
Submitted:
26 December 2023
Posted:
27 December 2023
You are already at the latest version
Abstract
Parkinson's disease (PD) is a neurodegenerative disorder characterized by the loss of dopamin-ergic neurons responsible for unintended or uncontrollable movements. Mutations in leu-cine-rich repeat kinase 2 locus, contributes to genetic forms of PD. The fruit fly Drosophila mela-nogaster carrying this mutation (LRRK2-Dm), is an in vivo model of PD that develops motor impairment and stands for an eligible non mammalian paradigm to test novel therapeutic ap-proaches. Dehydrozingerone (DHZ) is a natural phenolic compound isolated from ginger. Pre-sents anti-inflammatory, antioxidant and neuroprotective properties making it a potential therapeutic target for PD. We administered DHZ and its C2-symmetric dimer (DHZ-DIM) at 0.5 and 1mM, for 14 and 21 days, in the LRRK2-Dm, with the aim to assess changes for rescuing motor behavior, brain dopaminergic neurons, mitochondria and synapses (T-Bars). The shorter treat-ment with both molecules revealed efficacy at the higher dose improving climbing behavior with a prevention of dopaminergic neuronal demise. After 21 days, a recovery of the motor disability, dopaminergic neuron loss, mitochondrial damage and T-Bars failure was observed with the DHZ-DIM. Our data indicates that the DHZ-DIM exerts a more potent neuroprotective effect respect to the monomer in LRRK2-Dm, prompting further investigation of these compounds in rodent models of PD.
Keywords:
Parkinson’s Disease
; nutraceutical compounds
; dehydrozingerone
; hydroxylated biphenyls
; nat-ural compounds
; Drosophila Melanogaster
; LRRK2
; neuroprotection
1. Introduction
Parkinson’s disease (PD) is an age-related neurodegenerative disorder with typical manifestation of motor symptoms which includes bradykinesia, rigidity, postural instability, and tremor associated to several non-motor symptoms, namely cognitive impairment, depression, sleep disturbance and olfactory deficits [1,2,3,4,5]. Pathological hallmark of the disease is the loss of dopaminergic neurons in the substantia nigra and even though the processes underlying dopaminergic cell death remain unclear as well as the exact etiology. However, some biochemical mechanisms are recognized as contributors to the neuropathology. First of all, an abnormal and toxic intracellular accumulation and aggregation of misfolded proteins, such as α-synuclein and parkin, which converge into the core of Lewy bodies. Moreover, neuroinflammation characterized by reactive microgliosis, oxidative stress caused by overproduction of reactive oxygen species, in combination with reactive metabolites of dopamine, and mitochondrial dysfunction are significantly present [6,7,8] .
With regard to available therapies for PD, effective preventive neuroprotective or disease-modifying cure is currently lacking, highlighting urgent need for novel drugs or alternative strategies at least to halt the progression of the disease. In the recent decades, many studies have adopted a more holistic approach based on metabolic amelioration achieved by specific dietetic programs as possible preventive therapy for neurodegenerative diseases. As matter of fact, many nutraceuticals and food-derived bioactive compounds, in virtue of their intrinsic properties, could have a dramatic role to reduce the risk factors for the arising of chronic neurodegenerative diseases. Focused emphasis has been given for the use of polyphenols, present in most vegetables, e.g. flavonoids, phenolic acids or curcuminoids as therapeutic natural compounds against inflammation, neurodegeneration, and oxidative stress [9,10,11].
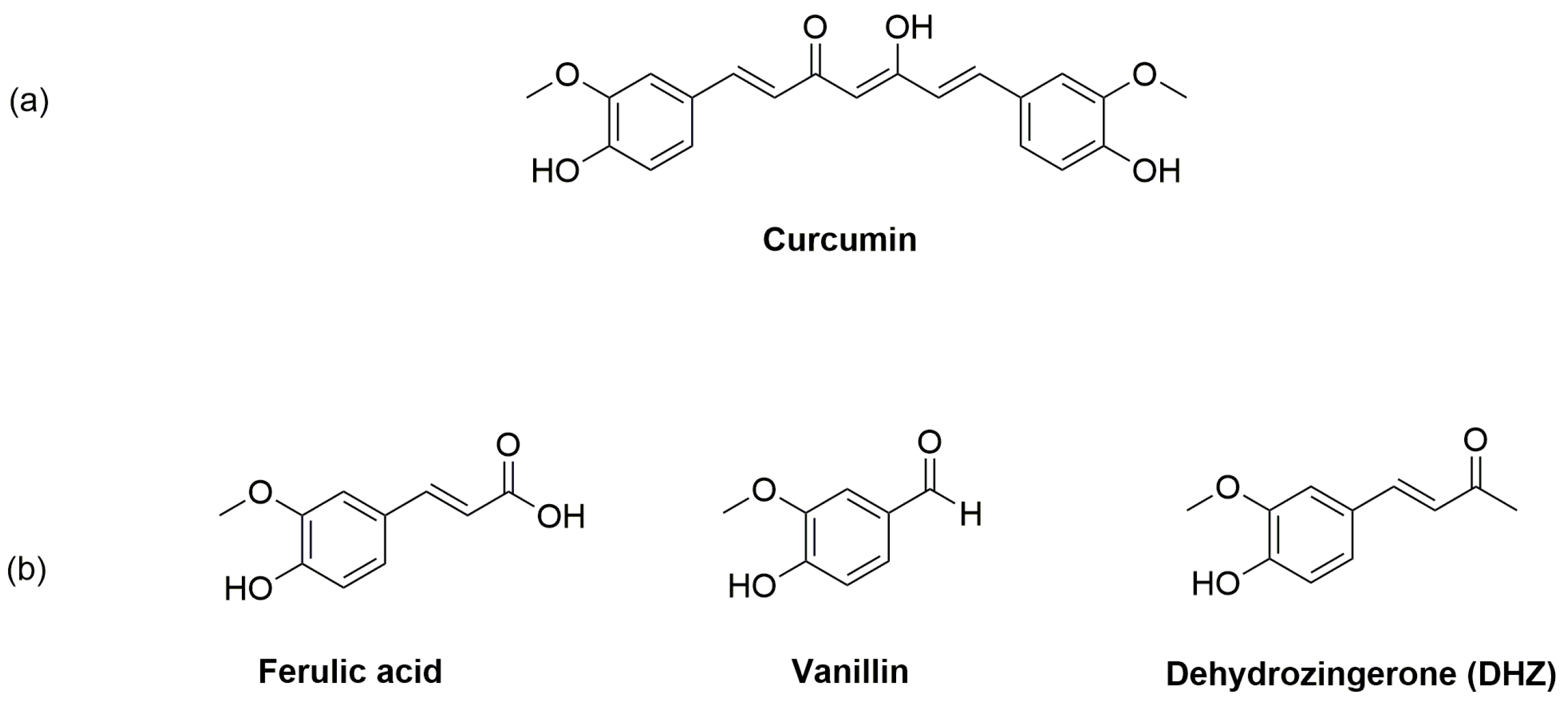
Curcumin (Figure 1a), also known as diferuloylmethane is the active component of Curcuma longa (Zingiberaceae family) rhizome. This rhizome meets large appreciation as a spice in Indian curries and has garnered specific attention for its wide-range of pharmacological activities [12,13,14]. Many studies in vitro and in vivo showed that curcumin possesses neuroprotection properties [15,16]. Moreover, it has been reported that curcumin is able to suppress PD-like phenotypes in flies [17,18,19].
Unfortunately, the pharmacological potential of curcumin, is widely restricted because of its poor bioavailability due to its, chemical instability, and rapid metabolic degradation into ferulic acid, vanillin and dehydrozingerone (DHZ) at physiological pH (Figure 1b) [17]. Therefore, it makes sense to explore the development of curcumin derivatives with enhanced bioavailability with consequent translational benefits to hinder PD by identification and production of more potent compounds in the context of phytotherapic options.
As above mentioned, DHZ, a structural half analogue of curcumin (CUR), is a natural phenolic compound extracted from ginger (Zingiber officinale) rhizome that exhibits enhanced solubility and stability in water compared to CUR. This property gives to DHZ a tremendous advantage for biomedical applications where water solubility and stability are crucial factors.
It has been demonstrated that DHZ, exhibits many biological activities and pharmacological properties, including anti-inflammatory, antioxidant, anti-obesity, anticancer, tyrosinase inhibitory, neuroprotective, antidepressant and anti-fungal effects [18,19,20,21,22,23,24,25,26,27].
Often, hydroxylated biphenyls, have higher biological activities respect to the corresponding monomer. Hydroxylated biphenyls are widely distributed in the plant kingdom and play an important role in biosystems due to their unique pharmacophore structure that is made up of two aromatic rings bridged by a single C–C bond. The high level of specificity of the hydroxylated biphenyls scaffold with the catalytic domain of different proteins [28,29,30]. Consequently, the hydroxylated biphenyl framework offers an ideal molecular structure for structural modifications in the development of potential drug candidates [31].
In prior studies, we established that both DHZ and its symmetric dimer (DHZ-DIM) exhibit protective effects against lipid autoxidation [32] which is an important factor in the development of neurodegenerative disorders such as Alzheimer's and Parkinson's disease. Furthermore, DHZ-DIM exerted a potent anti-inflammatory, antioxidant, and antithrombotic activity on endothelial cells in combination with antiaggregating and cytoprotective properties as demonstrated by its ability to partially inhibit the aggregation process of alpha-synuclein [33,34]. For these reasons DHZ and DHZ-DIM could represent interesting candidates to reverse the symptoms of PD.
Although most PD cases seem sporadic, genetic factors may play a role in favoring the disease, and mutations in several specific genes have been related with familiar forms of PD. Among them, mutations in the leucine-rich repeat kinase 2 (LRRK2) gene have been correlated to late-onset autosomal dominant PD. Pathological mutations of LRRK2 have been found not only in 13% of familial forms, but also in approximately 1-2% of idiopathic PD cases [35,36]. Furthermore, familial forms related to mutations in the LRRK2 gene show clinical symptoms indistinguishable from idiopathic forms of PD [37].
LRRK2 is a protein widely expressed within different brain areas, such as the cortex, striatum, hippocampus, cerebellum, and substantia nigra [38,39]. It is involved in GTPase, and kinase activities associated to signal transduction cascades for synaptic vesicle trafficking, mitochondrial metabolism, and autophagy [40,41]. LRRK2 holds a double enzymatic core in the N-terminal and the C-terminal WD40 domain [42]. In particular, the coding variants G2019S, diffuse in Caucasian individuals [43], and G2385R present in Asian population [44], in the WD40 domain, resulted in about 50% loss of kinase activity leading to a partial loss-of-function of LRRK2 [45]. These missense substitutions double the risk of getting sick with PD [46,47,48].
Regarding to the PD genetic approach, Drosophila melanogaster (Dm), commonly known as the fruit fly, is a powerful translational animal model for studying neurodegenerative diseases because of carrying nearly 75% homology with human disease genes [49]. Moreover, Dm has several advantages compared to mammalian models, from expandable population, quick life cycles, and easy genetic manipulation, to low maintenance costs, and less rigid ethical concerns. Notably, most of the genes implicated in familial forms of PD have an evolutionary counterpart in Dm [50,51]. Therefore, Dm carrying the LRRK2 loss of function mutation in the WD40 domain [52] develops the essential traits of the pathology such as motor impairment, dopaminergic neuronal cell loss and mitochondrial abnormalities [53,54,55] providing precious information regarding PD pathophysiology mechanisms and a bona fide tool to firstly test novel therapeutic approaches to the disease.
Thus, the aim of this paper was to investigate the neuroprotective potential of DHZ and DHZ-DIM, prepared following new sustainable procedures, carrying out the reactions under microwave irradiation. These two compounds were tested on the Drosophila PD strain LRRK2 loss-of-function (LRRK), compared to Wild-Type (w1118) in terms of physiological and brain morphological parameters that are severely compromised in the mutant flies, such as longevity, motor activity, integrity of brain dopaminergic neurons as well as brain mitochondria and synapses abnormalities. After treatment with both molecules at the dose of 0.5-1 mM and Vehicle for 14 and 21 days after eclosion, there was a significant amelioration of motor performances, and a prevention of dopaminergic neuron loss, mitochondrial damage, and synapses (T-Bars) failure in LRRK mutant. Specifically, our data indicate that the DHZ-DIM exerts a more potent neuroprotective effect with respect to the monomer in this fruit fly PD model suggesting that these curcumin-related compounds could be promising medicaments for novel therapeutic scenario toward LRRK2-linked PD.
2. Materials and Methods
2.1. General
Reagents were obtained from Sigma Aldrich (Munich, Germany) and were used without further purification. Microwave reactions were carried out on a MW instrument CEM-Discover SP MW (Matthews, NC, USA). 1H-NMR and 13C-NMR spectra were recorded in CDCl3 at 600 and 150 MHz, respectively, with a 600 MHz NMR spectrometer Bruker Avance III HD (Palo Alto, CA, USA). Chemical shifts are given in ppm (δ); multiplicities are indicated by s (singlet), d (doublet), or dd (doublet of doublets). Elemental analysis was performed using an elemental analyser model 240 C Perkin Elmer (Walthan, MA, USA). Flash chromatography was carried out with silica gel 60 (230–400 mesh) VWR (Radnor, AF, USA) eluting with the appropriate solution in the stated v:v proportions. Reaction was monitored by analytical thin-layer chromatography (TLC) with 0.25 mm thick silica gel plates 60 F 254 Sigma Aldrich (Munich, Germany). Melting point was determined on a 530 apparatus Büchi (Flawil, Switzerland) and is uncorrected. The purity of new compounds was judged to be >98% by 1H-NMR spectral determination.
2.2. Fly Stocks
Adult wild type (WT; w1118) and LRRKWD40 (LRRK) mutant (LRRKex1, #34750, from Bloomington Stock Center) Dm males were used. After emergence from pupae, WT or LRRK mutant males were separated from females. WT and mutant flies were reared on a standard cornmeal-yeast-agar medium in controlled environmental conditions (24-25°C; 60% relative humidity; light/dark = 12/12 hours). For the treatment, groups of mutant and WT flies were reared on a standard medium supplemented with two concentrations of DHZ and DHZ-DIM (0.5 and 1 mM) for 14 and 21 days.
2.3. Drugs
DHZ and DHZ-DIM were dissolved in DMSO (final concentration 0.5%) and added to the diet at the concentrations reported.
2.4. Climbing Assay
The climbing assay (negative geotaxis assay) was used to assess locomotor ability [56] WT and LRRK mutants were treated for 14 or 21 days with DHZ, DHZ-DIM or Vehicle. Cohorts of at least 30 flies from each group, in three independent experiments, were subjected to the assay. Flies were placed individually in a vertically positioned plastic tube (length 10 cm; diameter 1.5 cm) and tapped to the bottom. Climbing time (sec) was recorded upon crossing a line drawn at 6 cm from the bottom. The number of flies that could climb unto or above this line within 10 seconds was recorded and expressed as a percentage of total flies tested. Data were expressed as the average ± standard error of the mean (SEM) from three experiment replications.
2.5. Survival Curves
According to previous reports [57] WT and LRRK male flies were separated from females after emergence from pupae under CO2 anesthesia. Cohorts of 60 flies from each group, were collected in groups of 10-15 in vials containing standard diet with drugs at 1mM or VEH, monitored daily at 25°C and changed with frequency throughout adult life. Data were collected from eclosion to death. For mortality analysis, Kaplan-Meier survival curves and statistical comparisons Log-rank (Mantel-Cox) test and Gehan-Breslow-Wilcoxon test were utilized.
2.6. Immunohistochemistry
Six to ten flies from each experimental group were selected to performed free-floating fluorescent immunostaining for Tyrosine Hydroxylase (TH). Animals were anesthetized on ice, and brains were rapidly dissected and fixed in 4% paraformaldehyde in phosphate buffered saline (PBS) for 2 hours. Brains were then incubated with the TH rabbit primary antibody (1:100; AB 152 Millipore) and 10% normal donkey serum in PBS + 0.3% Tween 20 (PBST), at 4 °C for 72 h. After rinsing, the brains were incubated with a donkey anti-rabbit Alexa Fluor 594 secondary antibody (1: 200 Jackson ImmunoResearch) and 10% normal donkey serum in PBST at 4 °C for another 72 h. Subsequently, the brains were mounted on glass slides and coverslipped with Vectashield and analyzed under a fluorescence spinning disk confocal microscope (Crisel Instruments). The brains were scanned through Z-stacks (63X objective, stack thickness 0.5 µm) and the number of TH-positive neurons of different clusters (PPL1, PPL2, PPM1/2, PPM3) in both hemispheres, was counted via NIH ImageJ software.
2.7. Electron Microscopy Analysis
The electron microscopy studies were performed in strict accordance with the general methodological procedures indicated by Casu et al. (2020) [55].
Briefly, flies from each experimental group (n=5), were anesthetized on ice, and brains were rapidly dissected and fixed in a mixture of 1% paraformaldehyde and 1.25% glutaraldehyde in 0.15 M cacodylate buffer for 2 hours. Brains were then post-fixed with 1% osmium tetroxide for 1 h, and stained overnight with 0.5% uranyl acetate at 4°C. After dehydration in a graded acetone series, brains were embedded in EPON resin. To identify the protocerebrum, where the dopaminergic posterior clusters reside, 1 µm semi-thin coronal sections of the whole brain were stained with toluidine blue. Ultrathin sections (90 nm) cut with a Reichert Supernova ultramicrotome, were counterstained with uranyl acetate and lead citrate and observed under a JEOL JEM 1400 Plus electron microscope, equipped with a CCD camera, at an acceleration voltage of 80 kV.
Morphometric analysis: the mitochondria (total number), the percentage of mitochondria with swollen cristae (percentage of mitochondria displaying swollen cristae versus total mitochondria with discernible cristae) and the T-bars density was analyzed in the unitary area (25 µm2) in the protocerebrum. Thirty to forty unitary fields were evaluated for each brain. In total, about 17500 mitochondria and 4000 T-bars were randomly sampled on 792 non-overlapping micrographs at a magnification of 8000X. Swollen cristae were considered when the distance between two contiguous membranes of one crista, doubled the average cristae size. T-bars were unambiguously identified at presynaptic active zones by the presence of T-shaped electron-dense projections.
2.8. Statistics
Data are presented as means ± SEM. Two group comparisons have been analyzed by factorial two-way ANOVA with the Strain and Treatment as between group factors. Before performing the analyses, datasets have been checked for normal distribution by using the Shapiro-Wilks test and for homogeneity of variances between the experimental groups with the Bartlett’s test. When the normal distribution of data and homogeneous variances across the experimental groups was obtained in all datasets, thus, the factorial ANOVA was applied.
In all the other cases, when the transformation data didn’t reveal homogeneity of variances, the non-parametric analysis by Kruskal-Wallis comparisons test was performed. When parametric two-way ANOVA revealed statistically significant interactions, sources of significance were ascertained by pairwise post-hoc analyses by using the HSD Tukey’s test. For mortality analysis, Kaplan-Meier survival and statistical comparisons Gehan-Breslow-Wilcoxon test were used. Statistical analyses were all carried out with PRISM, GraphPad 8 Software, with the significance level set at P < 0.05.
3. Results
3.1. Chemistry
DHZ and DHZ-DIM were synthesized with comparable yields and purity, employing a method previously outlined by our group [34]. Notably, we followed more sustainable procedures, utilizing microwave irradiation to significantly reduce the reaction time from 12 hours to just 30 minutes. DHZ was prepared by Claisen-Schmidt condensation reaction of vanillin and acetone in presence of 1N NaOH as base (90% yield). DHZ-DIM was prepared under the same conditions starting from vanillin dimer [34] and acetone in presence of 1N LiOH as base (83% yield) (Scheme 1).
3.1.1. Synthetic Procedures
[(E)-4-(4-hydroxy-3-methoxyphenyl) but-3-en-2-one] (DHZ)
An aqueous solution of NaOH 1N (5.2 mL, 5.2 mmol) was added to a vanillin solution (0.2 g, 1.3 mmol) in acetone (7 mL). Reaction mixture was stirred under MW irradiation at 100 °C for 30 min. The solvent was then roto-evaporated and 10% HCl were cautiously added. The obtained heterogeneous solution was extracted with ether, the organic phase dried over Na2SO4 and evaporated to afford a brown solid compound. The crude material was finally purified by flash chromatography using of CH2Cl2 as eluent, to give DHZ as yellow solid (0.23 g, 90 %): mp 126–127 °C; 1H NMR δ 2.39 (s, 3H), 3.85 (s, 3H), 5.99 (bs, 1H), 6.52 (d, J = 16.0 Hz, 1H), 6.90 (d, J = 8.0 Hz, Ar, 1H), 7.00 (d, J = 1.6 Hz, Ar, 1H), 7.06 (dd, J = 1.6, 8.0 Hz, Ar, 1H), 7.40 (d, J = 16.0 Hz, 1H); 13C NMR δ 27.29, 56.93, 109.30, 114.80, 123.94, 124.99, 126.95, 143.76, 146.84, 148.26, 198.46; Anal. Calcd for C11H12O3: C, 68.74; H, 6.29; Found: C, 68.93; H, 6.41.
[(3E,3′E)-4,4′-(6,6′-dihydroxy-5,5′-dimethoxy-[1,1′-biphenyl]-3,3′-diyl) bis(but-3-en-2-one)] (DHZ-DIM)
An aqueous solution of LiOH 1N (4.0 mL, 4.0 mmol) was added to vanillin dimer (0.2 g, 0.66 mmol) in acetone (5 mL). Reaction mixture was stirred under MW irradiation at 100 °C for 30 min. The solvent was then roto-evaporated and 10% HCl were cautious-ly added. The obtained precipitate was filtered, washed with water and dried to afford a DHZ-DIM as yellow solid compound (0.18 g, 83 %): mp 242–243 °C; 1H NMR δ 2.35 (s, 6H), 3.99 (s, 6H), 5.28 (bs, 2H), 6.61 (d, J = 16.0 Hz, 2H), 7.1 (d, J = 2.0 Hz, Ar, 2H), 7.15 (d, J = 2.0 Hz, Ar, 2H), 7.47 (d, J = 16.0 Hz, 2H); 13C NMR δ 27.35, 56.20, 108.67, 123.59, 125.28, 125.44, 126.60, 143.52, 145.45, 147.36, 198.28; Anal. Calcd for C22H22O6: C, 69.10; H, 5.80; Found: C, 69.47; H, 5.70.
3.2. DHZ and DHZ-DIM Prevent the Motor Impairment in LRRK: Climbing Test
Groups of LRRK and WT flies received both DHZ or DHZ-DIM at the two doses of 0.5 and 1mM in their diet for 14 and 21 days to evaluate the effect of both molecules on locomotor ability. The climbing activity and the percentage of flies reaching the target within 10s was measured. Remarkably, after 14 days of both molecules’ treatment, it was observed a significant improvement of the climbing behavior of LRRK only with the highest dose of 1 mM if compared to LRRK vehicle (p< 0.0001 Figure 2a-b). On the other hand, at the same dose (1 mM) only DHZ-DIM was able to induce a recovery of the motor disability after 21 days of treatment (p<0.0001). The administration of the same drugs at both concentrations did not significantly affect in the w1118 (Figure 2a-b-c-d).
In search for an additional parameter of motor ability, we counted the percentage of WT and LRRK flies completing the climbing test within 10 s (Figure 3). The resulting relative percentages confirmed the improvement in mutants treated for 14-21 days, regardless of DHZ and DHZ-DIM at the concentration of 0.5 and 1 mM. In addition, the DHZ-DIM was more effective at 1 mM highest dose at both timeline treatment.
3.3. DHZ and DHZ-DIM Extended Longevity
Assessment of fly lifespan by means of survival curves, allowed monitoring the effect of drugs on Dm survival throughout adulthood. LRRK displayed a shorter life span compared to WT w1118 (p < 0.0001, Figure 4a) since mutant flies encountered a dramatic decay in their survival rate (about 50%) at 35-40 days after enclosure (with median survival at 47 days). In the attempt to prolong the LRRK lifespan, aiming at verifying to that extent the studied molecules were ad hoc effective, mutant flies were supplied with DHZ and DHZ-DIM at the highest concentration of 1 mM and compared with vehicle-treated flies. Both compounds significatively extended longevity of LRRK (p < 0.0001, Figure 4b) although flies treated with DHZ-DIM showed a lower decay in the survival rate with the median survival about 70 days with respect to DHZ (around 54 days). Moreover, a small group of LRRK flies (about 20%) treated with the dimer achieved a survival rate at 80 days like w1118.
3.4. DHZ and DHZ-DIM Prevent the Loss of Dopaminergic Neurons
After the climbing behavioral test, flies were sacrificed to evaluate the effect of DHZ and DHZ-DIM treatment on TH positive neurons in the brain posterior dopaminergic clusters (PPL1, PPL2, PPM1/2 and PPM3 (Figure 5). Since both compounds displayed a maximal behavioral effect at 1 mM concentration, only brains from flies exposed for 14 and 21 days to the above concentration of DHZ and DHZ-DIM were processed for TH immunohistochemistry.
The quantitative analysis of TH immunofluorescent neurons showed that both compounds at 1 mM significantly prevented the loss of dopaminergic neurons in all four posterior clusters after 14 days of treatment (p<0.0001). However, at 21 days, a significant prevention of dopaminergic neuronal demise was surprisingly detected, only with the DHZ-DIM treatment (p<0.0001) (Figure 6).
3.5. DHZ-DIM is More Effective than Monomer in Preventing the Mitochondrial Damage and the Loss of T-Bars in LRRK2 Drosophila
Transmission electron microscopy (TEM) analysis was conducted on LRRK and WT brain flies treated with vehicle, DHZ or DHZ-DIM at the dose of 1 mM for 21 days since immunohistochemical results showed that both compounds elicited the maximal effects at 1 mM concentration, with particular reference at the latest time point treatment. As previously described, ultrastructural brain morphology showed axons and terminal boutons, mitochondria and T-bar presynaptic densities (Figure 7a-b-c-d). In particular, in vehicle-treated LRRK brain flies several mitochondria displaying swollen cristae were present (Figure 7a).
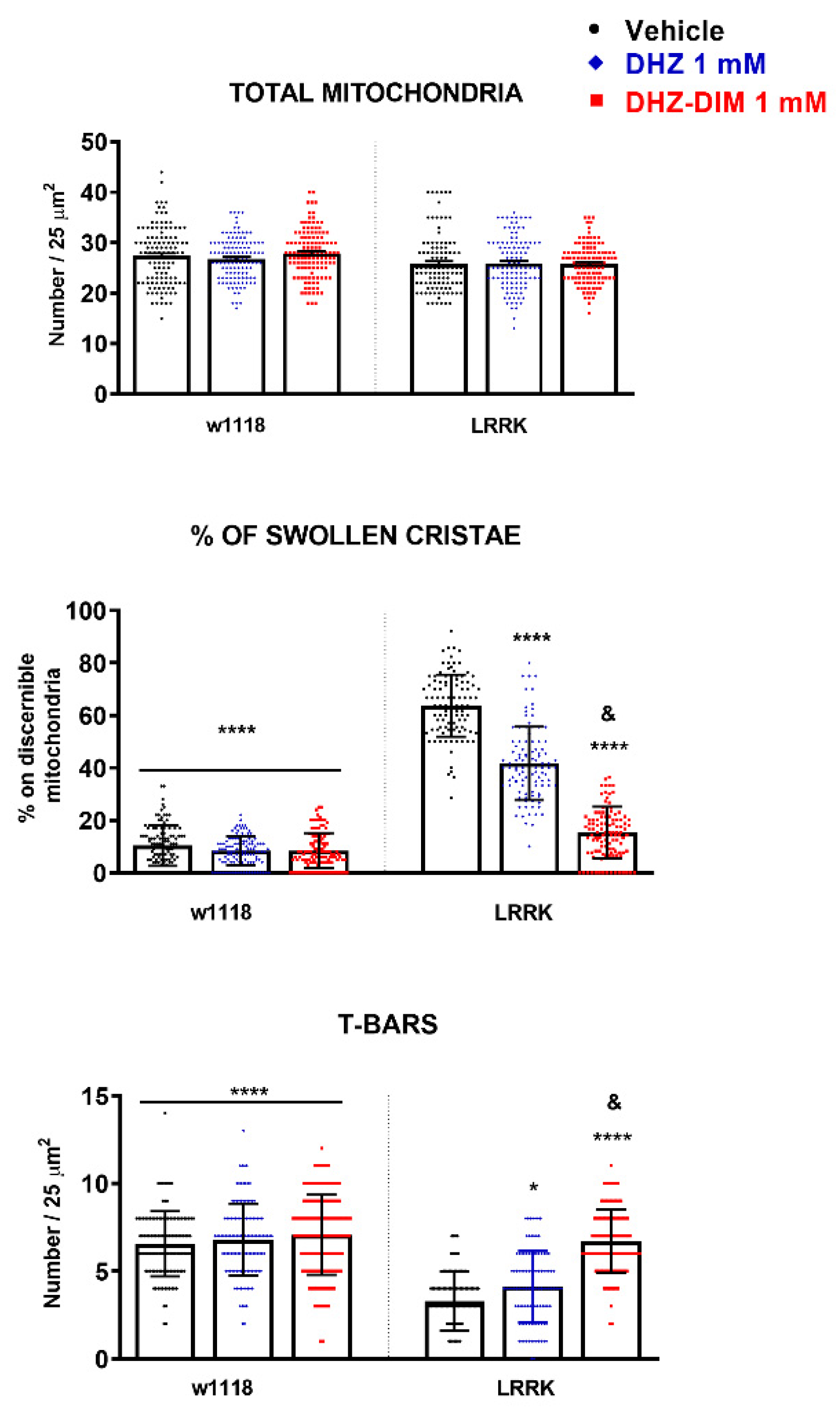
The morphometric analysis did not reveal any difference in the number of total mitochondria between control and LRRK treated with DHZ or DHZ-DIM (Figure 8). The administration of both substances clearly decreased the occurrence of mitochondria with swollen cristae as compared with vehicle administration, (p < 0.0001) and, more relevant, in accordance with other morphological and functional parameters, the treatment with DHZ-DIM was more worthwhile than DHZ in avoiding the formation of swollen mitochondria (Figure 8). Similarly, using both compounds, it was observed a significant increase in the number of T-bar in the presynaptic bouton active zones as compared to vehicle treatment (p < 0.01; Figure 8) but, definitely, the contribution of the dimer resulted more impactful. By contrast, the exposure of WT flies to both monomer and dimer did change neither mitochondria number and morphology, nor the number of T-bars.
4. Discussion
In the present study we investigated the neuroprotective activity of the two curcumin derivatives DHZ and its C2 dimer in a transgenic Drosophila model of PD. We improved process sustainability of DHZ and DHZ-DIM synthesis carrying out the reactions under microwave (MW) irradiation. As matter of fact, MW technology presents numerous advantages in various chemical synthesis processes. This efficiency is attributed to the ability of MW in heating the reaction mixture both rapidly and uniformly, promoting faster and more complete chemical transformations compared to conventional methods. Moreover, this technology minimizes the risk of hazardous reactions or the release of volatile substances, contributing to a safer working environment. The utilization of MW aligns with the broader goals of sustainability, promoting energy efficiency and reducing overall environmental impact. To our knowledge, this is the first study evaluating DHZ and DHZ-DIM as neuroprotective agents using an in vivo model of PD. Our data demonstrate that the exposure to both compounds prevented motor deficits and protected against the progressive loss of dopaminergic neurons. The neuroprotective efficacy of the DHZ and DHZ-DIM could be related to their antioxidant action since in previous studies, we found that both DHZ and its symmetric dimer (DHZ-DIM) exhibit protective effects against lipid autoxidation when used in combination with conventional antioxidants [32]. The antioxidant activity of DHZ-DIM is also associated with antiaggregating and cytoprotective properties ascertained by its ability to partially inhibit the aggregation process of 𝛼-synuclein [34].
The methodological approach to the morphological study, has taken into account that in the Drosophila brain, DA neurons are organized in distinct bilateral symmetric clusters with projections onto specific brain areas [58,59]. These dopaminergic neurons of posterior clusters, analyzed in this study, such as PPL1, PPM1/2 and PPL2 innervate distinct regions of the mushroom bodies that are implicated with learning and memory [60]. Moreover, PPM3 neurons innervate the central complex, which is the area related to the control of motor activity.
Therefore, the neuronal rescue of the above anatomical circuits upon DHZ and DHZ-DIM directly correlates to the protective role for contrasting those brain areas dramatically affected by the progression of PD. In this context, DA neurons’ degeneration in PD is further boosted by oxidative stress mechanisms involving also DA facing a rapid oxidation. The Dopamine autoxidation produced dopamine quinones and free radicals. Moreover, the cyclization of dopamine quinones forms aminochrome, which generates superoxide and down-regulates antioxidative nicotinamide adenine dinucleotide phosphate (NADPH) [59,60]. The susceptibility of the brain to oxidative stress is augmented by various factors such as high oxygen demands, higher rates of oxidative metabolism and lower levels of protective antioxidant system.
This critical scenario is further worsened in the PD genetic models, where LRRK2 mutation caused increased generation of ROS and cell toxicity. A proof of concept has been offered by a recent in vitro work showing that LRRK2 knockout provides resistance to oxidative stress and apoptosis suggesting LRRK2 as a proapoptotic kinase [61]. Moreover, previous studies demonstrated that deletion of the WD40 domain prevents autophosphorylation [62,63] and, in the other hand, the G2385R polymorphism in the WD40 domain, expressed in our drosophila PD model, increases the sensitivity of cells to hydrogen peroxide suggesting a pro-apoptotic mechanism [64]. Considering that our compounds act as antioxidants the observed neuroprotective ability on dopaminergic neurons well matched with the prevention on motor impairment in mutant flies.
It is important to note that after 14 days of treatment, DHZ and its dimer both effectively hindered the symptoms of Parkinson’s diseases in LRRK flies but only after a longer treatment of 21 days, the DHZ dimer is superior to the monomer in avoiding motor impairment and loss of dopaminergic neurons. These results are reasonably related to the aging progression that, by wide scientific consensus, is one of the major risk factors for developing PD. This is also suggested by the fact that there are many common features between PD and normal aging [65] including protein aggregation [46], increased oxidative stress [66], decreased mitochondrial function [67], dysfunction of the proteasome [68], and impairment of autophagy [69]. Therefore, in aged parkinsonian flies, the DHZ-DIM demonstrates greater efficacy, compared to the monomer. This enhanced DHZ-DIM activity could be ascribed to the differences in the chemical structure and lipophilicity between the two molecules making sense to draw the hypothesis that the DHZ-DIM has a higher ability to cross the cell membrane and to interact with cell components more efficiently than the corresponding monomer. It has been also demonstrated the superior ability of DHZ-DIM in protecting lipids from autoxidation and, additionally, its higher antioxidant properties and reactivity when compared to its corresponding monomer [32,70].
In our study, DHZ-DIM has been proved to be more decisive than monomer also in extending longevity of parkinsonian flies. This finding replicates the conclusion of several studies showing that dietary supplementation with compounds rich in polyphenols such as Avocado Persea americana, grape and grape seed extracts, gallic acid and very high doses of curcumin, enhanced the lifespan of drosophila models of Parkinson disease [71,72,73]. Polyphenols can delay oxidative reactions in cells by rapidly donating protons to radicals or by forming complexes with pro-oxidant metals. Furthermore, polyphenols can interact with receptors or enzymes in signal translation, promoting an antioxidant condition [10,74].
The neuroprotective effect on motor improvement and brain dopaminergic neurons of our compounds fits very well with the reduced mitochondrial damage in LRRK brain flies detected in our investigation confirming previous findings related to the presence of damaged mitochondria in LRRK mutant flies, as indicated by multiple dilated cristae [55]. On that regard, oxidative phosphorylation within the mitochondria accounts for the majority of ATP production in neurons required for the transmission of nerve impulse. Various studies have suggested that mitochondrial dysfunction can contribute to increased levels of oxidative stress and can affect neuronal degeneration [75,76,77]. In postmortem studies, high oxidation of proteins and DNA stimulated parallel levels of lipid peroxidation so that reduction of glutathione has been found in the substantia nigra in PD patients [78]. Moreover, mitochondrial complex I inhibition has been observed of PD patients suggesting that the increased presence of ROS through complex I inhibition is one of the major contributors for the DAergic neuronal cell death in PD, reinforcing the concept that this type of stress is dramatically involved in the pathology [1]. On top of such considerations, it cannot be overlooked that several genetic mutations, including LRRK2, are linked to mitochondrial dysfunction in PD pathogenesis. For example, LRRK2 interacts with the mitochondrial fusion proteins and mitochondrial outer membrane proteins [79,80]. LRRK2 mutations determined alterations in mitochondrial fusion and fission mechanisms, mitophagy and in mitochondrial DNA damage [6,81] all elements that increase ROS production, inhibition of peroxidase activity, and a consequent increment of oxidative stress [82].
The association of mitochondrial dysfunction and production of ROS represent a potential target for treating PD. Mitochondria-targeted antioxidants and flavonoids such as alpha-lipoic acid, hesperidin (flavanone rich in citrus), the flavonoid baicalein, the carotenoid lycopene and CUR have produced positive outcomes in vitro and in vivo studies. Indeed, it has been demonstrated that these compounds can act on mitochondrial integrity, ATP production, mitochondrial membrane potentials, GSH levels, halting increased ROS production and apoptosis and mitigating mitochondrial alterated mechanisms [83,84,85,86,87,88,89].
In our study, after 21 days of treatment, DHZ-DIM has been proved to be more effective than monomer in reducing mitochondrial damage in LRRK brain flies suggesting that DHZ-DIM could act suppressing mitochondrial dysfunctions as already demonstrated for the above antioxidants.
Although numerous studies in cell and animal models support the potential of antioxidants in treating PD, many of these results cannot be reproduced in humans.
As a matter of fact, clinical trials have not demonstrated any efficacy of creatine or coenzyme Q10 in patient with PD [90,91]. A possible explanation for these negative clinical results is that oxidative stress could be a downstream effect of mitochondrial dysfunction rather than a direct cause of PD neurodegeneration. Otherwise, novel drug delivered approaches may be required.
Finally, we noticed the recovering in the loss of T-bars in the mutant flies after the chronic treatment with DHZ and more significatively pronounced with DHZ-DIM. T-bars are the presynaptic active zones involved in neurotransmitter release in Drosophila [92]. LRRK2 binds synaptic vesicles through specific protein-protein interactions in the WD40 domain [93]. Remarkably, synaptic proteomic analysis showed that the G2385R variant impairs LRRK2 binding to key synaptic proteins including synapsin which may explain the loss of T-bars in the LRRK2 WD40 Drosophila model [48]. Moreover, evidence supports the role of mitochondria in synaptic plasticity by maintaining cytosolic calcium within physiological ranges [94]. Therefore, also a reduced mitochondrial antioxidant function could be linked to synaptic loss. The neuroprotective effect of DHZ-DIM could be related to its capacity in preventing mitochondrial damage and synaptic loss by its antioxidant activity.
5. Conclusions
In summary, the overall findings from this study indicated that DHZ-DIM, more than its monomer, possessed a strong neuroprotective effect through its ability to ameliorate the PD-like phenotypes in our Drosophila PD model. If validated in mammalian models of PD, it could be considered a promising compound for the design and development of novel nutraceutical agents with neuroprotective properties to be evaluated in humans. However, on a daily basis, it should be also considered both the additional demands to counteract pathological conditions and the practical inability to provide with dietary intake the necessary “overdose” of nutraceuticals present only in traces in available aliments possibly decisive for a real impact against neurodegenerative diseases.
Although there is strong evidence that suggests to what extent dietary intake of natural compound with antioxidant and anti-inflammatory properties may inhibit neurodegeneration in PD, more detailed studies are needed exploiting alternative neurodegenerative disease models in both mammals and Drosophila also in combination with clinical trials. These further experiments will contribute to give hope for future effective therapies against PD with natural compounds.
Author Contributions
Conceptualization, M.A.C., I.M, M.D.S and M.A.D.; methodology, M.A.C., I.M, M.D.S, M.A.D, D.F. and P.C.; formal analysis, I.M, M.D.S. D.F. and P.C.; investigation, M.A.C., I.M, M.D.S, M.A.D, D.F., P.C. and P.M.; resources, M.A.C., M.A.D.; data curation M.D.S, I.M., D.F., P.C.; writing—original draft preparation, M.A.C.; writing—review and editing, M.A.C., I.M., M.D.S., M.A.D., D.F., P.C., and A.D. All authors have read and agreed to the published version of the manuscript.
Funding
This research was founded by the intramural research program at the University of Cagliari, Italy, and by the CNR project FOE-2021 DBA.AD005.225" (Institute of Biomolecular Chemistry, Unit of Sassari, Italy)
Institutional Review Board Statement
Not applicable here.
Data Availability Statement
The data presented in this study are available on request from the corresponding author.
Acknowledgments
We acknowledge the CeSAR (Centro Servizi d'Ateneo per la Ricerca) of the University of Cagliari, Italy, for the fluorescent image stacks performed with a fluorescence spinning disk confocal microscope (Crisel Instruments) and for the electron microscopy analysis (JEOL JEM 1400 Plus electron microscope).
Conflicts of Interest
The authors declare no conflict of interest.
References
- Dauer, W.; Przedborski, S. Parkinson’s Disease: Mechanisms and Models. Neuron 2003, 39, 889–909. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez-Oroz, M.C.; Jahanshahi, M.; Krack, P.; Litvan, I.; Macias, R.; Bezard, E.; Obeso, J.A. Initial Clinical Manifestations of Parkinson’s Disease: Features and Pathophysiological Mechanisms. Lancet Neurol 2009, 8, 1128–1139. [Google Scholar] [CrossRef] [PubMed]
- Lees, A.J.; Hardy, J.; Revesz, T. Parkinson’s Disease. Lancet 2009, 373, 2055–2066. [Google Scholar] [CrossRef] [PubMed]
- Chaudhuri, K.R.; Odin, P. The Challenge of Non-Motor Symptoms in Parkinson’s Disease. Prog Brain Res 2010, 184, 325–341. [Google Scholar] [CrossRef] [PubMed]
- Erkkinen, M.G.; Kim, M.-O.; Geschwind, M.D. Clinical Neurology and Epidemiology of the Major Neurodegenerative Diseases. Cold Spring Harb Perspect Biol 2018, 10, a033118. [Google Scholar] [CrossRef] [PubMed]
- Park, J.-S.; Davis, R.L.; Sue, C.M. Mitochondrial Dysfunction in Parkinson’s Disease: New Mechanistic Insights and Therapeutic Perspectives. Curr Neurol Neurosci Rep 2018, 18, 21. [Google Scholar] [CrossRef] [PubMed]
- Blesa, J.; Trigo-Damas, I.; Quiroga-Varela, A.; Jackson-Lewis, V.R. Oxidative Stress and Parkinson’s Disease. Front Neuroanat 2015, 9, 91. [Google Scholar] [CrossRef] [PubMed]
- Hauser, D.N.; Hastings, T.G. Mitochondrial Dysfunction and Oxidative Stress in Parkinson’s Disease and Monogenic Parkinsonism. Neurobiol Dis 2013, 51, 35–42. [Google Scholar] [CrossRef] [PubMed]
- Scalbert, A.; Manach, C.; Morand, C.; Rémésy, C.; Jiménez, L. Dietary Polyphenols and the Prevention of Diseases. Crit Rev Food Sci Nutr 2005, 45, 287–306. [Google Scholar] [CrossRef]
- Arias-Sánchez, R.A.; Torner, L.; Fenton Navarro, B. Polyphenols and Neurodegenerative Diseases: Potential Effects and Mechanisms of Neuroprotection. Molecules 2023, 28, 5415. [Google Scholar] [CrossRef]
- Aryal, S.; Skinner, T.; Bridges, B.; Weber, J.T. The Pathology of Parkinson’s Disease and Potential Benefit of Dietary Polyphenols. Molecules 2020, 25, 4382. [Google Scholar] [CrossRef] [PubMed]
- Gupta, S.C.; Patchva, S.; Koh, W.; Aggarwal, B.B. Discovery of Curcumin, a Component of Golden Spice, and Its Miraculous Biological Activities. Clin Exp Pharma Physio 2012, 39, 283–299. [Google Scholar] [CrossRef] [PubMed]
- Maheshwari, R.K.; Singh, A.K.; Gaddipati, J.; Srimal, R.C. Multiple Biological Activities of Curcumin: A Short Review. Life Sciences 2006, 78, 2081–2087. [Google Scholar] [CrossRef]
- Aggarwal, B.B.; Sung, B. Pharmacological Basis for the Role of Curcumin in Chronic Diseases: An Age-Old Spice with Modern Targets. Trends Pharmacol Sci 2009, 30, 85–94. [Google Scholar] [CrossRef] [PubMed]
- Di Meo, F.; Margarucci, S.; Galderisi, U.; Crispi, S.; Peluso, G. Curcumin, Gut Microbiota, and Neuroprotection. Nutrients 2019, 11, 2426. [Google Scholar] [CrossRef] [PubMed]
- Bhat, A.; Mahalakshmi, A.M.; Ray, B.; Tuladhar, S.; Hediyal, T.A.; Manthiannem, E.; Padamati, J.; Chandra, R.; Chidambaram, S.B.; Sakharkar, M.K. Benefits of Curcumin in Brain Disorders. Biofactors 2019, 45, 666–689. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.J.; Pan, M.H.; Cheng, A.L.; Lin, L.I.; Ho, Y.S.; Hsieh, C.Y.; Lin, J.K. Stability of Curcumin in Buffer Solutions and Characterization of Its Degradation Products. J Pharm Biomed Anal 1997, 15, 1867–1876. [Google Scholar] [CrossRef]
- Parihar, V.K.; Dhawan, J.; Kumar, S.; Manjula, S.N.; Subramanian, G.; Unnikrishnan, M.K.; Rao, C.M. Free Radical Scavenging and Radioprotective Activity of Dehydrozingerone against Whole Body Gamma Irradiation in Swiss Albino Mice. Chem Biol Interact 2007, 170, 49–58. [Google Scholar] [CrossRef]
- Kim, S.J.; Kim, H.M.; Lee, E.S.; Kim, N.; Lee, J.O.; Lee, H.J.; Park, N.Y.; Jo, J.Y.; Ham, B.Y.; Han, S.H.; et al. Dehydrozingerone Exerts Beneficial Metabolic Effects in High-Fat Diet-Induced Obese Mice via AMPK Activation in Skeletal Muscle. J Cell Mol Med 2015, 19, 620–629. [Google Scholar] [CrossRef]
- Dettori, M.A.; Pisano, M.; Rozzo, C.; Delogu, G.; Fabbri, D. Synthesis of Hydroxylated Biphenyl Derivatives Bearing an Alpha,Beta-Unsaturated Ketone as a Lead Structure for the Development of Drug Candidates against Malignant Melanoma. CHEMMEDCHEM 2021, 16, 1022–1033. [Google Scholar] [CrossRef]
- Hampannavar, G.A.; Karpoormath, R.; Palkar, M.B.; Shaikh, M.S. An Appraisal on Recent Medicinal Perspective of Curcumin Degradant: Dehydrozingerone (DZG). Bioorganic & Medicinal Chemistry 2016, 24, 501–520. [Google Scholar] [CrossRef]
- Moorkoth, S.; Prathyusha, N.S.; Manandhar, S.; Xue, Y.; Sankhe, R.; Pai, K.S.R.; Kumar, N. Antidepressant-like Effect of Dehydrozingerone from Zingiber Officinale by Elevating Monoamines in Brain: In Silico and in Vivo Studies. Pharmacol. Rep 2021, 73, 1273–1286. [Google Scholar] [CrossRef]
- Lee, E.S.; Kang, J.S.; Kim, H.M.; Kim, S.J.; Kim, N.; Lee, J.O.; Kim, H.S.; Lee, E.Y.; Chung, C.H. Dehydrozingerone Inhibits Renal Lipotoxicity in High-fat Diet–Induced Obese Mice. J Cell Mol Med 2021, 25, 8725–8733. [Google Scholar] [CrossRef] [PubMed]
- Pathak, N.; Cheruku, S.P.; Rao, V.; Vibhavari, R.J.A.; Sumalatha, S.; Gourishetti, K.; Rao, C.M.; Kumar, N. Dehydrozingerone Protects Temozolomide-Induced Cognitive Impairment in Normal and C6 Glioma Rats besides Enhancing Its Anticancer Potential. 3 Biotech 2020, 10, 438. [Google Scholar] [CrossRef] [PubMed]
- Tirunavalli, S.K.; Gourishetti, K.; Kotipalli, R.S.S.; Kuncha, M.; Kathirvel, M.; Kaur, R.; Jerald, M.K.; Sistla, R.; Andugulapati, S.B. Dehydrozingerone Ameliorates Lipopolysaccharide Induced Acute Respiratory Distress Syndrome by Inhibiting Cytokine Storm, Oxidative Stress via Modulating the MAPK/NF-κB Pathway. Phytomedicine 2021, 92, 153729. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Li, Y.; Wen, C.; Yan, Z.; Olatunji, O.J.; Yin, Z. Dehydrozingerone Alleviates Hyperalgesia, Oxidative Stress and Inflammatory Factors in Complete Freund’s Adjuvant-Induced Arthritic Rats. Drug Des Devel Ther 2022, 16, 3015–3022. [Google Scholar] [CrossRef] [PubMed]
- Choi, J.G.; Kim, S.Y.; Jeong, M.; Oh, M.S. Pharmacotherapeutic Potential of Ginger and Its Compounds in Age-Related Neurological Disorders. Pharmacol Ther 2018, 182, 56–69. [Google Scholar] [CrossRef] [PubMed]
- Hajduk, P.J.; Bures, M.; Praestgaard, J.; Fesik, S.W. Privileged Molecules for Protein Binding Identified from NMR-Based Screening. J Med Chem 2000, 43, 3443–3447. [Google Scholar] [CrossRef]
- Bringmann, G.; Gulder, T.; Gulder, T.A.M.; Breuning, M. Atroposelective Total Synthesis of Axially Chiral Biaryl Natural Products. Chem. Rev. 2011, 111, 563–639. [Google Scholar] [CrossRef]
- Paquin, A.; Reyes-Moreno, C.; Bérubé, G. Recent Advances in the Use of the Dimerization Strategy as a Means to Increase the Biological Potential of Natural or Synthetic Molecules. Molecules 2021, 26, 2340. [Google Scholar] [CrossRef]
- Pisano, M.; Pagnan, G.; Dettori, M.A.; Cossu, S.; Caffa, I.; Sassu, I.; Emionite, L.; Fabbri, D.; Cilli, M.; Pastorino, F.; et al. Enhanced Anti-Tumor Activity of a New Curcumin-Related Compound against Melanoma and Neuroblastoma Cells. Molecular Cancer 2010, 9, 137. [Google Scholar] [CrossRef] [PubMed]
- Kancheva, V.; Slavova-Kazakova, A.; Fabbri, D.; Dettori, M.A.; Delogu, G.; Janiak, M.; Amarowicz, R. Protective Effects of Equimolar Mixtures of Monomer and Dimer of Dehydrozingerone with α-Tocopherol and/or Ascorbyl Palmitate during Bulk Lipid Autoxidation. Food Chemistry 2014, 157, 263–274. [Google Scholar] [CrossRef] [PubMed]
- Profumo, E.; Buttari, B.; D’Arcangelo, D.; Tinaburri, L.; Dettori, M.A.; Fabbri, D.; Delogu, G.; Riganò, R. The Nutraceutical Dehydrozingerone and Its Dimer Counteract Inflammation- and Oxidative Stress-Induced Dysfunction of In Vitro Cultured Human Endothelial Cells: A Novel Perspective for the Prevention and Therapy of Atherosclerosis. Oxid Med Cell Longev 2016, 2016, 1246485. [Google Scholar] [CrossRef]
- Marchiani, A.; Mammi, S.; Siligardi, G.; Hussain, R.; Tessari, I.; Bubacco, L.; Delogu, G.; Fabbri, D.; Dettori, M.A.; Sanna, D.; et al. Small Molecules Interacting with α-Synuclein: Antiaggregating and Cytoprotective Properties. Amino Acids 2013, 45, 327–338. [Google Scholar] [CrossRef]
- Kumari, U.; Tan, E.K. LRRK2 in Parkinson’s Disease: Genetic and Clinical Studies from Patients. FEBS J 2009, 276, 6455–6463. [Google Scholar] [CrossRef] [PubMed]
- Hernandez, D.G.; Reed, X.; Singleton, A.B. Genetics in Parkinson Disease: Mendelian versus Non-Mendelian Inheritance. J Neurochem 2016, 139 Suppl 1, 59–74. [Google Scholar] [CrossRef]
- Trinh, J.; Zeldenrust, F.M.J.; Huang, J.; Kasten, M.; Schaake, S.; Petkovic, S.; Madoev, H.; Grünewald, A.; Almuammar, S.; König, I.R.; et al. Genotype-Phenotype Relations for the Parkinson’s Disease Genes SNCA, LRRK2, VPS35: MDSGene Systematic Review. Mov Disord 2018, 33, 1857–1870. [Google Scholar] [CrossRef] [PubMed]
- Biskup, S.; Moore, D.J.; Celsi, F.; Higashi, S.; West, A.B.; Andrabi, S.A.; Kurkinen, K.; Yu, S.-W.; Savitt, J.M.; Waldvogel, H.J.; et al. Localization of LRRK2 to Membranous and Vesicular Structures in Mammalian Brain. Ann Neurol 2006, 60, 557–569. [Google Scholar] [CrossRef]
- Taymans, J.-M.; Van den Haute, C.; Baekelandt, V. Distribution of PINK1 and LRRK2 in Rat and Mouse Brain. J Neurochem 2006, 98, 951–961. [Google Scholar] [CrossRef]
- Wallings, R.; Manzoni, C.; Bandopadhyay, R. Cellular Processes Associated with LRRK2 Function and Dysfunction. FEBS J 2015, 282, 2806–2826. [Google Scholar] [CrossRef]
- Price, A.; Manzoni, C.; Cookson, M.R.; Lewis, P.A. The LRRK2 Signalling System. Cell Tissue Res 2018, 373, 39–50. [Google Scholar] [CrossRef]
- Mills, R.D.; Mulhern, T.D.; Cheng, H.-C.; Culvenor, J.G. Analysis of LRRK2 Accessory Repeat Domains: Prediction of Repeat Length, Number and Sites of Parkinson’s Disease Mutations. Biochem Soc Trans 2012, 40, 1086–1089. [Google Scholar] [CrossRef] [PubMed]
- Berg, D.; Schweitzer, K.J.; Leitner, P.; Zimprich, A.; Lichtner, P.; Belcredi, P.; Brüssel, T.; Schulte, C.; Maass, S.; Nägele, T.; et al. Type and Frequency of Mutations in the LRRK2 Gene in Familial and Sporadic Parkinson’s Disease*. Brain 2005, 128, 3000–3011. [Google Scholar] [CrossRef] [PubMed]
- Tan, E.K.; Schapira, A.H. Uniting Chinese across Asia: The LRRK2 Gly2385Arg Risk Variant. Eur J Neurol 2008, 15, 203–204. [Google Scholar] [CrossRef] [PubMed]
- Rudenko, I.N.; Kaganovich, A.; Hauser, D.N.; Beylina, A.; Chia, R.; Ding, J.; Maric, D.; Jaffe, H.; Cookson, M.R. The G2385R Variant of Leucine-Rich Repeat Kinase 2 Associated with Parkinson’s Disease Is a Partial Loss-of-Function Mutation. Biochem J 2012, 446, 99–111. [Google Scholar] [CrossRef] [PubMed]
- Tan, E.K.; Peng, R.; Wu, Y.R.; Wu, R.M.; Wu-Chou, Y.H.; Tan, L.C.; An, X.K.; Chen, C.M.; Fook-Chong, S.; Lu, C.S. LRRK2 G2385R Modulates Age at Onset in Parkinson’s Disease: A Multi-Center Pooled Analysis. Am J Med Genet B Neuropsychiatr Genet 2009, 150B, 1022–1023. [Google Scholar] [CrossRef] [PubMed]
- Ross, O.A.; Soto-Ortolaza, A.I.; Heckman, M.G.; Aasly, J.O.; Abahuni, N.; Annesi, G.; Bacon, J.A.; Bardien, S.; Bozi, M.; Brice, A.; et al. Association of LRRK2 Exonic Variants with Susceptibility to Parkinson’s Disease: A Case-Control Study. Lancet Neurol 2011, 10, 898–908. [Google Scholar] [CrossRef]
- Carrion, M.D.P.; Marsicano, S.; Daniele, F.; Marte, A.; Pischedda, F.; Di Cairano, E.; Piovesana, E.; von Zweydorf, F.; Kremmer, E.; Gloeckner, C.J.; et al. The LRRK2 G2385R Variant Is a Partial Loss-of-Function Mutation That Affects Synaptic Vesicle Trafficking through Altered Protein Interactions. Sci Rep 2017, 7, 5377. [Google Scholar] [CrossRef]
- Bilen, J.; Bonini, N.M. Drosophila as a Model for Human Neurodegenerative Disease. Annu Rev Genet 2005, 39, 153–171. [Google Scholar] [CrossRef]
- Reiter, L.T.; Potocki, L.; Chien, S.; Gribskov, M.; Bier, E. A Systematic Analysis of Human Disease-Associated Gene Sequences in Drosophila Melanogaster. Genome Res 2001, 11, 1114–1125. [Google Scholar] [CrossRef]
- Aryal, B.; Lee, Y. Disease Model Organism for Parkinson Disease: Drosophila Melanogaster. BMB Rep 2019, 52, 250–258. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.; Imai, Y.; Gehrke, S.; Liu, S.; Lu, B. The Synaptic Function of LRRK2. Biochem Soc Trans 2012, 40, 1047–1051. [Google Scholar] [CrossRef] [PubMed]
- Hewitt, V.L.; Whitworth, A.J. Mechanisms of Parkinson’s Disease: Lessons from Drosophila. Curr Top Dev Biol 2017, 121, 173–200. [Google Scholar] [CrossRef]
- De Rose, F.; Marotta, R.; Poddighe, S.; Talani, G.; Catelani, T.; Setzu, M.D.; Solla, P.; Marrosu, F.; Sanna, E.; Kasture, S.; et al. Functional and Morphological Correlates in the Drosophila LRRK2 Loss-of-Function Model of Parkinson’s Disease: Drug Effects of Withania Somnifera (Dunal) Administration. PLoS One 2016, 11, e0146140. [Google Scholar] [CrossRef]
- Casu, M.A.; Mocci, I.; Isola, R.; Pisanu, A.; Boi, L.; Mulas, G.; Greig, N.H.; Setzu, M.D.; Carta, A.R. Neuroprotection by the Immunomodulatory Drug Pomalidomide in the Drosophila LRRK2WD40 Genetic Model of Parkinson’s Disease. Front Aging Neurosci 2020, 12, 31. [Google Scholar] [CrossRef]
- Diana, A.; Collu, M.; Casu, M.A.; Mocci, I.; Aguilar-Santelises, M.; Setzu, M.D. Improvements of Motor Performances in the Drosophila LRRK2 Loss-of-Function Model of Parkinson’s Disease: Effects of Dialyzed Leucocyte Extracts from Human Serum. Brain Sci 2020, 10, 45. [Google Scholar] [CrossRef]
- Mocci, I.; Casu, M.A.; Sogos, V.; Liscia, A.; Angius, R.; Cadeddu, F.; Fanti, M.; Muroni, P.; Talani, G.; Diana, A.; et al. Effects of Memantine on Mania-like Phenotypes Exhibited by Drosophila Shaker Mutants. CNS Neurosci Ther 2023, 29, 1750–1761. [Google Scholar] [CrossRef] [PubMed]
- Monastirioti, M. Biogenic Amine Systems in the Fruit Fly Drosophila Melanogaster. Microsc Res Tech 1999, 45, 106–121. [Google Scholar] [CrossRef]
- Lima, S.Q.; Miesenböck, G. Remote Control of Behavior through Genetically Targeted Photostimulation of Neurons. Cell 2005, 121, 141–152. [Google Scholar] [CrossRef]
- Zars, T. Behavioral Functions of the Insect Mushroom Bodies. Curr Opin Neurobiol 2000, 10, 790–795. [Google Scholar] [CrossRef]
- Quintero-Espinosa, D.A.; Sanchez-Hernandez, S.; Velez-Pardo, C.; Martin, F.; Jimenez-Del-Rio, M. LRRK2 Knockout Confers Resistance in HEK-293 Cells to Rotenone-Induced Oxidative Stress, Mitochondrial Damage, and Apoptosis. Int J Mol Sci 2023, 24, 10474. [Google Scholar] [CrossRef] [PubMed]
- Jaleel, M.; Nichols, R.J.; Deak, M.; Campbell, D.G.; Gillardon, F.; Knebel, A.; Alessi, D.R. LRRK2 Phosphorylates Moesin at Threonine-558: Characterization of How Parkinson’s Disease Mutants Affect Kinase Activity. Biochem J 2007, 405, 307–317. [Google Scholar] [CrossRef] [PubMed]
- Jorgensen, N.D.; Peng, Y.; Ho, C.C.-Y.; Rideout, H.J.; Petrey, D.; Liu, P.; Dauer, W.T. The WD40 Domain Is Required for LRRK2 Neurotoxicity. PLOS ONE 2009, 4, e8463. [Google Scholar] [CrossRef] [PubMed]
- Tan, E.K.; Zhao, Y.; Skipper, L.; Tan, M.G.; Di Fonzo, A.; Sun, L.; Fook-Chong, S.; Tang, S.; Chua, E.; Yuen, Y.; et al. The LRRK2 Gly2385Arg Variant Is Associated with Parkinson’s Disease: Genetic and Functional Evidence. Hum Genet 2007, 120, 857–863. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez, M.; Rodriguez-Sabate, C.; Morales, I.; Sanchez, A.; Sabate, M. Parkinson’s Disease as a Result of Aging. Aging Cell 2015, 14, 293–308. [Google Scholar] [CrossRef]
- Zhou, C.; Huang, Y.; Przedborski, S. Oxidative Stress in Parkinson’s Disease: A Mechanism of Pathogenic and Therapeutic Significance. Ann N Y Acad Sci 2008, 1147, 93–104. [Google Scholar] [CrossRef] [PubMed]
- Henchcliffe, C.; Beal, M.F. Mitochondrial Biology and Oxidative Stress in Parkinson Disease Pathogenesis. Nat Clin Pract Neurol 2008, 4, 600–609. [Google Scholar] [CrossRef] [PubMed]
- Cook C, Petrucelli L, 2009 A Critical Evaluation of the Ubiquitin-Proteasome System in Parkinson’s Disease. Biochim Biophys Acta 1792, 664–675.
- Pan, T.; Kondo, S.; Le, W.; Jankovic, J. The Role of Autophagy-Lysosome Pathway in Neurodegeneration Associated with Parkinson’s Disease. Brain 2008, 131, 1969–1978. [Google Scholar] [CrossRef]
- Slavova-Kazakova, A.K.; Koleva, L.; Kancheva, V.D.; Delogu, G. Comparative Study of Antioxidant Potential of Curcumin and Its Degradation Products–Vanillin, Ferulic Acid and Dehydrozingerone. Bulg Chem Commun, 2018, 50, 158–163. [Google Scholar]
- Ortega-Arellano, H.F.; Jimenez-Del-Rio, M.; Velez-Pardo, C. Dmp53, Basket and drICE Gene Knockdown and Polyphenol Gallic Acid Increase Life Span and Locomotor Activity in a Drosophila Parkinson’s Disease Model. Genet Mol Biol 2013, 36, 608–615. [Google Scholar] [CrossRef] [PubMed]
- Ortega-Arellano, H.F.; Jimenez-Del-Rio, M.; Velez-Pardo, C. Neuroprotective Effects of Methanolic Extract of Avocado Persea Americana (Var. Colinred) Peel on Paraquat-Induced Locomotor Impairment, Lipid Peroxidation and Shortage of Life Span in Transgenic Knockdown Parkin Drosophila Melanogaster. Neurochem Res 2019, 44, 1986–1998. [Google Scholar] [CrossRef] [PubMed]
- Siddique, Y.H.; Naz, F.; Jyoti, S. Effect of Curcumin on Lifespan, Activity Pattern, Oxidative Stress, and Apoptosis in the Brains of Transgenic Drosophila Model of Parkinson’s Disease. Biomed Res Int 2014, 2014, 606928. [Google Scholar] [CrossRef] [PubMed]
- Mandel, S.; Youdim, M.B.H. Catechin Polyphenols: Neurodegeneration and Neuroprotection in Neurodegenerative Diseases. Free Radic Biol Med 2004, 37, 304–317. [Google Scholar] [CrossRef] [PubMed]
- Tsang, A.H.K.; Chung, K.K.K. Oxidative and Nitrosative Stress in Parkinson’s Disease. Biochim Biophys Acta 2009, 1792, 643–650. [Google Scholar] [CrossRef] [PubMed]
- Kausar, S.; Wang, F.; Cui, H. The Role of Mitochondria in Reactive Oxygen Species Generation and Its Implications for Neurodegenerative Diseases. Cells 2018, 7, 274. [Google Scholar] [CrossRef] [PubMed]
- Yang, D.; Li, T.; Liu, Z.; Arbez, N.; Yan, J.; Moran, T.H.; Ross, C.A.; Smith, W.W. LRRK2 Kinase Activity Mediates Toxic Interactions between Genetic Mutation and Oxidative Stress in a Drosophila Model: Suppression by Curcumin. Neurobiol Dis 2012, 47, 385–392. [Google Scholar] [CrossRef]
- Reale, M.; Pesce, M.; Priyadarshini, M.; Kamal, M.A.; Patruno, A. Mitochondria as an Easy Target to Oxidative Stress Events in Parkinson’s Disease. CNS Neurol Disord Drug Targets 2012, 11, 430–438. [Google Scholar] [CrossRef]
- Wang, X.; Yan, M.H.; Fujioka, H.; Liu, J.; Wilson-Delfosse, A.; Chen, S.G.; Perry, G.; Casadesus, G.; Zhu, X. LRRK2 Regulates Mitochondrial Dynamics and Function through Direct Interaction with DLP1. Hum Mol Genet 2012, 21, 1931–1944. [Google Scholar] [CrossRef]
- Hsieh, C.-H.; Shaltouki, A.; Gonzalez, A.E.; Bettencourt da Cruz, A.; Burbulla, L.F.; St Lawrence, E.; Schüle, B.; Krainc, D.; Palmer, T.D.; Wang, X. Functional Impairment in Miro Degradation and Mitophagy Is a Shared Feature in Familial and Sporadic Parkinson’s Disease. Cell Stem Cell 2016, 19, 709–724. [Google Scholar] [CrossRef]
- Mancini, A.; Mazzocchetti, P.; Sciaccaluga, M.; Megaro, A.; Bellingacci, L.; Beccano-Kelly, D.A.; Di Filippo, M.; Tozzi, A.; Calabresi, P. From Synaptic Dysfunction to Neuroprotective Strategies in Genetic Parkinson’s Disease: Lessons From LRRK2. Front Cell Neurosci 2020, 14, 158. [Google Scholar] [CrossRef] [PubMed]
- Angeles, D.C.; Ho, P.; Chua, L.L.; Wang, C.; Yap, Y.W.; Ng, C.; Zhou, Z. dong; Lim, K.-L.; Wszolek, Z.K.; Wang, H.Y.; et al. Thiol Peroxidases Ameliorate LRRK2 Mutant-Induced Mitochondrial and Dopaminergic Neuronal Degeneration in Drosophila. Hum Mol Genet 2014, 23, 3157–3165. [Google Scholar] [CrossRef] [PubMed]
- Jin, H.; Kanthasamy, A.; Ghosh, A.; Anantharam, V.; Kalyanaraman, B.; Kanthasamy, A.G. Mitochondria-Targeted Antioxidants for Treatment of Parkinson’s Disease: Preclinical and Clinical Outcomes. Biochim Biophys Acta 2014, 1842, 1282–1294. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Jia, H.; Liu, J.; Ao, N.; Yan, B.; Shen, W.; Wang, X.; Li, X.; Luo, C.; Liu, J. Combined R-α–Lipoic Acid and Acetyl-L-Carnitine Exerts Efficient Preventative Effects in a Cellular Model of Parkinson’s Disease. J Cell Mol Med 2010, 14, 215–225. [Google Scholar] [CrossRef]
- Abdin, A.A.; Sarhan, N.I. Intervention of Mitochondrial Dysfunction-Oxidative Stress-Dependent Apoptosis as a Possible Neuroprotective Mechanism of α-Lipoic Acid against Rotenone-Induced Parkinsonism and L-Dopa Toxicity. Neurosci Res 2011, 71, 387–395. [Google Scholar] [CrossRef] [PubMed]
- Tamilselvam, K.; Braidy, N.; Manivasagam, T.; Essa, M.M.; Prasad, N.R.; Karthikeyan, S.; Thenmozhi, A.J.; Selvaraju, S.; Guillemin, G.J. Neuroprotective Effects of Hesperidin, a Plant Flavanone, on Rotenone-Induced Oxidative Stress and Apoptosis in a Cellular Model for Parkinson’s Disease. Oxid Med Cell Longev 2013, 2013, 102741. [Google Scholar] [CrossRef]
- Wang, Y.-H.; Yu, H.-T.; Pu, X.-P.; Du, G.-H. Baicalein Prevents 6-Hydroxydopamine-Induced Mitochondrial Dysfunction in SH-SY5Y Cells via Inhibition of Mitochondrial Oxidation and up-Regulation of DJ-1 Protein Expression. Molecules 2013, 18, 14726–14738. [Google Scholar] [CrossRef]
- Kaur, H.; Chauhan, S.; Sandhir, R. Protective Effect of Lycopene on Oxidative Stress and Cognitive Decline in Rotenone Induced Model of Parkinson’s Disease. Neurochem Res 2011, 36, 1435–1443. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Yu, Y.; Li, X.; Ross, C.A.; Smith, W.W. Curcumin Protects against A53T Alpha-Synuclein-Induced Toxicity in a PC12 Inducible Cell Model for Parkinsonism. Pharmacol Res 2011, 63, 439–444. [Google Scholar] [CrossRef]
- Parkinson Study Group QE3 Investigators; Beal, M.F.; Oakes, D.; Shoulson, I.; Henchcliffe, C.; Galpern, W.R.; Haas, R.; Juncos, J.L.; Nutt, J.G.; Voss, T.S.; et al. A Randomized Clinical Trial of High-Dosage Coenzyme Q10 in Early Parkinson Disease: No Evidence of Benefit. JAMA Neurol 2014, 71, 543–552. [CrossRef]
- Writing Group for the NINDS Exploratory Trials in Parkinson Disease (NET-PD) Investigators; Kieburtz, K.; Tilley, B.C.; Elm, J.J.; Babcock, D.; Hauser, R.; Ross, G.W.; Augustine, A.H.; Augustine, E.U.; Aminoff, M.J.; et al. Effect of Creatine Monohydrate on Clinical Progression in Patients with Parkinson Disease: A Randomized Clinical Trial. JAMA 2015, 313, 584–593. [CrossRef]
- Fouquet, W.; Owald, D.; Wichmann, C.; Mertel, S.; Depner, H.; Dyba, M.; Hallermann, S.; Kittel, R.J.; Eimer, S.; Sigrist, S.J. Maturation of Active Zone Assembly by Drosophila Bruchpilot. J Cell Biol 2009, 186, 129–145. [Google Scholar] [CrossRef]
- Piccoli, G.; Onofri, F.; Cirnaru, M.D.; Kaiser, C.J.O.; Jagtap, P.; Kastenmüller, A.; Pischedda, F.; Marte, A.; von Zweydorf, F.; Vogt, A.; et al. Leucine-Rich Repeat Kinase 2 Binds to Neuronal Vesicles through Protein Interactions Mediated by Its C-Terminal WD40 Domain. Mol Cell Biol 2014, 34, 2147–2161. [Google Scholar] [CrossRef]
- Zaichick, S.V.; McGrath, K.M.; Caraveo, G. The Role of Ca2+ Signaling in Parkinson’s Disease. Disease Models & Mechanisms 2017, 10, 519–535. [Google Scholar] [CrossRef]
Figure 1.
Chemical structures of: (a) Curcumin; (b) Ferulic acid, Vanillin and Dehydrozingerone (DHZ).
Figure 1.
Chemical structures of: (a) Curcumin; (b) Ferulic acid, Vanillin and Dehydrozingerone (DHZ).
Scheme 1.
Synthesis of Dehydrozingerone (DHZ) and its dimer (DHZ-DIM).
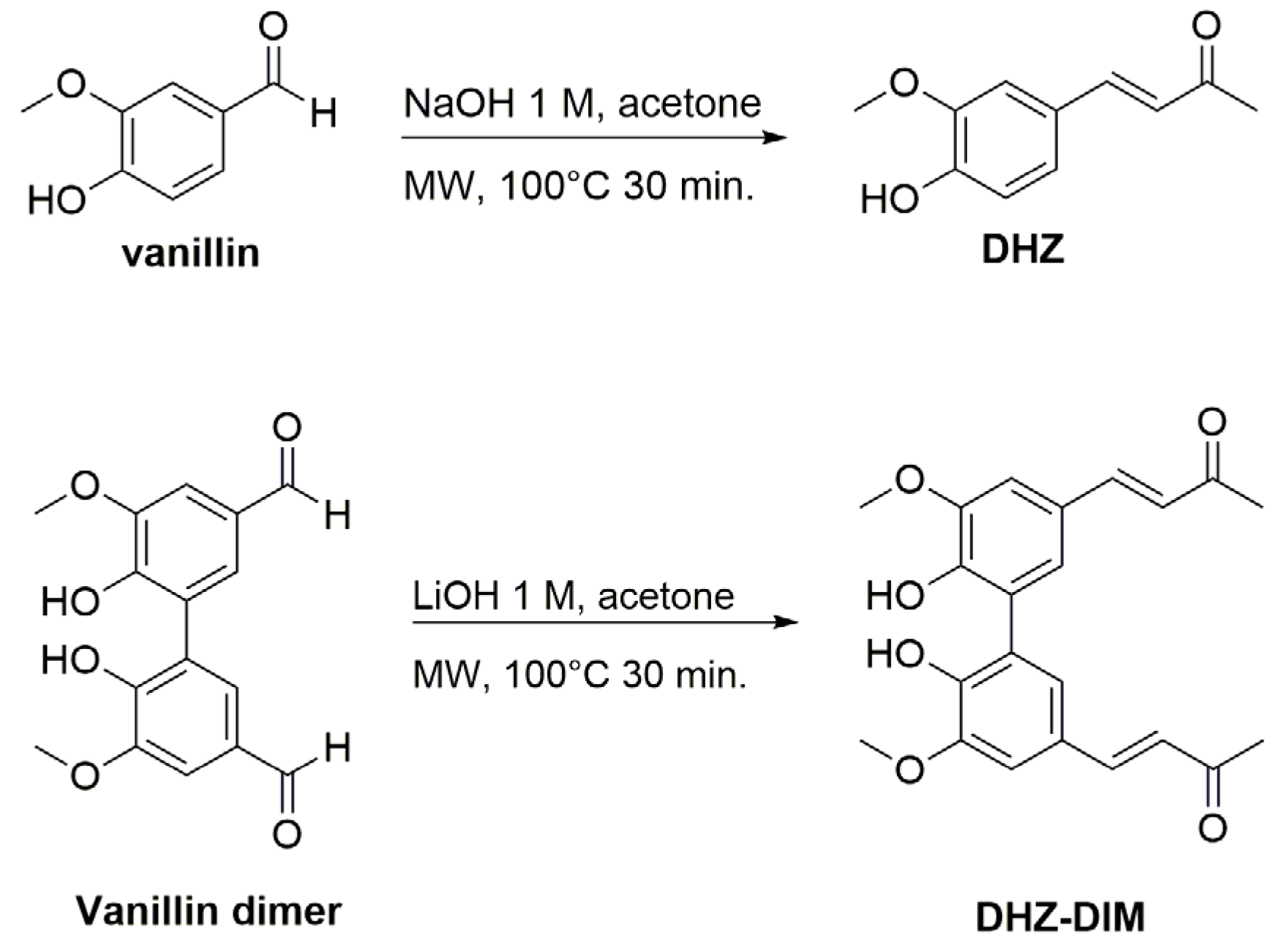
Figure 2.
Effect of DHZ and DHZ-DIM at 0.5 mM and 1 mM on the climbing behavior parameters in both LRRK and w1118 WT flies and their respective control groups at 14 (a-b) and 21 days (c-d) of treatment. § and ****p<0,0001 vs LRRK vehicle; **p<0,05 vs LRRK vehicle.
Figure 2.
Effect of DHZ and DHZ-DIM at 0.5 mM and 1 mM on the climbing behavior parameters in both LRRK and w1118 WT flies and their respective control groups at 14 (a-b) and 21 days (c-d) of treatment. § and ****p<0,0001 vs LRRK vehicle; **p<0,05 vs LRRK vehicle.
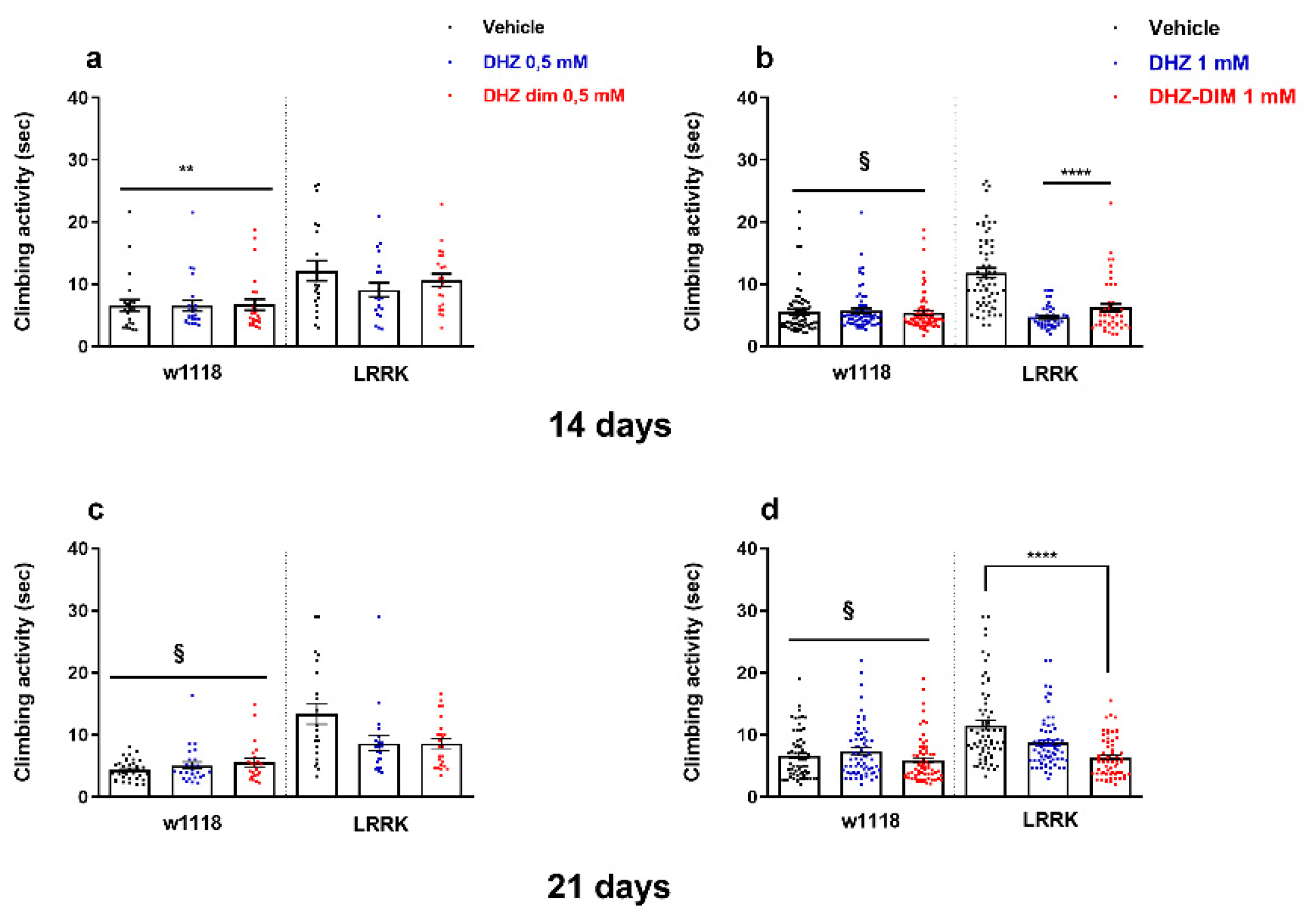
Figure 3.
Percentage of w1118 and LRRK individuals reaching the target within 10 s.
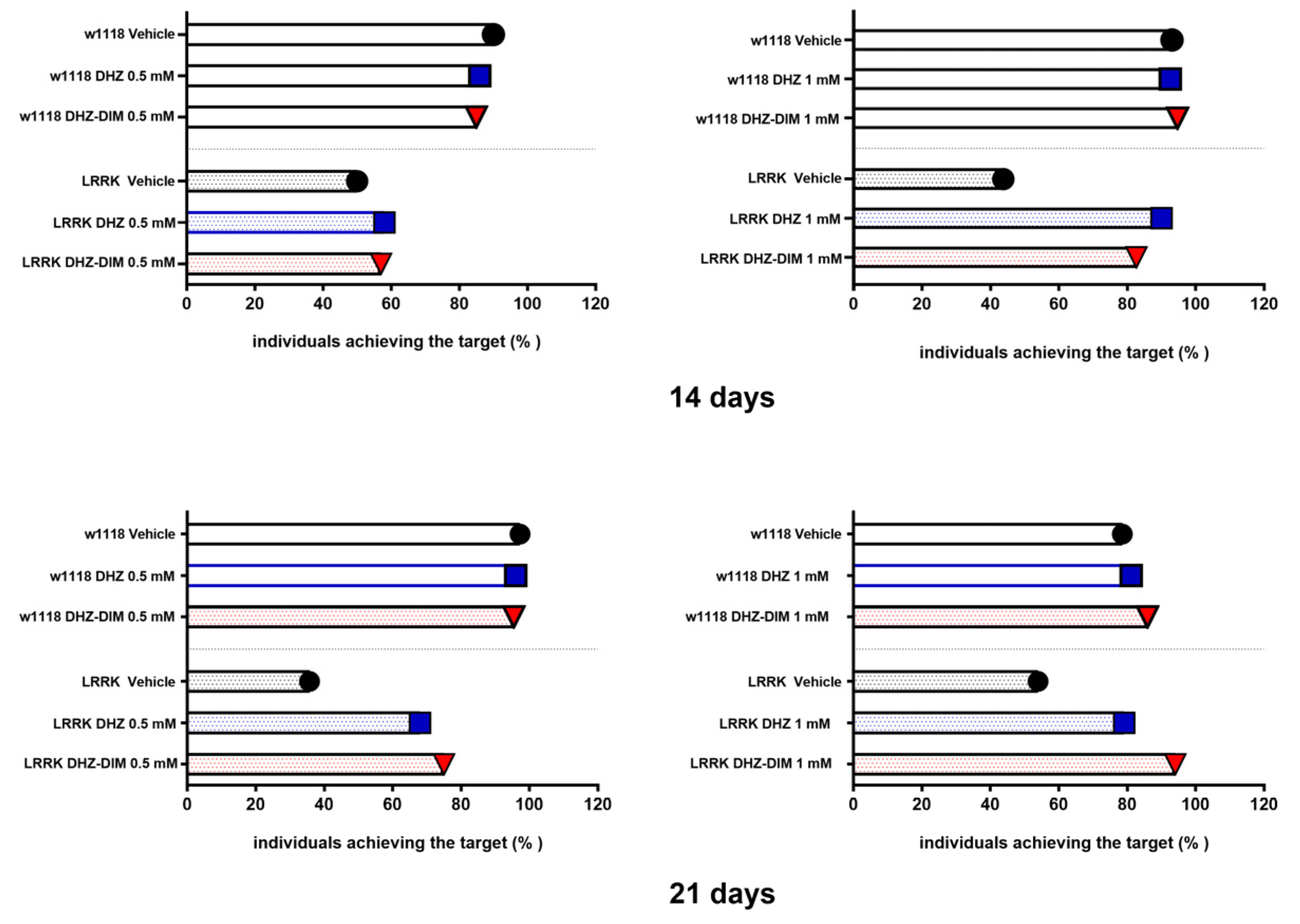
Figure 4.
Life span. a: reduced lifespan in LRRK compared to w1118 flies both treated with Vehicle; b: survival rate observed in DHZ and DHZ-DIM-treated LRRK at 1mM compared to WT Vehicle. Cumulative survival curves data are expressed as mean ± SEM. ***p<0.001 indicate significant difference between LRRK vehicle and WT vehicle. ****p<0.01 LRRK vehicle vs LRRK treated with DHZ-DIM. (test Gehan-Breslow-Wilcoxon).
Figure 4.
Life span. a: reduced lifespan in LRRK compared to w1118 flies both treated with Vehicle; b: survival rate observed in DHZ and DHZ-DIM-treated LRRK at 1mM compared to WT Vehicle. Cumulative survival curves data are expressed as mean ± SEM. ***p<0.001 indicate significant difference between LRRK vehicle and WT vehicle. ****p<0.01 LRRK vehicle vs LRRK treated with DHZ-DIM. (test Gehan-Breslow-Wilcoxon).
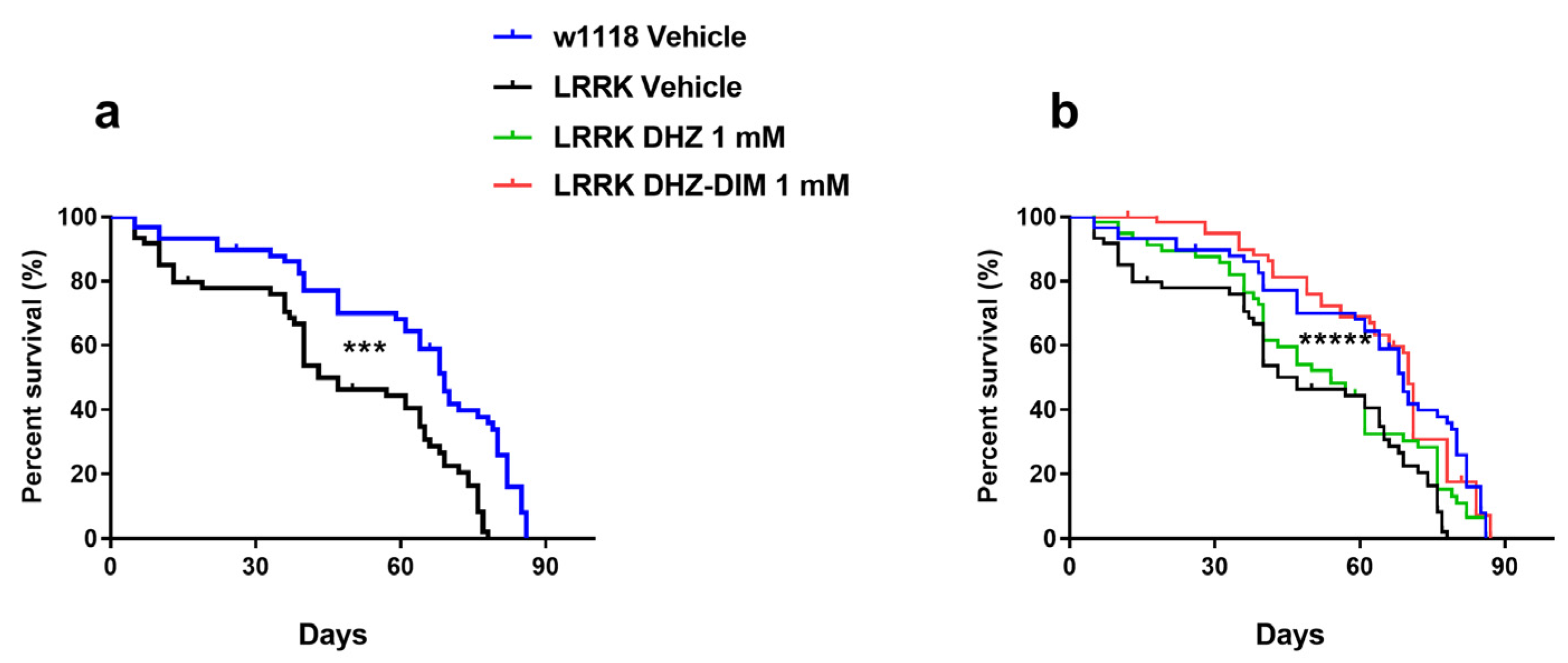
Figure 5.
Representative image stacks showing the entire brain (a: 10X scale bar: 100µm) and the dopaminergic brain neurons in all posterior clusters. (b: 63X, scale bar: 10µm).
Figure 5.
Representative image stacks showing the entire brain (a: 10X scale bar: 100µm) and the dopaminergic brain neurons in all posterior clusters. (b: 63X, scale bar: 10µm).
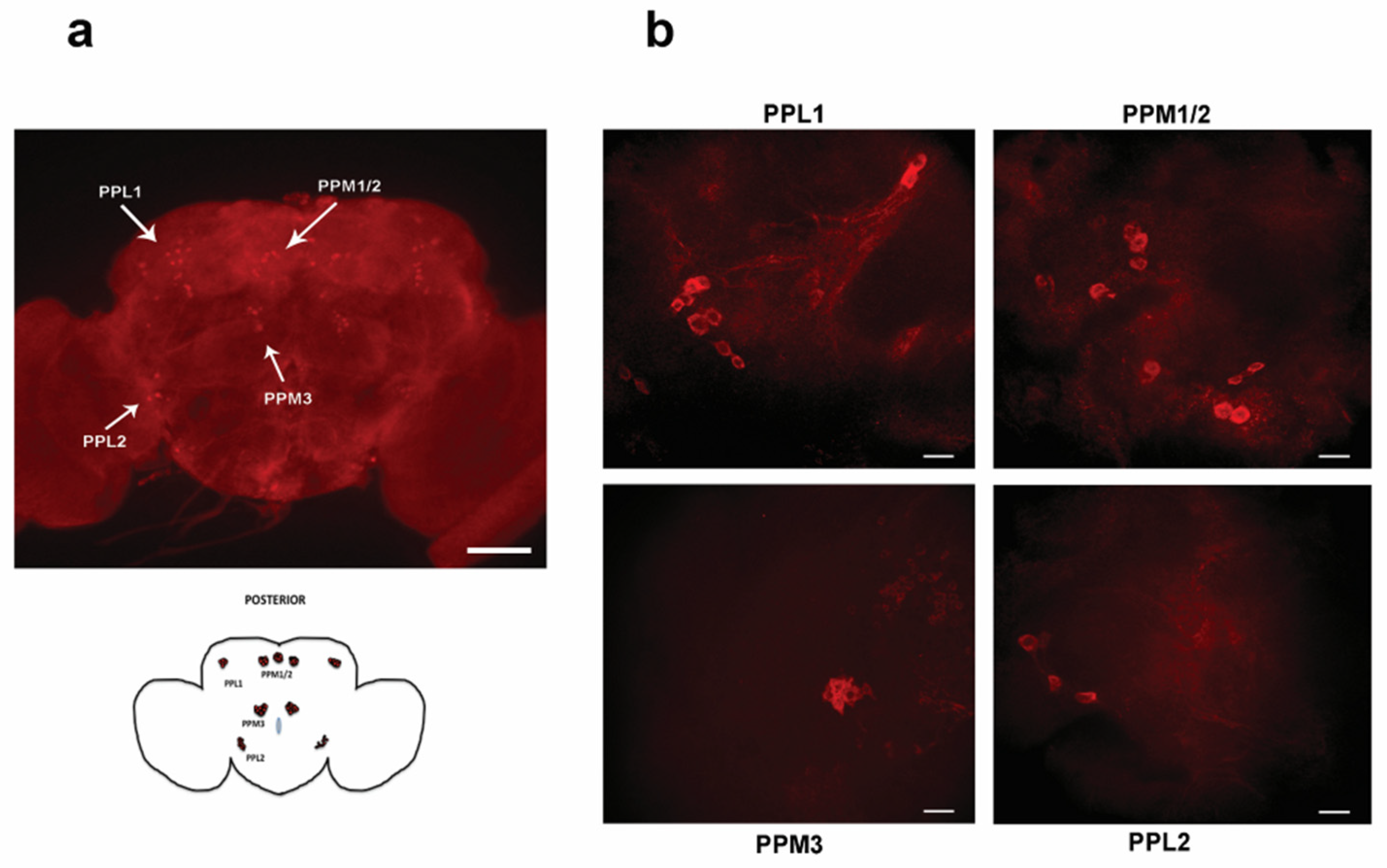
Figure 6.
DHZ and DHZ-DIM prevent the loss of dopaminergic neurons. Both compounds at 1 mM significantly prevented the loss of dopaminergic neurons in all posterior clusters at 14 days of treatment whereas, at 21 days, a significant prevention of dopaminergic neuron loss was observed only with the DHZ-DIM treatment. **p<0.01, ***p<0.001 and ****p<0.0001 indicate significant difference between LRRK2 vehicle and LRRK2 treated with DHZ, DHZ-DIM or vehicle and treated WT groups. &p<0.0001 LRRK treated with DHZ-DIM vs LRRK treated with DHZ.
Figure 6.
DHZ and DHZ-DIM prevent the loss of dopaminergic neurons. Both compounds at 1 mM significantly prevented the loss of dopaminergic neurons in all posterior clusters at 14 days of treatment whereas, at 21 days, a significant prevention of dopaminergic neuron loss was observed only with the DHZ-DIM treatment. **p<0.01, ***p<0.001 and ****p<0.0001 indicate significant difference between LRRK2 vehicle and LRRK2 treated with DHZ, DHZ-DIM or vehicle and treated WT groups. &p<0.0001 LRRK treated with DHZ-DIM vs LRRK treated with DHZ.
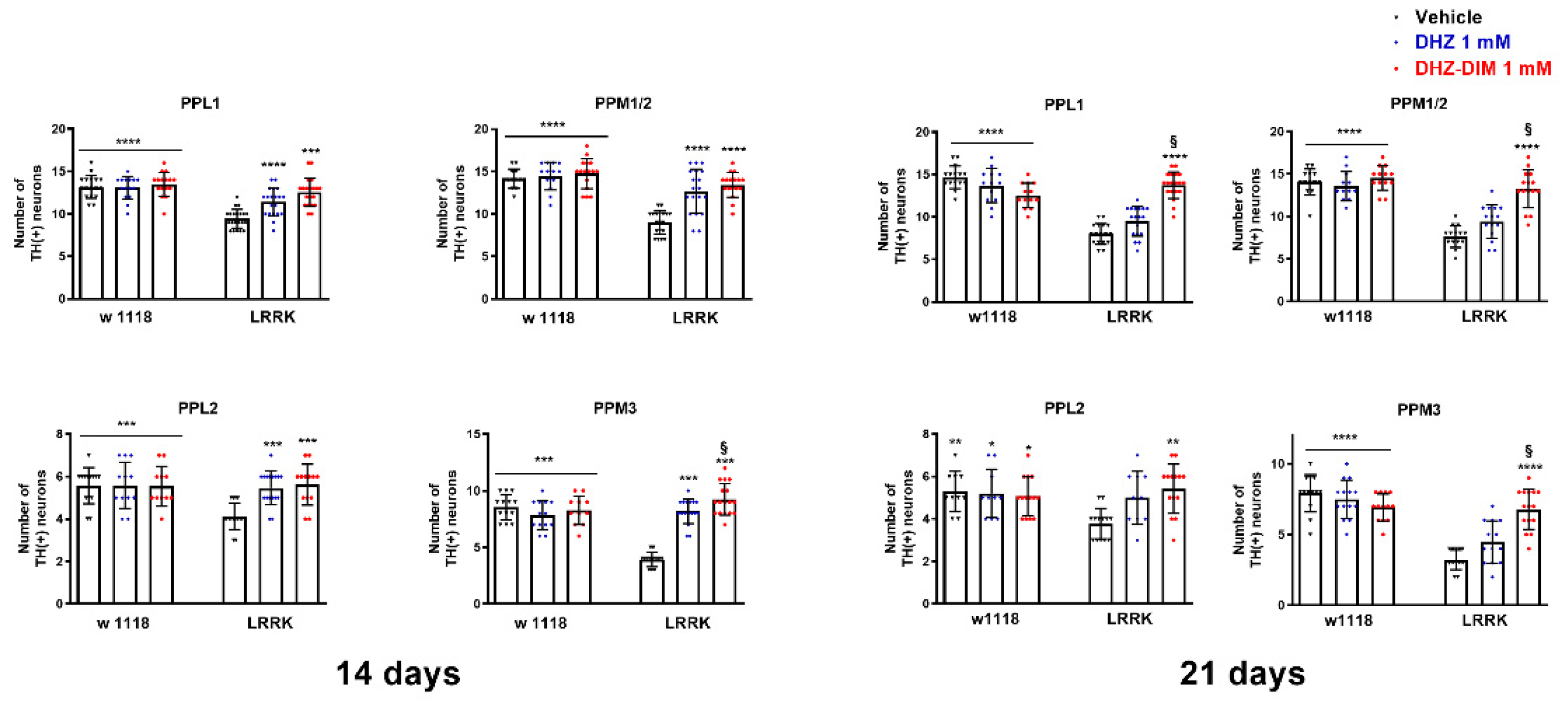
Figure 7.
Representative images of mitochondria (8,000X) acquired from the protocerebrum of LRRK and w1118 treated with vehicle (a-d), LRRK treated with DHZ (b) and DHZ-DIM (c) for 21 days. Red Arrows: Mitochondria with swollen cristae; Black Arrows: Mitochondria with normal cristae; White Arrows: T-Bars.
Figure 7.
Representative images of mitochondria (8,000X) acquired from the protocerebrum of LRRK and w1118 treated with vehicle (a-d), LRRK treated with DHZ (b) and DHZ-DIM (c) for 21 days. Red Arrows: Mitochondria with swollen cristae; Black Arrows: Mitochondria with normal cristae; White Arrows: T-Bars.
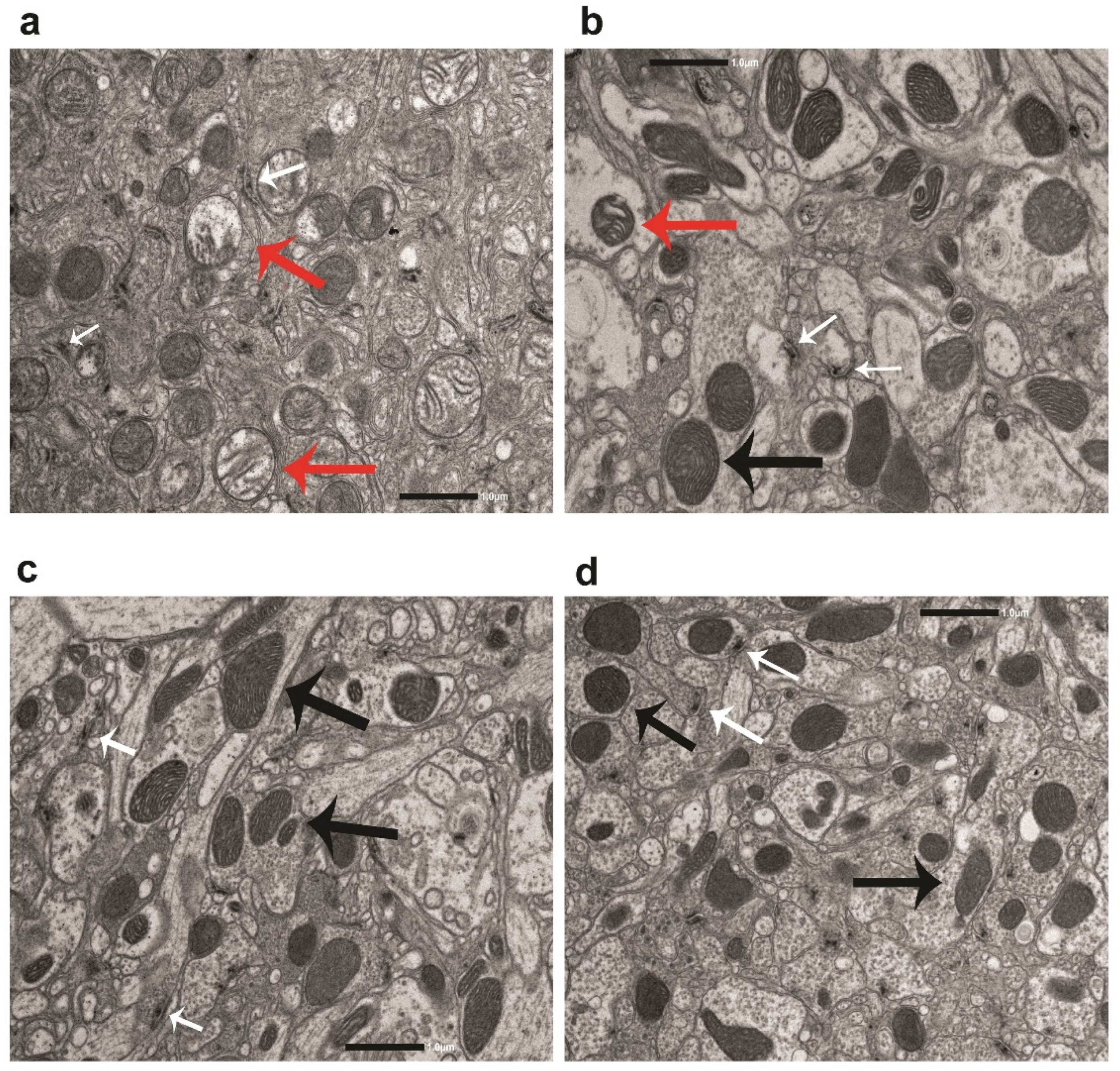
Figure 8.
DHZ and DHZ-DIM effect on the number of total mitochondria, percentage of cristae and T-Bars in the protocerebrum of LRRK after 21 days of treatment. *p<0.05 and ****p<0.0001 indicate significant difference between LRRK vehicle and LRRK treated with DHZ, DHZ-DIM or vehicle and treated WT groups. &p<0.0001 LRRK treated with DHZ-DIM vs LRRK treated with DHZ.
Figure 8.
DHZ and DHZ-DIM effect on the number of total mitochondria, percentage of cristae and T-Bars in the protocerebrum of LRRK after 21 days of treatment. *p<0.05 and ****p<0.0001 indicate significant difference between LRRK vehicle and LRRK treated with DHZ, DHZ-DIM or vehicle and treated WT groups. &p<0.0001 LRRK treated with DHZ-DIM vs LRRK treated with DHZ.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



