
Submitted:
28 December 2023
Posted:
29 December 2023
You are already at the latest version
Abstract
The use of biostimulants consisting of plant growth promoting rhizobacteria (PGPR) is rapidly expanding in horticulture in recent years. In the current study, a novel mix of six Bacillus sp. strains was tested as a PGPR biostimulant in two experiments with zucchini squash (Cucurbita pepo L.) cultivated in the greenhouse soil and in an open field, respectively. In both experiments, seeds of the local landrace ‘Kompokolokytho’ and the commercial hybrid ‘ARO-800’ were either inoculated or non-inoculated with the PGPR biostimulant. The application of the six Bacillus sp. strains increased both the vegetative growth and the yield of zucchini squash, and these effects were associated with significantly higher shoot phosphorus levels in both experiments and both genotypes. Furthermore, at the end of the cultivation, the colony forming units of Bacillus sp. were appreciably higher in plants originating from inoculated compared to non-inoculated seeds, indicating that the tested mix of Bacillus sp can be successfully applied through seed inoculation. ‘ARO-800’ produced more vegetative and fruit biomass than ‘kompokolokytho’ under greenhouse cropping conditions, while in the open-field crop both genotypes performed equally, presumably because the local landrace is traditionally grown in open fields, and thus it is not well adapted to greenhouse cropping.
Keywords:
biostimulants
; PGPR
; integrated crop management
; greenhouse
; zucchini squash
; landrace.
1. Introduction
Nowadays, the deleterious effects of climate change on agricultural systems, coupled with the escalating global population, are anticipated to elevate global hunger by 30% by the year 2050 [1]. Considering this complex scenario, agriculture has to face the dual challenge of fulfilling the escalating demand for food production while concurrently alleviating its environmental impact on natural ecosystems and human health [2]. Currently, a spectrum of both traditional and innovative eco-friendly management practices is being widely implemented to augment crop yields and increase food safety standards [3].
Horticulture, and especially vegetable production, heavily depends on external inputs of mineral nutrients, primarily in the form of synthetic fertilizers. The relatively low nutrient use efficiency exhibited by many vegetable crops leads to an excess application of nutrients beyond what the crops actually require, thereby causing substantial environmental impacts [4,5]. This excess application not only raises concerns about resource sustainability but also contributes to environmental issues such as nutrient runoff, soil degradation and water pollution [6]. Addressing the nutrient use efficiency in vegetable production is crucial for mitigating these environmental challenges and promoting sustainable agricultural practices. Recognizing these concerns, the European Commission has set an ambitious goal to substitute 30% of synthetic fertilizers with greener alternatives by 2050 [7]. Plant biostimulants, acknowledged as natural products, have emerged as eco-friendly tools for the reduction of synthetic fertilizer use, as they can enhance nutrient use efficiency [8]. Their application not only contribute in reducing reliance on chemical fertilizers but also helps plants cope with stress induced by climate change [9,10]. Therefore, plant biostimulants hold the promise of fostering a more sustainable agriculture, ensuring crop yields under lower inputs, and represent a crucial step towards environmental responsibility [8,11,12,13,14].
Non-pathogenic soil microorganisms with beneficial effects on plant growth and crop production can be used as microbial biostimulants in commercial crops [15]. Bacterial strains living in the rhizosphere or even within the root tissues with beneficial effects on plant growth are generally termed “plant growth promoting rhizobacteria” (PGPR). Due to their beneficial effects on crop yield, several PGPR have been licensed for commercial distribution as biostimulants [16]. PGPR are usually applied either by inoculating the seeds before sowing or by soil drenching after planting or plant emergence.
The treatment of seeds with microbial preparations is based on specific techniques, which may have different effectiveness–success in coating the seeds, depending on the size, weight, and texture of each seed. Biopriming is a seed-presoaking technique along with the inoculation of beneficial microorganisms. It combines both the biological agent (microorganisms) and physiological soaking (seed hydration) phase [17]. The exudates released from the seed may serve as source of energy and nutrients to the biocontrol agents during biopriming [18], thus facilitating the proliferation and the colonization of these biocontrol agents over the surface of seeds which facilitate the nutrient/water uptake. Film coating seed treatment is based on seed coating with a thin film, which contains the active substance-microorganisms and one or two inert materials that function either as a carrier to preserve the microorganisms and/or as adhesives. This film is created after treating the seeds with a liquid-dense solution or suspension [19].
Zucchini squash (Cucurbita pepo L.), a member of the Cucurbitaceae family, is a popular vegetable worldwide [20] of great economic importance due to its high nutritional value [21]. Zucchini squash is widely cultivated both in fields and in greenhouses in the Mediterranean region [22] and in arid climatic zones characterized by limited water resources and hot weather conditions as well as in soils with low organic matter content [23].
Taking the above into consideration, an investigation was designed to assess the effectiveness of a novel inoculant contained the six Bacillus sp. strains in improving nutrient uptake and concomitantly plant growth and yield when used as a PGPR biostimulant through biopriming of zucchini squash seeds.
2. Materials and Methods
2.1. Biostimulant Application
2.1.1. Microbial Inoculant
The microbial inoculant used as biostimulant contained the following six Bacillus species: Bacillus subtilis, Bacillus pumilus, Bacillus megaterium, Bacillus amyloliquefaciens, Bacillus velezensis, and Bacillus licheniformis at a concentration of 1*1012CFU (Colony Forming Unit) ml−1 of each one. Every species was, separately, cultivated at 37 oC, aerobically, in a liquid growth medium containing 0.1g L−1 K2HPO4, 0.4 g L−1 KH2PO4, 0.01 g L−1 FeSO4 x 7 H2O, 0.20 g L−1 MgSO4 x 7 H2O, 0.20 g L−1 MnSO4, 0.10 g L−1 NaCl, 0.02 g L−1 CaCl2 x 2 H2O and 2g L−1 glucose. The cultures were stopped 10 h after entering stationary growth phase. Then the cultures were centrifuged and the biomass from each culture was redissolved with 0.01% PBS (Phosphate Buffered Saline). The six biomasses were mixed and subsequently diluted in PBS, so that the final microbial inoculant contained bacteria of the Bacillus genus at a concentration of 6*1012 CFU L−1.
2.1.2. Seed Treatment with Microbial Inoculant
Zucchini seeds were treated using the biopriming technique. Seeds were placed in a sealed plastic bag, covered with the liquid microbial inoculant and hydrated at 15-20 oC for 1 h. After that, seeds were air dried for 24 h. At the end of the process, the seeds were checked for both their germination and their microbial load concentration. Each zucchini seed possessed with bacteria of the Bacillus genus at a concentration of 1*105 CFU.
2.1.3. Estimation of Root Colonization by Bacillus sp.
At the end of the greenhouse experiment, the roots of one randomly selects plant per treatment were placed in a sealed plastic bag and immediately transferred to the laboratory to determine the concentration of Bacillus sp. bacteria on the surface and interior of the roots. In the laboratory, the zucchini roots were rinsed with sterile distilled water to remove adhering soil particles. Then, roots were cut into smaller segments (about 1-2 cm in length) and placed in segments in sterile containers and added 10 mL of 0.01 M phosphate buffer solution (PBS) to the containers. Dilutions of the root homogenate were conducted by transferring known volumes into sterile tubes containing sterile PBS (10−1, 10−2, 10−3, etc.). An amount of 0.1 mL from each dilution was transferred onto separate sterile agar plates with TSA medium and incubated plates were left in a microbiology incubator, aerobic, at 37 °C for three days, to allow colonies’ growth. After incubation, plates were examined, and Bacillus colonies were counted on each plate.
2.2. Plant Material and Experimental Layout
Two consecutive experiments were conducted at the Laboratory of Vegetable Production at the Agricultural University of Athens (latitude 37°98’ N, longitude 23°70’ E, altitude 24 m). The first experiment was conducted in the natural soil of a greenhouse from November 2020 till February 2021, while the second experiment was conducted in an open experimental field in summer 2022. More specifically, in the greenhouse experiment, zucchini seedlings were transplanted to the greenhouse soil on the 28th of November 2020 and the experiment was terminated on the 23rd of February 2021, while in the open field experiment planting was performed on the 15th of June 2022 and the experiment was terminated on the 4th of August 2022. In both experiments, integrated crop management (ICM) practices were consistently applied in all treatments.
Two zucchini squash (Cucurbita pepo L.) genotypes were tested. The first genotype of zucchini was the local landrace ‘KOMPOKOLOKYTHO’ from Agrogen S.A., while the second genotype was the commercial hybrid, ‘ARO-800′ from AROSEED (https://aroseed.gr/). In both experiments, half of zucchini seeds were inoculated with PGPR (mix of six Bacillus sp.) before sowing, while the rest were not inoculated in both genotypes. Consequently, the resulting treatments were as follows:
a) ‘KOMPOKOLOKYTHO’, +PGPR;
b) ‘KOMPOKOLOKYTHO’, -PGPR
c) ‘ARO-800′, +PGPR,
d) ‘ARO-800′, -PGPR.
The experiments were designed and analyzed as a two-factorial design (PGPR, genotype) with two levels for each factor (PGPR: inoculation or not; genotype: landrace ‘Kompokolokytho’ or ‘ARO-800′) and the experimental design was randomized complete blocks in both trials. Each of the 2×2=4 experimental treatments was replicated 4 times in one of the 4 groups formed.
Irrigation was performed daily with drip irrigation pipelines in both experiments. Morning irrigation was constant throughout the experiment, while additional afternoon irrigation was applied when needed. Fertilization was applied 3 times a week. Fertilizers were not applied in same amounts each time, as fertilization was continuously adjusted using the Decision Support System NUTRISENSE (https://nutrisense.online/), which is specialized for fertigation management in horticulture.
2.3. Determination of Plant Biomass and Total Yield
In both experiments, for the determination of plants fresh and dry biomass, one zucchini plant per replicate was sampled before commencement of harvest (1st sampling date) and at the end of the experiment (2nd sampling date). When plant samples were collected, shoot and root fresh biomass was recorded. Shoot biomass was the sum of leaves and stems. Then samples were oven-dried at 65 oC for at least 72 h until constant weight was reached. Dried samples were then used to determine plants dry weight and nutrient content. For the estimation of plants total yield, zucchini fruits were harvested when they reached marketable size (length over 12 cm) [24]. In the greenhouse experiment, the harvesting started on the 3rd of January 2021 and terminated on the 23rd of February 2021. In the open field experiment, harvesting started on the 9th of July 2022 and terminated on the 4th of August 2022.
2.4. Shoot Mineral Analysis
Nutrient analyses of both experiments were carried out at the Laboratory of Vegetable Production of the Agricultural University of Athens. Dried leaf tissues were ground in a Wiley mill to pass through a 20-mesh screen and 1 g of the dried tissues was analysed for the macronutrients: N, P, K. The N concentration of leaf tissues was determined after mineralization with sulfuric acid by “Regular Kjeldahl method” [25], whereas P and K concentrations were determined by dry ashing at 550°C for 8 h. Then, extraction for the measurement of nutrients was carried out with a solution of HCl 1Ν placed into the capsule. The solution contained in the capsule was filtered with Whatman No 42 filters into 100 mL volumetric flasks and distilled water was added up to 100 mL. Potassium was measured by placing, diluted or undiluted extraction solution, in the flame photometer (Sherwood 410, Cambridge, UK), while phosphorus was determined as phosphomolybdate blue complex at 880 nm using a spectrophotometer (Anthos Zenyth 200; Biochrom, Cambridge, United Kingdom).
2.5. Statistical Analysis
Two-factor analysis of variance (ANOVA) was applied to evaluate the two selected zucchini genotypes and the pre-inoculation of zucchini seeds with plant growth promoting rhizobacteria, and the interactions between them. Data were analyzed as means ± SE (n = 5). A multiple range (Duncan) test was conducted for all parameters at a p ≤ 0.05 level of significance. Statistical analysis was performed using the STATISTICA software package for Windows 12.0 (Tulsa, OK, USA).
3. Results
3.1. Greenhouse Zucchini Crop
The seed inoculation with the PGPR used as biostimulant significantly increased the vegetative growth of zucchini squash grown the natural soil in a greenhouse. This is indicated by the substantial increase of the fresh and dry shoot weight in both genotypes tested in the current study (Table 1). The fresh and dry shoot mass of the commercial hybrid ‘ARO-800′ was higher than that of the local landrace, irrespective of inoculating the seeds with the PGPR biostimulant or not. The dry matter content in the shoot was not influenced by the treatment of the seed with PGPR but was significantly higher in the commercial hybrid compared with that measured in the local landrace.
The length of the zucchini fruit was not influenced by the inoculation of the seeds with PGPR in the greenhouse crop, while it was similar in both genotypes tested in the current study (Table 2). However, the total fruit yield was significantly enhanced by seed priming with PGPR in both the commercial variety and the local landrace. The commercial hybrid rendered a significantly higher yield than the local landrace, without any interaction between seed priming with PGPR and genotype. The yield increase in the greenhouse zucchini crop was exclusively a result of a higher fruit number per plant, while the mean fruit weight was not influenced by PGPR application in the seeds in both genotypes.
The inoculation of the seeds with PGPR in the greenhouse crop of zucchini squash had no impact on the shoot nitrogen and potassium levels but significantly increased the phosphorus concentration at both sampling dates (Table 3). The shoot P concentration was significantly higher in the landrace on the first sampling date compared to the commercial hybrid, but this difference disappeared on the 2nd sampling date at crop termination. On the other hand, the shoot K concentration was similar at both genotypes on the 1st sampling date, while it was significantly lower in the local landrace at crop termination compared to the commercial hybrid. No interaction between inoculation and genotype was found in the shoot N, P, and K concentrations.
3.2. Open Field Zucchini Squash Crop
The inoculation of the seeds with PGPR stimulated the vegetative growth of zucchini squash cultivated conventionally in an open field, as indicated by the increased fresh and dry shoot weight in both inoculated genotypes compared to non-inoculation (Table 4). Similarly with the greenhouse crop, in the open field crop the commercial hybrid ‘ARO-800′ produced more shoot biomass compared to that rendered by the local landrace, irrespective of seed inoculation with PGPR or not. The dry matter content in the shoot of zucchini squash grown in the open field was not influenced by the treatment of the seed with PGPR but was significantly higher in the commercial hybrid compared with that measured in the local landrace.
The total fruit yield of zucchini squash cultivated in an open field was significantly enhanced by seed priming with PGPR in both the commercial variety and the local landrace (Table 5), in agreement with the respective results in the greenhouse crop. However, unlike in the greenhouse crop, in the open field the commercial hybrid rendered a similar yield with the local landrace. The yield increase imposed by seed inoculation with PGPR in the open field crop was exclusively a result of a higher fruit number per plant in both genotypes, similarly with the greenhouse zucchini crop. The mean fruit weight was not influenced by PGPR application in the seeds.
The inoculation of the seeds with PGPR in the conventional cultivation of zucchini squash in open field significantly increased the phosphorus concentration at both sampling dates, while it had no impact on the shoot nitrogen and potassium levels (Table 6). Similarly with the greenhouse crop, the shoot P concentration in the open field crop was significantly higher in the landrace on the first sampling date compared to the commercial hybrid, but not on the 2nd sampling date. Furthermore, the shoot K concentration was significantly higher in the local landrace than in the commercial hybrid at both sampling dates.
3.3. Root Colonization by PGPR
As shown in Figure 1, the number of Bacillus sp. bacteria counted at crop termination on the root system of zucchini squash was appreciably higher in plants originating from seeds treated with the PGPR biostimulant than in plants originating from untreated seeds. The differences were highly significant in both genotypes tested in the current study.
4. Discussion
In the current study, the inoculation of zucchini squash seeds with the novel mix of Bacillus sp. strains improved substantially both the vegetative growth and the fruit production of the plants. These results clearly show that this mix of Bacillus strains includes plant growth promoting rhizobacteria (PGPR) and thus it could be used as a microbial biostimulant. Soil microorganisms have an active role in natural processes that affect soil fertility and soil quality, such as atmospheric nitrogen fixation, organic matter decomposition and mineral nutrition [26]. These processes are intimately associated not only with the growth and yield of cultivated plants but also with the quality of the obtained products [27]. Several studies have shown that the use of beneficial rhizobacteria (PGPR) as root inoculants in vegetables promotes plant growth, while it can improve soil composition or resistance to pests and diseases. Therefore, many PGPR strains of mixtures of them are used as commercial biostimulants in agriculture and horticulture. According to the European Council Regulation (2019/1009), biostimulants are defined as plant substances, mixtures, and products of microorganisms, which “stimulate plant nutrition processes independently of the nutrient content in the product with the sole aim of improving one or more of the following characteristics of the plant or the plant rhizosphere: i) nutrient use efficiency, ii) tolerance to abiotic stress, iii) quality traits, or iv) availability of confined nutrients in the soil or rhizosphere” [28]. The PGPR can be classified into the following two main categories: extracellular root-forming bacteria (ePGPR) and intracellular root-forming bacteria (iPGPR) [29]. ePGPR live in the rhizosphere or in the spaces between the cells of the root cortex, whereas iPGPR live mainly within specialized nodal structures of the root cells. Also due to their ability to bind to soil nutrients, they improve the levels of nutrients involved in plant cellular processes [30]. Furthermore, some PGPR strains promote the development of disease suppressants or produce phytohormones that increase plant tolerance to biotic stress thereby acting as biostimulants [31].
The tested Bacillus sp. strains mix used as a microbial biostimulant in the current study effectively colonized the zucchini roots, as Bacillus sp. were present at substantially higher populations in the roots of inoculated plants at the end of the experiment, compared to the non-inoculated plants. Patakioutas et al. [32] has reported a significant reduction of the population density for Bacillus amyloliquefaciens when it was used as biostimulant in a soilless tomato crop in a greenhouse. In the current study, the population density was also lower than the initial inoculant and this result indicates that repeated PGPR applications by drenching during the cropping period could potentially increase their effectiveness in terms of crop productivity.
The tested mix of Bacillus sp. strains improved specifically the uptake of P and its transport to the aboveground shoots, as indicated by the higher leaf P levels in plants from seeds inoculated with the tested PGPR mix. Phosphorus is an essential macronutrient for plants but its availability to plants is marginal in most soils [33,34]. Although total P is abundant in the soil [35], its concentration in the soil solution is mostly lower than 0.1 mg L−1 (i.e., 0.1 ppm) because of the poor solubility of the phosphoric salts occurring in the soil, which are mostly complexes of calcium phosphates and magnesium phosphates [36]. As a result, the water-soluble P is quickly depleted in the rhizosphere, and this causes a concentration gradient between the rhizosphere and the bulk of the soil. This gradient contributes to a continuous diffusion of P from the bulk of the soil to the root surface. However, due to the low soluble P concentration in the soil solution, the contribution of diffusion to delivery of P to the root hairs can hardly cover the plant needs, thereby constituting a bottleneck in plant nutrient status [37]. More specifically, as has been reported by Olsen and Watanabe [38], the diffusion coefficient for soil P is relatively low, especially when the P concentrations in the soil solution are less than 0.2 mg kg−1 [38]. Mass flow also contributes to transport of P from the soil bulk to the surface of the root hairs. However, due to the low P concentration in the soil solution the transport of this macronutrient to the root surface through mass flow is negligible. Indeed, zucchini plants generally require from 0,6 to 1,6 L of water per day [39]. Thus, even with a P concentration of 1 mg L−1 in the soil solution, the P delivery via mass flow cannot exceed 1.6 mg day−1, which is far below the actual plant needs.
Given these limitations in P delivery from the bulk of the soil to the root surface through natural processes, plants have evolved other P transport pathways beyond mass flow and diffusion to fully cover their P needs. These pathways are mostly associated with beneficial effects of soil microorganisms. The most widely known alternative pathway of P transport from the soil bulk to the plant roots is through the hyphae of beneficial fungi, such as the arbuscular mycorrhizal fungi which form symbiotic relationships with the plant roots [24]. Furthermore, several studies have concluded that many beneficial microorganisms, including bacteria [40] and fungi [24] living in the soil, are capable of solubilizing otherwise insoluble soil P [41]. Thus, through natural selection, phosphate-solubilizing bacteria and plants have evolved mutual relationships of reciprocal symbiosis [42]. Soil microorganisms can affect P transformation and thus influence the subsequent transportation efficiency of P [43]. The above statement was confirmed in the study of Gravel et al. [44] who reported that inoculation of tomato roots with specific PGPR lead to increased P levels in plants shoots when plants were cultivated on an organic medium, while the same plants had no impact on their leaf P status when they were grown on rockwool. The lack of any positive response of plants grown on rockwool to the PGPR is reasonable, since the P concentrations in the nutrient solutions supplied to rockwool-grown crops is mostly higher than 30 mg L−1 [10], i.e., much higher than those occurring in the soil solution. Considering the increased shoot P levels in the plants obtained from seed inoculated with the mix of the 6 Bacillus sp. strains in the current study, compared to those from non-inoculated seed, it is reasonable to conclude that this mix acted as a biostimulant contributing to a higher P availability for the roots of zucchini squash. Nevertheless, the current study did not reveal whether the tested mix of microorganisms increased the P availability to the zucchini squash crop through an increased solubility of soil P or through other mechanisms. Hence, further research is needed to unravel the mechanisms deployed by the microorganisms tested in the current study to increase the soil P availability in soil-grown zucchini crops.
Adequate P levels in zucchini shoots range between 4 and 6 mg g−1 [45]. Considering this P range, the shoot P levels measured in non-inoculated plants in the current study are considered marginally low, while seed priming with PGPR shifted the shoot P to levels well within the adequacy range. Hence, plants originating from seed treated with PGPR increased plant growth and fruit production, a result which is in agreement with Batista et al. [46] and Souza et al. [47] who concluded that increased P availability and absorption leads to yield increase in zucchini plants.
Regarding N concentration, several studies have found that PGPR can increase the N levels in tomato and zucchini shoots [10,48,49]. However, in the present study, PGPR application did not increase the N levels in plant shoots in both experiments. The lack of any effect of the tested mix of PGPR on plant N status was presumably a result of the adequate N supply to all treatments, which was ensured by using the DSS NUTRISENSE to calculate a balanced fertigation scheme throughout the experiment. Thus, the PGPR tested in the current study could not provide an additional benefit to the plants from inoculated seed. Furthermore, the tested mix of PGPR did not increase the potassium levels in the shoot of plants originating from inoculated seed. In agreement with our results, the potassium status in cultivated vegetable plants is usually not affected by the presence of PGPR in their roots [10,44,50].
Organically cultivated zucchini produced from 1.2 to 1.7 kg of fruit per m-2 [51], while plants in soilless production systems can reached up to 3.5-6.0 kg m−2 [39]. In the current study the application of PGPR in a conventional zucchini crop resulted in substantially higher yield levels than that reported by Montemurro et al. [51] in organically cultivated zucchini, but lower than in the soilless zucchini crop. Nevertheless, the yield performance of fruit vegetables is also depending on the duration of the harvesting season, which can vary widely depending on the cropping season, the cropping system, and the local cultivation practices.
The comparison of the two genotypes tested in the current study revealed that the local landrace ‘Kompokolokytho’ produced lower shoot fresh and dry biomass and lower total yield compared to the commercial hybrid under greenhouse conditions. However, under open field conditions, the local landrace, and the hybrid ‘ARO-800′ resulted in similar total yields. A likely explanation for the different responses of the two genotypes is the season of the year, which was winter in the greenhouse crop but summer in the open field crop. Presumably, the local landrace has either a lower adaptability than the commercial hybrid under cold conditions or a higher adaptability to hot summer conditions. The latter is more likely, as ‘Kompokolokytho’ is a native landrace of Greece, a country with a hot summer season, and this landrace is traditionally grown in the open field.
5. Conclusions
The current study showed that the tested mix of Bacillus sp. strains possesses a biostimulant activity, as its application through seed priming had a positive effect on both the vegetative growth and the yield of zucchini squash grown conventionally under both greenhouse and open-field conditions. The increased biomass and fruit production in the plants originating from inoculated seeds compared to those obtained from non-inoculated plants was associated with significantly higher shoot P levels in both the greenhouse and the open-field crop and in both tested genotypes, i.e., the local landrace and the commercial hybrid. Furthermore, at the end of the cultivation, the colony forming units of Bacillus sp. were appreciably higher in the plants originating from inoculated seeds compared to those from non-inoculated seeds. This finding suggests that the roots of zucchini squash can be effectively colonized by the tested mix of Bacillus sp. microorganisms when this is applied through seed inoculation. Furthermore, the effective colonization of the zucchini squash roots with the tested PGPR points to causal relationships between the tested Bacillus sp. strains and the increased shoot P levels and, in turn, between the higher shoot P levels and the increased vegetative and fruit biomass production. Nevertheless, further research is needed to unravel the mechanisms underlying the increased P availability for the roots of zucchini squash that is achieved after seed priming with the tested PGPR mix.
The commercial zucchini squash hybrid ‘ARO-800′ produced more vegetative and fruit biomass than the local landrace ‘kompokolokytho’ under greenhouse cropping conditions while in the open-field crop both genotypes performed equally, presumably because the local landrace is traditionally grown in, and thus adapted to, open-field cropping systems.
Author Contributions
Conceptualization, D.S. and G.N.; methodology, D.S. and G.N.; software, D.S.; validation, D.S.; formal analysis, D.S. and G.N.; investigation, D.S., D.Y. and G.N.; resources, D.S.; data curation, P.M. and D.Y.; writing—original draft preparation, P.M., D.Y. and P.K.; writing—review and editing, D.S, P.K. and G.N.; visualization, D.S., P.M., D.Y., P.K. and G.N.; supervision, D.S. and G.N.; project administration, D.S. and G.N.; funding acquisition, D.S. All authors have read and agreed to the published version of the manuscript.
Funding
This research was co-financed by the Greek General Secretariat of Research and Innovation and the European Union through the Operational Program “Research–Create–Innovate” in the context of a project granted to the company HUMOFERT S.A., which was titled “A microbial inoculant specialized for priming of vegetable seeds, and emerging seeds. Two new agricultural supplies aiming at improving yield and quality of vegetables within the context of sustainable agriculture”.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Van Dijk, M.; Morley, T.; Rau, M.L.; Saghai, Y. A meta-analysis of projected global food demand and population at risk of hunger for the period 2010–2050. Nat Food, 2021, 2, 494–501. [Google Scholar] [CrossRef]
- Kastner, T.; Chaudhary, A.; Gingrich, S.; Marques, A.; Persson, U.M.; Bidoglio, G.; Provost, G.; Schwarzmüller, F. Global agricultural trade and land system sustainability: Implications for ecosystem carbon storage, biodiversity, and human nutrition. One Earth, 2021, 4, 1425–1443. [Google Scholar] [CrossRef]
- Bailey-Serres, J.; Parker, J.E.; Ainsworth, E.A.; Oldroyd, G.E.; Schroeder, J.I. Genetic strategies for improving crop yields. Nature, 2019, 575, 109–118. [Google Scholar] [CrossRef]
- Koli, P.; Bhardwaj, N.R.; Mahawer, S.K. Agrochemicals: Harmful and beneficial effects of climate changing scenarios. In Climate Change and Agricultural Ecosystems; K. K. Choudhary, A. Kumar, and A. K. Singh, Ed.; Elsevier: Duxford, 2019; pp. 65–94. [Google Scholar]
- Thompson, R.B.; Incrocci, L.; van Ruijven, J.; Massa, D. Reducing contamination of water bodies from European vegetable production systems Agric Water Manage. 2020, 240, 106258. [Google Scholar]
- Tei, F.; De Neve, S.; de Haan, J.; Kristensen, H.L. Nitrogen management of vegetable crops. Agric Water Manage 2020, 240, 106316. [Google Scholar] [CrossRef]
- Li, J.; van Gerrewey, T.; Geelen, D. A Meta-Analysis of Biostimulant Yield Effectiveness in Field Trials. Front Plant Sci, 2022, 13, 836702. [Google Scholar] [CrossRef]
- du Jardin, P. Plant biostimulants: Definition, concept, main categories and regulation. Sci Hort, 2015, 196, 3–14. [Google Scholar] [CrossRef]
- Hunter, M.C.; Smith, R.G.; Schipanski, M.E.; Atwood, L.W.; Mortensen, D.A. Agriculture in 2050: Recalibrating targets for sustainable intensification. Biosci 2017, 67, 386–391. [Google Scholar] [CrossRef]
- Kalozoumis, P.; Savvas, D.; Aliferis, K.; Ntatsi, G.; Marakis, G.; Simou, E.; Tampakaki, A.; Karapanos, I. Impact of plant growth-promoting rhizobacteria inoculation and grafting on tolerance of tomato to combined water and nutrient stress assessed via metabolomics analysis. Front Plant Sci, 2021, 12, 670236. [Google Scholar] [CrossRef]
- Rouphael, Y.; Colla, G. Editorial: Biostimulants in agriculture. Front Plant Sci, 2020, 11, 40. [Google Scholar] [CrossRef]
- Sabatino, L.; Consentino, B.B.; Ntatsi, G.; La Bella, S.; Baldassano, S.; Rouphael, Y. Stand-Alone or Combinatorial Effects of Grafting and Microbial and Non-Microbial Derived Compounds on Vigour, Yield and Nutritive and Functional Quality of Greenhouse Eggplant. Plants, 2022, 11, 1175. [Google Scholar] [CrossRef]
- Consentino, B.B.; Virga, G.; La Placa, G.G.; Sabatino, L.; Rouphael, Y.; Ntatsi, G.; Iapichino, G.; La Bella, S.; Mauro, R.P.; D’Anna, F.; et al. Celery (Apium graveolens L.) Performances as Subjected to Different Sources of Protein Hydrolysates. Plants, 2020, 9, 1633. [Google Scholar] [CrossRef]
- Consentino, B.B.; Aprile, S.; Rouphael, Y.; Ntatsi, G.; De Pasquale, C.; Iapichino, G.; Alibrandi, P.; Sabatino, L. Application of PGPB Combined with Variable N Doses Affects Growth, Yield-Related Traits, N-Fertilizer Efficiency and Nutritional Status of Lettuce Grown under Controlled Condition. Agron, 2022, 12, 236. [Google Scholar] [CrossRef]
- Yakhin, O.I.; Lubyanov, A.A.; Yakhin, I.A.; Brown, P.H. Biostimulants in plant science: A global perspective. Front Plant Sci, 2017, 7, 2049. [Google Scholar] [CrossRef]
- Ruzzi, M.; Aroca, R. Plant growth-promoting rhizobacteria act as biostimulants in horticulture. Sci Hortic, 2015, 196, 124–134. [Google Scholar] [CrossRef]
- Rakshit, A.; Singh, H.B.; Sen, A. Nutrient use efficiency: From basics to advances; Springer: New Delhi, 2015. [Google Scholar]
- Wright. B.; Rowse, H.R.; Whipps, J.M. Application of beneficial microorganisms to seeds during drum priming. Biomed Sci Technol, 2003, 13, 599–614. [Google Scholar]
- Pedrini, S.; Merritt, D.; Stevens, J.; Dixon, K. Seed Coating: Science or Marketing Spin? Trends Plant Sci, 2017, 22, 106–116. [Google Scholar] [CrossRef]
- Liu, J.; Wang, B.; Li, Y.; Huang, L.; Zhang, Q.; Zhu, H.; Wen, Q. RNA sequencing analysis of low temperature and low light intensity-responsive transcriptomes of zucchini (Cucurbita pepo L.). Sci Hort, 2020, 265, 109263. [Google Scholar] [CrossRef]
- Martínez-Valdivieso, D.; Gómez, P.; Font, R.; Río-Celestino, M.D. Mineral composition and potential nutritional contribution of 34 genotypes from different summer squash morphotypes. Eur Food Res Technol, 2015, 240, 71–81. [Google Scholar] [CrossRef]
- Liopa-Tsakalidi, A.; Savvas, D.; Beligiannis, G.N. Modelling the Richards function using Evolutionary Algorithms on the effect of electrical conductivity of nutrient solution on zucchini growth in hydroponic culture. Simu Model Practice Theory 2010, 18, 1266–1273. [Google Scholar] [CrossRef]
- Farid, I.M.; Siam, H.S.; Abbas, M.H.; Mohamed, I.; Mahmoud, S.A.; Tolba, M.; Abbas, H.H.; Yang, X.; Antoniadis, V.; Rinklebe, J.; Shaheen, S.M. Co-composted biochar derived from rice straw and sugarcane bagasse improved soil properties, carbon balance, and zucchini growth in a sandy soil: A trial for enhancing the health of low fertile arid soils. Chemosphere 2022, 292, 133389. [Google Scholar] [CrossRef] [PubMed]
- Colla, G.; Rouphael, Y.; Cardarelli, M.; Tullio, M.; Rivera, C.M.; Rea, E. Alleviation of salt stress by arbuscular mycorrhizal in zucchini plants grown at low and high phosphorus concentration. Biol Fertility Soils 2008, 44, 501–509. [Google Scholar] [CrossRef]
- Bremner, J.M. Total nitrogen. Meth Soil Anal: Part 2 chemical and microbiological properties, 1965, 9, 1149–1178. [Google Scholar]
- Rouphael, Y.; Colla, G. Toward a sustainable agriculture through plant biostimulants: From experimental data to practical applications. Agron, 2020, 10, 1461. [Google Scholar] [CrossRef]
- Bucki, P.; Regdos, K.; Siwek, P.; Domagała-Świątkiewicz, I.; Kaszycki, P. Impact of soil management practices on yield quality, weed infestation and soil microbiota abundance in organic zucchini production. Sci Hort, 2021, 281, 109989. [Google Scholar] [CrossRef]
- EU. (2019). Regulation of the european parliament and of the council laying down rules on the making available on the market of EU fertilising products and amending Regulations (EC) No 1069/2009 and (EC) No 1107/2009 and repealing Regulation (EC) No 2003/2003. https://eur-lex.europa.eu/legalcontent/EN/TXT/?uri=OJ:L:2019:170:TOC.
- Viveros, O.M.; Jorquera, M.A.; Crowley, D.E.; Gajardo, G.; Mora, M.L. Mechanisms and practical considerations involved in plant growth promotion by rhizobacteria. J Soil Sci Plant Nutr, 2010, 10, 293–319. [Google Scholar]
- Khatoon, Z.; Huang, S.; Rafique, M.; Fakhar, A.; Kamran, M.A.; Santoyo, G. Unlocking the potential of plant growth-promoting rhizobacteria on soil health and the sustainability of agricultural systems. J Environ Manage, 2020, 273, 111118. [Google Scholar] [CrossRef] [PubMed]
- Mena-Violante, H.G.; Olalde-Portugal, V. Alteration of tomato fruit quality by root inoculation with plant growth-promoting rhizobacteria (PGPR): Bacillus subtilis BEB-13bs. Sci Hort, 2007, 113, 103–106. [Google Scholar] [CrossRef]
- Patakioutas, G.; Dimou, D.; Yfanti, P.; Karras, G.; Ntatsi, G.; Savvas, D. Root inoculation with beneficial micro-organisms as a means to control Fusarium oxysporum f. sp. lycopersici in two Greek landraces of tomato grown on perlite. Acta Hort, 2015, 1107, 277–286. [Google Scholar] [CrossRef]
- Azziz, G.; Bajsa, N.; Haghjou, T.; Taulé, C.; Valverde, Á.; Igual, J.M.; Arias, A. Abundance, diversity and prospecting of culturable phosphate solubilizing bacteria on soils under crop–pasture rotations in a no-tillage regime in Uruguay. Appl Soil Ecol, 2012, 61, 320–326. [Google Scholar] [CrossRef]
- Tak, H.I.; Ahmad, F.; Babalola, O. , Inam, A. Growth, photosynthesis and yield of chickpea as influenced by urban wastewater and different levels of phosphorus. Int J Plant Res, 2012, 2, 6–13. [Google Scholar] [CrossRef]
- Richardson, A.E.; Barea, J.M.; McNeill, A.M.; Prigent-Combaret, C. Acquisition of phosphorus and nitrogen in the rhizosphere and plant growth promotion by microorganisms. Plant Soil 2009, 321, 305–339. [Google Scholar] [CrossRef]
- Shah, A.; Nazari, M.; Antar, M.; Msimbira, L.A.; Naamala, J.; Lyu, D.; Rabileh, M.; Zajonc, J.; Smith, D.L. PGPR in agriculture: A sustainable approach to increasing climate change resilience. Front Sust Food Sys, 2021, 5, 667546. [Google Scholar] [CrossRef]
- Larsen, S. Soil phosphorus. Adv Agron, 1967, 19, 151–210. [Google Scholar]
- Olsen, S.R.; Watanabe, F.S. Diffusion of phosphorus as related to soil texture and plant uptake. Soil Sci Soc Amer J, 1963, 27, 648–653. [Google Scholar] [CrossRef]
- Rouphael, Y.; Colla, G. Growth, yield, fruit quality and nutrient uptake of hydroponically cultivated zucchini squash as affected by irrigation systems and growing seasons. Sci Hort, 2005, 105, 177–195. [Google Scholar] [CrossRef]
- Maheshwari, D.K.; Dheeman, S. Field crops: Sustainable management by PGPR. Springer 2019, 458. [Google Scholar]
- Bechtaoui, N.; Raklami, A.; Benidire, L.; Tahiri, A.I.; Göttfert, M.; Oufdou, K. Effects of PGPR co-inoculation on growth, phosphorus nutrition and phosphatase/phytase activities of faba bean under different phosphorus availability conditions. Pol J Environ Stud, 2020, 29, 1557–1565. [Google Scholar] [CrossRef] [PubMed]
- Berde, C.V.; Salvi, S.P.; Kajarekar, K.V.; Joshi, S.A.; Berde, V.B. Insight into the animal models for microbiome studies. Springer, 2021, Chapter 13.
- Richardson, A.E. Prospects for using soil microorganisms to improve the acquisition of phosphorus by plants. Funct Plant Biol, 2001, 28, 897–906. [Google Scholar] [CrossRef]
- Gravel, V.; Antoun, H.; Tweddell, R.J. Growth stimulation and fruit yield improvement of greenhouse tomato plants by inoculation with Pseudomonas putida or Trichoderma atroviride: Possible role of indole acetic acid (IAA). Soil Biol Biochem, 2007, 39, 1968–1977. [Google Scholar] [CrossRef]
- B. van RAIJ. (1996). Recomendações de adubação e calagem para o Estado de São Paulo (Vol. 100, p. 285p). Campinas: IAC.
- Batista, C.M.; da Mota, W.F.; Pegoraro, R. 9.; Gonçalves, R.E.M.; Aspiazú, I. Production of italian zucchini in response to N and P fertilization. Rev Brasileira Ciências Agr 2020, 15, 1–6. [Google Scholar]
- Souza, F.I.D.; Grangeiro, L.C.; de Souza, V.D.F.; Gonçalvez, F.D.C.; de Oliveira, F.H.; de Jesus, P.M. X Agronomic performance of Italian zucchini as a function of phosphate fertilization. Revista Brasileira de Engenharia Agrícola e Ambiental, 2018, 22, 206–211. [Google Scholar] [CrossRef]
- Tchiaze, A.I.; Taffouo, V.D.; Fankem, H.; Kenne, M.; Baziramakenga, R.; Ekodeck, G.E.; Antoun, H. Influence of Nitrogen Sources and Plant Growth-Promoting Rhizobacteria Inoculation on Growth, Crude Fiber and Nutrient Uptake in Squash (Cucurbita moschata Duchesne ex Poir.) Plants. Notulae Bot Hort Agroboti Cluj-Napoca, 2016, 44, 53–59. [Google Scholar] [CrossRef]
- Cordero, I.; Balaguer, L.; Rincón, A.; Pueyo, J.J. Inoculation of tomato plants with selected PGPR represents a feasible alternative to chemical fertilization under salt stress. J Plant Nutr Soil Sci, 2018, 181, 694–703. [Google Scholar] [CrossRef]
- Novello, G.; Cesaro, P.; Bona, E.; Massa, N.; Gosetti, F.; Scarafoni, A.; Todeschini, V.; Berta, G.; Lingua, G.; Gamalero, E. The effects of plant growth-promoting bacteria with biostimulant features on the growth of a local onion cultivar and a commercial zucchini variety. Agron 2021, 11, 888. [Google Scholar] [CrossRef]
- Montemurro, F.; Fiore, A.; Campanelli, G.; Tittarelli, F.; Ledda, L.; Canali, S. Organic fertilization, green manure, and vetch mulch to improve organic zucchini yield and quality. HortSci, 2013, 48, 1027–1033. [Google Scholar] [CrossRef]
Figure 1.
Number of Bacillus sp. bacteria counted at crop termination on the root system of two zucchini squash genotypes (landrace ‘Kompokolokytho’ or ‘ARO-800′ F1) originating from seeds either treated or non-treated with the PGPR biostimulant.
Figure 1.
Number of Bacillus sp. bacteria counted at crop termination on the root system of two zucchini squash genotypes (landrace ‘Kompokolokytho’ or ‘ARO-800′ F1) originating from seeds either treated or non-treated with the PGPR biostimulant.
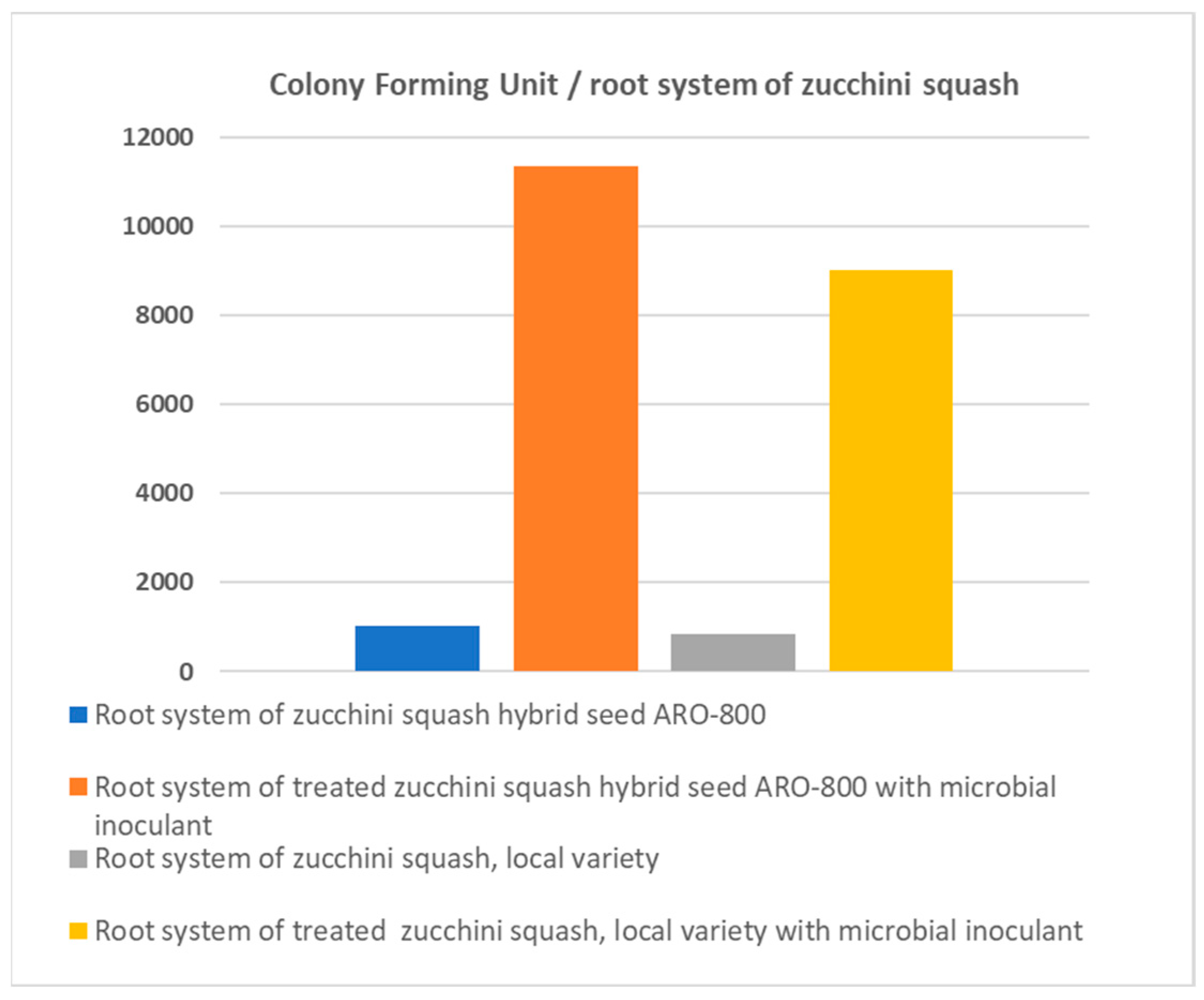

Table 1.
Impact of seed inoculation with a PGPR strain used as biostimulant on shoot fresh weight, shoot dry weight and shoot dry matter content of two zucchini squash genotypes (landrace ‘Kompokolokytho’ or ‘ARO-800′ F1) grown in the soil in a greenhouse.
Table 1.
Impact of seed inoculation with a PGPR strain used as biostimulant on shoot fresh weight, shoot dry weight and shoot dry matter content of two zucchini squash genotypes (landrace ‘Kompokolokytho’ or ‘ARO-800′ F1) grown in the soil in a greenhouse.
| Genotype | PGPR | Shoot Fresh Weight (kg/plant) | Shoot Dry Weight (g/Plant) | Shoot Dry Matter Content (%) |
|---|---|---|---|---|
| Landrace | -PGPR | 1.373 | 106.4 | 7.8 |
| +PGPR | 1.699 | 130.6 | 7.8 | |
| ‘ARO-800′ | -PGPR | 1.616 | 137.8 | 8.6 |
| +PGPR | 1.983 | 160.9 | 8.2 | |
| Main Effects | ||||
| PGPR | ||||
| -PGPR | 1.494 b | 122.1 b | 8.2 | |
| +PGPR | 1.841 a | 145.8 a | 8.0 | |
| Genotype | ||||
| Landrace | 1.536 b | 118.5 b | 7.8 b | |
| ‘ARO-800′ | 1.799 a | 149.4 a | 8.4 a | |
| Significance | ||||
| PGPR | ** | ** | n.s. | |
| Genotype | * | *** | * | |
| PGPR × genotype | n.s. | n.s. | n.s. | |
Means (n = 5) followed by different letters are significant according to ANOVA. The symbols *, **, and *** indicate that the differences were significant at P<0.05, P<0.01 και P<0.001, respectively, while n.s. denotes non-significant differences.
Table 2.
Impact of seed inoculation with a PGPR strain used as biostimulant on mean fruit length, mean fruit weight, and total yield in two zucchini squash genotypes (landrace ‘Kompokolokytho’ or ‘ARO-800′ F1) grown in the soil in a greenhouse.
Table 2.
Impact of seed inoculation with a PGPR strain used as biostimulant on mean fruit length, mean fruit weight, and total yield in two zucchini squash genotypes (landrace ‘Kompokolokytho’ or ‘ARO-800′ F1) grown in the soil in a greenhouse.
| Genotype | PGPR | Mean Fruit Length (cm) | Mean Fruit Weight (g) | Total Yield (kg m−2) |
|---|---|---|---|---|
| Landrace | -PGPR | 16.9 | 146.4 | 1.392 |
| +PGPR | 16.7 | 149.6 | 1.765 | |
| ‘ARO-800′ | -PGPR | 16.6 | 152.4 | 1.991 |
| +PGPR | 16.8 | 156.1 | 2.355 | |
| Main Effects | ||||
| PGPR | ||||
| -PGPR | 16.7 | 149.4 | 1.692 b | |
| +PGPR | 16.8 | 152.8 | 2.060 a | |
| Genotype | ||||
| Landrace | 16.8 | 147.6 b | 1.578 b | |
| ‘ARO-800′ | 16.7 | 154.3 a | 2.173 a | |
| Significance | ||||
| PGPR | n.s. | n.s. | ** | |
| Genotype | n.s. | * | *** | |
| PGPR × genotype | n.s. | n.s. | n.s. | |
Means (n = 5) followed by different letters are significant according to ANOVA. The symbols *, **, and *** indicate that the differences were significant at P<0.05, P<0.01 και P<0.001, respectively, while n.s. denotes non-significant differences.
Table 3.
Impact of seed inoculation with a PGPR strain used as biostimulant on N, P and K concentrations in shoot samples collected at two sampling dates (1st SD and 2nd SD, respectively) from two zucchini squash genotypes (landrace ’Kompokolokytho’ or ‘ARO-800′ F1) in the soil in a greenhouse.
Table 3.
Impact of seed inoculation with a PGPR strain used as biostimulant on N, P and K concentrations in shoot samples collected at two sampling dates (1st SD and 2nd SD, respectively) from two zucchini squash genotypes (landrace ’Kompokolokytho’ or ‘ARO-800′ F1) in the soil in a greenhouse.
| Genotype | PGPR | Shoot N (mg g−1 d.wt.) |
Shoot P (mg g−1 d.wt.) |
Shoot K (mg g−1 d.wt.) |
|||||
|---|---|---|---|---|---|---|---|---|---|
| 1st SD | 2nd SD | 1st SD | 2nd SD | 1st SD | 2nd SD | ||||
| Landrace | -PGPR | 3.38 | 3.40 | 3.29 | 2.88 | 35.2 | 25.0 | ||
| +PGPR | 3.58 | 3.36 | 4.14 | 3.30 | 35.4 | 28.1 | |||
| ‘ARO-800′ | -PGPR | 3.52 | 3.49 | 2.73 | 3.08 | 35.4 | 29.3 | ||
| +PGPR | 3.68 | 3.51 | 2.82 | 3.33 | 37.2 | 32.9 | |||
| Main Effects | |||||||||
| PGPR | |||||||||
| -PGPR | 3.45 | 3.45 | 3.01 b | 2.98 b | 35.3 | 27.1 | |||
| +PGPR | 3.63 | 3.44 | 3.48 a | 3.32 a | 36.3 | 30.5 | |||
| Genotype | |||||||||
| Landrace | 3.48 | 3.38 | 3.71 a | 3.09 | 35.3 | 26.5 b | |||
| ‘ARO-800′ | 3.60 | 3.50 | 2.77 b | 3.20 | 36.3 | 31.1 a | |||
| Significance | |||||||||
| PGPR | n.s. | n.s. | * | * | n.s. | n.s. | |||
| Genotype | n.s. | n.s. | *** | n.s. | n.s. | * | |||
| PGPR × genotype | n.s. | n.s. | n.s. | n.s. | n.s. | n.s. | |||
Means (n = 5) followed by different letters are significant according to ANOVA. The symbols *, and *** indicate that the differences were significant at P<0.05 and P<0.001, respectively, while n.s. denotes non-significant differences.
Table 4.
Impact of seed inoculation with a PGPR strain used as biostimulant on shoot fresh weight, shoot dry weight and shoot dry matter content of two zucchini squash genotypes (landrace ’Kompokolokytho’ or ‘ARO-800′ F1) grown in an open field according to conventional farming practices.
Table 4.
Impact of seed inoculation with a PGPR strain used as biostimulant on shoot fresh weight, shoot dry weight and shoot dry matter content of two zucchini squash genotypes (landrace ’Kompokolokytho’ or ‘ARO-800′ F1) grown in an open field according to conventional farming practices.
| Genotype | PGPR | Shoot Fresh Weight (kg/Plant) | Shoot Dry Weight (g/Plant) | Shoot Dry Matter Content (%) |
|---|---|---|---|---|
| Landrace | -PGPR |
1.373 | 106.4 | 7.8 |
| +PGPR | 1.699 | 130.6 | 7.8 | |
| ‘ARO-800′ | -PGPR |
1.616 | 137.8 | 8.6 |
| +PGPR | 1.983 | 160.9 | 8.2 | |
| Main Effects | ||||
| PGPR | ||||
| -PGPR σπόρου |
1.494 b | 122.1 b | 8.2 | |
| +PGPR | 1.841 a | 145.8 a | 8.0 | |
| Genotype | ||||
| Landrace | 1.536 b | 118.5 b | 7.8 b | |
| ‘ARO-800′ | 1.799 a | 149.4 a | 8.4 a | |
| Significance | ||||
| PGPR σπόρου σπόρου |
** | ** | n.s. | |
| Genotype | * | *** | * | |
| PGPR × genotype σπόρου σπόρου |
n.s. | n.s. | n.s. | |
Means (n = 5) followed by different letters are significant according to ANOVA. The symbols *, **, and *** indicate that the differences were significant at P<0.05, P<0.01 and P<0.001, respectively, while n.s. denotes non-significant differences.
Table 5.
Impact of seed inoculation with a PGPR strain used as biostimulant on mean fruit length, mean fruit weight, and total yield in two zucchini squash genotypes (landrace ’Kompokolokytho’ or ‘ARO-800′ F1) grown in an open field according to conventional farming practices.
Table 5.
Impact of seed inoculation with a PGPR strain used as biostimulant on mean fruit length, mean fruit weight, and total yield in two zucchini squash genotypes (landrace ’Kompokolokytho’ or ‘ARO-800′ F1) grown in an open field according to conventional farming practices.
| Genotype | PGPR | Total Yield (kg m−2) | Fruit Number per m2 | Mean Fruit Weight (g) |
|---|---|---|---|---|
| Landrace | -PGPR |
1.592 | 14.00 | 113.5 |
| +PGPR | 1.883 | 16.36 | 115.2 | |
| ‘ARO-800′ | -PGPR |
1.601 | 13.67 | 117.2 |
| +PGPR | 2.014 | 17.00 | 118.6 | |
| Main Effects | ||||
| PGPR | ||||
| -PGPR σπόρου |
1.587 b | 13.83 b | 115.3 | |
| +PGPR | 1.942 a | 16.68 a | 116.9 | |
| Genotype | ||||
| Landrace | 1.743 | 15.18 | 114.3 | |
| ‘ARO-800′ | 1.807 | 15.33 | 117.4 | |
| Significance | ||||
| PGPR σπόρου σπόρου |
** | * | n.s. | |
| Genotype | n.s. | n.s. | n.s. | |
| PGPR × genotype σπόρου σπόρου |
n.s. | n.s. | n.s. | |
Means (n = 5) followed by different letters are significant according to ANOVA. The symbols * and ** indicate that the differences were significant at P<0.05 and P<0.01 respectively, while n.s. denotes non-significant differences.
Table 6.
Impact of seed inoculation with a PGPR strain used as biostimulant on N, P and K concentrations in shoot samples collected at two sampling dates (1st SD and 2nd SD, respectively) from two zucchini squash genotypes (landrace ’Kompokolokytho’ or ‘ARO-800′ F1) grown in an open field according to conventional farming practices.
Table 6.
Impact of seed inoculation with a PGPR strain used as biostimulant on N, P and K concentrations in shoot samples collected at two sampling dates (1st SD and 2nd SD, respectively) from two zucchini squash genotypes (landrace ’Kompokolokytho’ or ‘ARO-800′ F1) grown in an open field according to conventional farming practices.
| Genotype | PGPR | Shoot N (mg g−1 d.wt.) |
Shoot P (mg g−1 d.wt.) |
Shoot K (mg g−1 d.wt.) |
|||||
|---|---|---|---|---|---|---|---|---|---|
| 1st SD | 2nd SD | 1st SD | 2nd SD | 1st SD | 2nd SD | ||||
| Landrace | -PGPR | 4.78 | 3.40 | 4.90 b | 3.98 | 42.0 | 34.7 | ||
| +PGPR | 5.48 | 3.66 | 6.45 a | 4.14 | 42.5 | 34.3 | |||
| ‘ARO-800′ | -PGPR | 3.99 | 2.97 | 4.32 bc | 3.35 | 33.5 | 25.0 | ||
| +PGPR | 4.08 | 3.58 | 3.69 cd | 4.49 | 36.0 | 29.3 | |||
| Main Effects | |||||||||
| PGPR | |||||||||
| -PGPR | 4.38 | 3.19 | 4.61 | 3.67 b | 37.8 | 29.1 | |||
| +PGPR | 4.78 | 3.62 | 5.07 | 4.32 a | 39.3 | 31.8 | |||
| Genotype | |||||||||
| Landrace | 5.12 | 3.53 | 5.67 | 4.06 | 42.3 a | 34.4 a | |||
| ‘ARO-800′ | 4.03 | 3.28 | 4.00 | 3.92 | 34.8 b | 27.1 b | |||
| Significance | |||||||||
| PGPR | n.s. | n.s. | ns | * | n.s. | n.s. | |||
| Genotype | n.s. | n.s. | * | n.s. | * | * | |||
| PGPR × genotype | n.s. | n.s. | * | n.s. | n.s. | n.s. | |||
Means (n = 5) followed by different letters are significant according to ANOVA. The symbols * indicate that the differences were significant at P<0.05, while n.s. denotes non-significant differences.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



