
Submitted:
22 December 2023
Posted:
25 December 2023
You are already at the latest version
Abstract
Serine is widely involved in antioxidant synthesis, immune response, and fat metabolism. However, it remains unclear whether dietary serine supplementation affects fat deposition in the skeletal muscles of pigs. Thus, we explored the effects of dietary serine supplementation on growth performance, meat quality, and composition of gut microbes and their metabolites in growing-finishing pigs. Forty-eight boars weighing approximately 20 kg were fed either a basal diet or a basal diet supplemented with 0.15% serine. The results showed that dietary serine increased the daily weight gain and improved the antioxidant ability of pigs, as indicated by the decreased malondialdehyde content and increased glutathione and superoxide dismutase content. Pigs supplemented with serine had better meat quality, as shown by the lower drip loss and higher IMF content. Furthermore, dietary serine increased the relative abundance of Streptococcus and Lactobacillus and decreased the relative abundance of Clostridium_sensu_stricto_1 and Terrisporobacter. Differential microbial metabolites were mostly enriched in metabolic pathways related to lipid synthesis, such as alpha-linolenic acid metabolism and steroid hormone biosynthesis. Correlation analysis showed that the altered metabolites were closely related to the intestinal microbiota. In conclusion, our results suggested that serine serves as a potential additive for improving meat quality in growing-finishing pigs.
Keywords:
Finishing pig
; Gut microbe
; Meat quality
; Metabolite
; Serine
Introduction
Markets have preferred pig breeds with fast growth rates and have neglected the importance of meat quality under intensive cultivation conditions for the past decades to meet the needs of the masses [1,2,3]. However, with economic development and the increasing focus on health, more attention has been paid to meat quality. Dietary nutrients can improve meat quality in pigs [4]. For example, adding β-glucan in finishing pigs can increase intramuscular fat (IMF) content and adjust the ratio of saturated and unsaturated fatty acids [5]; fermented okara can improve meat color, glutathione peroxidase (GSH-PX) and total superoxide dismutase (T-SOD) activity, which are beneficial to meat quality [6]. Moreover, certain functional amino acids, including tryptophan, threonine, arginine, and leucine, improve meat quality [7]. Additionally, IMF content increased in growing-finishing pigs when methionine intake was restricted after weaning [8], meat color and fatty acid composition improved, and IMF content increased in growing-finishing pigs supplemented with arginine and glutamic acid [9].
Serine participates in various metabolic pathways, including glutathione, purine, and pyrimidine synthesis [10]. It enhances the host antioxidant abilities by promoting the synthesis of GSH and folate-dependent NADPH [11,12]. Our previous study showed that the ratio of serine to glycine affected the IMF content in growing-finishing pigs fed a low-protein diet [13]. It has been demonstrated that both exogenous and endogenous serine residues inhibit lipid deposition in the liver [14]. However, whether dietary supplementation with serine alone affects fat deposition in the skeletal muscle of pigs remains unknown.
The intestinal microbiota composition has been reported to mediate the beneficial effects of dietary functional nutrients on meat quality. For instance, the byproduct of rice distilling improved meat drip loss by influencing the relative abundance of Erysipelotricchaceae and Porphyromonadaceae in growing-finishing pigs [15], and dietary yeast β-glucan improved meat pH value and water-holding capacity by decreasing relative abundance of Candidatus Eremiobacterota [16]. We previously found that serine increased the relative abundance of Clostridia and Firmicutes and the number of operational taxonomic units (OTUs) in mice [17], indicating its effects on microbiota composition. However, whether serine affects meat quality by modulating intestinal microbiota composition in growing-finishing pigs remains unknown. In this study, we aimed to explore the effects of serine on the growth performance, meat quality, and serum biochemical parameters of growing-finishing pigs. Furthermore, we determined the composition of intestinal microbes and their metabolites to investigate whether microbes were involved in the effects of serine supplementation on growing-finishing pigs.
Materials and Methods
Experimental design
In this study, 48 healthy crossbred (Duroc × Landrace × Yorkshire) male pigs with an average initial body weight (BW) of approximately 20 kg were selected. Pigs were randomly divided into two groups (six pens per treatment and four pigs per pen) and fed either a basal diet (CONT) or a basal diet supplemented with 0.15% serine (SER). The animals had ad libitum access to water and feed during the 17-week experimental period. The initial and final BW and dietary consumption of each pen were recorded during the experiment. The average daily feed intake (ADFI), average daily gain (ADG), and feed-to-gain (F/G) ratio were calculated. All nutrients conformed to the requirements of the National Research Council (NRC) (2012), and the diet composition and nutrient level is shown in Table 1. The experimental protocol was approved by the Protocol Management and Review Committee of the Institute of Subtropical Agriculture, Chinese Academy of Sciences.
Sample collection
Fecal samples were collected from all the pigs the day before slaughter. All fecal samples were transferred into sterile microcentrifuge tubes immediately and then stored at −80 ℃ for further microbial and metabolomic analysis [18]. Blood samples were collected through anterior vena cava puncture before slaughter and centrifuged at 3000 × g at 4 ℃ for 10 minutes to collect serum samples. All serum samples were stored at −80 ℃ until analysis. Longissimus dorsi muscle samples between the 6th and 7th rib were obtained and then stored at 4 ℃ for 24 hours for meat quality analysis.
Biochemical parameters
Commercially available kits (Jiancheng, Nanjing, China) were used to measure malondialdehyde (MDA) content, and kits (BYabscience, Nanjing, China) were used to detect glutathione (GSH) and superoxide dismutase (SOD) content.
Meat quality
Initial and ultimate pH values (pH45mi and pH24h) post-mortem were measured, and meat color was evaluated on a freshly cut surface of longissimus dorsi using a colorimeter with the parameters L* (brightness), a*(redness), and b* (yellowness) [19,20]. Longissimus dorsi samples were weighed before and after 24-hour storing at 4 ℃ (W1 and W2, respectively), and drip loss was calculated as ((W1-W2)/W1) [8,21]. Intramuscular fat was defined as the ratio of crude fat weight to the longissimus dorsi muscle weight. Crude fat was extracted using Soxhlet extraction.
Fecal microbiota profiling
Fecal DNA was obtained using a Magnetic Soil and Stool DNA Kit (TianGen, Shanghai, China; Catalog #: DP712). Specific primers were selected for the V3-V4 region of 16S rDNA. Phusion High-Fidelity PCR Master Mix with GC Buffer was provided by New England Biolabs Company (USA) for PCR. Amplicons were purified using a Universal DNA Purification Kit (TianGen, Shanghai, China, Catalog #: DP214) and sequenced on Illumina Novaseq6000 platforms. Fastp (version 0.23.1) software was used for quality filtering to acquire high-quality clean tags. Microbial sequences were categorized into OTUs based on 97% identity. Species annotations for each representative sequence were used to obtain species messages, including microbial relative abundance and distribution, evenness messages, and mutual or specific OTUs. Intestinal microbial alpha and beta diversity were analyzed using QIIME software. Moreover, microbial community differences were directly displayed in dimensionality reduction maps, including principal coordinate analysis (PCoA) through OTUs multi-sequence comparison.
Determination of fecal metabolites
Mixtures of fecal samples and an 80% methanol solution were centrifuged to obtain the supernatant, and metabolites were analyzed using liquid chromatography-mass spectrometry (LC-MS). Metabolites were detected using a high-resolution series mass spectrometer and annotated after matching the exact molecular mass data (m/z) based on the online Human Metabolome Database (HMDB). Differential metabolites were detected using Student’s t-test. Multiple tests were implemented using false discovery rate (FDR) to assess significantly altered metabolites in accordance with the P-value. We performed a correlation analysis based on the Pearson correlation coefficient after obtaining microbial and metabolomic data to explore the relative extent of metabolite and microbial species diversity.
Statistical analysis
All data were analyzed using one-way ANOVA followed by Duncan’s multiple comparison test. Data statistics software (SPSS 25.0) was used, and a probability value (P-value) < 0.05 was set as a statistically significant difference.
Results
Growth performance and serum biochemical parameters
The growth performance of the pigs is shown in Table 2. Compared to control pigs, pigs in the SER group had higher final BW and ADFI and lower F/G; however, the differences were not significant. ADG was significantly higher in pigs in the SER group than in those in the CONT group. As shown in Table 3, serine significantly decreased the MDA content and increased the GSH and SOD contents in the serum of growing-finishing pigs (P < 0.05).
Meat quality
As shown in Table 4, we did not observe significant differences in meat color (L, a, and b), pH45min, or pH24h value (P > 0.05) between the treatment groups. However, drip loss was significantly lower (P < 0.05) and IMF content was significantly higher (P < 0.05) in pigs supplemented with 0.15% serine than in the control pigs.
Fecal microbiota composition
The fecal microbiota composition was compared using a 16S rDNA phylogenetic approach. The Venn diagram indicated that pigs in the CONT group had 1827 unique OTUs, whereas pigs in the SER group had 2062 unique OTUs and 1489 universal OTUs (Figure 1A). Fecal microbial α-diversity had no significant difference between different groups, as indicated using Chao1 index (Figure 1B). The major microbes at the class level included Clostridia, Bacilli, Bacteroidia, Gammaproteobacteria, and Spirochaetia, and serine increased the relative abundance of Bacilli and Bacteroidia and decreased Clostridia relative abundance (Figure 1C). The major microbes at the genus level included Streptococcus, Clostridium_sensu_stricto_1, Lactobacillus, and Terrisporobacter. Serine supplementation increased Streptococcus and Lactobacillus relative abundance, and decreased Clostridium_sensu_stricto_1 and Terrisporobacter relative abundance (Figure 1D).
As indicated by the results of the weighted UniFrac t-test and PCoA (Figure 2A) based on the Jaccard distance matrix (Figure 2B), the beta diversity was distinctively different between pigs in the CONT and SER groups. We then filtered out the significantly different microbes at the genus and species levels using a statistical t−test. The results showed that at the genus level, the relative abundances of Lactobacillus, Prevotellaceae_UCG-001, and Phascolarctobacterium were significantly increased, whereas the relative abundances of Clostridium_sensu_stricto-1, Terrisporobacter, Eubacterium_coprostanollgenes group, Listeria, Candidatus_Soleaferrea, and UCG-009 were significantly decreased by serine supplementation (Figure 2C). At the species level, the relative abundance of Clostridium butyricum was significantly decreased, and that of Lactobacillus delbrueckii was significantly increased by serine supplementation (Figure 2D).
Fecal metabolites
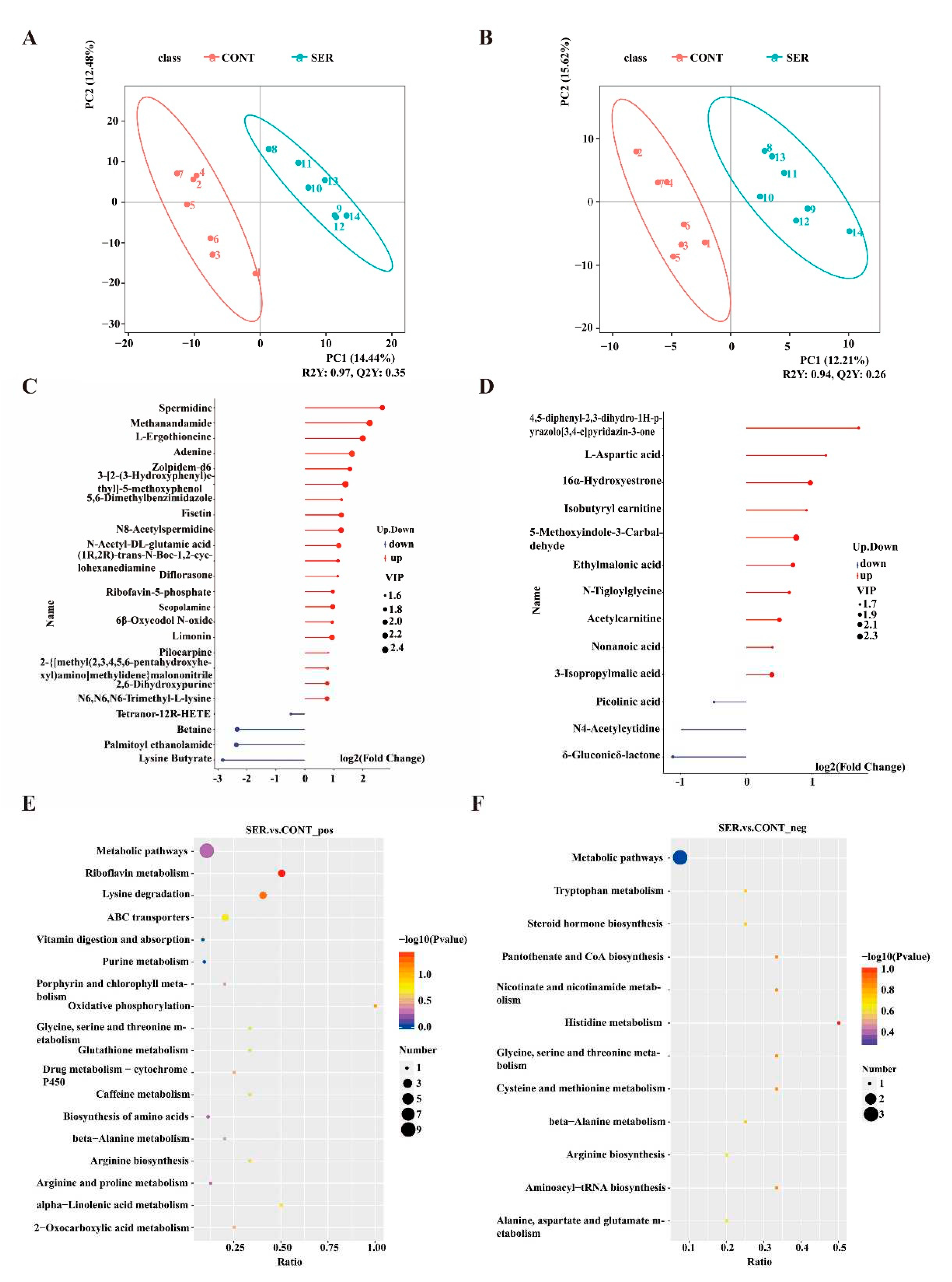
Untargeted metabolomics was used to explore differences in fecal metabolite composition. The data were processed using partial least squares discriminant analysis (PLS-DA), visually demonstrating an obvious discrepancy between pigs in the CONT and SER groups (Figure 3A,B). The threshold was constructed as VIP > 1.0, fold change (FC) > 1.2, or FC < 0.833 and P < 0.05 to obtain the differentially expressed metabolites (Figure 3C,D). For the positive ion, significant differences were detected in 24 metabolites, with 20 metabolites increased and four metabolites decreased. For the negative ion, remarkable differences were observed in 13 metabolites, with 10 increased and 3 decreased.
KEGG pathway annotation and enrichment analysis revealed the affected pathways by serine supplementation. As shown in the KEGG enrichment map, apparent differences were observed between the CONT and SER groups (Figure 3E,F). In the positive ion mode, differential metabolites were mainly enriched in riboflavin metabolism; lysine degradation; oxidative phosphorylation; glycine, serine, and threonine metabolism; glutathione metabolism; amino acid biosynthesis; arginine biosynthesis; and alpha-linolenic acid metabolism. In negative ion mode, differential metabolism was mainly enriched in tryptophan metabolism, glycine, serine, and threonine metabolism, steroid hormone biosynthesis, pantothenate and CoA biosynthesis, alanine, aspartate, and glutamate metabolism, histidine metabolism, arginine biosynthesis, cysteine and methionine metabolism, and aminoacyl-tRNA biosynthesis.
Figure 3.
The composition of fecal metabolites. Partial least-square discriminant analysis (PLS-DA) of positive (A) and negative (B) ion mode. Differential metabolites stem graph in the positive (C) and negative (D) ion mode. KEGG pathway analysis of metabolism in positive (E) and negative (F) ion mode between treatment groups. CONT, pigs fed a basal diet. SER, pigs fed a basal diet supplemented with 0.15% serine.
Figure 3.
The composition of fecal metabolites. Partial least-square discriminant analysis (PLS-DA) of positive (A) and negative (B) ion mode. Differential metabolites stem graph in the positive (C) and negative (D) ion mode. KEGG pathway analysis of metabolism in positive (E) and negative (F) ion mode between treatment groups. CONT, pigs fed a basal diet. SER, pigs fed a basal diet supplemented with 0.15% serine.
Correlation between microbiota and metabolites
Pearson’s correlation analysis was performed to explore the correlation between the top five differentially expressed fecal microbes and the top 10 differentially expressed fecal metabolites between the CONT and SER groups. The correlation Sankey diagram analysis clearly indicated a relationship between the intestinal microbiota and metabolites (Figure 4 A,B). As shown in Figure 4C, there was a significant positive correlation between Clostridium_sensu_stricto_1 and its metabolites (in the positive ion mode) including N8-Acetylspermidine, limonin, 6-phenyl-1,2,3,4-tetrahydro-2,5-benzodiazocin-1-one, 7-Methylxanthine, L-Ergothioneine, and spermidine (P < 0.05). As shown in Figure 4D, there was a significant positive correlation between Anaerorhabdus_furcosa_group and metabolites (in the negative ion mode) including16-Hydroxyestrone, 5-Methoxyindole-3-Carbaldehyde, Ethylmalonic acid, and L-aspartic acid (P < 0.05). Clostridium_sensu_stricto_1 and UCG-009 were also positively correlated with certain differential metabolites, including 6-Hydroxyestrone and 3-Isopropylmalic acid, respectively (P < 0.05). Phascolarctobacterium and Lactobacillus were negatively correlated with the differential metabolites, as shown in Figure 4C,D (P < 0.05).
Discussion
Serine is beneficial for anti-inflammatory action and antioxidation and can influence the composition of intestinal microbes, lipid metabolism, and protection of the intestinal mucosa [22]. In this study, 0.15% serine was added to the diet of finishing pigs to explore its beneficial effects. Serine increased growth performance, promoted meat quality, characterized by higher IMF content, less drip loss, enhanced antioxidant capability, and altered the composition of intestinal microbes and metabolites in growing-finishing pigs. Correlation analysis indicated a significant correlation between Clostridium_sensu_stricto_1 and Lactobacillus and certain lipid metabolites. These results indicated that serine might increase growth performance and meat quality by influencing intestinal microbes and their metabolism in growing-finishing pigs.
The IMF and drip loss are two critical indices for evaluating meat quality. We previously found that pigs had higher IMF content when a suitable ratio of serine to glycine was maintained in their diet [13]. In this study, dietary supplementation with serine alone increased IMF content, further suggesting that serine may positively affect IMF deposition. Previous studies have demonstrated the role of serine in lipid metabolism [23], and serine deficiency increases fat deposition in the liver [14]. Serine participates critically in one-carbon metabolism and promotes NADPH synthesis, which is involved in the regulation of lipid metabolism [24]. We previously found that serine activates mTOR signaling, which promotes lipogenesis [25,26].
Increased antioxidant capability can improve the integrity of the cell membrane, which helps cells hold water [27]. Serine can significantly enhance antioxidant capability by acting as an indirect precursor of GSH to promote GSH synthesis [28]. In this study, we further confirmed that serine supplementation increased serum GSH content in growing-finishing pigs. Moreover, the content of other antioxidant enzymes, such as SOD, also increased, and the content of MDA, a biomarker of oxidative stress [29], decreased, illustrating that pigs supplemented with serine had better antioxidative capability. These pigs also exhibited lower meat drip loss. Thus, serine may decrease drip loss via its strong ability to maintain the oxidative balance.
A variety of dynamic microbiota exist in the enteric canal and are mainly associated with meat quality by regulating lipid metabolism [30]. A previous study showed that serine alters the alpha and beta diversity of gut microbes [17]. Similarly, in this study, serine supplementation altered the β-diversity of intestinal microbiota in growing-finishing pigs, which indicated that serine could modulate the composition of gut microbes. Moreover, our results showed that Streptococcus was the predominant bacterial genus in growing-finishing pigs. The relative abundances of Lactobacillus and Streptococcus increased, whereas the relative abundances of Terrisporobacter and Clostridium_sensu_stricto_1 were decreased in the intestines of growing-finishing pigs supplemented with serine. Notably, the relative abundances of Lactobacillus and Prevotellaceae UCG-009, which are positively correlated with IMF content [31], were increased by dietary supplementation with serine. Therefore, the alteration of gut microbes caused by serine supplementation may also be one of the reasons for the increased IMF content.
Significant differences were observed in the metabolites among pigs in different groups according to the metabolomic results. The metabolites that differed between the two groups were mainly lipids and lipid-like molecules, including acetyl-carnitine, ethylmalonic acid, nonanoic acid, organic acids, and derivatives, including N8-acetylspermidine and L-aspartic acid. Among these differential metabolites, acetylcarnitine can promote fat synthesis by providing acetyl groups to several substances, including acetyl-CoA [30]. Ethylmalonic acid participates in fatty acid synthesis by acting as a cofactor for ethylmalonyl-CoA decarboxylase [32]. The level of l-aspartic acid, which is increased by serine supplementation, positively correlates with the IMF content [33,34]. Moreover, the differentially expressed metabolites were mostly enriched in metabolic pathways related to lipid biosynthesis, including alpha-linolenic acid metabolism, tryptophan metabolism [35,36], and steroid hormone biosynthesis. It is not only intestinal microbes that affect IMF content but also their metabolites [37]. Thus, enhanced metabolite content and metabolic pathways associated with lipid metabolism may be other factors promoting IMF accumulation in pigs supplemented with serine. Importantly, the correlation analysis between microbes and metabolites indicated that the alteration of metabolites might be related to changes in the intestinal microbiota, as microbes, including Anaerorhabdus_furcosa_group, Clostridium_sensu stricto_1, Lactobacillus, Phascolarctobacterium and UCG-009, were significantly altered, and most of them were strongly linked with metabolites associated with lipid metabolism. Nevertheless, further studies are needed to explore whether serine directly or indirectly influences meat quality by modulating the intestinal composition of microbes and their metabolites.
In summary, serine supplementation in diets significantly increased the growth performance, meat quality, and antioxidant ability of growing-finishing pigs. Additionally, serine affects intestinal microbiota composition and their metabolites, which may mediate its advantageous effects on growing-finishing pigs. Consequently, our results suggest that serine can serve as an additive to improve the meat quality of growing-finishing pigs.
- Animal Welfare Statement:The study was conducted according to the principles of the animal welfare committee of the Institute of Subtropical Agriculture, Chinese Academy of Sciences and was approved by the animal welfare committee of the Institute of Subtropical Agriculture, Chinese Academy of Sciences.
- Informed Consent Statement: Informed consent was obtained from all individual participants included in the study.
- Data Availability Statement: The 16S rDNA gene sequence data have been deposited in the NCBI Bioproject database (https://www.ncbi.nlm.nih.gov/bioproject/), registration number for PRJNA994863.
- Acknowledgments: This work was supported by the Hunan Provincial Science and Technology Department(2021JJ30320), Science and Technology Innovation Program of Hunan Province (2023RC1074), Youth Innovation Promotion Association CAS and China Agriculture Research System of MOF and MARA (CARS-35).
References
- Liu, B.; Chen, Y.; Li, Q.; Zhong, Z.; Tan, Y.; Zhang, S.; Zhu, L. Effects of Breed and Gender Effect on Pork Quality Traits. Southwest China Journal of Agricultural Sciences 2019, 32, 2222–2225. [Google Scholar]
- Wang, Y.; Thakali, K.; Morse, P.; Shelby, S.; Chen, J.; Apple, J.; Huang, Y. Comparison of Growth Performance and Meat Quality Traits of Commercial Cross-Bred Pigs versus the Large Black Pig Breed. Animals (Basel) 2021, 11. [Google Scholar] [CrossRef]
- Chen, B.L.; Li, D.Y.; Leng, D.; Kui, H.; Bai, X.; Wang, T. Gut microbiota and meat quality. Frontiers in Microbiology 2022, 13. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Wang, Y.; Huang, Z.; Su, Y.; Li, F.; Yin, J. Research Progress of Nutritional Regulation for Meat Quality of Pigs. Chinese Journal of Animal Nutrition 2020, 32, 4555–4564. [Google Scholar]
- Luo, J.; Zeng, D.; Cheng, L.; Mao, X.; Yu, J.; Yu, B.; Chen, D. Dietary beta-glucan supplementation improves growth performance, carcass traits and meat quality of finishing pigs. Anim Nutr 2019, 5, 380–385. [Google Scholar] [CrossRef] [PubMed]
- Tian, Z.; Deng, D.; Cui, Y.; Chen, W.; Yu, M.; Ma, X. Diet supplemented with fermented okara improved growth performance, meat quality, and amino acid profiles in growing pigs. Food Sci Nutr 2020, 8, 5650–5659. [Google Scholar] [CrossRef]
- Ma, X.; Yu, M.; Liu, Z.; Deng, D.; Cui, Y.; Tian, Z.; Wang, G. Effect of amino acids and their derivatives on meat quality of finishing pigs. J Food Sci Technol 2020, 57, 404–412. [Google Scholar] [CrossRef]
- Wu, L.; Zhang, H.; Na, L.; Zhou, X.; Li, X.; Zhao, Y.; Wen, Z.; He, Q. Methionine restriction at the post-weanling period promotes muscle fiber transition in piglets and improves intramuscular fat content in growing-finishing pigs. Amino Acids 2019, 51, 1657–1666. [Google Scholar] [CrossRef]
- Hu, C.J.; Jiang, Q.Y.; Zhang, T.; Yin, Y.L.; Li, F.N.; Deng, J.P.; Wu, G.Y.; Kong, X.F. Dietary supplementation with arginine and glutamic acid modifies growth performance, carcass traits, and meat quality in growing-finishing pigs. Journal of animal science 2017, 95. [Google Scholar] [CrossRef]
- He, L.; Long, J.; Zhou, X.; Liu, Y.; Li, T.; Wu, X. Serine is required for the maintenance of redox balance and proliferation in the intestine under oxidative stress [J]. FASEB J 2020, 34, 4702–4717. [Google Scholar] [CrossRef]
- He, L.; Ding, Y.; Zhou, X.; Li, T.; Yin, Y. Serine signaling governs metabolic homeostasis and health. Trends Endocrinol Metab 2023, 34, 361–372. [Google Scholar] [CrossRef]
- Lionaki, E.; Gkikas, I.; Daskalaki, I.; Ioannidi, M.-K.; Klapa, M.I.; Tavernarakis, N. Mitochondrial protein import determines lifespan through metabolic reprogramming and de novo serine biosynthesis. Nature Communications 2022, 13. [Google Scholar] [CrossRef]
- Zhou, X.; Liu, Y.; Zhang, L.; Kong, X.; Li, F. Serine-to-glycine ratios in low-protein diets regulate intramuscular fat by affecting lipid metabolism and myofiber type transition in the skeletal muscle of growing-finishing pigs. Anim Nutr 2021, 7, 384–392. [Google Scholar] [CrossRef]
- He, L.; Liu, Y.; Liu, D.; Feng, Y.; Yin, J.; Zhou, X. Exogenous and Endogenous Serine Deficiency Exacerbates Hepatic Lipid Accumulation. Oxid Med Cell Longev 2021, 2021, 4232704. [Google Scholar] [CrossRef]
- Nguyen Cong, O.; Bernard, T.; Pham Kim, D.; Do Duc, L.; Nassim, M.; Nguyen Thi, H.; Nguyen Hoang, T.; Georges, D.; Jerome, B.; Vu Dinh, T.; et al. Growth performance, carcass quality characteristics and colonic microbiota profiles in finishing pigs fed diets with different inclusion levels of rice distillers' by-product [J]. Animal science journal = Nihon chikusan Gakkaiho 2019, 90, 948–960. [Google Scholar] [CrossRef] [PubMed]
- He, L.; Guo, J.; Wang, Y.; Wang, L.; Xu, D.; Yan, E.; Zhang, X.; Yin, J. Effects of Dietary Yeast beta-Glucan Supplementation on Meat Quality, Antioxidant Capacity and Gut Microbiota of Finishing Pigs [J]. Antioxidants (Basel) 2022, 11. [Google Scholar] [CrossRef]
- Zhang, H.; Hua, R.; Zhang, B.; Zhang, X.; Yang, H.; Zhou, X. Serine Alleviates Dextran Sulfate Sodium-Induced Colitis and Regulates the Gut Microbiota in Mice [J]. Front Microbiol 2018, 9, 3062. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.B.; Du, W.; Fu, A.K.; Zhang, X.P.; Huang, Y.; Lee, K.H.; Yu, K.; Li, W.F.; Li, Y.L. Intestinal microbiota and oral administration of Enterococcus faecium associated with the growth performance of new-born piglets. Benef Microbes 2016, 7, 529–538. [Google Scholar] [CrossRef] [PubMed]
- Long, C.; Zhou, X.; Wang, Q.; Xie, C.; Li, F.; Fan, Z.; Zhang, B.; Ruan, Z.; Chen, X.; Wu, X.; et al. Dietary supplementation of Lonicera macranthoides leaf powder improves amino acid profiles in serum and longissimus thoracis muscle of growing-finishing pigs. Anim Nutr 2016, 2, 271–275. [Google Scholar] [CrossRef] [PubMed]
- Driessen, B.; Van Beirendonck, S.; Buyse, J. The Impact of Grouping on Skin Lesions and Meat Quality of Pig Carcasses. Animals (Basel) 2020, 10. [Google Scholar] [CrossRef]
- Henriquez-Rodriguez, E.; Tor, M.; Pena, R.N.; Estany, J. A polymorphism in the stearoyl-CoA desaturase gene promoter increases monounsaturated fatty acid content in dry-cured ham. Meat Sci 2015, 106, 38–43. [Google Scholar] [CrossRef]
- He, Y.; Liu, Y.; Guan, P.; He, L.; Zhou, X. Serine Administration Improves Selenium Status, Oxidative Stress, and Mitochondrial Function in Longissimus Dorsi Muscle of Piglets with Intrauterine Growth Retardation. Biol Trace Elem Res 2023, 201, 1740–1747. [Google Scholar] [CrossRef]
- Ogbuagu, N.E.; Ayo, J.O.; Aluwong, T.; Akor-Dewu, M.B. L-serine improves lipid profile, performance, carcass weight and intestinal parameters in feed restricted broiler chickens during the hot-dry season. Trop. Anim. Health Prod. 2022, 54, 12. [Google Scholar] [CrossRef]
- Lewis, Caroline A.; Parker, Seth J.; Fiske, Brian P.; McCloskey, D.; Gui, Dan Y.; Green, Courtney R.; Vokes, Natalie I.; Feist, Adam M.; Vander Heiden, Matthew G.; Metallo, Christian M. Tracing Compartmentalized NADPH Metabolism in the Cytosol and Mitochondria of Mammalian Cells. Molecular Cell 2014, 55, 253–263. [Google Scholar] [CrossRef]
- Chen, S.; Xia, Y.; He, F.; Fu, J.; Xin, Z.; Deng, B.; He, L.; Zhou, X.; Ren, W. Serine Supports IL-1β Production in Macrophages Through mTOR Signaling. Frontiers in immunology 2020, 11. [Google Scholar] [CrossRef]
- Mao, Z.; Zhang, W. Role of mTOR in Glucose and Lipid Metabolism. International journal of molecular sciences 2018, 19. [Google Scholar] [CrossRef]
- Jiang, J.; Jin, F.; Lin, G.; Xiong, Y.L. Modulation of muscle antioxidant enzymes and fresh meat quality through feeding peptide-chelated trace minerals in swine production. Food Bioscience 2021, 42. [Google Scholar] [CrossRef]
- Zhou, X.; He, L.; Wu, C.; Zhang, Y.; Wu, X.; Yin, Y. Serine alleviates oxidative stress via supporting glutathione synthesis and methionine cycle in mice. Mol Nutr Food Res 2017, 61. [Google Scholar] [CrossRef]
- Wang, Y.; Li, Y.; Xie, J.; Zhang, Y.; Wang, J.; Sun, X.; Zhang, H. Protective effects of probiotic Lactobacillus casei Zhang against endotoxin- and d-galactosamine-induced liver injury in rats via anti-oxidative and anti-inflammatory capacities. Int Immunopharmacol 2013, 15, 30–37. [Google Scholar] [CrossRef] [PubMed]
- Yin, J.; Li, Y.; Tian, Y.; Zhou, F.; Ma, J.; Xia, S.; Yang, T.; Ma, L.; Zeng, Q.; Liu, G.; et al. Obese Ningxiang pig-derived microbiota rewires carnitine metabolism to promote muscle fatty acid deposition in lean DLY pigs. Innovation (Camb) 2023, 4, 100486. [Google Scholar] [CrossRef] [PubMed]
- Tang, S.; Xin, Y.; Ma, Y.; Xu, X.; Zhao, S.; Cao, J. Screening of Microbes Associated With Swine Growth and Fat Deposition Traits Across the Intestinal Tract [J]. Frontiers in Microbiology 2020, 11. [Google Scholar] [CrossRef]
- Izzo, L.T.; Trefely, S.; Demetriadou, C.; Drummond, J.M.; Mizukami, T.; Kuprasertkul, N.; Farria, A.T.; Nguyen, P.T.T.; Murali, N.; Reich, L.; et al. Acetylcarnitine shuttling links mitochondrial metabolism to histone acetylation and lipogenesis. Sci. Adv. 2023, 9, 20. [Google Scholar] [CrossRef]
- Chen, Z.; Sun, Y.; Chen, L.; Zhang, Y.; Wang, J.; Li, H.; Yan, X.; Xia, L.; Yao, G. Differences in meat quality between Angus cattle and Xinjiang brown cattle in association with gut microbiota and its lipid metabolism. Frontiers in Microbiology 2022, 13. [Google Scholar] [CrossRef]
- Wang, B.; Wang, Y.J.; Zuo, S.X.; Peng, S.J.; Wang, Z.J.; Zhang, Y.J.; Luo, H.L. Untargeted and Targeted Metabolomics Profiling of Muscle Reveals Enhanced Meat Quality in Artificial Pasture Grazing Tan Lambs via Rescheduling the Rumen Bacterial Community. J. Agric. Food Chem. 2021, 69, 846–858. [Google Scholar] [CrossRef] [PubMed]
- Goodarzi, P.; Habibi, M.; Roberts, K.; Sutton, J.; Shili, C.N.; Lin, D.; Pezeshki, A. Dietary Tryptophan Supplementation Alters Fat and Glucose Metabolism in a Low-Birthweight Piglet Model. Nutrients 2021, 13. [Google Scholar] [CrossRef] [PubMed]
- Xiao, P.; Goodarzi, P.; Pezeshki, A.; Hagen, D.E. RNA-seq reveals insights into molecular mechanisms of metabolic restoration via tryptophan supplementation in low birth weight piglet model. Journal of animal science 2022, 100. [Google Scholar] [CrossRef] [PubMed]
- Zubiri-Gaitán, A.; Blasco, A.; Hernández, P. Plasma metabolomic profiling in two rabbit lines divergently selected for intramuscular fat content. Communications Biology 2023, 6. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Bacteria community variety and richness. (A) Venn diagram. (B) Chao1 index. Relative abundance of top 10 microbiota community at class level (C) and genus level (D). CONT, pigs fed a basal diet. SER, pigs fed a basal diet supplemented with 0.15% serine.
Figure 1.
Bacteria community variety and richness. (A) Venn diagram. (B) Chao1 index. Relative abundance of top 10 microbiota community at class level (C) and genus level (D). CONT, pigs fed a basal diet. SER, pigs fed a basal diet supplemented with 0.15% serine.
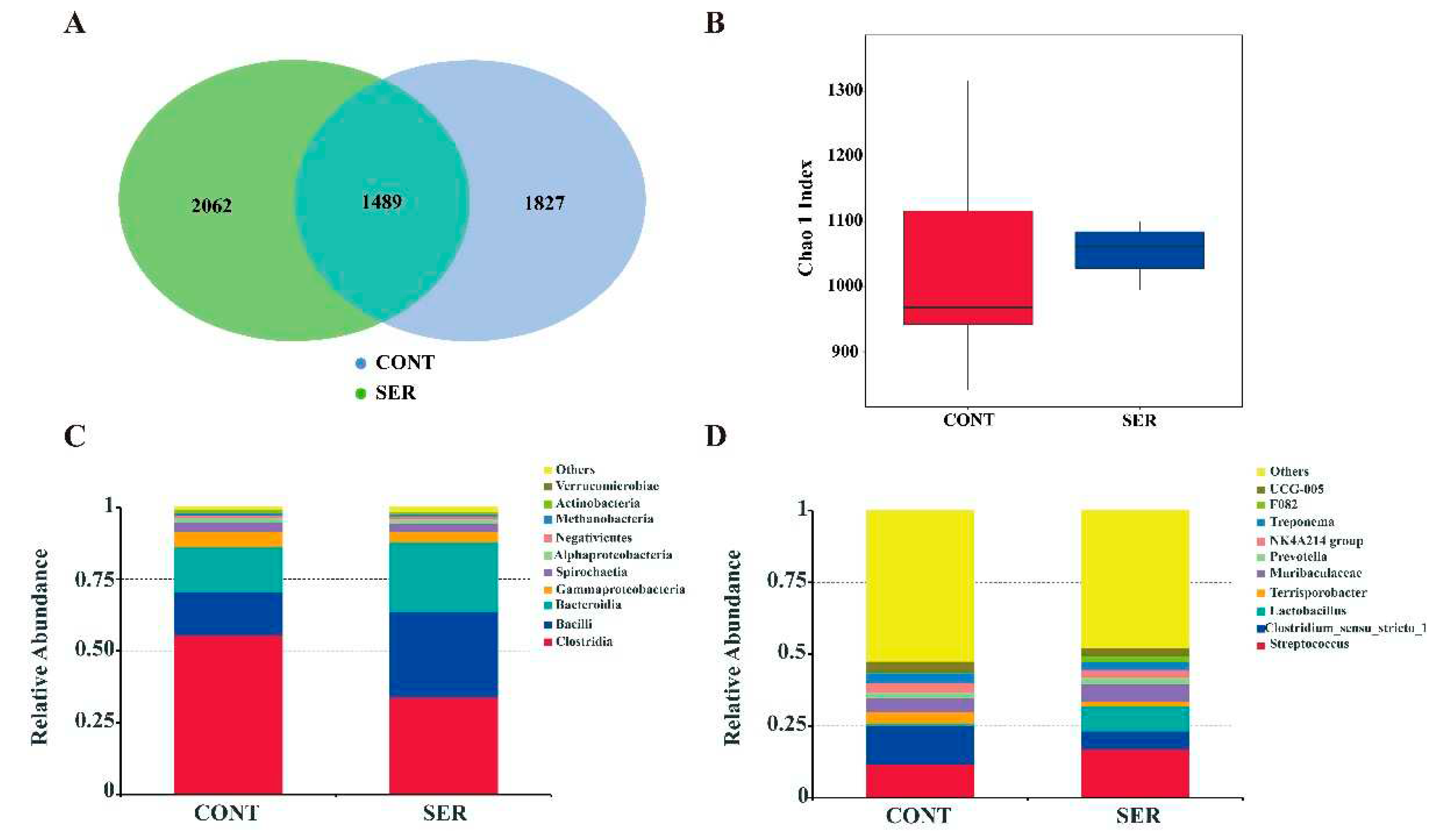
Figure 2.
Microbial β-diversity and different microbes between groups based on t-Test analysis. Principal Coordinate Analysis (PCoA) based on the jaccard distance matrix (A). Box plot, displayed by weighted Unifrac t-Test (B). *P < 0.05. Different microbes between groups at genus (C) and species (D) level, analyzed by T test. CONT, pigs fed a basal diet. SER, pigs fed a basal diet supplemented with 0.15% serine.
Figure 2.
Microbial β-diversity and different microbes between groups based on t-Test analysis. Principal Coordinate Analysis (PCoA) based on the jaccard distance matrix (A). Box plot, displayed by weighted Unifrac t-Test (B). *P < 0.05. Different microbes between groups at genus (C) and species (D) level, analyzed by T test. CONT, pigs fed a basal diet. SER, pigs fed a basal diet supplemented with 0.15% serine.
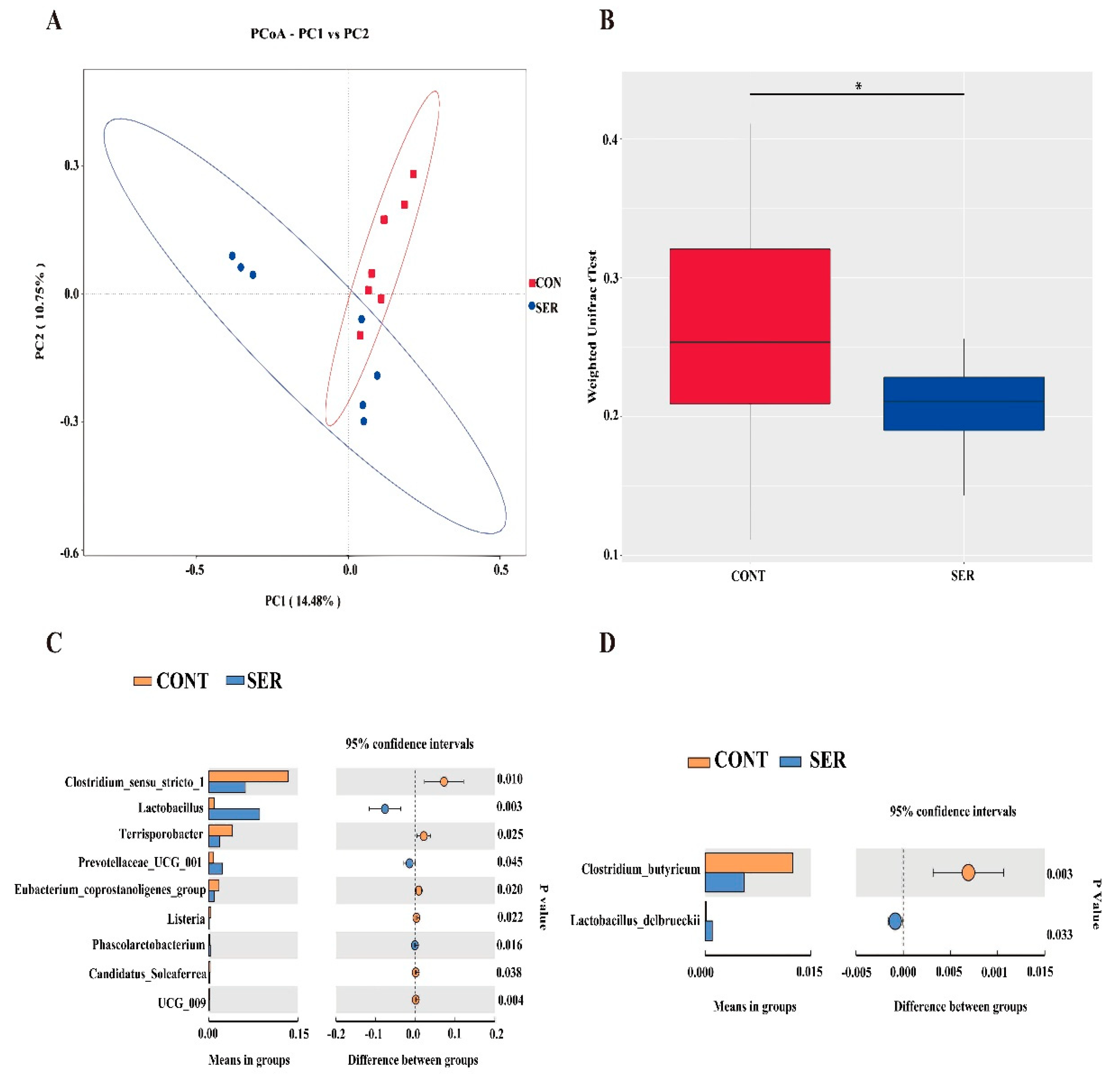
Figure 4.
Correlations between intestinal microbiota and metabolites. Sankey diagram of differential intestinal microbiota in the positive (A) and negative (B) ion mode. The correlation heat map of differential intestinal microbiota and metabolites in the positive (C) and negative (D) mode. *P < 0.05.
Figure 4.
Correlations between intestinal microbiota and metabolites. Sankey diagram of differential intestinal microbiota in the positive (A) and negative (B) ion mode. The correlation heat map of differential intestinal microbiota and metabolites in the positive (C) and negative (D) mode. *P < 0.05.
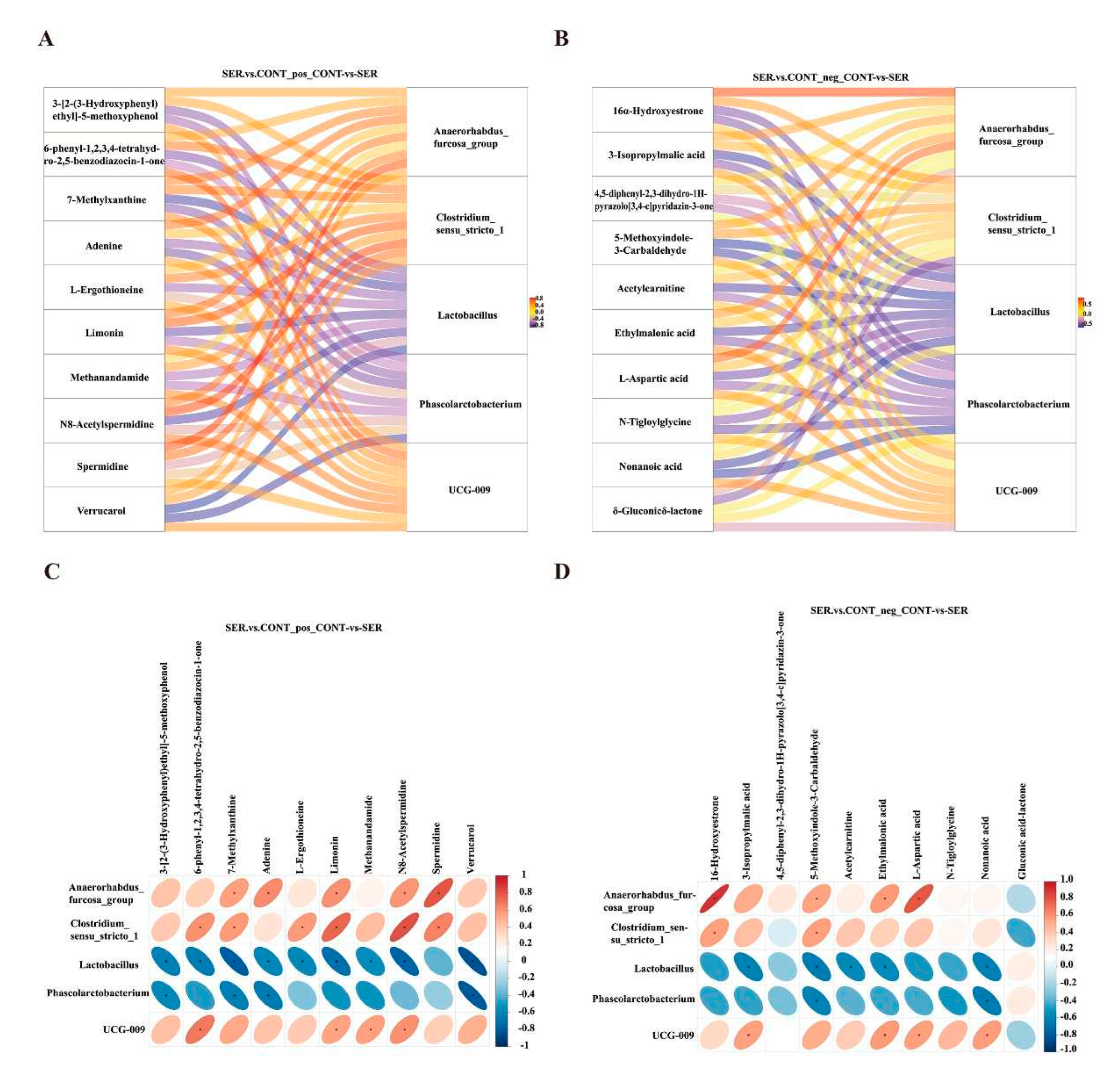

Table 1.
Diet composition and nutrient level.
| Item | 20~30 kg | 30~60kg | 60~120 kg |
|---|---|---|---|
| Corn, % | 65.00 | 72.30 | 59.00 |
| Wheat bran, % | 6.00 | 3.00 | / |
| Rice, % | / | / | 20.00 |
| Soybean meal#, % | 25.00 | 19.50 | 14.00 |
| Soybean oil, % | / | 1.20 | 3.00 |
| Limestone, % | 1.00 | 0.80 | 0.80 |
| calcium hydrogen phosphate, % | 0.80 | 0.70 | 0.50 |
| NaCl, % | 0.40 | 0.45 | 0.30 |
| L-lysine hydrochloride, % | 0.30 | 0.60 | 0.25 |
| DL-methionine, % | 0.10 | 0.10 | / |
| L-threonine, % | 0.20 | 0.20 | / |
| L-tryptophan, % | / | 0.03 | / |
| Zeolite powder | 0.08 | / | 1.03 |
| Premix*, % | 1.12 | 1.12 | 1.12 |
| Total, % | 100 | 100 | 100 |
| Calculated nutrient level | |||
| DE(MJ/kg) | 13.86 | 14.28 | 14.03 |
| CP, % | 17.51 | 15.57 | 12.55 |
| Ca, % | 0.64 | 0.53 | 0.46 |
| Total P, % | 0.50 | 0.43 | 0.38 |
| P, % | 0.23 | 0.20 | 0.17 |
| Lysine, % | 1.03 | 1.12 | 0.71 |
| Methionine, % | 0.36 | 0.33 | 0.2 |
| Threonine, % | 0.73 | 0.65 | 0.37 |
| Tryptophan, % | 0.15 | 0.15 | 0.10 |
CP = crude protein; DE = digestible energy. #Soybean meal contains 43% protein. *Provided for per kilogram of diet: Vitamin A, 13,000 IU; Vitamin D3, 4,000 IU; Vitamin E, 32 IU; Vitamin K, 4 mg; Vitamin B1, 4 mg; Vitamin B2, 10 mg; Vitamin B6, 6 mg; Vitamin B12, 6 mg; Vitamin B3,48 mg; Vitamin B5, 24 mg; folic acid 2 mg; biotin 0.2 mg; FeSO4 180 mg; CuSO4 12 mg; ZnSO4 140 mg; MnSO4 8 mg; Ca(IO3)2 0.4 mg; Na2SeO3 0.2 mg.
Table 2.
Growth performance of growing-finishing pigs.
| CONT | SER | |
|---|---|---|
| Initial BW, kg | 20.95±0.76 | 20.35±0.38 |
| Final BW, kg | 116.8±7.1 | 125.2±4.9 |
| ADG, kg | 0.806±0.059a | 0.871±0.026b |
| ADFI, kg | 2.25±0.16 | 2.29±0.04 |
| F/G | 2.80±0.05 | 2.63±0.10 |
a,b indicates a significant difference between the treatment groups (P < 0.05), n=6. Data were shown as Mean ± SD. CONT, pigs fed a basal diet. SER, pigs fed a basal diet supplemented with 0.15% serine. Initial BW, initial body weight; final BW, final body weight; ADG, average daily gain; ADFI, average daily feed intake; F/G, the ratio of feed to gain.
Table 3.
Serum biochemical parameters of growing-finishing pigs.
| CONT | SER | |
|---|---|---|
| GSH, ng/mL | 129.4±17.4a | 196.9±19.1b |
| SOD, ng/mL | 197.6±29.3a | 286.4±17.2b |
| MDA, nmol/mL | 4.06±0.40a | 1.91±0.24b |
a, b indicates a significant difference between the treatment groups (P < 0.05), n=6. Data were shown as Mean ± SD. CONT, pigs fed a basal diet. SER, pigs fed a basal diet supplemented with 0.15% serine. GSH, glutathione; SOD, superoxide dismutase; MDA, malondialdehyde.
Table 4.
Meat quality traits of the growing-finishing pigs.
| CONT | SER | |
|---|---|---|
| Color | ||
| L | 45.23±1.56 | 44.11±1.28 |
| a | 13.86±0.33 | 14.48±0.48 |
| b | 5.40±0.57 | 5.08±0.46 |
| pH45min | 6.60±0.09 | 6.42±0.11 |
| pH24h | 5.60±0.07 | 5.53±0.07 |
| Drip loss | 3.58±0.50a | 2.24±0.33b |
| Intramuscular fat | 2.26±0.18a | 2.94±0.17b |
a, b indicates a significant difference between the treatment groups (P < 0.05), n=6. Data were shown as Mean ± SD. CONT, pigs fed a basal diet. SER, pigs fed a basal diet supplemented with 0.15% serine.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



