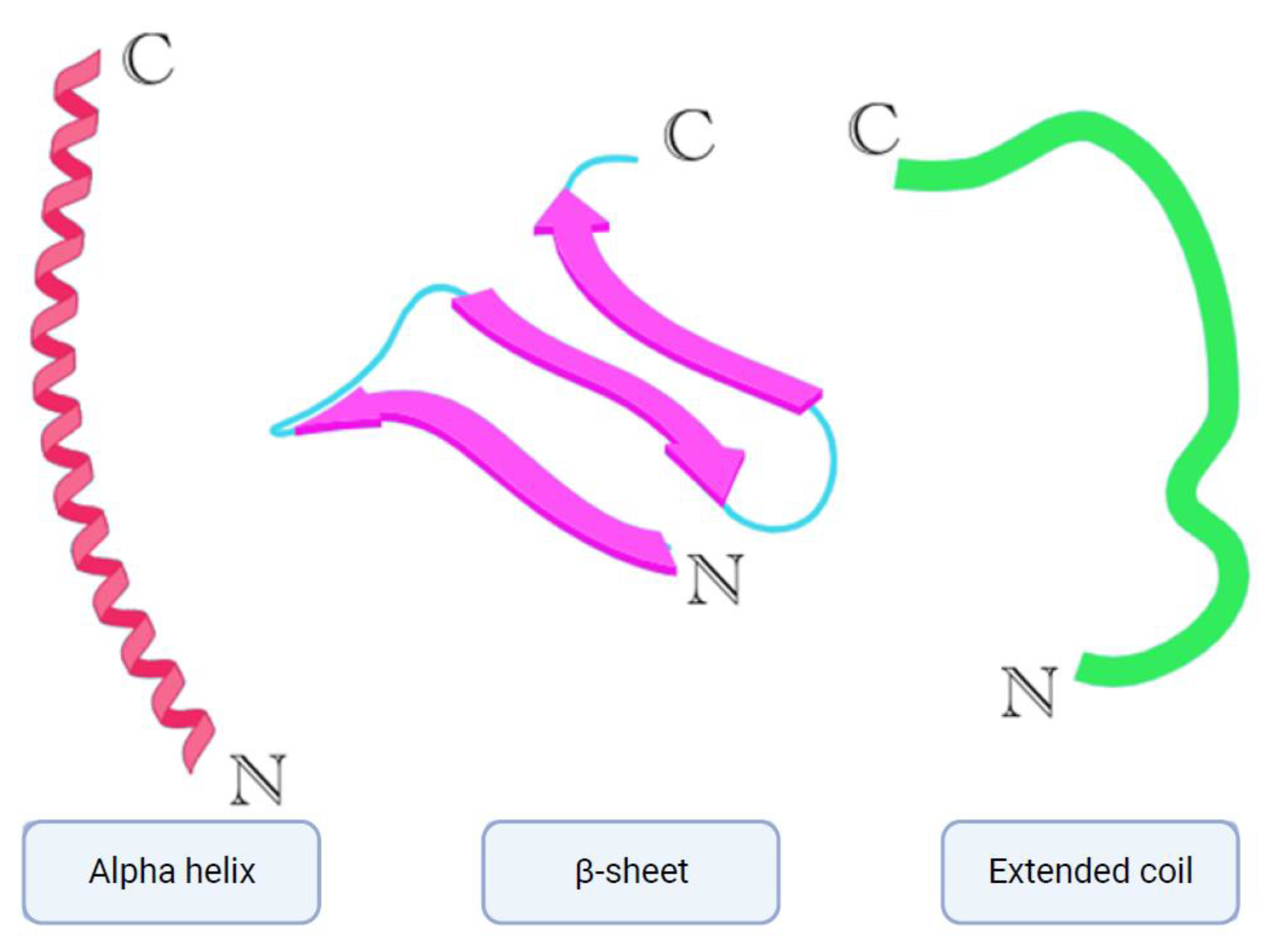
Figure 2.
3-D structure of 4.7 kDa defensin with antifungal activity derived from Pisum sativum (Source: RCSB PDB/ID 6NOM, Pinheiro-Aguiar et al. [21]).
Figure 2.
3-D structure of 4.7 kDa defensin with antifungal activity derived from Pisum sativum (Source: RCSB PDB/ID 6NOM, Pinheiro-Aguiar et al. [21]).
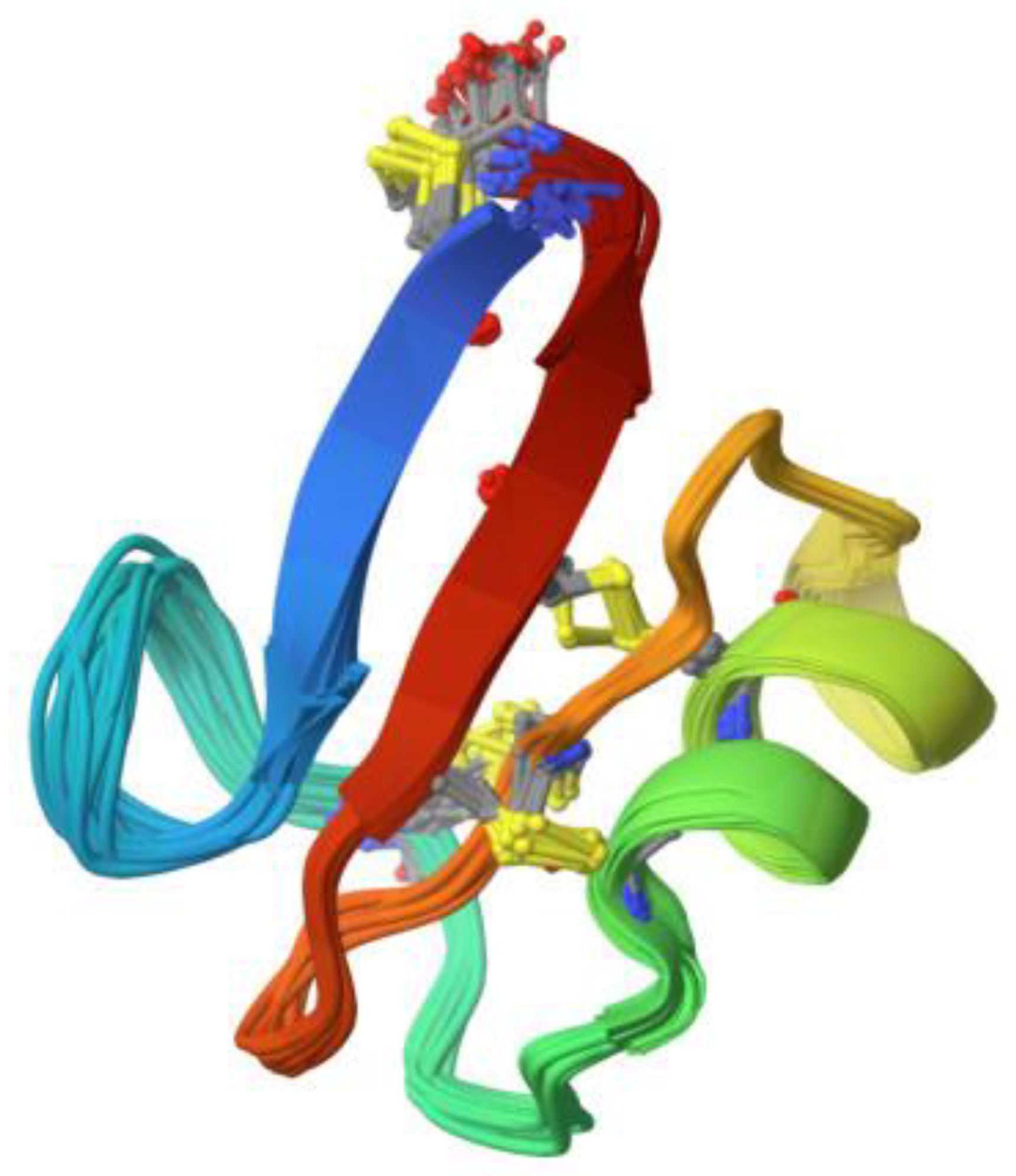
Figure 3.
3-D structure of 23-residue antibiotic magainin isolated from Xenopus laevis (Source: RCSB PDB/ID 2MAG, Gesell et al., 1997 [51].
Figure 3.
3-D structure of 23-residue antibiotic magainin isolated from Xenopus laevis (Source: RCSB PDB/ID 2MAG, Gesell et al., 1997 [51].

Figure 4.
3-D structure of unusually rich in tryptophan bovine antimicrobial peptide indolicidin (Source: RCSB PDB/ID 1G89, Rozek et al., 2000 [53].
Figure 4.
3-D structure of unusually rich in tryptophan bovine antimicrobial peptide indolicidin (Source: RCSB PDB/ID 1G89, Rozek et al., 2000 [53].
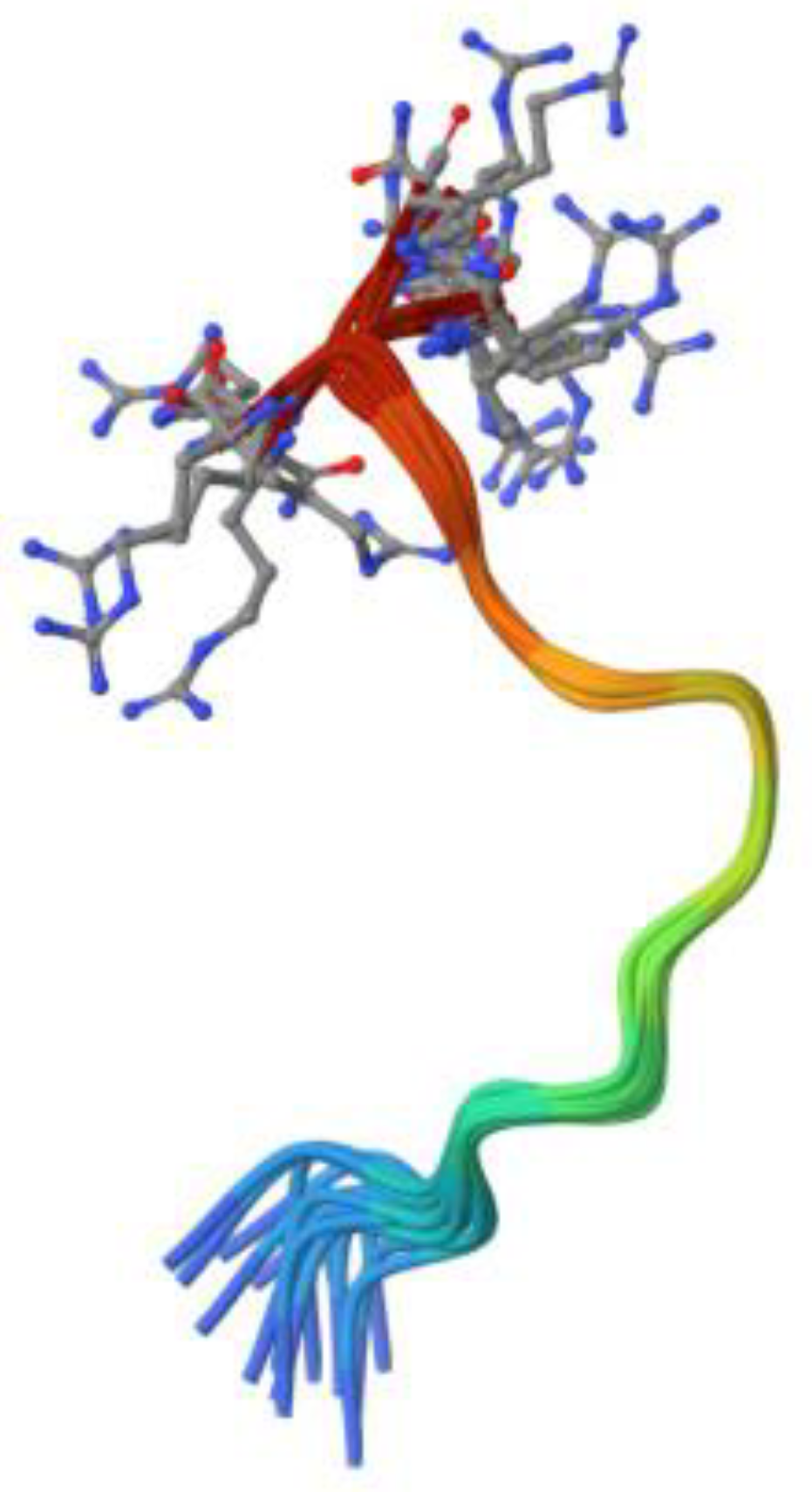
Figure 5.
Figure, showing the mechanisms of action of AMPs on a pathogenic cell. Two major types of peptides are pictured–membrane-bound and intracellular active peptides. The outer layer “1” is the cytoplasmic membrane of pathogen cell and the AMPs are marked as ”2”. Membrane-bound peptides are represented by five mechanisms: barrel-stave, toroidal, carpet, flood-gate and aggregate channel model. The intracellular AMPs are represented by numbers “3”- “7” and show the inhibition of enzymes required for binding of cell wall structural proteins, DNA and RNA synthesis, ribosomal functions and chaperone protein synthesis, and cellular respiration by ROS formation. Optimized after Kapil et al., 2020 [57]. Created in BioRender.com.
Figure 5.
Figure, showing the mechanisms of action of AMPs on a pathogenic cell. Two major types of peptides are pictured–membrane-bound and intracellular active peptides. The outer layer “1” is the cytoplasmic membrane of pathogen cell and the AMPs are marked as ”2”. Membrane-bound peptides are represented by five mechanisms: barrel-stave, toroidal, carpet, flood-gate and aggregate channel model. The intracellular AMPs are represented by numbers “3”- “7” and show the inhibition of enzymes required for binding of cell wall structural proteins, DNA and RNA synthesis, ribosomal functions and chaperone protein synthesis, and cellular respiration by ROS formation. Optimized after Kapil et al., 2020 [57]. Created in BioRender.com.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



