
Submitted:
26 December 2023
Posted:
27 December 2023
You are already at the latest version
Abstract
Recreational water activities are widely recognized to have a positive impact on our physical and mental well-being. However, recreational water sources and their management are also a risk factor for human health, due to different agents, including the overgrowth of cyanobacteria and algae. These organisms have the potential to metabolize organic matter and produce thermophilic and thermotolerant toxins. Different species of algae participate in biofilm formation and impact on maintenance of equipment for water treatment. We reviewed the state of the art by providing basic definitions, taxonomy and the epidemiological and clinical issues related to the recreational uses of waters. Methods for treatments and monitoring are summarized, considering both traditional and innovative strategies. Public health and surveillance perspectives impose to focus on the detection of toxins, the formation of biofilms, the understanding of the benthonic and planktonic components as part of a larger microbial biodiversity.
Keywords:
Algae
; Cyanobacteria
; Recreational water
; Swimming pool
; SPA
; Water treatments
; Monitoring
; Climate change
1. Introduction
Water is a fundamental element for human health and well-being [1]. Recreational water activities are widely known to positively affect our physical and mental health and fitness. These activities, such as swimming in oceans, lakes, rivers, pools, or spas, are popular leisure for several communities and major attractions for tourists and sporting events. However, freshwaters and coastal water bodies are experiencing rising pollution due to the escalating pressure of human activities, as well as the impact of climate changes. Contributing factors include untreated sewage overflows, animal excretion run-off from nearby farms, and algal blooms from high nutrient loads. Water sources safety represents one of the main factors influencing human health. Ensuring the proper management of waters intended for recreational uses implies a substantial economic impact in the sectors of tourism, wellness, sports events and a positive cascade across the entire water protection chain, by diffusing awareness of the resource and respect of policies. Climate change and anthropic activities also increasingly contribute to the impact on fresh and marine waters due to polluting discharges of untreated sewage, farming industries animal slurry, and algal blooms catalyzed by nutrients eutrophication phenomena [2]. A crucial and yet neglected aspect in recreational waters is linked to the cyanobacterial and algal overgrowth, whose pathogenic species are potentially capable of metabolizing organic matter, producing thermophilic and thermotolerant toxins, often biologically active neurotoxins, posing severe health risks for humans, animals and environments [3]. Among the several aquatic habitats, recreational waters and thermal mineral basins, indeed, represent a favorable reservoir for Algae growth. The uncontrolled proliferation of algal species on the surface layers of spa pools leads to the generation of unpleasant coloring, further affecting the appearance and smell of the water in pool, but impacting also on pipelines and treatment plants [4]. Besides being ubiquitous in water environments, low algae concentrations do not generally represent health risks for human health. The main concerns in pools are related to the visual impact, unpleasant odor dissuading bathers to access thermal or rehabilitative treatments, or activities in swimming pools or water parks [5,6]. Besides, the macroscopic presence of algae is often only the visible part of a more complex microflora component including different bacteria and protozoa species, that may represent a risk for health by contact, inhalation or drinking [1]. Therefore, the field is part of a huge world of studies on microalgae, that started in XIX century and it is still very complex and fragmented into different disciplines, methods and points of view, uncovering over 36000 publications, of which almost 160 related to recreational waters, mainly published in the last decade, as available at PubMed databases. Here, this general and heterogeneous issue is approached from a wider perspective of public health, providing the essential principles and bases to address the complex and not-yet solved question of algae in waters for recreational uses.
2. Definition and Classification of Algae in Recreational Waters
Algae are aquatic organisms devoid of true roots, stems, or leaves, and without distinct multicellular structures. Algae are ancient photosynthetic organisms that shaped the biosphere. Historically, algae were divided into two groups based on their size: macroalgae and microalgae. The brown algal seaweeds are the macroscopic ones, while green, red and golden-brown are considered microalgae [7]. Another classification is based on the presence of the nucleus. The cyanobacteria, known as the blue-green algae have a prokaryotic organization while all the other algal groups are eukaryotic [7]. The taxonomic diversity is also represented by metabolic factors the non-plant algal groups employ heterotrophy and phagotrophy, while the plant-related groups utilize photosynthesis.
2.1. Evolution of Eukaryotic Photosynthetic Algae
It is now widely accepted that the first eukaryote cell was produced by the engulfment of an alpha-proteobacterium by an archaeal cell, belonging to the Asgard lineage [8]. Around 4 billion years ago, indeed, there lived a microbe called LUCA — the Last Universal Common Ancestor (Figure 1) and the first eukaryotic common ancestor (FECA) was generated to take place about 2.5 billion years ago by the engulfment process [9]. FECA is the common ancestor of all eukaryotic cells that have ever existed, while LECA (the last eukaryotic common ancestor, dated to around 1.2 billion years ago) is the common ancestor of the crown group eukaryotes [9]. For the crown group eukaryotes, a red algae fossil (Bioangiomorpha pubescens) is the first attribution dating back to 1 billion years ago [10]. The first green alga in the fossil record is Proterocladus antiquus dated to around 950 million years ago, therefore the red and green algae lineages had separated by 950 million years ago leading to a significant increase in algal diversity around as demonstrated by biomolecular data [8,11]. Oxygen photosynthesis originated in the prokaryotic cyanobacteria, then it took place in eukaryotes at least 1 billion years ago from the uptake of the beta-cyanobacterium Gloeomargarita lithophora by a phagotrophic eukaryotic cell. Subsequently, the cyanobacterium evolved into the primary plastid as photosynthetic organelle [12]. This endosymbiotic event marks the beginning of the Archaeplastida, a monophyletic supergroup that includes three lineages: green algae plus land plants, red algae and glaucophytes [13,14]. One other independent primary photosynthetic symbiosis is known: the cyanobacterium belonging to the Synechoccus/Prochlorococcus group of alpha-cyanobacteria was engulfed by the cercozoan amoebae of the genus Paulinella and evolved into the chromatophore organelle [15]. Since 120 million years ago, secondary and tertiary endosymbiosis took place giving rise to complex plastid organization [7]. Green algal plastid endosymbiosis led to the establishment of euglenids and chlorarachniophytes. The event of the red algal plastid formed cryptophytes, haptophye, dinoflagellates and stramenopiles (diatoms, brown algae).
2.2. Algal Taxonomy and Phylogeny
The phylogeny and taxonomy of algae are constantly under review due to the increase of groupings, particularly within the golden-brown group where secondary endosymbiosis often occurs [7]. Algae classification follows two main approaches. The use taxonomic groupings not sticking to monophyly but providing an overarching system is recently proposed [11]. In contrast, Adl et al. [14] use a hierarchical system based on monophyly [14]. Chromista are included in the five-eukaryote kingdom (Protozoa, Chromista, Fungi, Plantae or Archaeplastida, Animalia). This kingdom comprises the golden-brown groups of algae (chlorophyll c containing plastids of red algae origin) and most marine algae with heterotrophic protists, Dinoflagellates. The original definition of the kingdom Chromista derived from Thomas Cavalieri-Smth: "all chomophyte algae where chloroplasts are separate from the cytosol by four topologically distinct membranes as well as all heterotrophic protists that descended secondarily from them by losing plastids”. This definition was updated to note that a chromista plastics are inside the endoplasmatic reticulum (ER), whereas plastid in the Plantae (Archaeplastida) are always found in cytosol [15]. Adl et al. do not use the traditional taxonomical higher categories (phylum, family etc.. ) but recognize two overarching domains in eukaryotes, Amorpha and Diaphretickes. For Adl et al. Archaeplastida are a monophyletic clade under the Diaphoretickes and they include Stramenopiles, Alveolata, Rhizaria. Dinoflagellates are in the Alveolata group, the seaweeds are most closely associated with the Stramenopiles, while seaweeds, diatoms and dinoflagellates are not related to the other traditional algal groups of Chloroplastida (red and green algae) [14].
2.3. Classification of Algae
Harvey is considered the algologist who proposed the first descriptive algal classification. Since W. H. Harvey, several classifications have been proposed based on a variety of characters including morphological, physiological, biochemical traits and more recently the molecular characters have also been considered. F. E. Fritsch (1935), also known as Father of Phycology, proposed the most acceptable and comprehensive algal classification. In his book “The Structure and Reproduction of the Algae” the algal classification is based on different characteristics as pigmentation, chemical nature of reserve food material, flagellar arrangement (kind, number and point of insertion), presence or absence of organized nucleus in cell and playback mode. He classified algae into 8 phyla and 11 classes (Figure 1 and Table S1) [16,17,18,19,20]. A recent study has demonstrated that based on genetic differences and not just morphological ones it is possible to classify algae into 12 phyla [19].
Algae can range in size from a single cell to millions of cells, spanning seven orders of magnitude. The organisms comprise unicellular organisms (microalgae), such as Chlorella, diatoms, and Prototheca, or multicellular organisms (macroalgae), for example, green and brown seaweeds [21]. The macroalgae species Macrocystis pyrifera, also known as giant kelp, reaches 60 m in length, while microalgae are a smaller heterogeneous group, with organisms of sizes that range from 1 µm to 1 mm, such as Chlorella, which lives in freshwater or the soil and has spherical cells with a diameter ranging from 2 µm to 10 µm [17]. Microalgae (microphytes) are single-celled organisms that can convert solar energy into chemical energy through photosynthesis [16]. Macroalgae (seaweed) have a multicellular organisation whit and developed anatomical arrangements that resemble stems, roots, and leaves of higher plants for functions such as anchorage, transport, photosynthesis, and reproduction. This specialization indicates a level of complexity and evolutionary advancement [16]. Algae depend on carbon, nitrogen, phosphorus, and micronutrients for metabolic processes [21]. Growth rate and cellular composition depend on several environmental factors such as light, temperature, pH, and salinity. [22]. Sustainable high productivity requires synergistic interactions between multiple environmental variables and nutritional factors. Photosynthetic algae have a broad distribution on Earth, and they grow naturally in their environments, resulting in biomass production. Algae are ubiquitous organisms and they can be categorized ecologically on their habitats. Planktonic algae grow suspended in the water, whereas neustonic algae grow on the water surface. Cryophilic algae occur in snow and ice, thermophilic algae live in hot springs; edaphic algae live on or in soil; epizoic algae grow on animal, such as turtles and sloths; epiphytic algae grow on fungi, land plants, or other algae; corticolous algae grow on the bark of trees; epilithic algae live on rocks; endolithic algae live in porous rocks or coral; and chasmolithic algae grow in rock fissures. Some algae live inside other organisms, and in a general sense these are called endosymbionts. Specifically, endozoic endosymbionts live in protozoa or animals such as shelled gastropods, whereas endophytic endosymbionts live in fungi, plants, or other algae.
2.2. Algae in Recreational Waters
In recreational waters, algal blooms can be harmful because some algal species contain secondary metabolites that are toxic to humans and animals [23,24]. Harmful algal blooms (HABs) result from the proliferation of diverse algal species, mainly including cyanobacteria (also known as blue-green algae), diatoms, dinoflagellates, green algae. These harmful blooms can be caused by many types of phytoplankton. However, three main types of phytoplankton produce most of blooms that make people and animals sick: i) cyanobacteria (sometimes called blue-green algae), ii) dinoflagellates (sometimes called microalgae or red tide) and iii) diatoms (sometimes called microalgae or red tide).
The two main toxin-producing algal groups are cyanobacteria (aka blue-green algae) and dinoflagellates: from an epidemiological point of view, algal blooms produced by cyanobacteria are usually isolated from freshwater, while dinoflagellates blooms are mostly detected in seawater [24]. [24]. With regards to spa pools (i.e., thermal water fed pools), the algal variety and the overgrowth generally depend on the composition of minerals and the temperature of the water, whose optimal value is about 30 °C [25,26]; while higher temperatures (> 55 °C) strongly inhibit microflora diversity and increase biomass production. Spa thermal water conditions, consisting of stagnant water, availability of organic matter and relatively high temperatures, foster the over-proliferation of Algae, which consume oxygen, alter the organoleptic properties of the water and produce toxins [27,28]. Furthermore, many, diatoms and dinoflagellates produce potent neurotoxins such as beta-N-methylamino-L-alanine (BMAA), saxitoxins, and their various isomers [29]. About 30 of the estimated 2000 dinoflagellate species on Earth produce toxins that cause human illness, mostly contracted from consuming shellfish or fish that bioaccumulated neurotoxic amino acids [29,30]. The World Health Organization realized the guidebook “Toxic cyanobacteria in water”, in 2021, to give detailed information on cyanotoxin related phenomenon. Recreational exposure to cyanotoxins is possible through: ingestion, aspiration, inhalation. Human fatalities are known only from exposure to cyanotoxins via haemodialysis. Although, a small number of severe health effects have been plausibly attributed to recreational exposure, many of the health effects that have been associated with recreational exposure to cyanobacteria are mild and self-limiting, such as irritation of the skin, mucous membranes and gastrointestinal tract; hay fever–like symptoms; nausea; and fever [23]. The main human health concern is ingestion or nasal uptake of the toxins that cyanobacterial scums may contain. Although no human deaths have been unequivocally attributed to recreational exposure, numerous deaths of livestock, pets and wild animals have been caused by consumption of water containing toxic cyanobacteria. This gives rise to concern regarding accidental ingestion of water containing cyanotoxins during recreational activities [23].
3. Environmental Epidemiology and Ecotoxicity
The attention of worldwide Health Authorities is particularly focused on HABs forming algae and cyanobacterial organisms are not directly pathogenic to humans. HABs are indeed not able to multiply in human body as the common pathogenic microorganisms, rather they produce secondary metabolites as toxins, showing elevate toxicity on both animals and humans [23]. The formation of algal blooms usually depends on the excessive release of phosphorous and nitrogen nutrients in water environments. However, although this represents the main determinant for algal blooms formation, in some cases it results restrictive for a proper evaluation of toxin risks: indeed, certain cyanobacteria, such as Planktothrix rubescens, decrease in response to eutrophic conditions instead [31]. HABs in recreational water might usually derive from the excess of fertilizers applied in soils for agricultural purposes [32]. It is essential to consider that toxins constitute a portion of algal cells lifecycle: as for Cyanobacteria, following cellular death, the lysis of the outer wall occurs, and cyanotoxins are generally released in water environments. Some species are also able to release the toxins before the cell lysis [33]. Cyanobacterial algal blooms are easily visible, producing a green-blue layer on water surfaces (i.e., planktonic cyanobacteria, typical of Microcystis sp., Dolichospermum sp. and Aphanizomenon sp. strains), grow on surfaces of rocks, sediments (i.e., benthic cyanobacteria) or they may result invisible whereas algal species sink underneath the surface [32,34].
A percentage ranging between 25% and 75% of naturally occurring CyanoHABs are potentially toxic [24]. HABs in recreational water sources (pools, spa, rivers, lake) are capable of directly impacting public health by way of water swallowing (algal toxicity is higher when ingestion route is involved), inhalation of contaminated aerosols and skin exposure (i.e., contact) during recreational activities (e.g., swimming) [23]. Beyond the capability of impacting aquatic ecosystems, the spectrum of clinical manifestations results highly variable, ranging from the mild skin irritation (i.e., dermatoxins, causing itching and rashes able to affect epidermis and dermis layers) to compromising neurological (i.e., neurotoxins) or gastrointestinal issues (since algal toxins are capable of migrating through the blood stream to liver (i.e., hepatoxins), brain, or nervous system), reaching severe neurodegenerative diseases such as Alzheimer’s disease, long-term liver damage, or cancer [32].
The global warming, and the relating climate change consistently contribute to the growth of cyanobacterial HABs across the world, affecting public health and aquatic biodiversity, by means of the increase in the leakage of toxins: a recent study has shown the augmented pathogenic Vibrio sp. occurrence in function of the increase of algal toxins [35].
3.1. Ecotoxicity of Algal Blooms and Related Toxins
HABs associated toxins are dangerous not only to humans and animals, but also to livestock. Nevertheless, few studies, focused on the characterization of potential impacts in terms of ecotoxicity, of cyanotoxins on terrestrial plants (or even, on other non-harmful algae), aquatic organisms and wildlife, are not well characterized; consequently, the effects of sustained exposure to cyanotoxins on aquatic life are poorly understood [36]. Studies, reporting data on algal and cyanobacterial toxins ecotoxicity, mainly focused on the effects of 5 cyanotoxin classes (i.e., predominantly microcystins, followed by cylindrospermopsin, anatoxin-a, saxitoxins, nodularin) on aquatic invertebrates, fishes, amphibians and birds, exposed to the toxins in freshwater habitats. The majority of research has been conducted employing a fish or aquatic invertebrate model, assessing mortality, bioaccumulation, and biochemical responses as measurable endpoints. The evaluation of the available data brings out that the 8 μg/L microcystins threshold value for recreational water regulated by U.S. Environmental Protection Agency (US EPA) is only protective for acute toxicity in aquatic organisms but not presumably against chronic toxicity [37]. Considering that in water ecosystems small invertebrates such as water fleas (Ceriodaphnia spp. and Daphnia spp.) feed on the blue-green algae, and that such invertebrates are themselves prey for other invertebrates and fish species, it is presumable that, whether small invertebrates are negatively affected by toxins produced by the blue-green algae, the entire food chain has the potential to be affected.
Experiments ,conducted by Shamohamadloo et al. (2020) on water fleas and mayfly nymphs (Hexagenia spp.) exposed to different levels of cyanobacterial toxins, showed that the exposure to toxic species did not affect mayfly nymphs, that may potentially have reduced the negative effects (possibly their larger size compared to water fleas, allows a better tolerability to toxins) [38]. On the contrary, some water flea’s species reached 100% mortality also at low toxins doses [38]. Similarly, Smutná et al. (2014), exposed Daphnia magna to varying cyanobacterial bloom samples (containing both cyanobacteria and cyanotoxins) in a series of acute (48 h) and chronic (21 day) toxicity assays [39]: 75% samples showed high acute toxicity on the crustacean bioindicator, and most samples evidenced significant lethal effects (with 35.6 mg/L biomass LC50 values) in chronic toxicity tests. Smutná et al (2014) underlined how toxicity levels were independent of the microcystin contents of the samples, consenting to hypothesize that not only toxins, but also other cyanobacterial components (e.g., lipopolysaccharides, peptides, and other unidentified metabolites) are responsible in the ecotoxicity of complex cyanobacterial blooms [39]. Indeed, previous studies evidenced how the increase of mesozooplankton was inversely correlated with the abundance of potentially toxic Cyanobacteria (e.g., Microcystis sp., Anabaena sp. and Cylindrospermopsis sp.); in contrast, microzooplankton community was not affected, rather was consistently present [40].
Palíková et al. (2007) assessed effects of different cyanobacterial biomasses (containing different concentrations of microcystins) on the embryolarval development of carp (Cyprinus carpio), demonstrating that, regardless of microcystin content or type in samples, analyzed biomasses were highly toxic. In particular, samples dominated by Aphanizomenon sp. and Planktothrix sp. strains resulted in a consistent mortality rate, while specimens in which Microcystis spp. were isolated induced lower effects [41], opening to the possibility that microcystins may not be the main responsible of HABs toxicity. This hypothesis is supported by a recent review research focusing attention on HABs threat to aquatic biota related to the potential bioaccumulation and toxicity in fish [42], nonetheless the bioaccumulation potential is lower compared to zooplankton or invertebrates [43]. Cyanotoxins were demonstrated to be negatively impacting the antioxidant system of fish species, beyond affecting the mitochondrial and endoplasmic reticulum (by means of the increase of intracellular reactive oxygen species) [44,45]. In addition, immunomodulatory, inflammatory, antimicrobial and endocrine responses were detected in animals exposed to both microcystins and cylindrospermopsins [42,46,47,48,49].
Most interestingly, less common algal groups, such as the haptophyte Prymnesium parvum and the euglenoid Euglena sanguinea, are also capable of producing harmful algal blooms (HABs), whose toxins dangerously affect aquatic biota, but did not show effects on human health. The specie Prymnesium parvum, in particular, was described as responsible for some of the worst HAB-related ecological disasters occurring in inland waters [50]. Also avian wildlife in both freshwater and marine ecosystems is heavily affected by HABs and related toxins: a monitoring conducted in Chesapeake Bay (USA) from 2000 to 2020 resulted in the notification of several mortality events associated with presumably toxic algae and HAB events [51].
3.2. Environmental Epidemiology of HABs, Algal and Cyanobacterial Toxins
Risk assessment strategies for the protection of public health connected to HABs mainly consist of avoiding that their environmental concentrations exceed hazardous levels at points of human exposure [34]. European Union drinking-water legislation framework consists of Drinking Water Directive, revised with the EU Directive 2184/2020 [52], establishing the WHO threshold guideline value of 1 μg/L for the parameter “total microcystin-LR (free plus cell-bound)”, to be mandatorily evaluated if HABs presence in treated drinking water is considered hazardous [53]. Microcystin-LR (produced by cyanobacterial strains of Microcystis sp., Anabaena sp., Anabaenopsis sp., Aphanizomenon sp., Planktothrix sp., Oscillatoria sp., Phormidium sp.) was selected as cyanobacterial and algal toxins indicator in water intended for human consumption since it represents the most widely distributed and dangerous toxin (classified by WHO as 2B carcinogen) [54,55,56]. However, taking into account that microcystin-LR may not be a reliable indicator for any other toxin, it is necessary to improve risk assessment methodologies to put in place tailored strategies to mitigate toxins release at various levels (by avoiding algal favorable proliferation conditions and select the proper water treatment, etc.) [57]. It is indeed essential to augment HABs surveillance systems worldwide, especially integrating national and international reporting activities with information regarding the algal and cyanobacterial genera responsible for harmful algal blooms events registered.
For example, since 2016, US Center for Disease Control and Prevention (US CDC) is supported by the One Health Harmful Algal Bloom System (OHHABS, until 2011 covered by Harmful Algal Bloom-related Illness Surveillance System, i.e. HABISS) in the collection of information to assess environmental risk and prevent illnesses caused by blooms of harmful algae and cyanobacteria [58]. In the 2016-2018 three-years-period, 18 US states reported 421 harmful algal bloom events, whose 81% were classified as confirmed. Summer season gathered the highest percentage (98%) of reported events, whose peaks were registered in July, with a 27% rate. Almost the totality of events was reported from freshwater reservoirs, whereas about 40% described the evidence of visible scum. The specific research of algal and cyanobacterial toxins was performed on 83% algal bloom events: overall 94% samples highlighted the presence of microcystins, while the latter samples were contaminated with other types of toxins.
HABs monitoring continued after 2018, improved with U.S. Environmental Protection Agency risk-based guidance for the quantification of algal and cyanobacterial toxins, aimed at increasing the completeness and accuracy of public health surveillance. Also in 2019, among the 242 reported algal blooms, the majority of events was predominantly evidenced between July and October; in 2019, environmental testing for algal toxins or species was performed for 88 HABs reported, and 53% confirmation of the presence of toxins, whose 93% were identified as microcystins, followed by anatoxin-a, cylindrospermopsin, saxitoxin, nodularin and haemolytic toxin. The analysis of the blooms composition at molecular level allowed the identification of Microcystis sp., Pseudo-nitzschia sp., Anabaena sp, Aphanizomenon sp., Lyngbya sp., Oscillatoria sp., Anabaenopsis sp., Gymnodiniales sp. (belonging to the toxins producer Dinoflagellate Family), Phormidium sp. and Planktothrix sp. In 2020, 227 HAB events were notified by 13 states: summer months, and in particular July (with a 26% reporting rate) resulted the periods with higher number of events. Strain identification analysis at genus level allowed the confirmation of the same genera isolated in 2019, beyond further dinoflagellates (Gonyuaulacles sp., and Dinophysos sp.) and xyanobacteria, such as Nodularia sp., connected to the detection of nodularin toxin in one sample. Last reported OHHABS data refer to 2021, where 368 HAB events were reported, with a 50% higher retrieval rate compared to the previous three years. Also in 2021, most events were registered during summer season, with a 25% peak in August, bringing out an overall 85% events confirmation. Environmental testing allowed to determine that the majority of HABs events reported in 2021 were caused by Cyanobacteria (mostly Microcystis spp., Cylindrospermopsin spp., Planktothrix spp.), dinoflagellates and Diatoms (mainly strains of Pseudo-nizschia sp.), predominantly producing microcystins, anatoxins-a, saxitoxins, and cylindrospermopsins [59]
The effect of toxins producing HABs on freshwater and marine ecosystems, and the impact on public health, are extremely harmful to some business sectors like tourism and fishing industries: indeed, in the EU, the impact on such industries in terms of annual cost of HABs is estimated exceeding 918 million euros [54]. A substantial amount of data has been gathered in the “Harmful Algal Event Database” (HAEDAT), a meta database collecting records of harmful algal events: HAEDAT contains records provided by the ICES area (North Atlantic) since 1985, and by the PICES area (North Pacific) since 2000. Last HAEDAT report shared useful data on the total number of harmful algae events globally notified by single countries during the 1980-2015 period: the highest number of notifications belonged to France (894 events), United States (620), Canada (521), Portugal (475), Japan (450), and Norway (305) [54,60].
Microcystin-LR monitoring of twenty-four recreational water reservoirs in Eastern Cuba brought out concentrations exceeding WHO limits for drinking water in about 30% sources [61]. The evaluation of HABs in a drinking water treatment plant Macapa (Brazil) reported values of 2.1 µg/L in 2015, coinciding with a sensible increase in Limnothrix planctonica (cyanobacterium) density [62]. A monitoring performed in Qatar on drinking water sources resulted in the detection of microcystin-LR concentrations exceeded WHO threshold value (up to 1.33 µg/L) [63]. Douma et al. (2010) conducted a study in Morocco, exposing mice to cyanobacterial HABs algal biomass extracted from freshwater, aimed at evaluating the toxicity calculating the lethal dose 50 (LD50) [64]: the results demonstrated how the toxicity in mice was positively associated with the quantity of microcystin-LR present in the biomass [64]. A research, conducted in southern Spain drinking water treatment plants from 2016 to 2019, showed the prevalence of strains of Merismopedia sp. (generally toxic, but no toxins were detected in the analyzed samples), Planktothrix agardhii (more typical of northern Spain, than southern), Woronichinia naegeliana and Microcystis aeruginosa (usually both rarely isolated) [65].
In Italy, the first observations on algal blooms and related toxins date back to 1977. Noteworthy an impressive blooming of toxic Cyanobacteria affecting, in 1985, two large artificial reservoirs of Sardinia Region (the Medio Flumendosa and the Mulargia, built to supply drinking water). From both sites the dominant presence of Planktothrix rubescens, Microcystis aeruginosa, and Dolichospermum planctonicum (formerly classified as Anabaena planctonica) [65,66]. Nowadays, this phenomenon is periodically documented by Italian Health Authorities in the peninsula, in regions such as Lombardy, Trentino-Alto Adige, Emilia Romagna, Tuscany, Lazio, Molise, Sicily, Calabria and Sardinia, occurring with increasing frequency even the other regions. The most common toxic species of cyanobacteria in Italy are: Anabaena solitaria f. planctonica; Aphanizomenon flos-aquae; Aphanocapsa spp.; Cylindrospermopsis raciborskii; Dolichospermum planctonicum, D. flos-aquae, D. lemmermannii, D. crassum, D. smithii; Lyngbya sp.; Microcystis aeruginosa, M. flos-aquae, M. wesenbergii, M. panniformis; Planktothrix rubescens, P. agardhii; Pseudoanabaena spp.; Woronichinia spp. As regards the genus Planktothrix, the species P. rubescens is usually isolated from Italian water sources; while for Microcystis genus, M. aeruginosa is generally [67]. It however is complicated to establish a list of the most widespread species, considering the lack of quantitative data on abundance, moreover mostly relating to sporadic episodes and not to annual series. Also, analyzing literature, the most common toxic species belong to the genera Microcystis, Planktothrix, Aphanizomenon, Dolichospermum, Anabaena and Cylindrospermopsis [68].
In a One Health approach, environmental epidemiologists, animal health experts, professionals involved in the protection of public health, academics and stakeholders should act in synergy to empower the awareness and understanding risks linked to harmful algal bloom events, generating networks and strategies to improve the capability to monitor, detect, and quickly notify HABs events and connected illnesses.

Table 1.
Main toxins producing genera, produced toxins, their classification and WHO threshold value in recreational water (Information collected from the Report “Algal Bloom and its economic impact”, JRC Technical Report, 2016) [54] (a : Reference values collected from “WHO Guidelines on Recreational Water Quality: Volume 1 Coastal and Fresh Waters” [23].
Table 1.
Main toxins producing genera, produced toxins, their classification and WHO threshold value in recreational water (Information collected from the Report “Algal Bloom and its economic impact”, JRC Technical Report, 2016) [54] (a : Reference values collected from “WHO Guidelines on Recreational Water Quality: Volume 1 Coastal and Fresh Waters” [23].
| Toxins procuding genera | Toxin category | Toxin classification (based on effects on humans and animals) | WHO guideline value in recreational watera | References |
|---|---|---|---|---|
| Anabaenopsis, Aphanizomenon, Dolichospermum (formerly, Ananbaena), Mycrocystis, Oscillatoria, Phormidium, Planktothrix | Microcystins | Hepatotoxins | 24 µg/L | [70,71] |
| Nodularia, Nostoc | Nodularins | Not established | [70,71,72] | |
| Aphanizomenon, Cylindrospermopsis, Dolichospermum (formerly, Ananbaena), Lyngbya, Oscillatoria, Raphidopsis, Umezakia | Cylindrospermopsin | Cytotoxins | 6 µg/L | [70,73] |
| Aphanizomenon, Cylindrospermopsis, Dolichospermum (formerly, Ananbaena), Oscillatoria | Anatoxins | Neurotoxins | 60 µg/L | [74,75] |
| Aphanizomenon, Cylindrospermopsis, Dolichospermum (formerly, Ananbaena), Lyngbya, Planktothrix Raphidiopsis | Saxitoxins | 30 µg/L | [70,76,77] | |
| Aphanizomenon, Dolichospermum (formerly, Ananbaena), Mycrocistis, Nodularia, Nostoc | b-Methylamino L-Alanine (BMAA) | Not established | [78,79] | |
| Anacystis, Dolichospermum (formerly, Ananbaena), Microcystis, Oscillatoria, Schizothrix, Synechococcus | Lypopolysaccharides | Dermatoxins | Not established | [80,81] |
| Lyngbya | Lyngbyatoxins | Not established | [82,83] | |
| Lyngbya, Oscillatoria, Schizothrix | Aplysiatoxins | Not established | [84] |
3.3. Algae and Cyanobacteria in Spa and Thermal Spring Water Sources
Thermal spring basins are globally exploited for recreational and medical purposes, being the thermal-mineral water and muds rich in minerals and bio-active compounds resulting from the metabolic pathways of the autochthonous microbiota and microflora [4], playing an essential role in the beneficial effects of these sources. Most health authorities worldwide and, in particular, European Union countries have established that thermal water springs (i.e., spa water) should be left untreated, nonetheless the risk of not employing disinfection treatments may pose health risks of bathing in thermal-mineral water pools [3] not undergoing continuous water renewal. Most national and international regulations assume that the employment of chemical disinfectants (e.g., sodium hypochlorite or hydrogen peroxide) on thermal basins is in contrast with their healing effects, considering the high sensitivity of microbiota and microflora composition to common environmental biocides [85]. Despite the substantial lack of studies focusing on the mode of action of the different spa waters, World Health Organization (WHO) recognizes therapeutical water, such as hydrothermal sources, among the oldest global healing means, as a traditional medicine strategy, intended for therapeutic and prophylactic purposes towards several pathologies [86], beyond being considered part of the cultural and medical traditions in most Central European countries [87]. Worldwide, Health Authorities [88,89,90] and, in particular, Italian Health Ministry, do not allow common drinking waters’ disinfection treatments on thermal-mineral waters: Health Ministry Decree 25/2012, regulating treatment strategies for water intended for human consumption, precises that thermal-mineral water sources are excluded from any disinfection treatment [91]. Also at regional level, chemical disinfection of thermal water is forbidden, while physical technologies are encouraged [91,92,93].
Nevertheless, water disinfection would sensibly reduce health risk, further ensuring the adequate health status of spa basins, the sanitation treatment may strongly affect non-pathogenic and autochthonous microbiota, leading to a strong reduction of the healing effect of the sources [3]. Moreover, while microbial diversity of spa water can control the survival and proliferation of the pathogenic microbiota [94,95,96] by means of the production of bacteriocin-like active molecules, demonstrated able to reduce both Coliforms and Staphylococci [97,98], the potentially beneficial microorganisms may not be capable of contrasting biofilm formation in pools’ walls and water distribution systems. Spa water microbiota may host virulent species of Legionella spp. and Mycobacterium spp., capable of surviving in amoebic organisms, exploiting the production of biofilms, literally protecting the pathogens from extremophilic conditions and hiding them from endogenous microbiota [99]. The most common family of algae occurring worldwide is Bacillariophyceae, dominant in both western and eastern Europe, according to the algological studies [4,100].
Kiliç et al. (2022) conducted a research on the Delicermik-Koprukoy hot spring water (Erzurum, North East of Turkey), a sodium-calcium-bicarbonate-carbon dioxide chemically classified sources, reporting the dene growth of algae belonging to Bacillariophyta, Chlorophyta, Cyanophyta and Euglenophyta [4,101]. Giorgio et al., (2018) studied the microflora composition of spring water and mud samples collected in a thermal-mineral basin in Naples (Regione Campania, Italy), reporting the isolation of 7 green Algae, 2 Cyanobacteria and 7 Diatoms taxa: above all, the most abundant benthic microalgae were strains of Amphora ovalis, Anabaena sp., Chlorella sp., Coccomyxa sp., Cocconeis placentula, Gomphonema acuminatum, Leptolyngbya sp., Monodus sp., Navicula cincta. Nitzschia palea Nostoc sp., Pseudococcomyxa simplex, Rhoicosphenia abbreviate, and Scenedesmus sp. [100].
The isolation of Cyanophyceae, Chlorophyceae and Bacillariophyceae, overall non-pathogenic microorganisms, was confirmed by similar studies reporting the ability of the algal species of producing macromolecules holding therapeutic properties [102,103,104,105]. The Cyanobacteria Anabaena sp., Leptolyngbya sp. and Nostoc sp. produce compounds showing cytotoxic effects towards human cancer lines [105,106]; Strains of Nostoc sp., are capable of producing lipophilic extracts showing antibacterial activities and antiviral proteins [107], while Chlorella sp. algal extracts have antitumoral and anti-immunostimulatory activities [108]. A polysaccharidic compound isolated from strains of Coccomyxa sp. resulted effective in contrasting the influenza A virus infection [109]. Species of Nostoc sp. have also been identified as producers of microcystins, harmful toxins used as indicators in water intended for human consumption (according to the Italian regulation D. Lgs. 18/2023). While Anabaena sp. strains produce anatoxin-a and anatoxin-b [110,111].
Stoyneva (2003) described Bulgarian algal flora in thermal springs, underlining the detection of over 200 species, consisting of cyanoprokaryotes, green, yellow-green and red Algae, Diatoms and Glaucophytes; in general, Chlorophyta, whose 75 taxa were isolated, were the most abundant [112]. The isolation of Diatoms was furthermore highlighted by Lai et al., (2023), analyzing 65 Diatoms genera (of which, 196 species) and their assemblages in thermal springs connected to rivers in Regione Sardegna (Italy) [113].
Ulcay et al. (2017) reported a monitoring aimed at identifying algal species in Germencik (Alangüllü) thermal water springs (Turkey): overall, 21 Cyanobacteria (more tolerant to the majority of ecological extreme conditions, such as high-water temperature), 5 Bacillariophyceae and 1 Conjugatophyceae taxa were isolated. Characterized species belonged to Aphanothece, Chroococcus, Pseudanabaena, Spirulina, Leptolyngbya, Heteroleibleinia, Phormidium, Oscillatoria, Navicula, Achnanthidium, Kamptonema, Rhopalodia, Amphora, Surilella and Spirogyra genera. The most interesting species were Leptolyngbya subtilis, Leptolyngbya thermobia and Heteroleibleinia kossinskajae [114]. Leptolyngbya species were also isolated from salty water springs in Hawaii, Sri Lanka, Australia, Japan, USA, Czech Republic [115], water sources in Netherlands [116], thermal water springs in Greece [117]. A monitoring of algal flora of thermal water springs located in Egypt allowed the isolation of 209 species from 8 algal Phyla: Cyanobacteria (91 species), Chlorophyta (59), Bacillariophyta (52), Chrysophyta, Xanthophyta, Euglenophyta, Dinophyta and Charophyta [118]. Gupta (2017) showed that most of the algal flora isolated from thermal water springs located in Jharkhand, India, belonged to Cyanophyceae (25 strains), Bacillariophyceae (4), Chlorophyceae (2) and Euglenophyceae (2) families [119].
Several studies have reported the isolation of toxin- producing algae, such as Synechocystis spp. and Cylindrospermopsis raciborskii from hot spring water in Europe: the possible chemical treatment of toxin-producing algae and cyanobacteria may lead to the cell disruption and consequent release of the harmful toxins in water environment, increasing the exposure to bathers [3,120,121].
4. Clinical Epidemiology of HABs, Algal and Cyanobacterial Toxins
Out of the over 5000 known extant species of phytoplankton, about 200 can harm human health and food security by producing toxins [122]. These toxins may interfere with the recreational use of coastal or inland waters and cause economic losses. The IOC-UNESCO Taxonomic Reference List currently lists 105 dinoflagellates, 37 marine cyanobacteria, 31 diatoms, 8 haptophytes, 6 raphidophytes, 3 dictyochophytes, and 2 pelagophytes that produce toxins [123]. In addition to toxic species, HABs can be related to non-toxic microalgae producing high biomass and causing seawater discolorations, anoxia, and mucilage that negatively affect the environment and human activities. The occurrences of harmful microalgal species are now routinely recorded in the Ocean Biodiversity Information System, OBIS (https://www.obis.org), a global database on the diversity, distribution, and abundance of all marine organisms. Since 1985 the OBIS reports of harmful algal events impacting on human society (Figure 2).
Harmful Algal Blooms (HABs) can cause several syndromes in humans that mostly depend on the mainly route of exposure: in fact, the illnesses vary from respiratory symptoms like throat irritation, nasal congestion, cough and wheeze, gastrointestinal signs like nausea, vomiting, diarrhea, abdominal cramps and dermatological symptoms like dermatitis, rash, eye and ear irritation. In addition, algal poisoning often seems to target the depurative organs like the liver and kidneys, leading to hepatoenteritis, acute kidney disease and organ failure with elevated serum enzyme levels [123]. In rare unlucky cases HABs can cause fatal events..
As regards the economic impact on healthcare, a study conducted by Kouakou et al. reports an estimation of the medical cost’s which are $86, $1,015 and $12,605, respectively, for mild, moderate and severe cases of digestive illnesses and $86, $1,235 and $14,600, respectively, for mild, moderate and severe respiratory illnesses [124]. However, due to the absence of reliable surveillance systems, accurate estimates of their impact on human health, particularly in the long term, are not available. Additionally, evidence regarding the potential lasting effects of both acute illness and chronic exposure to HABs is lacking [125]. Diseases are reported globally, but there is a dearth of information on outcomes from Africa and South America. This may indicate under-reporting or neglected diseases in the English language literature from these continents. Conversely, monitoring programs have been successful in Europe, where commercial activities such as aquaculture are more prevalent than wild harvesting. In a significant proportion of studies, exposure assessment was conducted to confirm the diagnosis. The assessment involved the use of proxy measures. About 25% of the papers used direct measures of toxin levels in the food consumed. In contrast, around 33% of the literature used levels of toxins and HAB species that were linked geographically and temporally to harvesting or direct exposure areas.
Despite the well-known toxicity of harmful algae, some people voluntarily consume certain types of algae, such as cyanobacteria, due to their high protein content and potential health benefits, including detoxification, elevated mood and energy, increased alertness and vivacity [126,127,128]. However, Roy-Lachapelle et al. (2017) discovered that certain algae dietary supplements contained cyanotoxins at levels that exceeded the tolerable daily intake values [129]. The literature contains numerous articles discussing the health effects of exposure to harmful algae. Indeed, research related to “Algae” and “Human health” has exponentially increased over the last decade (Table 2).
Table 2 summarizes the main characteristics of 49 studies. There is a clear trend of an increasing number of studies conducted over time. A small percentage (28,57%) of all studies reported cases of Cyanobacteria, followed by Paralytic Shellfish Poisoning (6,12%). The most recurrent outcome reported is damaged respiratory function described in 63,26% of the studies, followed by 42,85% about gastrointestinal illness and 36,73% dermatologic symptoms. Not all diseases reported due to contact with water potentially infected with algae are directly caused by algae. There are suspected cases (Figgatt et al. 2017). However, some cases are confirmed to be directly caused by toxic algae, including cyanobacteria. The studies were mainly comprised of case reports (53.9%), surveillance data, and ecological studies (23.2%), and anecdotal reports of illness (9.8%). The clinical outcome was most commonly reported in North America (44.2%), followed by Australasia/Oceania (21.8%) and Asia (17.1%). Ingestion through a food vector was the most commonly reported route of exposure (93.7% of studies). However, inhalation and direct contact with seawater were also reported as routes of transmission. A few studies, all describing Ciguatera Poisoning, reported possible transmission through breastfeeding, sexual contact, and placental transfer [178]. Todd (1989c) proposed that reported cases of disease caused by toxic algae from food contamination are underestimated [177]. He suggests a multiplier of 10 to account for the total number of cases.
5. Anti-Algae and Treatments
The maintenance of modern public swimming pools involves several chemical substances used to ensure water quality and users safety, including sodium and calcium hypochlorites, chloramine and trichloroisocyanuric acid as disinfectant agents and algaecides [178,179,180,181]. Algae growth in pool waters can be promoted by warm temperature (T), sunlight exposure time, pH value, carbon dioxide (CO2 level, phosphate level, amount of other organic contaminant and last by disinfection capacity [182]. Chlorine itself can be used to control algae level. It has been estimated that a chlorine’s free concentration of 10 mg/L for several hours is enough to kill off algae [183].
Algaecides are chemical substances designed to inhibit algae growth [183]. Their use is regulated by EU Biocidal Products Directive 98/8/EC implemented in the member states by May 2000. The control of algae in swimming pools is of great significance since their presence make the water green or cloud. Due to the increase of private and public swimming pool facilities, especially in urban areas and the rising awareness about the importance of pool hygiene and regular maintenance, the swimming pool algaecides trade is growing. According to Data Bridge Market Research Report (2022), the global algaecides market including application to surface water, home use, hotels and resorts, commercial pools & Spas, aquaculture and agriculture, is expected to account for USD 7,127.56 Million by 2029. Recent advancements allowed to develop multi-functional algaecides able to prevent the growth of algae and contemporary contribute to water clarification. The formulation of long-lasting algaecides with slow-release properties together with those specifically designed for saltwater pools represent the most emerging trends in this field.
Swimming pool algaecides market include compounds such as Quaternary Ammonium Compounds (QAC), Metallic algaecides and others. Factors affecting the effectiveness include: the species of the target algae concentration, specific characteristic of the product, the duration of exposure and different environmental parameters, such as alkalinity, pH, dissolved organic carbon (DOC), and conductivity [183].
Depending on specific characteristics, the use of these compounds can produce environmental and health implications. For instance, when QAC are used in combination of chlorine can become a precursor of disinfection byproducts (DBPs) [185]; copper-based algaecides can cause metal stains or turn the swimming pool water into a clear green solution when copper is oxidized by chlorine [182,186].
5.1. Quaternary Ammonium Compounds (QAC)
The substances referred as quaternary ammonium compounds (QACs) are characterized by a cationic headgroup and one hydrophobic hydrocarbon side chain at least [187]. QACs are typically classified in three main groups: alkyl-trimethyl-ammonium compounds (ATMACs), benzyl-alkyl-dimethyl- ammonium compounds (BACs) and dialkyl-dimethyl-ammonium compounds (DDACs) whose main characteristics have been listed in Table 3 [188].
The two homologues with 12 and 14 carbon atoms in the aliphatic chain (BAC-12 and BAC-14) represent the most used formulations in the wide list of commercially available BAC mixtures. The effectiveness of QACs depends on their structure but generally destabilize the cell membrane, causing leakage of intracellular low-molecular-weight material, proteins and nucleic acids, resulting in rapid cell lysis [191].
Recently QAC have been candidate to be recognized as persistent and mobile chemicals (PMs) due to their high mobility (LogKow < 4.5), water solubility and half-lives [188], as shown in Table 3. The concentration detected worldwide vary from less than 1 μg/L in surface water and to 100 μg/L in wastewater treatment plant effluents and can be as high as 1200 μg/L in raw wastewater. When released in marine ecosystem, QAC determine a reduced cellular viability and an alteration of the antioxidant mechanisms of the marine invertebrate Mytilus galloprovincialis at sub-lethal concentrations (~ 100 μg/L) [192].
Zheng et al. (2022) recently published the first detection of quaternary ammonium compounds in breast milk, registering concentrations ranging from 0.33 to 7.4 ng/mL, making of breastfeeding a significant QACs exposure for infants [193].
5.2. Metallic Pool Algaecides
Metallic pool algaecides contain copper or silver ions that effectively kill algae. Copper-based algaecides, such as CuSO4, were first introduced in 1904 and are the most widely used algaecides [194]. Copper compounds are highly effective against protista algae cells. Algaecides with copper compounds typically contain organocopper or chelated copper.
Bishop et al. (2014) evaluated responses of eight target algal species (Cymbella tumida, Ankistrodesmus falcatus, Haematococcus pluvialis, Pandorina charkowiensis, Eudorina elegans, Nostoc punctiforme, Microcystis aeruginosa and Desmidium sp.) (Figure 3a) [195] and five nontarget animal species (Daphnia magna, Ceriodaphnia dubia, Pimephales promelas, Hyallela azteca, Lepomis macrochirus) (Figure 3b) to exposures of a copper-based algaecide (a.i. copper 5%) in 96-h laboratory toxicity tests. The copper concentrations required to achieve control (i.e. 96-h EC90) of the targeted algae were significantly higher than those corresponding to 96-h LC50 for the selected animal species, indicating significant risks to nontarget species. . Among the tested, D. magna was the most sensitive invertebrate.
More recently, novel copper-based algaecides have been developed to reduce the toxicity towards non-target organisms and to improve efficiency against algae. These formulations include chelator agents that can pass through algal cells membranes and cause cell lysis rapidly [196]. However, the authors who tested these compounds reported the effectiveness and toxicity strictly dependent on water chemistry.
5.3. Algae Removal
The eutrophication caused by the releases of urban and industrial discharges containing high concentration of nutrients in receiving water bodies such as rivers, lakes and ponds leads to great threats to drinking water safety since the algal organic matter is a precursor of disinfection by-products (DBPs) during the chlorination process [197,198]. Moreover, the rupture of algal cells in water represents a further threat to human health due to the release of toxic substances, such as microcystins (MCs). To manage the eutrophication of lakes several technologies have been attempted in the last 30 years, including physical, biological and chemical methods [199]. Flushing and dilution, deep aeration, and mechanical algae removal are the most common physical methods used to remove algae from small lakes but are limited only to these environments due to their high cost and complex operation. Biological methods are based on plant allelopathic, stocking filter-feeding fish, biological activated carbon, biofilm pretreatment, feeding zooplankton, simulated artificial wetland. However, their use needs to be carefully evaluated to avoid causing damage of the biological structure and biodiversity of lakes. Chemical methods presents low cost and high efficiency. The addiction of chemical reagents into the algae-laden water allow the inactivation and algal cell’s death through chemical reactions.. Small size and low density make difficult the removal of harmful algae by water treatment facilities. However, more than 95% of algal cells can be removed by drinking water treatment plants (DWTPs) which are constituted by the following units: coagulation, sedimentation, filtration and disinfection. Coagulation-flocculation (C/F) process is recognized as one of the most efficient treatment methods to remove cyanobacterial blooms CyanoHABs in (DWTPs). Conventional coagulants and flocculants used for water purification allow to achieve high performance of removal, however their residuals (e.g., aluminum) in treated water are suspected to be related with neurodegenerative diseases such as Alzheimer's, as well as neurotoxic disorders and carcinogenic effects [200]. For algae-laden water a high dosage of coagulants is required. As consequence, a large amount of sludge is produced. The treatment and the management of this sludge determines an increase of the overall treating costs. Thus, in last decades, research studies focused on the development of new eco-friendly substances, safe, efficient, and cost-effective. Natural coagulants represent a promising and environmentally friendly alternative to the conventional ones [201,202]. El Bouaidi et al. (2020) reported as 0.5 g/L of Vicia faba seeds and 1 g/L of Opuntia ficus indica cladodes allowed to remove up to 85% of M. aeruginosa from treated water [203]. Moreover, the application of natural coagulants allows to reduce the sludge production from DWTPs and to use the produced sludge in agronomic application since its biodegradability and the missing of harmful substances. In order to upgrade DWTPs, the algae removal efficiency can be improved by adding pre-oxidation (pre-chlorination, potassium permanganate and ozone pre-oxidation), air flotation and advanced treatment processes (ultrafiltration membrane, ozone, activated carbon filtration and advanced oxidation process) [204,205].
6. Detection Methods of Algae
Several methods for detecting algae have been developed, but they lack a unified classification standard. Among technologies, there are morphological structure, cytochrome, immunoassays, and nucleic acid methods.
Traditionally, algae have been monitored using morphological structure-based detection methods, such as morphological observation under light microscopy [206]. These methods include traditional Microscopic Examination Technique (MET), considered the gold standard technique, which is capable of simultaneously providing data on morphology, composition, and abundance within real samples [207]. An evolution of MET is Image Identification Technique (ITT), developed to overcome the limitation of MET in the study of harmful algae characterized by too small size to be analysed without an optical microscope [208]. An application of IIT is Imaging Flow Cytometry (IFCM), characterized by combining FCM and computer image processing technology to realize the automation of cell image capture and recognition [209].
Focusing on the advantage of traditional methods of quantification and identification of algae in recreational waters, the utilization of microscopy allows both cell counting (commonly, using the relative abundance) and taxonomic analysis of algal cyanobacterial blooms. Usually, epifluorescence and inverted light microscopy with sedimentation chambers are employed, following samples staining with selective dyes (consenting to distinguish autotrophic and heterotrophic cells). Microscopy analysis however does not allow the distinction between toxic and non-toxic species (if not presumptive), resulting in a lack of information in the risk assessment of algal blooms [210]. Structure-based techniques have several limitations such as the need for highly trained personnel and it is not efficient for analysis in a short and timely manner, especially for many samples.
Therefore, multiple techniques have been developed over the years to achieve better performance from the point of view of speed and accuracy. It is possible to organize all these new strategies in three different macro-groups: i) Cytochrome-Based Detection Techniques (CBDT), ii) Immunoassays and iii) Nucleic acid-based detection.
The first group of methods includes techniques that are mainly based on the investigation of characteristic pigments and are named CBDT (cytochrome-based detection techniques) such as High-Performance Liquid Chromatography (HPLC), Absorption Spectral Analysis (ASA), and Fluorescence Spectral Analysis (FSA) [208]. HPLC is known for being able to perform both qualitative and quantitative type analysis with high reliability, as well as being able to perform parallel analysis. On the other hand, however, chromatographic techniques need sample pretreatment steps and often require potentially toxic organic solvents that limit their use [211]. ASA techniques measure the absorption of magnetic radiation as a function of a characteristic frequency and wavelength of different spectral ranges: IR and UV-VIS. The absorption of electromagnetic radiation by pigments will produce characteristic spectral bands with different shapes and intensities depending on the type and concentration of the specific pigment. It is therefore used as a diagnostic element while it is limited in quantification because of the nonlinearity of the response due to mutual interference of multiple pigments. The study of pigment transmission spectra and quantification of photosynthetic components also provide data on algal life; in fact, the amount of photosynthetic pigmentation is a growth index [212]. Many pigments exhibit the phenomenon of fluorescence, thus the emission of light following excitation; this technique has multiple advantages; first, it allows quantitative analysis due to the linearity of the response, it is also easy to use, it has high sensitivity and it does not require any pretreatments, allowing even on-site and instantaneous analysis due to the portable instrumentation. Moreover, algae have different cell wall composition, and for this reason, IR micro-spectroscopy allows us to distinguish different species by producing different spectra based on their major components [213].
Aimed to isolating, identifying and quantifying cyanobacteria, algae, cyanotoxins and algal toxins, worldwide Health Authorities sustain that the most useful analytical means are molecular methods, including immunoassays and nucleic acid-based detection [23]. Like any methodologies, molecular evaluations for HABs detection in recreational water hold both advantages and disadvantages: undoubtedly, limitations in “old generation” methods (e.g., through morphological and cultural classification) might be overcome by integrating the analysis with recently developed innovative methodologies, such as AI/machine learning, biosensors employment and satellite imaging utilization. The present paragraph aims at describing both conventional and recently designed techniques for Algae, Cyanobacteria, and toxins characterization, to be utilized for recreational water ecosystems, such as lysis methodologies for cyanobacterial cells, enzyme-linked immunosorbent assays (ELISA), polymerase chain reaction (PCR) and DNA sequencing techniques, etc. [214].
To perform an adequate analysis of cyanotoxins, the total lysis of cyanobacterial cells is needed, especially to measure the concentration of intracellular cyanotoxins. Indeed, the quantification of cyanobacterial density is usually assayed through direct (assaying cell mass) or indirect (analysing intracellular biomolecules) methodologies [215]. For example, US EPA requires measuring the total concentration of cyanotoxins, which is the sum of the intracellular and extracellular amounts [216,217]. Cell lysis can be performed using chemicals, ultrasonication, and physical methods [215].
Chemical lysis aimed at disrupting cell membranes exploits chemical compounds such as enzymes (such as proteinase K and lysozyme), surfactants (e.g., Triton-X 100), reducing agents (e.g., β-mercaptoethanol) or organic solvents (e.g., phenol) to disrupt the cell membrane [218]. Ultrasonic cell lysis consists of variating sonic pressure causing cavitation, able to break down both cell walls and membranes [219,220]. Physical methodologies to lyse algal and cyanobacterial cells consists in mechanical (e.g., bead beating, commonly using ceramic glass beads) and cryogenic strategies (mainly lyophilization and freeze–thawing) [218,221]. The advantage of using physical methods or ultrasonication is to avoid chemical contamination issues. In general, especially for molecular characterization and -omics methods, bead beating is considered the most efficient cell lysis method [221], although several studies reported the highest lysis yield by utilizing multiple lysis methods in succession, especially to disrupt membranes and walls of the most resilient algal and cyanobacterial species [222,223,224].
For toxins detection and quantification, in addition to microscopy methods, US EPA usually recommends high-performance liquid chromatography coupled with either mass spectrometry (i.e., LC-MS) or ultraviolet/photodiode array detectors, enzyme-linked immunosorbent assays (i.e., ELISA, exploiting competitive binding between antibodies and the targeted toxins antigens), and protein phosphatase inhibition assays (PPIA) [217,225]. The above-mentioned methods indeed are highly sensitive to microcystin detection: ELISA-based techniques, for example, are rapid (2 h) and user-friendly as well (not needing samples pre-concentration, and employing small sample volumes), being able to detect a minimum of 0.02–0.07 ng/mL microcystins. Most commercially available ELISA kits for microcystin detection are based on an assay able to bind the ADDA β-amino acid, (4E, 6E 3-amino-9-methoxy-2, 6, 8-trimethyl-10-phenyldeca-4, 6-dienoic acid), present in most of the microcystins and nodularins penta- and heptapeptide congeners [226]. Nevertheless, assays based on ELISA are not capable of differentiating microcystin congeners [227,228] and lack of specificity may be experienced due to cross-reactions with microcystin degradation byproducts and metabolites, bringing to overestimation and false-positive results in the assessment of cyanotoxicity [229,230]. In contrast, chromatography coupled with tandem mass spectrometry are highly sensitive, allowing to discriminate between different cyano-toxin congeners [231,232], using the proper standards, but only a few of them are commercially available [233]. LC-MS-based methods use chromatography (coupled with tandem mass spectrometry) to purify cyanotoxin by affinity-based, hydrophobic/hydrophilic, ionic mobility passing through a separation medium [232].
In the last decade, attention has been focused on the employment of the most sensitive molecular biology techniques, consisting of the “old generation” polymerase chain reaction (PCR), and the more advanced quantitative real-time PCR (qPCR), and DNA microarrays [217]. While conventional PCR works using the amplification of organism-specific genomic DNA sequences, providing qualitative outputs (i.e., presence/absence), qPCR and real-time PCR, exploiting a dye or a probe that incorporates DNA binding to quantify the DNA target (cyanobacterial or cyanotoxin gene copy number) in purified DNA (e.g., DNA extracted from an environmental sample, including algal blooms) [234]. In addition, the employment of reverse transcription qPCR (RT-qPCR) allows the analysis and characterization of the expression of transcribed cyanotoxin genes, consenting to diversifying active cells from quiescent [235].
Molecular methods to identify and quantify cyanotoxins use mcyE (microcystin gene cluster) as a target gene to detect all potential microcystin producer species, beyond the nodularin synthetase gene clusters [236]. Considering the most commonly used target genes, while assays targeting mcyA (Microcystin Synthetase A) gene allows the detection of the majority of microcystin-producing strains, assays targeting mcyE and mcyG genes are specific to Microcystis sp. and Planktothrix sp. Strains [237,238]. Gupta and Matthews (2010) targeted conserved signature proteins (CSPs) and conserved signature indels (CSIs), which were found to be highly specific to a particular clade of organisms: the protocols used consented to increase the detection specificity [239,240,241]. HABs monitoring, conducted in the Ohio inland lakes (Ohio, USA) between 2020 and 2022, allowed to demonstrate that qPCR and RT-qPCR can be considered extremely useful early warning tools to characterize cyanobacterial blooms and to identify cyanotoxin production [241,242,243]. Furthermore, compared to traditional microscopy and cell counting methods, qPCR is undoubtedly more rapid, providing results in 3 hours [242]. Recent advances, associating PCR techniques with followed by high-throughput sequencing of the 16s rRNA gene (e.g., Next Generation Sequencing), are being employed as risk assessment ameliorating strategies [243]. However, the issue in the exploitation of innovative molecular methodologies is mainly linked to the lack of standardization of such techniques for the detection of harmful algal blooms and related toxins.
Moreover, alternative high-throughput methods to monitor recreational waters include atomic force microscopy (i.e., AFM), which consents, in the presence of metal ions, to visualize the formation of microcystins strands, to investigate the adhesion properties of algal cells additionally [244,245,246].
Future directions point at the development of devices based on microfluidic (i.e., optical analysis) and DNA capture technologies: such tools allow to incorporate target-specific probes or fluorophores capable of binding to target molecules in cyanobacterial blooms extracts returning a specific signal related to the quantity of the investigated molecule [247,248]. The advantage of microfluidic devices is related mainly to their portability, especially to the high sensitivity and the possibility to use small sample volumes [249].
Certainly, the high increasing quantity of multiple data on recreational water ecosystems brings to consider artificial intelligence (i.e., AI), machine-learning, and -omics technologies algorithms as absolutely valuable tools in predicting the proliferation of harmful algal blooms and the consequent production of dangerous toxins (e.g., CyanoHABs tool) [250,251,252]: these tools, these tools, whether properly standardized through precise guidelines and policies, may reduce the necessity of onsite expertise in performing cyanobacterial monitoring [253,254].
7. Risk Assessment, Laws and Water Safety Plan
To support the risk assessment from recreational water exposure, WHO has established guideline values to cyanotoxins surveillance [255]. The potential for cyanobacterial bloom formation or the growth of algal species in a water body is determined by its environmental conditions, such as nutrients and light. Additionally, hydro-physical conditions, such as the rate of water exchange and vertical mixing of the water, can also have a role.
Due to the complexity of phytoplankton ecology in water bodies, it is highly recommended to involve limnological expertise when assessing the risk of occurrence of cyanobacterial blooms. Two chapters in Toxic cyanobacteria in water give guidance for this: Ibelings et al. (2021), on phytoplankton ecology, and Burch, Brookes & Chorus (2021), on assessing water body conditions and management [256,257].
In this sense, the most important pieces of EU legislation that apply to algae are the Habitats Directive (Council Directive 92/43/EEC), the Marine Strategy Framework Directive (Directive 2008/56/EC) and the Water Framework Directive (Directive 2000/60/EC).
The World Health Organization (WHO) guidebook Toxic cyanobacteria in water (Chorus & Welker, 2021) gives a comprehensive overview of the information and expertise needed to assess the risk of cyanotoxin occurrence, including for recreational water use, and for developing effective risk management strategies. The information below is largely summarized from specific chapters of this book, unless cited otherwise. The WHO background documents for four groups of cyanotoxins (microcystins, cylindrospermopsins, anatoxin-a and saxitoxins) give detailed information on the derivation of WHO guideline values, including for recreational exposure (WHO 2020a, b, c, d, e).
According to WHO (2021) secondary metabolites contained in some species of HABs are toxic to humans and animals [23]. Depending on water bodies HABs grow on the sediment or on the surface of submerged macrophytes, as in the case of fresh water or can be observed on clumps on the sediment down to depths of 30 m as in marine subtropical and tropical coastal areas. Skin blistering, oedema and deep skin lesions have been observed as result of the exposure to specific toxins from these filaments. Due to differences in growth conditions and health effects both risk assessment and effective management interventions need to be properly calibrated in freshwater and marine recreational sites. The first legislations on HABs are dated to 1995 when the local Galician Government legally established the monitoring network for marine biotoxins in bivalve mollusks grown in rafts in the Galician Rías (NW Spain) (Pazos and Maneiro, 1999). In 1998 the US Congress recognized the threats associated to HABs and hypoxia and established “Harmful Algal Bloom and Hypoxia Research and Control Act”. In Europe several Regulations mandate the control of HABs (Food and Hygiene Regulations [Regulation (EC) 853/2004; Regulation (EC) 2074/2005; Regulation (EU) 2019/627; Regulation (EU) 2021/1709])as well as several Directives including the Water Framework Directive (WFD) (European Commission, 2000), Bathing Water Directive (BWD) (European Commission, 2006) and the European Marine Strategy Framework Directive (MSFD) (European Commission, 2008) have considered the need to seriously evaluate the HABs in the evaluation of water quality. Considering the HABs a serious threat for health, these regulations prescribe the monitoring of marine biotoxins and toxic phytoplankton. Specific thresholds to trigger control measures including beach closures or seafood trade bans have been established (European Food Safety Authority, EFSA, 2009). Recently the algae growth inhibition assay for Skeletonema costatum and Phaeodactylum tricornutum marine has been established by UNI EN ISO 10253:2017. A report dealing with good practice for cyanobacterial and algal bloom detection across Europe has been recently published by Euopean Commission (Sanseverino et al., 2023). According to this report fourteen member states country (Italy is not included) have implemented national monitoring of cyanobacterial/algal blooms in bathing waters in the context of recreational exposure.
Regarding to algaecides, the REACH Regulation (EC 1907/2006), entered into force in 2007, to ensure a high level of protection of human health and the environment from the risks that can be posed by chemicals, the manufacturers and importers of chemicals substances must identify and manage risks linked to the compounds they produce and sale. When one or more tons per year are manufactured or imported, a registration dossier has to be submitted to the European Chemicals Agency (ECHA) that is responsible for the assessment of the effectiveness and risks of each chemical substance. On the ECHA website, a list of the authorized substances is published. The approval of algaecide, their availability and their use throughout Europe is established by the Regulation 528/2012 concerning the making available on the market and use of biocidal products.
8. Conclusions
Phytoplankton or suspended algae play a key role in the food webs in freshwater ecosystems as these organisms are the main source of organic matter but may also represent a risk for human and environmental health. Early detection and widespread monitoring of algal blooms are crucial for effective freshwater resource management and to mitigate negative environmental impacts. Harmful algal blooms, caused by microorganisms/phytoplankton, produce toxins that pose a serious threat to freshwater biodiversity. Scientific research is focused on developing several strategies for predicting, detecting in real-time, and monitoring the spatio-temporal occurrence of algal blooms in freshwater lakes or in different waters for recreational uses. The complex and diverse nature of microalgae necessitates a large amount of data and knowledge from multiple disciplines, including species selection, cultivation parameters, reactor design, and conversion technologies for integrating water treatment and plant engineering. Algae are undesirable an sometimes harmful in swimming pools or basins for several rehabilitative, sportive or ludic activities. However, they are an ubiquitous and fundamental part of the earth ecosystems, supporting life through water and light, since the beginning of life. The impact of Algae in recreational waters has been overhyped or overlooked. It deserves new attention to unravelling the underlying biology and redesigning the real role in public health, within a broader One Health perspective.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org. Table S1.
Author Contributions
Conceptualization, V.R.S.; methodology, F.V., F.C. and G.L.; formal analysis, F.V., F.C., V.V., A.B., M.G.I. and G.L.; data curation, F.V., F.C., V.V., A.B., M.G.I. and G.L.; writing—original draft preparation, F.V., F.C., V.V., A.B., M.G.I., M.G. and G.L.; writing—review and editing, , F.V., F.C., V.V., A.B., M.G.I., M.G. and G.L., and V.R.S.; supervision, G.M., and V.R.S. All authors have read and agreed to the published version of the manuscript.
Funding
This study was partially funded by the MIUR-Fund-PON R&I 2014-2020 React-EU and IUSM Projects [CUP H83C23000160001; Prot. 1007-2023].
Acknowledgments
The authors are grateful to Elena Scaramucci and Fabrizio Michetti for editing the manuscript and Manuela Camerino and Tiziana Zilli for library assistance.
References
- World Health Organization; 2003. Guidelines on recreational water quality: Volume 1 Coastal and fresh waters. WHO, Geneva.
- Summers, E. J.; Ryder, J. L. A critical review of operational strategies for the management of harmful algal blooms (HABs) in inland reservoirs. J. Environ. Manag. 2023, 330, 117141. [Google Scholar] [CrossRef] [PubMed]
- Varga, C. To treat or not to treat? Misbeliefs in spa water disinfection. Int J Biometeorol 2019, 63, 1135–1138. [Google Scholar] [CrossRef] [PubMed]
- Kiliç, S.; Kalkan, E.; Nadaroglu, H. Removal of algae from thermal mud pool: A case study in Koprukoy (Erzurum, Northeast (NE) Turkey) thermal spring area. Bull. Chem. Soc. Ethiop. 2022, 36, 545–553. [Google Scholar] [CrossRef]
- Xiao, F.; Xiao, P.; Wang, D. Influence of allochthonous organic matters on algae removal: Organic removal and floc characteristics. Colloids Surf A Physicochem Eng Asp 2019, 583, 123995. [Google Scholar] [CrossRef]
- Qi, J.; Lan, H.C.; Miao, S.Y.; Xu, Q.; Liu, R.; Liu, H.; Qu, J. KMnO4-Fe(II) pretreatment to enhance Microcystis aeruginosa removal by aluminum coagulation: Does it work after long distance transportation. Water Res 2010, 44, 3617–3624. [Google Scholar] [CrossRef]
- Gilmour, D.J. Diversity of algae and their biotechnological potential. Adv. Microb. Physiol. 2023, 82. [Google Scholar] [CrossRef]
- Cohen, P.A.; Kodner, R.B. The earliest history of eukaryotic life: Uncovering an evolutionary story through the integration of biological and geological data. Trends Ecol. Evol. 2022, 37, 246–256. [Google Scholar] [CrossRef] [PubMed]
- Butterfield, N.J. Bangiomorpha pubescens n. gen., n. sp.: Implication for the evolution of sex, multicellularity and the mesoproterozoic/neoproterozoic radiation of eukaryotes. J. Paleontol. 2011, 26, 1–20. [Google Scholar]
- Brocks, J.J. The transition from a cyanobacterial to alga world and the emergence of animals. Emerg. Top. Life Sci. 2018, 2, 181–190. [Google Scholar] [CrossRef]
- Adl, S. M.; Simpson, A. G.; Lane, C. E.; Lukeš, J.; Bass, D.; Bowser, S. S.; Brown, M. W.; Burki, F.; Dunthorn, M.; Hampl, V.; Heiss, A.; Hoppenrath, M.; Lara, E.; Le Gall, L.; Lynn, D. H.; McManus, H.; Mitchell, E. A.; Mozley-Stanridge, S. E.; Parfrey, L. W.; Pawlowski, J.; Spiegel, F. W. Revis. Classif. Eukaryotes. J. Eukaryot. Microbiol. 2012, 59, 429–493. [CrossRef]
- Bolton, J.J. What is aquatic botany? - And why algae are plants: The importance of non-taxonomic terms for groups of organisms. Aquat. Bot. 2016, 132, 1–4. [Google Scholar] [CrossRef]
- Ponce-Toledo, R.I.; Lopez-Garcia, P.; Moreira, D. Horizontal and endosymbiotic gene transfer in early plastid evolution. New Phytol 2019, 224, 618–624. [Google Scholar] [CrossRef]
- Adl, S. M.; Bass, D.; Lane, C. E.; Lukeš, J.; Schoch, C. L.; Smirnov, A.; Agatha, S.; Berney, C.; Brown, M. W.; Burki, F,; Cárdenas, P. ; Čepička, I.; Chistyakova, L.; Del Campo, J.; Dunthorn, M.; Edvardsen, B.; Eglit, Y.; Guillou, L.; Hampl, V.; Heiss, A. A.; Zhang, Q. Revisions to the classification, nomenclature and diversity of eukaryotes. J. Eukaryot. Microbiol. 2019, 66, 4–119. [Google Scholar] [CrossRef]
- Cavalier-Smith, T. The kingdom chromista: Origin and systematics. Prog. Phycol. Res. Biopress Bristol. 1986, 309–347. [Google Scholar]
- Javed, M.R.; Bilal, M.J.; Mehmood, M.A.; Nashat, N. Microalgae as a Feedstock for Biofuel Production: Current Status and Future Prospects. Energy Res. Dev. 2019, 3. [Google Scholar]
- Enamala, M.K.; Enamala, S.; Chavali, M.; Donepudi, J.; Yadavalli, R.; Kolapalli, B.; Aradhyula, T.V.; Velpuri, J.; Kuppam, C. Production of biofuels from microalgae - A review on cultivation, harvesting, lipid extraction, and numerous applications of microalgae. Renew. Sustain. Energy Rev. 2018, 94. [Google Scholar] [CrossRef]
- Nation Museum of Natural Hystory. The classification. Available online: https://naturalhistory.si.edu/research/botany/research/algae/algae-classification (accessed on 20 December 2023).
- Barsani, L.; and Gualtieri, P. Algae: Anatomy, Biochemistry, and Biotechnology. Third edition. (2023). By CRC Press.
- Fritsch, F.E. The structure and reproduction of algae, vol I. 1935. Cambridge University Press, London.
- Ghosh, A.; Kiran, B. Carbon Concentration in Algae: Reducing CO2 From Exhaust Gas. Trends Biotechnol. 2017, 35, 806–808. [Google Scholar] [CrossRef] [PubMed]
- Ambika, H.D. Positive and negative environmental impacts on algae. Algae Mater. 2023, 343–353. [Google Scholar] [CrossRef]
- World Health Organization; 2021a. Guidelines on Recreational Water Quality: Volume 1 Coastal and Fresh Waters. WHO, Geneva.
- Centers for Disease Control and Prevention, 2023. Available online: https://www.cdc.gov/habs/be-aware-habs.html (accessed on 7 November 2023).
- Dallas, H. Water temperature and riverine ecosystems: An overview of knowledge and approaches for assessing biotic responses, with special reference to South Africa. Water SA 2008, 34, 393–404. [Google Scholar] [CrossRef]
- Sompong, U.; Hawkins, P.R.; Besley, C.; Peerapornpisal, Y. The distribution of cyanobacteria across physical and chemical gradients in hot springs in northern Thailand. FEMS Microbiol Ecol 2005, 52, 365–376. [Google Scholar] [CrossRef]
- Lee, J.; Rai, P.K.; Jeon, Y.J.; Kim, K.H.; Kwon, E.E. (2017). The role of algae and cyanobacteria in the production and release of odorants in water. Environmental pollution (Barking, Essex: 1987) 2017, 227, 252–262. [Google Scholar] [CrossRef] [PubMed]
- Winterbourn, M. The distribution of algae and insects in hot spring thermal gradients at Waimangu, New Zealand. N. Z. J. Mar. Freshw. Res 1969, 3, 459–465. [Google Scholar] [CrossRef]
- Zhang, Y.; Whalen, J.K.; Cai, C.; Shan, K.; Zhou, H. Cyanobacteria-diatom/dinoflagellate blooms and their cyanotoxins in freshwaters: A nonnegligible chronic health and ecological hazard. Water Res 2023, 233, 119807. [Google Scholar] [CrossRef] [PubMed]
- Quiblier, C.; Wood, S.; Echenique-Subiabre, I.; Heath, M.; Villeneuve, A.; Humbert, J.-F. A review of current knowledge on toxic benthic freshwater cyanobacteria e ecology, toxin production and risk management. Water Res 2013, 47, 5464–5479. [Google Scholar] [CrossRef]
- Ibelings, B.W.; Kurmayer, R.; Azevedo, S.M.F.O.; Wood, S.A.; Chorus, I.; Welker, M. Understanding the occurrence of cyanobacteria and cyanotoxins. Toxic Cyanobacteria Water 2021, 213–294. [Google Scholar]
- Centers for Disease Control and Prevention, 2016. Harmful Algal Bloom (HAB) Associated Illness. May 27, 2016. Available online: http://www.cdc.gov/habs/general.html (accessed on 7 November 2023).
- Environmental Protection Agency, 2022. Available online: https://www.epa.gov/cyanohabs/learn-aboutcyanobacteria-and-cyanotoxins (accessed on 13 November 2023).
- Chorus, I. and Welker M. 2021 Toxic Cyanobacteria in Water - A guide to their public health consequences, monitoring and management. 2nd Edition. CRC Press, Boca Raton, FL. on behalf of the World Health Organization, Geneva. Taylor & Francis.
- Chatterjee, S.; More, M. Cyanobacterial Harmful Algal Bloom Toxin Microcystin and Increased Vibrio Occurrence as Climate-Change-Induced Biological Co-Stressors: Exposure and Disease Outcomes via Their Interaction with Gut-Liver-Brain Axis. Toxins (Basel) 2023, 15, 2. [Google Scholar] [CrossRef]
- Olker, J.; Banerji, A.; Benesh, K.; Kinziger, B.; Scott, T.; Karschnik, T.; Frisch, J.; Feist, T.; Pilli, A.; Hoff, D. Evidence Map for Ecological Toxicity of Cyanotoxins using ECOTOXicology Knowledgebase Systematic Protocols. Science Inventory 2023.
- Mehinto, A.C. , Smith, J., Wenger, E., Stanton, B., Linville, R., Brooks, B.W., Sutula, M.A., Howard, M.D.A. Synthesis of ecotoxicological studies on cyanotoxins in freshwater habitats - Evaluating the basis for developing thresholds protective of aquatic life in the United States. Sci Total Env. 2021, 795, 148864. [Google Scholar] [CrossRef]
- Shahmohamadloo R., S. , Poirier D. G., Almirall X. O., Bhavsar S. P., Sibley P. K. Assessing the toxicity of cell-bound microcystins on freshwater pelagic and benthic invertebrates. Ecotoxicol. Environ. Saf. 2020, 188, 109945. [Google Scholar] [CrossRef]
- Smutná, M. , Babica P., Jarque S., Hilscherová K., Maršálek B., Haeba M., Bláha L. Acute, chronic and reproductive toxicity of complex cyanobacterial blooms in Daphnia magna and the role of microcystins. Toxicon 2014, 79, 11–18. [Google Scholar] [CrossRef]
- Davis, T.W.; Koch, F.; Marcoval, M.A.; Wilhelm, S.W.; Gobler, C.J. Mesozooplankton and microzooplankton grazing during cyanobacterial blooms in the western basin of Lake Erie. Harmful Algae 2020, 15, 26–35. [Google Scholar] [CrossRef]
- Palíková, M.; Krejcí, R.; Hilscherová, K.; Babica, P.; Navrátil, S.; Kopp, R.; Bláha, L. Effect of different cyanobacterial biomasses and their fractions with variable microcystin content on embryonal development of carp (Cyprinus carpio L.). Aquat Toxicol. 2007, 81, 312–318. [Google Scholar] [CrossRef] [PubMed]
- Falfushynska, H.; Kasianchuk, N.; Siemens, E.; Henao, E.; Rzymski, P. A Review of Common Cyanotoxins and Their Effects on Fish. Toxics 2023, 11, 118. [Google Scholar] [CrossRef]
- Ferrão-Filho, A.D.S.; Kozlowsky-Suzuki, B. Cyanotoxins: Bioaccumulation and effects on aquatic animals. Mar. Drugs 2011, 9, 2729–2772. [Google Scholar] [CrossRef]
- Wu, J.L.; Liu, W.X.; Wen, C.G.; Qian, G.M.; Hu, B.Q.; Jian, S.Q.; Yang, G.; Dong, J. Effect of microcystin on the expression of Nrf2 and its downstream antioxidant genes from Cristaria plicata. Aquat. Toxicol. 2020, 225, 105526. [Google Scholar] [CrossRef] [PubMed]
- Lundqvist, J.; Pekar, H.; Oskarsson, A. Microcystins activate nuclear factor erythroid 2-related factor 2 (Nrf2) in human liver cells in vitro—Implications for an oxidative stress induction by microcystins. Toxicon 2017, 126, 47–50. [Google Scholar] [CrossRef]
- Shaw, G.R.; Seawright, A.A.; Moore, M.R.; Lam, P.K.S. Cylindrospermopsin, a cyanobacterial alkaloid: Evaluation of its toxicological activity. Ther. Drug Monit. 2000, 22, 89–92. [Google Scholar] [CrossRef]
- Shu, Y.; Jiang, H.; Yuen, C.N.T.; Wang, W.; He, J.; Zhang, H.; Liu, G.; Wei, L.; Chen, L.; Wu, H. Microcystin-leucine arginine induces skin barrier damage and reduces resistance to pathogenic bacteria in Lithobates catesbeianus tadpoles. Ecotoxicol. Environ. Saf. 2022, 238, 113584. [Google Scholar] [CrossRef]
- Xia, H.; Song, T.; Wang, L.; Jiang, L.; Zhou, Q.; Wang, W.; Liu, L.; Yang, P.; Zhang, X. Effects of dietary toxic cyanobacteria and ammonia exposure on immune function of blunt snout bream (Megalabrama amblycephala). Fish Shellfish Immun. 2018, 78, 383–391. [Google Scholar] [CrossRef]
- Qiao, Q.; Liang, H.; Zhang, X. Effect of cyanobacteria on immune function of crucian carp (Carassius auratus) via chronic exposure in diet. Chemosphere 2013, 90, 1167–1176. [Google Scholar] [CrossRef]
- Patiño, R.; Christensen, V.G.; Graham, J.L.; Rogosch, J.S.; Rosen, B.H. Toxic Algae in Inland Waters of the Conterminous United States—A Review and Synthesis. Water 2023, 15, 2808. [Google Scholar] [CrossRef]
- Rattner, B.A.; Wazniak, C.E.; Lankton, J.S.; McGowan, P.C.; Drovetski, S.V.; Egerton, T.A. Review of harmful algal bloom effects on birds with implications for avian wildlife in the Chesapeake Bay region. Harmful Algae. 2022, 120, 102319. [Google Scholar] [CrossRef]
- EU Council, 2020. Available online: https://eur-lex.europa.eu/legal-content/EN/TXT/PDF/?uri=CELEX:32020L2184&from=ES (accessed on 14 November 2023).
- Li, J.; Parkefelt, L.; Persson, K. M.; Pekar, H. Improving cyanobacteria and cyanotoxin monitoring in surface waters for drinking water supply. J. Water Secur. 2017, 3. [Google Scholar] [CrossRef]
- Sanseverino, I.; Conduto, D.; Pozzoli, L.; Dobricic, S. , Lettieri, T. Algal bloom and its economic impact. European Commission, JRC Technical Reports. 2016.
- World Health Organization, 2017. Guidelines for Drinking-Water Quality: Fourth Edition Incorporating the First Addendum. World Health Organization, Geneva.
- IARC, 2010. Ingested Nitrate and Nitrite, and Cyanobacterial Peptide Toxins. World Health Organization; International Agency for Research on Cancer; Lyon, France: 2010.
- Hurtado, I.; Pouget, L.; Fernández, S.; Cascales, P. Monitoring and forecasting cyanobacteria risk for a drinking water plant in Spain. Water Supply 2022, 22(7), 6296–6307. [Google Scholar] [CrossRef]
- Roberts, V.A.; Vigar, M.; Backer, L.; et al. Surveillance for Harmful Algal Bloom Events and Associated Human and Animal Illnesses — One Health Harmful Algal Bloom System, United States, 2016–2018. MMWR Morb Mortal Wkly Rep 2020, 69, 1889–1894. [Google Scholar] [CrossRef]
- Centers for Disease Control and Prevention (CDC), 2023. Summary Report – One Health Harmful Algal Bloom System (OHHABS), United States, 2021. Atlanta, Georgia: U.S. Department of Health and Human Services, CDC, 2023. Available online: https://www.cdc.gov/habs/data/2021-ohhabs-data-summary.html (accessed on 11 November 2023).
- Harmful Algae Event Database, 2023. Available online: http://haedat.iode.org/ (accessed on 13 November 2023).
- Tito, J.C.R.; Luna, L.M.G.; Noppe, W.N.; Hubert, I.A. First Report on Microcystin-LR Occurrence in Water Reservoirs of Eastern Cuba, and Environmental Trigger Factors. Toxins 2022, 14, 209. [Google Scholar] [CrossRef]
- Oliveira, E.D.C.; Castelo-Branco, R.; Silva, L.; Silva, N.; Azevedo, J.; Vasconcelos, V.; Faustino, S.; Cunha, A. First Detection of Microcystin-LR in the Amazon River at the Drinking Water Treatment Plant of the Municipality of Macapa, Brazil. Toxins 2019, 11, 669. [Google Scholar] [CrossRef]
- Chatziefthimiou, A.D.; Metcalf, J.S.; Glover, W.B.; Banack, S.A. , Dargham, S.R., Richer, R.A. Cyanobacteria and cyanotoxins are present in drinking water impoundments and groundwater wells in desert environments. Toxicon 2016, 114, 75–84. [Google Scholar] [CrossRef] [PubMed]
- Douma, M.; Ouahid, Y.; del Campo, F.F.; Loudiki, M.; Mouhri, K.; Oudra, B. Identification and quantification of cyanobacterial toxins (microcystins) in two Moroccan drinking-water reservoirs (Mansour Eddahbi, Almassira). Environ. Monit. Assess. 2010, 160, 439–450. [Google Scholar] [CrossRef] [PubMed]
- Bruno, M.; Gucci, P.M.B.; Pierdominici, E.; Sestili, P.; Joppolo, A.; Volterra, L. Anatoxin-a and previously unknown toxin in Anabaena planctonica from blooms found in Lake Mulargia (Italy). Toxicon 1994, 32, 369–73. [Google Scholar] [CrossRef] [PubMed]
- Bruno, M.; Gucci, P.M.B.; Pierdominici, E.; Sestili, P.; Joppolo, A.; Volterra, L. Production of microcystinlike toxins in different freshwater species of Oscillatoria. Toxicon 1992, 32, 1307–11. [Google Scholar] [CrossRef] [PubMed]
- Messineo V; Bogialli S; Melchiorre S; Sechi N; Luglié A; Casiddu P; Mariani MA; Padedda BM; Di Corcia A; Mazza R; Carloni E; Bruno M. Cyanobacterial toxins in Italian freshwaters. Limnologica 2009, 39, 95–106. [Google Scholar] [CrossRef]
- Istituto Superiore di Sanità, 2011. Rapporti ISTISAN 11/35 Pt.1. Cyanobacteria in water for human consumption. State of knowledge for risk assessment. Volume 1. Edited by Luca Lucentini and Massimo Ottaviani for “National Group for cyanobacteria risk management in water for human consumption” 2011, xxii, 165 p.
- S-3 EUROHAB Project, 2023. Sentinel-3 satellite products for detecting Eutrophication and Harmful Algal Bloom events in the French-English Channel. European Union. Official project website Available to: https://www.s3eurohab.eu/ (accessed on 14 November 2023). (accessed on 14 November 2023).
- Boopathi, T. and J.S. Ki, Impact of environmental factors on the regulation of cyanotoxin production. Toxins (Basel), 2014, 6, 1951–78. [Google Scholar] [CrossRef]
- Pearson, L. , et al., On the chemistry, toxicology and genetics of the cyanobacterial toxins, microcystin, nodularin, saxitoxin and cylindrospermopsin. Mar Drugs 2010, 8, 1650–80. [Google Scholar] [CrossRef]
- Weirich, C.A. and T.R. Miller. Freshwater harmful algal blooms: toxins and children's health. Curr Probl Pediatr Adolesc Health Care 2014, 44, 2–24. [Google Scholar] [CrossRef] [PubMed]
- Humpage, A.R. and I.R. Falconer, Oral toxicity of the cyanobacterial toxin cylindrospermopsin in male Swiss albino mice: determination of no observed adverse effect level for deriving a drinking water guideline value. Env. Toxicol 2003, 18, 94–103. [Google Scholar] [CrossRef]
- Astrachan, N.B.; Archer, B.G.; Hilbelink, D.R. Evaluation of the subacute toxicity and teratogenicity of anatoxin-a. Toxicon 1980, 18, 684-8. [Google Scholar] [CrossRef]
- Bumke-Vogt, C.; Mailahn, W.; Chorus, I. Anatoxin-a and neurotoxic cyanobacteria in German lakes and reservoirs. Environ. Toxicol. 1999, 14, 117–125. [Google Scholar] [CrossRef]
- Van Apeldoorn, M.E. , et al. Toxins of cyanobacteria. Mol Nutr Food Res 2007, 51, 7–60. [Google Scholar] [CrossRef]
- Strichartz, G. Structural determinants of the affinity of saxitoxin for neuronal sodium channels. Electrophysiological studies on frog peripheral nerve. J Gen Physiol 1984, 84, 281–305. [Google Scholar] [CrossRef]
- Holtcamp, W. The emerging science of BMAA: do cyanobacteria contribute to neurodegenerative disease? Env. Health Perspect 2012, 120, A110–6. [Google Scholar] [CrossRef] [PubMed]
- Jiang, L. , et al., Quantification of neurotoxin BMAA (beta-N-methylamino-Lalanine) in seafood from Swedish markets. Sci Rep 2014, 4, 6931. [Google Scholar] [CrossRef] [PubMed]
- Torokne, A.; Palovics, A. ; M. Bankine. Allergenic (sensitization, skin and eye irritation) effects of freshwater cyanobacteria--experimental evidence. Env. Toxicol 2001, 16, 512-6. [Google Scholar] [CrossRef] [PubMed]
- Blahova, L. , et al., The isolation and characterization of lipopolysaccharides from Microcystis aeruginosa, a prominent toxic water bloom forming cyanobacteria. Toxicon 2013, 76, 187–96. [Google Scholar] [CrossRef] [PubMed]
- Osborne, N.J.T.; Webb, P.M.; Shaw, G.R. The toxins of Lyngbya majuscule and their human and ecological health effects. Environ. Int. 2001, 27, 381–392. [Google Scholar] [CrossRef] [PubMed]
- Arthur, K. , et al., The exposure of green turtles (Chelonia mydas) to tumour promoting compounds produced by the cyanobacterium Lyngbya majuscula and their potential role in the aetiology of fibropapillomatosis. Harmful Algae 2008, 7, 114–125. [Google Scholar] [CrossRef]
- Churro, C.; Dias, E.; Valrio, E. Risk Assessment of Cyanobacteria and Cyanotoxins, the Particularities and Challenges of Planktothrix spp. Monitoring. 2012.
- Gere, D.; Róka, E.; Záray, G.; Vargha, M. Disinfection of Therapeutic Spa Waters: Applicability of Sodium Hypochlorite and Hydrogen Peroxide-Based Disinfectants. Water 2022, 14, 690. [Google Scholar] [CrossRef]
- World Health Organization. 2013. WHO traditional medicine strategy: 2014-2023. World Health Organization. ISBN: 9789241506090.
- Balegová, J. The mineral spa of Rudnok as described by staff colonel Henrik Mayer. Comm. Hist. Art. Med. 2008, 54, 87–92. [Google Scholar]
- World Health Organization, 2006. Available online: http://apps.who.int/iris/bitstream/10665/43336/1/9241546808_eng.pdf (accessed on 6 November 2023).
- Health and Safety Executive, 2014. Available online: www.hse.gov.uk/pubns/books/hsg282.htm (accessed on 6 November 2023).
- EU Council, 2020. Directive (EU) 2020/2184 of the European Parliament and of the Council of 16 December 2020 on the quality of water intended for human consumption. Official Journal of the European Union. Available online: https://eur-lex.europa.eu/legal-content/EN/TXT/PDF/?uri=CELEX:32020L2184&from=ES. (accessed on November 7th, 2023).
- Ministero della Salute. 2012. Decreto 7 febbraio 2012, n. 25 Disposizioni tecniche concernenti apparecchiature finalizzate al trattamento dell'acqua destinata al consumo umano. (12G0044) (GU Serie Generale n.69 del 22-03-2012) Available to: https://shorturl.at/gDM35. (Accessed on November 5th, 2023). 5 November.
- Regione Toscana, 2004. Legge Regionale 27 luglio 2004, n. 38 Norme per la disciplina della ricerca, della coltivazione e dell'utilizzazione delle acque minerali, di sorgente e termali. (GU 3a Serie Speciale Regioni n.45 del 13/11/2004) Available to: https://www.edizionieuropee.it/law/html/208/to4_04_012.html. (Accessed on November 5th, 2023). 5 November 2023.
- Regione Campania, 2008. Legge Regionale n. 8 del 29 luglio 2008. “Disciplina della ricerca ed utilizzazione delle acque minerali e termali, delle risorse geotermiche e delle acque di sorgente”. Available to: https://www.regione.campania.it/normativa/userFile/documents/attachments/1953_8-2008Storico.pdf (Accessed on November 5th, 2023). 5 November.
- Sevillano, D.; Romero-Lasta, C.I.; Alou, L.; Gonzáles, N. Impact of the biotic and abiotic components of low mineralized natural mineral waters on the growth of pathogenic bacteria of human origin: a key to self-control of spa water quality. J. Hydrol 2018, 566, 227–234. [Google Scholar] [CrossRef]
- Margarucci, L. M.; Romano Spica, V.; Gianfranceschi, G.; Valeriani, F. Untouchability of natural spa waters: Perspectives for treatments within a personalized water safety plan. Environ. Int. 2019, 133(Pt A), 105095. [Google Scholar] [CrossRef]
- Valeriani, F.; Gianfranceschi, G.; Romano Spica, V. The microbiota as a candidate biomarker for SPA pools and SPA thermal spring stability after seismic events. Environ. Int. 2020, 137, 105595. [Google Scholar] [CrossRef] [PubMed]
- Giampaoli, S.; Valeriani, F.; Gianfranceschi, G.; Vitali, M.; Delfini, M.; Festa, M.; Bottari, E.; Romano Spica, V. Hydrogen sulfide in thermal spring waters and its action on bacteria of human origin. Microchem. J. 2013, 108, 210–214. [Google Scholar] [CrossRef]
- Vachee, A.; Vincent, P.; Struijk, C.B.; Mossel, D.A.A.; Leclerc, H. Study of the fate of the autochtonous bacterial flora of still mineral waters by analysis of restriction fragment length polymorphism of genes coding for rRNA. Syst. Appl. Microbiol. 1997, 20, 492–503. [Google Scholar] [CrossRef]
- Lu, J.; Struewing, I.; Vereen, E.; Kirby, A.; Levy, K.; Moe, C.; Ashbolt, N. Molecular detection of Legionella spp. and their associations with Mycobacterium spp., Pseudomonas aeruginosa and amoeba hosts in a drinking water distribution system. J Appl Microbiol 2016, 120, 509–521. [Google Scholar] [CrossRef]
- Giorgio, A.; Carraturo, F.; Aliberti, F.; De Bonis, S.; Libralato, G.; Morra, M.; Guida, M. Characterization of microflora composition and antimicrobial activity of algal extracts from Italian thermal muds. J. Nat. Sci. Biol. Med. 2018, 9. [Google Scholar]
- Kalkan, E. Algae potential of the Delicermik thermal spring area (Köprüköy-Erzurum, NE Turkey). Int. J. Lat. Technol. Eng. Manag. Appl. Sci. 2019, 8, 98–101. [Google Scholar]
- Pabuccu, K. A research on Köprüköy-Deliçermik algal flora. Master Thesis, Graduate School of Natural and Applied Sciences, Atatürk University, Turkey, 1993.
- Gerwick, L; Gerwick, W.H.; Coates, R.C.; Engene, N.; Grindberg, R.V.; Jones A.C., et al.; et al. Giant marine cyanobacteria produce exciting potential pharmaceuticals. Microbe 2008, 3, 277–284. [Google Scholar] [CrossRef]
- Sivonen, K; Börner, T. Bioactive compounds produced by cyanobacteria. In: The Cyanobacteria: Molecular Biology, Genomics and Evolution. Norfolk: Caister Acad. Press 2008, 15997.
- Costa, M.; Garcia, M.; Costa Rodrigues, J.; Costa, M.S.; Ribeiro, M.J.; Fernandes, M.H.; et al. Exploring bioactive properties of marine cyanobacteria isolated from the Portuguese coast: High potential as a source of anticancer compounds. Mar Drugs 2013, 12, 98–114. [Google Scholar] [CrossRef] [PubMed]
- Surakka, A. , Sihvonen, L.M., Lehtimäki, J.M., Wahlsten, M., Vuorela, P., Sivonen, K., et al. Benthic cyanobacteria from the Baltic sea contain cytotoxic Anabaena, Nodularia, and Nostoc strains and a apoptosis inducing phormidium strain. Env. Toxicol 2005, 20, 285–92. [Google Scholar] [CrossRef] [PubMed]
- Jaki, B.; Orjala, J.; Heilmann, J.; Linden, A.; Vogler, B.; Sticher, O.; et al. Novel extracellular diterpenoids with biological activity from the Cyanobacterium nostoc commune. J Nat Prod 2000, 63, 339–43. [Google Scholar] [CrossRef] [PubMed]
- Kralovec, J.A.; Metera, K.L.; Kumar, J.R.; Watson, L.V.; Girouard, G.S.; Guan, Y., et al.; et al. Immunostimulatory principles from Chlorella pyrenoidosa – Part 1: Isolation and biological assessment in vitro. Phytomedicine 2007, 14, 57–64. [Google Scholar] [CrossRef] [PubMed]
- Komatsu, T.; Kido, N. ; Sugiyama, T; Yokochi, T. Antiviral activity of acidic polysaccharides from Coccomyxa gloeobotrydiformi, a green alga, against an in vitro human influenza A virus infection. Immunopharmacol Immunotoxicol 2013, 35, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Kust, A.; Urajová, P.; Hrouzek, P.; Čapková, K.; Štenclová, L.; Řeháková, K.; Kozlíková-Zapomělová, E.; Lepšová-Skácelová, O.; Lukešová, A.; Mareš, J. ; A new microcystin producing Nostoc strain discovered in broad toxicological screening of non-planktic Nostocaceae (cyanobacteria). Toxicon 2018, 150, 66–73. [Google Scholar] [CrossRef] [PubMed]
- Sanchez, J.A.; Otero, P.; Alfonso, A.; Ramos, V.; Vasconcelos, V.; Aráoz, R.; Botana, L.M. Detection of anatoxin-a and three analogs in Anabaena spp. cultures: new fluorescence polarization assay and toxin profile by LC-MS/MS. Toxins 2014, 6, 402–415. [Google Scholar] [CrossRef]
- Stoyneva, M. Survey on green algae of Bulgarian thermal springs. Biol. Bratisl. - Sect. Bot. 2003, 58, 563–574. [Google Scholar]
- Lai, G.G.; Wetzel, C.E.; Ector, L.; Lugliè, A.; Padedda, B.M. Composition, structure, and distribution of diatom assemblages in Mediterranean thermal spring ecotones affected by natural and human disturbances. Aquat Sci 2023, 85, 55. [Google Scholar] [CrossRef]
- Ulcay, S.; Öztürk, M.; Kurt, O. Algae Flora of Germencik-Alangüllü (Aydın, Turkey) Thermal Water. Celal Bayar Univ. J. Sci. 2017, 13, 601–608. [Google Scholar]
- Komárek, J.; Anagnostidis, K. Süßwasserflora von Mitteleuropa. Cyanoprokaryota; 2nd Part: Oscil. 2005, 19, 759.
- Veen, A.; Hof, C.H.J.; Kouwets, F.A.C.; Berkhout, T. Rijkswaterstaat Waterdienst, Informatiehuis Water [Taxa Watermanagement the Netherlands (TWN)] Available to: http://ipt.nlbif.nl/ipt/resource?r=checklist-twn. (Accessed November 7th, 2023). 7 November.
- Anagnostidis, K. Untersuchungen über die Cyanophyceen einiger Thermen in Griechenland. Institute Systematic Botany & Pflanzengeorge University. Thessalon. 1961, 7, 1–322.
- Shanab, M.M.A. Algal flora of Ain Helwan I. Algae of the Worm Spring. Egypt. J. Phycol. 2006, 7, 209–231. [Google Scholar] [CrossRef]
- Gupta, R. K. Mini review: Thermal algae of India with special reference to Jharkhand. Biospectra 2017, 12, 15–20. [Google Scholar]
- Varga, C. Water hygiene and toxicology: actual problems and new research trends in Hungary. Acta Biol Debr Oecol Hung 2012, 29, 9–120. [Google Scholar]
- Antunes, J.T.; Leao, P.N.; Vasconcelos, V.M. Cylindrospermopsis raciborskii: review of the distribution, phylogeography, and ecophysiology of a global invasive species. Front Microbiol 2015, 6, 473. [Google Scholar] [CrossRef]
- Simon, N.; Cras, A. L.; Foulon, E.; Lemée, R. Diversity and evolution of marine phytoplankton. Comptes rendus biologies 2009, 332, 159–170. [Google Scholar] [CrossRef]
- Poniedziałek, B.; Rzymski, P.; Wiktorowicz, K. Experimental immunologyFirst report of cylindrospermopsin effect on human peripheral blood lymphocytes proliferation in vitro. Cent. Eur. J. Immunol. 2012, 37, 314–317. [Google Scholar] [CrossRef]
- Kouakou CRC, Poder TG. Economic impact of harmful algal blooms on human health: a systematic review. J Water Health 2019, 17, 499–516. [Google Scholar] [CrossRef]
- Moore, S. K.; Trainer, V. L.; Mantua, N. J.; Parker, M. S.; Laws, E. A.; Backer, L.C.; Fleming, L.E. . Impacts of climate variability and future climate change on harmful algal blooms and human health. Environmental health : a global access science source 2008, 7, S4. [Google Scholar] [CrossRef] [PubMed]
- Dietrich, D.; Hoeger, S. Guidance values for microcystins in water and cyanobacterial supplement products (blue-green algal supplements): a reasonable or misguided approach? Toxicol. Appl. Pharmacol. 2005, 203, 273–289. [Google Scholar] [CrossRef]
- Drobac, D.; Tokodi, N.; Simeunović, J.; Baltić, V.; Stanić, D.; Svirčev, Z. Human exposure to cyanotoxins and their effects on health. Arh. Za Hig. Rada I Toksikol. 2013, 64, 119–130. [Google Scholar] [CrossRef]
- Ananya; Kamal, A.; Iffat, A. Cyanobacteria “the blue green algae” and its novel applications: A brief review. International J. Innov. Appl. Stud. 2014, 7, 251–261. [Google Scholar]
- Roy-Lachapelle, A.; Solliec, M.; Bouchard, M.F.; Sauvé, S. Detection of Cyanotoxins in Algae Dietary Supplements. Toxins 2017, 9, 76. [Google Scholar] [CrossRef]
- Codd, G.A.; Steffensen, D.A. Toxic blooms of cyanobacteria in Lake Alexandrina, South Australia - Learning from history. Mar. Freshw. Res. 1994, 45. [Google Scholar] [CrossRef]
- Dillenberg, H.O.; Dehnel, M.K. Toxic waterbloom in Saskatchewan, 1959. Can Med Assoc J 1960, 83, 1151–4. [Google Scholar] [PubMed]
- Griffiths, D.J.; Saker, M.L. The Palm Island mystery disease 20 years on: a review of research on the cyanotoxin cylindrospermopsin. Env. Toxicol 2003, 18, 78–93. [Google Scholar] [CrossRef] [PubMed]
- Hawkins, P.R.; Runnegar, M.T. Severe hepatotoxicity caused by the tropical cyanobacterium (blue-green alga) Cylindrospermopsis raciborskii (Woloszynska) Seenaya and Subba Raju isolated from a domestic water supply reservoir. Appl Env. Microbiol 1985, 50, 1292–5. [Google Scholar] [CrossRef] [PubMed]
- Jeffery, B.; Barlow, T. Amnesic shellfish poison. Food Chem. Toxicol. 2004, 42, 545–557. [Google Scholar] [CrossRef]
- Teixeira, M. da G.; Costa, M. da C. Gastroenteritis epidemic in the area of the Itaparica Dam, Bahia, Brazil. Bull Pan Am Health Organ 1993, 27, 244-53. [Google Scholar]
- Azevedo, S.M.; Carmichael, W.W. Human intoxication by microcystins during renal dialysis treatment in Caruaru-Brazil. Toxicology 2002, 441-6, 181–182. [Google Scholar] [CrossRef]
- Backer, L.C.; Fleming, L.E. Recreational exposure to aerosolized brevetoxins during Florida red tide events. Harmful Algae 2003, 2, 19–28. [Google Scholar] [CrossRef]
- Stewart, I.; Webb, P.M. Epidemiology of recreational exposure to freshwater cyanobacteria – an international prospective cohort study. BMC Public Health 2006, 6, 93. [Google Scholar] [CrossRef] [PubMed]
- Bean, J.A.; Fleming, L.E. Florida Red Tide Toxins (Brevetoxins) and Longitudinal Respiratory Effects in Asthmatics. Harmful Algae 2011, 10, 744–748. [Google Scholar] [CrossRef]
- Kirkpatrick, B.; Fleming, L.E. Aerosolized red tide toxins (brevetoxins) and asthma: Continued health effects after 1 hour beach exposure. Harmful Algae 2011, 10, 138–143. [Google Scholar] [CrossRef] [PubMed]
- Aifeng, L.; Jinggang, M. Toxins in mussels (Mytilus galloprovincialis) associated with diarrhetic shellfish poisoning episodes in China. Toxicon 2012, 60, 420–425. [Google Scholar]
- Giannuzzi, L.; Sedan, D. An acute case of intoxication with cyanobacteria and cyanotoxins in recreational water in Salto Grande Dam, Argentina. Mar. Drugs 2011, 9, 2164–2175. [Google Scholar] [CrossRef] [PubMed]
- Lévesque, B.; Gervais, M.C. Prospective study of acute health effects in relation to exposure to cyanobacteria. Sci Total Env. 2014, 1, 397–403. [Google Scholar] [CrossRef]
- Trevino-Garrison, I.; DeMent, J. Human illnesses and animal deaths associated with freshwater harmful algal blooms-Kansas. Toxins 2015, 7, 353–66. [Google Scholar] [CrossRef] [PubMed]
- Berdalet, E.; Pavaux, A. Environmental, human health and socioeconomic impacts of Ostreopsis spp. Blooms in the NW Mediterranean. Blooms in the NW Mediterranean. Harmful Algae 2022, 119. [Google Scholar] [CrossRef]
- Veldee, M.V. ; An Epidemiological Study of Suspected Water-borne Gastroenteritis. Am J Public Health Nations Health 1931, 21, 1227–35. [Google Scholar] [CrossRef]
- Backer, L.C.; Kirkpatrick, B. Occupational exposure to aerosolized brevetoxins during Florida red tide events: effects on a healthy worker population. Env. Health Perspect 2005, 113, 644–9. [Google Scholar] [CrossRef]
- Jiye, L.; So-Young, W. Dynamic calibration of phytoplankton blooms using the modified SWAT model. J. Clean. Prod. 2022, 343. [Google Scholar]
- Figgatt, M.; Muscatiello, N. Harmful Algal Bloom-Associated Illness Surveillance: Lessons From Reported Hospital Visits in New York, 2008-2014. Am J Public Health 2016, 106, 440–2. [Google Scholar] [CrossRef]
- Wu, J.; Hilborn, E.D. Acute health effects associated with satellite-determined cyanobacterial blooms in a drinking water source in Massachusetts. Env. Health 2021, 20, 83. [Google Scholar] [CrossRef] [PubMed]
- Hilborn, E.D.; Roberts, V.A. Centers for Disease Control and Prevention (CDC). Algal bloom-associated disease outbreaks among users of freshwater lakes--United States, 2009-2010. MMWR Morb Mortal Wkly Rep 2014, 63, 11-5. [Google Scholar] [PubMed]
- Roberts, V.A.; Vigar, M. Surveillance for Harmful Algal Bloom Events and Associated Human and Animal Illnesses - One Health Harmful Algal Bloom System, United States, 2016-2018. MMWR Morb Mortal Wkly Rep 2020, 69, 1889–1894. [Google Scholar] [CrossRef] [PubMed]
- Backer, L.C.; Carmichael, W. Recreational exposure to low concentrations of microcystins during an algal bloom in a small lake. Mar Drugs 2008, 6, 389–406. [Google Scholar] [CrossRef] [PubMed]
- Backer, L.C.; McNeel, S.V. Recreational exposure to microcystins during algal blooms in two California lakes. Toxicon 2010, 55, 909–921. [Google Scholar] [CrossRef]
- Stumpf, R.P.; Li, Y. Quantifying Karenia brevis bloom severity and respiratory irritation impact along the shoreline of Southwest Florida. PLoS One 2022, 17. [Google Scholar] [CrossRef] [PubMed]
- Diaz, R.E.; Friedman, M.A. Neurological illnesses associated with Florida red tide (Karenia brevis) blooms. Harmful Algae 2019, 82, 73–81. [Google Scholar] [CrossRef]
- Tichadou, L.; Glaizal, M.; Armengaud, A.; Grossel, H.; Lemée, R.; Kantin, R.; Lasalle, J.L.; Drouet, G.; Rambaud, L.; Malfait, P.; de Haro, L. Health impact of unicellular algae of the Ostreopsis genus blooms in the Mediterranean Sea: experience of the French Mediterranean coast surveillance network from 2006 to 2009. Clin Toxicol 2010, 48, 839–44. [Google Scholar] [CrossRef]
- Lavery, A.; Backer, L.; Daniel, J. Evaluation of Electronic Health Records to Monitor Illness From Harmful Algal Bloom Exposure in the United States. J Env. Health 2021, 839, 8–14. [Google Scholar]
- Abdullah, L.; Ferguson, S.; Niedospial, D.; Patterson, D.; Oberlin, S.; Nkiliza, A.; Bartenfelder, G.; Hahn-Townsend, C.; Parks, M.; Crawford, F.; Reich, A.; Keegan, A.; Kirkpatrick, B.; Mullan, M. Exposure-response relationship between K. brevis blooms and reporting of upper respiratory and neurotoxin-associated symptoms. Harmful Algae 2022, 117. [Google Scholar] [CrossRef]
- Mchau, G.J.; Makule, E.; Machunda, R.; Gong, Y.Y.; Kimanya, M. Harmful algal bloom and associated health risks among users of Lake Victoria freshwater: Ukerewe Island, Tanzania. J. Water Health 2019, 17, 826–836. [Google Scholar] [CrossRef]
- Vila, M.; Abós-Herràndiz, R.; Isern-Fontanet, J.; Alvarez, J.; Berdalet, E. Establishing the link between Ostreopsis cf. Ovata blooms and human health impacts using ecology and epidemiology. Ovata blooms and human health impacts using ecology and epidemiology. Sci. Mar. 2016, 80, 107–115. [Google Scholar] [CrossRef]
- Lavery, A.M.; Backer, L.C.; Roberts, V.A.; DeVies, J.; Daniel, J. ; (). Evaluation of Syndromic Surveillance Data for Studying Harmful Algal Bloom-Associated Illnesses - United States, 2017-2019. MMWR. Morb. Mortal. Wkly. Rep. 2021, 70, 1191–1194. [Google Scholar] [CrossRef]
- Illoul, H.; Hernández, F.R.; Vila, M.; Adjas, N.; Younes, A.A.; Bournissa, M.; Koroghli, A.; Marouf, N.; Rabia, S.; Ameur, F.L.K. The genus Ostreopsis along the Algerian coastal waters (SW Mediterranean Sea) associated with a human respiratory intoxication episode. Cryptogam. Algol. 2012, 2, 209–216. [Google Scholar] [CrossRef]
- Figgatt, M.; Hyde, J.; Dziewulski, D.; Wiegert, E.; Kishbaugh, S.; Zelin, G.; Wilson, L. Harmful Algal Bloom-Associated Illnesses in Humans and Dogs Identified Through a Pilot Surveillance System - New York, 2015. MMWR. Morb. Mortal. Wkly. Rep. 2017, 66, 1182–1184. [Google Scholar] [CrossRef]
- Fleming, L.E.; Kirkpatrick, B.; Backer, L.C.; Bean, J.A.; Wanner, A.; Dalpra, D.; Tamer, R.; Zaias, J.; Cheng, Y.S.; Pierce, R.; Naar, J.; Abraham, W. Clark, R.; Zhou, Y.; Henry, M.S.; Johnson, D.; Van De Bogart, G.; Bossart, G.D.; Harrington, M.; Baden, D.G. Initial evaluation of the effects of aerosolized Florida red tide toxins (brevetoxins) in persons with asthma. Env. Health Perspect 2005, 113, 650-7. [Google Scholar] [CrossRef] [PubMed]
- Fleming, L.E.; Bean, J.A.; Kirkpatrick, B.; Cheng, Y.S.; Pierce, R.; Naar, J.; Nierenberg, K.; Backer, L.C.; Wanner, A.; Reich, A.; Zhou, Y.; Watkins, S.; Henry, M.; Zaias, J.; Abraham, W.M.; Benson, J.; Cassedy, A.; Hollenbeck, J.; Kirkpatrick, G.; Clarke, T.; Baden, D.G. Exposure and effect assessment of aerosolized red tide toxins (brevetoxins) and asthma. Environ. Health Perspect. 2009, 117, 1095–1100. [Google Scholar] [CrossRef] [PubMed]
- Hoagland, P.; Jin, D.; Beet, A.; Kirkpatrick, B.; Reich, A.; Ullmann, S.; Fleming, L.E.; Kirkpatrick, G. The human health effects of Florida red tide (FRT) blooms: an expanded analysis. Environ. Int. 2014, 68, 144–153. [Google Scholar] [CrossRef] [PubMed]
- Hoagland, P.; Jin, D.; Polansky, L.Y.; Kirkpatrick, B.; Kirkpatrick, G.; Fleming, L.E.; Reich, A.; Watkins, S.M.; Ullmann, S.G.; Backer, L.C. The costs of respiratory illnesses arising from Florida gulf coast Karenia brevis blooms. Environ. Health Perspect. 2009, 117, 1239–1243. [Google Scholar] [CrossRef] [PubMed]
- Kirkpatrick, B.; Fleming, L.E.; Backer, L.C.; Bean, J.A.; Tamer, R.; Kirkpatrick, G.; Kane, T.; Wanner, A.; Dalpra, D.; Reich, A.; Baden, D.G. Environmental exposures to Florida red tides: Effects on emergency room respiratory diagnoses admissions. Harmful Algae 2006, 5, 526–533. [Google Scholar] [CrossRef]
- Chomérat, N.; Antajan, E.; Auby, I.; Bilien, G.; Carpentier, L.; Casamajor, M.-N.d.; Ganthy, F.; Hervé, F.; Labadie, M.; Méteigner, C.; et al. First Characterization of Ostreopsis cf. ovata (Dinophyceae) and Detection of Ovatoxins during a Multispecific and Toxic Ostreopsis Bloom on French Atlantic Coast. Mar. Drugs 2022, 20, 461. [Google Scholar] [CrossRef] [PubMed]
- Kirkpatrick, B.; Bean, J.A.; Fleming, L.E.; Kirkpatrick, G.; Grief, L.; Nierenberg, K.; Reich, A.; Watkins, S.; Naar, J. Gastrointestinal Emergency Room Admissions and Florida Red Tide Blooms. Harmful Algae 2010, 9, 82–86. [Google Scholar] [CrossRef] [PubMed]
- Vila, M.; Masó, M.; Sampedro, N.; et al. The genus Ostreopsis in recreational waters of the Catalan Coast and Balearic Islands (NW Mediterranean Sea): is this the origin of human respiratory difficulties? ISSHA IOC UNESCO 2008, 334–336. [Google Scholar]
- Gallitelli, M. , Ungaro, N. Respiratory illness as a reaction to tropical algal blooms occurring in a temperate climate. JAMA 2005, 293, 2599–2600. [Google Scholar]
- Durando, P.; Ansaldi, F.; Oreste, P.; Moscatelli, P.; Marensi, L.; Grillo, C.; Gasparini, R.; Icardi, G. ; Collaborative Group for the Ligurian Syndromic Algal Surveillance. Ostreopsis ovata and human health: epidemiological and clinical features of respiratory syndrome outbreaks from a two-year syndromic surveillance, 2005-06, in north-west Italy. Euro Surveill 2007, 12. [Google Scholar]
- Milian, A.; Nierenberg, K.; Fleming, L.E.; Bean, J.A.; Wanner, A.; Reich, A.; Backer, L.C.; Jayroe, D.; Kirkpatrick, B. Reported respiratory symptom intensity in asthmatics during exposure to aerosolized Florida red tide toxins. J. Asthma: Off. J. Assoc. Care Asthma 2007, 44, 583–587. [Google Scholar] [CrossRef]
- Fleming, L.E.; Kirkpatrick, B.; Backer, L.C.; Bean, J.A.; Wanner, A.; Reich, A.; Zaias, J.; Cheng, Y.S.; Pierce, R.; Naar, J.; Abraham, W.M.; Baden, D.G. Aerosolized red-tide toxins (brevetoxins) and asthma. Chest 2007, 131, 187–194. [Google Scholar] [CrossRef]
- Todd, E.C.D. Preliminary estimates of costs of foodborne disease in the United States. J. Food Prot. 1989, 52, 595–601. [Google Scholar] [CrossRef]
- Chowdhury, R.; Warnakula, S.; Kunutsor, S.; Crowe, F.; Ward, H. A.; Johnson, L.; Franco, O. H.; Butterworth, A. S.; Forouhi, N. G.; Thompson, S. G. , Khaw, K. T., Mozaffarian, D., Danesh, J., & Di Angelantonio, E. Association of dietary, circulating, and supplement fatty acids with coronary risk: a systematic review and meta-analysis. Annals of internal medicine 2014, 160, 398–406. [Google Scholar] [CrossRef]
- Carter, R. A A.; Joll, C. A. Occurrence and formation of disinfection by-products in the swimming pool environment: A critical review. J. Environ. Sci. (China) 2017, 58, 19–50. [Google Scholar] [CrossRef]
- Heilgeist, S.; Sahin, O.; Sekine, R.; Stewart, R.A. Mapping the Complex Journey of Swimming Pool Contaminants: A Multi-Method Systems Approach. Water 2022, 14, 2062. [Google Scholar] [CrossRef]
- PWTAG - Pool Water Treatment Advisory Group ( (2009). Swimming Pool Water: Treatment and Quality Standards for Pools and Spas. BCPublications.
- Bélanger, A. S.; Brouard, J. S.; Charlebois, P.; Otis, C.; Lemieux, C.; Turmel, M. Distinctive architecture of the chloroplast genome in the chlorophycean green alga Stigeoclonium helveticum. Mol. Genet. Genom. :MGG 2006, 276, 464–477. [Google Scholar] [CrossRef]
- Martić, A.; Čižmek, L.; Ul'yanovskii, N. V.; Paradžik, T.; Perković, L.; Matijević, G.; Vujović, T.; Baković, M.; Babić, S.; Kosyakov, D. S.; Trebše, P.; Čož-Rakovac, R. ; & Čož-Rakovac, R. Intra-Species Variations of Bioactive Compounds of Two Dictyota Species from the Adriatic Sea: Antioxidant, Antimicrobial, Dermatological, Dietary, and Neuroprotective Potential. Antioxid. (Basel Switz.) 2023, 12, 857. [Google Scholar] [CrossRef]
- Puetz, J.D. Swimming Pool Water Chemistry. The Care and Treatment of Swimming Pool Water; Arch Chemicals, Inc., Now a Part of Lonza: Norwalk, CT, USA, 2013.
- Bures, F. Quaternary ammonium compounds: simple in structure, complex in application. Top. Curr. Chem. (Cham) 2019, 377, 14. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Chen, S.; Zhang, H.; Wang, N.; Ma, B.; Liu, X.; Niu, L.; Yang, F.; Xu, Y.; Zhang, X. Effects of copper sulfate algaecide on the cell growth, physiological characteristics, the metabolic activity of Microcystis aeruginosa and raw water application. J. Hazard. Mater. 2023, 445, 130604. [Google Scholar] [CrossRef] [PubMed]
- Juergensen, L.; Busnarda, J.; Caux, P.Y.; Kent, R.A. Fate, behavior, and aquatic toxicity of the fungicide DDAC in the Canadian environment. Environ. Toxicol 2000, 15, 174–200. [Google Scholar] [CrossRef]
- Frank, T. Reregistration Eligibility Decision for Alkyl Dimethyl Benzyl Ammonium Chloride (ADBAC). US Environmental Protection Agency Office of Prevention, Pesticides, and Toxic Substances, Report, 2006, 10–44.
- Chapman, J.S. Biocide resistance mechanisms. Int. Biodeterior. Biodegrad 2003, 51, 133–138. [Google Scholar] [CrossRef]
- Faggio, C.; Pagano, M.; Alampi, R.; Vazzana, I.; Felice, M.R. Cytotoxicity, haemolymphatic parameters, and oxidative stress following exposure to sub-lethal concentrations of quaternium-15 in Mytilus galloprovincialis. Aquat. Toxicol 2006, 180, 258–265. [Google Scholar] [CrossRef] [PubMed]
- Michalek, K.; Ventura, A.; Sanders, T. Mytilus hybridisation and impact on aquaculture: A minireview. Mar. Genom. 2016, 27, 3–7. [Google Scholar] [CrossRef] [PubMed]
- Moore and Kellerman. U. S. Dept. Agri. Bureau Plant Industry, Bull. No. 1994, 64, I904.
- Bishop, W.M.; Johnson, B.M.; Rodgers Jr, J. H. Comparative responses of target and nontarget species to exposures of a copper-based algaecide. J Aquat Plant Manag 2014, 52, 65–70. [Google Scholar]
- Kang, L.; Mucci, M.; Fang, J.; Lürling, M. New is not always better: Toxicity of novel copper based algaecides to Daphnia magna. Ecotoxicol. Environ. Saf. 2022, 241, 113817. [Google Scholar] [CrossRef]
- Peng, Y.; Yang, X.; Ren, B.; Zhang, Z.; Deng, X.; Yin, W.; Yang, S. Algae removal characteristics of the ultrasonic radiation enhanced drinking water treatment process. J. Water Process Eng. 2023, 55, 104154. [Google Scholar] [CrossRef]
- Sheng, D.; Bu, L.; Zhu, S.; Wu, R.; Shi, Z.; Zhou, S. Pre-oxidation coupled with charged covalent organic framework membranes for highly efficient removal of organic chloramines precursors in algae-containing water treatment. Chemosphere 2023, 333, 138982. [Google Scholar] [CrossRef]
- Zhang, Y. , Luo, P., Zhao, S., Kang, S., Wang, P., Zhou, M., Lyu, J. Control and remediation methods for eutrophic lakes in the past 30 years. Control and remediation methods for eutrophic lakes in the past 30 years. Water Sci. Technol. 2020, 81, 1099–1113. [Google Scholar]
- Rondeau, V.; Commenges, D.; Jacqmin-Gadda, H.; Dartigues, J.F. Relation between aluminum concentrations in drinking water and Alzheimer's disease: an 8-year follow-up study. Am. J. Epidemiol. 2000, 152, 59–66. [Google Scholar] [CrossRef]
- El Bouaidi, W.; Libralato, G.; Tazart, Z.; Enaime, G.; Douma, M.; Ounas, A.; Loudiki, M. Nature-based coagulants for drinking water treatment: An ecotoxicological overview. Water Environ. Res. 2022, 94, e10782. [Google Scholar] [CrossRef]
- El Bouaidi, W.; Libralato, G.; Douma, M.; Ounas, A.; Yaacoubi, A.; Lofrano, G.; Loudiki, M. A review of plant-based coagulants for turbidity and cyanobacteria blooms removal. Environ. Sci. Pollut. Res. 2022, 29, 42601–42615. [Google Scholar] [CrossRef] [PubMed]
- El Bouaidi, W.; Essalhi, S.; Douma, M.; Tazart, Z.; Ounas, A.; Enaime, G. ; Yaacoubi, A;, Loudiki, M. Evaluation of the potentiality of Vicia faba and Opuntia ficus indica as eco-friendly coagulants to mitigate Microcystis aeruginosa blooms. Desalin WATER Treat 2020, 196, 198–213. [Google Scholar] [CrossRef]
- Ma, J.; Jia, B.; Li, S.; Kong, Y.; Nie, Y.; Zhang, H.; Gao, T. Enhanced coagulation of covalent composite coagulant with potassium permanganate oxidation for algae laden water treatment: Algae and extracellular organic matter removal. Chem. Eng. J. Adv. 2023, 13, 100427. [Google Scholar] [CrossRef]
- Zeng, S.; Liu, Y.; Wang, Y.; Wang, Y.; Zhou, Y.; Li, L.; Ren, L. Photo-Fenton self-cleaning carbon fibers membrane supported with Zr-MOF@ Fe2O3 for effective phosphate removal from algae-rich water. Chemosphere 2023, 323, 138175. [Google Scholar] [CrossRef]
- Zeng, S.; Liu, Y.; Wang, Y.; Wang, Y.; Zhou, Y.; Li, L.; Ren, L. Photo-Fenton self-cleaning carbon fibers membrane supported with Zr-MOF@ Fe2O3 for effective phosphate removal from algae-rich water. Chemosphere 2023, 323, 138175. [Google Scholar] [CrossRef]
- Barkallah, M.; Elleuch, J.; Smith, K.F.; Chaari, S.; Ben Neila, I.; Fendri, I.; Michaud, P.; Abdelkafi, S. Development and application of a real-time PCR assay for the sensitive detection of diarrheic toxin producer Prorocentrum lima. J. Microbiol. Meth. 2020, 178, 106081. [Google Scholar] [CrossRef]
- Chin Chwan Chuong, J.J.; Rahman, M.; Ibrahim, N.; Heng, L.Y.; Tan, L.L.; Ahmad, A. Harmful Microalgae Detection: Biosensors versus Some Conventional Methods. Sens. (Basel) 2022, 22, 3144. [Google Scholar] [CrossRef]
- Liu, F.; Zhang, C.; Wang, Y.; Chen, G. A review of the current and emerging detection methods of marine harmful microalgae. Sci Total Environ. 2022, 815, 152913. [Google Scholar] [CrossRef]
- Park, J.; Kim, Y.; Kim, M.; Lee, W.H. A novel method for cell counting of microcystis colonies in water resources using a digital imaging flow cytometer and microscope. Environ. Eng. Res. 2019, 24, 397–403. [Google Scholar] [CrossRef]
- Sanseverino, I.; Conduto António, D.; Loo, R.; Lettieri, T. Cyanotoxins: methods and approaches for their analysis and detection. Jt. Res. Cent. (JRC) Tech. Rep. Eur. Comm. 2017. [Google Scholar]
- Ghosh, P.; Mukherji, S. Fate, detection technologies and toxicity of heterocyclic PAHs in the aquatic and soil environments. Sci Total Env. 2023, 892, 164499. [Google Scholar] [CrossRef] [PubMed]
- Jing-Yan, L.; Li-Hua, Z.; Zhen-Hui, R. Recent application of spectroscopy for the detection of microalgae life information: A review. Applied Spectroscopy Reviews 55, 1, 26–59. [Google Scholar]
- Murdock, J. N.; Wetzel, D. L. FT-IR Microspectroscopy Enhances Biological and Ecological Analysis of Algae. Appl. Spectrosc. Rev. 2009, 44, 335–361. [Google Scholar] [CrossRef]
- Saleem, F.; Jiang, J.L.; Atrache, R.; Paschos, A.; Edge, T.A.; Schellhorn, H.E. Cyanobacterial Algal Bloom Monitoring: Molecular Methods and Technologies for Freshwater Ecosystems. Microorganisms 2023, 11, 851. [Google Scholar] [CrossRef] [PubMed]
- Saleem, F.; Jiang, J.L.; Atrache, R.; Paschos, A.; Edge, T.A.; Schellhorn, H.E. Cyanobacterial Algal Bloom Monitoring: Molecular Methods and Technologies for Freshwater Ecosystems. Microorganisms 2023, 11, 851. [Google Scholar] [CrossRef]
- Jin, C.; Mesquita, M.M.F.; Deglint, J.L.; Emelko, M.B.; Wong, A. Quantification of Cyanobacterial Cells via a Novel Imaging-Driven Technique with an Integrated Fluorescence Signature. Sci. Rep. 2018, 8, 9055. [Google Scholar] [CrossRef] [PubMed]
- United States Environmental Protection Agency. Determination of Cyanotoxins in Drinking and Ambient Freshwaters. (Accessed on December 15th, 2023). Available to: https://www.epa.gov/cyanohabs/determination-cyanotoxins-drinking-and-ambient-freshwaters. 15 December.
- Islam, M.S.; Aryasomayajula, A. A Review on Macroscale and Microscale Cell Lysis Methods. Micromachines. Micromachines. Selvaganapathy P.R 2017, 8, 83. [Google Scholar] [CrossRef]
- Pandur, Ž.; Dular, M.; Kostanjšek, R.; Stopar, D. Bacterial Cell Wall Material Properties Determine E. coli Resistance to Sonolysis. coli Resistance to Sonolysis. Ultrason. Sonochem 2022, 83, 105919. [Google Scholar] [CrossRef] [PubMed]
- Brown, R.B.; Audet, J. Current Techniques for Single-Cell Lysis. J. R. Soc. Interface 2008, 5, S131–S138. [Google Scholar] [CrossRef] [PubMed]
- Danaeifar, M. New Horizons in Developing Cell Lysis Methods: A Review. Biotechnol. Bioeng. 2022, 119, 3007–3021. [Google Scholar] [CrossRef] [PubMed]
- Dahlgren, K.K.; Gates, C.; Lee, T.; Cameron, J.C. Proximity-Based Proteomics Reveals the Thylakoid Lumen Proteome in the Cyanobacterium Synechococcus Sp. PCC 7002. PCC 7002. Photosynth. Res. 2021, 147, 177–195. [Google Scholar] [CrossRef] [PubMed]
- Mehta, K.K.; Evitt, N.H.; Swartz, J.R. Chemical Lysis of Cyanobacteria. J. Biol. Eng. 2015, 9, 10. [Google Scholar] [CrossRef] [PubMed]
- Morin, N.; Vallaeys, T.; Hendrickx, L.; Natalie, L.; Wilmotte, A. An Efficient DNA Isolation Protocol for Filamentous Cyanobacteria of the Genus Arthrospira. J. Microbiol. Methods 2010, 80, 148–154. [Google Scholar] [CrossRef]
- Qian, S.S.; Chaffin, J.D.; DuFour, M.R; Sherman, J.J.; Golnick, P.C.; Collier, C.D.; Nummer, S.A.; Margida, M.G. Quantifying and Reducing Uncertainty in Estimated Microcystin Concentrations from the ELISA Method. Environ. Sci. Technol. 2015, 49, 14221–14229. [Google Scholar] [CrossRef]
- 225. Fischer,W.J.; Garthwaite, I.; Miles, C.O.; Ross, K.M.; Aggen ,J.B.; Chamberlin, A.R.; Towers, N.R.; Dietrich, D.R. Congener-Independent Immunoassay for Microcystins and Nodularins. Environ. Sci. Technol. 2001, 35, 4849–4856.
- Shoemaker, J.A.; Tettenhorst, D.R.; de la Cruz, A. Single Laboratory Validated Method for Determination of Microcystins and Nodularin in Ambient Freshwaters by Solid Phase Extraction and Liquid Chromatography/ Tandem Mass Spectrometry (LC/MS/MS) (Accessed on December 16th 2023). Available to: https://www.epa.gov/sites/default/files/2017-11/documents/microcystin_method_for_ambient_water_nov_2017.pdf. 16 December.
- Zaffiro, A.; Rosenblum, L.; Wendelken, S.C. Method 546: Determination of Total Microcystins and Nodularins in Drinking Water and Ambient Water by Adda Enzyme-Linked Immunosorbent Assay (Accessed on th 2023). Available to: https://www.epa.gov/sites/default/files/2016-09/documents/method-546-determination-total-microcystins-nodularins-drinking-water-ambient-water-adda-enzyme-linked-immunosorbent-assay.pdf. 16 December.
- He, X.; Stanford, B.D.; Adams, C.; Rosenfeldt, E.J.; Wert, E.C. Varied Influence of Microcystin Structural Difference on ELISA Cross-Reactivity and Chlorination Efficiency of Congener Mixtures. Water Res. 2017, 126, 515–523. [Google Scholar] [CrossRef] [PubMed]
- Mountfort, D.O.; Holland, P.; Sprosen, J. Method for Detecting Classes of Microcystins by Combination of Protein Phosphatase Inhibition Assay and ELISA: Comparison with LC-MS. Toxicon 2005, 45, 199–206. [Google Scholar] [CrossRef] [PubMed]
- Greer, B.; McNamee, S. E.; Boots, B.; Cimarelli, L.; Guillebault, D.; Helmi, K.; Marcheggiani, S.; Panaiotov, S.; Breitenbach, U.; Akçaalan, R.; Medlin, L. K.; Kittler, K.; Elliott, C. T.; Campbell, K. A Validated UPLC-MS/MS Method for the Surveillance of Ten Aquatic Biotoxins in European Brackish and Freshwater Systems. Harmful Algae 2016, 55, 31–40. [Google Scholar] [CrossRef] [PubMed]
- Coskun, O. Separation Techniques: Chromatography. North. Clin. Istanb. 2016, 3, 156–160. [Google Scholar]
- 232. Kumar, P; Rautela, A.; Kesari, V.; Szlag, D.; Westrick, J.; Kumar, S. Recent Developments in the Methods of Quantitative Analysis of Microcystins. J. Biochem. Mol. Toxicol. 2020, 34, e22582. [CrossRef]
- Kralik, P.; Ricchi, M. A Basic Guide to Real Time PCR in Microbial Diagnostics: Definitions, Parameters, and Everything. Front. Microbiol. 2017, 8, 108. [Google Scholar] [CrossRef]
- Barón-Sola, Á.; Campo, F.F.d.; Sanz-Alférez, S. Dynamics of Cylindrospermopsin Production and Toxin Gene Expression in Aphanizomenon Ovalisporum. Adv. Microbiol. 2016, 6, 381–390. [Google Scholar] [CrossRef]
- Jungblut, A.-D.; Neilan, B.A. Molecular Identification and Evolution of the Cyclic Peptide Hepatotoxins, Microcystin and Nodularin, Synthetase Genes in Three Orders of Cyanobacteria. Arch. Microbiol. 2006, 185, 107–114. [Google Scholar] [CrossRef]
- Ngwa, F.; Madramootoo, C.; Jabaji, S. Monitoring Toxigenic Microcystis Strains in the Missisquoi Bay, Quebec, by PCR Targeting Multiple Toxic Gene Loci. Environ. Toxicol. 2014, 29, 440–451. [Google Scholar] [CrossRef]
- Furukawa, K.; Noda, N.; Tsuneda, S.; Saito, T.; Itayama, T.; Inamori, Y. Highly Sensitive Real-Time PCR Assay for Quantification of Toxic Cyanobacteria Based on Microcystin Synthetase A Gene. J. Biosci. Bioeng. 2006, 102, 90–96. [Google Scholar] [CrossRef]
- Gupta, R.S.; Mathews, D.W. Signature Proteins for the Major Clades of Cyanobacteria. BMC Evol. Biol. 2010, 10, 24. [Google Scholar] [CrossRef] [PubMed]
- Gupta, R.S. Impact of Genomics on Clarifying the Evolutionary Relationships amongst Mycobacteria: Identification of Molecular Signatures Specific for the Tuberculosis-Complex of Bacteria with Potential Applications for Novel Diagnostics and Therapeutics. High-Throughput 2018, 7, 31. [Google Scholar] [CrossRef] [PubMed]
- Duan, X.; Zhang, C.; Struewing, I.; Li, X.; Allen, J.; Lu, J. Cyanotoxin-Encoding Genes as Powerful Predictors of Cyanotoxin Production during Harmful Cyanobacterial Blooms in an Inland Freshwater Lake: Evaluating a Novel Early-Warning System. Sci. Total Environ. 2022, 830, 154568. [Google Scholar] [CrossRef] [PubMed]
- Lu, J.; Struewing, I.; Wymer, L.; Tettenhorst, D.R.; Shoemaker, J.; Allen, J. Use of qPCR and RT-qPCR for Monitoring Variations of Microcystin Producers and as an Early Warning System to Predict Toxin Production in an Ohio Inland Lake. Water Res. 2020, 170, 115262. [Google Scholar] [CrossRef] [PubMed]
- Crevecoeur, S; Edge, T.A.; Watson, L.C.; Watson, S.B.; Greer, C.W.; Ciborowski, J.J.H.; Diep, N.; Dove, A.; Drouillard, K.G.; Frenken, T.; McKay, R.M.; Zastepa, A.; Comte, J. Spatio-Temporal Connectivity of the Aquatic Microbiome Associated with Cyanobacterial Blooms along a Great Lake Riverine-Lacustrine Continuum. Front. Microbiol. 2023, 14. [Google Scholar] [CrossRef]
- Marcuello, C. Present and Future Opportunities in the Use of Atomic Force Microscopy to Address the Physico-Chemical Properties of Aquatic Ecosystems at the Nanoscale Level. Int. Aquat. Res. 2022, 14, 231–240. [Google Scholar] [CrossRef]
- Pillet, F.; Dague, E.; Ilić, J.P.; Ružić, I.; Rols, M.-P.; DeNardis, N.I. Changes in Nanomechanical Properties and Adhesion Dynamics of Algal Cells during Their Growth. Bioelectrochem. Amst. Neth. 2019, 127, 154–162. [Google Scholar] [CrossRef]
- Ceballos-Laita, L.; Marcuello, C.; Lostao, A.; Calvo-Begueria, L.; Velazquez-Campoy, A.; Bes, M.T.; Fillat, M.F.; Peleato, M.-L. Microcystin-LR Binds Iron, and Iron Promotes Self-Assembly. Environ. Sci. Technol. 2017, 51, 4841–4850. [Google Scholar] [CrossRef]
- Wei, Y.-J.; Zhao, Y.-N.; Zhang, X.; Wei, X.; Chen, M.-L.; Chen, X.-W. Biochemical Analysis Based on Optical Detection Integrated Microfluidic Chip. TrAC Trends Anal. Chem. 2023, 158, 116865. [Google Scholar] [CrossRef]
- Knob, R.; Hanson, R.L.; Tateoka, O.B.; Wood, R.L.; Guerrero-Arguero, I.; Robison, R.A.; Pitt, W.G.; Woolley, A.T. Sequence-Specific Sepsis-Related DNA Capture and Fluorescent Labeling in Monoliths Prepared by Single-Step Photopolymerization in Microfluidic Devices. J. Chromatogr. A. 2018, 1562, 12–18. [Google Scholar] [CrossRef] [PubMed]
- Khan, M.; Mao, S.; Li, W.; Lin, J.-M. Microfluidic Devices in the Fast-Growing Domain of Single-Cell Analysis. Chem.—Eur. J. 2018, 24, 15398–15420. [Google Scholar] [CrossRef]
- Gaur, A.; Pant, G.; Jalal, A.S. Computer-Aided Cyanobacterial Harmful Algae Blooms (CyanoHABs) Studies Based on Fused Artificial Intelligence (AI) Models. Algal Res. 2022, 67, 102842. [Google Scholar] [CrossRef]
- Baek, S.-S.; Pyo, J.; Pachepsky, Y. , Park, Y., Ligaray, M., Ahn, C.-Y.; Kim, Y.-H.; Ahn Chun, J.; Hwa Cho, K. Identification and Enumeration of Cyanobacteria Species Using a Deep Neural Network. Identification and Enumeration of Cyanobacteria Species Using a Deep Neural Network. Ecol. Indic. 2020, 115, 106395. [Google Scholar] [CrossRef]
- Hennon, G.M.M.; Dyhrman, S.T. Progress and promise of omics for predicting the impacts of climate change on harmful algal blooms. Harmful Algae 2020, 91, 101587. [Google Scholar] [CrossRef]
- Zhu, M.; Wang, J.; Yang, X.; Zhang, Y.; Zhang, L.; Ren, H.; Wu, B.; Ye, L. A Review of the Application of Machine Learning in Water Quality Evaluation. Eco-Environ. Health 2022, 1, 107–107. [Google Scholar] [CrossRef]
- Almuhtaram, H.; Kibuye, F.A.; Ajjampur, S.; Glover, C.M. , Hofmann, R.; Gaget,V.; Owen, C.; Wert, E.C.; Zamyadi, A. State of Knowledge on Early Warning Tools for Cyanobacteria Detection. State of Knowledge on Early Warning Tools for Cyanobacteria Detection. Ecol. Indic. 2021, 133, 108442. [Google Scholar] [CrossRef]
- WHO, World Health Organization. Guidelines for safe recreational water environments: swimming pool and similar environment. Geneva: WHO 2006.
- Ibelings BW, Kurmayer R, Azevedo SMFO, Wood SA, Chorus I.; Welker M. Understanding the occurrence of cyanobacteria and cyanotoxins. In: Chorus I, Welker M, editors. Toxic cyanobacteria in water, second edition. Geneva: World Health Organization 2021.
- Burch, M.; Brookes, J., Chorus, I. Assessing and controlling the risk of cyanobacterial blooms: waterbody conditions. In: Chorus I, Welker M, editors. Toxic cyanobacteria in water, second edition. Geneva: World Health Organization 2021.
Figure 1.
Schematic representation of evolution of algae according to primary and secondary endosymbiotic events [9,15] and classification according to National Museum of Botany Smithsonian based on morphological traits [8,11,15]. Source of the images are public repositories and Wikipedia.
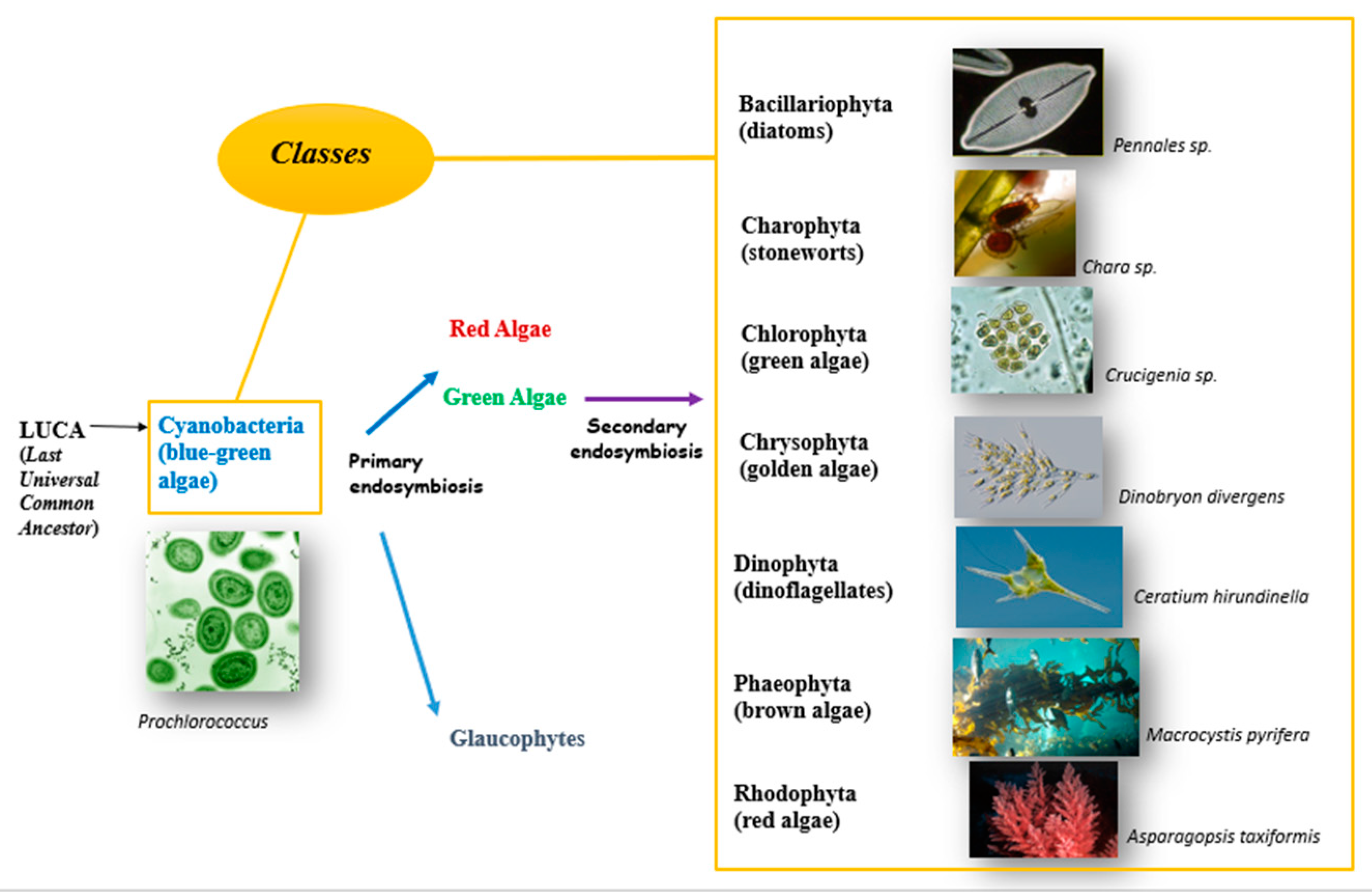
Figure 2.
Temporal trends in harmful algal bloom (HAB) events showed an increase in the number of reported events per year, with statistically significant increases observed from 1980 onwards (Ocean Biodiversity Information System, www.obis.org).
Figure 2.
Temporal trends in harmful algal bloom (HAB) events showed an increase in the number of reported events per year, with statistically significant increases observed from 1980 onwards (Ocean Biodiversity Information System, www.obis.org).
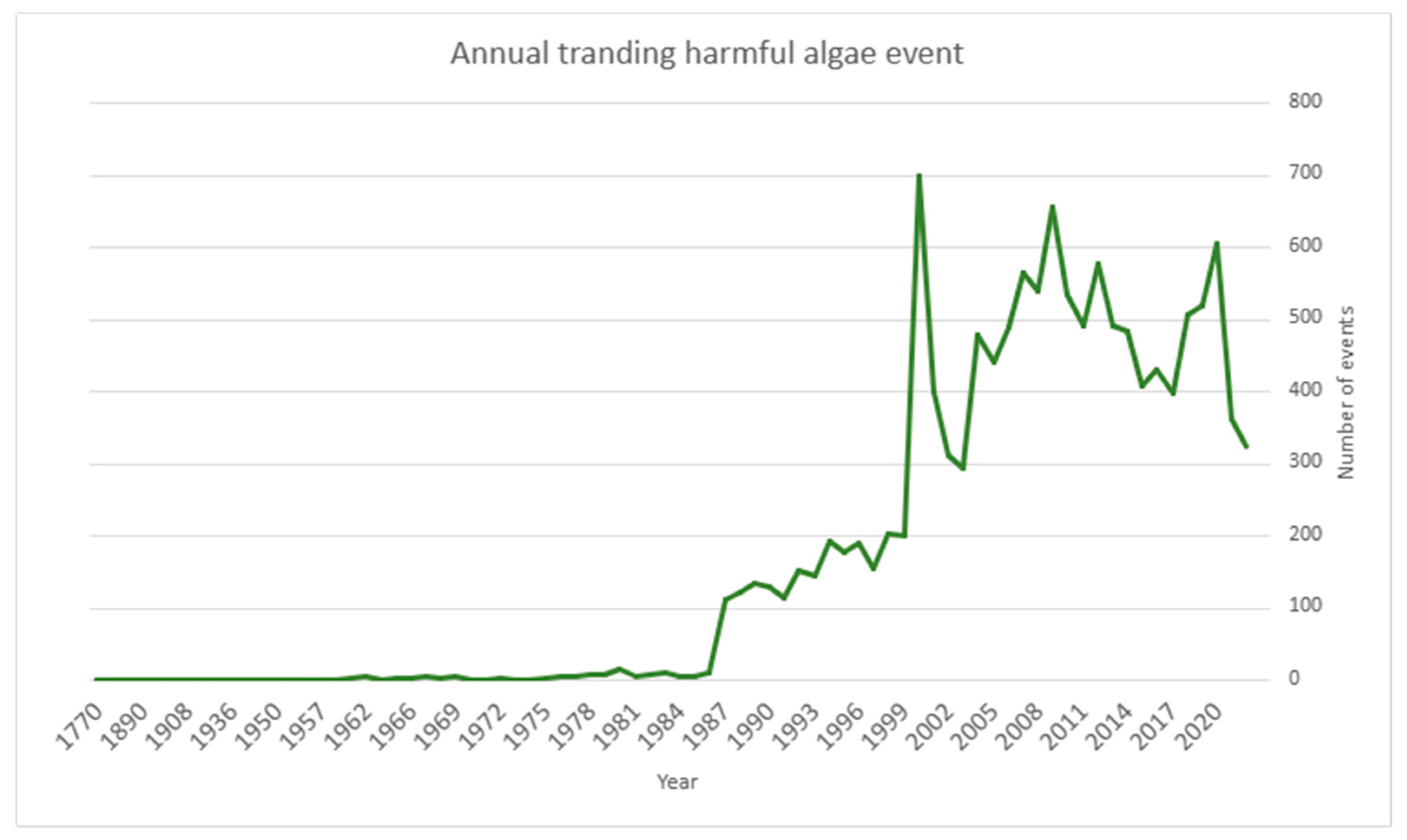
Figure 3.
Responses of eight target algal species (a) and five nontarget animal species (b) to exposures of a copper-based algaecide [Data from ref. 194].
Figure 3.
Responses of eight target algal species (a) and five nontarget animal species (b) to exposures of a copper-based algaecide [Data from ref. 194].
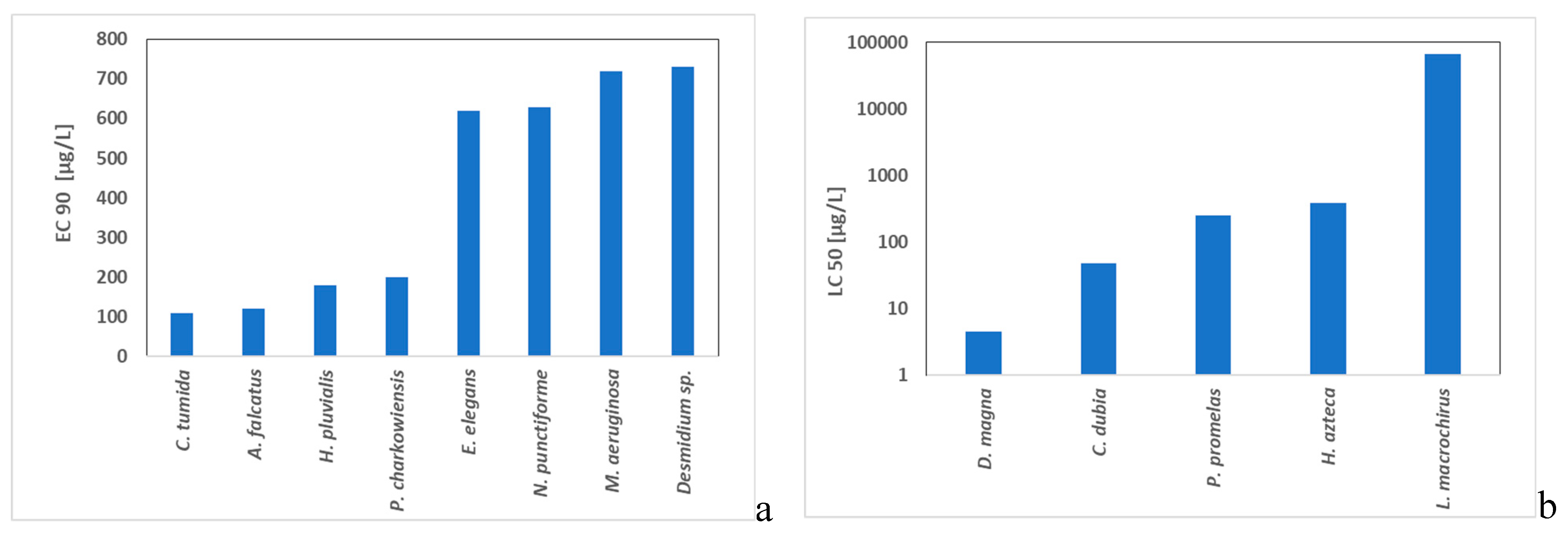
Table 2.
Main suspected and confirmed cases of human illnesses resulting from exposure to cyanobacteria or algae.
Table 2.
Main suspected and confirmed cases of human illnesses resulting from exposure to cyanobacteria or algae.
| Author | Year | Country | Types of Recreational Water | Kingdom | Exposure route | Population | Outcome syndrome |
|---|---|---|---|---|---|---|---|
| Codd, G.A. 1994 ]130] |
1878 | Australia | Lake water | Cyanobacteria Nodularia spumigena | Ingestion | Several hundred livestock deaths; 1 human |
Undescribed illness in human |
| Dillenberg, H.O. 1960 [131] |
1959 | Saskatchewan, Canada | Lake water | Cyanobacteria Anabaena circinalis |
Ingestion | Multiple livestock, fish, geese, dogs died >2 human events |
Gastrointestinal illness |
| Griffiths 2003 [132] |
1979 | Palm Island, Australia | Potable water reservoir | Cyanobacterium Cylindrospermopsis raciborskii | - | 140 children and 10 adults hospitalized |
Hepatoenteritis, acute kidney disease and liver failure |
| Hawkins 1985 [133] |
1979 | Australia | Fresh water | Cylindrospermopsis raciborskii | 138 people | Hepatoenteritis | |
| Jeffery 2004 [134] |
1987 | Canada | Sea water | Pseudo-nitzschia spp. Nitzschia spp. |
Ingestion | 150 people with 19 hospitalization and 4 deaths | Gastrointestinal effects and/or neurological signs |
| Texeira 1993 [135] |
1988 | Brazil | Fresh water | Chlorophyll-a | Drinkingwater from the Itaparicareservoir | 88 human fatalities and 2000 people poisoned |
|
| Azevedo 2002 [136] |
1996 | Brazil | In the filtration system | Cyanobacterium | Dyalisis | 116 human cases | Headache, blurred vision, eye pain, nausea,and vomiting. Of these, 100 patients developed acute liver failure and more than half of them died |
| Backer 2003 [137] |
1999 | Sarasota and Jacksonville, Florida, USA |
Sea water | Karenia brevis | Inalation | 129 adults (≥18 years) who spent time (10 min to ∼8 h; average of 71 min) on beaches during February and October 1999 | Upper and lower respiratory irritation |
| Stewart 2006 [138] |
2006 | Southern Queensland and the Myall Lakes area of New South Wales, Australia; Northeast and central Florida, USA | Lake water | Phytoplankton | Ingestion | 1331 study participants (n = 216 in Florida) comprised of adults and children |
Difficulty breathing, dry and productive cough, runny nose, unusual sneezing sore throat, wheezy breathing, cutaneous, fever |
| Bean 2011 [139] |
2011 | Sarasota, Florida, USA | Sea water | Karenia brevis | Inalation | 125 asthmatic patients residents of the Sarasota, Florida for ≥6 months | |
| Kirkpatrick 2011 [140] |
2011 | Sarasota, Florida, USA | Sea water | Inalation | 52 participants (≥12 years) with self-report of physician-diagnosedasthma |
Increased symptoms of asthma and suppressed respiratory function, suppression for several days after 1 h of exposure to the brevetoxin aerosols | |
| Li 2012 [141] |
2011 | China | Sea water | Amphidoma languida, Azadinium spinosum, Protoceratium reticulatum, Lingulodinium polyedrum, Gonyaulax spinifera, Dinophysis spp, Prorocentrum lima, Dinophysis spp | Consumption of contaminated shellfish | More than 200 people | Diarrheic shellfish poisoning: diarrhea, nausea, vomiting, and abdominal pain |
| Giannuzzi 2011 [142] |
2011 | Uruguay | Fresh water | Cyanobacteria | Drinking water | Human acute intoxication |
Intoxication |
| Levesque 2014 [143] |
2014 | Quebec province, Canada | Lake water | Cyanobacteria | 267 families consisting of 466 subjects |
Gastrointestinal symptoms | |
| Trevino-Garrison 2015 [144] |
2015 | Kansas, USA | Lake water | Cyanobacteria | Direct contact, 100% (7/7), ingestion, 43% (3/7), inhalation, 14% (1/7). The median time from exposure to symptom onset was 24 h, with a range of 3–48 h | 13 humans. Seven confirmed illnesses, median age 40 years | Nausea, vomiting, diarrhea, cough, sore throat, rash and liver damage |
| Berdalet 2022 [145] |
2022 | Sant Andreu de Llavaneres, Catalonia, Spain |
Sea water | Ostreopsis | 16 staff of a nearby indoor-outdoor restaurant directly exposed to marine aerosols at least 8 h daily |
Malaise, runny nose, sore eyes, headaches | |
| Veldee 1931 ]146] |
1930–1931 | United States | River water | Cyanobacteria Anabaena flos-aquae |
drinking water | > 6 events | Gastrointestinal illness |
| Backer 2005 [147] |
2001 and 2003 | Sarasota and Manatee, Florida, USA |
Sea water | Exposure to aerosolized brevetoxins |
28 healthy lifeguards (≥18 years) who are occupationally exposed to red tide toxins during daily work activities | Upper respiratory symptoms and headache | |
| Lee 2022 [148] |
2005–2017 | South Korea | Fresh water | Neurodegenerative disease outcomes including motor neuron disease, Alzheimer’s disease, and Parkinson’s disease | |||
| Figgatt 2016 [149] |
2008 - 2014 | New York, USA | Fresh water | 228 hospital visits: 5.3% of them were hospitalized | |||
| Wu 2021 [150] |
2008–2011 | Greater Boston area, Massachusetts, USA | Fresh water | Cyanobacteria | Human patients who visited the emergency rooms of approximately 70 Massachusetts hospitals | Gastrointestinal, respiratory and dermal illnesses | |
| Hilborn 2014 [151] |
2009–2010 | USA | Lake water | Cyanobacteria | Recreational activities | 41 cases | Gastrointestinal, general, dermatologic, eye/ear, neurologic, respiratory symptoms |
| Roberts 2020 [152] |
2016–2018 | USA | 366 fresh water (94%), 1 brackish, 22 unknown (6%) | Cyanobacteria, Dinoflagellates, Raphidophyceans, Diatoms | Exposure: Public outdoor area 235 (60%), Beach 63 (16%), Private residence 53 (14%), Other 57 (15%), Park 27 (7%), Unknown 22 (6%) | 389 human cases: 341 probable (88%), 14 confirmed, 34 suspected, 153 were < 18 years | Gastrointestinal 262 (67%), generalized 169 (43%), dermatologic 104 (27%), ear, nose, or throat 62 (16%), neurologic 56 (14%), cardiopulmonary 41 (11%), ophthalmologic 30 (8%), other 30 (8%), musculoskeletal 13 (3%), genitourinary 6 (2%), unknown 9 (2%). No deaths were reported |
| Backer 2008 [153] |
3 days in August of 2006 | Unspecified lakes, USA | Lake water | Microcystisaeruginosa | 97 participants | Respiratory (sore throat; congestion; cough; throat irritation; eye irritation; other), dermatologic (itchy skin, red skin, hives, skin irritation, rash, other); other (earache, agitation, headache, abdominal pain, diarrhoea) | |
| Backer 2010 [154] |
August 2007 | Two lakes in Siskiyou County, California, USA |
Lake water | Respiratory (sore throat; congestion; cough; throat irritation; eye irritation; other), dermatologic (itchy skin, red skin, hives, skin irritation, rash, other); other (earache, agitation, headache, abdominal pain, diarrhea) | |||
| Stumpf 2022 [155] |
August 2006–January 2019 |
Sarasota and Manatee Counties, Florida, USA |
Sea water | Volunteers | Respiratory irritation conditions | ||
| Diaz 2019 [156] |
Between 2005 and 2009 | 6 counties in Southwest Florida, USA |
Sea water | Karenia brevis | Emergency department (ED) visitsin patients ≥55 years | Headache | |
| Tichadou 2010 [157] |
Between 2006 and 2009 | French Mediterranean coast | Sea water | French Mediterranean Ostreopsis | 47 patients | General flu-like symptoms, including headache, joint pain, vertigo, fever, fatigue, and diarrhea | |
| Lavery 2021 [158] |
Between January 2009 and April 2019 | USA | Lake water | 380 records in the Commercial database and 178 records in the Medicaid database | Cough, allergy, other malaise and fatigue, headache, routine general medical examination, shortness of breath, hypertension, acute pharyngitis, and acute upper respiratory infection | ||
| Abdullah 2022 [159] |
Between June 2019 and August 2021 |
5 counties in Southwest Florida, USA |
Sea water | Karenia brevis | Exposure to aerosolizedbrevetoxins | 258 participants | Respiratory tract symptoms (coughing, sneezing, sore throat,nasal congestion); neurological (headaches and dizziness); andsystemic outcomes (fever, chills, nausea) |
| Mchau 2019 [160] |
December 2018 | Ukerewe Island, Tanzania |
Lake water | Phytoplankton Chlorophyceae, Cyanophyceae, Bacillariophyceae and Dinophyceae | 134 subjects (31%) used lake water, 229 (53%) used well water and 69 (16%) used treated supplied pipe water as their main source of water for drinking and other domestic uses |
432 subjects > 18 years (mean age 42) | Vomiting, diarrhea, skin irritation, eye irritation, throat irritation and stomach upset |
| Vila 2016 [161] |
From June toDecember 2013 | Sant Andreu de Llavaneres,Catalonia, Spain | Sea water | Ostreopsis | 16 staff of a nearby indoor-outdoor restaurant directly exposed to marine aerosols at least 8 h daily |
Eye irritation, nose irritation, rhinorrhoea, and general malaise | |
| Lavery 2021 [162] |
January 1, 2017–December 31, 2019 | USA | Lake water | Unspecified Cyanobacteria | Inhalation, ingestionor skin contact | Patients aged 18–44 years (37%) and 45–64 years (30%) | 133 respiratory (41%), 44 gastrointestinal (14%), 33 neurologic (10%), and 27 dermatologic (8%) |
| Illoul 2012 [163] |
July 2009 | Beaches in Western area ofAlgiers Wilaya, Algeria | Sea water | Ostreopsis | 300 patients | Fever, rhinitis, conjunctivitis, cough and skin irritations | |
| Figgatt M. 2015 [164] |
June–September 2015 | New York, USA | Fresh waters | Unspecified Cyanobacteria | Recreational activities included swimming (28 patients; 88%); wakeboarding, jet skiing, waterskiing or tubing (seven; 22%); boating (seven; 22%); wading (six; 19%); and using personalwatercraft (four; 13%) |
32 human cases, median age 24 years (2–63 years) |
Skin problems, cough, abdominal pain, diarrhea, nausea, vomiting and other symptoms (e.g. chills, muscle aches, or watery eyes), fatigue/general weakness, Sore throat, Neuroglogic (e.g., headache or seizure); no hospitalizations or deaths were reported |
| Fleming 2005 [165] |
March 2003 | Sarasota, Florida, USA | Sea water | Exposure to aerosolizedbrevetoxins during red tide events | 59 persons (≥12 years) with physician-diagnosed asthma |
Respiratory symptoms and respiratory impairment |
|
| Fleming 2009 [166] |
March 2005 and September 2006 | Sarasota, Florida, USA | Sea water | Exposure to aerosolized brevetoxins |
87 persons (≥12 years) with physician-diagnosed asthma |
Respiratory symptmos like cough, wheezing, shortness of breath and other like throat irritation, nasal congestion, eye irritation, headache, itchy skin, diarrhoea |
|
| Hoagland 2014 [167] |
Monthly emergency department (ED) data 2005–2009 and monthly hospital inpatient data from 1999 to 2009 | 6 counties in Southwest Florida, USA |
Sea water | Karenia brevis | Emergency department (ED) visits in patients ≥55 years |
Adverse respiratory and digestivehealth effects | |
| Hoagland 2009 [168] |
October 2001–September 2006 | Sarasota, Florida, USA | Sea water | Karenia brevis | Exposure to aerosolized brevetoxins |
Daily emergency department (ED) visits |
Pneumonia, bronchitis, asthma, upper airway disease UAD |
| Kirkpatrick 2006 [169] |
October 1–December 31, 2001 (red tide period) and October 1–December31, 2002 (non-red tide period) |
Sarasota, Florida, USA | Sea water | Red tide | Emergency room (ER) visits | Pneumonia 31%, bronchitis 56%, asthma 44%, and upper airway disease 64% |
|
| Chomerat 2022 [170] |
September 2020 and 2021 | Several beaches on the French Basque coast |
Sea water | Ostreopsis | Direct contact with water and/or after inhalation of aerosols | 674 confirmed cases | Difficulty breathing, irritations, headaches, dry cough, nose irritation, eye irritation and/or dermatitis, and general malaise |
| Kirkpatrick 2010 [171] |
September 1–December 31, 2001, and thenon-bloom period during the fall of 2002, September 1–December 31, 2002 |
Sarasota, Florida, USA | Sea water | Exposure to aerosolized brevetoxins |
Number of emergency room (ER) admissions during the Florida red tide bloom |
Gastritis, duodenitis, and non-infectious enteritis and colitis (acute, chronic, unspecified, and other) |
|
| Vila 2008 [172] |
Summer 2004 | Sant Andreu de Llavaneres, Catalonia, Spain |
Sea water | 62 people | Rhinorrhea (74.2%), nose irritation (66.1%), throat irritation (62.9%), coughing (59.7%), expectoration (51.6%), eye irritation (41.4%) and migraine (40.3%) | ||
| Galitelli 2006 [173] |
Summers of 2003 and 2004 |
Coasts of Bari, Italy | Sea water | Ostreopsisgenus | Recreational or working activities on the beach |
28 patients ( 1 child; mean age, 38.6 years) |
Rhinorrhoea 100%; cough 43%; fever 14%; and bronchoconstriction with mild dyspnoea and wheezes 43%. Conjunctivitis 11% |
| Bresciani & Durando 2007 [174] |
Summers of 2005 and 2006 | Genova and La Spezia, Italy |
Sea water | Ostreopsis ovata | 209 patients (mean age 35.9 ± 20.1 years, range 1–89 years). 43 patients seeking medical help at the emergency departments needed hospitalization | Fever (63.6%), sore throat (50.2%), cough (40.2%), and dyspnoea (38.8%) | |
| Milian 2007 [175] |
Unexposed period: March 2003 and March 2005; exposed period: January 2003, May 2004 October 2004 |
Sarasota, Florida, USA | Sea water | Exposure to aerosolized brevetoxins |
97 persons (≥12 years) with physician-diagnosed asthma |
Respiratory symptoms like eye and/or throat irritation; nasal congestion; cough; wheeze; chest tightness |
|
| Fleming 2007 [176] |
Unexposed period: March 2003 and March 2005; exposed period: January 2003, May 2004 October 2004 |
Sarasota, Florida, USA | Sea water | Exposure to aerosolized brevetoxins |
97 persons (≥12 years) with physician-diagnosed asthma |
Respiratory symptoms like eye and/or throat irritation; nasal congestion; cough; wheeze; chest tightness |
Table 3.
Chemical characteristics of QAC .
| QAC | Formula | CAS-Number | Molecular weight |
Half-life | Log Koc | Log Dow/Kow |
| ATMACs | CH3(CH3)7-17 N(CH3)3Cl | C8:10108-86-8 C10: 10108-86-9 C12: 112-00-5 C14: 4574-04-3 C16: 112-02-7 C18: 112-03-8 |
C8: 207.8 C10: 235.8 C12: 263.9 C14: 291.9 C16: 320.0 C18: 348.0 |
- | C8: 2.78 C10: 3.30 C12: 3.82 C14: 4.34 C16: 4.36 C18: 5.38 |
C8: -1.05 C10: -0.189 C12: 0.857 C14: 1.77 C16: 2.43 C18: 3.25 |
| DDACs | [CH3(CH3)7-17]2 N(CH3)2Cl | C8: 5538-94-3 C10: 7173-51-5 C12: 3401-74-9 C14: 10108-91-5 C16: 1812-53-9 C18:107-64-2 |
C8: 306.0 C10: 362.1 C12: 418.2 C14: 474.3 C16: 530.4 C18: 586.5 |
180 days in river water (EPA, 2017) 1048 days in soil [189] |
C8:4.64 C10: 5.68 C12: 6.73 C14: 7.71 C16: 8.81 C18: 9.86 |
C8: 1.57 C10: 2.59 C12: 4.31 C14: 6.25 C16: 9.98 C18: 9.52 |
| BACs | CH3(CH3)5-17N(Cl) (CH3)2 CH2 C6H5 | C6: 22559-57-5 C8: 959-55-7 C10: 965-32-2 C12: 139-07-1 C14: 139-08-2 C16: 122-18-9 C18: 122-19-0 |
C6: 255.8 C8: 283.9 C10: 311.9 C12: 340.0 C14: 368.0 C16: 396.1 C18: 424.1 |
379 days (in water at pH 9) [190] |
C6: 3.87 C8: 4.39 C10: 4.91 C12: 5.43 C14: 5.96 C16: 6.48 C18: 7.00 |
C6: -0.763 C8: 0.233 C10: 1.31 C12: 2.10 C14: 2.78 C16: 3.54 C18: 4.28 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



