Submitted:
25 December 2023
Posted:
26 December 2023
You are already at the latest version
Abstract
Intestinal mucositis (IM) is a common adverse effect of chemotherapy, limiting its clinical application. Edible Chinese medicine, specifically Codonopsis pilosula-derived CP-A, with anti-inflammatory and gastrointestinal protective effects. The present work was to investigate CP-A's role in ameliorating IM and its mechanism using in vitro and rat models. Western blot, immunohistochemical (IHC) and real-time PCR (RT-PCR) analyses were used to assess protein expression related to the extracellular-regulated protein kinases (ERK)/myosin light chain kinase (MLCK)/myosin light chain 2 (MLC2) signaling pathway and tight junction proteins. Enzyme-linked immunosorbent assay (ELISA) was conducted to measure inflammatory factors, and 16S rRNA Amplicon Sequencing was employed for cecum content analysis. The results indicated that CP-A could restore body weight, food intake and histopathological changes in IM rats. Besides, abnormal MLCK activation induced by 5-fluorouracil (5-FU) was attenuated by CP-A via the ERK/MLCK/MLC2 pathway. CP-A treatment improved tight junction protein levels and reduced inflammatory factor expression. On the other hand, intestinal flora experiment demonstrated that CP-A intervention could increase the abundance of lactobacillus. In conclusion, CP-A mitigates 5-FU-induced IM by inhibiting the ERK/MLCK/MLC2 pathway, reducing the expression of inflammatory factors, improving the intestinal mucosal barrier and regulating intestinal flora. This study sheds light on CP-A's therapeutic potential in IM treatment, providing insights for future research.
Keywords:
inulin-type fructan CP-A
; intestinal mucositis
; inflammation
; ERK/MLCK/MLC2 signaling pathway
; mucosal barrier
; intestinal microbiota
1. Introduction
Many cancers, including stomach, esophageal, breast and head cancer, rely on 5-FU treatment [1,2]. 5-FU is the first-line treatment for colorectal cancer [3,4]. However, the use of 5-FU can result in severe adverse reactions, such as IM [5]. The clinical manifestations of IM include diarrhea, nausea and vomiting, which cause intense discomfort to patients and seriously limit the use of chemotherapeutic drugs [6,7]. Unfortunately, there are currently no effective medications for alleviating the pain associated with intestinal mucositis, therefore, it is necessary to identify and develop treatments that can effectively alleviate the symptoms and enhance the quality of life for patients undergoing chemotherapy. 5-FU exhibits cytotoxic effects on intestinal epithelial cells, resulting in damage to the intestinal mucosal barrier and the release of pro-inflammatory factors [8]. Tight junction proteins are essential components of the intestinal barrier [9]. The primary mode through which 5-FU reaches the intestinal mucosal barrier is by degrading the tight junction proteins [10]. In addition, 5-FU disrupts the intestinal flora [11,12]. These factors offer potential avenues for addressing IM.
Proinflammatory factors play an important role in the occurrence and development of inflammation [13,14]. Overexpression of tumor necrosis factor-alpha (TNF-α) and interferon gamma (IFN-γ) is often detected in cases of IM [15,16]. ERK and MLCK play central roles in the protection of intestinal epithelial tight junction [17]. By promoting the phosphorylation of MLC2, MLCK is activated by Phospho-ERK1/2, leading to the destruction of intestinal tight junction and degradation of tight junction proteins such as Claudin-1, Occludin, F-actin and Zonula occludens-1 (ZO-1) [18,19]. TNF-α activation of ERK1/2 promotes ERK phosphorylation, thus activating the ERK/MLCK/MLC2 pathway [19]. In addition, the overexpression of IFN-γ is involved in the activation of MLCK which up-regulate the expression of Claudin-2 and promote interleukin (IL)-13 release [20]. Once the intestinal barrier is breached, an increase in intestinal epithelium permeability permits endotoxins and macromolecules to enter the cells freely, causing the release of IL-6, IL-8, TNF-α and IFN-γ, which will aggravate inflammatory response [18]. Moreover, the destruction of the intestinal mucosal barrier is accompanied by disturbances in the intestinal flora [9,21]. Therefore, the inhibition of TNF-α and IFN-γ overexpression and the regulation of ERK/MLCK/MLC2 pathway protect the intestinal mucosal barrier and reduce inflammation. A stable and healthy microbiome is essential for the proper functioning of the gut [8]. Many studies have indicated that chemotherapy-induced gastrointestinal diseases are often accompanied by disturbance of intestinal flora [10]. Dysregulation of intestinal flora leads to disruption of intestinal homeostasis and intestinal mucus barrier integrity [9,11]. There is increasing evidence that 5-FU-induced IM can cause the imbalance of intestinal flora, the abundance of harmful bacteria increases and the abundance of probiotics decreases [10,12]. Thus, regulating intestinal flora and increasing the abundance of probiotics may be a potential treatment for 5-FU-induced IM.
Traditional Chinese Medicine (TCM) and natural products extracted from TCM have a long history of clinical application in the treatment of various diseases, including IM [22]. Codonopsis pilosula, an edible herbal medicine widely used for improving poor gastrointestinal function in traditional Chinese medicine, is derived from the roots of Codonopsis pilosula (Franch.) Nanf., Codonopsis pilosula (Franch.) Nannf. var. modesta (Nannf.) L.T. Shen and Codonopsis tangshen Oliv. In recent days, the edible plant has been listed by National Health Commission of China as a substance that is both food and traditional Chinese medicine. Over the past years, Codonopsis pilosula, the plant homologous to food and medicine is known for its anti-ulcerative colitis effects, its role in regulating intestinal flora, and its ability to decrease inflammatory factor levels [23,24]. The main active constituents of Codonopsis pilosula are polysaccharides, lignans and polyacetylenes [25]. Additionally, the anti-inflammatory effects of polysaccharides from Codonopsis pilosula have been reported [24,26]. CP-A is an inulin-type fructan and represents the most active ingredient in Codonopsis pilosulae polysaccharides. Nevertheless, research on IM treatment is rare. Meanwhile, it’s in vivo and in vitro anti-IM mechanisms remain unclear. Hence, this study selected IEC-6 cells and Sprague-Dawley (SD) rats as experimental subjects to investigate the mechanism of CP-A treatment for 5-FU-induced chemotherapeutic enteritis. This study will provide valuable insights for the development of improved treatments for IM, addressing a critical need in clinical oncology.
2. Materials and Methods
2.1. Chemicals and Reagents
CP-A (Figure 1a, MW: 3.6kDa), an inulin-type fructan, was isolated from Codonopsis pilosula as previously described [27]. 5-FU (F6173) was purchased from Macklin Biochemical Co., Ltd. (Shanghai, China). Bifid. Triple Viable Capsules Dissolving at Intestines (BTC) were purchased from Jincheng HEALTH Pharmaceutical Co., Ltd. (Jincheng, Shanxi, China). Primary antibodies against rat MLC2 (DF7911 1:2000), Phospho-MLC2 (Ser15 AF8618 1:1000) were purchased from Affinity Biosciences Co., Ltd. (OH, United States). MLCK (21642-1-AP 1:1000), ERK1/2 (11257-1-AP 1:5000), Phospho-ERK1/2 (Thr202/Tyr204 28733-1-AP 1:2000), Beta Actin (20536-1-AP, 1:2000), Claudin-2 (26912-1-AP, 1:1000), Occludin (27260-1-AP, 1:5000), ZO-1 (21773-1-AP, 1:5000) were purchased from Proteintech Group, Inc. (Wuhan, China). Claudin-1 (bs-1428R, 1:1000) and F-actin (bs-1571R) were obtained from Beijing Biosynthesis Biotechnology Co. Ltd. (Beijing, China). The secondary antibodies were obtained from ImmunoWay Biotechnology (TX, United States). All ELISA kits were acquired from Jiangsu Meimian Industrial Co., Ltd. (Jiangsu, China). All the other materials were of analytical grade.
2.2. Cells Culture and MTT Assay
The rat intestinal epithelial cell line IEC-6 was acquired from the BeNa Culture Collection (BNCC100548) and maintained in complete high-glucose DMEM culture medium (Thermo Fisher Gibco, MA, USA, Cat. No. C11995500BT) with 10% fetal bovine serum (FBS, AusGeneX, Australia, Cat. No. 10100147) and 1% streptomycin/penicillin (Gibco, NY, USA, Cat. No. 15240-062) at 37°C in 95% air and 5% CO2. Cell viability was assessed using the 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT) assay after treatment with different concentrations of CP-A (0, 1, 5, 10, 20, 50, 100, 200 and 500 μg/mL) for 24 h. Briefly, IEC-6 cells (2.0 × 103 cells/well) in the logarithmic (log) growth phase were inoculated into 96-well plates (100 μL/well) and made to adhere for 24 h. Thereafter, the medium was substituted with CP-A and treated for 24 h. Then, the different concentrations of CP-A were substituted with MTT (100 μL) to each well at a final concentration of 5 mg/mL and incubated for 4 h. Subsequently, removing the medium and Dimethyl sulfoxide (DMSO, 100 μL) was added to each well. After shaking the plates for 10 min, cell viability was evaluated at 570 nm. The MTT assay was repeated three times to ensure data reproducibility.
2.3. Total RNA Extraction and Quantitative Real-Time PCR
IEC-6 cells were seeded in 6-well plates (2.0 × 105 cells/well) and cultured for 24 h to adhere. Cells were treated with different concentrations of CP-A for 1 h and then incubated with or without 5-FU (10 μg/mL) for 24 h [28]. Total RNA was extracted from IEC-6 cells using TransZol Up reagent (Transgen, Beijing, China, Cat. No. ET111-01). RNA concentration was detected using an Eppendorf Bioluminometer D30 (Roche, Mannheim, Germany). Subsequently, a cDNA Synthesis Supermix kit (Transgen, Beijing, China, Cat. No. AT311-02) was used to reverse transcribe total RNA into cDNA according to the manufacturer's instructions. Finally, Tip Green qPCR Supermix (Transgen, Beijing, China, Cat. No. AQ141-01) was used to conduct RT-PCR analysis on a Light Cycler96 Real-Time PCR System (Roche, Mannheim, Germany). The experiment was conducted according to the reaction program (one cycle of 94 °C for 30 s, followed by 40 cycles of 94 °C for 5 s and 60 °C for 30 s). The primer sequences for the target genes, such as MLCK, ERK1/2, TNF-α, IL-4, IL-6 and IL-10 used in this study are shown in Table S1 (see Supplementary material 1). The relative quantification of all mRNA expression was performed using the 2−ΔΔCt method. GAPDH was purchased from Sangon Biotech (Shanghai, China, Cat. No. B661204-0001) as an internal reference for the normalization of the target genes.
2.4. Experimental Animals
Male SD rats [SPF grade, weighing 200±20 g, approval number: SCXK (Jing) 2019-0010] were provided by Beijing Sipeifu Biotechnology Co., Ltd. All animals used in this study were fed with 12 h light/dark cycle and allowed free access to food and clean water under a regulated condition of room temperature (20-24 °C) and relatively constant humidity. All experimental protocols were approved by the Animal Experimental Ethics Committee of the Shanxi Medical University. In accordance with the regulations on human care and use of laboratory animals, all reasonable measures were taken to minimize animal suffering.
2.5. Establishment of 5-FU-Induced Intestinal Mucositis
All rats were allowed to acclimate for a week and then randomly divided into six groups (n=12): control group, 5-FU group (50 mg/kg, intraperitoneal injection, i.p.), BTC group (250 mg/kg, intragastric administration, i.g.) and CP-A groups (15, 30, 60 mg/kg, i.g.). In the control group, rats were intraperitoneally injected with 0.8 mL 0.9% saline solution per 200g of body weight, while the other group rats were administered 5-FU over the first three days. The vehicle, BTC, or different doses of CP-A were orally administered to all rats daily during the experiment (Figure 2a). Rats were deeply anesthetized with 10% chloral hydrate 24 h after the final treatment. Blood samples were obtained from the abdominal aorta for biochemical analysis, and the cecal contents were collected to analyze the intestinal flora. Small-intestinal tissues were rapidly harvested and rinsed with cold PBS. Ileum samples of all rats (0.5 × 0.5 cm) were fixed in 4% paraformaldehyde and embedded in paraffin for subsequent experiments, while other intestinal tissues were stored at -80 °C for biochemical analysis.
2.6. Histopathological Evaluation
Paraffin blocks containing tissue sections of 5-μm-thickness were cut and stained with hematoxylin and eosin (H&E) to evaluate ileum tissue damage using an Olympus light microscope (Tokyo, Japan).
2.7. Immunohistochemical Staining
IEC-6 cells were inoculated into clean microscope cover glass placed in 6-well plates (2.0 × 105 cells/well), followed by treatment as before. The glass plates containing the cells were removed for immunohistochemical analysis.
Paraffin sections and slides of the cells were dewaxed. After blocking endogenous peroxidase activity, sections were incubated overnight at 4 °C with ZO-1, Occludin, Claudin-1, F-actin, MLCK and Phospho-MLC2 antibodies, followed by the addition of secondary anti-rabbit antibodies. This was followed by one-hour incubation indoors. DAB staining was performed after 40 min, followed by restraint using hematoxylin. Images were captured using a Nikon Eclipse Ci scanning microscope (Nikon Instruments, Melville, NY).
2.8. ELISA Analysis
Fresh blood samples were centrifuged to collect serum (1000 rpm for 10 min at 4 ℃), which the IEC-6 cells seeded onto six-well plates as previously described. The cell supernatant was collected for analysis. Serum and IEC-6 supernatant levels of TNF-α, IL-4, IL-6, IL-8, IL-10, IL-13 and IFN-γ were measured using ELISA according to the manufacturer's instructions.
2.9. Western Blotting Experiments
Total protein from cell samples or small intestine tissues was extracted using RIPA Lysis buffer (containing 10 μL 100× PMSF, 10 μL phosphatase inhibitor per 1mL) via ultrasonic crusher (Scientz, Zhejiang, China) and the tissue grinder (Servicebio, Wuhan, China). The BCA Protein Assay Kit (Keygen Biotech, Jiangsu, China, Cat. No. KGP902) was used to quantify the protein concentration. Equal amounts of protein were separated using electrophoresis on sodium dodecyl sulfate-polyacrylamide gels (SDS-PAGE) and transferred onto polyvinylidene difluoride (PVDF) membranes. The membranes were then blocked with a rapid blocking buffer for 15 min before being incubated overnight at 4°C with various antibodies, including MLC2, Phospho-MLC2, MLCK, ERK1/2, Phospho-ERK1/2, ZO-1, Occludin, Claudin-1, Claudin-2 and β-actin. Finally, the membranes were incubated with secondary antibodies for 1 h. The membranes were washed three times with TBST solution for 5 min each time, followed by incubation with primary and secondary antibodies. The Bio-Rad ChemiDoc XRS imaging system was used to detect protein band intensity after the addition of Electro-Chemi-Luminescence (ECL) reagent (Seven Beijing, China). The analysis was conducted using Image Lab Software version 6.0 (Bio-Rad Laboratories).
2.10. Gut Microbiota Analysis
Sequencing was performed by Shanghai Personal Biotechnology Co., Ltd. (Shanghai, China). Briefly, total genomic DNA was isolated from the cecal contents using the OMEGA Soil DNA Kit (M5635-02) (Omega Bio-Tek, Norcross, GA, USA), according to the manufacturer's instructions. The V3–V4 regions of the 16S rRNA were amplified using forward primer 338F (5'-ACTCCTACGGGAGGCAGCA-3') and reverse primer 806R (5'-GGACTACHVGGGTWTCTAAT-3'). The DADA2 plug-in was used for quality filtering, denoising, merging and chimera removal. QIIME2 and R packages (v3.2.0) were utilized for processing sequence data, while ASV-level alpha diversity indices such as Chao1 Shannon Simpson were calculated via ASV table analysis. Beta diversity analysis [Bray-Curtis metrics via principal coordinate analysis (PCoA)] showed the structural variation of microbial communities in each sample group, while LDA Effect Size (LEfSe) detected differentially abundant taxa across groups using default parameters.
2.11. Data Analysis
All study outcomes were presented as the Mean ± Standard error of mean (SEM) from three independent experiments. Statistical analyses were performed using SPSS software (version 26.0; IBM, New York, NY, USA) and GraphPad Prism software (version 8.0; San Diego, USA). Student's t-test was used to compare two groups, while one-way analysis of variance (ANOVA) was employed for multiple comparisons of three or more groups, with p-values (p < 0.05) considered statistically significant.
3. Reaults
3.1. Cytotoxic Effects of CP-A on IEC-6 Cells
IEC-6 cells were treated with varying concentrations of CP-A for either 24 or 48 h, and no significant cytotoxicity was observed based on the MTT assay results, as shown in Figure 1b. Low and high doses of CP-A at 100 μg/mL and 200 μg/mL, respectively, were selected for subsequent in vitro studies.
3.2. Protective Effects of CP-A on 5-FU-Induced Intestinal Mucositis in Rats
The study established a rat model involving continuous intraperitoneal injection of 5-FU for 3 days, starting from the first day. The experimental design is displayed in Figure 2a. Apart from the control group, rats in the other groups experienced weight loss from the second day. Following BTC or CP-A administration, this trend was reversed. Rats in the BTC, CP-A medium and high-dose groups gradually gained weight from the fifth day, and the rats in the CP-A low-dose group gradually gained weight from the sixth day. The rats in the control group continued to gain weight during the experiment, while those in the 5-FU group rats continued to lose weight at the second day (Figure 2b). Food intake and diarrhea showed similar changes (Figure 2c and 2d). Histopathological results revealed varying degrees of damaged intestinal epithelium caused by 5-FU. The rats in the 5-FU group had the most severe intestinal damage, with significant mucosal necrosis compared to the control group. Administration of BTC and CP-A resulted in a dose-dependent alleviation of intestinal damage, as depicted in Figure 2e, while the histopathological scores showed a notable reduction compared to the group administered with only 5-FU (p < 0.01), as presented in Figure 2f.
3.3. Effect of CP-A on Inflammatory Cytokines
Assays conducted on IEC-6 cells revealed that following the administration of 5-FU, there was an up-regulation of expression levels of TNF-α, IFN-γ, IL-6, IL-8, and IL-13. Conversely, levels of IL-4 and IL-10 were significantly reduced. However, the intervention by BTC and CP-A reversed this trend, especially at a concentration level of CP-A (200 μg/mL and 60 mg/kg) (p < 0.01) (Figure 3).
3.4. Effect of CP-A on ERK/MLCK/MLC2 Signaling Pathway
After demonstrating that CP-A improved inflammatory cytokines levels, western blot analysis revealed a change in ERK/MLCK/MLC2 pathway following CP-A administration. We detected various proteins (ERK1/2, p-ERK1/2, MLCK, MLC2 and p-MLC2) associated with ERK/MLCK/MLC2 pathway in both IEC-6 cells and small intestinal tissues. As shown in Figure 4a-d and 4f-i, p-ERK1/2 was activated in the 5-FU group, and this activation was diminished after CP-A administration (p < 0.01). MLCK which is downstream of p-ERK1/2, was significantly downregulated compared to the group administered only 5-FU (p < 0.01). Western blot results obtained from rats treated with 5-FU and IEC-6 cells treated with 5-FU were similar. Protein immunohistochemical staining also yielded results consistent with those of the western blot analysis, as depicted in Figure 4e and 4j.
3.5. Effect of CP-A on Intestinal Mucosal Barrier Proteins
Western blotting was performed to determine the expression of tight junction proteins in IEC-6 cells and immunohistochemical staining to detect the expression of the proteins in the small intestine tissues. The results proved that 5-FU significantly downregulated the expression of Occludin, ZO-1 and Claudin-1 and upregulated the expression of Claudin-2. Nevertheless, these effects were reversed by CP-A. Similarly, 5-FU remarkably reduced the concentrations of Occludin, ZO-1, Claudin-1 and F-actin compared to the control group. After CP-A and BTC administration, the concentrations of these tight junction proteins increased (Figure 5a-f). The above data manifested that CP-A plays an important role in the treatment of cellular and intestinal tissue barrier dysfunction.
Then, RT-PCR was used to assess the mRNA levels of MLCK, ERK1/2, TNF-α, IL-4, IL-6 and IL-10. In the 5-FU group, there was a significant increase in the expression of MLCK, TNF-α and IL-6 compared to the control group. However, the expression of ERK1/2, IL-4 and IL-10 showed the opposite pattern. The RT-PCR results were similar to those of ELISA, western blotting and immunohistochemical staining (Figure 5g-l).
3.6. Effect of CP-A on Microbial Diversity
The rarefaction curve of the Shannon index reflects the detection depth of microorganisms in the sample (Figure 6a). The Venn diagram shows the number of ASVs/OTUs common and unique to each group (Figure 6b). Overlap analysis showed that 549 ASV/OTUs were present in all six groups. In addition, 8996, 9081, 8513, 7989, 8431 and 6856 unique ASVs/OTUs were observed in the control, 5-FU, BTC and CP-A (L, M, and H) groups, respectively. To evaluate the richness and diversity of the microbiota community, we utilized the Chao 1, Shannon and Simpson indices for alpha diversity analysis (Figure 6c). The rats in the 5-FU group had the highest Chao 1, Shannon and Simpson indices. After CP-A treatment, these indices decreased. These results suggested that 5-FU increased the diversity of the microbial community. Beta diversity was used to measure the similarity of microbial communities in each group. PCoA based on the Bray-Curtis distance showed the largest differences between the control, 5-FU and high-dose CP-A groups (Figure 6d). This result indicates that CP-A treatment might affect community structure.
3.7. Taxonomic Composition Analysis of Microbial Community
The gut microbial composition was determined at the Phylum, Class, Order, Family, and Genus levels. At the Phylum level, Firmicutes, Proteobacteria, Bacteroidetes, and Actinobacteria were the most abundant, and their abundance were more than 95% (Figure 7a and Table S2). After 5-FU injection, the abundance of Firmicutes and Bacteroidetes decreased from 89.49% to 62.86% and from 6.97% to 6.54%, respectively. In addition, the abundances of Proteobacteria and Actinobacteria increased to 27.03% and 2.57% by 5-FU. After CP-A intervention, their abundance recovered to normal levels.
We calculated the flora abundance at the class level, and the results are shown in Figure 7b and Table S3. Bacilli, Clostridia, Bacteroidia and Erysipelotrichi were the most abundant in the control group, but the abundance of Gammaproteobacteria was the second highest in the 5-FU group (p < 0.01 vs. control and CP-A H). After 5-FU administration, the abundance of Bacilli decreased notably (p < 0.01), whereas that of Clostridia increased to 43.28%, making it the highest in the 5-FU group. At the Order level, Lactobacillales, Clostridiales, Bacteroidales and Erysipelotrichales were the most abundant in control rats, whereas Clostridiales and Enterobacteriales were the two most abundant in 5-FU rats (Figure 7c and Table S4). Furthermore, the abundance of Enterobacteriales increased markedly in the 5-FU group (p < 0.01 vs. control and CP-A H). Lactobacillaceae were the dominant bacteria in the control group, and Enterobacteriaceae were the dominant bacteria in the 5-FU group at the Family level (Figure 7d and Table S5). After CP-A and BTC treatment, Enterobacteriaceae was observably decreased (p < 0.05). Lactobacillus was the characteristic bacteria in the control group, and Shigella was the characteristic bacteria in the 5-FU group at the genus level. Lactobacillus decreased notably in the 5-FU group compared to that in the control group (p < 0.01). After CP-A and BTC treatment, Lactobacillus was observably increased (p < 0.01) (Figure 7e and Table S6).
3.8. Effect of CP-A on Intestinal Mucosal Barrier Proteins
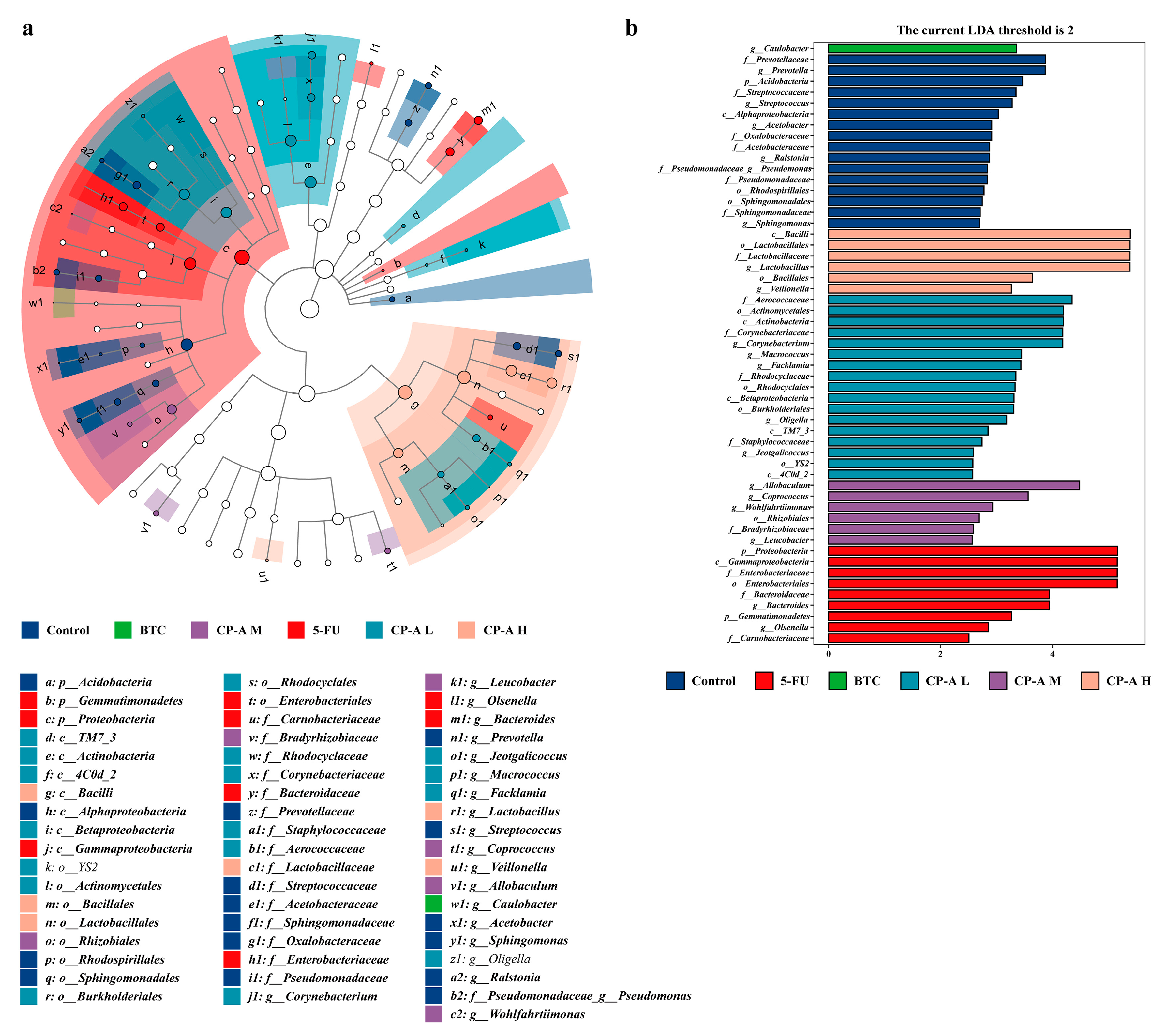
LEfSe analysis was used to identify biomarkers in each group (Figure 8a and 8b). g_Prevotella, g_Streptococcus, g_Acetobacter and g_Ralstonia were marker bacterial genera in the control group, whereas g_Olsenella and g_Bacteroides were highly expressed in the 5-FU group. After BTC and CP-A administration, g_Caulobacter was the main bacterium in the BTC group, while g_Lactobacillus and g_Veillonella were the marker bacteria in the CP-A H group.
4. Discussion
The main manifestations of IM induced by 5-FU were diarrhea and decreased food intake [29]. This is primarily attributed to the damage inflicted upon the integrity of the intestinal mucosa by 5-FU [30]. Previous research has indicated that the mucosa lining of the small intestine is particularly susceptible to the detrimental effects of 5-FU treatment [31,32]. Our study aimed to investigate the effect of CP-A on inflammation induced by 5-FU, both in vitro and in vivo. Our findings demonstrated that CP-A effectively restored body weight and food intake, alleviated diarrhea symptoms and reduced intestinal mucosal damage in rats with 5-FU-induced IM. These results suggest that CP-A has an inhibitory effect on 5-FU-induced IM and probably plays a role in inhibiting the ERK/MLCK/MLC2 signaling pathway and regulating microbial structure.
Inflammation can occur due to the release of pro-inflammatory factors and often accompanies various immune responses and processes [33,34]. TNF-α, an inflammatory cytokine, is produced by macrophages and monocytes during inflammation, and it is involved in the conduction of various intracellular signals, leading to cell necrosis or apoptosis [35]. TNF-α is also an essential pro-inflammatory cytokine detectable in various inflammatory conditions [36,37]. Endotoxins and exotoxins are the most common inducers of cytokine production [38]. As inflammation progresses, the generation of inflammatory cytokines including, TNF-α and IFN-γ, orchestrates the innate immune response against infections [39]. However, excessive expression of TNF-α and IFN-γ is harmful, and IL-6 and IL-8 also contribute to inflammation [40]. Furthermore, both IL-4 and IL-10 were found to downregulate MCP-1 production, which plays a crucial role in intestinal inflammation, not only in Caco2 cells but also in freshly isolated epithelial cells [41]. Our in vitro and in vivo experiments revealed that CP-A exhibited remarkable anti-inflammatory effects by modulating inflammatory cytokine levels in IEC-6 cells stimulated with 5-FU and in rats with intestinal mucositis. CP-A significantly reduced the levels of IL-6, IL-8, TNF-α and IFN-γ, while enhancing IL-4 and IL-10 expression in both cell cultures and the rat model rat.
Tight-junction proteins play a protective role in the intestinal mucosal barrier, and their degradation is often observed during intestinal inflammation [9]. An increasing number of studies have shown that the degradation of tight junction proteins is accompanied by the activation of ERK and MLCK [17,20]. MLCK promotes the phosphorylation of MLC2, which increases the expression of p-MLC2 and produces ATP, which degrades tight junction proteins [36]. Moreover, a 5-FU-induced increase in TNF-α and IFN-γ expression not only promotes the occurrence of inflammation but also acts as an activator of ERK and MLCK [9]. Furthermore, the activation of MLCK leads to increased secretion of IL-13 and Claudin-2 expression [20,42]. The ERK/MLCK/MLC2 signaling pathway becomes activated due to elevated p-ERK1/2 levels and MLC2 phosphorylation following 5-FU administration in both cellular and rat models. However, treatment with CP-A significantly reduced p-ERK1/2 levels, along with MLC2 phosphorylation and MLCK expression, thereby deactivating the ERK/MLCK/MLC2 signaling pathway. Thus, the expression of Claudin-1, F-actin, Occludin, and ZO-1 in the cells and small intestine was upregulated, whereas the expression of Claudin-2 and IL-13 were downregulated.
To assess the potential cytotoxicity of CP-A on IEC-6 cells, we performed an MTT assay, which indicated that CP-A promotes cell growth without causing cytotoxic effects. Another study also supported CP-A's lack of cytotoxicity [43]. Western blot and IHC results revealed that CP-A significantly restores Claudin-1, Occludin and ZO-1 levels while down-regulating IL-6, IL-8, TNF-α and IFN-γ.
Additionally, the gut microbiota plays a crucial role in maintaining intestinal homeostasis as it acts as an ecological barrier [44]. Imbalances within the gut microbiota can lead to various adverse effects, such as diarrhea and indigestion, associated with inflammatory bowel diseases, such as ulcerative colitis or Crohn's disease [4,45]. Interestingly, our sequencing data revealed that 5-FU disrupted the overall structure and abundance of the gut microbiota and altered the relative abundance of specific bacterial genera known to inhabit the intestine [11,12,21]. The results of α-diversity analysis indicated that high-dose CP-A decreased the richness and diversity of the flora. However, the results of β-diversity analysis indicated that the high-dose CP-A group and the control group had similar microflora structures, whereas the 5-FU group displayed a significantly distinct microflora structure. Hence, CP-A effectively stabilized the microbial community.
Firmicutes, Proteobacteria, Bacteroidetes and Actinobacteria are the four dominant bacteria in normal organisms, with Firmicutes having the highest relative abundance. Our findings are consistent with those of previous studies showing that 5-FU can alter the gut microbiota composition by decreasing Firmicutes abundance while increasing Proteobacteria prevalence. This shift in composition may affect dietary energy absorption, given Firmicutes's role in this process [46]. Research has shown that dysregulation of the gut flora accompanies 5-FU-induced intestinal mucositis, leading to increased Proteobacteria abundance and gastrointestinal disorders such as enteritis and diarrhea [4]. Interestingly, CP-A reversed these changes in the abundance of Firmicutes and Proteobacteria, offering a potential avenue for mitigation.
To further investigate bacterial genera specifically altered by 5-FU and CP-A treatment, we examined their relative abundances among the rats used in this study. 5-FU significantly increased the relative abundance of Shigella and decreased the relative abundance of Lactobacillus; however, CP-A restored the relative abundance of these genera. Notably, high-dose CP-A treatment resulted in a significant increase in the relative abundance of Lactobacillus compared to that in the other groups. Lactobacillus is an intestinal probiotic that effectively combats acute diarrhea and intestinal mucosal lesions [47]. Shigella is a small gram-negative bacterium that invades the intestinal epithelium and causes shigellosis, also known as bacillary dysentery. The main cause of bacterial diarrhea is Shigella infection, which is highly lethal in malnourished people [48]. Therefore, CP-A administration was found to alleviate 5-FU-induced intestinal mucositis in rats by reducing the prevalence of Shigella and increasing the prevalence of Lactobacillus.
5. Conclusions
These results suggested that CP-A may exert its beneficial effects by decreasing inflammatory factors, improving intestinal mucosal barrier damage and modulating the gut microbiota. Specifically, CP-A was able to inhibit MLCK expression, suppress TNF-α and IFN-γ signaling pathways, and prevent degradation of Claudin-1, F-actin, Occludin and ZO-1 proteins while also reducing Claudin-2 and IL-13 expression levels. CP-A reduces pro-inflammatory cytokine levels (IL-6, IL-8, TNF-α and IFN-γ), increases anti-inflammatory cytokine levels (IL-4 and IL-10), stabilizes microbial structure, and balances microbial communities by up-regulating Lactobacillus relative abundance while down-regulating Shigella relative abundance. In conclusion, our work provided potential novel strategy for treatment of chemotherapy-induced IM by Codonopsis pilosula. Moreover, it also provided experimental evidence for the application of in patients with IM caused by chemotherapy. Nevertheless, as this study was only performed the molecular mechanism and changes in the gut microbiota in rats unilaterally, which may have limitations. In our future endeavor, more in-depth investigations on the anti-IM effect of CP-A, including genetically defective animals or fecal bacteria transplantation would be performed. These future studies should be conducted to gain further insight into the mechanism of action and highlight the potential of CP-A as a promising candidate drug.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org. Supplementary Material 1: Table S1 Sequences of the primers used in the RT-PCR. Table S2 Relative abundance of the most representative phyla in experimental rats. Table S3 Relative abundance of the most representative classes in experimental rats. Table S4 Relative abundance of the most representative orders in experimental rats. Table S5 Relative abundance of the most representative families in experimental rats. Table S6 Relative abundance of the most representative families in experimental rats. Supplementary Material 2: Original gels used in Figure 4 and Figure 5.
Author Contributions
Jiangtao Zhou: Conceptualisation, Methodology, Investigation, Writing – original draft, Writing-review & editing, Funding acquisition. Deyun Li: Conceptualisation, Investigation, Resources, Writing – original draft, Writing- review & editing, Data curation. Jiajing Wang: Data curation, Formal Analysis, Resources, Investigation. Zhuoyang Cheng, Changjian Wang, Xuepeng Zhang, Xiexin Xu: Investigation, Resources. Jianping Gao: Conceptualization, Methodology, Writing-original draft, Writing-review & editing, Funding acquisition.
Funding
This work was supported by National Natural Science Foundation of China (81904031), National Key Research and Development Program of China (2019YFC1710800), Natural Science Foundation of Shanxi Province (201901D211325) and Science Research Start-up Fund for Doctor of Shanxi Medical University (XD1802).
Institutional Review Board Statement
The animal experimental scheme strictly followed the ethical code of Animal Care and Use Committee of Shanxi Medical University and the Guidelines for The Care and Use of Laboratory Animal (approval number: SYXK (Jing) 2017-0005).
Data Availability Statement
The cecum microbiota gene sequence dataset presented in this study has been made available at the NCBI online repository: NCBI; Submission ID: SUB14100411; BioProject ID: PRJNA1054780.
Conflicts of Interest
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
Abbreviations
| IM | Intestinal mucositis |
| 5-FU | 5-fluorouracil |
| IHC | Immunohistochemical |
| RT-PCR | Real-time PCR |
| ERK | Extracellular-regulated protein kinases |
| MLCK | Myosin light chain kinase |
| MLC2 | Myosin light chain 2 |
| ELISA | Enzyme-linked immunosorbent assay |
| TNF-α | Tumor necrosis factor-alpha |
| IFN-γ | Interferon gamma |
| ZO-1 | Zonula occludens-1 |
| IL | Interleukin |
| BTC | Bifid. Triple Viable Capsules Dissolving at Intestines |
| MTT | 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide |
| DMSO | Dimethyl sulfoxide |
| PBS | Phosphate-buffered saline |
| H&E | Hematoxylin and eosin |
| PCoA | Principal coordinate analysis |
| LEfSe | LDA Effect Size |
| FBS | Fetal bovine serum |
| BSA | Bovine serum albumin |
| SPF | Specific pathogen-free |
| SD | Sprague–Dawley |
| TCM | Traditional Chinese Medicine |
| IEC-6 | Rat intestinal epithelial cell line-6 |
| PVDF | polyvinylidene difluoride |
| ECL | Electro-Chemi-Luminescence |
References
- D. B. Longley, D. P. Harkin and P. G. Johnston, 5-fluorouracil: mechanisms of action and clinical strategies. Nat. Rev. Cancer 2003, 3, 330–338. [Google Scholar] [CrossRef] [PubMed]
- R. A. Ribeiro, C. W. Wanderley, D. V. Wong, J. M. Mota, C. A. Leite, M. H. Souza, F. Q. Cunha and R. C. Lima-Junior, Irinotecan- and 5-fluorouracil-induced intestinal mucositis: insights into pathogenesis and therapeutic perspectives. Cancer Chemother. Pharmacol. 2016, 78, 881–893. [Google Scholar] [CrossRef] [PubMed]
- J. Y. Douillard, D. Cunningham, A. D. Roth, M. Navarro, R. D. James, P. Karasek, P. Jandik, T. Iveson, J. Carmichael, M. Alakl, G. Gruia, L. Awad and P. Rougier, Irinotecan combined with fluorouracil compared with fluorouracil alone as first-line treatment for metastatic colorectal cancer: a multicentre randomised trial. Lancet 2000, 355, 1041–1047. [Google Scholar]
- J. Wang, W. Feng, S. Zhang, L. Chen, Y. Sheng, F. Tang, J. He, X. Xu, H. Ao and C. Peng, Ameliorative effect of Atractylodes macrocephala essential oil combined with Panax ginseng total saponins on 5-fluorouracil induced diarrhea is associated with gut microbial modulation. J. Ethnopharmacol. 2019, 238, 111887. [Google Scholar]
- R. V. Lalla, M. M. Schubert, R. J. Bensadoun and D. Keefe, Anti-inflammatory agents in the management of alimentary mucositis. Support. Care Cancer 2006, 14, 558–565. [Google Scholar] [CrossRef] [PubMed]
- D. M. Keefe, Intestinal mucositis: mechanisms and management. Curr. Opin. Oncol. 2007, 19, 323–327. [Google Scholar] [CrossRef]
- R. Sharma, P. Tobin and S. J. Clarke, Management of chemotherapy-induced nausea, vomiting, oral mucositis, and diarrhoea. Lancet Oncol. 2005, 6, 93–102. [Google Scholar] [CrossRef] [PubMed]
- B. Cai, J. Pan, H. Chen, X. Chen, Z. Ye, H. Yuan, H. Sun and P. Wan, Oyster polysaccharides ameliorate intestinal mucositis and improve metabolism in 5-fluorouracil-treated S180 tumour-bearing mice. Carbohydr. Polym. 2021, 256, 117545. [Google Scholar] [CrossRef] [PubMed]
- J. Wu, Y. Gan, M. Li, L. Chen, J. Liang, J. Zhuo, H. Luo, N. Xu, X. Wu, Q. Wu, Z. Lin, Z. Su and Y. Liu, Patchouli alcohol attenuates 5-fluorouracil-induced intestinal mucositis via TLR2/MyD88/NF-kB pathway and regulation of microbiota. Biomed. Pharmacother. 2020, 124, 109883. [Google Scholar]
- M. B. Zeisel, P. Dhawan and T. F. Baumert, Tight junction proteins in gastrointestinal and liver disease. Gut 2019, 68, 547–561. [Google Scholar] [CrossRef]
- H. Chen, F. Zhang, R. Li, Y. Liu, X. Wang, X. Zhang, C. Xu, Y. Li, Y. Guo and Q. Yao, Berberine regulates fecal metabolites to ameliorate 5-fluorouracil induced intestinal mucositis through modulating gut microbiota. Biomed. Pharmacother. 2020, 124, 109829. [Google Scholar]
- H. L. Li, L. Lu, X. S. Wang, L. Y. Qin, P. Wang, S. P. Qiu, H. Wu, F. Huang, B. B. Zhang, H. L. Shi and X. J. Wu, Alteration of Gut Microbiota and Inflammatory Cytokine/Chemokine Profiles in 5-Fluorouracil Induced Intestinal Mucositis. Front. Cell Infect. Microbiol. 2017, 7, 455. [Google Scholar]
- S. Deng, D. Wu, L. Li, J. Li and Y. Xu, TBHQ attenuates ferroptosis against 5-fluorouracil-induced intestinal epithelial cell injury and intestinal mucositis via activation of Nrf2. Cell Mol. Biol. Lett. 2021, 26, 48. [Google Scholar]
- S. Kato, S. Hayashi, Y. Kitahara, K. Nagasawa, H. Aono, J. Shibata, D. Utsumi, K. Amagase and M. Kadowaki, Saireito (TJ-114), a Japanese traditional herbal medicine, reduces 5-fluorouracil-induced intestinal mucositis in mice by inhibiting cytokine-mediated apoptosis in intestinal crypt cells. PLoS ONE 2015, 10, e0116213. [Google Scholar]
- Atiq, B. Shal, M. Naveed, A. Khan, J. Ali, S. Zeeshan, S. D. Al-Sharari, Y. S. Kim and S. Khan, Diadzein ameliorates 5-fluorouracil-induced intestinal mucositis by suppressing oxidative stress and inflammatory mediators in rodents. Eur. J. Pharmacol. 2019, 843, 292–306. [Google Scholar] [CrossRef] [PubMed]
- J. Su, C. Li, X. Yu, G. Yang, J. Deng, Z. Su, H. Zeng, J. Chen, X. Zhang and X. Lai, Protective Effect of Pogostone on 2,4,6-Trinitrobenzenesulfonic Acid-Induced Experimental Colitis via Inhibition of T Helper Cell. Front. Pharmacol. 2017, 8, 829. [Google Scholar] [CrossRef] [PubMed]
- J. Sui, C. Zhang, X. Fang, J. Wang, Y. Li, J. Wang, L. Wang, J. Dong, Z. Zhou, C. Li, J. Chen, T. Ma and D. Chen, Dual role of Ca(2+)-activated Cl(-) channel transmembrane member 16A in lipopolysaccharide-induced intestinal epithelial barrier dysfunction in vitro. Cell Death Dis. 2020, 11, 404. [Google Scholar]
- L. Du, J. J. Kim, J. Shen and N. Dai, Crosstalk between Inflammation and ROCK/MLCK Signaling Pathways in Gastrointestinal Disorders with Intestinal Hyperpermeability. Gastroenterol. Res. Pract. 2016, 2016, 7374197. [Google Scholar]
- R. Al-Sadi, S. Guo, D. Ye and T. Y. Ma, TNF-alpha modulation of intestinal epithelial tight junction barrier is regulated by ERK1/2 activation of Elk-1. Am. J. Pathol. 2013, 183, 1871–1884. [Google Scholar] [CrossRef]
- C. R. Weber, D. R. Raleigh, L. Su, L. Shen, E. A. Sullivan, Y. Wang and J. R. Turner, Epithelial myosin light chain kinase activation induces mucosal interleukin-13 expression to alter tight junction ion selectivity. J. Biol. Chem. 2010, 285, 12037–12046. [Google Scholar] [CrossRef]
- S. Chen, K. Qian, G. Zhang and M. Zhang, Akkermansia muciniphila and its outer membrane protein Amuc_1100 prophylactically attenuate 5-fluorouracil-induced intestinal mucositis. Biochem. Biophys. Res. Commun. 2022, 614, 34–40. [Google Scholar] [CrossRef] [PubMed]
- T. Zhang, S. H. Lu, Q. Bi, L. Liang, Y. F. Wang, X. X. Yang, W. Gu and J. Yu, Volatile Oil from Amomi Fructus Attenuates 5-Fluorouracil-Induced Intestinal Mucositis. Front. Pharmacol. 2017, 8, 786. [Google Scholar] [CrossRef] [PubMed]
- X. Chu, X. J. Liu, J. M. Qiu, X. L. Zeng, H. R. Bao and J. Shu, Effects of Astragalus and Codonopsis pilosula polysaccharides on alveolar macrophage phagocytosis and inflammation in chronic obstructive pulmonary disease mice exposed to PM2.5. Environ. Toxicol. Pharmacol. 2016, 48, 76–84. [Google Scholar] [CrossRef]
- Y. Meng, Y. Xu, C. Chang, Z. Qiu, J. Hu, Y. Wu, B. Zhang and G. Zheng, Extraction, characterization and anti-inflammatory activities of an inulin-type fructan from Codonopsis pilosula. Int. J. Biol. Macromol. 2020, 163, 1677–1686. [Google Scholar] [CrossRef]
- S. M. Gao, J. S. Liu, M. Wang, T. T. Cao, Y. D. Qi, B. G. Zhang, X. B. Sun, H. T. Liu and P. G. Xiao, Traditional uses, phytochemistry, pharmacology and toxicology of Codonopsis: A review. J. Ethnopharmacol. 2018, 219, 50–70. [Google Scholar]
- Y. Zou, H. Yan, C. Li, F. Wen, X. Jize, C. Zhang, S. Liu, Y. Zhao, Y. Fu, L. Li, F. Liu, J. Chen, R. Li, X. Chen and M. Tian, A Pectic Polysaccharide from Codonopsis pilosula Alleviates Inflammatory Response and Oxidative Stress of Aging Mice via Modulating Intestinal Microbiota-Related Gut–Liver Axis. Antioxidants 2023, 12. [Google Scholar]
- J. Li, T. Wang, Z. Zhu, F. Yang, L. Cao and J. Gao, Structure Features and Anti-Gastric Ulcer Effects of Inulin-Type Fructan CP-A from the Roots of Codonopsis pilosula (Franch.) Nannf. Molecules 2017, 22. [Google Scholar]
- G. Pepe, S. F. Rapa, E. Salviati, A. Bertamino, G. Auriemma, S. Cascioferro, G. Autore, A. Quaroni, P. Campiglia and S. Marzocco, Bioactive Polyphenols from Pomegranate Juice Reduce 5-Fluorouracil-Induced Intestinal Mucositis in Intestinal Epithelial Cells. Antioxidants 2020, 9. [Google Scholar]
- J. H. Liu, C. H. Hsieh, C. Y. Liu, C. W. Chang, Y. J. Chen and T. H. Tsai, Anti-inflammatory effects of Radix Aucklandiae herbal preparation ameliorate intestinal mucositis induced by 5-fluorouracil in mice. J. Ethnopharmacol. 2021, 271, 113912. [Google Scholar] [CrossRef]
- D. Korenaga, M. Honda, M. Yasuda, S. Inutsuka, T. Nozoe and H. Tashiro, Increased intestinal permeability correlates with gastrointestinal toxicity among formulations of the fluorouracil analogue tegafur in rats. Eur. Surg. Res. 2002, 34, 351–356. [Google Scholar] [CrossRef]
- C. Tefas, L. Ciobanu, C. Berce, A. Mester, S. Onica, C. Toma, M. Tantau and M. Taulescu, Beneficial effect of oral administration of zinc sulfate on 5-fluorouracil-induced gastrointestinal mucositis in rats. Eur. Rev. Med. Pharmacol. Sci. 2020, 24, 11365–11373. [Google Scholar]
- X. Y. Wang, B. Zhang, Y. Lu, L. Xu, Y. J. Wang, B. Y. Cai and Q. H. Yao, RNA-seq and In Vitro Experiments Reveal the Protective Effect of Curcumin against 5-Fluorouracil-Induced Intestinal Mucositis via IL-6/STAT3 Signaling Pathway. J. Immunol. Res. 2021, 2021, 8286189. [Google Scholar]
- D. C. Xiang, J. Y. Yang, Y. J. Xu, S. Zhang, M. Li, C. Zhu, C. L. Zhang and D. Liu, Protective effect of Andrographolide on 5-Fu induced intestinal mucositis by regulating p38 MAPK signaling pathway. Life Sci. 2020, 252, 117612. [Google Scholar] [CrossRef]
- X. Li, Q. Li, B. Xiong, H. Chen, X. Wang and D. Zhang, Discoidin domain receptor 1(DDR1) promote intestinal barrier disruption in Ulcerative Colitis through tight junction proteins degradation and epithelium apoptosis. Pharmacol. Res. 2022, 183, 106368. [Google Scholar] [CrossRef] [PubMed]
- H. T. Idriss and J. H. Naismith, TNF? and the TNF receptor superfamily: Structure-function relationship(s). Microsc. Res. Tech. 2000, 50, 184–195. [Google Scholar] [CrossRef]
- S. Huang, Y. Fu, B. Xu, C. Liu, Q. Wang, S. Luo, F. Nong, X. Wang, S. Huang, J. Chen, L. Zhou and X. Luo, Wogonoside alleviates colitis by improving intestinal epithelial barrier function via the MLCK/pMLC2 pathway. Phytomedicine 2020, 68, 153179. [Google Scholar] [CrossRef] [PubMed]
- J. Wu, Y. Gan, H. Luo, N. Xu, L. Chen, M. Li, F. Guan, Z. Su, Z. Lin, J. Xie and Y. Liu, beta-Patchoulene Ameliorates Water Transport and the Mucus Barrier in 5-Fluorouracil-Induced Intestinal Mucositis Rats via the cAMP/PKA/CREB Signaling Pathway. Front. Pharmacol. 2021, 12, 689491. [Google Scholar] [CrossRef] [PubMed]
- J. M. Cavaillon, Exotoxins and endotoxins: Inducers of inflammatory cytokines. Toxicon 2018, 149, 45–53. [Google Scholar] [CrossRef]
- P. C. Heinrich, I. Behrmann, S. Haan, H. M. Hermanns, G. Muller-Newen and F. Schaper, Principles of interleukin (IL)-6-type cytokine signalling and its regulation. Biochem. J. 2003, 374, 1–20. [Google Scholar] [CrossRef]
- N. Nishimoto and T. Kishimoto, Inhibition of IL-6 for the treatment of inflammatory diseases. Curr. Opin. Pharmacol. 2004, 4, 386–391. [Google Scholar] [CrossRef]
- T. Kucharzik, N. Lugering, H. G. Pauels, W. Domschke and R. Stoll, IL-4, IL-10 and IL-13 down-regulate monocyte-chemoattracting protein-1 (MCP-1) production in activated intestinal epithelial cells. Clin. Exp. Immunol. 1998, 111, 152–157. [Google Scholar]
- X. Liu, J. Xu, Q. Mei, L. Han and J. Huang, Myosin Light Chain Kinase Inhibitor Inhibits Dextran Sulfate Sodium-Induced Colitis in Mice. Dig. Dis. Sci. 2012, 58, 107–114. [Google Scholar]
- Y. Y. Shao, Y. N. Zhao, Y. F. Sun, Y. Guo, X. Zhang, Z. P. Chang, R. G. Hou and J. Gao, Investigation of the internalization and transport mechanism of Codonopsis Radix polysaccharide both in mice and Caco-2 cells. Int. J. Biol. Macromol. 2022, 215, 23–35. [Google Scholar] [CrossRef] [PubMed]
- J. -h. Chen, C.-l. Zhao, Y.-s. Li, Y.-b. Yang, J.-g. Luo, C. Zhang and L. Wang, Moutai Distiller's grains Polyphenol extracts and rutin alleviate DSS-induced colitis in mice: Modulation of gut microbiota and intestinal barrier function (R2). Heliyon 2023, 9. [Google Scholar]
- Adak and M., R. Khan, An insight into gut microbiota and its functionalities. Cell Mol. Life Sci. 2019, 76, 473–493. [Google Scholar] [CrossRef] [PubMed]
- Kong, R. Gao, X. Yan, L. Huang, J. He, H. Li, J. You and H. Qin, Alterations in intestinal microbiota of colorectal cancer patients receiving radical surgery combined with adjuvant CapeOx therapy. Sci. China Life Sci. 2019, 62, 1178–1193. [Google Scholar] [CrossRef] [PubMed]
- E. Kim, D. B. Kim and J. Y. Park, Changes of Mouse Gut Microbiota Diversity and Composition by Modulating Dietary Protein and Carbohydrate Contents: A Pilot Study. Prev. Nutr. Food Sci. 2016, 21, 57–61. [Google Scholar] [CrossRef]
- M. Qasim, M. Wrage, B. Nuse and J. Mattner, Shigella Outer Membrane Vesicles as Promising Targets for Vaccination. Int. J. Mol. Sci. 2022, 23. [Google Scholar]
Figure 1.
Cytotoxicity of CP-A to IEC-6 cells (a) The chemical structure of CP-A (b) Cell viability was measured by MTT. Values were represented as mean±SEM (n = 3).
Figure 1.
Cytotoxicity of CP-A to IEC-6 cells (a) The chemical structure of CP-A (b) Cell viability was measured by MTT. Values were represented as mean±SEM (n = 3).
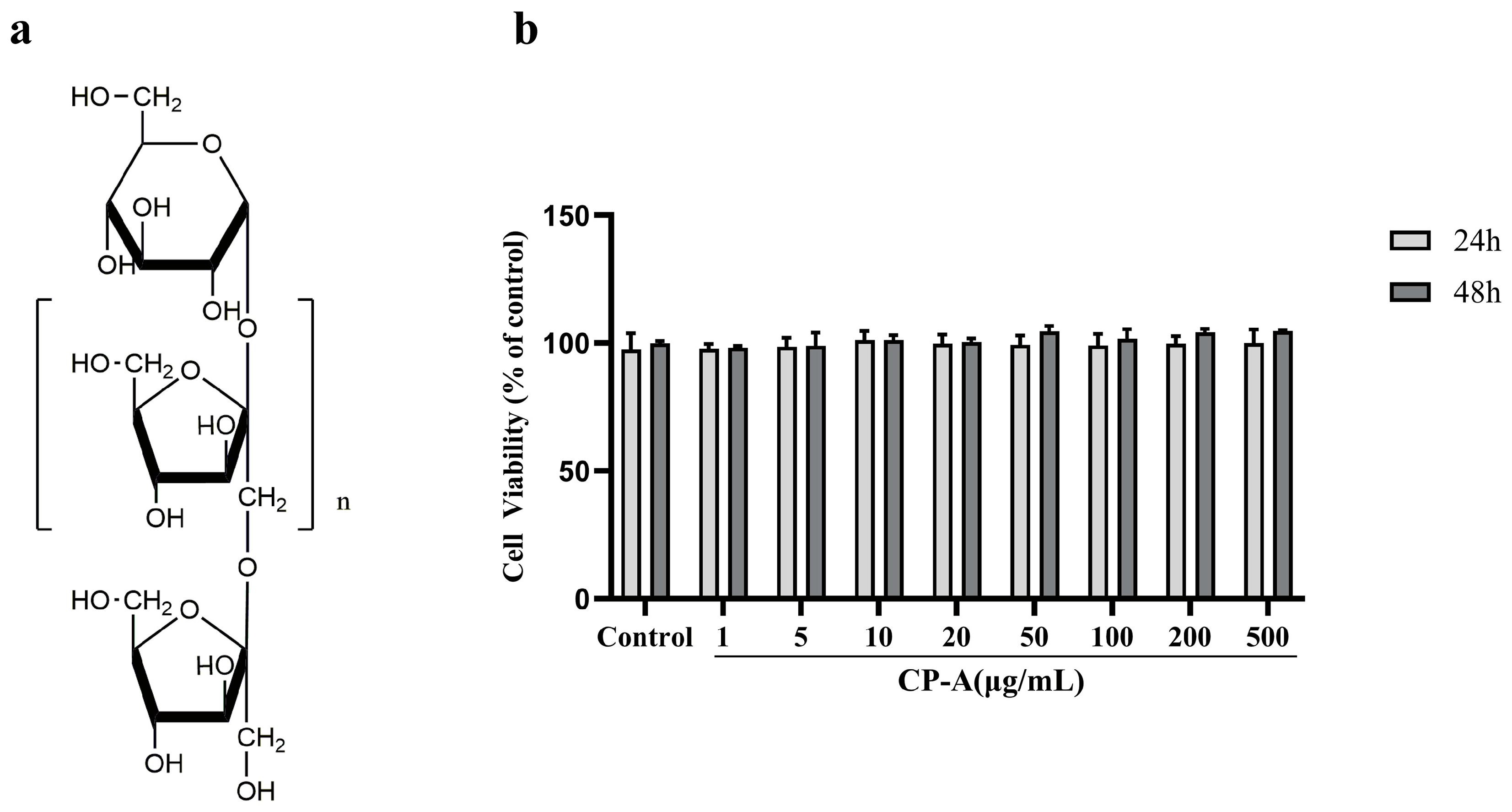
Figure 2.
CP-A alleviates the macroscopic manifestation of intestinal mucositis in rats. (a) Experimental design and study grouping. (b) Body weight changes (n = 6) (c) Food intake (n = 6). (d) Diarrhea scores (n = 6). (e) H&E-stained sections from ileums (n = 3, magnification 50×). (f) Histopathological scores. CP-A L, low dose of CP-A (15 mg/kg, i.g.); CP-A M, medium dose of CP-A (30 mg/kg, i.g.); CP-A H, high dose of CP-A (60 mg/kg, i.g). Values were represented as mean±SEM. #p < 0.05 and ##p < 0.01 versus control group, *p < 0.05 and **p < 0.01 versus 5-FU group.
Figure 2.
CP-A alleviates the macroscopic manifestation of intestinal mucositis in rats. (a) Experimental design and study grouping. (b) Body weight changes (n = 6) (c) Food intake (n = 6). (d) Diarrhea scores (n = 6). (e) H&E-stained sections from ileums (n = 3, magnification 50×). (f) Histopathological scores. CP-A L, low dose of CP-A (15 mg/kg, i.g.); CP-A M, medium dose of CP-A (30 mg/kg, i.g.); CP-A H, high dose of CP-A (60 mg/kg, i.g). Values were represented as mean±SEM. #p < 0.05 and ##p < 0.01 versus control group, *p < 0.05 and **p < 0.01 versus 5-FU group.
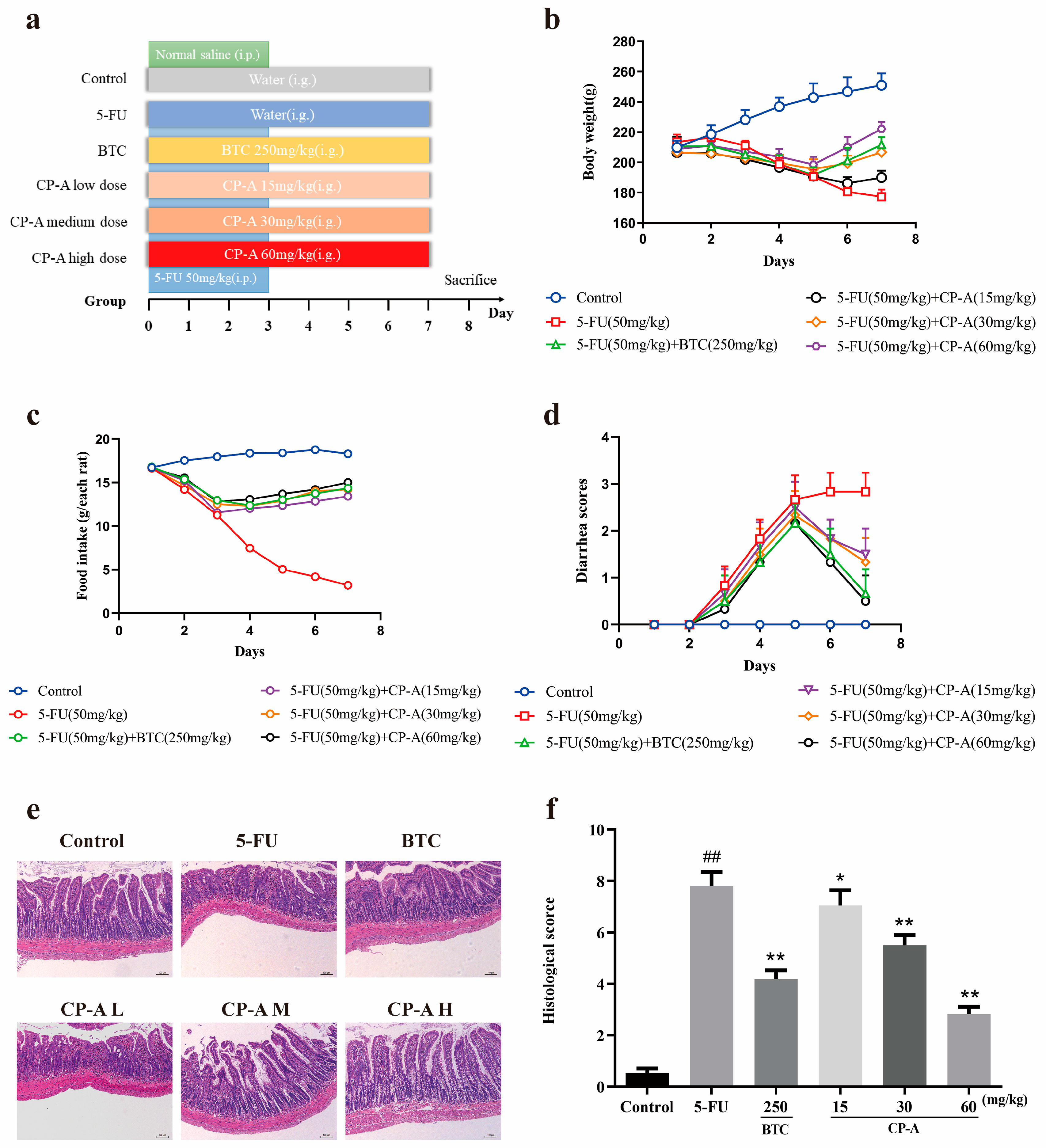
Figure 3.
Effects of CP-A on inflammatory cytokines in 5-FU induced rats and IEC-6 cells. (a-g) The levels of TNF-α, IFN-γ, IL-4, IL-6, IL-8, IL-10 and IL-13 in rat serum (n = 6). (h-m) The levels of TNF-α, IFN-γ, IL-4, IL-6, IL-8 and IL-13 in cellular supernatant (n = 6). Values were represented as mean±SEM. #p < 0.05 and ##p < 0.01 versus control group, *p < 0.05 and **p < 0.01 versus 5-FU group.
Figure 3.
Effects of CP-A on inflammatory cytokines in 5-FU induced rats and IEC-6 cells. (a-g) The levels of TNF-α, IFN-γ, IL-4, IL-6, IL-8, IL-10 and IL-13 in rat serum (n = 6). (h-m) The levels of TNF-α, IFN-γ, IL-4, IL-6, IL-8 and IL-13 in cellular supernatant (n = 6). Values were represented as mean±SEM. #p < 0.05 and ##p < 0.01 versus control group, *p < 0.05 and **p < 0.01 versus 5-FU group.
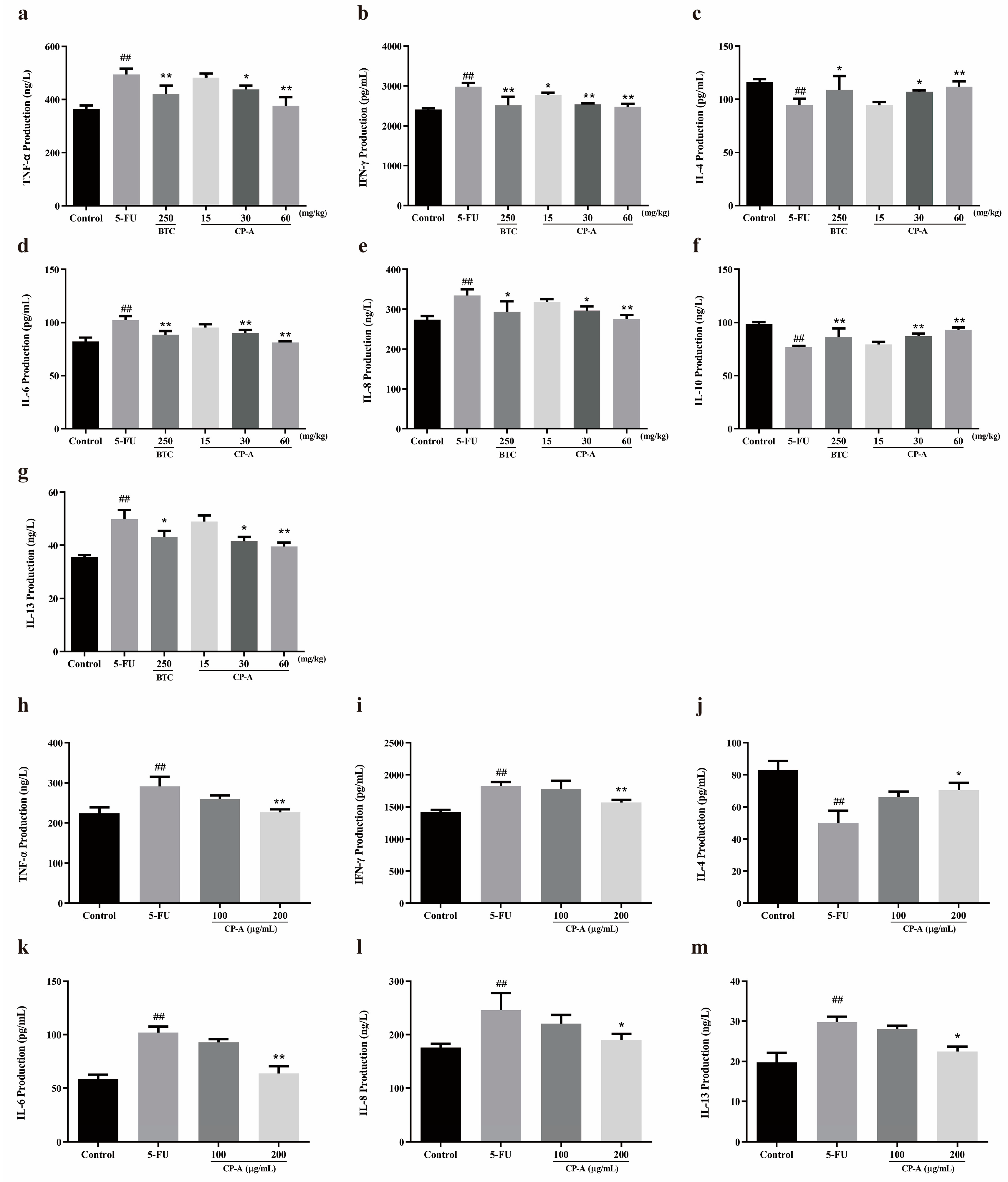
Figure 4.
Effects of CP-A on ERK/MLCK/MLC2 signaling pathway proteins in 5-FU induced rats and IEC-6 cells. (a-d) Expression of MLC2, p-MLC2, MLCK, ERK1/2 and p-ERK1/2 in IEC-6 cells. (e) MLCK protein in IEC-6 cells (magnification 100×). (f-i) Expression of MLC2, p-MLC2, MLCK, ERK1/2 and p-ERK1/2 in intestinal. (j) p-MLC2 protein in IEC-6 cells (magnification 100x). Values were represented as mean±SEM (n = 3). #p < 0.05 and ##p < 0.01 versus control group, *p < 0.05 and **p < 0.01 versus 5-FU group.
Figure 4.
Effects of CP-A on ERK/MLCK/MLC2 signaling pathway proteins in 5-FU induced rats and IEC-6 cells. (a-d) Expression of MLC2, p-MLC2, MLCK, ERK1/2 and p-ERK1/2 in IEC-6 cells. (e) MLCK protein in IEC-6 cells (magnification 100×). (f-i) Expression of MLC2, p-MLC2, MLCK, ERK1/2 and p-ERK1/2 in intestinal. (j) p-MLC2 protein in IEC-6 cells (magnification 100x). Values were represented as mean±SEM (n = 3). #p < 0.05 and ##p < 0.01 versus control group, *p < 0.05 and **p < 0.01 versus 5-FU group.
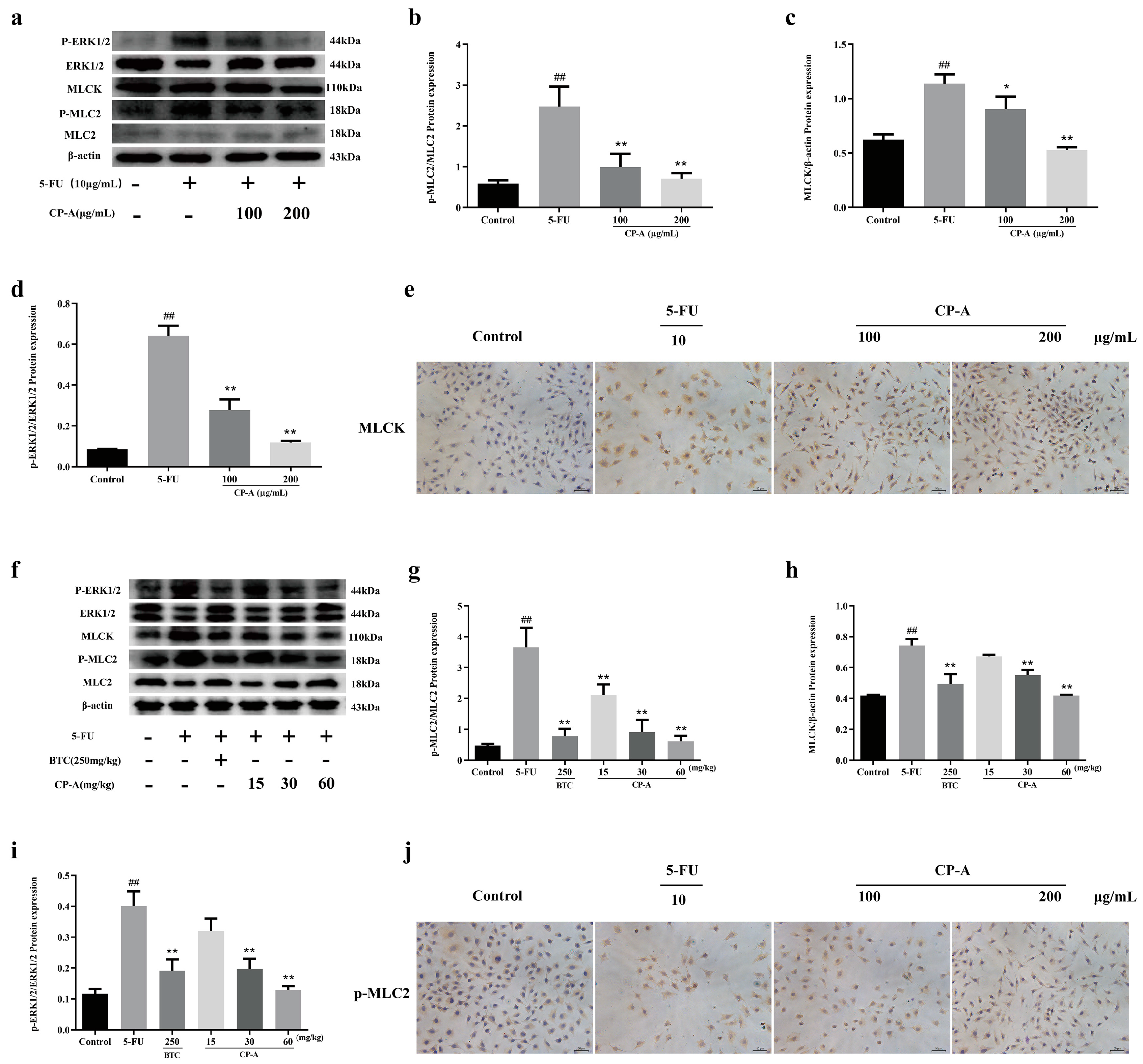
Figure 5.
Effects of CP-A on intestinal mucosal barrier proteins. (a-e) Expression of Claudin-1, Claudin-2, Occludin and ZO-1 proteins in IEC-6 cells. (f) Expression of Claudin-1, F-actin, Occludin and ZO-1 proteins in intestinal (magnification 50x). (g-l) Relative mRNA expression of ERK1/2, MLCK, TNF-α, IL-4, IL-6 and IL-10. Values were represented as mean±SEM (n = 3). #p < 0.05 and ##p < 0.01 versus control group, *p < 0.05 and **p < 0.01 versus 5-FU group.
Figure 5.
Effects of CP-A on intestinal mucosal barrier proteins. (a-e) Expression of Claudin-1, Claudin-2, Occludin and ZO-1 proteins in IEC-6 cells. (f) Expression of Claudin-1, F-actin, Occludin and ZO-1 proteins in intestinal (magnification 50x). (g-l) Relative mRNA expression of ERK1/2, MLCK, TNF-α, IL-4, IL-6 and IL-10. Values were represented as mean±SEM (n = 3). #p < 0.05 and ##p < 0.01 versus control group, *p < 0.05 and **p < 0.01 versus 5-FU group.
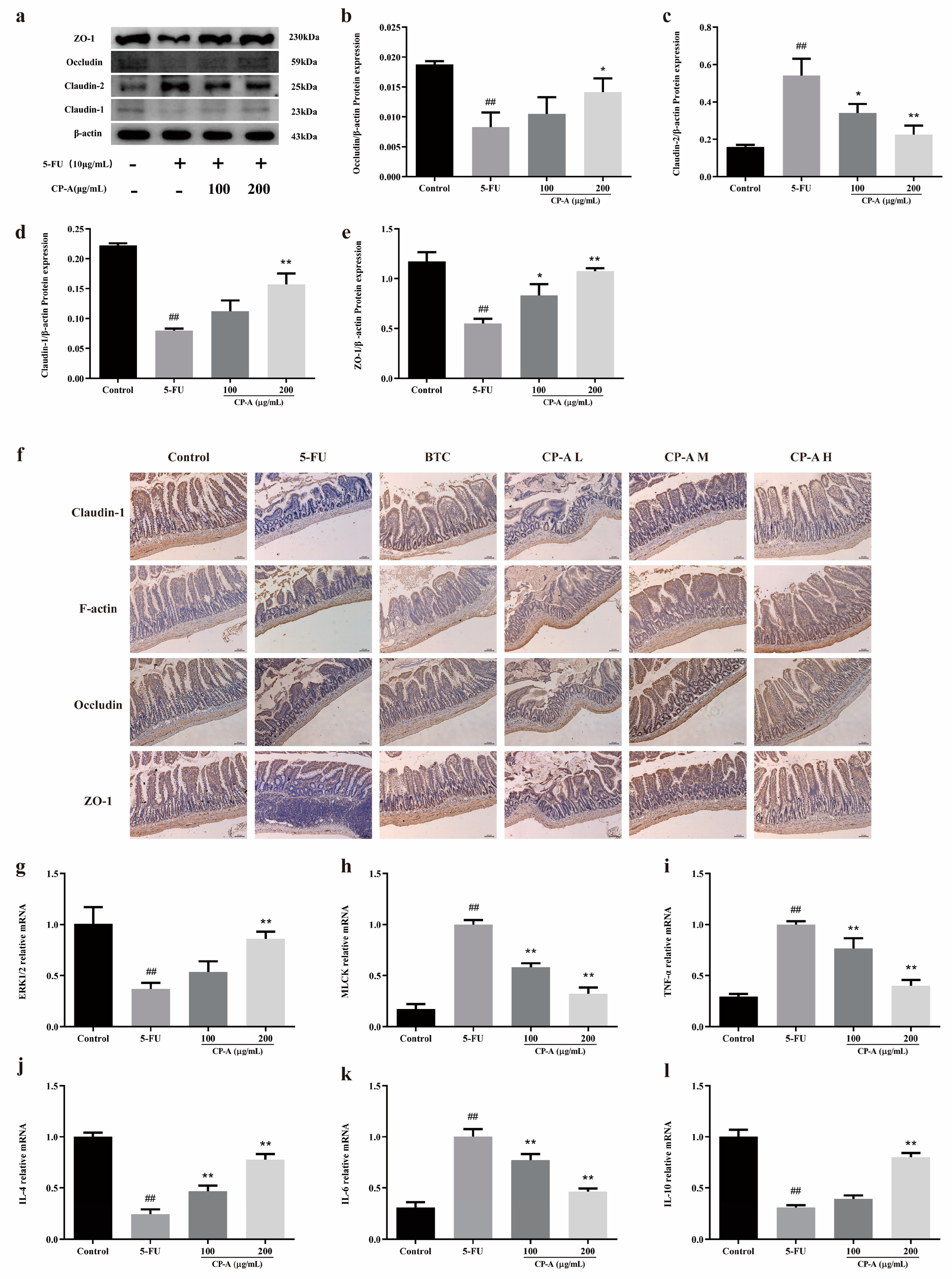
Figure 6.
Effects of CP-A on ASV and microbial diversity analysis. (a) rarefaction curve of Shannon index. (b) Venn diagram of ASVs. (c) Alpha diversity (Chao1 index, Shannon index, Simpson index); (d) Beta diversity (bray-curtis). Values were represented as mean±SEM (n = 6).
Figure 6.
Effects of CP-A on ASV and microbial diversity analysis. (a) rarefaction curve of Shannon index. (b) Venn diagram of ASVs. (c) Alpha diversity (Chao1 index, Shannon index, Simpson index); (d) Beta diversity (bray-curtis). Values were represented as mean±SEM (n = 6).
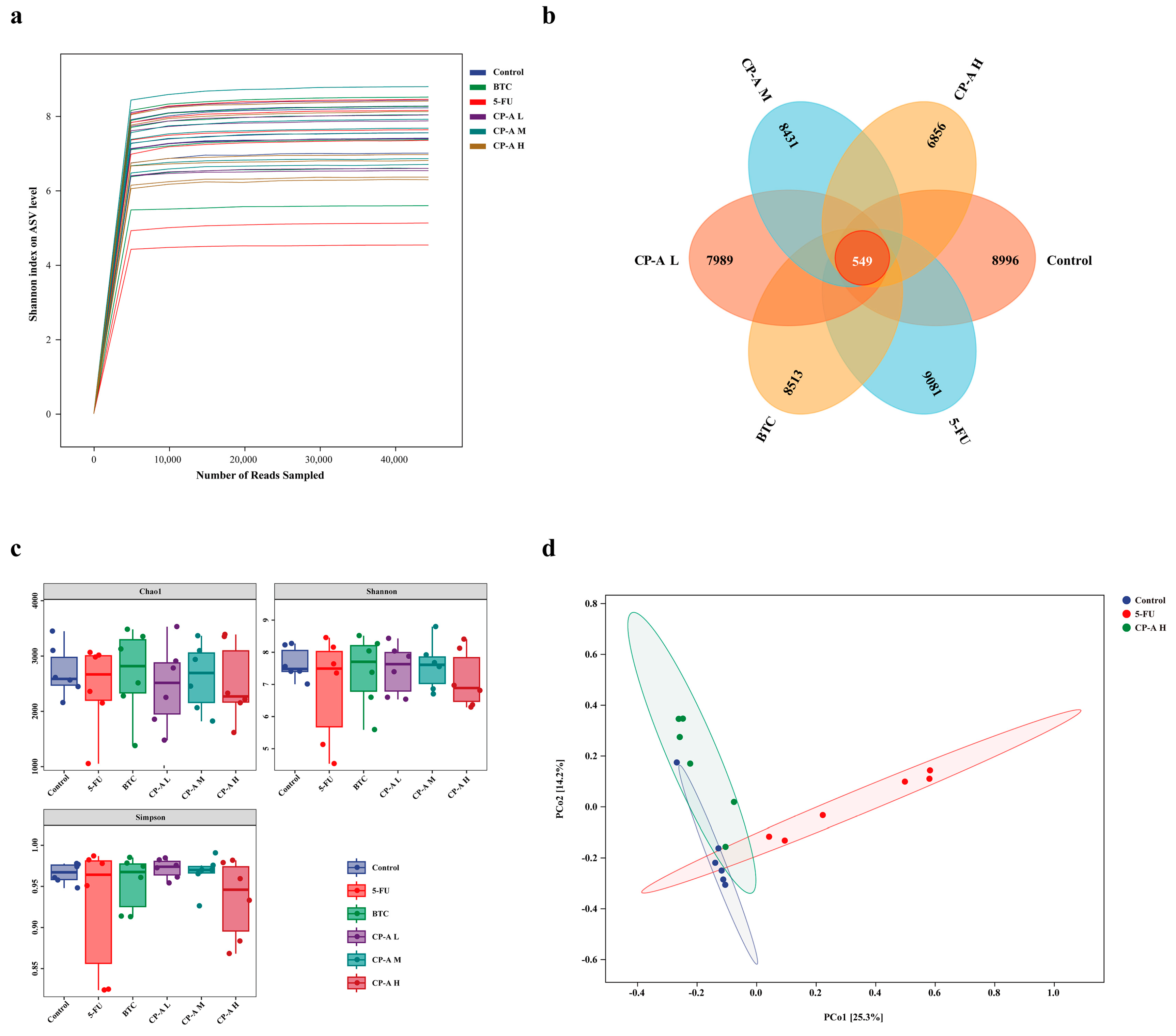
Figure 7.
Taxonomy analysis of microbiota community at Phylum, Class, Order, Family and Genus levels (n = 6).
Figure 7.
Taxonomy analysis of microbiota community at Phylum, Class, Order, Family and Genus levels (n = 6).
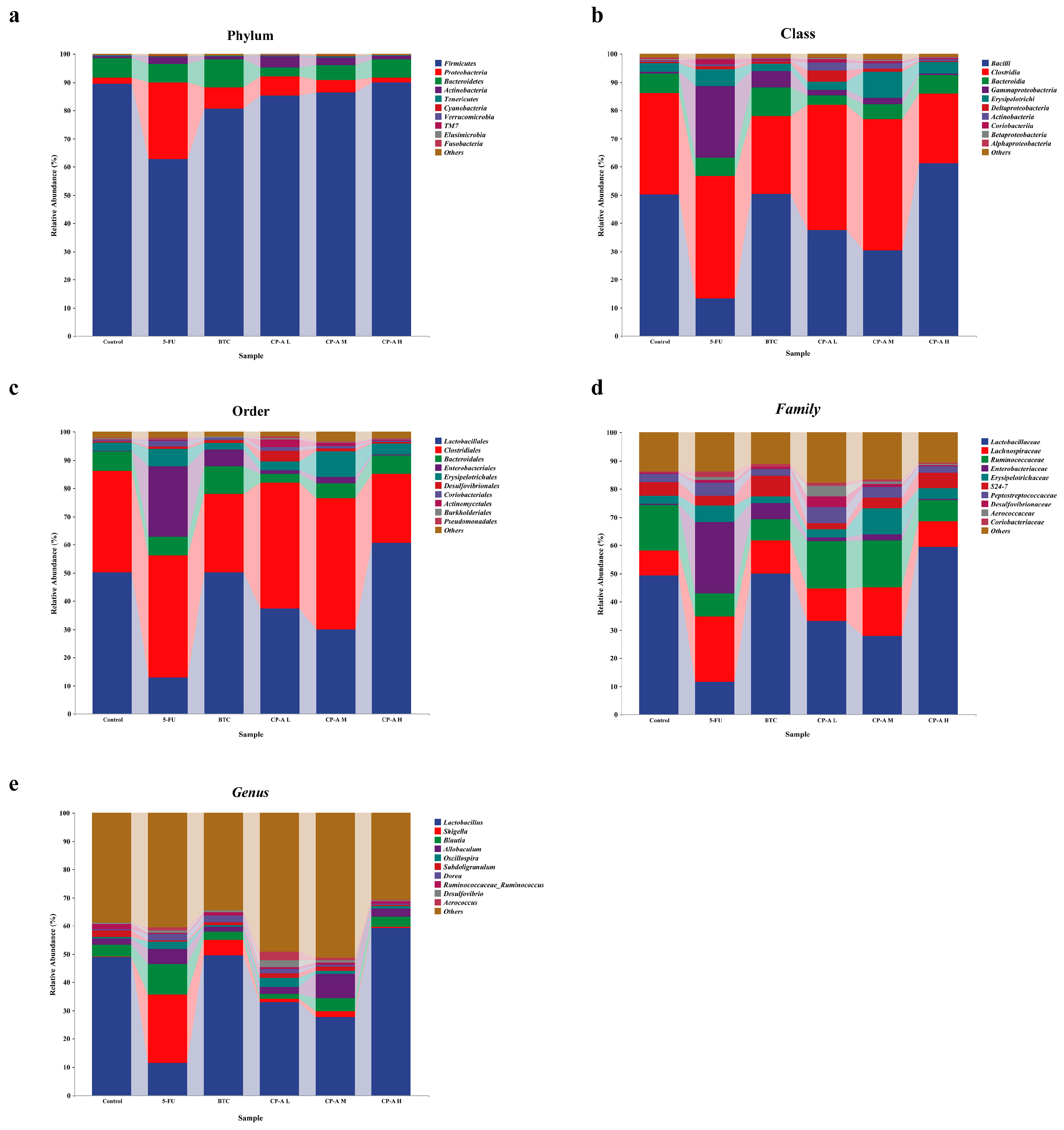
Figure 8.
LEfSe analysis (n = 6) (a) Each group of LEfSe analysis by cladogram. (b) LDA distribution histogram of significantly different species in each group.
Figure 8.
LEfSe analysis (n = 6) (a) Each group of LEfSe analysis by cladogram. (b) LDA distribution histogram of significantly different species in each group.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



