
Submitted:
27 December 2023
Posted:
28 December 2023
You are already at the latest version
Abstract
We investigated by Real-time PCR and end point PCR the presence of 10 potential zoonotic viruses in 91 small mammals sold as pets in pet shops of Southern Italy. The animals were analysed for: aichivirus, sapovirus, astrovirus, hepatitis A virus, noroviruses (GI and GII), rotavirus, circovirus, coronaviruses. Among the 9 species of small mammals analysed (golden hamsters, Java squirrels, mongolian gerbils, peruvian guinea pigs, pet mice, pet rats, dwarf rabbits, african hedgehogs and sugar gliders) 12 pet rats, 11 pet mice and 1 golden hamster tested positive to a PCR amplifying a conserved gene (RdRp) of coronavirus. However further characterizaton by sequencing gave not conclusive results. A pet rat resulted instead positive to aichivirus and its sequence showed similarity with a murine kobuvirus-1 strain identified in China. To our knowledge, this is the first study reporting the detection of aichivirus in rodents in Italy, the virus probably introduced through the importation of animals from Asia.
Since living in close contact to humans, expecially with children, the continuous monitoring of the small mammals is very important to avoid the spread of new pathologies among animals and humans and further investigation is desirable in order to safeguard public health.
Keywords:
Aichivirus
; Rattus norvegicus
; non conventional pets
; small mammals
; kobuvirus
1. Introduction
During the last decades, the global demand for non conventional pets has greatly increased. It has been calculated that only in Italy there are at least about 30 millions of ornamental fishes, 1.4 millions of reptiles, 12.9 millions of ornamental birds and almost 1.8 million of small mammals like chinchillas, guinea pigs, ferrets, prairie dog, meerkats, golden hamsters, Java squirrels, mongolian gerbils, peruvian guinea pigs, pet mice, pet rats, dwarf rabbits, african hedgehogs, sugar gliders, etc. [Boros et al. 2019, Chomel et al. 2007, d'Ovidio & Santoro 2023, Firth et al. 2014, Riley et al.2005, Haake et al. 2020]. The close social interaction between animals and humans provide a strong motivation to study the virome composition of unconventional pets, as these animals could represent a relevant source of infection due to little-known or even unknown virus agents [Chan et al. 2020, Chomel et al. 2007, Mira et al. 2018]. Furthermore, the illegal trafficking of companion animals contributes to increase the risk of introducing in free areas new pathogens transmissible to other animals and/or humans. As a matter of fact, hundreds of pets are destined for the black market whose turnover is second only to drug trafficking (Zoomafia Observatory Anti-Vivisection League). Animals purchased on the black market are indeed not subjected to any health check and may come from endemic areas [Alfano et al. 2019, Alfano et al. 2020, Firth et al. 2014]. Open borders and a constant increase in human and animal movement could trigger the rapid emergence and spread of new pathogens on a global scale, as recently happened for SARS-CoV-2, so the identification of new viral agents, especially in animal species in close contact to humans, is of fundamental importance for the assessment of the zoonotic risk.
Several pathogens, which were considered to be pathogens exclusively to animals, have instead shown zoonotic potential [Chomel 2007, Reperant 2016]. In recent years, the trend is definitely growing. For instance, according to Johnson [2020], at least 142 animal viruses could be able to cause diseases in humans, not least SARS-CoV-2, responsible for the recent COVID-19 pandemic. A more complete characterization of the viruses became recently possible thanks to the use of innovative molecular diagnostic techniques and to the development of a metagenomic approach. As a result, new viral agents and new genetic lineages of already known viruses (rotaviruses as an example ) were discovered and for some of them zoonotic potential was detected [Matthijnssens 2006]. The aim of our work was to study the circulation, by molecular assay, of some potential zoonotic viruses like aichivirus, sapovirus, astrovirus, hepatitis A virus, noroviruses (GI and GII), rotavirus and circovirus, in non-conventional pets imported in Italy from various EU and non-EU countries in order to expand the limited knowlwdge currently available on this topic.
Aichiviruses (AiVs), belong to the Kobuvirus genus of the Picornaviridae family. Kobuviruses were associated with gastrointestinal diseases and fecal-oral infection, not only in humans, but also in dogs, cats, cattle and pigs [Abi et al. 2022, Bergallo 2017, Diakoudi et al. 2020, Kaikkonen 2010].
Astrovirus (AsV) is single-stranded, non-enveloped RNA virus belonging to the Astroviridae family. There are 8 known serotypes, of which serotype 1 (AsV) is the most widespread and mainly associated with episodes of gastroenteritis in humans and children [Le Cann et al. 2004].
Noroviruses are non-enveloped single-stranded RNA viruses belonging to the Caliciviridae family, they are very widespread viruses causing highly contagious gastroenteritis, with GI and GII the genogroups more frequently involved in gastroenteritis in humans [Chhabra 2019].
Sapovirus (SaV), another member of the Caliciviridae family is characterized by a single-stranded RNA genome. It has been classified in 5 known genogroups, identified on the basis of the complete sequence of the capsid; genogroups I, II, IV and V include human strains while genogroup III has been identified only in pigs. Sapovirus is an etiological agent responsible for human gastroenteritis, especially in children and it is considered an emerging contaminant that poses a risk to public health due to its presence in aquatic environments [Varela et al. 2016].
Rotavirus (RV) belonging to the Reoviridae family is characterized by a segmented double-stranded RNA genome. Nine groups/species (A-I) and a further tentative group J [Banyai et al., 2017]. Serogroups (A-G) are known, of which serogroup A is the one mainly associated with episodes of human gastroenteritis, causing severe symptoms in children [Zeng et al. 2008].
Hepatitis A virus (HAV), small non-enveloped virus characterized by a single-stranded RNA genome, belonging to the Enterovirus genus, Picornaviridae family. It is a very persistent virus in the environment and causes hepatitis and serious liver damage in humans [Costafreda 2006].
Circovirus (PCV) is a small non-enveloped icosahedral single-stranded DNA virus, member of the Circoviridae family and the Circovirus genus. There are currently 11 species of known circoviruses that infect a wide variety of birds and mammals [Kim et al. 2017].
Apart from the above named viruses, we posed our attention also on the search of coronaviruses. Coronavirus (CoV), order Nidovirales, family Coronaviridae, is characterized by single-stranded RNA genome with positive polarity, they are responsible for enteric and/or respiratory diseases in a large variety of animals. Their ability to evolve giving rise to new viral genotypes and to overcome the species barrier infecting new hosts is also well known in the humans, where some coronaviruses, identified in recent years, were of zoonotic origin.
After the emergencies related to SARS [Drosten et al. 2003] and MERS [Chafekar 2018] and the subsequent recent pandemic (COVID-19), the international scientific community received a further boost to research activities on coronaviruses [Dakroub et al. 2022 ].
2. Materials and Methods
Animals investigated
In the period 2021–2022 the organs (637) of 91 small mammals, kept as pets in Italy and mostly died from unknown causes, were analyzed for the presence of various zoonotic viruses by the Istituto Zooprofilattico Sperimentale del Mezzogiorno (IZSM), during the activities of the Project IZS ME 04/19 RC.
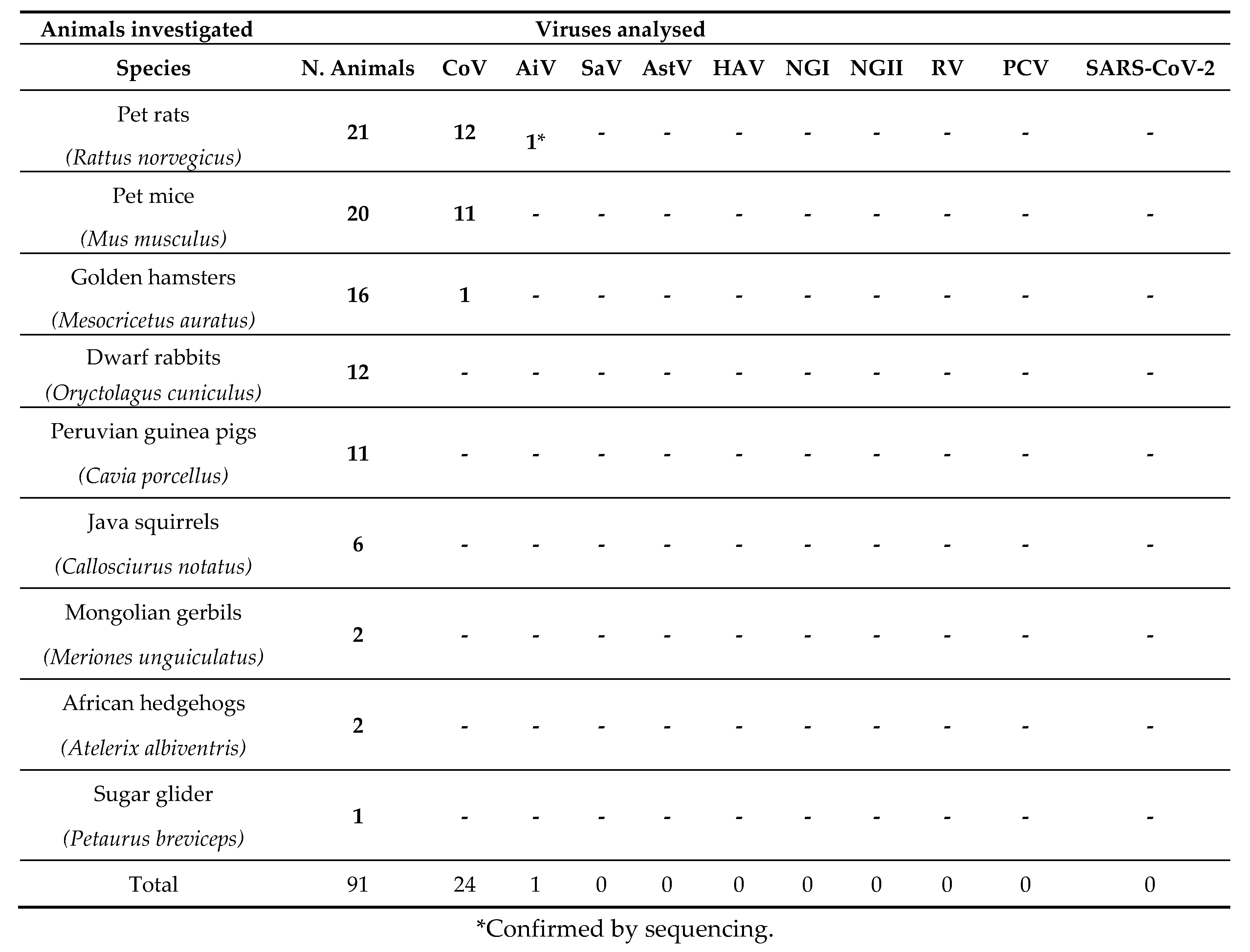
In details the following species were tested: golden hamster (Mesocricetus auratus n = 16), Java squirrel (Callosciurus notatus n =6), mongolian gerbil (Meriones unguiculatus n =2), peruvian guinea pig (Cavia porcellus n =11), pet mouse (Mus musculus n =20), pet rat (Rattus norvegicus n =21), dwarf rabbit (Oryctolagus cuniculus n =12), african hedgehog (Atelerix albiventris n =2) and sugar glider (Petaurus breviceps n =1).
Necropsy analysis
All analyzed animals were exotic companion animals gathered from veterinary clinics, pet shops and breeders. In particular hedgehogs came from Africa arrived first in central Europe (usually in the Czech Republic) and then were brought in Italy by european importers. Squirrels were imported directly from Malaysia and Thailand. The other animals (golden hamsters, mongolian gerbils, peruvian guinea pigs, pet mice, pet rats, dwarf rabbits, and sugar gliders) were bred and marketed in Italy. Causes of death where not clearely acclared. Most of these animals arrived in large loads and died from stress during transport because of infectious conditions that arised as a result of stress and promiscuity. Other possible death reasons could be related to peracute infections in subclinically infected animals or interspecies transmission of infectious diseases enhanced by strict cohabitation of different species particularly in pet shops.
All individuals underwent necropsy carried out by professional staff (veterinarians and laboratory technicians) in a necropsy room. Organs (brain, lungs, heart, liver, spleen, kidney and intestine) were removed with sterile scalpels, dissected, split out in sterile tubes and delivered within 24h to the laboratory for virological investigations. Samples were immediately analysed or stored at −80°C before DNA/RNA extraction.
Nucleic acids extraction
For nucleic acid extraction, 25 mg of tissue were homogenated by Tissue Lyser (Qiagen GmbH, Hilden, Germany) in 2 mL Eppendorf safe-lock tubes containing 1 mL phosphate buffered saline solution (PBS) as described previously [Amoroso et al 2021]. Extraction was carried out on 200 µl of homogenate by using a QIAsymphony automated extraction system (Qiagen GmbH, Hilden, Germany) with the DSP Virus/Pathogen Mini kit (Qiagen GmbH, Hilden, Germany) according to the manufacturer’s instructions. A sample made with 200 µL of PBS instead of homogenate was used as a negative process control (NPC). PCR inhibitors likely present in the samples were monitored by adding an external process control (EPC), namely, murine norovirus [Tofani et al 2021], 5 µL of which (107 PFUmL-1) was spiked in each sample prior to extraction. EPC was amplified by real-time PCR as indicated in the literature [Baaert et al 2008]. Results were analyzed as follows: if the threshold cycle (Ct) of the EPC in the eluted sample was comparable to that of the EPC in the NPC, the sample was analyzed as undiluted. If, instead, the difference between the two Cts was at least 3 or a multiple of 3, all the analyses were carried out on the sample diluted 1:10 or more [Amoroso et al 2021].
Biomolecular analysis
Extracted samples were analyzed by Real-time PCR and Real-time Reverse Transcription PCR (RT-PCR) for the presence of the following 10 viruses: aichivirus (AiV), sapovirus (SaV), astrovirus (AsV), hepatitis A virus (HAV), norovirus GI (NGI), norovirus GII (NGII), rotavirus (RV), circovirus (PCV) and SARS-CoV-2, with primers and probes indicated in Table n. 1. The analysis for SARS-COV-2 detection was carried out using different diagnostic kits: 1) RADI COVID-19 Detection Kit-Menarini Diagnostics; 2) ARGENE® SARS-COV-2 R-GENE® BIOMÉRIEUX; 3) TaqMan™ 2019nCoV Assay Kit v1 - Thermo Fisher Scientific; 4) Allplex™ SARS-CoV-2 Assay–Seegene.
Furthermore, a generic end point PCR was carried out to detect any coronavirus likely present in the samples. In details, 5 uL of nucleic acids were amplified by a RT-PCR reaction targeting the partial sequence of a 456 bp fragment of the RNA-dependent RNA polymerase (RdRp) gene. PCR reaction was carried out with the primers indicated in Table 1 using the AGPATH reaction kit (Thermo Fisher Scientific) as already described [Amoroso et al 2020]. PCR products were analyzed by Tape station (Agilent Technologies—Santa Clara, CA, USA) using the D 1000 kit. Amplicons of the positive samples were sequenced by capillary electrophoresis as previously described [Amoroso et al 2013].
Aichivirus characterization
To further characterize the aichivirus identified, an RT-PCR end-point was performed using the one-step RT-PCR kit (Qiagen), followed by nested-PCR using the VP1-F/VP1-R primers pairs, which amplifies a fragment of approximately 296 bp within the VP1 (capsid protein) [Lodder et al. 2013].
The PCR product with the expected size was then purified through enzymatic reaction (ExoSAP-IT, Affymetrix, USB) and subjected to Sanger sequencing (Eurofins Genomic Milano, Italy).
The sequence named MuKb_PO207_ITA23 was submitted to NCBI database under the accession number OR828459.
The 200bp VP1 fragment of the strain MuKb_PO207_ITA23 was aligned with sequences retrieved from NCBI database by MEGA X software [Kumar et al. 2018]. The phylogenetic tree was built using the Neighbor-joining method with 500x bootstraps (MEGA X).
3. Results
Results of the investigation carried out are shown in Table 2.
Interestingly 1 of the 91 small mammals analysed, one pet rat (Rattus norvegicus) tested positive for aichivirus in the brain. The aichivirus identified underwent sequencing and phylogenetic analysis. The obtained partial VP1 sequence was compared with the public sequence database using BlastN and results confirmed the AiV to be member of the species Aichivirus A, genus Kobuvirus murine kobuvirus (MuKVs). The sequence showed the highest nucleotide identity of 96% (100% aa. Id.) with a murine kobuvirus 1 MuKV/YN27/CHN (MW292480) strain identified in China (see Figure 1).
The detected strain belongs to a group of strains exclusively found in China, while strains identified in the USA and in Hungary are less closely related (<90%nd. Id). The strain displayed a limited identity with human strains with 85% aa.id.
As reported in Table 2, 24 animals (12/21 pet rats, 11/20 pet mice and 1/16 golden hamster) tested positive to the broad range coronavirus PCR (RdRp gene). Positive samples, showing a unique band of around 456 bp, underwent sequencing analysis which unfortunately gave not conclusive results, so they couldn’t be confirmed and further characterised. None of the other viruses investigated (AstV, HAV, SaV, NGI, NGII, PCV, RV, SARS-CoV-2), resulted positive to Real-time PCR analysis.
4. Discussion
Most of the outbreaks involving novel pathogens are caused by infectious agents transmitted to humans from animals [Han et al. 2015]. The identification of the reservoir of zoonotic pathogens is therefore of utmost importance for the control of the diseases spread and for the prevention of new epidemic or pandemic events. This represents a priority in habitats in which the animals coexist with humans. On this regard small mammals are more often used as pets and therefore frequently live in very close contact with humans. This growing trend poses the attention on their role in maintaining or transmitting zoonotic pathogens to humans. In Europe, more than 20 zoonoses (caused by bacteria or parasites) involving small mammals (in particular rodents) are known [Viouque et al. 2022, Han et al., 2015]. Some of these diseases, transmitted by rodents directly or as vectors, may have serious human health consequences [Centers for Disease Control and Prevention, 2010]. Scarce information is instead currently available on the presence and epidemiology of viral pathogens with zoonotic potential in these small mammals. In recent years, the advancements in molecular techniques have revolutionized the field of virology, allowing the discover of many new viral species [Plyusnin, 2020], more than 1000 being officially recognized by the International Committee on Taxonomy of Viruses (ICTV) only in 2020 [Dance 2021]. Furthermore, thanks to modern techniques, many genetic reassortments between animal and human viruses have been documented and several viruses have been recently detected in both mammals and humans [Vijgen 2005, Riley 2005, Khamrin 2006, Matthijinsens2006, Chomel 2007, Firth 2014, Reperant 2016, Chafekar 2018].
The best example of virus able to infect multiple species is represented by coronavirus, which infects a wide range of wild and domestic animals as well as humans. CoVs indeed exhibited a penchant for jumping species barriers often with devastating effects. The emergence of highly pathogenic or infectious CoVs in humans over the past 20 years, including severe acute respiratory syndrome CoV (SARS-CoV) [Drosten et al. 2003], Middle East respiratory syndrome CoV (MERS-CoV) [Chafekar 2018], and the more recent severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2) [Dakroub et al. 2022], underscores the significant threat that CoV spillovers pose to public health.
Other examples of potential zoonotic viruses are represented by AsV and RVA which are known human pathogens involved in gastroenteritis episodes in children [Bergallo 2018, Albano. 2007]. Mammalian AsV was also identified in animals like bats [Amoroso et al. 2018] while for RVA there is a large literature describing its spillover from different animal species to humans or viceversa (from porcine [Maneekarn 2006], simian [Khamrin 2006], bovine [Cooney, 2001], feline/canine [Isegawa 1992], caprine [Khamrin 2006], lapine [Matthijnssens 2006] ovine [Banyai 2010], equine [Malasao et al. 2015].
We analysed 91 micromammal pets belonging to 9 different species for the presence of potential zoonotic viruses. The results obtained showed positivity to PCR for 25 animals, 24 of which, positive to CoV RdRp gene, couldn’t be confirmed by sequencing and will not be further discussed. Interestingly, the brain of a pet rat, tested positive for aichivirus, with partial sequence showing a notable identity (96% nt. id.) with the murine kobuvirus-1 strain MuKV/YN27/CHN (MW292480), a strain identified in a rat (MW292480). Interestingly, the identified strain clusters with a group of strains identified exclusively in China [Riley et al 2005, Zhang et al. 2021].
The MuKV is distinct from human strains, despite all belonging to Aichivirus A [Reuter, Boros, and Pankovics 2011]. However, the obtained fragment within VP1 does not allow for an in-depth analysis of genomic variability. It is also noteworthy that the available sequences are predominantly from China, which influences the comparison results.
In rat, detection of AiV was reported in China [You et al. 2020, Zhang et al. 2021, Gao et al. 2022], in America [Firth et al. 2014], in Europe, MuKVs were identified in Hungary [Boros et al 2019] and various studies in Asia and America describe their presence in the feces of different rodents, including Apodemus agrarius, Rattus norvegicus, Rattus losea, Rattus argentiventer, and Mus musculus [Lodder et al 2013,You et al. 2020; Williams et al. 2018; Lu et al. 2018, d'Ovidio et al 2023].
All the pet rats we analysed were born in Italy and bread to be sold as pets. Some of them, during their permanence in the pet shops came into contact with various other rodents also from non EU countries like Malaysia and Thailand. Since according to literature, no other AiV was described in pet rats in Italy, we hypothesized that the virus was introduced in our country through the importation of other small mammals from Asia, (as already suggested for other viral agents [Alfano et al 2019, Lu et al 2018] which infected the rat by coming into close contact (may be in pet shops) with it.
AiV was found in different animals in Europe: in Hungary in healthy piglets (Sus scrofa domestica) [Reuter et al. 2009], in Italy in Mussels and Shellfish [Fusco et al. 2017 and 2019], in dogs [Kapoor et al. 2011, Di Martino et al. 2013], in cats [Di Martino et al. 2015], pipistrelle bat [Diakoudi et al. 2020], in domestic and wild ungulates [Di Martino et al. 2021] and in goats [Abi et al. 2022]. Some authors [Kapoor et al. 2011) identified in domestic dogs (with gastroenteritis) a kobuvirus closely related to human aichivirus thus witnessing cross-species transmission in cohabiting species.
Aichivirus was also found in the environment of different countries. It was indeed described in sewage samples, in Africa: Tunisia, [Sdiri et al 2010], in marine bathing and non-bathing waters in Italy [Bonadonna et al. 2019, Cioffi et al. 2021].
Furthermore the virus was described as involved in children gastroenteritis in different countries: in South America: Brasile [Oh et al. 2006], in Europe: Germany [Oh et al. 2006], Finlandia [Kaikkonen et al. 2010], Italy [Bergallo et al. 2017, Biscaro et al. 2018]; in Asia: Bangladesh, Thailand, Vietnam, and Japan [Pham et al. 2007].
The aichiviruses, since being ubiquitary in the environment and also associated to gastroenteritis in children [Bergallo 2017, Rivadulla 2020] must be taken into due account as potential zoonotic pathogen and further investigations are desirable to obtain a clearer picture of its circulation.
None of the other viruses analysed (AstV, HAV, SaV, NGI, NGII, PCV, RV, SARS-CoV-2), resulted positive to Real-time PCR analysis. These data reassure about the absence of almost all the viruses investigated in the animals analysed. Since these animals are treated as pets and always live in close contact with children, it is important to monitor and guarantee their health and the safety of their “employment” as pets. However, even though we researched for 10 potential zoonotic viruses and investigated around one hundred animals, there are a lot of other viruses (with also unknown zoonotic potential) which, if searched, could be found. Level of attention must be therefore always kept high also because these animals usually make a huge ride before arriving in our houses and are subject to a high risk of contact with infectious agents.
Concluding our study underlines the importance of monitoring unconventional pets, which role in the transmission of zoonotic agents has been almost neglected till now. Further investigation on a large number of animals including other viruses is crucial in order to guarantee public health expecially for children since they are used to play and to closely interact with these pets.
Author Contributions
Conceptualization, F.A.; methodology, F.A., M.G.A., D.dO, M.S., M.G.L., I.DB; investigation, F.A., S.R., M.M., F.S., M.L., F.P., A.G; writing—original draft preparation, F.A.; writing—review and editing, M.G.A; project administration, G.F., E.DC; funding acquisition, F.A. All authors have read and agreed to the published version of the manuscript.
Funding
This study was funded by the Italian Ministry of Health, Ricerca Corrente 2019, IZS ME, 04/19 RC, recipient Flora Alfano.
Institutional Review Board Statement
Ethical review and approval were waived for this study, as samples were part of the diagnostic activity of Istituto Zooprofilattico Sperimentale del Mezzogiorno, statal public institution that operate within the national health service.
Informed Consent Statement
Not applicable.
Data Availability Statement
All data supporting the present study is reported in this manuscript. Sequence data presented in this study are openly available in the GenBank database.
Acknowledgments
We thank Amalia Gallo and Clementina Auriemma for their kind and excellent assistance.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Abi, K.M.; Yang, C.; Tang, C.; Jing, Z.Z. Aichivirus C isolate is a diarrhoea-causing pathogen in goats. Transbound Emerg Dis 2022, 69, e2268-e2275. Epub 2022 May 11. PMID: 35502695. [CrossRef]
- Albano, F.; Bruzzese, E.; Bella, A.; Cascio, A.; Titone, L.; Arista, S.; Izzi, G.; Virdis, R.; Pecco, P.; Principi, N.; Fontana, M.; Guarino, A. Rotavirus and not age determines gastroenteritis severity in children: a hospital-based study. Eur J Pediatr 2007, 166, 241-7. Epub 2006 Aug 29. PMID: 16941130. [CrossRef]
- Alfano, F.; Dowgier, G.; Valentino, M.P.; Galiero, G.; Tinelli, A.; Decaro, N.; Fusco, G. Identification of Pantropic Canine Coronavirus in a Wolf (Canis lupus italicus) in Italy. J Wildl Dis 2019, 55, 504-508. [CrossRef]
- Alfano, F.; Fusco, G.; Mari, V.; Occhiogrosso, L.; Miletti, G.; Brunetti, R.; Galiero, G.; Desario, C.; Cirilli, M.; Decaro, N. Circulation of pantropic canine coronavirus in autochthonous and imported dogs, Italy. Transbound Emerg Dis 2020, 67, 1991–9. [CrossRef]
- Amoroso, MG.; Lucifora, G.; Degli Uberti, B.; Serra, F.; De Luca, G.; Borriello, G.; De Domenico, A.; Brandi, S.; Cuomo, MC.; Bove, F.; Riccardi, MG.; Galiero, G.; Fusco, G. Fatal Interstitial Pneumonia Associated with Bovine Coronavirus in Cows from Southern Italy. Viruses 2020, 12, 1331. PMID: 33228210; PMCID: PMC7699522. [CrossRef]
- Amoroso, MG.; Serra, F.; Esposito, C.; D'Alessio, N.; Ferrara, G.; Cioffi, B.; Anzalone, A.; Pagnini, U.; De Carlo, E.; Fusco, G.; Montagnaro, S. Prevalence of Infection with Porcine Circovirus Types 2 and 3 in the Wild Boar Population in the Campania Region (Southern Italy). Animals (Basel) 2021, 11,3215. 10: ,. [CrossRef]
- Amoroso, M.G.; Corrado, F.; De Carlo, E.; Lucibelli, M.G.; Martucciello, A.; Guarino, A.; Galiero, G. Bubaline Herpesvirus 1 Associated with Abortion in a Mediterranean Water Buffalo. Res Vet Sci 2013, 94, 813–816. [CrossRef]
- Amoroso, MG.; Russo, D.; Lanave, G.; et al. Detection and phylogenetic characterization of astroviruses in insectivorous bats from Central-Southern Italy. Zoonoses Public Health 2018, 65, 702–710. [CrossRef]
- Baaert, L.; Wobus, C.E.; Van Coillie, E.; Thackray, L.B.; Debevere, J.; Uyttendaele, M. Detection of murine norovirus 1 by using plaque assay, transfection assay, and real-time reverse transcription-PCR before and after heat exposure. Appl Environ Microbiol 2008, 74, 543–546. [CrossRef]
- Bányai, K.; Papp, H.; Dandár, E.; Molnár, P.; Mihály, I.; Van Ranst, M.; Martella, V.; Matthijnssens, J. Whole genome sequencing and phylogenetic analysis of a zoonotic human G8P[14] rotavirus strain. Infect Genet Evol 2010, 10, 1140-4. 1016. [CrossRef]
- Bányai, K.; Kemenesi, G.; Budinski, I.; Földes, F.; Zana, B.; Marton, S.; Varga-Kugler, R.; Oldal, M.; Kurucz, K.; Jakab, F. Candidate new rotavirus species in Schreiber's bats, Serbia. Infect Genet Evol 2017, 48, 19-26. Epub 2016 Dec 6. PMID: 27932285; PMCID: PMC7106153. 1016. [CrossRef]
- Bergallo, M.; Galliano, I.; Montanari, P.; Rassu, M.; Daprà, V. Aichivirus in Children with Diarrhea in Northern Italy. Intervirology 2017, 60, 196-200. Epub 2018 Mar 2. PMID: 29502122. [CrossRef]
- Bergallo, M.; Galliano, I.; Daprà, V.; Rassu, M.; Montanari, P.; Tovo, PA. Molecular Detection of Human Astrovirus in Children With Gastroenteritis, Northern Italy. Pediatr Infect Dis J 2018, 37, 738-742. PMID: 30004391. [CrossRef]
- Biscaro, V.; Piccinelli, G.; Gargiulo, F.; Ianiro, G.; Caruso, A.; Caccuri, F.; De Francesco, M.A. Detection and molecular characterization of enteric viruses in children with acute gastroenteritis in Northern Italy. Infect Genet Evol 2018, 60, 35-41. doi: 10.1016/j.meegid.2018.02.011. Epub 2018 Feb 10. PMID: 29438743. [CrossRef]
- Bonadonna, L.; Briancesco, R.; Suffredini, E.; Coccia, A.; Della Libera, S.; Carducci, A.; Verani, M.; Federigi, I.; Iaconelli, M.; Bonanno Ferraro, G.; Mancini, P.; Veneri, C.; Ferretti, E.; Lucentini, L.; Gramaccioni, L.; La Rosa, G. Enteric viruses, somatic coliphages and Vibrio species in marine bathing and non-bathing waters in Italy. Mar Pollut Bull 2019, 149, 110570. Epub 2019 Sep 8. PMID: 31542593. [CrossRef]
- Boros, Á.; Orlovácz, K.; Pankovics, P.; Szekeres, S.; Földvári, G.; Fahsbender, E.; Delwart, E.; Reuter, G. Diverse picornaviruses are prevalent among free-living and laboratory rats (Rattus norvegicus) in Hungary and can cause disseminated infections. Infect Genet Evol 2019, 75, 103988. Epub 2019 Aug 1. PMID: 31377399. [CrossRef]
- Centers for Disease Control and Prevention. Diseases from rodents. www.cdc.gov/rodents/diseases/index.html (2010).
- Chhabra, P.; de Graaf, M.; Parra, G.I.; Chan, M.C.; Green, K.; Martella, V.; Wang, Q.; White, P.A.; Katayama, K.; Vennema, H.; Koopmans, M.P.G.; Vinjé, J. Updated classification of norovirus genogroups and genotypes. J Gen Virol 2019, 100, 1393-1406. Erratum in: J Gen Virol 2020, 101, 893. PMID: 31483239; PMCID: PMC7011714. [CrossRef]
- Chafekar, A.; Fielding, B.C. MERS-CoV: Understanding the Latest Human Coronavirus Threat. Viruses 2018, 10, 93. PMID: 29495250; PMCID: PMC5850400. Viruses 2018, 10. [CrossRef]
- Chan, J.F.; Yuan, S.; Kok, K.H.; To, K.K.; Chu, H.; Yang, J.; Xing, F.; Liu, J.; Yip, C.C.; Poon, R.W.; Tsoi, H.W.; Lo, S.K.; Chan, K.H.; Poon, V.K.; Chan, W.M.; Ip, J.D.; Cai, J.P.; Cheng, V.C.; Chen, H.; Hui, C.K.; Yuen, K.Y. A familial cluster of pneumonia associated with the 2019 novel coronavirus indicating person-to-person transmission: a study of a family cluster. Lancet 2020, 395, 514-523. Epub 2020 Jan 24. PMID: 31986261; PMCID: PMC7159286. 1016. [CrossRef]
- Chomel, B.B.; Belotto, A.; Meslin, F.X. Wildlife, exotic pets, and emerging zoonoses. Emerg Infect Dis 2007, 13, 6-11.
- Cioffi, B.; Ianiro, G.; Iaccarino, D.; D'Apice, F.; Ferraro, A.; Race, M.; Spasiano, D.; Esposito, E.; Monini, M.; Serra, F.; Cozza, D.; Di Nocera, F.; De Maio, L.; Amoroso, M.G.; De Carlo E.; Fusco, G. A potential risk assessment tool to monitor pathogens circulation in coastal waters. Environ Res 2021, 200, 111748. Epub 2021 Jul 22. PMID: 34303676. [CrossRef]
- Cooney, M.A.; Gorrell, R.J.; Palombo, E.A. Characterisation and phylogenetic analysis of the VP7 proteins of serotype G6 and G8 human rotaviruses. J Med Microbiol 2001, 50, 462-467. PMID: 11339255. [CrossRef]
- Costafreda, M.I.; Bosch, A.; Pintó, R.M. Development, evaluation, and standardization of a real-time TaqMan reverse transcription-PCR assay for quantification of hepatitis A virus in clinical and shellfish samples. Appl Environ Microbiol 2006, 72, 3846-55. PMID: 16751488; PMCID: PMC1489592. [CrossRef]
- Dakroub, H.; Russo, D.; Cistrone, L.; Serra, F.; Fusco, G.; De Carlo E.; Amoroso, M.G. A First Assessment of SARS-CoV-2 Circulation in Bats of Central-Southern Italy. Pathogens. 2022, 11, 742. PMID: 35889988 Free PMC article. [CrossRef]
- Dance, A. Beyond coronavirus: The virus discoveries transforming biology. Nature 2021, 595, 22–25.
- Di Martino, B.; Di Felice, E.; Ceci, C.; Di Profio, F.; Marsilio, F. Canine kobuviruses in diarrhoeic dogs in Italy. Vet Microbiol 2013, 166, 246-249. Epub 2013 Jun 6. PMID: 23806200; PMCID: PMC7117211. [CrossRef]
- Di Martino, B.; Di Profio, F.; Melegari, I.; Marsilio, F.; Martella, V. Detection of feline kobuviruses in diarrhoeic cats, Italy. Vet Microbiol 2015, 176, 186-189. Epub 2015 Jan 17. PMID: 25631253; PMCID: PMC7117564. [CrossRef]
- Di Martino, B.; Di Profio, F.; Robetto, S.; Fruci, P.; Sarchese, V.; Palombieri, A.; Melegari, I.; Orusa, R.; Martella, V.; Marsilio, F. Molecular Survey on Kobuviruses in Domestic and Wild Ungulates From Northwestern Italian Alps. Front Vet Sci 2021, 8, 679337. PMID: 34195249; PMCID: PMC8237713. [CrossRef]
- Diakoudi, G.; Jamnikar-Ciglenečki, U.; Lanave, G.; Lelli, D.; Martella, V.; Kuhar, U. Genome sequence of an aichivirus detected in a common pipistrelle bat (Pipistrellus pipistrellus). Arch Virol 2020, 165, 1019-1022. Epub 2020 Feb 13. PMID: 32056001. 32056001. [CrossRef]
- d'Ovidio, D.; Santoro, D. Zoonotic Dermatoses of Exotic Companion Mammals. Vet Clin North Am Exot Anim Pract 2023, 26, 511-523. [CrossRef]
- Drosten, C.; Gunther, S.; Preiser, W.; van der Werf, S.; Brodt, H.R.; Becker, S.; Rabenau, H.; Panning, M.; Kolesnikova, L.; Fouchier, R.A.; et al. Identification of a Novel Coronavirus in Patients with Severe Acute Respiratory Syndrome. N. Engl. J. Med 2003, 348, 1967–1976. [CrossRef]
- Firth, C.; Bhat, M.; Firth, M.A.; Williams, S.H.; Frye M.J.; Simmonds, P.; Conte, J.M.; Ng, J.; Garcia, J.; Bhuva, N.P.; Lee, B.; Che, X.; Quan, P.L.; Lipkin, W.I. Detection of zoonotic pathogens and characterization of novel viruses carried by commensal Rattus norvegicus in New York City. mBio 2014, 5, e01933-14. PMID: 25316698; PMCID: PMC4205793. [CrossRef]
- Fusco, G.; Di Bartolo, I.; Cioffi, B.; Ianiro, G.; Palermo, P.; Monini, M.; Amoroso, M.G. Prevalence of Foodborne Viruses in Mussels in Southern Italy. Food Environ Virol 2017, 9, 187-194. Epub 2017 Jan 4. PMID: 28054332. [CrossRef]
- Fusco, G.; Anastasio, A.; Kingsley, D.H.; Amoroso, M.G.; Pepe, T.; Fratamico, P.M.; Cioffi, B.; Rossi, R.; La Rosa, G.; Boccia, F. Detection of Hepatitis A Virus and Other Enteric Viruses in Shellfish Collected in the Gulf of Naples, Italy. Int J Environ Res Public Health 2019, 16, 2588. PMID: 31331104; PMCID: PMC6678136. [CrossRef]
- Gao, Y.; He, W.; Fu, J.; Li, Y.; He, H.; Chen, Q. Epidemiological Evidence for Fecal-Oral Transmission of Murine Kobuvirus. Front Public Health 2022, 10, 865605. PMID: 35517645; PMCID: PMC9062591. [CrossRef]
- Haake, C.; Cook, S.; Pusterla, N.; Murphy, B. Coronavirus Infections in Companion Animals: Virology, Epidemiology, Clinical and Pathologic Features. Viruses 2020, 12, 1023.
- Han, B.A.; Schmidt, J.P.; Bowden, S.E. & Drake, J.M. Rodentreser-voirsoffuture zoonotic diseases. Proceedings of the National Academy of Sciences of the United States of America, 2015, 112, 7039–7044. [CrossRef]
- Isegawa, Y.; Nakagomi, O.; Nakagomi, T.; Ueda, S. A. VP4 sequence highly conserved in human rotavirus strain AU-1 and feline rotavirus strain FRV-1. J Gen Virol 1992, 73, 1939-46. PMID: 1322955. [CrossRef]
- Johnson, C.K.; Hitchens, P.L.; Pandit, P.S.; Rushmore, J.; Evans, T.S.; Young, C.C.W.; Doyle, M.M. Global shifts in mammalian population trends reveal key predictors of virus spillover risk. Proc Biol Sci 2020, 287, 20192736. Epub 2020 Apr 8. PMID: 32259475; PMCID: PMC7209068. [CrossRef]
- Kaikkonen, S.; Rasanen, S.; Ramet, M.; et al: Aichivirus infection in children with acute gastroenteritis in Finland. Epidemiol Infect 2010, 138, 1166–1171.
- Kapoor, A.; Simmonds, P.; Dubovi, E.J.; Qaisar, N.; Henriquez, JA.; Medina, J.; Shields, S.; Lipkin, W.I. Characterization of a canine homolog of human Aichivirus. J Virol 2011, 85, 11520-5. Epub 2011 Aug 31. PMID: 21880761; PMCID: PMC3194941. [CrossRef]
- Khamrin, P.; Maneekarn, N.; Peerakome, S.; Yagyu, F.; Okitsu, S.; Ushijima, H. Molecular characterization of a rare G3P[3] human rotavirus reassortant strain reveals evidence for multiple human-animal interspecies transmissions. J Med Virol 2006, 78, 986-94. PMID: 16721863. [CrossRef]
- Kim, H.R.; Park, Y.R.; Lim, D.R.; Park, M.J.; Park, J.Y.; Kim, S.H.; Lee, K.K.; Lyoo, Y.S.; Park, C.K. Multiplex real-time poly-merase chain reaction for the differential detection of porcine circovirus 2 and 3. J. Virol. Methods 2017, 250, 11–16. [Google Scholar] [CrossRef] [PubMed]
- Kitajima, M.; Hata, A.; Yamashita, T.; Haramoto, E.; Minagawa, H.; Katayama, H. Development of a reverse transcription-quantitative PCR system for detection and genotyping of aichi viruses in clinical and environmental samples. Appl Environ Microbiol 2013, 79, 3952-8. Epub 2013 Apr 19. PMID: 23603673; PMCID: PMC3697579. [CrossRef]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across Computing Platforms. Mol Biol Evol 2018, 35, 1547–1549. [Google Scholar] [CrossRef] [PubMed]
- Le Cann, P.; Ranarijaona, S.; Monpoeho, S.; Le Guyader, F.; Ferré, V. Quantification of human astroviruses in sewage using real-time RT-PCR. Res Microbiol 2004, 155, 11-5. PMID: 14759703. [CrossRef]
- Lodder, W.J., Rutjes, S.A.; Takumi, K.; de Roda Husman, A.M. Aichi virus in sewage and surface water, the Netherlands. Emerg Infect Dis 2013, 19, 1222-30. [CrossRef]
- Lu, L.; Van Dung, N.; Ivens, A.; Bogaardt, C.; O'Toole, A.; Bryant, J.E.; Carrique-Mas, J.; Van Cuong, N.; Anh, P.H.; Rabaa, M.A., et al. Genetic diversity and cross-species transmission of kobuviruses in Vietnam. Virus Evol 2018, 4, vey002. [CrossRef]
- Malasao, R.; Saito, M.; Suzuki, A.; Imagawa, T.; Nukiwa-Soma, N.; Tohma, K.; Liu, X.; Okamoto, M.; Chaimongkol, N.; Dapat, C.; Kawamura, K.; Kayama, Y.; Masago, Y.; Omura, T.; Oshitani, H. Human G3P[4] rotavirus obtained in Japan, 2013, possibly emerged through a human-equine rotavirus reassortment event. Virus Genes 2015, 50, 129-33. Epub 2014 Oct 29. PMID: 25352228; PMCID: PMC4349953. [CrossRef]
- Maneekarn, N.; Khamrin, P.; Chan-it, W.; Peerakome, S.; Sukchai, S.; Pringprao, K.; Ushijima, H. Detection of rare G3P[19] porcine rotavirus strains in Chiang Mai, Thailand, provides evidence for origin of the VP4 genes of Mc323 and Mc345 human rotaviruses. J Clin Microbiol 2006, 44, 4113-9. Epub 2006 Sep 20. PMID: 16988014; PMCID: PMC1698310. [CrossRef]
- Matthijnssens, J.; Rahman, M.; Martella, V.; Xuelei, Y.; De Vos, S.; De Leener, K.; Ciarlet, M.; Buonavoglia, C.; Van Ranst, M. Full genomic analysis of human rotavirus strain B4106 and lapine rotavirus strain 30/96 provides evidence for interspecies transmission. J Virol 2006, 80, 3801-10. PMID: 16571797; PMCID: PMC1440464. [CrossRef]
- Mira, F.; Purpari, G.; Lorusso, E.; Di Bella, S.; Gucciardi, F.; Desario, C.; Macaluso, G.; Decaro, N.; Guercio, A. Introduction of Asian canine parvovirus in Europe through dog importation. Transbound Emerg Dis 2018, 65, 16-21. [CrossRef]
- Oh, D.Y.; Silva, P.A.; Hauroeder, B.; et al: Molecular characterization of the first Aichi viruses isolated in Europe and in South America. Arch Virol 2006, 151, 1199–1206. [CrossRef]
- Pham, N.T.; Khamrin, P.; Nguyen, T.A.; et al: Isolation and molecular characterization of Aichi viruses from fecal specimens collected in Japan.; Bangladesh, Thailand, and Vietnam. J Clin Microbiol 2007, 45, 2287–2288. [CrossRef]
- Plyusnin, I.; Kant, R.; Jääskeläinen, A.J.; Sironen, T.; Holm, L.; Vapalahti, O.; Smura, T. Novel NGS pipeline for virus discovery from a wide spectrum of hosts and sample types. Virus Evol 2020, 6, veaa091. [CrossRef]
- Reperant, L.A.; Brown, I.H.; Haenen, O.L.; de Jong, M.D.; Osterhaus, A.D.; Papa, A.; Rimstad, E.; Valarcher, J.F.; Kuiken, T. Companion Animals as a Source of Viruses for Human Beings and Food Production Animals. J Comp Pathol 2016, 155, S41-53. Epub 2016 Aug 10. PMID: 27522300. [CrossRef]
- Reuter, G.; Boldizsár, A.; Pankovics, P. Complete nucleotide and amino acid sequences and genetic organization of porcine kobuvirus.; a member of a new species in the genus Kobuvirus.; family Picornaviridae. Arch Virol. 2009, 154, 101-108. Epub 2008 Dec 19. PMID: 19096904. [CrossRef]
- Reuter, G.; Boros, A. and Pankovics, P. 'Kobuviruses - a comprehensive review'. Rev Med Virol 2011, 21, 32-41. [CrossRef]
- Riley, P.Y, Chomel, B.B. Hedgehog zoonoses. Emerg Infect Dis 2005, 11, 1-5.
- Rivadulla, E.; Romalde, J.L. A Comprehensive Review on Human Aichi Virus. Virol Sin 2020, 35, 501-516. Epub 2020 Apr 27. Erratum in: Virol Sin. 2020 Dec 2;: PMID: 32342286; PMCID: PMC7223127. [CrossRef]
- Sdiri-Loulizi, K.; Hassine, M.; Aouni, Z.; et al: First molecular detection of Aichi virus in sewage and shellfish samples in the Monastir region of Tunisia. Arch Virol 2010, 155, 1509–1513. [CrossRef]
- Tofani, S.; Ianiro, G.; De Sabato, L.; Monini, M.; Angeloni, G.; Ponterio, E.; D’Agostino, C.; Di Bari, M.A.; Valeri, M.; Di Bartolo, I. Detection and whole genome sequencing of murine norovirus in animal facility in Italy. Anim. Biotechnol 2021, 29, 1–8. 34. [CrossRef]
- UNI EN ISO 15216-2:2019.
- Varela, M.F.; Hooper, A.S.; Rivadulla, E.; Romalde, J.L. Human Sapovirus in Mussels from Ría do Burgo, A Coruña (Spain). Food Environ Virol 2016, 8, 187–193. Epub 2016 May 7. PMID: 27156175. [CrossRef]
- Vijgen, L.; Keyaerts, E.; Moës, E.; Thoelen, I.; Wollants, E.; Lemey, P.; Vandamme, A.M.; Van Ranst, M. Complete genomic sequence of human coronavirus OC43: molecular clock analysis suggests a relatively recent zoonotic coronavirus transmission event. J Virol 2005, 79, 1595-1604. PMID: 15650185; PMCID: PMC544107. [CrossRef]
- Vioque, F.; Dashti, A.; Santín, M.; Ruiz-Fons, F.; Köster, P.C.; Hernández-Castro, C.; García, J.T.; Bailo, B.; Ortega, S.; Olea, P.P.; Arce, F.; Chicharro, C.; Nieto, J.; González, F.; Viñuela, J.; Carmena, D.; González-Barrio, D. Wild micromammal host spectrum of zoonotic eukaryotic parasites in Spain. Occurrence and genetic characterisation. Transbound Emerg Dis 2022, 69, e2926-e2942. Epub 2022 Jul 8. PMID: 35752461. [CrossRef]
- Williams, S.H.; Che, X.; Garcia, J.A.; Klena, J.D.; Lee, B.; Muller, D.; Ulrich, W.; Corrigan, R.M.; Nichol, S.; Jain, K.; Lipkin, W.I. Viral Diversity of House Mice in New York City. mBio 2018, 9, e01354-17. PMID: 29666290; PMCID: PMC5904411. [CrossRef]
- You, F.F.; Zhang, M.Y.; He, H.; He, W.Q.; Li, Y.Z.; Chen, Q. Kobuviruses carried by Rattus norvegicus in Guangdong.; China. BMC Microbiol 2020, 20, 94. PMID: 32295529; PMCID: PMC7161169. [CrossRef]
- Zeng, S.Q.; Halkosalo, A.; Salminen, M.; Szakal, E.D.; Puustinen, L.; Vesikari, T. One-step quantitative RT-PCR for the detection of rotavirus in acute gastroenteritis. J Virol Methods 2008, 153, 238-40. Epub 2008 Sep 17. PMID: 18765254. [CrossRef]
- Zhang, M.; You, F.; Wu, F.; He, H.; Li, Q.; Chen Q. Epidemiology and genetic characteristics of murine kobuvirus from faecal samples of Rattus losea, Rattus tanezumi and Rattus norvegicus in southern China. J Gen Virol 2021, 102, 001646. PMID: 34486970; PMCID: PMC8567428. [CrossRef]
Figure 1.
Phylogenetic tree based on the alignments of the 200bp VP1 partial fragments. Accession numbers, strain name and country of detection are reported for each entry. The strain identified in this study is indicated by black dot.
Figure 1.
Phylogenetic tree based on the alignments of the 200bp VP1 partial fragments. Accession numbers, strain name and country of detection are reported for each entry. The strain identified in this study is indicated by black dot.
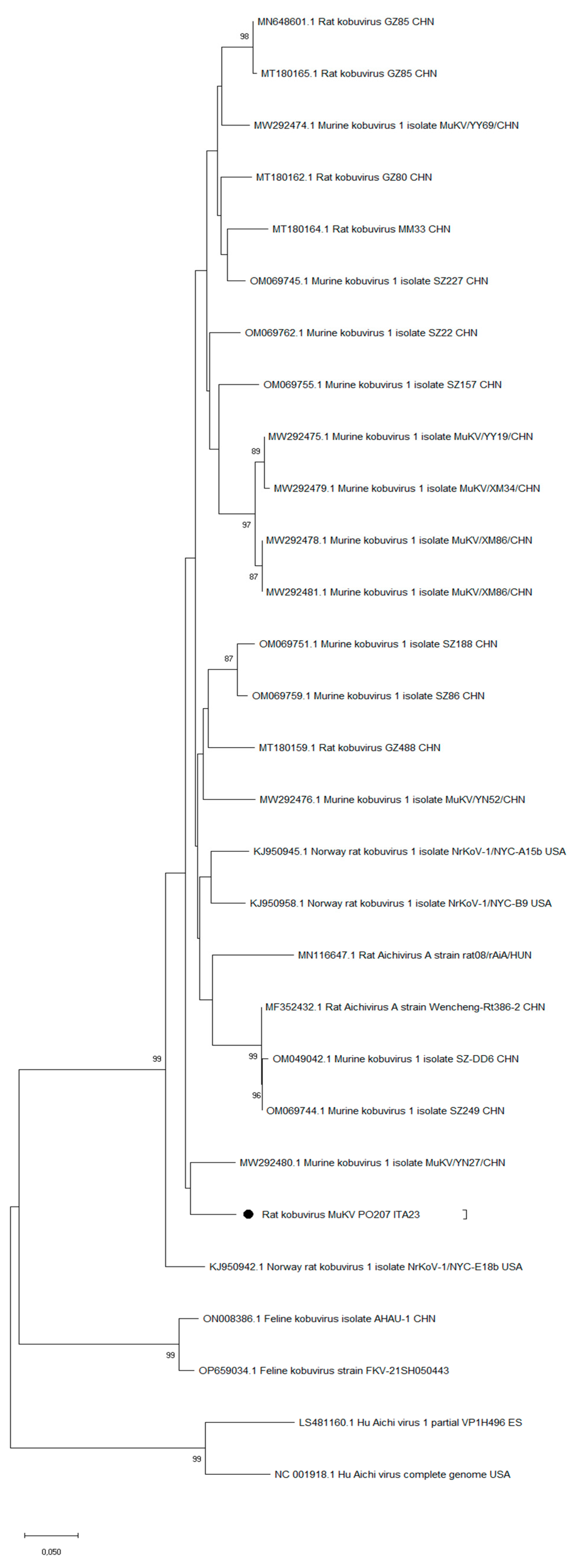

Table 1.
Primer pairs and specific probe used for virus detection.
| Virus | Primers | Bibliography | |
|---|---|---|---|
| Astrovirus | Primer forward | 5’-CCGAGTAGGATCGAGGGT-3’ | Le Cann et al. 2004 |
| Primer reverse | 5’-GCTTCTGATTAAATCAATTTTAA-3’ | ||
| Probe | FAM: 5’-CTTTTCTGTCTCTGTTTAGATTATTTTAATCACC-3’ Tamra | ||
| Aichivirus | Primer forward | 5’-GTCTCCACHGACACYAAYTGGAC-3’ | Kitajima et al. 2013 |
| Primer reverse | 5’- GTTGTACATRGCAGCCCAGG-3’ | ||
| Probe | 5’-FAM-TTYTCCTTYGTGCGTGC- 3’NFQ (MGB) | ||
| Sapovirus | Primer forward | 5’-GAYCASGCTCTCGCYACCTAC-3’ | Varela et al. 2016 |
| Primer reverse | 5’-CCCTCCATYTCAAACACTA-3’ | ||
| Probe | 5’- FAM-CCCCTATRAACCA-3’NFQ (MGB) | ||
| Rotavirus | Primer forward | 5’-ACCATCTWCACRTRACCCTCTATGAG-3’ | Zeng et al. 2008 |
| Primer reverse | 5’-GGTCACATAACGCCCCTATAGC-3’ | ||
| Probe | 5’- FAM-AGTTAAAAGCTAACACTGTCAAA-3’(MGB) | ||
| Norovirus GI | Primer forward | 5’-CGCTGGATGCGNTTCCAT-3’ | ISO 15216 |
| Primer reverse | 5’-CCTTAGACGCCATCATCATTTAC-3’ | ||
| Probe | FAM-5’-TGGACAGGAGAYCGCRATCT-3’TAMRA | ||
| Norovirus GII | Primer forward | 5’-ATGTTCAGRTGGATGAGRTTCTCWGA-3’ | ISO 15216 |
| Primer reverse | 5’-TCGACGCCATCTTCATTCACA-3’ | ||
| Probe | FAM-5’-AGCACGTGGGAGGGCGATCG-3’-MGB/NFQ | ||
| Hepatitis A Virus | Primer forward | 5’-TCACCGCCGTTTGCCTAG-3’ | ISO 15216 |
| Primer reverse | 5’-GGAGAGCCCTGGAAGAAAG-3’ | ||
| Probe | FAM-CCTGAACCTGCAGGAATTAA-3’-MGB/NFQ | ||
| Porcine Circovirus | Primer forward | ACGTCCCTTTACTTTCAATTCACA | Kim et al. 2017 |
| Primer reverse | TATACTTGGTACACACATCCAGAGTCA | ||
| Probe | FAM-TGAGTTGATTACTGGCACGCCTAAACCAC-BHQ | ||
| Coronavirus | Primer forward | GGGTTGGGACTATCCTAAGTGTGA | Drosten et al. 2003 |
| Primer reverse | TAACACACAAACACCATCATCA | ||
Table 2.
Species analyzed and viral positive detection by biomolecular assays.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



