
Submitted:
27 December 2023
Posted:
28 December 2023
You are already at the latest version
Abstract
Anthracnose stands as the primary obstacle to lupin cultivation, impeding development despite the considerable agronomic, ecological, and economic potential of such legume crops. This review explores recent efforts to unravel the complexities of anthracnose in domesticated lupins, focusing on both the plant perspective and the causative pathogenic agent, Colletotrichum lupini. Leveraging cutting-edge technologies has yielded crucial insights into various facets of this devastating disease, encompassing plant and pathogen biology, genetic and molecular regulations of the interaction, fungal diversity and population dynamics, and screening of plant genetic resources for anthracnose resistance. The lack of effective disease control measures, relying primarily on the use of disease-free seeds, highlights the need to develop anthracnose-resistant varieties. However, challenges arise from the intricacy of lupin's response to the disease, influenced by polygenic inheritance, in spite of loci with major effects, and environmental factors. The slow pace of genetic improvement underscores the need for more efficient breeding processes, including biotechnological approaches. This review offers a comprehensive overview of current progress and knowledge gaps, stressing the urgent need to further enhance understanding of C. lupini pathogenic mechanisms and lupin‘s resistance. Integrating advanced technologies and accelerated research efforts is paramount for achieving efficient disease management and sustainable lupin cultivation in the face of anthracnose challenges.
Keywords:
Colletotrichum lupini
; Lupinus spp
; disease management
; early molecular pathogen detection
; infection mechanisms
; molecular regulatory pathways
; prophylaxis
; genetic resistance
; breeding
1. Anthracnose: A Foremost Devastating Threat in Lupins
Lupins (Lupinus spp.) are economically important as versatile crops with applications in animal feed, human consumption, and biofuel production, contributing to agricultural diversification and economic sustainability. Agronomically, lupins enhance soil fertility through nitrogen fixation and promote sustainable farming practices, while ecologically, they play a crucial role in crop rotation systems, breaking pest cycles and fostering biodiversity in agricultural landscapes. Prominent cultivated lupin species encompass Lupinus angustifolius (narrow-leaved blue lupin), Lupinus albus (white lupin), Lupinus luteus (yellow lupin), and Lupinus mutabilis Sweet (sweet pearl lupin, also recognized as the Andean lupin or tarwi). These are pulse legumes, cultivated primarily for their seeds known for their elevated protein content. In 2020, worldwide lupins cultivation covered a total harvested area of 888,507 hectares; Australia contributed to approximately 55% of this production, while Europe accounted for 32%, according to the FAO [1].
Cultivation of lupins faces constraints due to their susceptibility to diseases, resulting in substantial declines in both yield quantity and quality. Among the pressing challenges, anthracnose caused by the seed- and air-borne fungal pathogen Colletotrichum lupini stands out as the most important disease affecting lupin cultivation worldwide [2]. Anthracnose manifests as sunken, necrotic lesions that can develop on any aerial part of the lupin plant (Figure 1; [3]).
The failure of the lupin industry to address anthracnose threats on white lupins (L. albus L.) in France (1982) and Ukraine (1983) led to subsequent outbreaks in major lupin-producing countries, including Australia, Poland, and Germany [4,5,6]. This negligence resulted in a ban on white lupin cultivation in Australia until 1998 due to its proven susceptibility and challenges in finding resistance [5,7]. The spread of the disease in Australia in 1996 prompted the development of disease management strategies, focusing on the identification of resistant plant varieties [5]. Currently, white lupin holds significant promise as a local alternative to imported soybean in Europe, owing to its high protein content suitable for the food and feed industry. However, its cultivation is still hindered by the prevalence of anthracnose disease that results in substantial yield losses, emphasizing the need for identifying novel genetic resistance sources and developing new prophylactic and early detection tools to effectively manage the disease.
2. Colletotrichum lupini: The Causative Agent for Anthracnose in Lupins
In the search for resistance against a pathogen and for effective disease management, a comprehensive understanding of various facets of the pathogen, including its taxonomy, lifestyle, and the strategies employed for plant infection and establishment, becomes imperative [8].
Anthracnose is attributed to the Colletotrichum genus (phylum: Ascomycota, class: Sordariomycetes), an economically significant group of pathogenic fungi. It affects a diverse array of plants, including fruits, vegetables, cereals, ornamentals, and tree species, resulting in considerable pre- and post-harvest losses [6,9,10,11,12,13,14]. Although generally considered a host-specific pathogen of lupins [6,15], C. lupini has been reported in non-typical hosts, including olive trees in Morocco [16] and Ceylon cinnamon trees [17]. Colonies of C. lupini grown on Potato Dextrose Agar (PDA) in the dark exhibit a grey to brown coloration with a woolly appearance, producing dark mycelia with an entire margin (Figure 1a). These strains generate pink or orange conidia on both simple and branched conidiophores, often accompanied by melanized appressoria [9,17,18].
2.1. C. lupini Belongs to the Colletotrichum Acutatum Species Complex (CAsc)
C. lupini belongs to the Colletotrichum acutatum species complex (CAsc) [6,19,20,21,22], which is a diverse complex [23] of more than 50 species as of 2023 [24,25]. The C. acutatum species complex (CAsc) infects a diverse range of hosts and has been proposed as a model system for studying the evolution of plant-pathogen adaptations due to the diversity of mechanisms present in the complex [22]. It is the most recently diverged lineage within the Colletotrichum genus, having emerged approximately 15.69 million years ago [25]. Morphologically distinct from other members of the Colletotrichum genus, CAsc is characterized by conidia with acute ends [10,23,26]. Additionally, with an average genome size of 50.37 Mb compared to the varied genome sizes of different Colletotrichum species ranging from 44.15 to 109.66 Mb, the C. acutatum complex has the lowest repetitive DNA content (approximately 2.43%) [25]. Molecular loci such as glyceraldehyde-3-phosphate dehydrogenase (GAPDH) and β-tubulin 2 (tub2) have proven valuable for classifying most species within the complex, while the Apn2-Mat1-2-1 intergenic locus (APN/MAT1) is particularly useful for intraspecies segregation [24,25].
2.2. Colletotrichum lupini Reproduces Clonally, Exhibiting Distinct Morphologies and Virulence Patterns Both between and within Clonal Lineages
Understanding C. lupini population structure and evolutionary potential is crucial to design successful disease management strategies. Figure 2 provides a chronological overview of the key milestones in the taxonomic classification and population genetics of the species C. lupini, accompanied by a progression of the methodologies employed throughout different periods.
The most comprehensive population genetic survey was recently published, describing a refined population structure within C. lupini species [27]. It is based on 3D-RADseq (triple digest restriction site-associated DNA sequencing) high-throughput genotyping of a worldwide collection of 76 Colletotrichum isolates including 67 C. lupini strains, achieving an unprecedented resolution level. Phylogenetic and structural analyses revealed four distinct lineages (I-IV), highlighting a robust population structure, and a notably high standardized index of association (r̅d), indicative of clonal reproduction in C. lupini. Nevertheless, varied morphologies and virulence patterns on white lupin (L. albus) and Andean lupin (L. mutabilis) were observed among and within clonal lineages. Lineage II isolates exhibited a unique minichromosome, partly shared with lineages III and IV but absent in lineage I isolates, suggesting a potential role in host-pathogen interactions. All four lineages were identified in the South American Andes region, recognized as the species' center of origin. Since the 1990s, only lineage II members have been found outside South America, designating it as the current pandemic population. To effectively manage the disease, strategies should prevent the dissemination of other lineages from South America while prioritizing resistance development against lineage II beyond this region, considering Andean lupins as promising untapped resistance sources.
Figure 2.
Timeline illustrating the evolution of the characterization and classification of Colletotrichum lupini isolates. Historical methods for the identification and classification of Colletotrichum isolates affecting lupines have involved a combination of morphological and molecular techniques, along with the vegetative compatibility group (VCG) system. Such methods allowed Yang and Sweetingham [28] to delineate three groups, with COL2 emerging as the predominant one. Initially designated as Colletotrichum gloeosporioides, C. lupini was subsequently reclassified into the Colletotrichum acutatum species complex (CAsc) by Talhinhas et al. [18] as strongly supported by ITS, Tub2, His4 markers. Isolates derived from anthracnose-infected lupins formed a distinct group, designated as Group A1, subsequently acknowledged as the clade representing the C. lupini species [23]. Nirenberg et al. [9] first classified C. lupini as a distinct species within CAsc, identifying two varieties: C. lupini var. lupini and C. lupini var. setosum. Dubrulle et al. [17] utilized multilocus sequencing to characterize 71 isolates from western France along with 16 reference lines of C. lupini. The findings reconciled the earlier classification systems, as they revealed the existence of two distinct groups belonging to the A1 group described by Talhinhas et al. [18]: Group I, a smaller cluster encompassed only three strains, two of which were recognized as C. lupini var. lupini by Nirenberg et al. [9]; in contrast, the remaining isolates formed the larger group (group II), which included one strain identified as C. lupini var. setosum by Nirenberg et al. [9] and another belonging to the most abundant subgroup of A1 as described by Talhinhas et al. [18]. The identification of only one haplotype for C. lupini var. setosum suggested clonal dispersion leading to global dissemination [19]. Isolates from Chile exhibited high genetic variability, highlighting the rich intra-species diversity of C. lupini within South America, which is most likely its center of origin [6,29]. A recent study involving strains from five continents revealed six genetically diverse groups with ten morphotypes, all found within South America but geographically separated [30]. Further advances in understanding C. lupini's population structure, utilizing high-throughput genotyping through Next-Generation Sequencing (NGS) on a global Colletotrichum isolates collection, have finally unveiled four lineages (I-IV) and underscored the dynamics of C. lupini populations, pinpointing the clonal lineage II population as the primary driver behind the global lupin anthracnose pandemic [27]. Additionally, the recently unveiled whole-genome sequences for two reference strains provides an opportunity to deepen our understanding of C. lupini's genetic architecture, particularly with respect to host speciation and pathogenicity factors [14]. The icons were created using BioRender (https://biorender.com/).
Figure 2.
Timeline illustrating the evolution of the characterization and classification of Colletotrichum lupini isolates. Historical methods for the identification and classification of Colletotrichum isolates affecting lupines have involved a combination of morphological and molecular techniques, along with the vegetative compatibility group (VCG) system. Such methods allowed Yang and Sweetingham [28] to delineate three groups, with COL2 emerging as the predominant one. Initially designated as Colletotrichum gloeosporioides, C. lupini was subsequently reclassified into the Colletotrichum acutatum species complex (CAsc) by Talhinhas et al. [18] as strongly supported by ITS, Tub2, His4 markers. Isolates derived from anthracnose-infected lupins formed a distinct group, designated as Group A1, subsequently acknowledged as the clade representing the C. lupini species [23]. Nirenberg et al. [9] first classified C. lupini as a distinct species within CAsc, identifying two varieties: C. lupini var. lupini and C. lupini var. setosum. Dubrulle et al. [17] utilized multilocus sequencing to characterize 71 isolates from western France along with 16 reference lines of C. lupini. The findings reconciled the earlier classification systems, as they revealed the existence of two distinct groups belonging to the A1 group described by Talhinhas et al. [18]: Group I, a smaller cluster encompassed only three strains, two of which were recognized as C. lupini var. lupini by Nirenberg et al. [9]; in contrast, the remaining isolates formed the larger group (group II), which included one strain identified as C. lupini var. setosum by Nirenberg et al. [9] and another belonging to the most abundant subgroup of A1 as described by Talhinhas et al. [18]. The identification of only one haplotype for C. lupini var. setosum suggested clonal dispersion leading to global dissemination [19]. Isolates from Chile exhibited high genetic variability, highlighting the rich intra-species diversity of C. lupini within South America, which is most likely its center of origin [6,29]. A recent study involving strains from five continents revealed six genetically diverse groups with ten morphotypes, all found within South America but geographically separated [30]. Further advances in understanding C. lupini's population structure, utilizing high-throughput genotyping through Next-Generation Sequencing (NGS) on a global Colletotrichum isolates collection, have finally unveiled four lineages (I-IV) and underscored the dynamics of C. lupini populations, pinpointing the clonal lineage II population as the primary driver behind the global lupin anthracnose pandemic [27]. Additionally, the recently unveiled whole-genome sequences for two reference strains provides an opportunity to deepen our understanding of C. lupini's genetic architecture, particularly with respect to host speciation and pathogenicity factors [14]. The icons were created using BioRender (https://biorender.com/).

2.3. Recently Unveiled Complete Genome Sequences May Provide Substantial Potential for Uncovering the Pathogenicity Factors of This Devastating Plant Pathogen
The first available genome sequence information of C. lupini was the 36.554 kb mitogenome (mtDNA) of C. lupini var. setosum (COL-2), isolated from South African white lupin, encompassing 46 genes, including 15 protein-coding genes, two ribosomal RNAs, and 29 tRNA genes [31]. Presently, two C. lupini strains have complete nuclear genomic sequences available (https://mycocosm.jgi.doe.gov/mycocosm/home/releases?flt=colletotrichum+lupini). In 2021, the genome of strain RB221, obtained from symptomatic white lupin in France and representative of the predominant C. lupini population globally (lineage II), was sequenced. The total assembly size comprised 63.41 Mb, distributed across 11 nuclear chromosomes, and an additional 36.55 Mb for one circular mtDNA [14]. Functional annotation predicted over 18,000 genes, including 39 specific to C. lupini. The genome of the Ukrainian strain CBS 109225 (belonging to lineage I [30]) was recently sequenced, resulting in a 58.76 Mbp assembly length and 15,365 predicted genes (https://mycocosm.jgi.doe.gov/Collu1/Collu1.info.html; Baroncelli et al., 2022).
3. Pathogenesis and Epidemiology of Anthracnose in Lupins: Insights into Primary and Secondary Infection Mechanisms
3.1. Dissemination of Anthracnose through Primary Infection: Asymptomatic Infected Seeds as the Principal Mode of Spread
Anthracnose primarily spreads through primary infection via asymptomatic infected seeds [6,35], with the pathogen primarily concentrated on the seed coat, while sparing the embryo [57]. Although infected seeds may appear healthy, they often exhibit a wrinkled seed coat and may display necrosis (Figure 1e); nonetheless, these seeds retain the capacity to germinate. Extensive agricultural losses, impacting both lupin field crops and storage, have been attributed to the fungi's ability to persist in a latent state within asymptomatic seeds and plants, remaining undetected and contributing to the widespread dissemination of the disease [15,35]. A spatio-temporal modeling estimated that even a small proportion (0.01%) of infected lupin seeds could potentially compromise at least 15% of susceptible crops in a field under optimal climatic conditions, characterized by suitable temperature and high rainfall [6,36].
Secondary infections typically occur due to splashing raindrops, leading to the dispersal of conidia from infected seedlings [6,37]. The disease can affect all aerial parts of lupin plants at any developmental stage, causing symptoms such as the curling and twisting of leaves, petioles, pods, shoots, and flowers, resulting in the bending of stems resembling a 'shepherd's hook' and eventual plant death (Figure 1). Additionally, the presence of dark spots, necrotic lesions, and orange conidia on aerial plant parts has been observed [6,11,23]. In Australia, yield loss has been significantly associated with seasonal rainfall and warm temperatures, where wet leaves create favorable conditions for secondary infection [5,6,32,38]. The lack of resistance remains a major contributing factor; without proper management, anthracnose has the potential to devastate entire fields of susceptible cultivars [34].
3.2. Colletotrichum lupini Adopts a Hemibiotrophic Lifestyle with an Initial Brief Biotrophic Phase Succeeded by a Highly Destructive Necrotrophic Development
The initial phase of plant infection by C. lupini involves a biotrophic process, utilizing large intracellular primary hyphae. Colletotrichum spp. exhibit specialized features facilitating the infection of host tissues, including germ tubes, appressoria, intracellular hyphae, and narrower secondary hyphae. Once entering host tissues, the fungus employs various colonization strategies, ranging from intracellular hemibiotrophy to subcuticular intramural necrotrophy, where the pathogen enters a quiescent stage. Some endophytic behaviors have also been documented [2,39,40,41,42].
The infection pathway of C. lupini in L. mutabilis (Andean lupin) petiole and stem tissues has been recently described [2]. C. lupini conidia undergo germination, forming melanized dome-shaped appressoria within 12 hours post-infection (Figure 2) [43]. These appressoria penetrate host tissue through a combination of mechanical and enzymatic degradation [39,41]. The formation of the penetration peg, marking the initiation of host colonization, is succeeded by the development of primary hyphae, marking the onset of the biotrophic phase [44,45].
In the subcuticular intramural necrotrophy stage, a brief symptomless period (latent stage) precedes the penetration into cells, rapidly transitioning to necrotrophy within a day of plant penetration. The shift to the necrotrophic phase involves hyphal extension into epidermal, cortex, and vascular cells [2,3,39,40,42,45]. Within the epidermal cells, cytoplasm disorganization and plasma membrane breakdown occur, accompanied by the degradation of organelles such as the chloroplast. Concurrently, acervuli develop during this stage, producing conidia that serve as potential initiators for secondary infections, perpetuating the cycle [2,39,44,45].
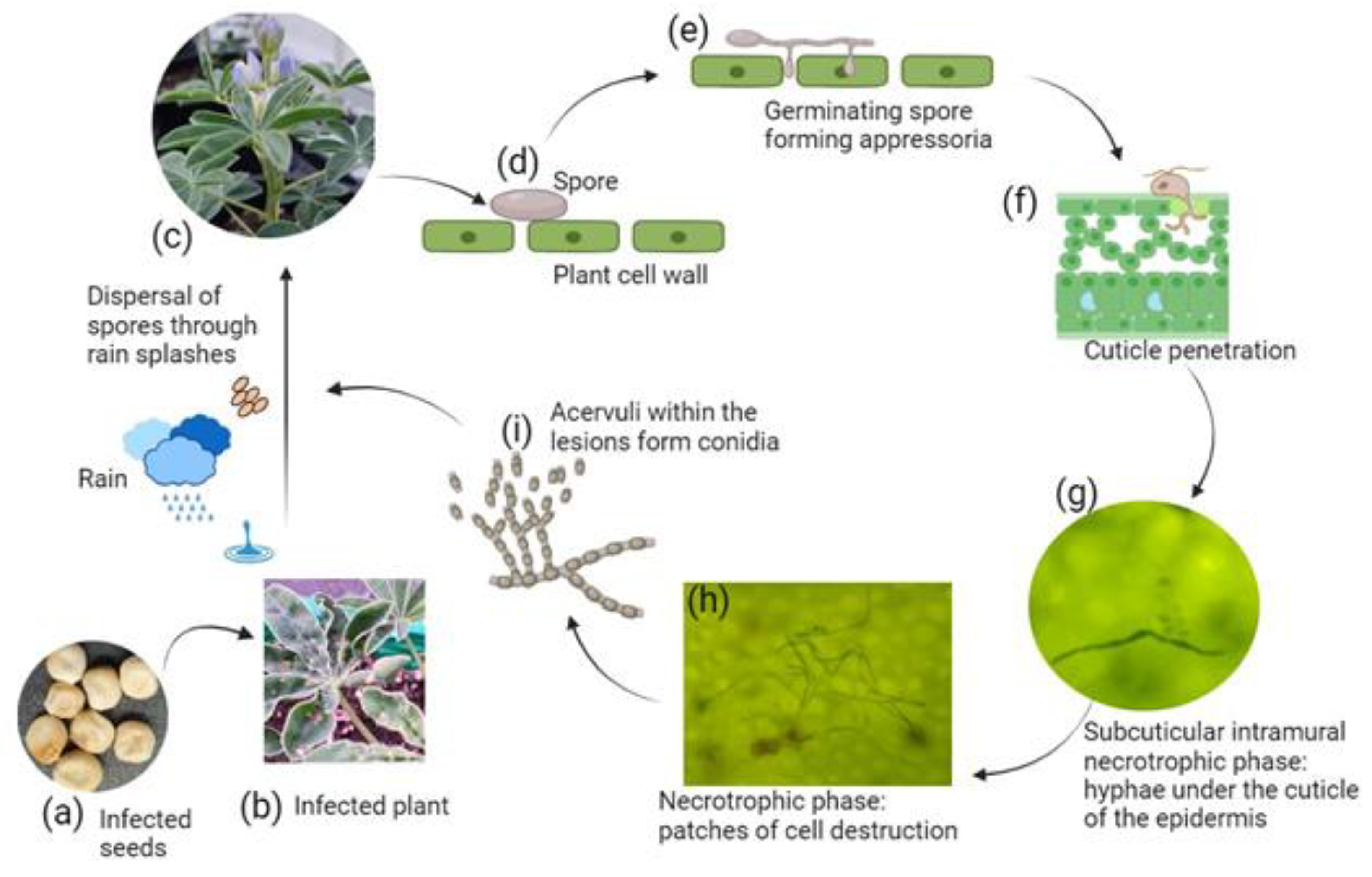
Figure 3.
The hemibiotrophic lifecycle of C. lupini during colonization of lupine plants. Anthracnose predominantly disseminates through primary infection from asymptomatic infected seeds (a). Rain droplets in the field disperse spores from infected plants (b) onto uninfected ones (c). This phenomenon is replicated in laboratory conditions by infecting healthy seeds and plants using spore inoculum. Spores settle on the cell surface (d) and undergo germination to form melanized appressoria (e) [2]. Appressoria have been observed as early as 12 hours post infection [43]. By 24 hours post infection, the biotrophic phase commences with the penetration of germinated spores into cuticle (f) [2,43]. In the subcuticular intramural necrotrophic phase (g), C. lupini produces hyphae beneath the cuticle, expanding within the epidermal cell walls without entering the protoplasts, maintaining this stage for up to 48 hours [2]. Subsequently, the fungus transitions to the necrotrophic phase within tissues (h), resulting in cell disruption and death [2]. Dark sunken lesions on the stems become evident at 60 to 84 hours post infection, with disrupted cells appearing as brown to black lesions under microscopic examination. Acervuli within the lesions generate conidia (i), which are released and transferred to healthy plants through rain splashes (c). The figure was generated using BioRender (https://biorender.com/), and microscopic images (g) and (h) were captured under x40 magnification.
Figure 3.
The hemibiotrophic lifecycle of C. lupini during colonization of lupine plants. Anthracnose predominantly disseminates through primary infection from asymptomatic infected seeds (a). Rain droplets in the field disperse spores from infected plants (b) onto uninfected ones (c). This phenomenon is replicated in laboratory conditions by infecting healthy seeds and plants using spore inoculum. Spores settle on the cell surface (d) and undergo germination to form melanized appressoria (e) [2]. Appressoria have been observed as early as 12 hours post infection [43]. By 24 hours post infection, the biotrophic phase commences with the penetration of germinated spores into cuticle (f) [2,43]. In the subcuticular intramural necrotrophic phase (g), C. lupini produces hyphae beneath the cuticle, expanding within the epidermal cell walls without entering the protoplasts, maintaining this stage for up to 48 hours [2]. Subsequently, the fungus transitions to the necrotrophic phase within tissues (h), resulting in cell disruption and death [2]. Dark sunken lesions on the stems become evident at 60 to 84 hours post infection, with disrupted cells appearing as brown to black lesions under microscopic examination. Acervuli within the lesions generate conidia (i), which are released and transferred to healthy plants through rain splashes (c). The figure was generated using BioRender (https://biorender.com/), and microscopic images (g) and (h) were captured under x40 magnification.
3.3. Colletotrichum lupini Exhibits a Wide Temperature Tolerance Range
C. lupini demonstrates a broad temperature tolerance, surviving within the range of 5°C to 35°C, with an optimum temperature around 25°C [9,17,32]. At low temperatures, the strain enters an extended latent state, resulting in slowed growth both in vitro and in vivo, with full recovery of growth rate when exposed to optimal temperature. The capacity to survive a latent phase in non-optimal conditions poses challenges for disease management, as spores from previous infections can endure, awaiting favorable conditions to germinate. Persistent infections from year to year, arising from dormant infections in nurseries and seeds, pose a significant threat, despite a decreased disease frequency at cold temperatures (5°C to 10°C) across various cultivars, including susceptible ones from L. albus (white lupin) and L. angustifolius (narrow-leaf lupin) species [17,32].
Conversely, at elevated temperatures, mycelia growth stops after 40 days at 37°C, and the failure to recover upon returning to the optimal temperature implies potential non-survival or significant slowdown of C. lupini in planta [17].Given these findings, high-temperature seed treatments have been proposed as a potential strategy to control the spread of the pathogen [33,34].
4. Molecular Mechanisms Governing the Interaction between C. lupini and Lupins
Molecular studies play a central role in revealing variations in the metabolomic, proteomic, and transcriptomic profiles of the host plant and fungal pathogen throughout the infection process. This improves our understanding of the intricate regulatory biological pathways governing plant-pathogen interaction, influencing the development or prevention of anthracnose. In addition, these studies make it possible to identify potential breeding targets for the development of new resistant varieties, subject to prior functional validation of the identified candidate genes.
4.1. Unveiling the Main Molecular Regulatory Pathways in the Interaction between C. lupini and lupins: Contributions from -Omics Analysis
4.1.1. Omics Analysis of the Hemibiotrophic Lifestyle of C. lupini to Uncover Putative Key Virulence Factors
Dubrulle et al. [43] investigated the hemibiotrophic lifestyle of C. lupini during white lupin (L. albus) infection through transcriptomic and proteomic analyzes, revealing a finely tuned temporal regulation of the fungal pathogen molecular pathways guiding the transition from biotrophic to necrotrophic stages. The infection initiated with appressorium development within the first 24 hours post-inoculation, accompanied by increased expression of genes related to cellular processes. The transition to biotrophic-necrotrophic phases prompts a sudden increase in the expression of genes associated with pathogenicity and stress response, including Necrosis- and Ethylene-inducing Proteins (NEP) and Nudix. This increase extends to genes covering diverse protein families linked to the infectious process, such as carbohydrate-active enzymes (CAZymes) that may degrade the host plant cell wall, peptidases, transmembrane transporters, and candidate effectors—mobile small secreted peptides enabling the pathogen to manipulate host cell metabolism and counteract defense responses. This increase aligned with protein quantification within the same families and correlated with the appearance of necrotic lesions on infected plants. Additionally, a few toxin-encoding genes were upregulated in the necrotrophic phase [43].
4.1.2. –Omics Analysis of the Host Plant Response to Highlight Major Defense Pathways Involved
Lupins deploy multiple complementary defense pathways to combat anthracnose. Beyond the initial basal defense line featuring physical barriers against pathogens and the continuous expression and activity of certain general defense genes, a secondary defense layer involves the activation of plant molecular signaling cascades. These cascades are initiated by the recognition of pathogenic molecules, such as microbe or pathogen-associated molecular patterns (PAMPs/MAMPs), triggering PAMPs-triggered immunity (PTI) or by secreted effector proteins, inducing effector-triggered immunity (ETI), a process dependent on the presence of R genes [7,46,47,48,49].
Recently, four lines of narrow-leafed lupin (L. angustifolius L.) exhibiting varying levels of anthracnose resistance and distinct allelic compositions of the two Lanr1/AnMan resistance loci were selected for a differential gene expression profiling of defense responses through transcriptomic analysis. The effective defense observed in Lanr1 gene-mediated resistance involves the coordinated reprogramming of oxidation–reduction, photosynthesis, and pathogenesis-related protein genes shortly after the plant-pathogen contact. A similar defense response mediated by AnMan gene, albeit delayed, proved less effective in protecting against the disease. Anthracnose resistance driven by Lanr1 gene resembles a rapid R gene response (ETI), while the AnMan gene likely imparts a horizontal response (PTI), providing a moderate level of resistance [7].
Bitarishvili et al. [50] analyzed metabolic profiles of two white lupin (L. albus) cultivars in response to primary infection caused by two C. lupini strains isolated from a single Russian field. The results highlighted the importance of nitrogen mobilization in response to C. lupini attack. Fumaric acid and Gamma-aminobutyric acid (GABA) (a ubiquitous four-carbon, non-protein amino acid) were pointed out as potential markers of C. lupini infection.
4.2. Plant and Fungal Secondary Metabolites as Key Regulators of the Pathogenic Interaction
Pathogenic fungi deploy an array of key secondary metabolites crucial for attacking host plants, encompassing polyketides, alkaloids, and terpenes [51]. Transitions between fungal lifestyles involve the orchestrated regulation of various gene families; this regulatory network activates enzymes and metabolites, leading to changes in morphology and infection styles [41,43]. Masi et al. [21] detected and characterized two novel secondary metabolites, lupindolinone and lupinlactone, produced by a strain of C. lupini isolated from symptomatic stems of white lupin (L. albus), demonstrating significant biological activities.
Conversely, plant secondary metabolites assume pivotal roles in the dynamics of plant-pathogen interactions. Terpenoids, flavanols, flavones, and various other compounds, constituting stress-inducible phytochemicals, emerge as key contributors to the development of the plant immune response [52]. Plant secondary metabolites produced in response to C. lupini were investigated in narrow-leafed lupin (L. angustifolius L., NLL) [47,53]. NLL, when pre-exposed to fungal phytotoxic metabolites, demonstrates enhanced production of antifungal compounds in its leaves, suggesting that prior exposure to fungal toxins can potentially bolster resistance against subsequent pathogenic attacks [53].
5. Disease Management through Prophylaxis
5.1. The Utilization of Disease-Free Seeds as a Primary Prophylactic Measure for Mitigating Anthracnose Spread
Presently, the predominant method for minimizing disease spread involves utilizing disease-free seeds, subjected to rigorous phytosanitary controls, with PCR methods employed to quantify seed infections [5,17,54,55]. In Australia, this is complemented by additional seed treatments such as seed dressing with foliar fungicides [5]. In Europe, due to bans on specific fungicides, alternative treatments like dry heat and UV processing have been explored. While these methods reduced germination rates in Andean lupin (L. mutabilis), dry heat and long-term storage have proven successful in narrow-leaved lupins (L. angustifolius L.) [5,6,33,56]. Additionally, various treatments for reducing C. lupini viability in white lupin (L. albus L.), such as hot water, steam, elevated partial pressure of oxygen (EPPO), vinegar, long-term storage, and Biological Control Agents (BCA), were investigated [57]. Results indicated that long-term storage and vinegar reduced infection without significantly impacting germination rates. While BCAs showed promise in controlled conditions, their effectiveness in the field required further research for practical application.
As a seedborne pathogen, C. lupini primarily spreads through infected yet symptomless seeds, underscoring the critical need for phytosanitary measures to prevent future outbreaks, especially involving strains currently restricted to South America [27].
5.2. Enhancing Precision and Efficiency in C. lupini Detection: Advances in Molecular Methodologies and Tools
Some infected seeds may remain asymptomatic while retaining the ability to germinate, thus serving as potential sources of inoculum for the further dissemination of infection. Historically, the microscopic inspection of germinating seeds for fungal features such as appressoria was a laborious and time-consuming process. The development of a robust, rapid, and sensitive technique for detecting C. lupini with minimal damage is pivotal for effective disease management. Molecular assays stand as the predominant diagnostic tools in plant pathology, with nuclear ribosomal cluster genes, including 18S, 5.8S, and 28S, commonly employed as markers for differentiating plant pathogenic fungi. Due to the limitations of Internal Transcribed Spacer (ITS) regions in distinguishing certain Colletotrichum complexes, the polymorphic intergenic spacer (IGS) has been utilized as a molecular diagnostic assay for C. lupini [54]. To enhance the detection sensitivity, an enrichment step involving the incubation of seeds with amended Yeast Malt Broth was introduced to increase the C. lupini biomass in infected seeds. However, this method proved insufficient for quantifying disease severity. Consequently, a quantitative polymerase chain reaction (qPCR)-based assay, as demonstrated by Dubrulle et al. [17], was employed to quantify C. lupini, particularly on asymptomatic plants cultivated at lower temperatures, positioning it as an early detection tool [17]. Subsequently, a TaqMan qPCR assay targeting the Glyceraldehyde 3-phosphate dehydrogenase (GADPH) gene was introduced by Kamber et al. [55], proving effective alternative method to detect C. lupini in lupin plants [57].
6. Insights into the Genetics of Anthracnose Resistance in Domesticated Lupin Species and Implications for Crop Improvement
In the absence of an effective and sustainable disease treatment option [57], breeding for plant host resistance appears to be the most promising solution for the lupine industry. Over the past two decades, substantial efforts have been directed towards identifying genetic sources of resistance, as well as resistance loci and genes. This has encompassed the exploration of diverse germplasm, including cultivars, breeding lines, wild accessions, and mapping populations, while leveraging modern techniques such as high-throughput genotyping through Next-Generation Sequencing (NGS) technologies and genome-wide association studies (GWAS). The different genetic sources of resistance as well as loci and genes known to be involved in anthracnose resistance across the lupin species are summarized in Table 1.
Loci associated with anthracnose resistance in lupins were initially identified in the narrow leaf lupin (L. angustifolius, NLL) using two Australian mapping populations. The first population (D x D) resulted from a cross between two domesticated (D) cultivars, 'Tanjil' (resistant) and 'Unicrop' (susceptible), leading to the development of co-dominant microsatellite-anchored fragment length polymorphism (MFLP) markers for anthracnose resistance associated to the Lanr1 locus [46,61]. The second population (W x D) originated from a cross between the Australian domesticated resistant (D) breeding line '83A:476' and a Moroccan susceptible wild germplasm type (W) named 'P27255’; this cross resulted in enhanced detection of resistant loci [66,67]. Five molecular markers, including two flanking the Lanr1 gene at 0.9 cM, were implemented into breeding programs [68]. Subsequent refinements to the map, utilizing improved marker systems [69,70,71], narrowed down the position of the Lanr1 locus to a 388 Kb fragment containing 41 predicted genes, including an NBS-LRR resistance gene [70]. Another locus, designated AnMan, was identified from a cross between ‘Mandalup’ (resistant) and ‘Quilinock’ (susceptible) cultivars [62]. The resistant alleles (Lanr1 and AnMan) were mapped to distinct positions on the same chromosome, NLL-11. The Lanr1 allele, predominantly present in Australian germplasm, was rare in European germplasm. Its prevalence in Australian germplasm s thought to result from marker-assisted selection (MAS) [7]. While 'Tanjil' and 'Mandalup' showed resistance relative to standards, they proved susceptible in the German environment [32,49,63]. A search for a resistant locus in German cultivars led to the discovery of a single dominant locus named LanrBo [63].
Historically, yellow lupin (L. Luteus) has posed challenges to the breeding industry due to its high susceptibility to anthracnose [72]. Renewed interest in yellow lupins grew, driven by research revealing resistance to diseases such as Pleiochaeta root rot and brown spot, coupled with thriving growth in high rainfall regions, particularly the southwest part of Australia [72,73]. Screening germplasm identified various sources of resistance to anthracnose [72]. 'Gyulatanyai' (P20856), a Hungarian cultivar, exhibited anthracnose resistance in New Zealand, leading to its inclusion in the Australian breeding program. Initially resistant, seven Russian lines experienced an increase in susceptibility with elevated disease pressure and advancing plant age. A Portuguese line from USDA collection, P28716, exhibited resistance despite wild-type traits like hard seeds, pod shattering, late flowering, vernalization requirement, and high alkaloid content. With superior resistance compared to lines from Russia and Hungary, P28716 was selected for breeding, yielding progeny with domesticated traits [72]. A major quantitative trait locus (QTL) for anthracnose resistance, explaining 75% of the phenotypic variance, was identified on L. luteus LG13. This mapping was conducted using an F2 segregating population derived from a cross between the susceptible cultivar AluProt-CGNA, developed and grown in Chile, and the resistant wild accession Core 98 (PI385149). Notably, this major QTL was located in a syntenic region with L. angustifolius (NLL), and marker sequences flanking the QTL exhibited high homology with the Lanr1 locus, suggesting an orthologous gene.
A genetic source for anthracnose resistance in white lupin (L. albus) has been first identified in Ethiopian landraces representing a unique genepool [74,75]. Among these, P27174, P27175, and P27178 stand out as the three most resistant accessions. The recombinant inbred line (RIL) mapping population, derived from the Kiev Mutant×P27174 cross, has historically served as the basis for anthracnose resistance QTL mapping, initially using low marker density [58], and more recently, with a high-density consensus linkage map based on transcriptome-anchored markers [59]. Both studies consistently identified two major QTLs located on linkage groups ALB02 and ALB04, jointly explaining over 40% of the phenotypic variance, and molecular markers associated with P27174 anthracnose resistance loci are available for use in breeding progenies. Notably, the QTLs identified in white lupin did not align with previously mapped narrow-leafed lupin loci conferring anthracnose resistance [59]. Recently, high-throughput phenotyping methods have been developed to efficiently identify field-relevant anthracnose resistance in white lupin [76]. In a recent genome-wide association study (GWAS) involving 181 white lupin accessions from the central regions of domestication and traditional cultivation, 15 highly resistant accessions were identified [60]. The study revealed two significant single nucleotide polymorphisms (SNPs) associated with anthracnose resistance, located on the Lalb_Chr05_g0216161 gene, which encodes a RING zinc-finger E3 ubiquitin ligase and is potentially implicated in plant immunity. The outcomes of this research, offering insights into marker-assisted selection, genomic prediction, and the elucidation of anthracnose resistance mechanisms in white lupin, hold promise for advancing crop improvement.
7. Addressing Technical Challenges to Improve Lupin Biotechnologies for Facilitating Anthracnose-Resistant Variety Breeding
Functional analysis and validation of candidate genes and loci for anthracnose resistance require biotechnological approaches. Additionally, plant biotechnologies, including micropropagation and in vitro regeneration [77], along with advanced gene editing methods, offer attractive opportunities for accelerating breeding processes. Genetic engineering in lupins is challenging due to the crop's recalcitrance to biotechnological methods [78,79]. While genetic transformation has shown some success in all domesticated lupin species, including L. angustifolius [80,81,82], L. luteus [83,84], L. mutabilis [85,86], and more recently in L. albus [79,87,88,89], efficiency rates remain low. Recently, the CRISPR/Cas9 system was implemented to functionally validate the multidrug and toxic compound extrusion (MATE) and aluminum-activated malate transporter (ALMT) genes in white lupin [90,91]. This system was further optimized for a multiplex genome editing approach using Agrobacterium rhizogenes-mediated transformation to study the function of the Lalb_Chr05g0223881 gene, encoding a putative trehalase [92]. These advancements present promising avenues for lupin genetic improvement and breeding for anthracnose resistance.
8. Conclusions & Perspectives
To date, anthracnose is one of the most prevalent challenges that hampered the development of lupin cultivation, despite the great agronomical and ecological interests and economical potential of these legume plants. With a renewed interest in the crop as an alternative to soybean [93,94,95] and coupled with latest technologies, many researchers are now trying to shed more light into the comprehension of the disease. The studies address both the plant perspective [7,35,49,60] and the causative pathogenic agent, C. lupini [2,17,21,27,30,43]. The approaches also encompass the establishment of resources and techniques aimed at comprehending the complexity of this particular plant-pathogen interaction [2,6,14,31,60,76]. The work is catching pace.
A critical objective is to undertake thorough investigations specifically focused on the C. lupini species, alongside extrapolating insights from the C. acutatum complex (CAsc) to which it belongs. Despite the existing wealth of knowledge, including sequenced genomes, -omics data, and phylogenetic diversity, there is a requirement for more comprehensive documentation regarding the species' lifestyle, pathogenicity, and virulence distribution within the species. Elucidating how these factors impact the plant and contribute to the onset of anthracnose is essential for gaining a comprehensive understanding of the pathogenic mechanisms employed by the pathogen. Acquiring such information is also pivotal for the early detection and efficient management of the disease. This knowledge will be particularly beneficial for breeders and farmers seeking sustainable alternatives to reduce dependence on fungicides. Consequently, it can help mitigate operational costs and the environmental impact associated with lupin cultivation.
Developing anthracnose-resistant varieties is the most promising solution to solve this critical problem. Nevertheless, the quest for (partial) resistance against anthracnose proves to be a formidable task, primarily attributed to the intricate nature of the host's response to the disease. This complexity arises from various endogenous factors encompassing metabolic and physiological components, alongside environmental factors that influence the plant host's defense mechanisms against C. lupini. The limitations also include the polygenetic inheritance of the resistance in lupins, despite the presence of loci with major effects.
Whole-genome approaches in lupin, including biparental mapping population analysis, genome-wide association studies (GWAS) and –omics technologies, have been combined towards the discovery of molecular markers, which have been further used in MAS. Nevertheless, our comprehension of the host-pathogen interaction between lupins and C. lupini remains incomplete, particularly concerning the genetic, molecular, and cellular components of the plant's resistance mechanism, including its capacity to detect and respond to C. lupini attacks. As with many legume crops [96], the rate of genetic improvement of lupins has been slow. The introduction in narrow-leaf lupin breeding lines of the lanr1 resistance locus was one of the successes in the advancement of the lupin industry. However, releasing few cultivars carrying an incremental improvement of anthracnose resistance took decades of traditional breeding [74,97], stressing the needs for faster and more efficient breeding processes and technologies. Since the discovery of additional loci in new germplasms, there is a need to re-explore this arena and to functionally analyze the loci, a key step which is still lacking. An urgent task is to establish improved biotechnology methods in lupins, such as the use of genome editing, to study the candidate gene functions and to deliver resistance, building on the validation of genes and pathways that can be modified to improve resistance. While work have been initiated, a considerable amount of efforts remains, and this review delineates both current progress and existing knowledge gaps within this domain.
Author Contributions
Writing—original draft preparation, C.B., L.G. and S.S.; writing—review and editing, E.B., M.R and P.V.; visualization, C.B. and S.S.; supervision, C.B.; project administration, C.B. and E.B.; funding acquisition, C.B., E.B., L.G., and P.V. All authors have read and agreed to the published version of the manuscript.
Funding
This work was financially supported by the Ministry of Science and Higher Education of the Russian Federation as a part of Federal Scientific and Technical Program for the Development of Genetic Technologies for 2019-2027 (Grant № 075-15-2021-1068 of 28.09.2021).
Acknowledgments
We apologize to our colleagues whose contributions could not be referenced due to space constraints.
Conflicts of Interest
The authors declare no conflict of interest. The funders had no role in the writing of the manuscript.
References
- FAOSTAT Food and Agricultural Organization of the United Nations; 2021;
- Guilengue, N.; Silva, M. do C.; Talhinhas, P.; Neves-Martins, J.; Loureiro, A. Subcuticular–Intracellular Hemibiotrophy of Colletotrichum Lupini in Lupinus Mutabilis. Plants 2022, 11. [CrossRef]
- Peres, N.A.; Timmer, L.W.; Adaskaveg, J.E.; Correll, J.C. Lifestyles of Colletotrichum Acutatum. Plant Disease 2005, 89, 784–796. [CrossRef]
- Sweetingham, M.W.; Cowling, W.A.; Buirchell, B.J.; Brown, A.G.P.; Shivas, R.G. Anthracnose of Lupins in Western Australia. Australasian Plant Pathology 1995, 24, 271–271. [CrossRef]
- Shea, G.; Thomas, G.; Buirchell, B.; Salam, M.; McKirdy, S.; Sweetingham, M. Case Study: Industry Response to the Lupin Anthracnose Incursion in Western Australia.; Palta, J.A., Berger, J.B., Ed.; International Lupin Association: Fremantle, Western Australia, September 2008; pp. 425–431.
- Talhinhas, P.; Baroncelli, R.; Le Floch, G. Anthracnose of Lupins Caused by Colletotrichum Lupini : A Recent Disease and a Successful Worldwide Pathogen. Journal of Plant Pathology 2016, 98, 5–14. [CrossRef]
- Książkiewicz, M.; Rychel-Bielska, S.; Plewiński, P.; Bielski, W.; Nuc, M.; Kozak, B.; Krajewski, P.; Jędryczka, M. A Successful Defense of the Narrow-Leafed Lupin against Anthracnose Involves Quick and Orchestrated Reprogramming of Oxidation–Reduction, Photosynthesis and Pathogenesis-Related Genes. Scientific Reports 2022, 12, 8164–8164. [CrossRef]
- Pandey, A.K.; Kumar, A.; Mbeyagala, E.K.; Barbetti, M.J.; Basandrai, A.; Basandrai, D.; Nair, R.M.; Lamichhane, J.R. Anthracnose Resistance in Legumes for Cropping System Diversification. Critical Reviews in Plant Sciences 2023, 42, 177–216. [CrossRef]
- Nirenberg, H.I.; Feiler, U.; Hagedorn, G. Description of Colletotrichum Lupini Comb. Nov. in Modern Terms. Mycologia 2002, 94, 307–320. [CrossRef]
- Sreenivasaprasad, S.; Talhinhas, P. Genotypic and Phenotypic Diversity in Colletotrichum Acutatum, a Cosmopolitan Pathogen Causing Anthracnose on a Wide Range of Hosts. Molecular Plant Pathology 2005, 6, 361–378. [CrossRef]
- Dean, R.; Van Kan, J.A.L.; Pretorius, Z.A.; Hammond-Kosack, K.E.; Di Pietro, A.; Spanu, P.D.; Rudd, J.J.; Dickman, M.; Kahmann, R.; Ellis, J.; et al. The Top 10 Fungal Pathogens in Molecular Plant Pathology. Molecular Plant Pathology 2012, 13, 414–430. [CrossRef]
- Gresta, F.; Wink, M.; Prins, U.; Abberton, M.; Caprato, J.; Scarafoni, A.; Hill, G. Lupins in European Cropping Systems. In Legumes in cropping systems; Murphy-Bokern, D., Stoddard, F.L., Watson, C.A., Eds.; CABI International 2017, 2017; Vol. Chapter 6, pp. 88–108.
- Abreha, K.B.; Ortiz, R.; Carlsson, A.S.; Geleta, M. Understanding the Sorghum–Colletotrichum Sublineola Interactions for Enhanced Host Resistance. Frontiers in Plant Science 2021, 12. [CrossRef]
- Baroncelli, R.; Pensec, F.; Lio, D.D.; Boufleur, T.; Vicente, I.; Sarrocco, S.; Picot, A.; Baraldi, E.; Sukno, S.; Thon, M.; et al. Complete Genome Sequence of the Plant-Pathogenic Fungus Colletotrichum Lupini. Molecular Plant-Microbe Interactions 2021, 34, 1461–1464. [CrossRef]
- Talhinhas, P.; Baroncelli, R. Colletotrichum Species and Complexes: Geographic Distribution, Host Range and Conservation Status; Fungal Diversity; Springer Netherlands, 2021; Vol. 110, p. 198; ISBN 1322502100491.
- Msairi, S.; Chliyeh, M.; Touhami, A.O.; Abdelaziz, M.; Alaoui, E. FIRST REPORT OF Colletotrichum Lupini CAUSING ANTHRACNOSE DISEASE ON THE OLIVE FRUITS IN MOROCCO; Article in PLANT CELL BIOTECHNOLOGY AND MOLECULAR BIOLOGY; 2020;
- Dubrulle, G.; Pensec, F.; Picot, A.; Rigalma, K.; Pawtowski, A.; Nicolleau, S.; Harzic, N.; Nodet, P.; Baroncelli, R.; Le Floch, G. Phylogenetic Diversity and Effect of Temperature on Pathogenicity of Colletotrichum Lupini. Plant Disease 2020, 104, 938–950. [CrossRef]
- Talhinhas, P.; Sreenivasaprasad, S.; Neves-Martins, J.; Oliveira, H. Genetic and Morphological Characterization of Colletotrichum Acutatum Causing Anthracnose of Lupins. Phytopathology® 2002, 92, 986–996. [CrossRef]
- Lotter, H.C.; Berger, D.K. Anthracnose of Lupins in South Africa Is Caused by Colletotrichum Lupini Var. Setosum. Australasian Plant Pathology 2005, 34, 385–392. [CrossRef]
- Jayawardena, R.S.; Hyde, K.D.; Damm, U.; Cai, L.; Liu, M.; Li, X.H.; Zhang, W.; Zhao, W.S.; Yan, J.Y. Notes on Currently Accepted Species of Colletotrichum. Mycosphere 2016, 7, 1192–1260. [CrossRef]
- Masi, M.; Nocera, P.; Boari, A.; Zonno, M.C.; Pescitelli, G.; Sarrocco, S.; Baroncelli, R.; Vannacci, G.; Vurro, M.; Evidente, A. Secondary Metabolites Produced by Colletotrichum Lupini, the Causal Agent of Anthachnose of Lupin (Lupinus Spp.). Mycologia 2020, 112, 533–542. [CrossRef]
- Baroncelli, R.; Talhinhas, P.; Pensec, F.; Sukno, S.A.; Floch, G.L.; Thon, M.R. The Colletotrichum Acutatum Species Complex as a Model System to Study Evolution and Host Specialization in Plant Pathogens. Frontiers in Microbiology 2017, 8. [CrossRef]
- Damm, U.; Cannon, P.F.; Woudenberg, J.H.C.; Crous, P.W. The Colletotrichum Acutatum Species Complex. Studies in Mycology 2012, 73, 37–113. [CrossRef]
- Bhunjun, C.S.; Phukhamsakda, C.; Jayawardena, R.S.; Jeewon, R.; Promputtha, I.; Hyde, K.D. Investigating Species Boundaries in Colletotrichum. Fungal Diversity 2021, 107, 107–127. [CrossRef]
- Chen, Y. peng; Wu, T.; Tian, W.H.; Ilyukhin, E.; Hyde, K.D.; Maharachchikumbura S. S. N. Comparative Genomics Provides New Insights into the Evolution of Colletotrichum. Mycosphere 2022, 13, 134–187. [CrossRef]
- Lardner, R.; Johnston, P.R.; Plummer, K.M.; Pearson, M.N. Morphological and Molecular Analysis of Colletotrichum Acutatum Sensu Lato. Mycological Research 1999, 103, 275–285. [CrossRef]
- Alkemade, J.A.; Baroncelli, R.; Messmer, M.M.; Hohmann, P. Attack of the Clones: Population Genetics Reveals Clonality of Colletotrichum Lupini, the Causal Agent of Lupin Anthracnose. Molecular Plant Pathology 2023. [CrossRef]
- Yang, H.A.; Sweetingham, M.W. The Taxonomy of Colletotrichum Isolates Associated with Lupin Anthracnose. Australian Journal of Agricultural Research 1998, 49, 1213–1213. [CrossRef]
- Riegel, R.; Véliz, D.; Von Baer, I.; Quitral, Y.; Muñoz, M. Genetic Diversity and Virulence of Colletotrichum Lupini Isolates Collected in Chile. Tropical Plant Pathology 2010, 35, 144–152. [CrossRef]
- Alkemade, J.A.; Messmer, M.M.; Voegele, R.T.; Finckh, M.R.; Hohmann, P. Genetic Diversity of Colletotrichum Lupini and Its Virulence on White and Andean Lupin. Scientific Reports 2021, 11. [CrossRef]
- Pszczółkowska, A.; Okorski, A.; Jastrzębski, J.P.; Paukszto; Fordoński, G. The Complete Mitogenome of Colletotrichum Lupini Var. Setosum. Mitochondrial DNA Part B: Resources 2016, 1, 37–38. [CrossRef]
- Thomas, G.J.; Sweetingham, M.W.; Yang, H.A.; Speijers, J. Effect of Temperature on Growth of Colletotrichum Lupini and on Anthracnose Infection and Resistance in Lupins. Australasian Plant Pathology 2008, 37, 35–35. [CrossRef]
- Falconí, C.E.; Yánez–Mendizábal, V. Dry Heat Treatment of Andean Lupin Seed to Reduce Anthracnose Infection. Crop Protection 2016, 89, 178–183. [CrossRef]
- Falconí, C.E.; Yánez-Mendizábal, V. Available Strategies for the Management of Andean Lupin Anthracnose. Plants 2022, 11. [CrossRef]
- Falconí, C.E.; Visser, R.G.F.; Van Heusden, S. Influence of Plant Growth Stage on Resistance to Anthracnose in Andean Lupin (Lupinus Mutabilis). Crop and Pasture Science 2015, 66, 729–734. [CrossRef]
- Diggle, A.J.; Salam, M.U.; Thomas, G.J.; Yang, H.A.; O’connell, M.; Sweetingham, M.W. Analytical and Theoretical Plant Pathology AnthracnoseTracer: A Spatiotemporal Model for Simulating the Spread of Anthracnose in a Lupin Field; 2002;
- Semaškienė, R.; Brazauskienė, I.; Lisova, R.; Liepienė, N.; Maknickienė, Z. The Incidence of Anthracnose (Colletotrichum Spp.) on Lupine Seed. Zemdirbyste-Agriculture 2008, 144–150.
- Thomas, G.J.; Sweetingham, M.W. Cultivar and Environment Influence the Development of Lupin Anthracnose Caused by Colletotrichum Lupini. Australasian Plant Pathology 2004, 33, 571–577. [CrossRef]
- Perfect, S.E.; Bleddyn Hughes, H.; O’connell, R.J.; Green, J.R. Colletotrichum: A Model Genus for Studies on Pathology and Fungal-Plant Interactions; 1999. [CrossRef]
- Cannon, P.F.; Damm, U.; Johnston, P.R.; Weir, B.S. Colletotrichum - Current Status and Future Directions. Studies in Mycology 2012, 73, 181–213. [CrossRef]
- O’Connell, R.J.; Thon, M.R.; Hacquard, S.; Amyotte, S.G.; Kleemann, J.; Torres, M.F.; Damm, U.; Buiate, E.A.; Epstein, L.; Alkan, N.; et al. Lifestyle Transitions in Plant Pathogenic Colletotrichum Fungi Deciphered by Genome and Transcriptome Analyses. Nature Genetics 2012, 44, 1060–1065. [CrossRef]
- De Silva, D.D.; Crous, P.W.; Ades, P.K.; Hyde, K.D.; Taylor, P.W.J. Life Styles of Colletotrichum Species and Implications for Plant Biosecurity. Fungal Biology Reviews 2017, 31, 155–168. [CrossRef]
- Dubrulle, G.; Picot, A.; Madec, S.; Corre, E.; Pawtowski, A.; Baroncelli, R.; Zivy, M.; Balliau, T.; Le Floch, G.; Pensec, F. Deciphering the Infectious Process of Colletotrichum Lupini in Lupin through Transcriptomic and Proteomic Analysis. Microorganisms 2020, 8, 1621–1621. [CrossRef]
- Diéguez-Uribeondo, J.; Förster, H.; Adaskaveg, J.E. Digital Image Analysis of Internal Light Spots of Appressoria of Colletotrichum Acutatum; 2003; Vol. 93, pp. 923–923;.
- Diéguez-Uribeondo, J.; Förster, H.; Soto-Estrada, A.; Adaskaveg, J.E. Subcuticular-Intracellular Hemibiotrophic and Intercellular Necrotrophic Development of Colletotrichum Acutatum on Almond. Phytopathology® 2005, 95, 751–758. [CrossRef]
- You, M.; Buirchell, B.; M Siddique, K.H.; Sweetingham, M. A PCR-Based Molecular Marker Applicable for Marker-Assisted Selection for Anthracnose Disease Resistance in Lupin Breeding. Cellular & Molecular Biology Letters 2005, 10, 123–134.
- Wojakowska, A.; Muth, D.; Narozna, D.; Mądrzak, C.; Stobiecki, M.; Kachlicki, P. Changes of Phenolic Secondary Metabolite Profiles in the Reaction of Narrow Leaf Lupin (Lupinus Angustifolius) Plants to Infections with Colletotrichum Lupini Fungus or Treatment with Its Toxin. Metabolomics 2013, 9, 575–589. [CrossRef]
- Yang, H.; Tao, Y.; Zheng, Z.; Shao, D.; Li, Z.; Sweetingham, M.W.; Buirchell, B.J.; Li, C. Rapid Development of Molecular Markers by Next-Generation Sequencing Linked to a Gene Conferring Phomopsis Stem Blight Disease Resistance for Marker-Assisted Selection in Lupin (Lupinus Angustifolius L.) Breeding. Theoretical and Applied Genetics 2013, 126, 511–522. [CrossRef]
- Fischer, K.; Dieterich, R.; Nelson, M.N.; Kamphuis, L.G.; Singh, K.B.; Rotter, B.; Krezdorn, N.; Winter, P.; Wehling, P.; Ruge-Wehling, B. Characterization and Mapping of LanrBo: A Locus Conferring Anthracnose Resistance in Narrow-Leafed Lupin (Lupinus Angustifolius L.). Theoretical and Applied Genetics 2015, 128, 2121–2130. [CrossRef]
- Bitarishvili, S.; Samad, S.; Boldyrev, S.; Ben, C.; Volkova, P.; Shavarda, A.; Lukashevich, M.; Bondarenko, E. Metabolic Profiling Reveals Fumaric Acid and GABA as Possible Markers of Colletotrichum Lupini Infection of White Lupin. Physiological and Molecular Plant Pathology 2023, 128. [CrossRef]
- Moraga, J.; Gomes, W.; Pinedo, C.; Cantoral, J.M.; Hanson, J.R.; Carbú, M.; Garrido, C.; Durán-Patrón, R.; Collado, I.G. The Current Status on Secondary Metabolites Produced by Plant Pathogenic Colletotrichum Species. Phytochemistry Reviews 2019, 18, 215–239. [CrossRef]
- Anjali; Kumar, S.; Korra, T.; Thakur, R.; Arutselvan, R.; Kashyap, A.S.; Nehela, Y.; Chaplygin, V.; Minkina, T.; Keswani, C. Role of Plant Secondary Metabolites in Defence and Transcriptional Regulation in Response to Biotic Stress. Plant Stress 2023, 8, 100154. [CrossRef]
- Wojakowska, A.; Kułak, K.; Jasiński, M.; Kachlicki, P.; Stawiński, S.; Stobiecki, M. Metabolic Response of Narrow Leaf Lupine (Lupinus Angustifolius) Plants to Elicitation and Infection with Colletotrichum Lupini under Field Conditions. Acta Physiologiae Plantarum 2015, 37. [CrossRef]
- Pecchia, S.; Caggiano, B.; Da Lio, D.; Cafà, G.; Le Floch, G.; Baroncelli, R. Molecular Detection of the Seed-Borne Pathogen Colletotrichum Lupini Targeting the Hyper-Variable IGS Region of the Ribosomal Cluster. Plants 2019, 8, 4–6. [CrossRef]
- Kamber, T.; Malpica-López, N.; Messmer, M.M.; Oberhänsli, T.; Arncken, C.; Alkemade, J.A.; Hohmann, P. A Qpcr Assay for the Fast Detection and Quantification of Colletotrichum Lupini. Plants 2021, 10, 2–9. [CrossRef]
- Falconí, C.E.; Yánez-Mendizábal, V. Efficacy of UV-C Radiation to Reduce Seedborne Anthracnose (Colletotrichum Acutatum) from Andean Lupin (Lupinus Mutabilis). Plant Pathology 2018, 67, 831–838. [CrossRef]
- Alkemade, J.A.; Arncken, C.; Hirschvogel, C.; Messmer, M.M.; Leska, A.; Voegele, R.T.; Finckh, M.R.; Kölliker, R.; Groot, S.P.C.; Hohmann, P. The Potential of Alternative Seed Treatments to Control Anthracnose Disease in White Lupin. Crop Protection 2022, 158. [CrossRef]
- Phan, H.T.T.; Ellwood, S.R.; Adhikari, K.; Nelson, M.N.; Oliver, R.P. The First Genetic and Comparative Map of White Lupin (Lupinus Albus L.): Identification of QTLs for Anthracnose Resistance and Flowering Time, and a Locus for Alkaloid Content. DNA Research 2007, 14, 59–70. [CrossRef]
- Książkiewicz, M.; Nazzicari, N.; Yang, H.; Nelson, M.N.; Renshaw, D.; Rychel, S.; Ferrari, B.; Carelli, M.; Tomaszewska, M.; Stawiński, S.; et al. A High-Density Consensus Linkage Map of White Lupin Highlights Synteny with Narrow-Leafed Lupin and Provides Markers Tagging Key Agronomic Traits. Scientific Reports 2017, 7, 15335–15335. [CrossRef]
- Alkemade, J.A.; Nazzicari, N.; Messmer, M.M.; Annicchiarico, P.; Ferrari, B.; Voegele, R.T.; Finckh, M.R.; Arncken, C.; Hohmann, P. Genome-Wide Association Study Reveals White Lupin Candidate Gene Involved in Anthracnose Resistance. Theoretical and Applied Genetics 2022, 135, 1011–1024. [CrossRef]
- Yang, H.; Boersma, J.G.; You, M.; Buirchell, B.J.; Sweetingham, M.W. Development and Implementation of a Sequence-Specific PCR Marker Linked to a Gene Conferring Resistance to Anthracnose Disease in Narrow-Leafed Lupin (Lupinus Angustifolius L.). Molecular Breeding 2004, 14, 145–151. [CrossRef]
- Yang, H.; Renshaw, D.; Thomas, G.; Buirchell, B.; Sweetingham, M. A Strategy to Develop Molecular Markers Applicable to a Wide Range of Crosses for Marker Assisted Selection in Plant Breeding: A Case Study on Anthracnose Disease Resistance in Lupin (Lupinus Angustifolius L.). Molecular Breeding 2008, 21, 473–483. [CrossRef]
- Ruge-Wehling, B.; Dieterich, R.; Thiele, C.; Eickmeyer, F.; Wehling, P. Resistance to Anthracnose in Narrow-Leafed Lupin (Lupinus Angustifolius L.): Sources of Resistance and Development of Molecular Markers. Journal Für Kulturpflanzen 2009, 61, 62–65.
- Lichtin, N.; Salvo-Garrido, H.; Till, B.; Caligari, P.D.S.; Rupayan, A.; Westermeyer, F.; Olivos, M. Genetic and Comparative Mapping of Lupinus Luteus L. Highlight Syntenic Regions with Major Orthologous Genes Controlling Anthracnose Resistance and Flowering Time. Scientific Reports 2020, 10. [CrossRef]
- Guilengue, N.; Neves-Martins, J.; Talhinhas, P. Response to Anthracnose in a Tarwi (Lupinus Mutabilis) Collection Is Influenced by Anthocyanin Pigmentation. Plants 2020, 9. [CrossRef]
- Boersma, J.G.; Pallotta, M.; Li, C.D.; Buirchell, B.J.; Sivasithamparam, K.; Yang Huaan Construction of a Genetic Linkage Map Using MFLP and Identification of Molecular Markers Linked to Domestication Genes in Narrow-Leafed Lupin (Lupinus Angustifolius L.). Cell. Mol. Biol. Lett. 2005, 10, 331–344.
- Nelson, M.N.; Phan, H.T.T.; Ellwood, S.R.; Moolhuijzen, P.M.; Hane, J.; Williams, A.; O’Lone, C.E.; Fosu-Nyarko, J.; Scobie, M.; Cakir, M.; et al. The First Gene-Based Map of Lupinus Angustifolius L. -Location of Domestication Genes and Conserved Synteny with Medicago Truncatula. Theoretical and Applied Genetics 2006, 113, 225–238. [CrossRef]
- Yang, H.; Tao, Y.; Zheng, Z.; Li, C.; Sweetingham, M.W.; Howieson, J.G. Application of Next-Generation Sequencing for Rapid Marker Development in Molecular Plant Breeding: A Case Study on Anthracnose Disease Resistance in Lupinus Angustifolius L. BMC Genomics 2012, 13, 318–318. [CrossRef]
- Kamphuis, L.G.; Hane, J.K.; Nelson, M.N.; Gao, L.; Atkins, C.A.; Singh, K.B. Transcriptome Sequencing of Different Narrow-leafed Lupin Tissue Types Provides a Comprehensive Uni-gene Assembly and Extensive Gene-based Molecular Markers. Plant Biotechnology Journal 2015, 13, 14–25. [CrossRef]
- Hane, J.K.; Ming, Y.; Kamphuis, L.G.; Nelson, M.N.; Garg, G.; Atkins, C.A.; Bayer, P.E.; Bravo, A.; Bringans, S.; Cannon, S.; et al. A Comprehensive Draft Genome Sequence for Lupin (Lupinus Angustifolius), an Emerging Health Food: Insights into Plant–Microbe Interactions and Legume Evolution. Plant Biotechnology Journal 2017, 15, 318–330. [CrossRef]
- Plewiński, P.; Książkiewicz, M.; Rychel-Bielska, S.; Rudy, E.; Wolko, B. Candidate Domestication-Related Genes Revealed by Expression Quantitative Trait Loci Mapping of Narrow-Leafed Lupin (Lupinus Angustifolius L.). International Journal of Molecular Sciences 2019, 20. [CrossRef]
- Adhikari, K.N.; Thomas, G.; Buirchell, B.J.; Sweetingham, M.W. Identification of Anthracnose Resistance in Yellow Lupin (Lupinus Luteus L.) and Its Incorporation into Breeding Lines. Plant Breeding 2011, 130, 660–664. [CrossRef]
- French, R.J.; Sweetingham, M.W.; Shea, G.G. A Comparison of the Adaptation of Yellow Lupin (Lupinus Luteus L.) and Narrow-Leafed Lupin (L. Angustifolius L.) to Acid Sandplain Soils in Low Rainfall Agricultural Areas of Western Australia. Australian Journal of Agricultural Research 2001, 52, 945–954. [CrossRef]
- Adhikari, K.N.; Buirchell, B.J.; Thomas, G.J.; Sweetingham, M.W.; Yang, H. Identification of Anthracnose Resistance in Lupinus Albus L. and Its Transfer from Landraces to Modern Cultivars. Crop and Pasture Science 2009, 60, 472–479. [CrossRef]
- Raman, R.; Cowley, R.B.; Raman, H.; Luckett, D.J. Analyses Using SSR and DArT Molecular Markers Reveal That Ethiopian Accessions of White Lupin (Lupinus Albus L.) Represent a Unique Genepool. Open Journal of Genetics 2014, 04, 87–98. [CrossRef]
- Alkemade, J.A.; Messmer, M.M.; Arncken, C.; Leska, A.; Annicchiarico, P.; Nazzicari, N.; Książkiewicz, M.; Voegele, R.T.; Finckh, M.R.; Hohmann, P. A High-Throughput Phenotyping Tool to Identify Field-Relevant Anthracnose Resistance in White Lupin. Plant Disease 2021, 105, 1719–1727. [CrossRef]
- Mulin, M.; Bellio-Spataru, A. Organogenesis from Hypocotyl Thin Cell Layers of Lupinus Mutabilis and Lupinus Albus; Plant Growth Regulation; 2000; Vol. 30, pp. 177–183;.
- Nguyen, A.H.; Hodgson, L.M.; Erskine, W.; Barker, S.J. An Approach to Overcoming Regeneration Recalcitrance in Genetic Transformation of Lupins and Other Legumes. Plant Cell, Tissue and Organ Culture 2016, 127, 623–635. [CrossRef]
- Aslam, M.M.; Karanja, J.K.; Zhang, Q.; Lin, H.; Xia, T.; Akhtar, K.; Liu, J.; Miao, R.; Xu, F.; Xu, W. In Vitro Regeneration Potential of White Lupin (Lupinus Albus) from Cotyledonary Nodes. Plants 2020, 9, 1–14. [CrossRef]
- Molvig, L.; Tabe, L.M.; Eggum, B.O.; Moore, A.E.; Craig, S.; Spencer, D.; Higgins, T.J.V. Enhanced Methionine Levels and Increased Nutritive Value of Seeds of Transgenic Lupins ( Lupinus Angustifolius L.) Expressing a Sunflower Seed Albumin Gene. Proceedings of the National Academy of Sciences 1997, 94, 8393–8398. [CrossRef]
- Pigeaire, A.; Abernethy, D.; Smith, P.M.; Simpson, K.; Fletcher, N.; Lu, C.-Y.; Atkins, C.A.; Cornish, E. Transformation of a Grain Legume (Lupinus Angustifolius L.) via Agrobacterium Tumefaciens-Mediated Gene Transfer to Shoot Apices; Molecular Breeding; Kluwer Academic Publishers, 1997; Vol. 3, pp. 341–349;.
- Barker, S.J.; Si, P.; Hodgson, L.; Ferguson-Hunt, M.; Khentry, Y.; Krishnamurthy, P.; Averis, S.; Mebus, K.; O’Lone, C.; Dalugoda, D.; et al. Regeneration Selection Improves Transformation Efficiency in Narrow-Leaf Lupin. Plant Cell, Tissue and Organ Culture 2016, 126, 219–228. [CrossRef]
- Li, H.; Wylie, S.J.; Jones, M.G.K. Transgenic Yellow Lupin (Lupinus Luteus); Plant Cell Reports; Springer-Verlag, 2000; Vol. 19, pp. 634–637;.
- Pniewski, T.; Kapusta, J.; Płucienniczak, A. Agrobacterium-Mediated Transformation of Yellow Lupin to Generate Callus Tissue Producing HBV Surface Antigen in a Long-Term Culture. Journal of Applied Genetics 2006, 47, 309–318. [CrossRef]
- Babaoglu, M.; Mccabe, M.S.; Power, J.B.; Dave, M.R. Agrobacterium-Mediated Transformation of Lupinus Mutabilis L. Using Shoot Apical Explants; ACTA PHYSIOLOGIAE PLANTARUM; 2000; Vol. 22, pp. 111–119;.
- Polowick, P.L.; Loukanina, N.N.; Doshi, K.M. Agrobacterium-Mediated Transformation of Tarwi (Lupinus Mutabilis Sweet), a Potential Platform for the Production of Plant-Made Proteins. In Vitro Cellular & Developmental Biology - Plant 2014, 50, 401–411. [CrossRef]
- Xu, W.; Zhang, Q.; Yuan, W.; Xu, F.; Muhammad Aslam, M.; Miao, R.; Li, Y.; Wang, Q.; Li, X.; Zhang, X.; et al. The Genome Evolution and Low-Phosphorus Adaptation in White Lupin. Nature Communications 2020, 11. [CrossRef]
- Aslam, M.M.; Waseem, M.; Zhang, Q.; Ke, W.; Zhang, J.; Xu, W. Identification of ABC Transporter G Subfamily in White Lupin and Functional Characterization of L.albABGC29 in Phosphorus Use. BMC Genomics 2021, 22. [CrossRef]
- Aslam, M.M.; Fritschi, F.B.; Zhang, D.; Wang, G.; Li, H.; Lam, H.; Waseem, M.; Xu, W.; Zhang, J. Overexpression of LaGRAS Enhances Phosphorus Acquisition via Increased Root Growth of Phosphorus-deficient White Lupin. Physiologia Plantarum 2023. [CrossRef]
- Zhou, Y.; Olt, P.; Neuhäuser, B.; Moradtalab, N.; Bautista, W.; Uhde-Stone, C.; Neumann, G.; Ludewig, U. Loss of LaMATE Impairs Isoflavonoid Release from Cluster Roots of Phosphorus-Deficient White Lupin. Physiologia Plantarum 2021, 173, 1207–1220. [CrossRef]
- Zhou, Y.; Neuhäuser, B.; Neumann, G.; Ludewig, U. LaALMT1 Mediates Malate Release from Phosphorus-Deficient White Lupin Root Tips and Metal Root to Shoot Translocation. Plant Cell and Environment 2020, 43, 1691–1706. [CrossRef]
- Zhu, X.; Xu, W.; Liu, B.; Zhan, Y.; Xia, T. Adaptation of High-Efficiency CRISPR/Cas9-Based Multiplex Genome Editing System in White Lupin by Using Endogenous Promoters. Physiologia Plantarum 2023, 175. [CrossRef]
- Monteiro, A.; Miranda, C.; Trindade, H. Mediterranean Lupines as an Alternative Protein Source to Soybean.; MDPI: Basel Switzerland, May 2021; pp. 38–38.
- Szczepański, A.; Adamek-Urbańska, D.; Kasprzak, R.; Szudrowicz, H.; Śliwiński, J.; Kamaszewski, M. Lupin: A Promising Alternative Protein Source for Aquaculture Feeds? Aquaculture Reports 2022, 26. [CrossRef]
- Gresta, F.; Oteri, M.; Scordia, D.; Costale, A.; Armone, R.; Meineri, G.; Chiofalo, B. White Lupin (Lupinus Albus L.), an Alternative Legume for Animal Feeding in the Mediterranean Area. Agriculture (Switzerland) 2023, 13. [CrossRef]
- Foyer, C.H.; Lam, H.-M.; Nguyen, H.T.; Siddique, K.H.M.; Varshney, R.K.; Colmer, T.D.; Cowling, W.; Bramley, H.; Mori, T.A.; Hodgson, J.M.; et al. Neglecting Legumes Has Compromised Human Health and Sustainable Food Production. Nature Plants 2016, 2, 16112. [CrossRef]
- Adhikari, K.N.; Thomas, G.; Diepeveen, D.; Trethowan, R. Overcoming the Barriers of Combining Early Flowering and Anthracnose Resistance in White Lupin (Lupinus Albus L.) for the Northern Agricultural Region of Western Australia. Crop and Pasture Science 2013, 64, 914–921. [CrossRef]
Figure 1.
Colletotrichum lupini, the pathogenic agent responsible for anthracnose in lupins, can impact lupin plants throughout all developmental stages. (a) Morphologically diverse strains of C. lupini cultivated under dark conditions on potato dextrose agar (PDA). (b-f) Manifestation of characteristic anthracnose symptoms induced by C. lupini observed on distinct white lupin organs, including leaf curling (b), stem necrotic lesions (c), pod (d) and seed (e) wrinkling and necrosis, and seedling radical necrosis with observable signs of twisting (f).
Figure 1.
Colletotrichum lupini, the pathogenic agent responsible for anthracnose in lupins, can impact lupin plants throughout all developmental stages. (a) Morphologically diverse strains of C. lupini cultivated under dark conditions on potato dextrose agar (PDA). (b-f) Manifestation of characteristic anthracnose symptoms induced by C. lupini observed on distinct white lupin organs, including leaf curling (b), stem necrotic lesions (c), pod (d) and seed (e) wrinkling and necrosis, and seedling radical necrosis with observable signs of twisting (f).


Table 1.
Summary of current knowledge on genetic sources and genomic loci involved in anthracnose resistance across domesticated lupin species. NA: Non available.
Table 1.
Summary of current knowledge on genetic sources and genomic loci involved in anthracnose resistance across domesticated lupin species. NA: Non available.
| Lupin species | Source for resistance (Cultivar / Line) | Origin | Locus/ candidate gene | Reference |
|
Lupinus albus (White lupin) |
P27174 P27175 P27178 Blu-25 |
Ethiopia Ethiopia Ethiopia Chile |
2 QTLs on ALB02 and ALB04 linkage groups NA NA Lalb_Chr05_g0216161 |
[58,59,60] |
| Rumbo Baer(a) | Chile | “ | ||
| Rumbo Baer(b) | Chile | “ | ||
|
Lupinus angustifolius (Narrow-leaved lupine) |
Tanjil | Australia | Lanr1 | [43,61,62,63] |
| Wonga Mandelup Bo7212 |
Australia Australia Germany |
Lanr1 AnMan LanrBo |
||
|
Lupinus luteus (Yellow lupin) |
Core 98 | Chile | Lanr1 homolog | [64,72] |
| Gyulatanya P20856 | Hungary | NA | ||
| P28716 | Portugese | NA | ||
|
Lupinus mutabilis (Andean lupin) |
LM34 | Chile | NA | [65] |
| I82 | Chile | NA |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



