
Submitted:
27 December 2023
Posted:
28 December 2023
You are already at the latest version
Preprints on COVID-19 and SARS-CoV-2
Abstract
Several methods can be used to evaluate antibody avidity, one of which is the modified ELISA, in which a chaotropic agent is added to disrupt the binding between the antigen and the antibody. Although the avidity index provides relevant information about the functionality of antibodies, it does not allow us to characterize which antigens the antibodies are strongly bound to. To this end, we adapted the Immunoblot and Dot-Blot techniques by adding potassium thiocyanate (KSCN) after incubating nitrocellulose membranes with serum from mice immunized with Neisseria meningitidis outer membrane vesicles (OMVs) or serum from mice immunized with recombinant receptor-binding domain (RBD) of SARS-CoV-2, with different adjuvants for each antigen. In terms of results, when standardizing the incubation time of sera with KSCN, we observed that 5 minutes was the ideal time for both assays, however, when using different dilutions of serum from mice immunized with RBD, there was an influence on the strength of antigen-antibody binding. For the N. meningitidis assay, the antibodies maintained high avidity for the antigens present in the OMVs, PorA, NadA, and Opa, especially in the presence of the aluminum hydroxide adjuvant. The research showed that standardizing the Immunoblot and Dot-Blot in the same assay optimizes time and resources, allowing qualitative analysis for two different antigens, and resulting in more robust information for immunization studies against these pathogens.
Keywords:
Avidity
; Immunoblot
; Dot-Blot
; Neisseria meningitidis
; SARS-CoV-2.
1. Introduction
Antibodies are produced by differentiated B cells (plasma cells) as part of humoral immunity, following antigen presentation. This occurs either in response to natural infections or after stimulation by vaccination to fight the infection and/or prepare the immune system before it occurs [5,19].
The antibody interacts with the antigen via a non-covalent binding that is specific to the epitope and the paratope. The binding strength between the paratope and the epitope determines the affinity of the antibody, while the overall binding strength between multiple antibodies and multiple binding sites is called avidity [12]. It is well known that IgG antibodies tend to bind strongly to antigens because of a process called affinity maturation, when the efficiency of the immune response improves after a second exposure to the pathogen, contributing to the generation of antibodies with high affinity and specificity [35].
Antibody avidity is an important criterion for assessing the immune response, as it provides information on the functionality of the antibody, which can be used as a parameter to measure vaccine efficacy or the evolution of the stage of a disease, e.g., whether the infection is in its acute or latent stage [29,32].
Several methods can be used to assess antibody avidity. A very common method is the modified ELISA, which consists of adding a chaotropic agent to disrupt the bond between the antigen and the antibody. Urea and potassium thiocyanate (KSCN) are examples of agents that have been widely used for ELISA avidity [28,30].
Several factors can influence the result of the modified ELISA for avidity assay, among which we can mention: different types of plates, types of antigens, different types of substrates and, more importantly, the chaotropic agent used, its concentration, time and temperature incubation [8].
Recently, a study in our laboratory [26] analyzed the effects of using different antigens of Neisseria meningitidis (whole cell suspensions and Outer membrane vesicles), different chaotropic agents (urea and KSCN), and incubation temperatures (4 °C, 25 °C, and 37 °C) to evaluate the avidity of mice serum. However, this study did not evaluate the time of incubation with the chaotropic agents, which we believe is an important parameter. In addition, avidity in this study was assessed using the conventional ELISA method.
The avidity is a relevant criteria for humoral immunity against N. meningitidis and its index positively correlated with bactericidal activity [34], but a common ELISA-avidity does not allow characterization of which antigens present in the outer membrane vesicles (OMVs) of N. meningitidis induced antibodies with elevated binding strength, which would be interesting for the evaluation of potential candidate antigens that could be included in a new vaccine formulation .
Based on this, Immunoblot, a method commonly used to characterize proteins, has already been adapted to determine the avidity of antibodies against recombinant antigens (NP, RBD, and S1) of SARS-CoV-2 using human sera [1]. To accomplish our objectives, we modified the Immunoblot assay into a qualitative in-house Immunoblot avidity assay. We used the potassium thiocyanate (KSCN) 1.5 M as chaotropic agent, like in the modified ELISA [26].
Since the avidity assay has already been standardized using human sera, we were interested in evaluating the methodology previously described [1] in an experimental model, using sera from mice immunized with OMVs of N. meningitidis or with recombinant Receptor-binding domain (RBD) of SARS-CoV-2, with different adjuvants for each antigen. As we know, it is very important to evaluate an assay using more than one method, for example, in a case using immunodiagnostic. So, in addition to Immunoblot, we also included the Dot-Blot technique in the same assay.
In this context, this study aimed to analyze the effect of different incubation times to evaluate the avidity between the antibodies and the OMVs or RBD antigens. Not only this manuscript supports the use of Immunoblot to test avidity, but it also shows the relevance of the method for pointing which N. meningitidis antigens induced antibodies of higher avidity, an important information when predicting ideal antigens for vaccines.
2. Materials and Methods
2.1. Antigenic preparations and Serum
To achieve our goal, we used antigens and mice sera available in our laboratory (Laboratory for the Development of Vaccine Platforms, Adjuvants, and Monoclonal Antibodies, Immunology Center, Adolfo Lutz Institute) from previous studies [15,33], already approved by the Ethics Committee for the Use of Animals (CEUA/IAL number 02/2022 and CEUA IAL/Pasteur protocol number 06/2012.)
For N. meningitidis evaluation, we used sera from middle-aged female mice (5 to 7 animals per group) of the Swiss strain, that were immunized with three subcutaneous doses (with an interval of 15 days between the first and second dose and 20 days between the second and third dose) containing 2 µg of OMVs of the B:4:P1.9 strain complexed with aluminum hydroxide (AH) or dioctadecyldimethylammonium bromide in bilayer fragments (DDA-BF) as adjuvants. Control groups were immunized with OMVs, AH, or DDA-BF alone. The sera used in this study was collected 15 days after the 3rd dose [33]. We chose the 3rd dose of the animals' sera because it had a higher antibody titer, and it was expected that the affinity maturation process had already occurred [26].
Meanwhile, for SARS-CoV-2 assays, we used sera from adult Swiss mice (5 animals per group) immunized with two intramuscular doses (with a 15-day interval between doses) and two intranasal doses (with a 7-day interval between doses) formulated with 3 µg recombinant receptor binding domain (RBD) complexed with 10 µg/ml OMVs of N. meningitidis strain C:2a:P1.5 and AH. The samples evaluated in this study were collected 15 days after the 4th dose. We chose the 4th dose of animal serum because it also showed antibodies with good avidity and viral neutralization [15].
2.2. Immunoblot
The OMVs and RBD antigens were separated by polyacrylamide gel electrophoresis in the presence of sodium dodecyl sulfate (SDS-PAGE) in a 10% gel, and a molecular weight marker (25-245 kDa) (New England Biolabs, Ipswich, MA) was added to the electrophoresis run. Proteins were then transferred to 0.45 µm nitrocellulose membranes (Bio-Rad Laboratories, Hercules, CA) in a transfer buffer (25 mM Tris, 192 mM glycine, 20% methanol, pH 8.3).
The membranes were stained with Ponceau red to verify the transfer of proteins in the gel and washed with PBS to remove all dye (Sigma Chemical Company, St. Louis, MO). The membranes containing approximately 25 µg/ml OMVs and 1.6 µg/ml RBD on each strip were then blocked with 5% PBS-Leite (La Sereníssima, Argentina) for 2 hours at room temperature (20-25 °C) under agitation.
The strips were washed 5 times with PBS again and then incubated overnight, at 4 °C, with pooled sera of mice immunized with N. meningitidis OMVs and adjuvants (OMV+AH, OMV+DDA, OMV, AH and DDA groups), diluted in 2.5% PBS-milk at a 1:1000 dilution. Meanwhile, the pooled sera of mice immunized with SARS-CoV-2 RBD (RBD + OMV + AH, RBD, AH and OMV groups) were incubated at a 1:100 dilution.
The membranes were then washed with PBS and an anti-mouse IgG γ chain antibody (Kirkegaard & Perry Laboratories, KLP, Gaithersburg, MD) conjugated with peroxidase was added at 1:10,000 dilution and incubated for 2 hours at room temperature (20-25 °C).
For detection, the substrate 4-chloro-1-naphthol (Sigma-Aldrich) was added for 20 minutes at room temperature (20-25 °C). The enzymatic reaction was stopped by washing the strips with distilled water.
2.3. Dot-Blot
The Dot-Blot technique was incorporated into the same Immunoblot assay by adding 1 µl of each antigen (0,71 µg of RBD and 0,83 µg of OMVs) in its native state on top (above the bands proteins) of each dry strip, after the electrophoretic transfer. The following steps were the same as the avidity assay using the Immunoblot assay described below, as the assays were performed on the same membrane.
2.4. Strategy used in the study to assess avidity on nitrocellulose membranes
The strategy used in the study to evaluate avidity on nitrocellulose membranes is based on previous research carried out in our laboratory. These studies, using Dot-Blot and Immunoblot techniques carried out separately, characterized the immune response. This characterization was carried out for both experimental and clinical applications
In an experimental context, a recent study carried out in our laboratory used the Dot-Blot and Immunoblot technique to evaluate IgG antibodies in mice immunized with a new vaccine platform containing the RBD of SARS-CoV-2, adjuvanted with a mixture of DDA and Saponin [20]. In the diagnostic field, we have developed a simple method for transporting and storing monoclonal antibodies (Mabs) on filter paper, which is viable both at room temperature and at -20 °C. This method was used for serotyping N. meningitidis outer membrane antigens using ELISA and Dot-Blot, as a practical approach for epidemiological screening [14]. In addition, our laboratory has applied in a clinical and experimental studies using Dot-Blot and Immunoblot techniques [3,33].
The strategy adopted in our work consisted of combining two different antigenic preparations of the same pathogen on a single nitrocellulose membrane. To do this, we first carried out electrophoresis to separate the proteins, then transferred them to the nitrocellulose membrane (Immunoblot). The strips were then dried on Whatman filter paper and, once dry, we applied the two selected antigens to the top of the strip (Dot-Blot).
2.5. Anti-IgG Antibody Avidity Assessment
The IgG antibody avidity assay was based on the method described [1], which consisted of using nitrocellulose strips with recombinant SARS-CoV-2 proteins (NP, RBD, and S1) incubated with human sera followed by incubation with the chaotropic agent urea 6 M for 3 minutes.
The procedure followed the same steps as the conventional Immunoblot and Dot-Blot described above with the addition of the chaotropic agent potassium thiocyanate (Sigma Chemical Company, St. Louis, MO) [KSCN 1.5 M] [34]. Our assay after the membrane was incubated with the serum and before the addition of the mouse anti-IgG secondary antibody as illustrated in (Figure 1) the the chaotropic agent added in different incubation times and were evaluated for both the N. meningitidis OMVs and SARS-CoV-2 RBD assays to analyze interference in the binding force between the antigen and antibody at room temperature with nitrocellulose membrane.
After determining the ideal incubation time with the chaotropic agent for both pathogens, we investigate the antigens to which the antibodies remained bound after the addition of the chaotropic agent were analyzed for Neisseria meningitidis. Another point investigated in the SARS-CoV-2 assay, since only the RBD antigen was used, was related to different dilutions that could affect the binding strength between the antigen and the antibody. The assay was interpreted qualitatively based on the intensity (strong or weak) of the bands and staining spots.
3. Results and Discussion
3.1. Immunoblot and Dot-Blot for N. meningitidis OMVs
OMVs are composed of several antigens, including the Neisseria Adhesin A (NadA), Transferrin binding protein (Tbp), Porin A (PorA), Opacity protein (Opa), and Neisseria surface protein A (NspA), which have different molecular weights [25] and shown in (Figure 2).
To standardize the incubation time for the caotropic agent using a mouse IgG antibody avidity assay using the Immunoblot and Dot-Blot technique that has not previously been described in mice, we tested three different incubation times using the 1.5 M KSCN caotropic agent in the avidity assay (Figure 3).
Regarding the main objective of our work, the standardization of the incubation time was proceeded with the sera from the group immunized with OMV+AH, since it was the one that showed the highest antibody titers against OMVs and the highest avidity index with the KSCN chaotropic agent in the ELISA avidity [26,33].
It was observed that time affected the appearance of the bands and spots. Visually, 5 minutes was the most appropriate incubation time, as the bands and spots remained with stronger color intensity, while 20 minutes of incubation resulted in almost complete disappearance of the bands, as shown in (Figure 3).
Once the time was set, we tested the sera from other immunization schedules for 5 minutes incubation. This allowed us to check the specificity of the antibodies with avidity and to predict which antigens would induce antibodies with higher binding strength. We used mouse serum of groups immunized with OMVs + DDA, OMVs, AH, and DDA (Figure 4).
As expected, the controls did not recognize any bands on the Immunoblot. For the sera from animals immunized with OMVs + DDA, the antibodies recognized the NadA (170-190 kDa), PorA (46 kDa), and Tbp (80 kDa) bands. The 46 kDa/PorA band remained stained intensely after KSCN incubation, while the high molecular weight bands - 80 kDa/Tbp and NadA/170-190 kDa - showed less intense staining (Figure 4). We can assume that the PorA protein induces antibodies with higher binding strength, while NadA and Tbp induce antibodies with lower binding strength.
Sera from mice immunized with OMVs + AH recognized more antigens, including NadA (180 kDa), Tbp (85-95 kDa), PorA (46 kDa), PorB (33-42 kDa), and Opa (25-30 kDa) (Figure 4). However, the NadA, PorA, and Opa bands were more intensely stained, whereas Tbp and PorB were less intensely stained. This suggests that, when formulated with AH, the antigens PorA, NadA, and Opa induced antibodies with higher binding strength, whereas Tbp and PorB induced antibodies with lower binding strength.
In this sense, aluminum hydroxide as an adjuvant seems to contribute to the presentation of more antigens, the activation of the immune response, and the induction of antibodies with higher binding strength, as observed in previous studies [6,26,33].
The Porin A is one of the trimeric proteins located on the outer membrane of N. meningitidis [25]. PorA has been described in the literature as an immunogenic protein that induces the production of antibodies with elevated avidity and bactericidal activity [18].
As for NadA, it is a protein exposed on the surface of the bacterium that adheres to the cell epithelium and provokes a strong response of protective bactericidal antibodies and cross-reactivity with strains of different serosubtypes. Moreover, the expression of NadA was associated with 50% of the strains causing meningococcal disease [7]. Anti-NadA antibodies were detected only in the presence of AH and ainduced antibodies with high avidity (Figure 4). Interestingly, NadA is a high molecular weight antigen (Figure 2) that induces cross-reactivity against different meningococcal serosubtypes and contributes to protection against multiple strain variants, making it an important antigen for vaccine preparations [7,18].
Opa is a protein located on the outer membrane of meningococci that adheres to and invades the surface of epithelial cells. Although it has been reported in the literature to be antigenically variable, this antigen was able to induce high avidity IgG antibodies (Figure 4). This reinforces the fact that antibodies can block bacterial adhesion and invasion to host cells, thereby favoring neutrophil phagocytic activity, as described by several authors [13,17,23,25,36].
During our study, no bactericidal assay was performed with the mice sera, but our results showed that the antibodies triggered by OMV+AH are likely to have high avidity against more than one antigen (PorA, Opa and NadA), which are immunogenic and induced bactericidal antibodies in other manuscripts, as described by some authors, which is important for protecting against meningococcal disease [16,25,34]. On the other hand, OMV+DDA induced antibodies that remained bounded with PorA and, which is a relevant antigen as well, like described above. OMVs, without any adjuvant, the antibodies show reactivity and good avidity only against PorA, confirming that the presence of adjuvants favors the presentation of more than one antigen of the bacterium and induces IgG antibodies of high avidity, as our data show.
In general, several studies have shown that aluminum hydroxide in vaccines induces high concentrations and prolonged antibody responses. AH adjuvant is also included in the formulation of other vaccines against other pathogens and diseases, such as hepatitis A and B, tetanus, HPV, diphtheria and tetanus toxoid (DTP), anthrax, rabies, Haemophilus influenzae type b, and also against diphtheria, tetanus and pertussis (DTaP), which are recommended for their proven efficacy and safety [11,27]. In addition, other authors have reported that aluminum hydroxide as an adjuvant in vaccine preparations leads to the generation of memory B cells and effector T cells [22], which likely contributed to the generation of highly avidity antibodies against several antigens in the OMVs + AH group.
Among meningococcal vaccines licensed for human use that contain aluminum hydroxide in their formulation, we have Bexsero®, a vaccine for the prevention of disease caused by N. meningitidis from serogroup B. This vaccine has been shown to be safe, effective, and highly immunogenic in children and infants. It contains in its formulation the recombinant antigens heparin-binding protein (NHBA), factor H binding protein (fHbp), NadA, and OMVs expressing Porin A, and is recommended for clinical use in several countries, including high-risk groups, adolescents, routine childhood immunization, and outbreak control [2,24,27].
For meningococcal disease prevention, interestingly, that more than one epitope is recognized for antibodies of different specificities, provided that it may favor opsonization and subsequent lysis by the complement system, as described by some authors [25].
Considering the Dot-Blot (Figure 4), the area remained stained, probably due to the high avidity antibodies remaining on the membrane. This confirms our results regarding the high avidity of mice IgG antibodies and the contribution of the adjuvants. However, we believe that Immunoblot would be a more interesting technique, allowing a better characterization of the response by separating the antigens according to molecular weight.
3.2. Standardization of SARS-CoV-2 Immunoblot and Dot-Blot for RBD
We used the recombinant RBD antigen that was provided by the Immunobiotechnology Laboratory of the Immunology Center of the Adolfo Lutz Central Institute. The recombinant RBD protein was expressed, and the antigen was purified according to [9]. As described in the literature, the recombinant RBD antigen of SARS-CoV-2 used for immunization is approximately 32 kDa [31] (Standlbauer et al., 2020). We also standardized the assay for SARS-CoV-2 using sera from mice immunized with RBD + OMVs + AH from previous work in our laboratory [15].
For the serum from the RBD + OMV + AH immunization scheme, we also tested different incubation times for 5, 10, and 20 minutes using the chaotropic KSCN as shown in (Figure 5). Similarly to OMV, 5 minutes seems to be ideal, while in a 20-minute incubation time the band almost disappeared from the membrane and the dot disappears completely from the membrane.
Since 5 minutes seemed to be the ideal incubation time and immunization with RBD resulted in the recognition of only one band, it would be more reliable to observe the interference of the dilution with this sera. For this purpose, different dilutions were used: 1/50, 1/100, 1/200, and 1/400, to evaluate whether it would interfere with the binding strength as shown in the (Figure 6).
Unlike the N. meningitidis study samples, where we were able to evaluate the results of the assay at a 1/1000 dilution with KSCN within 5 minutes, the SARS-CoV-2 study samples required a dilution of 1/100 (Figure 6). The type of immune response will vary depending on the components of the antigenic preparation (antigens and adjuvants used), the concentration, route and number of doses administered. That means that the results may be different and non-replicable depending on the various factors involved, so it is important to standardize the protocol within each study. In this sense, it is necessary to characterize the response before moving to a functional assay, adapting the assay according to the samples used and the experimental design. This again highlights the importance of standardizing the avidity assay [8].
In our study, a dilution of 1/50 to 1/200 would be appropriate for evaluating the avidity of the Immunoblot and a dilution of 1/50 would be appropriate for evaluating the avidity of the dot assay only, because at a dilution of 1/400, the band and the dot disappears completely in both the Immunoblot and Dot-Blot assays (Figure 6). The study [1] used another chaotropic agent, urea 6 M, to evaluate the avidity of antibodies present in the serum of humans infected with COVID-19. He showed that the use of different concentrations of urea can result in a high degree of variation in the avidity assay, thus demonstrating how variables can affect the results of a study.
However, at the 1/100 and 1/200 dilutions, the color of the band became less intense, suggesting that as the serum was diluted, antibodies with good avidity disappeared. Compared to the 1/50 dilution, the band was more intensely stained, suggesting that high avidity antibodies remain at this dilution (Figure 6). Similarly, the results of the characterization of these sera in our previous study [15] showed, by ELISA-Avidity, that the antibodies had an intermediate avidity index, close to the high avidity threshold, as well as a high neutralization index; together, these results show that the antibodies were functional. Our results are consistent with those obtained previously because, although it is not possible to quantify the avidity index by Immunoblot, the permanence of the stained band suggests that the antibodies had good binding strength to the antigen.
For COVID-19, it has already been correlated that antibodies with high avidity are good indicators of viral neutralization [4]. Also, some authors showed that three doses of mRNA vaccines against the Omicron variant (which is a variant of concern due to its high mutability rate) increased the avidity of antibodies directed against the RBD (where most mutations occur), thus ensuring protection against different variants of SARS-CoV-2 [10,21]. Seeing the relevance of avidity in the context of COVID-19 prevention, it is interesting to standardize different avidity assays that can be used in different conditions and locations. In this way, the avidity technique, which is simpler than neutralization, could be disseminated and provide more information on the immune response against SARS-CoV-2.
4. Conclusions
Our results showed that KSCN 1.5 M can be used for Immunoblot and Dot-Blot-avidity, at an incubation time of 5 minutes, using OMVs from N. meningitidis and RBD from SARS-CoV-2 as antigens. Thus, titration of sera from different immunization protocols is required before testing avidity, to assess the adequate dilution. Considering immunization with N. meningitidis OMVs, the Immunoblot-avidity is particularly interesting to suggest the proteins that induced antibodies of higher avidity. In our study, AH was a better adjuvant than DDA to support the recognition of various antigens with higher avidity. Extending this assay to other pathogens of public health concern is imperative.
Author Contributions
Conceptualization, E.D.G.; methodology, A.F.S, E.D.G.; (RBD) antigen production H.H.M.C, C.R.P ; investigation, A.F.S.,A.I.P and E.D.G; formal analysis, A.F.S. and E.D.G.; resources, E.D.G and C.R.P.; data curation, A.F.S. and E.D.G.; writing—original draft preparation, A.F.S, A.I.P. and E.D.G.; writing—review and editing, A.F.S., A.I.P. and E.D.G.; supervision, E.D.G.; project administration, E.D.G.; funding acquisition, E.D.G and C.R.P. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by Fundação de Amparo à Pesquisa do Estado de São Paulo (FAPESP) grants numbers 2012/15568-0, 2018/04202-0, 2021/11936-3, 2022/05566-1; Conselho de Desenvolvimento Científico e Tecnológico (CNPq), grant number 305301/2022-5; Coordenação de Aperfeiçoamento de Pessoal de Nível Superior (CAPES), finance code 001; and Secretaria de Estado da Saúde de São Paulo (SES/SP).
Institutional Review Board Statement
The animal study protocol was approved by the Ethics Committee of Animal Use of Adolfo Lutz Institute (CEUA/IAL) numbers 06/2012 (N. meningitidis immunization) and 02/2022 (SARS-CoV-2 immunization),CEUA IAL/Pasteur protocol number 2012.
Informed Consent Statement
Not applicable.
Data Availability Statement
Data from this article is available upon reasonable request with the corresponding author.
Acknowledgements
The authors would like to thank Dr Florian Krammer (Icahn School of Medicine, Mount Sinai Hospital, New York, USA) for donating the plasmids for RBD production; and MSc. Gabriela Trzewikoswki de Lima, for providing sera samples used in her Master's thesis.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Bauer, G. et al. The challenge of avidity determination in SARS-CoV-2 serology. J. Med. Virol., 2021, 93, 3092–3104. [CrossRef] [PubMed]
- Bekkat-Berkani et al. Public health perspective of a pentavalent meningococcal vaccine combining antigens of MenACWY-CRM and 4CMenB. J. Infect., 2022, 85, 481–491. [CrossRef] [PubMed]
- Belo, E.F.T.; Farhat, C.K.; De Gaspari, E.N. Comparision of dot-ELISA and standard ELISA for detection of Neisseria meningitidis outer membrane complex-specific antibodies. Braz. J. Infect. Dis., 2010, 14, 35–40. [Google Scholar] [PubMed]
- Benner, S.E. et al. SARS-CoV-2 antibody avidity responses in COVID-19 patients and convalescent plasma donors. J. Infect. Dis., 2020, 222, 1974–1984. [CrossRef]
- Brady, A. M.; Unger, E. R.; Panicker, G. Description of a novel multiplex avidity assay for evaluating HPV antibodies. J. Immunol. Methods, 2017, 447, 31–26. [Google Scholar] [CrossRef]
- Brito, L.T. et al. Study of different routes of immunization using outer membrane vesicles of Neisseria meningitidis B and comparison of two adjuvants. Vaccine, 2020, 38, 7674–7682. [CrossRef]
- Comanducci, M. et al. NadA, a novel vaccine candidate of Neisseria meningitidis. J. Exp. Med., 2002, 195, 1445–1454. [CrossRef]
- Correa, V.A.; Rodrigues, T.S.; Portilho, A.I.; Trzewikoswki de Lima, G.; De Gaspari, E. Modified ELISA for antibody avidity evaluation: The need for standardization. Biomed. J., 2021, 44, 433–348. [Google Scholar] [CrossRef]
- Da Costa, H.H.M. et al. RBD and Spike DNA-Based Immunization in Rabbits Elicited IgG Avidity Maturation and High Neutralizing Antibody Responses against SARS-CoV-2. Viruses, 2023, 15, 555. [CrossRef]
- Da Porto, F. et al. Antibody avidity and neutralizing response against SARS-CoV-2 omicron variant after infection or vaccination. J. Immunol. Res., 2022, 2022, 1–9. [CrossRef]
- Eickhoff, T.C.; Myers, M. Workshop summary. Aluminum in vaccines. Vaccine, 2002, 20, S1–S4. [Google Scholar] [CrossRef] [PubMed]
- Eisen, H. N. Affinity enhancement of antibodies: how low-affinity antibodies produced early in immune responses are followed by high-affinity antibodies later and in memory B-cell responses. Cancer Immunol. Res., 2014, 2, 381–392. [Google Scholar] [CrossRef] [PubMed]
- Estabrook, M.M.; Zhou,D.; Apicella, M.A.A. Nonopsonic phagocytosis of group C Neisseria meningitidis by human neutrophils. Infect. and Immun., 1998, 66, 1028–1036. [CrossRef] [PubMed]
- Ferraz, A. S. et al. Storage and stability of IgG and IgM monoclonal antibodies dried on filter paper and utility in Neisseria meningitidis serotyping by Dot-blot ELISA. BMC Infect. Dis., 2008, 8, 1–8. [CrossRef]
- Gaspar, E.B.; De Gaspari, E. Avidity assay to test the functionality of anti-SARS-Cov-2 antibodies. Vaccine, 2021, 39, 1473. [Google Scholar] [CrossRef]
- Granoff, D.M., et al. A modified enzyme-linked immunosorbent assay for measurement of antibody responses to meningococcal C polysaccharide that correlate with bactericidal responses. Clin. Diagn. Lab. Immunol., 1998, 5, 479–485. [CrossRef]
- Heyderman, R. S. et al. Neutrophil response to Neisseria meningitidis: inhibition of adhesion molecule expression and phagocytosis by recombinant bactericidal/permeability-increasing protein (rBPI21). J. Infect. Dis., 1999, 179, 1288–1292. [CrossRef]
- Humphries, H.E. et al. Multivalent liposome-based vaccines containing different serosubtypes of PorA protein induce cross-protective bactericidal immune responses against Neisseria meningitidis. Vaccine, 2006, 24, 36–44. [CrossRef]
- Ise, W.; Kurosaki, T. Regulation of Plasma Cell Differentiation. B Cells in Immun. Tol., 2020, 1254, 63–74. [Google Scholar] [CrossRef]
- Lima, G.G.; Portilho, A.I.; De Gaspari, E. Adjuvants to increase immunogenicity of SARS-CoV-2 RBD and support maternal–fetal transference of antibodies in mice. Pathog. Dis., 2022, 80, ftac038. [Google Scholar] [CrossRef]
- Luczkowiak, J. et al. Cross neutralization of SARS-CoV-2 omicron subvariants after repeated doses of COVID-19 mRNA vaccines. J. Med.Virol., 2023, 95, e28268. [CrossRef]
- Marrack, P.; Mckee, A.S.; Munks, M. W. Towards an understanding of the adjuvant action of aluminum. Nat. Rev. Immunol., 2009, 9, 287–293. [CrossRef]
- McNeil, G.; Virji, M. Phenotypic variants of meningococci and their potential in phagocytic interactions: the influence of opacity proteins, pili, PilC and surface sialic acids. Microb. Pathog., 1997, 22, 295–304. [Google Scholar] [CrossRef] [PubMed]
- Nompari, L. et al. Quality by design approach in the development of an ultra-high-performance liquid chromatography method for the Bexsero meningococcal group B vaccine. Talanta, 2018, 178, 552–562. [CrossRef] [PubMed]
- Pollard, A. J.; Frasch, C. Development of natural immunity to Neisseria meningitidis. Vaccine, 2001, 19, 1327–1346. [Google Scholar] [CrossRef] [PubMed]
- Portilho, A.I.; Santos, J.S.; Trzewikoswki de Lima, G.; Lima, G.G. ; De Gaspari,, E. Study of avidity-ELISA: Comparison of chaotropic agents, incubation temperature and affinity maturation after meningococcal immunization. J. Immunol. Methods, 2023, 512, 11337–2023. [Google Scholar] [CrossRef]
- Principi, N.; Esposito, S. Aluminum in vaccines: Does it create a safety problem? . Vaccine, 2018, 36, 5825–5831. [Google Scholar] [CrossRef] [PubMed]
- Pullen, G. R.; Fitzgerald, M. G.; Hosking, C. S. Antibody avidity determination by ELISA using thiocyanate elution. J. Immunol. Methods, 1986, 86, 83–87. [Google Scholar] [CrossRef]
- Reddy, S.B et al. Affinity antibodies to Plasmodium falciparum merozoite antigens are associated with protection from malaria. PLoS One, 2012, 7, 232242. [CrossRef]
- Smolander, H. et al. A novel antibody avidity methodology for rapid point-of-care serological diagnosis. J. Virol. Methods, 2010, 166, 86–91. [CrossRef]
- Standlbauer, D. et al. SARS-CoV-2 seroconversion in humans: a detailed protocol for a serological assay, antigen production, and test setup. Curr. Protoc. Microbiol., 2020, 57, e100. [CrossRef]
- Steward, M.W; Stanley, C.M.; Dimarchi, R.; Mulcahy, G.; Doel, T.R. High-affinity antibody induced by immunization with a synthetic peptide is associated with protection of cattle against foot-and-mouth disease. Immunology, 1991, 72, 79. [Google Scholar]
- Trzewikoswki de Lima, G.; Rodrigues, T.S.; Portilho, A. I.; Correa, V.A.; Gaspar, E.B.; De Gaspari, E. Immune responses of meningococcal B outer membrane vesicles in middle-aged mice. Pathog. Dis., 2020, 78, ftaa028. [Google Scholar] [CrossRef]
- Vermont, C.L.; Van Dijken, H.H.; Van Limpt, C.J.; De Groot, R.; Van Alphen, L.; Van Den Dobbelsteen, G.P. Antibody avidity and immunoglobulin G isotype distribution following immunization with a monovalent meningococcal B outer membrane vesicle vaccine. Infect. Immun., 2002, 70, 584–590. [Google Scholar] [CrossRef] [PubMed]
- Victora, G.D.; Nussenzweig, M.C. Germinal centers. Annu. Revi. Immunol., 2022, 40, 413–442. [Google Scholar] [CrossRef] [PubMed]
- Virji, M. et al. The N-domain of the human CD66a adhesion molecule is a target for Opa proteins of Neisseria meningitidis and Neisseria gonorrhoeae. Mol. Microbiol., 1996, 22, 929–939. [CrossRef]
- Watson, P.S.; Turner, D.P.J. Clinical experience with the meningococcal B vaccine, Bexsero®: Prospects for reducing the burden of meningococcal serogroup B disease. Vaccine, 2015, 34, 875–880. [Google Scholar] [CrossRef]
- Weynants, V. E. et al. Additive and synergistic bactericidal activity of antibodies directed against minor outer membrane proteins of Neisseria meningitidis. Infect. and Immun., 2007, 75, 5434–5442. [CrossRef]
Figure 1.
The figure describes the methodology used in this investigation, where Dot-Blot and Immunoblot methods were used on nitrocellulose membranes, with mice serum as primary antibody and anti-IgG antibody specific to mouse as the secondary antibody labelled with peroxidase. For the avidity assay, KSCN was used on the nitrocellulose membrane with the antigen in its native "Dot-Blot" form and in its denatured "Immunoblot" form.
Figure 1.
The figure describes the methodology used in this investigation, where Dot-Blot and Immunoblot methods were used on nitrocellulose membranes, with mice serum as primary antibody and anti-IgG antibody specific to mouse as the secondary antibody labelled with peroxidase. For the avidity assay, KSCN was used on the nitrocellulose membrane with the antigen in its native "Dot-Blot" form and in its denatured "Immunoblot" form.
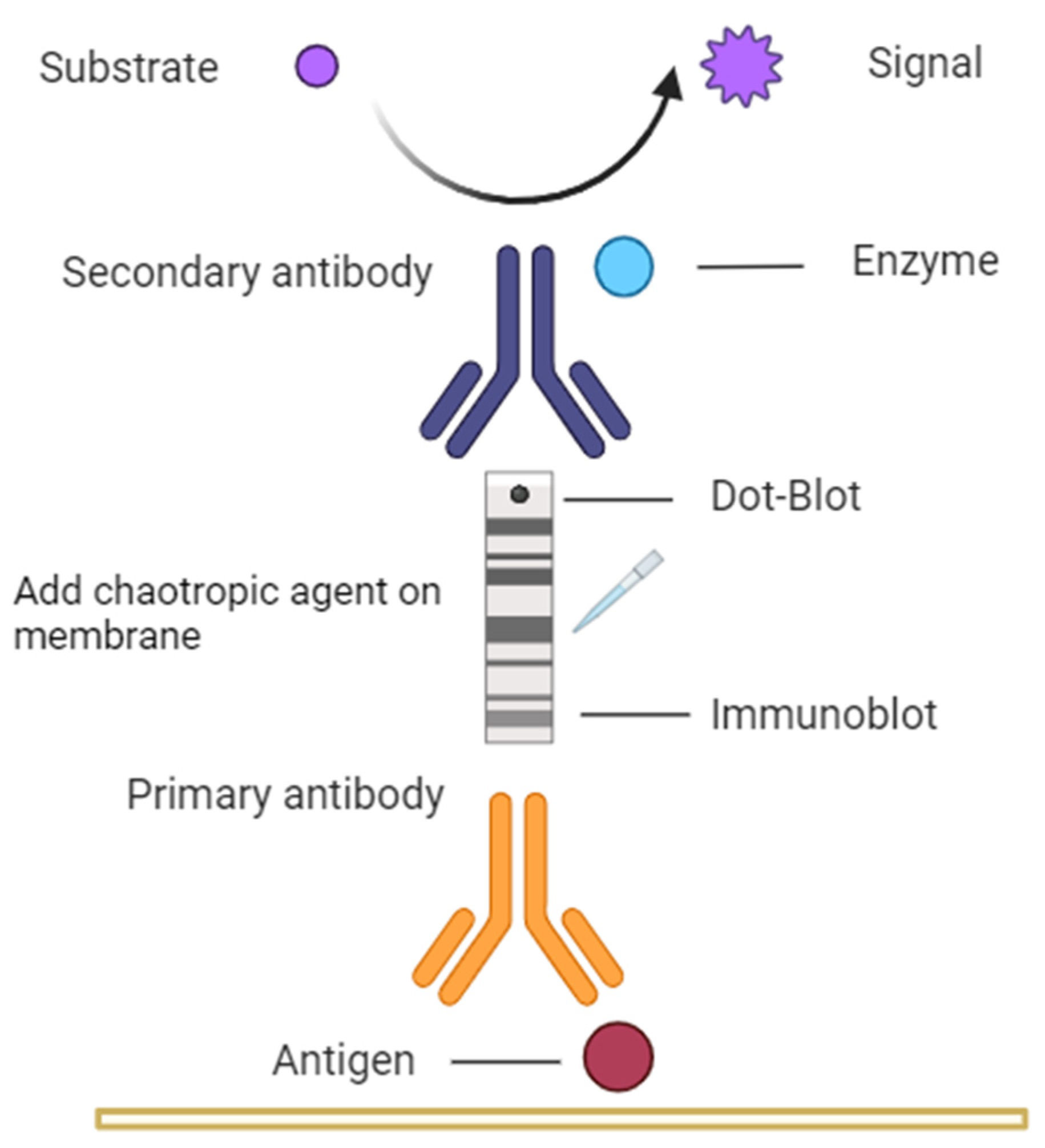
Figure 2.
- Characterization of OMVs by SDS-PAGE (10% polyacrylamide). On the left, molecular weight; on the right, antigens presented in N. meningitidis OMVs as suggested in the literature.
Figure 2.
- Characterization of OMVs by SDS-PAGE (10% polyacrylamide). On the left, molecular weight; on the right, antigens presented in N. meningitidis OMVs as suggested in the literature.
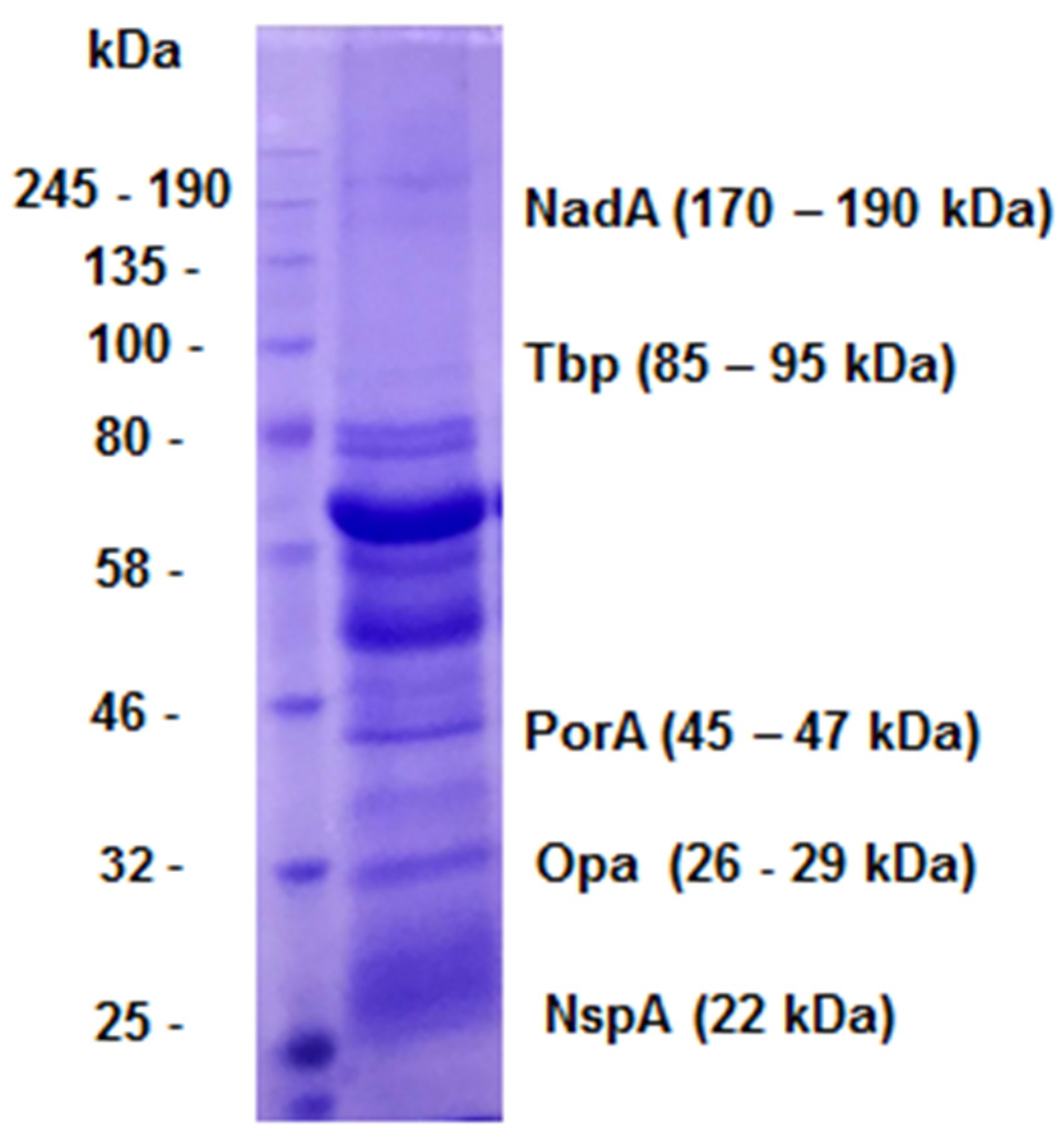
Figure 3.
- Evaluation of incubation time with chaotropic agent for Immunoblot and Dot-Blot techniques. The nitrocellulose membranes containing OMVs of N. meningitidis incubated with KSCN for different times (5, 10, and 20 minutes). C refers to the control (the conventional assay without the addition of the chaotropic agent). The molecular weight marker is on the left.
Figure 3.
- Evaluation of incubation time with chaotropic agent for Immunoblot and Dot-Blot techniques. The nitrocellulose membranes containing OMVs of N. meningitidis incubated with KSCN for different times (5, 10, and 20 minutes). C refers to the control (the conventional assay without the addition of the chaotropic agent). The molecular weight marker is on the left.
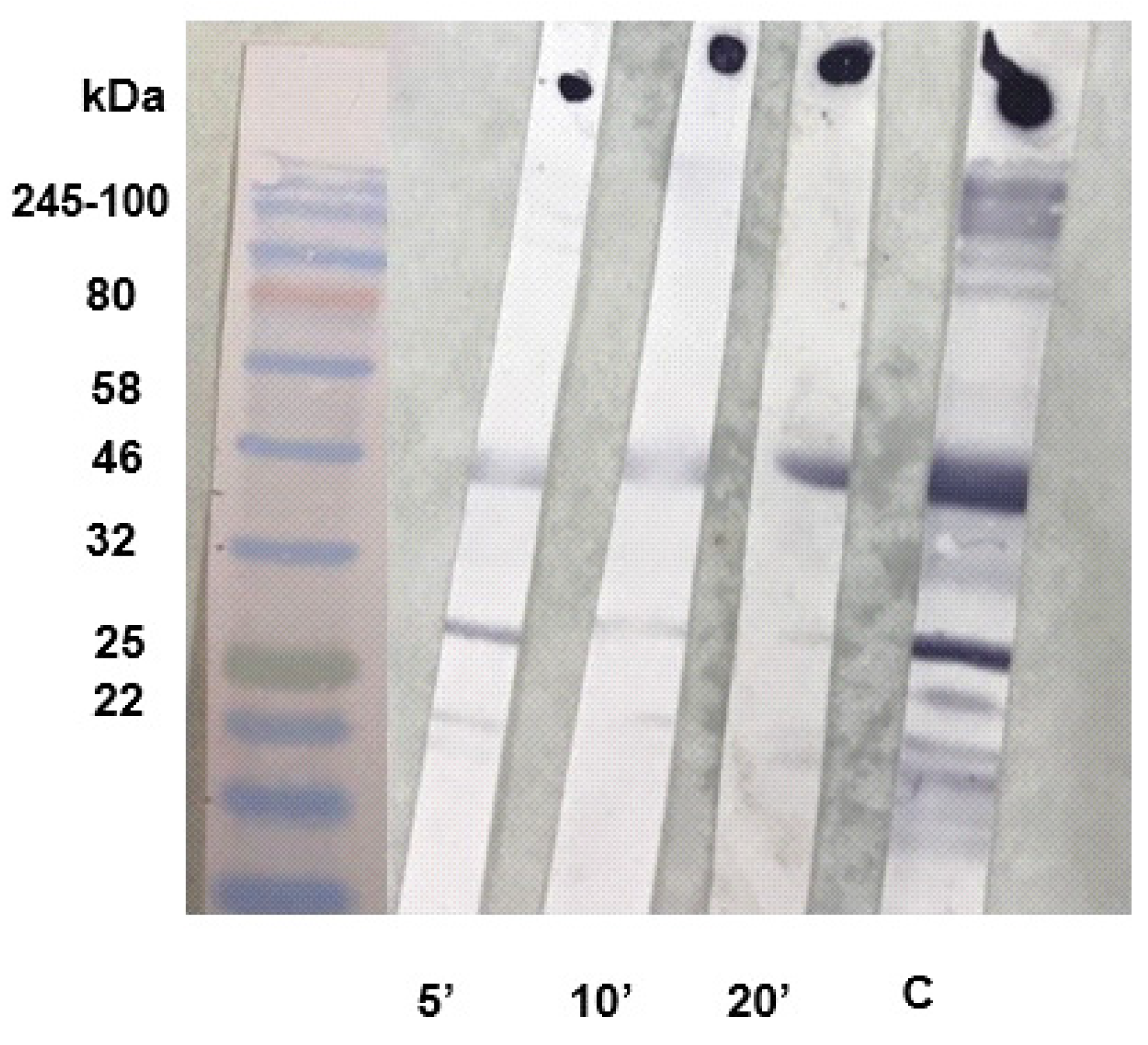
Figure 4.
- Evaluation of different sera without and with the KSCN. We can see the difference in reactivity of the different sera evaluated. For each group, the left strip was not incubated with KSCN and the right one was. In the sera of mice immunized with OMV+AH, we observed a higher antibody avidity for several antigens when compared with OMV+DDA, where only PorA remained intensely stained. The sera from control groups AH and DDA did not recognize any antigen in the band without the KSCN.
Figure 4.
- Evaluation of different sera without and with the KSCN. We can see the difference in reactivity of the different sera evaluated. For each group, the left strip was not incubated with KSCN and the right one was. In the sera of mice immunized with OMV+AH, we observed a higher antibody avidity for several antigens when compared with OMV+DDA, where only PorA remained intensely stained. The sera from control groups AH and DDA did not recognize any antigen in the band without the KSCN.
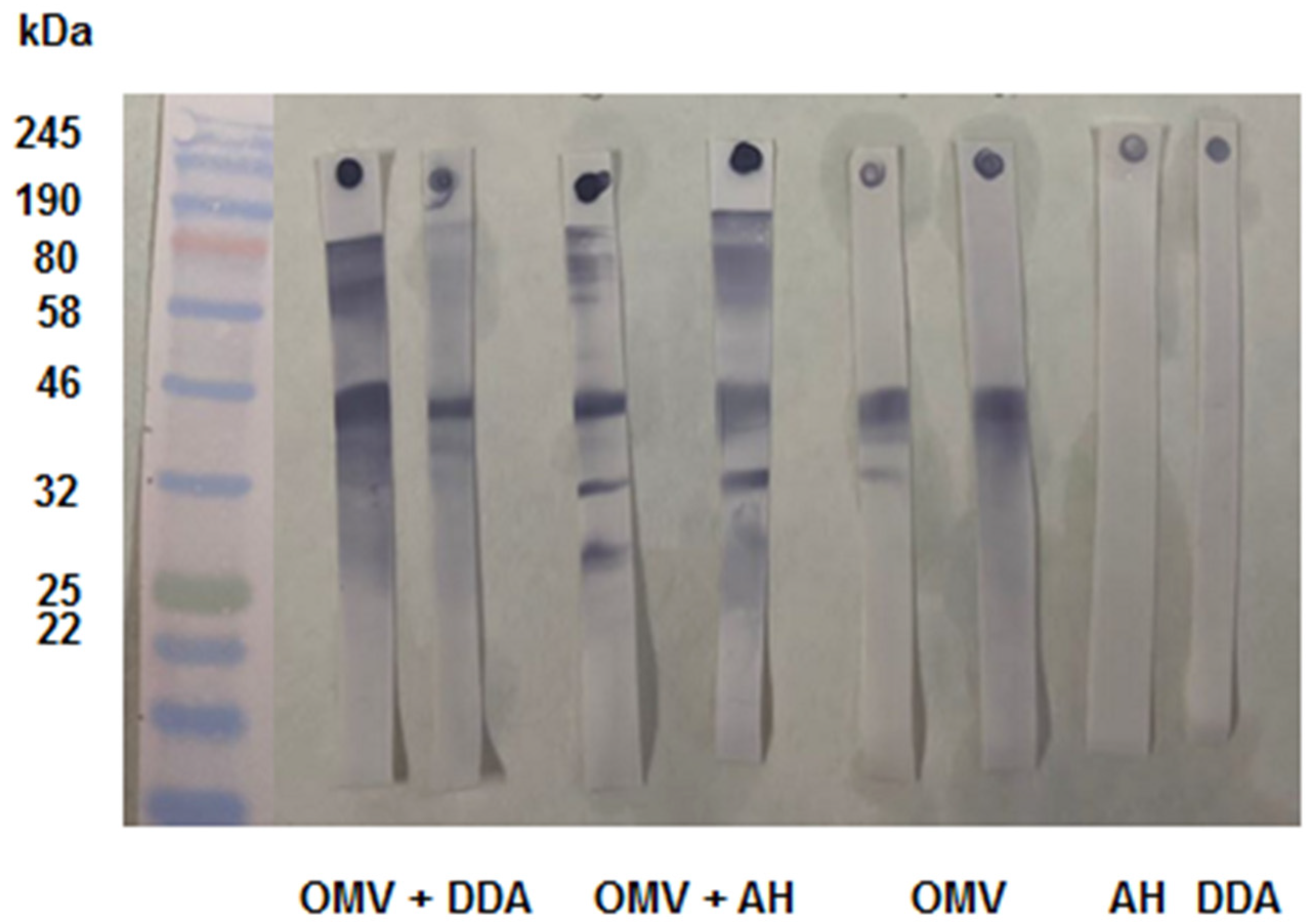
Figure 5.
– Evaluation of incubation time with KSCN for Immunoblot and Dot-Blot techniques. We observed that the nitrocellulose membranes containing RBD of SARS-CoV-2 showed different results at different incubation times (5, 10, and 20 minutes). C refers to the control (the conventional assay without the addition of the chaotropic agent). The molecular weight marker is on the left.
Figure 5.
– Evaluation of incubation time with KSCN for Immunoblot and Dot-Blot techniques. We observed that the nitrocellulose membranes containing RBD of SARS-CoV-2 showed different results at different incubation times (5, 10, and 20 minutes). C refers to the control (the conventional assay without the addition of the chaotropic agent). The molecular weight marker is on the left.
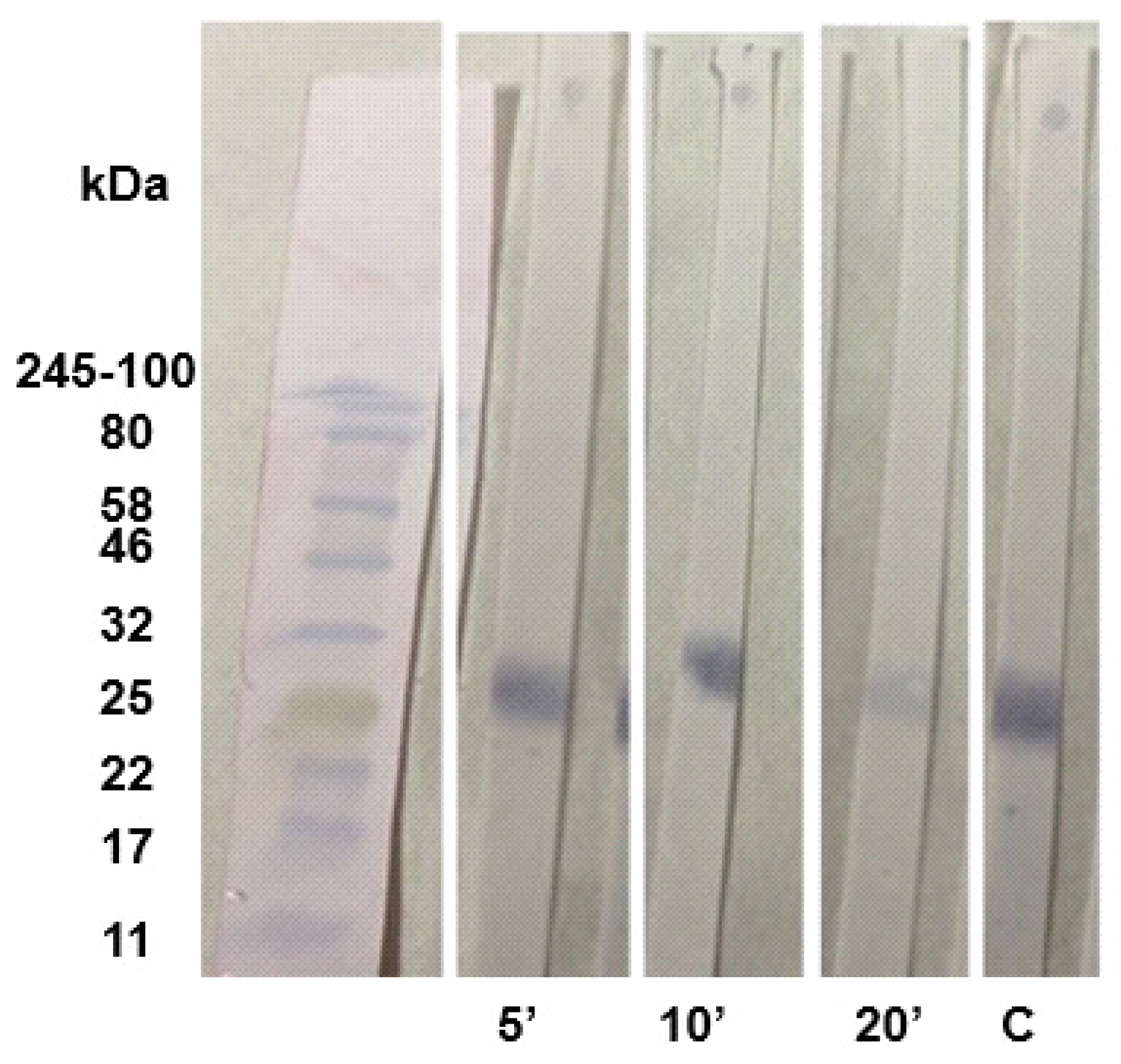
Figure 6.
- Nitrocellulose membranes with RBD antigen incubated with different concentrations of anti-RBD sera (1/50, 1/100, 1/200, and 1/400). For each dilution, the strip on the left was not incubated with KSCN and the strip on the right was.
Figure 6.
- Nitrocellulose membranes with RBD antigen incubated with different concentrations of anti-RBD sera (1/50, 1/100, 1/200, and 1/400). For each dilution, the strip on the left was not incubated with KSCN and the strip on the right was.

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



