
Submitted:
28 December 2023
Posted:
29 December 2023
You are already at the latest version
Abstract
Viruses are silent enemies that intrude and take control of the plant cell's machinery for their own multiplication. Infection by viruses and the resulting damage is still a major challenge in the agriculture sector. Plants have the ability to fight back, but the ability of viruses to counteract and mutate at a fast rate helps them to hide from the host response. Therefore, classical approaches for introgressing resistance genes by breeding have obtained limited success in counteracting the virus menace. Genetic modification (GM) based strategies have been successful in engineering artificial resistance in plants. Several different approaches based on pathogen derived resistance, antisense constructs, hairpin RNA, double stranded RNA etc. have been used to enhance plant resistance to the virus. Recently, genome editing (GE) strategies mainly involving the CRISPR/Cas mediated modifications are being used for virus control. In this review, we discuss the developments and advancements in the GM and GE based methods in tackling virus infection in plants.
Keywords:
Pathogen Derived Resistance
; RNA silencing
; artificial miRNA
; tasiRNA
; Epigenetic editing
; prime editing
; Multiplexing CRISPR/Cas editing
1. Introduction
Viruses constitute nearly half of the microbial pathogens that are accountable for emerging and re-emerging the epidemics worldwide [1]. They also pose serious challenges to agriculture, by causing major plant diseases like bunchy top of banana, tobacco mosaic, wheat streak mosaic, potato tuber necrotic ring spot, cucumber green mottle mosaic, African cassava mosaic etc. So far, 131 families containing 803 genera and 4853 species of plant virus are known (ICTV, 2018). The viruses are responsible for a worldwide crop loss of $30 billion annually and it is predicted to reach up to $60-80 billion [2,3,4]. The plant viral diseases threaten global food security as they can cause severe food limitations for the world population, which is projected to elevate from 8 billion in 2022 to 10 billion by 2050 [5,6].
For resolution of any problem, it is important to implement appropriate prevention and control measures. However, the viruses cannot be controlled by chemicals like pesticides, although their carriers or vectors can be restrained to some extent [7] This simply leaves the possibility of finding or generating plant species with inherent resistance to the virus infections. The silver lining came from the McKinney’s observation in 1929, that mild infection caused by Tobacco Mosaic Virus (TMV) green mosaic strain protected the tobacco plants from severe infection of TMV yellow mosaic strain, pointed towards the existence of mechanism(s) for cross protection [8]. This phenomenon involves the formation of a memory response in the host plant to combat the challenge posed by related virus [9] These early observations prompted search for host-based resistance genes and detailed understanding of plant-virus interactions [10,11].
The selection of dominant and recessive genes associated with virus resistance played an important role in breeding of naturally resistant plant varieties as a cheaper and better alternative for tackling viral diseases [4]. Classical approaches such as introgression of resistance genes by marker assisted selection (MAS), QTL mapping and gene pyramiding were extensively used to tackle the menance of viruses. However, there were several bottlenecks such as non-availability of markers near the resistance genes, transfer of undesired traits with resistance genes, and screening of plants for several generations, which made the process tedious and time consuming ands limited the success of these techniques [12].
The era of genetic engineering accelerated the process by allowing incorporation of genes in the plant genome as per the requirement. In the early 1980s, the concept of Pathogen Derived Resistance (PDR) laid the foundation of artificial resistance. PDR employed the use of virus proteins like coat protein (CP) to disrupt the virus replication by inactivating the viral components [10]. Subsequent studies unravelled the role of RNA silencing or RNA interference (RNAi) in symptom recovery and cross protection [13,14]. It also strengthened the possibilities for employing RNA silencing for generating virus resistance in plants. However, the silencing pathway has some limitations due to presence of silencing suppression activity in the viral encoded proteins. In recent times, CRISPR/Cas based tools are being used to edit the plant genome for generating virus resistant plants. In this review, we have discussed the improvements and advancements of gene modifications (GM) and genome editing (GE) approaches in managing plant virus infections.
2. GM approaches
These approaches involve the over-expression or knock-down of genes using a variety of genetic engineering tools. The methodology usually involves cloning the gene of interest (trans-gene) into the desired vector in either sense or antisense orientation followed by its transformation in plants [15]. Many old reports are available on engineering virus resistance in plants via this transgenic approach [16,17].
2.1. Pathogen Derived Resistance
In this approach the pathogen’s own genetic material is used to generate resistance in the host plant. The concept of PDR was first presented by Sanford and Johnston (1985) and a year later it was proven to be successful in case of virus infections. The transgenic plants expressing coat protein (CP) gene of TMV were shown to exhibit resistance against TMV infection [18]. This ground-breaking discovery initiated the development of various virus-resistant transgenic plants [5]. PDR can be classified into different types depending on the strategy used to generate resistance towards the virus in the host plant.
2.1.1. Protein-Mediated Resistance
The process entails the expression of viral encoded proteins like CP, MP (movement protein) or Rep (replicase) in the plants to counteract the virus [5]. In 1986, Beachy’s group first demonstrated that the expression of the CP gene of TMV in transgenic tobacco plants could provide considerable protection against virus infection [19]. It was observed that the expression of these viral proteins interfered with the disassembly of virus by formation of pseudo-capsids [5] Several transgenics were made using CP of Cucumber mosaic virus (CMV), Papaya ringspot virus (PRSV), Potato Virus X (PVX), Plum pox virus (PPV), Zucchini yellow mosaic virus (ZYMV), Watermelon mosaic virus (WMV), Tomato yellow leaf curl virus (TYLCV), Tomato mosaic virus (ToMV) and so on [20]
PDR also provided cross-protection against infection by related viruses. This technique was more effective in case of closely related viruses than those which are distantly related [21]. When TMV CP-transgenic plants, were infected by different viruses of the Tobamovirus family their inhibition was differentially compromised. Some viruses like Tomato mosaic virus (ToMV), Pepper mild mosaic virus (PMMV) and Tomato mild green mosaic virus (TMGMV) were inhibited by ~98% while no resistance was observed against others like Cucumber mosaic virus (CMV), African mosaic virus (AMV), Potato virus X (PVX) and Potato virus Y (PVY), though there was decrease in symptoms and spread [22]. In field trials, CP-TMV expressing tomato and CP-PVX/PVY expressing potato transgenics performed better than the wild-type [23].
In case of PVX infection, it was shown that the interaction of the 5' end of viral RNA with CP expressed in the host plant suppressed the expression of RNA dependent RNA polymerase (RdRP) encoded by the virus [24,25] The region and quantity of protein expression were important determiners for the generation of resistance. For instance, if CP was expressed in mesophyll but the viral RNA was introduced in epidermal cells then resistance did not develop [26]. Expression of mutant CP in the transgenics also resulted in resistance against wild-type CP, as the mutant CP could interact with the wild-type CP subunits or the host factors, which were involved in virion disassembly [27,28]. The extent of aggregation of CP correlated with level of Coat protein mediated resistance (CPMR) [29]. This observation suggested that homology between the amino acid sequences of the challenging viruses provided cross-protection in the transgenic [5].
Likewise, the MPs, which have role in cell to cell spread of the virus, were also utilized for triggering PDR. This technique worked better for large genera of viruses irrespective of the viral MP expressed in plants. The expression of defective or mutant MPs could also block virus transmission, since the mutant MPs competed with wild-type MPs of the virus for the plasmodesmatal sites [30]. Mutant TMV-MP provided resistance against Potexviruses, Tobamoviruses and Cucumoviruses [31]. Several groups tried to multiplex this approach by generating transgenics with more than one type of MP, by using overlapping regions of the ORFs which are also known as Triple gene block (TGB). However this approach did not provide wider resistance, possibly due to insufficient interaction of TGB with the plasmodesmata [32].
In subsequent studies, it was shown that use of modified non-structural proteins like Rep/RdRP were able to provide better results even at low levels of expression [33]. Mutations in the ori or NTP-binding sites of Rep gene (AC1) provided resistance against Geminiviruses [34,35]. The Rep/RdRP proteins could directly interfere with virus replication or bind with host factors involved in the viral replication process [36]. Detailed molecular mechanisms behind the responses were established for few viruses like TMV. Although later reports demonstrated that these proteins mediated resistance mainly through the RNA silencing machinery instead of direct protein interactions [37,38].
Despite several examples of the successful implementation of this technology, it faced some challenges due to evolutionary pressure provided by the expression of transgene. The genomic content of the virus interacting with the transgenic got altered due to recombination between transgenic and viral CP transcripts [39]. This trans-capsidation of the CP in transgenics imparted a new potential to the infecting virus isolate. Mixed infection by two unrelated viruses was another issue as it led to antagonism and synergism of defence responses. For instance, different outcomes were produced during mixed infection with Papaya ringspot virus (PRSV) and Papaya mosaic virus (PapMV). In the case when plants were first infected with PRSV followed by PapMV or simultaneously inoculated with both a synergistic response was observed. However, when first inoculation of PapMV was followed by PRSV an antagonistic response was observed, which caused reduction in PRSV titres due to activation of defence responses [40].
2.1.2. Defective interfering virus mediated resistance
Plant viruses are often associated with subgenomic/subviral RNAs known variably as satellite (sat) or defective interfering (DI) virus [41]. The DI viruses lack a critical portion of the virus genome, which makes them defective for replication. They require the presence of the complete functional virus, known as the helper virus, for their replication and encapsidation [42]. DI viruses are basically the defective versions of helper viruses formed by mainly deletions, duplication and inversion processes [43]. There is often a synergistic interaction between the DI virus and its helper as they can produce mild symptoms on one host plant but elicit severe symptoms on another host or in combination with a different strain of helper virus [44,45]. The observations that DI virus can attenuate symptoms of unrelated helper virus in co-infected cells by inhibiting the production and long-distance movement of helper virus [46,47]. It was used in transgenic approaches to decrease virus symptoms and/or titers [42]. These sat RNAs compete with the viral genome for binding to replicase for replication which indirectly suppresses the virus replication [47]. In 2015, it was shown that CMV sat-Y derived siRNAs directed against the VSR, 2b of CMV and P10 of Tombusvirus, caused displacement of the VSR-bound siRNAs resulting in down regulation of the viral target genes [48]. The major limitation of this strategy was a probable risk that the DI virus could mutate to a virulent form in conjunction with the infecting virus in the transgenic plants.
2.2. RNA silencing as a major antiviral weapon
The discovery of non-coding small RNAs and their role in gene silencing provided a paradigm shift in the understanding of host defense to virus and generation of virus resistance. RNA silencing is an evolutionarily conserved, fundamental cellular mechanism for regulating gene expression [49]. The process involves small RNA mediated degradation or inactivation of specific mRNA in sequence dependent manner [50]. In plant immunity, it contributes as first line defences against virus infection as is often referred to as virus induced gene silencing (VIGS). Earlier it was understood that the genome of RNA viruses and viroids were subject to RNA silencing [51,52] but later it was shown to be effective against the DNA viruses such as Geminiviruses (having ssDNA) and Caulimoviruses, Tungroviruses (having dsDNA with RNA intermediate) [53,54].
The RNA silencing machinery is triggered by the production of double stranded (ds) RNA either as replicative intermediate or self-folded complementary region of single strand (ss) viral transcript (Figure 1). The dsRNA is diced by the microprocessor protein complex containing Dicer Like Proteins (DCLs) to generate 20-24 nt small RNAs [55]. The small RNAs derived from the virus are normally categorised as viral small interfering RNA (vsiRNA). They are loaded onto RNA induced silencing complex (RISC) to direct Transcriptional Gene Silencing (TGS) or Post- Transcriptional Gene Silencing (PTGS) [55,56]. In TGS, the target gene gets methylated to block transcription [57,58], whereas in PTGS, the target mRNA is cleaved to block translation [59]. The visiRNAs also act as mobile signals as they can spread from cell to cell [60] [61]. These primary siRNAs also get amplified with the help of SGS3 and RDR6 to produce secondary siRNAs to provide protection throughout the plant [55,62].
2.2.1. Role of TGS in virus infection
RNA silencing has a great impact on regulating viral genes as well as host genome via epigenetic modifications. The small RNAs initiate RNA directed DNA methylation (RdDM) of complementary DNA sequences [63,64], which reduces or blocks the transcription of mRNA, so the gene undergoes silencing [65,66]. Some proteins like AGO4, DCL3, RNA polymerase IVa, RDR2 which are involved in TGS would also lead to initiates systemic silencing. Many small RNAs as well as dsRNA were reported to have a role in targeted methylation by triggering RdDM.
Studies have shown that a typical small RNA mediated methylation pathway involves transcription from the target locus by RNA polymerase IVa [67]. The ssRNA is converted into dsRNA by action of RdRP/RDR2, which is then diced by DCL3. The siRNAs thus generated, associate with AGO4 containing RISC and polymerase IVb complexes to search for the target DNA sequences. At the target site RNA polymerase V is recruited to form AGO4:siRNA:PolV complex which attracts de novo methyltransferases [68]. Normally cytosine methyltransferases like DOMAIN REARRANGED METHYLTRANSFERASE 1/2 (DRM1/2), CHROMOMETHYLASE 2/3 (CMT2/3) (for CHG/CHH methylation), METHYLTRANSFERASE 1 (MET1) (for CG methylation) and KRYPTONITE 2 (KYP2) get recruited at the target site alongwith DEFECTIVE IN RNA-DIRECTED DNA METHYLATION 1 (DRD1), DEFECTIVE IN MERISTEM SILENCING 3 (DMS3) and DEFICIENT IN DNA METHYLATION 1 (DDM1) [69]. Cytosine methylation engages the histone deacetylases and other methyltransferases to initiate histone modifications [70]. It has been shown that methylation in promoter region, rather than coding region, is more effective for silencing [71]. Moreover, CG and CHG methylation can be maintained after cell division and without RNA triggers but CHH methylation cannot. [63,72,73].
DNA viruses can be easily targeted through TGS, because of their ssDNA and dsDNA replication process (Figure 1). The replication intermediates of Pararetroviruses and Geminiviruses form mini chromosomes, which are common targets of TGS [74,75,76]. VIGS was shown to initiate RdDM and methylation deficient mutants of Arabidopsis were hypersusceptible to infection [69]. The accumulation of visiRNAs to threshold levels at the target DNA recruits DRM2 for initiating de novo methylation [77]. Methionine cycle (MTC) plays a crucial role in small RNA mediated methylation during virus infection by maintaining continuous supply of S-adenosyl methionine (SAM) and removal of methyl transferase inhibitor, S-adenosyl-homocysteine (SAH). S-adenosyl-L-homocysteine hydrolase (SAHH) converts SAH to L-homocysteine (HCY) and then HCY converted into methionine by Methionine synthase (MS). SAH has been shown to inhibit Hua enhancer 1 (HEN1), which is responsible for siRNA stability through 2’-O-methylation of their 3’ pentose sugar (Mäkinen & De, 2019). It was hypothesized that viral encoded RNA silencing suppressor proteins like HC-Pro interact with SAM synthease and reduce its activity which results in decreased levels of SAM [78]. Mixed virus infections of potyvirus and potexvirus create imbalances in MTC and GSH biosynthesis which induce oxidative burst and lead to more symptoms in infected plants [79].
2.2.2. Role of PTGS in virus infection
The replicative intermediates of RNA viruses and the transcripts of DNA viruses are targets of PTGS in the cytoplasm [76]. Plants show symptoms of recovery when the vsiRNAs are generated and spread through the vascular system. Arabidopsis mutants of sgs3, ago1, sgs2 showed greater susceptibility to virus infection [80]. Studies with Tomato bushy stunt virus (TBSV) infection in Nicotiana benthamiana and PVX and Turnip crinkle virus (TCV) in Arabidopsis have shown that the antiviral defense involves the AGO2 pathway [81,82,83].
To counteract RNA silencing, viral proteins contain suppressor of RNA silencing (VSR) activity (Figure 1). The VSRs interact with the components of PTGS at multiple points. They can interrupt silencing process by making dsRNAs unavailable for DCL action, inhibiting the siRNAs transport, quenching the siRNAs to prevent their loading in RISC and inactivation of RISC [84]. For instance, 2b protein, the VSR of Tomato aspermy virus (TAV), shows non-specific binding with siRNA and dsRNA [85]. Whereas, V2, the VSR is of Tomato yellow leaf curl virus binds with SGS3 which has an important role in dsRNA amplification [86,87].
2.2.3. small RNA shields in virus resistance
RNA silencing has been effectively used for generating virus resistant plants [49,88]. A variety of small RNAs have been used through various strategies to trigger the RNA silencing mediated defense responses against viruses (Table 1). The regions of the viral genome that encode VSRs eg. P69, HC-Pro and AC2 are usually selected as the targets of the small RNAs. Expression of small RNAs can be from various precursors, which may be in cis or trans and single or polycistronic [89,90]. Initial GM approaches involved the use of antisense constructs or hp-RNAs as precursors to generate a pool of siRNAs, which could target the viral transcripts. Subsequently, these were replaced by more specific tools through the use of constructs expressing antiviral artificial microRNAs (amiRNAs) and synthetic trans-acting small interfering RNAs (syn-tasiRNAs).
1) microRNA shield
MicroRNAs (miRNAs) are genome encoded small RNAs that are processed by DCL1 from imperfectly paired, hairpin precursor RNAs [114,115]. The miRNAs belong to large gene families and each miRNA can modulate plethora of targets involved in plant immunity and phytohormonal pathways [116]. High throughput sequencing tools have provided useful insights for identifying the miRNAs involved in virus resistance. miR156, miR395, miR159, miR166, miR168, miR160 and miR444 have important roles in virus attack [117,118,119,120]. Recent report has shown that a novel miRNA, Seq 119 is down-regulated during Rice stripe virus (RSV) infection. Overexpression of this miRNA reverses the symptoms caused by RSV [121].
In many cases, amiRNA have been used to over-express a synthetic sequence designed to target a key viral gene. In Arabidopsis plants, amiRNAs were used first to target TYMV and TuMV [122]. This technique has the advantage of enhanced specificity and reduce off target effects. The amiRNA is composed of two components namely, a miRNA precursor (pre-miR) scaffold and a synthetic RNA insert. Special consideration is placed on selection of the pre-miR backbone to ensure that the correct amiRNA is processed by the cellular machinery [123]. Studies with TuMV population dynamics showed that inadequate levels of amiRNA build up more mutations in the target site and reduce the efficacy of virus resistance [124].
To synthesize the amiRNA, the miRNA/miRNA* sequence of the pre-miR is replaced by a synthetic sequence complementary to the viral target by performing overlapping PCR [125]. The sequence of the amiRNA is designed to have maximum complementarity with the target and minimum hybridization energy while ensuring sufficient mismatches to retain the secondary structures or bulges [126,127]. The other parameters include avoidance of mismatches at 10 or 11th nucleotides and instability at the 5’ end. The design of an effective amiRNA also necessitates incorporation of Uracil (U) at first base and Adenine (A) or U at 10th nucleotide position for making amiRNA biologically active [126,128]. Targeting an exposed region in a viral genome can increase the chances of DCL accessibility and efficiency of the amiRNA [129]. Tomato transgenics overproducing the amiRNAs to silence the conserved regions of ToLCV- AV2/AC2 genes could tolerate various leaf curl viruses of tomato [130]. The amiRNA technique faced the limitations of stringent design principles and the level of amiRNA expression.
2) siRNA shield
The siRNAs are 21-24 nt in length and are derived from perfectly paired dsRNA molecules [131,132]. The siRNA-mediated gene silencing serves as a primary defense mechanism against plant viruses [76,133,134,135,136]. Transitivity of siRNAs is useful in propagating the silencing signal through the production of secondary siRNAs [55,137].
It was observed that better virus resistance was achieved when a dsRNA of the viral sequence was expressed in plant cell compared to the case of expressing either sense or antisense viral RNA [138]. This resulted in construction of intron splicable hairpin RNA (hpRNA) constructs with viral gene or gene-fragments. The hpRNA constructs offered the strongest resistance to RNA and DNA viruses [139,140,141]. The GM approaches involve expression of short-hairpin RNA (shRNA) constructs as precursors of the siRNAs in the plants to tackle the virus. The precursors are mobilized into the Agrobacterium and stably integrated into plants [142]. In majority of the cases, the VSR gene (s) have been used as a silencing target to generate virus resistance [140]. In Brazil, transgenic bean resistant to a Bean golden mosaic virus (BGMV) was created by expressing a hpRNA of Rep gene and it is being commercially cultivated [139].
Tomato plants expressing hpRNA corresponding to the TYLCV- Rep coding sequence produce 21- and 22-nt siRNAs and are resistant to TYLCV [143]. Similarly, Cassava transgenics containing hpRNA homologous to the overlapping region of South African cassava mosaic virus (SACMV)-Rep and the AC1/AC4, were tolerant to cassava mosaic disease [144]. In another study, three distinct intron hpRNA constructs comprising sequences of AC2, AC4, and fusion of AC2 and AC4 (AC2+AC4) of seven begomoviruses were used. All transgenic lines showed resistance against MYMIV infection when compared with untransformed controls [145].
3) Synthetic tasiRNA shield
The tasiRNAs are derived from non-protein-coding TAS transcripts that are capped as well as poly-adenylated and contain a binding site mostly for 22-nt miRNA [146]. The miRNA cleavage products are stabilized by Suppressor of Gene Silencing 3 (SGS3) and converted into a dsRNA form by RDR6 [147,148]. The dsRNA intermediate is then processed by DCL4 and dedicated dsRNA-Binding Protein 4 (DRB4) to phased 21-nt siRNAs in a ‘head-to-tail’ phased pattern. The transitive siRNAs are incorporated into AGO-RISC for targeting complementary sequences [147,148,149,150]. In Arabidopsis thaliana eight tasiRNA producing loci have been identified that fall into four TAS groups (TAS1-TAS4).
Since TAS precursors are able to generate multiple secondary siRNAs once they are cleaved by the miRNA, they work more efficiently against the rapidly evolving viruses that can evade the single amiRNA approach. The Arabidopsis TAS DNA sequences can be engineered to silence targeted viral sequences, by replacing some of the phased tasiRNA producing sequences by single or multiple siRNAs of different sequences but of equivalent length of base-pairs. When the replacing siRNAs are processed in the tasiRNA pathway from the engineered vectors as desired, these are called as the synthetic-tasiRNAs (syn-tasiRNAs), which consequently silence their targets in a usual manner using the RNA silencing machinery of the host plants. It was observed that most plants expressing higher levels of syn-tasiRNA were resistant to TSWV (Tomato Spotted Wilt Virus), while plants with particularly low levels of syn-tasiRNAs were infected. Several automated design tools have been used for designing syn-tasiRNA constructs, which are able to target distinct locations within a single or multiple viral RNAs. More efficient, long-lasting, and widespread resistance could be produced by simultaneous co-expression of multiple syn-tasiRNAs [151,152].
4) long non-coding RNA shield
Long non coding RNAs (lncRNAs) are 200 nt in length and are mainly produced by the action of polymerase II [153]. The lncRNAs resemble the mRNAs in having a 5'm7G cap and 3’poly(A) tail [154], [155], though their sequences show less conservation [156]. On the basis of origin they can be categorised as intergenic, intronic or exonic. They can act either in cis or trans and may have sense or antisense orientations [157,158]. NGS analysis has played an important role in identification of differentially expressed lncRNAs and their roles in host interaction studies [159]. RNA-seq of TYLCV-resistant tomato cultivar showed that some lncRNAs such as slylnc0048, slylnc00449, slylnc0483, slylnc0531 and slylnc0934 were up regulated while slylnc0475, slylnc0476, slylnc0673 and slylnc1052 were down regulated after virus infection [160].
LncRNAs can act as sources for the generation of small RNAs and regulate DNA or histone methylation by guiding the DNA methyltransferase. LMT1 encoded by Citrus tristeza virus (CTV) plays an important role in susceptibility of host plants. Mutation in LMT1 caused increase in the SA levels and enhanced virus resistance [161]. Recent reports have shown a relation between RNA silencing and viroid replication. Viroids are free RNA molecules which do not code for any protein and these are like circular lncRNAs [162]. Several horticulture crops such as apple, avocado, grapevine, peach, tomato etc are facing viroid infections. Potato spindle tuber viroid (PSTVd) is a good model for studying viroid-host interactions. PSTVd induced small RNAs cause the cleavage of several host transcripts coding for pyrophosphatase, callose etc [163,164].
The lncRNAs can also quench the function of small RNAs and act as target mimics of the miRNAs [165], [157]. SlLNR1 (lncRNA of tomato) involved in regulating leaf growth and development, contained sequences complementary to the vsiRNA derived from intergenic region (IR) of the virus. In susceptible lines, viral intergenic siRNAs targeted the SlLNR1 resulting in stunted growth and curling of leaves [166].
3. GE approaches
Genome editing (GE) offers the most precise tool to modify a target gene by using the endonucleases to create breaks in the DNA strands that lead to both target-specific mutagenesis (addition, removal or alteration) and gene replacement [167,168]. It offers the advantage of being less expensive, simple to design and execute and better acceptable. Several GE techniques have been implemented against different viruses and have produced better outcomes (Table 2). By complementing it with NGS, it will become easier to identify the fast mutating viral sequences and redesign the targets which influence the susceptibility response in host-virus interactions. [169].
Genome editing is now emerging and expanding as a modern tool in biotechnology. From basic techniques such as Meganucleases (MNs), ZFNs (zinc finger nucleases), TALENS (transcription activator-like effector nucleases to advanced techniques like CRISPR/Cas system. GE methods rely on the introduction of specific mutations to inactivate certain target genes. Modifications like CRISPR ON and OFF, use of Base Editors and Prime editors restricts the possibilities of accidental host targets.
3.1. Different GE tools
The onset of GE lies in the use of specialized restriction endonucleases known as meganucleases (MNs), which could target broad recognition sites of around 12-40 bp [170,171]. These nucleases create the double-stranded breaks (DSBs) that were repaired by non-homologous end joining (NHEJ), resulting in gene mutations [172]. MNs have been studied in model plant systems such as Arabidopsis, Gossypium and maize. They exhibited high efficiency of target recognition and moderate mutation rate. The length of target sequence recognised by the MN was around 68-88 bp, which was quite long as compared to others and it did not require another endonuclease (Ahmar S et al 2020). However, the major limitation was that production of these enzymes was costlier and time consuming. It was not easy to manipulate the MNs as their catalytic and DNA binding domains were intermingled and become difficult to detach [173].
Zinc-finger nucleases (ZFN) emerged as specialized cutter that could cleave dsDNA sequences [174], [175,176]. ZFN-induced double-strand breaks were subject to cellular DNA repair processes. ZFNs were engineered by fusing the non-specific DNA cleavage domain of Fok1 (Flavobacterium okeanokoites 1) with the Cys2-His2 zinc finger (ZF) DNA Binding domain [177,178]. Fok1 is a type II endonuclease that can bind to DNA regions in a non-specific manner, but it can cleave the double strand only when two monomers bind to the target sequence (overall 9-18 bp). Each monomer binds to 5-6 bp long sequences within the target region, so 4 monomers were required for generating 2 cleavage sites [179]. The breaks are repaired by homology directed repair (HDR) or NHEJ (when homologous sequences are not found) resulting in errors [180]. NHEJ creates deletions or insertions at target sites that further leads to frame shift and results in gene knockout while HDJ can insert new sequences at the break site that result in knock-in [181].
ZFNs were used to mutate the AC1 gene, which encodes Rep protein, in Tomato yellow leaf curl China virus [182]. Artificial ZFN proteins (AZPs) have been designed using conserved sequences for blocking the DNA binding sites, which are used by viruses [183,184]. For instance, AZPs bind to the IR region of Beet severe curly top virus (BSCTV) and interfere with binding of Rep thus suppressing replication of virus in the host [185]. The ZFN based technique allowed easy manipulation and could target any site but the efficiency was limited by chances of off-target cleavage. ZFN mediated gene editing has been used in many crops such as Arabidopsis, tobacco, maize, and soybean etc. [172,177,186,187].
TALENS (Transcription Activator-like effectors Nucleases) were produced by fusing DNA binding domains of transcription activator-like (TAL) effectors to DNA cleavage domains of Fok1 to induced dsDNA breaks that could be repaired by NHEJ or HDR repair mechanisms [188]. TAL effectors are produced by bacteria Xanthomonas in plants through the type III secretion system. [189]. These effectors alter the transcripts of the host by binding with the promoter region via repetitive amino acid residues in the central domain and activating gene expression [190,191]. TALENs were used to generate disease resistant plants by mutagenizing the susceptible genes in host plants [192]. TALENs were used to develop resistance of tobacco plants against Tobacco curly shoot virus (TbCSV) and Tomato leaf curl China virus (TYLCCNV) [193].
Recently, CRISPR-Cas has become popular in the scientific community as it has tremendous potential to improve crop traits (section 3.2). The attractive features of this technology include ease, efficiency, and rapidity of genetic manipulations. CRISPR system is rapidly becoming diverse and now includes different types of base editors, prime editors and endogenous tRNA systems. These modifications have given greater specificity and multiplex targeting abilities, which ultimately makes GE more precise (Table 2).
3.2. Manipulation of genome through CRISPR/Cas system
The CRISPR-Cas system was first identified as unique strategy used by bacteria and archaea, for adaptive immune response against invading phages or plasmids [194,195,196]. In this system, the prokaryotes were able to acquire small pieces of the infecting virus and then use it as memory for protection against further attacks. CRISPR stands for Cluster Regularly Interspaced Short Palindromic Repeats and the Cas protein is the RNA dependent DNA endonuclease. The Cas protein interacts with two special non-coding RNAs namely CRISPR RNA (crRNA) and transactivating crRNA (tracrRNA). The crRNA function as a short guide as it consists of a unique 20-base sequence that enables target recognition. The tracrRNA is a trans-encoded small RNA, which provides the stem loop structure [197]. The fusion of crRNA and tracrRNA generates a synthetic RNA of around 100 nucleotides (nt), called the single guide RNA (sgRNA).
In the initial step, Cas9 protein binds to sgRNA to form a catalytically active Cas9-sgRNA duplex [198,199]. The Cas 9 protein contains two REC domains, an arginine-rich Bridge Helix, a proto-spacer adjacent motif (PAM) interacting domain, and two DNA nuclease domains (HNH and Ruv C). The REC I domain is responsible for binding to the sgRNA, while the bridge helix is crucial for initiating cleavage activity upon binding of target DNA [200]. The PAM domain helps in recognizing the consensus NGG (N, any nucleotide; G, guanine) at 3’ end of the target DNA strand for initiating binding of Cas9 to target DNA [197,201,202,203]. After binding the Cas9 unzips the DNA to allow crRNA (within sgRNA) to anneal to the target DNA. After annealing, both the strands are cleaved 3 bases upstream to the PAM thereby creating blunt ends at the target site. The DSB can be repaired by the error prone NHEJ or HDJ to generate the mutations.
3.2.1. Cas9 editing
This technique has been beneficial to target DNA and RNA viruses with precision and high efficiency (Table 2) [204,205]. Studies using Arabidopsis thaliana or Nicotiana benthamiana showed that sgRNAs targeting the stem-loop sequence within the geminiviral genome or the non-coding intergenic region (IR) showed a stronger interference as compared those targeting the viral genes like CP or Rep. When gRNAs were used to target non-coding IRs in CLCuKoV and TYLCSV their systemic movement in host was blocked [206]. The sgRNAs also rendered a broad-spectrum resistance to different mono- & bipartite geminiviruses [207]. Combination of two sgRNAs against IR and Rep of CLCuMuV in a single plasmid enhanced the overall resistance of tobacco plants [208].
Several extensions or modifications of the CRISPR/Cas system have been developed either by mutating the Cas 9 enzyme or by using other variants of Cas. One of the class 2 effectors, Cas12a (also called Cpf1) has only RuvC domain and no HNH domain, hence it generates staggered DSBs instead of blunt [209,210]. Staggered DSB is near to the 3’ end of target sequence, which creates a 5’ overhang. This could be beneficial for knock-in experiments and also helpful in insertion of DNA fragments precisely and with perfect orientation through complementary ends by HDR [210]. Also it requires shorter crRNA than Cas9 and there is no requirement for tracrRNA as per the current reports [211].
3.2.2. Epigentic editors
An engineered Cas variant known as dead Cas9 (dCas9) contains inactivated HNH and RuvC nuclease domains [180]. It targets specific genomic region by recruiting chromatin modifiers and effector proteins, which leads to programmable epigenetic editing [212,213]. There is enormous potential for dCas9 to gain insights on plant-virus interactions and for performing transcriptional engineering and chromatin alterations [214].
3.2.3. RNA editors
The FnCas9 (class 2, Type II) derived from Francisella novicida, has RNA targeting and editing properties [215,216]. The FnCas9 with RNA-targeting gRNA has been used to cleave the ssRNA viruses for the resistance against CMV and TMV in N. benthamiana and A. thaliana [217]. CRISPR techniques especially CRISPR-Cas12a and CRISPR-Cas13a/d are also useful in detection of DNA and RNA virus amplicons generated through isothermal amplification [218].
Cas13 is a new type of protein isolated from Ruminococcus flavefaciens that comes in class 2 type VI effectors [219]. It has programmable RNA-targeting properties, which broaden the application of CRISPR/Cas system [204]. Recent studies have demonstrated the effectiveness of using various Cas13 systems successfully for targeting multiple RNA viruses [220]. Transgenic rice and tobacco harbouring LshCas13a (Leptotrichia shahii Cas13) with specific protospacer showed resistance to Rice stripe mosaic virus (RSMV), Tobacco mosaic virus (TMV), and Southern rice black-streaked dwarf virus (SRBSDV) [221]. The effectiveness of using LshCas13a to target the helper component proteinase (HC-Pro) and coat protein (CP) segments of the Turnip Mosaic Virus (TuMV) RNA genome in Nicotiana benthamiana and Arabidopsis thaliana, respectively has been demonstrated. Cas13 has many variants of which Cas13d is more advantageous than Cas13a, Cas13b, or others [222]. CRISPR techniques especially CRISPR-Cas12a and CRISPR-Cas13a/d are also useful in detection of DNA and RNA virus amplicons generated through isothermal amplification [218].
3.2.4. Base editors
These are formed by the fusion of enzyme deaminase with Cas nuclease domain. Base editors are used to create point mutations without the requirement of donor template and DSBs [223,224]. This approach is useful for targeting genes responsible for host susceptibility [225]. Cytosine base editors (CBE) and Adenosine base editors (ABE) are two commonly used editors in base editing. The former converts C:G to T:A and later converts A:T to G:C [226]. In this process, the availability of PAM is important to edit the target DNA. Advancements in the base editing technology such as variants of CBEs and ABEs are being used to create randomized mutations in targets [227,228]. In Arabidopsis resistance to Clover yellow vein virus (CIYVV) was obtained by using a novel CBE to convert a susceptible allele of eIF4E1 into a resistance allele by a point mutation of N176K [225].
3.2.5. Prime editors
Prime editing was achieved by using an engineered nCas9 fused with a reverse transcriptase and prime-editing guide RNA (pegRNA). The pegRNA has protospacer sequence that directs Cas nuclease to cleave target region and add extra sequences, which are responsible for specific changes [229,230]. Like base editing, this technique does not require double stranded breaks (DSBs).
3.2.6. Multiplexing CRISPR/Cas editing
Targeting multiple sites on a virus in a single go is being developed as an effective strategy to minimize the chances of virus escape. In cotton, three sgRNAs were designed to target three viral regions for modifying six overlapping genes (AV2/AV1, AC2/AC3, AC1/AC4). The analysis showed relatively low virus titres in the plants transformed with all three gRNAs as compared to plants expressing single gRNA and the wild-type plants [231].
This includes simultaneous delivery of Cas9 mRNA with multiple gRNAs expression cassettes in one plasmid, artificial gRNAs with Cas9 flanked by Csy4/HH-HDV ribozymes for cleavage and release of gRNAs, multiple gRNA precursors with Cas9 flanked by tRNAs etc and so on [232]. Coupling of CRISPR/Cas13d with an endogenous tRNA system for expression of four sgRNA was found to be effective against PVY, PVX, PVS or PLRV viruses in potato [233]. Combination of dCas9 with cytosine/adenosine deaminase and gRNA complex was used for generating point mutations in targets [223,234].
Multiple gRNA strategy is also helpful in case of mixed viral infections [235]. In Potato, CRISPR/Cas13a was coupled with an endogenous tRNA system and four different gRNAs to produce a polycistronic tRNA-gRNA (PTG) against PVY. However the transgenic lines exhibited similar levels of tolerance as observed in case of single gRNA lines, indicating that multiple target sites did not affect viral interference [233].
Major hurdles in the GE approaches are the generation of off-targets due to the constitutive expression of Cas nuclease or because of sequence mismatches in sgRNA (Mushtaq, M et al., 2020). Error prone repairing process creates selection pressure on viruses which results in generation of viral variants having resistance towards editing. Replacement of constitutive promoter by a virus inducible promoter for expression of Cas nuclease is quite useful to bring down the off-targets [236].

Table 2.
List of GE approaches available against Plant DNA and RNA viruses.
| Strategy | Virus | Plant | References |
|---|---|---|---|
| For DNA Viruses | |||
| ZFN | TYLCCNV, TbCSV BeYDV |
Tobacco |
[182] [237] |
| TALENS | TYLCCNV, TbCSV, TLCYV | [193] | |
| AZP | BSCTV, TYLCV RTBV |
[183], [238], [239], [240], [241], [242] | |
| CRISPR/Cas9 | TYLCV, CLcKV, BSCTV, BCTV, BeYDV, CLcV CaMV, BSCTV WDV BSV ACMV |
Tobacco Arabidopsis Barley Banana Cassava |
[243], [206] [244] [245] [246] [247] [248], [249] |
| For RNA Viruses | |||
| MNs | TRV | Tobacco | [250] |
| SpCas9 | BYSMV SYNV |
Tobacco | [251] [252] |
| FnCas9 | TMV, CMV CMV |
Tobacco Arabidopsis |
[217] |
| LshCas13a | TuMV TMV SRBSDV, RSMV PVY |
Tobacco Rice Potato |
[205] [221] |
| Cas13d/PTG | PVX or PLRV, PVY, PVS | Potato | [233] |
| Cas13a/PTG | PVY | Potato | [233] |
| Cas13a with multiplex gRNAs | PVY | Potato | [253] |
| LbCas12a | CLcMV TEV |
Cotton Tobacco |
[254] [255] |
| Cas12f | CLCuV | Cotton | [256] |
| Multiplex Cas9-gRNAs (9 duplex and 2 triplex) | ChiLCV | Tobacco | [245] |
Summary and Future perspectives
Viruses invade plant cells and hijack the host cellular machineries for their own multiplication. The plants fight back by activating the cellular defense responses resulting in genetic and epigenetic events to block viral gene expression. Several attempts were made to introgress resistance to virus in cultivated plants by breeding with wild relatives. After decades of failure, strategies for targeting the virus to confer resistance offered great promise in saving the crops from virus infection. Therefore, GM and GE have emerged as effective strategies to engineer resistance to the viruses (Figure 1).
The early GM approaches were based on engineering PDR by over-expressing CP or other viral proteins and it proved to be effective against many viruses. However, with the discovery and understanding of the underlying principles of RNA silencing and its role in providing resistance to plant viruses, there was a paradigm shift in the strategies. The first generation silencing tools employed the dsRNA or hpRNA precursors directed against specific viral genes. The transgenic papayas expressing siRNAs against Papaya ringspot virus (PRSV) proved to be a boon for the Hawaian papaya industry. However, the siRNAs had the limitations of being less specific and generating undesirable off targets [260]. This resulted in development of amiRNA tools by engineering the natural pre-miRNA backbones. These second generation tools were more specific but had limited targeting capacity, so syn-tasiRNA vectors were favoured and developed as the third generation tools. The GM technology has helped to save the papaya grown in Hawaii and reduce potato crop losses in USA and Canada. It holds the potential to prevent losses in other crop to virus and other diseases.
However, the scepticism regarding acceptance of GM crops by the public has hindered the active implementation of the technology. This branched off the search for exogenous application of dsRNA precursors or small RNAs to activate the host silencing pathways (Table 3). Several reports have highlighted the use of high pressure (4-6 bar) for efficient transfer of dsRNA in the nucleus for spray induced epigenetic modifications (SPIEM). When high pressure spraying was used to transfer 333 bp dsRNA in N. benthamiana, there was no induction of methylation at the target region at 3 days post spraying (dps), while 28% samples were methylated at 6 dps and 40% of samples were methylated at 10 dps [262]. However, spraying with low pressure did not result in any methylation. The RNA based bioformulations are in various stages of development and are anticipated to prove useful in tackling the virus infections.
The emergence of GE tools has added another arsenal in the fight against viruses infecting the crop plants. CRISPR/Cas9 provides a robust system to generate more effective and prolonged resistance against viruses. The recent additions of Cas13 and Cas12 variants have proved much effective in targeting the RNA viruses. The efficacy of CRISPR-based approaches primarily depends on the method of delivery and design of the sgRNA sequence. A number of delivery methods including Agrobacterium-mediated T-DNA transformation, protoplast transfection, microprojectile bombardment and virus-based methods have been deployed to introduce CRISPR/Cas components in plants. As viruses have the tendency to mutate with the faster rate, the multiplexing technologies can contribute for precise editing and broadening the range of the targeted viruses. Although the use of CRISPR/Cas for development of viral-resistant plants has been demonstrated to be effective, further improvements in the technology are awaited to attain efficient, durable and broad-spectrum resistance.
Author Contributions
A.R. wrote the manuscript and drew the figures. N.S.-M. edited and improved the manuscript. Both authors have read and approved the manuscript.
Funding
This work was financially supported by grants from ICGEB. A.R. acknowledges the Fellowship support from CSIR.
Institutional Review Board Statement
Not applicable
Informed Consent Statement
Not applicable
Data Availability Statement
Not applicable
Conflicts of Interest
The authors declare no conflict of interest
References
- Anderson, P.K.; Cunningham, A.A.; Patel, N.G.; Morales, F.J.; Epstein, P.R.; Daszak, P. Emerging infectious diseases of plants: pathogen pollution, climate change and agrotechnology drivers. Trends in ecology & evolution 2004, 19, 535–544. [Google Scholar]
- Jones, R.A. Global plant virus disease pandemics and epidemics. Plants 2021 10, 233. [CrossRef]
- Sastry, K.S.; A. Zitter, T.; Sastry, K.S.; Zitter, T.A. Management of virus and viroid diseases of crops in the tropics. Plant Virus and Viroid Diseases in the Tropics: Volume 2: Epidemiology and Management 2014, 149-480.
- Zhao, Y.; Yang, X.; Zhou, G.; Zhang, T. Engineering plant virus resistance: from RNA silencing to genome editing strategies. Plant Biotechnology Journal 2020, 18, 328–336. [Google Scholar] [CrossRef] [PubMed]
- Johnson, A.A.; Gopal, D.S.; Sudhakar, C. GM Crops for Plant Virus Resistance: A Review. Genetically Modified Crops: Current Status, Prospects and Challenges 2021, 2, 257-337. [Google Scholar]
- Tilman, D.; Balzer, C.; Hill, J.; Befort, B.L. Global food demand and the sustainable intensification of agriculture. Proceedings of the national academy of sciences 2011, 108, 20260–20264. [Google Scholar] [CrossRef] [PubMed]
- Legg, J.P.; Shirima, R.; Tajebe, L.S.; Guastella, D.; Boniface, S.; Jeremiah, S.; Nsami, E.; Chikoti, P.; Rapisarda, C. Biology and management of Bemisia whitefly vectors of cassava virus pandemics in Africa. Pest management science 2014, 70, 1446–1453. [Google Scholar] [CrossRef] [PubMed]
- Lu, B.; Stubbs, G.; Culver, J.N. Coat protein interactions involved in tobacco mosaic tobamovirus cross-protection. Virology 1998, 248, 188–198. [Google Scholar] [CrossRef] [PubMed]
- Muller, G.W.; Rezende, J.A. Preimmunization: Applications and perspectives in virus disease control. In Diseases of Fruits and Vegetables Volume I: Diagnosis and Management, Dordrecht: Springer Netherlands 2004, (pp. 361-395).
- Sanford, J.C.; Johnston, S.A. The concept of parasite-derived resistance—deriving resistance genes from the parasite's own genome. Journal of Theoretical Biology 1985, 113, 395–405. [Google Scholar] [CrossRef]
- Nicaise, V. Crop immunity against viruses: outcomes and future challenges. Frontiers in plant science 2014, 5, 660. [Google Scholar] [CrossRef] [PubMed]
- Galvez, L.C.; Banerjee, J.; Pinar, H.; Mitra, A. Engineered plant virus resistance. Plant Science 2014, 228, 11–25. [Google Scholar] [CrossRef] [PubMed]
- Baulcombe, D. RNA silencing in plants. Nature 2004, 431, 356–363. [Google Scholar] [CrossRef]
- Csorba, T.; Pantaleo, V.; Burgyán, J. RNA silencing: an antiviral mechanism. Advances in virus research 2009, 75, 35–230. [Google Scholar] [PubMed]
- Sharma, K.K.; Bhatnagar-Mathur, P.; Thorpe, T.A. Genetic transformation technology: status and problems. In Vitro Cellular & Developmental Biology-Plant 2005, 41, 102–112. [Google Scholar]
- Fitchen, J.H.; Beachy, R.N. Genetically engineered protection against viruses in transgenic plants. Annual Review of Microbiology 1993, 47, 739–763. [Google Scholar] [CrossRef] [PubMed]
- Baulcombe, D. Novel strategies for engineering virus resistance in plants. Current Opinion in Biotechnology 1994, 5, 117–124. [Google Scholar] [CrossRef]
- Abel, P.P.; Nelson, R.S.; De, B.; Hoffmann, N.; Rogers, S.G.; Fraley, R.T.; Beachy, R.N. Delay of disease development in transgenic plants that express the tobacco mosaic virus coat protein gene. Science 1986, 232, 738–743. [Google Scholar] [CrossRef] [PubMed]
- Beachy, R.N. Coat–protein–mediated resistance to tobacco mosaic virus: discovery mechanisms and exploitation. Philosophical Transactions of the Royal Society of London. Series B: Biological Sciences 1999, 354, 659–664. [Google Scholar] [CrossRef]
- Dasgupta, I.; Malathi, V.G.; Mukherjee, S.K. Genetic engineering for virus resistance. Current science 2003, 84, 341–354. [Google Scholar]
- Nejidat, A.; Beachy, R.N. Transgenic tobacco plants expressing a coat protein gene. Molecular Plant-Microbe Interactions 1990, 3, 247–251. [Google Scholar] [CrossRef]
- Anderson, E.J.; Stark, D.M.; Nelson, R.S.; Powell, P.A.; Tumer, N.E.; Beachy, R.N. Transgenic plants that express the coat protein genes of tobacco mosaic virus or alfalfa mosaic virus interfere with disease development of some nonrelated viruses. Phytopathology 1989, 79, 1284–1290. [Google Scholar] [CrossRef]
- Lawson, C.; Kaniewski, W.; Haley, L.; Rozman, R.; Newell, C.; Sanders, P.; Tumer, N.E. Engineering resistance to mixed virus infection in a commercial potato cultivar: resistance to potato virus X and potato virus Y in transgenic Russet Burbank. Bio/technology 1990, 8, 127–134. [Google Scholar] [CrossRef] [PubMed]
- Chapman, S.; Kavanagh, T.; Baulcombe, D. Potato virus X as a vector for gene expression in plants. The Plant Journal 1992, 2, 549–557. [Google Scholar] [CrossRef] [PubMed]
- Taschner, P.E.; Van Marle, G.; Brederode, F.T.; Tumer, N.E.; Bol, J.F. Plants transformed with a mutant alfalfa mosaic virus coat protein gene are resistant to the mutant but not to wild-type virus. Virology 1994, 203, 269–276. [Google Scholar] [CrossRef] [PubMed]
- Reimann-Philipp, U.; Beachy, R.N. December. The mechanism (s) of coat protein-mediated resistance against tobacco mosaic virus. In Seminars in Virology, Academic Press 1993, (Vol. 4, No. 6, pp. 349-356).
- Yusibov, V.M.; Loesch-Fries, L.S. N-terminal basic amino acids of alfalfa mosaic virus coat protein involved in the initiation of infection. Virology 1995, 208, 405–407. [Google Scholar] [CrossRef] [PubMed]
- Clark, W.G.; Fitchen, J.; Nejidat, A.; Deom, C.M.; Beachy, R.N. Studies of coat protein-mediated resistance to tobacco mosaic virus (TMV). II. Challenge by a mutant with altered virion surface does not overcome resistance conferred by TMV coat protein. Journal of general virology 1995, 76, 2613–2617. [Google Scholar] [CrossRef] [PubMed]
- Asurmendi, S.; Berg, R.H.; Smith, T.J.; Bendahmane, M.; Beachy, R.N. Aggregation of TMV CP plays a role in CP functions and in coat-protein-mediated resistance. Virology 2007, 366, 98–106. [Google Scholar] [CrossRef]
- Lapidot, M.; Gafny, R.; Ding, B.; Wolf, S.; Lucas, W.J.; Beachy, R.N. A dysfunctional movement protein of tobacco mosaic virus that partially modifies the plasmodesmata and limits virus spread in transgenic plants. The Plant Journal 1993, 4, 959–970. [Google Scholar] [CrossRef]
- Cooper, B.; Lapidot, M.; Heick, J.A.; Dodds, J.A.; Beachy, R.N. A defective movement protein of TMV in transgenic plants confers resistance to multipleviruses whereas the functional analog increases susceptibility. Virology 1995, 206, 307–313. [Google Scholar] [CrossRef]
- Beck, D.L.; Van Dolleweerd, C.J.; Lough, T.J.; Balmori, E.; Voot, D.M.; Andersen, M.T.; O'Brien, I.E.; Forster, R.L. Disruption of virus movement confers broad-spectrum resistance against systemic infection by plant viruses with a triple gene block. Proceedings of the National Academy of Sciences 1994, 91, 10310–10314. [Google Scholar] [CrossRef] [PubMed]
- Golemboski, D.B.; Lomonossoff, G.P.; Zaitlin, M. Plants transformed with a tobacco mosaic virus nonstructural gene sequence are resistant to the virus. Proceedings of the National Academy of Sciences 1990, 87, 6311–6315. [Google Scholar] [CrossRef]
- Hanson, S.F.; Maxwell, D.P. trans-Dominant inhibition of geminiviral DNA replication by bean golden mosaic geminivirus rep gene mutants. Phytopathology 1999, 89, 480–486. [Google Scholar] [CrossRef] [PubMed]
- Sangaré, A.; Deng, D.; Fauquet, C.M.; Beachy, R.N. Resistance to African cassava mosaic virus conferred by a mutant of the putative NTP-binding domain of the Rep gene (AC1) in Nicotiana benthamiana. Molecular Breeding 1999, 5, 95–102. [Google Scholar] [CrossRef]
- Donson, J.; Kearney, C.M.; Turpen, T.H.; Khan, I.A.; Kurath, G.; Turpen, A.M.; Jones, G.E.; Dawson, W.O.; Lewandowski, D.J. Broad resistance to tobamoviruses is mediated by a modified tobacco mosaic virus replicase transgene. Molecular plant-microbe interactions: MPMI 1993, 6, 635–642. [Google Scholar] [CrossRef]
- Tenllado, F.; García-Luque, I.; Serra, M.T.; Díaz-Ruíz, J.R. Resistance to pepper mild mottle tobamovirus conferred by the 54-kDa gene sequence in transgenic plants does not require expression of the wild-type 54-kDa protein. Virology 1996, 219, 330–335. [Google Scholar] [CrossRef] [PubMed]
- Marano, M.R.; Baulcombe, D. Pathogen-derived resistance targeted against the negative-strand RNA of tobacco mosaic virus: RNA strand-specific gene silencing? The Plant Journal 1998, 13, 537–546. [Google Scholar] [CrossRef]
- Greene, A.E.; Allison, R.F. Recombination between viral RNA and transgenic plant transcripts. Science 1994, 263, 1423–1425. [Google Scholar] [CrossRef] [PubMed]
- Chávez-Calvillo, G.; Contreras-Paredes, C.A.; Mora-Macias, J.; Noa-Carrazana, J.C.; Serrano-Rubio, A.A.; Dinkova, T.D.; Carrillo-Tripp, M.; Silva-Rosales, L. Antagonism or synergism between papaya ringspot virus and papaya mosaic virus in Carica papaya is determined by their order of infection. Virology 2016, 489, 179–191. [Google Scholar] [CrossRef]
- Hu, C.C.; Hsu, Y.H.; Lin, N.S. Satellite RNAs and satellite viruses of plants. Viruses 2009, 1, 1325–1350. [Google Scholar] [CrossRef] [PubMed]
- Palukaitis, P.; Roossinck, M.J.; Dietzgen, R.G.; Francki, R.I. Cucumber mosaic virus. Advances in virus research 1992, 41, 281–348. [Google Scholar]
- Patil, B.L.; Dasgupta, I. Defective interfering DNAs of plant viruses. Critical reviews in plant sciences 2006, 25, 47–64. [Google Scholar] [CrossRef]
- Burgyan, J.; Grieco, F.; Russo, M. A defective interfering RNA molecule in cymbidium ringspot virus infections. Journal of General Virology 1989, 70, 235–239. [Google Scholar] [CrossRef]
- Stanley, J.; Frischmuth, T.; Ellwood, S. Defective viral DNA ameliorates symptoms of geminivirus infection in transgenic plants. Proceedings of the National Academy of Sciences 1990, 87, 6291–6295. [Google Scholar] [CrossRef] [PubMed]
- Kong, Q.; Wang, J.; Simon, A.E. Satellite RNA-mediated resistance to turnip crinkle virus in Arabidopsis involves a reduction in virus movement. The Plant Cell 1997, 9, 2051–2063. [Google Scholar] [PubMed]
- Budzyńska, D.; Zwart, M.P.; Hasiów-Jaroszewska, B. Defective RNA Particles of Plant Viruses—Origin, Structure and Role in Pathogenesis. Viruses 2022, 14, 2814. [Google Scholar] [CrossRef] [PubMed]
- Shen, W.X.; Au, P.C.K.; Shi, B.J.; Smith, N.A.; Dennis, E.S.; Guo, H.S.; Zhou, C.Y.; Wang, M.B. Satellite RNAs interfere with the function of viral RNA silencing suppressors. Frontiers in Plant Science 2015, 6, 281. [Google Scholar] [CrossRef]
- Ibrahim, A.B.; Aragão, F.J. RNAi-mediated resistance to viruses in genetically engineered plants. Plant Gene Silencing: Methods and Protocols. 2015, pp.81-92.
- Mueller, E.; Gilbert, J.; Davenport, G.; Brigneti, G.; Baulcombe, D.C. Homology-dependent resistance: transgenic virus resistance in plants related to homology-dependent gene silencing. The Plant Journal 1995, 7, 1001–1013. [Google Scholar] [CrossRef]
- Schwind, N.; Zwiebel, M.; Itaya, A.; Ding, B.; WANG, M.B.; Krczal, G.; Wassenegger, M. RNAi-mediated resistance to Potato spindle tuber viroid in transgenic tomato expressing a viroid hairpin RNA construct. Molecular plant pathology 2009, 10, 459–469. [Google Scholar] [CrossRef] [PubMed]
- Prins, M.; Goldbach, R. The emerging problem of tospovirus infection and nonconventional methods of control. Trends in microbiology 1998, 6, 31–35. [Google Scholar] [CrossRef] [PubMed]
- Tyagi, H.; Rajasubramaniam, S.; Rajam, M.V.; Dasgupta, I. RNA-interference in rice against Rice tungro bacilliform virus results in its decreased accumulation in inoculated rice plants. Transgenic research 2008, 17, 897–904. [Google Scholar] [CrossRef] [PubMed]
- Shepherd, D.N.; Martin, D.P.; Thomson, J.A. Transgenic strategies for developing crops resistant to geminiviruses. Plant Science 2009, 176, 1–11. [Google Scholar] [CrossRef]
- Sanan-Mishra, N.; Abdul Kader Jailani, A.; Mandal, B.; Mukherjee, S.K. Secondary siRNAs in plants: biosynthesis, various functions, and applications in virology. Frontiers in Plant Science 2021, 12, 610283. [Google Scholar] [CrossRef] [PubMed]
- Tabassum, B.; Nasir, I.A.; Aslam, U.; Husnain, T. How RNA interference combat viruses in plants. Rijeka: InTech, Functional Genomics 2012, pp.113-130. [Google Scholar]
- Mette, M.F.; Aufsatz, W.; Van der Winden, J.; Matzke, M.A.; Matzke, A.J.M. Transcriptional silencing and promoter methylation triggered by double-stranded RNA. The EMBO journal 2000, 19, 5194–5201. [Google Scholar] [CrossRef]
- Kooter, J.M.; Matzke, M.A.; Meyer, P. Listening to the silent genes: transgene silencing, gene regulation and pathogen control. Trends in plant science 1999, 4, 340–347. [Google Scholar] [CrossRef] [PubMed]
- Brodersen, P.; Voinnet, O. The diversity of RNA silencing pathways in plants. TRENDS in Genetics 2006, 22, 268–280. [Google Scholar] [CrossRef]
- Palauqui, J.C.; Elmayan, T.; Pollien, J.M.; Vaucheret, H. Systemic acquired silencing: transgene-specific post-transcriptional silencing is transmitted by grafting from silenced stocks to non-silenced scions. The EMBO journal 1997, 16, 4738–4745. [Google Scholar] [CrossRef] [PubMed]
- Voinnet, O.; Vain, P.; Angell, S.; Baulcombe, D.C. Systemic spread of sequence-specific transgene RNA degradation in plants is initiated by localized introduction of ectopic promoterless DNA. Cell 1998, 95, 177–187. [Google Scholar] [CrossRef] [PubMed]
- Kumakura, N.; Takeda, A.; Fujioka, Y.; Motose, H.; Takano, R.; Watanabe, Y. SGS3 and RDR6 interact and colocalize in cytoplasmic SGS3/RDR6-bodies 2009, FEBS letters, 583, 1261-1266.
- Pélissier, T.; Thalmeir, S.; Kempe, D.; Sänger, H.L.; Wassenegger, M. Heavy de novo methylation at symmetrical and non-symmetrical sites is a hallmark of RNA-directed DNA methylation. Nucleic acids research 1999, 27, 1625–1634. [Google Scholar] [CrossRef] [PubMed]
- Wassenegger, M. RNA-directed DNA methylation. Plant Gene Silencing 2000, 83–100. [Google Scholar]
- Melnyk, C.W.; Molnar, A.; Bassett, A.; Baulcombe, D.C. Mobile 24 nt small RNAs direct transcriptional gene silencing in the root meristems of Arabidopsis thaliana. Current Biology 2011, 21, 1678–1683. [Google Scholar] [CrossRef] [PubMed]
- Brosnan, C.A.; Mitter, N.; Christie, M.; Smith, N.A.; Waterhouse, P.M.; Carroll, B.J. Nuclear gene silencing directs reception of long-distance mRNA silencing in Arabidopsis. Proceedings of the National Academy of Sciences 2007, 104, 14741–14746. [Google Scholar] [CrossRef] [PubMed]
- Pikaard, C.S.; Haag, J.R.; Ream, T.; Wierzbicki, A.T. Roles of RNA polymerase IV in gene silencing. Trends in plant science 2008, 13, 390–397. [Google Scholar] [CrossRef] [PubMed]
- Sigman, M.J.; Panda, K.; Kirchner, R.; McLain, L.L.; Payne, H.; Peasari, J.R.; Husbands, A.Y.; Slotkin, R.K.; McCue, A.D. An siRNA-guided ARGONAUTE protein directs RNA polymerase V to initiate DNA methylation. Nature Plants 2021, 7, 1461–1474. [Google Scholar] [CrossRef]
- Raja, P.; Sanville, B.C.; Buchmann, R.C.; Bisaro, D.M. Viral genome methylation as an epigenetic defense against geminiviruses. Journal of virology 2008, 82, 8997–9007. [Google Scholar] [CrossRef]
- Mirouze, M.; Paszkowski, J. Epigenetic contribution to stress adaptation in plants. Current opinion in plant biology 2011, 14, 267–274. [Google Scholar] [CrossRef]
- Wassenegger, M. The role of the RNAi machinery in heterochromatin formation. Cell 2005, 122, 13–16. [Google Scholar] [CrossRef] [PubMed]
- Aufsatz, W.; Mette, M.; Matzke, A.; Matzke, M. The role of MET1 in RNA-directed de novo and maintenance methylation of CG dinucleotides. Plant molecular biology 2004, 54, 793–804. [Google Scholar] [CrossRef] [PubMed]
- Cao, X.; Aufsatz, W.; Zilberman, D.; Mette, M.F.; Huang, M.S.; Matzke, M.; Jacobsen, S.E. Role of the DRM and CMT3 methyltransferases in RNA-directed DNA methylation. Current biology 2003, 13, 2212–2217. [Google Scholar] [CrossRef] [PubMed]
- Pooggin, M.M. How can plant DNA viruses evade siRNA-directed DNA methylation and silencing? International journal of molecular sciences 2013, 14, 15233–15259. [Google Scholar] [CrossRef]
- Raja, P.; Wolf, J.N.; Bisaro, D.M. RNA silencing directed against geminiviruses: post-transcriptional and epigenetic components. Biochimica et Biophysica Acta (BBA)-Gene Regulatory Mechanisms 2010, 1799, 337–351. [Google Scholar] [CrossRef] [PubMed]
- Ghoshal, B.; Sanfaçon, H. Symptom recovery in virus-infected plants: revisiting the role of RNA silencing mechanisms. Virology 2015, 479, 167–179. [Google Scholar] [CrossRef] [PubMed]
- Zhai, J.; Bischof, S.; Wang, H.; Feng, S.; Lee, T.F.; Teng, C.; Chen, X.; Park, S.Y.; Liu, L.; Gallego-Bartolome, J.; Liu, W. A one precursor one siRNA model for Pol IV-dependent siRNA biogenesis. Cell 2015, 163, 445–455. [Google Scholar] [CrossRef] [PubMed]
- Ivanov, K.I.; Eskelin, K.; Bašić, M.; De, S.; Lõhmus, A.; Varjosalo, M.; Mäkinen, K. Molecular insights into the function of the viral RNA silencing suppressor HC-Pro. The Plant Journal 2016, 85, 30–45. [Google Scholar] [CrossRef]
- Mäkinen, K.; De, S. The significance of methionine cycle enzymes in plant virus infections. Current opinion in plant biology 2019, 50, 67–75. [Google Scholar] [CrossRef]
- Vaucheret, H.; Béclin, C.; Fagard, M. Post-transcriptional gene silencing in plants. Journal of cell science 2001, 114, 3083–3091. [Google Scholar] [CrossRef] [PubMed]
- Scholthof, K.B.G. Taking Some of the Mystery out of Host∶ Virus Interactions. PLoS Pathogens 2011, 7, e1002033. [Google Scholar] [CrossRef] [PubMed]
- Jaubert, M.; Bhattacharjee, S.; Mello, A.F.; Perry, K.L.; Moffett, P. ARGONAUTE2 mediates RNA-silencing antiviral defenses against Potato virus X in Arabidopsis. Plant physiology 2011, 156, 1556–1564. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Zhang, X.; Singh, J.; Li, D.; Qu, F. Temperature-dependent survival of Turnip crinkle virus-infected arabidopsis plants relies on an RNA silencing-based defense that requires dcl2, AGO2, and HEN1. Journal of virology 2012, 86, 6847–6854. [Google Scholar] [CrossRef] [PubMed]
- Wieczorek, P.; Obrępalska-Stęplowska, A. A single amino acid substitution in movement protein of tomato torrado virus influences ToTV infectivity in Solanum lycopersicum. Virus research 2016, 213, 32–36. [Google Scholar] [CrossRef] [PubMed]
- Chen, P.Y.; Weinmann, L.; Gaidatzis, D.; Pei, Y.; Zavolan, M.; Tuschl, T.; Meister, G. Strand-specific 5′-O-methylation of siRNA duplexes controls guide strand selection and targeting specificity. Rna 2008, 14, 263–274. [Google Scholar] [CrossRef] [PubMed]
- Fukunaga, R.; Doudna, J.A. dsRNA with 5′ overhangs contributes to endogenous and antiviral RNA silencing pathways in plants. The EMBO journal 2009, 28, 545–555. [Google Scholar] [CrossRef]
- Glick, E.; Zrachya, A.; Levy, Y.; Mett, A.; Gidoni, D.; Belausov, E.; Citovsky, V.; Gafni, Y. Interaction with host SGS3 is required for suppression of RNA silencing by tomato yellow leaf curl virus V2 protein. Proceedings of the National Academy of Sciences 2008, 105, 157–161. [Google Scholar] [CrossRef] [PubMed]
- Voinnet, O. Induction and suppression of RNA silencing: insights from viral infections. Nature Reviews Genetics 2005, 6, 206–220. [Google Scholar] [CrossRef] [PubMed]
- Fahim, M.; Millar, A.A.; Wood, C.C.; Larkin, P.J. Resistance to Wheat streak mosaic virus generated by expression of an artificial polycistronic microRNA in wheat. Plant biotechnology journal 2012, 10, 150–163. [Google Scholar] [CrossRef] [PubMed]
- KUNG, Y.J.; LIN, S.S.; HUANG, Y.L.; CHEN, T.C.; Harish, S.S.; CHUA, N.H.; YEH, S.D. Multiple artificial microRNAs targeting conserved motifs of the replicase gene confer robust transgenic resistance to negative-sense single-stranded RNA plant virus. Molecular plant pathology 2012, 13, 303–317. [Google Scholar] [CrossRef] [PubMed]
- Hu, Q.; Niu, Y.; Zhang, K.; Liu, Y.; Zhou, X. Virus-derived transgenes expressing hairpin RNA give immunity to Tobacco mosaic virus and Cucumber mosaic virus. Virology journal 2011, 8, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Nicola-Negri, E.D.; Brunetti, A.; Tavazza, M.; Ilardi, V. Hairpin RNA-mediated silencing of Plum pox virus P1 and HC-Pro genes for efficient and predictable resistance to the virus. Transgenic Research 2005, 14, 989–994. [Google Scholar] [CrossRef] [PubMed]
- Kamachi, S.; Mochizuki, A.; Nishiguchi, M.; Tabei, Y. Transgenic Nicotiana benthamiana plants resistant to cucumber green mottle mosaic virus based on RNA silencing. Plant Cell Reports 2007, 26, 1283–1288. [Google Scholar] [CrossRef] [PubMed]
- Schwind, N.; Zwiebel, M.; Itaya, A.; Ding, B.; WANG, M.B.; Krczal, G.; Wassenegger, M. RNAi-mediated resistance to Potato spindle tuber viroid in transgenic tomato expressing a viroid hairpin RNA construct. Molecular plant pathology 2009, 10, 459–469. [Google Scholar] [CrossRef] [PubMed]
- Fuentes, A.; Ramos, P.L.; Fiallo, E.; Callard, D.; Sánchez, Y.; Peral, R.; Rodríguez, R.; Pujol, M. Intron–hairpin RNA derived from replication associated protein C1 gene confers immunity to Tomato yellow leaf curl virus infection in transgenic tomato plants. Transgenic Research 2006, 15, 291–304. [Google Scholar] [CrossRef] [PubMed]
- Zrachya, A.; Kumar, P.P.; Ramakrishnan, U.; Levy, Y.; Loyter, A.; Arazi, T.; Lapidot, M.; Gafni, Y. Production of siRNA targeted against TYLCV coat protein transcripts leads to silencing of its expression and resistance to the virus. Transgenic research 2007, 16, 385–398. [Google Scholar] [CrossRef]
- Arif, M.; Azhar, U.; Arshad, M.; Zafar, Y.; Mansoor, S.; Asad, S. Engineering broad-spectrum resistance against RNA viruses in potato. Transgenic research 2012, 21, 303–311. [Google Scholar] [CrossRef] [PubMed]
- Krubphachaya, P.; Juricek, M.; Kertbundit, S. Induction of RNA-mediated resistance to papaya ringspot virus type W. BMB Reports 2007, 40, 404–411. [Google Scholar] [CrossRef] [PubMed]
- Tougou, M.; Furutani, N.; Yamagishi, N.; Shizukawa, Y.; Takahata, Y.; Hidaka, S. Development of resistant transgenic soybeans with inverted repeat-coat protein genes of soybean dwarf virus. Plant cell reports 2006, 25, 1213–1218. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.J.; Kim, M.J.; Pak, J.H.; Im, H.H.; Lee, D.H.; Kim, K.H.; Lee, J.H.; Kim, D.H.; Choi, H.K.; Jung, H.W.; Chung, Y.S. RNAi-mediated Soybean mosaic virus (SMV) resistance of a Korean Soybean cultivar. Plant Biotechnology Reports 2016, 10, 257–267. [Google Scholar] [CrossRef]
- Ludlow, E.J.; Mouradov, A.; Spangenberg, G.C. Post-transcriptional gene silencing as an efficient tool for engineering resistance to white clover mosaic virus in white clover (Trifolium repens). Journal of Plant Physiology 2009, 166, 1557–1567. [Google Scholar] [CrossRef] [PubMed]
- Vanderschuren, H.; Alder, A.; Zhang, P.; Gruissem, W. Dose-dependent RNAi-mediated geminivirus resistance in the tropical root crop cassava. Plant molecular biology 2009, 70, 265–272. [Google Scholar] [CrossRef] [PubMed]
- Shekhawat, U.K.; Ganapathi, T.R.; Hadapad, A.B. Transgenic banana plants expressing small interfering RNAs targeted against viral replication initiation gene display high-level resistance to banana bunchy top virus infection. Journal of general virology 2012, 93, 1804–1813. [Google Scholar] [CrossRef] [PubMed]
- Soler, N.; Plomer, M.; Fagoaga, C.; Moreno, P.; Navarro, L.; Flores, R.; Pena, L. Transformation of Mexican lime with an intron-hairpin construct expressing untranslatable versions of the genes coding for the three silencing suppressors of Citrus tristeza virus confers complete resistance to the virus. Plant Biotechnology Journal 2012, 10, 597–608. [Google Scholar] [CrossRef]
- Winterhagen, P.; Dubois, C.; Sinn, M.; Wetzel, T.; Reustle, G.M. Gene silencing and virus resistance based on defective interfering constructs in transgenic Nicotiana benthamiana is not linked to accumulation of siRNA. Plant Physiology and Biochemistry 2009, 47, 739–742. [Google Scholar] [CrossRef] [PubMed]
- Patil, B.L.; Bagewadi, B.; Yadav, J.S.; Fauquet, C.M. Mapping and identification of cassava mosaic geminivirus DNA-A and DNA-B genome sequences for efficient siRNA expression and RNAi based virus resistance by transient agro-infiltration studies. Virus research 2016, 213, 109–115. [Google Scholar] [CrossRef] [PubMed]
- Shimizu, T.; Ogamino, T.; Hiraguri, A.; Nakazono-Nagaoka, E.; Uehara-Ichiki, T.; Nakajima, M.; Akutsu, K.; Omura, T.; Sasaya, T. Strong resistance against Rice grassy stunt virus is induced in transgenic rice plants expressing double-stranded RNA of the viral genes for nucleocapsid or movement proteins as targets for RNA interference. Phytopathology 2013, 103, 513–519. [Google Scholar] [CrossRef] [PubMed]
- Liang, C.; Liu, H.; Hao, J.; Li, J.; Luo, L. Expression profiling and regulatory network of cucumber microRNAs and their putative target genes in response to cucumber green mottle mosaic virus infection. Archives of virology 2019, 164, 1121–1134. [Google Scholar] [CrossRef] [PubMed]
- Ai, T.; Zhang, L.; Gao, Z.; Zhu, C.X.; Guo, X. Highly efficient virus resistance mediated by artificial microRNAs that target the suppressor of PVX and PVY in plants. Plant Biology 2011, 13, 304–316. [Google Scholar] [CrossRef] [PubMed]
- Lafforgue, G.; Martínez, F.; Niu, Q.W.; Chua, N.H.; Daròs, J.A.; Elena, S.F. Improving the effectiveness of artificial microRNA (amiR)-mediated resistance against Turnip mosaic virus by combining two amiRs or by targeting highly conserved viral genomic regions. Journal of Virology 2013, 87, 8254–8256. [Google Scholar] [CrossRef] [PubMed]
- Gago-Zachert, S.; Schuck, J.; Weinholdt, C.; Knoblich, M.; Pantaleo, V.; Grosse, I.; Gursinsky, T.; Behrens, S.E. Highly efficacious antiviral protection of plants by small interfering RNAs identified in vitro. Nucleic acids research 2019, 47, 9343–9357. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Cheng, X.; Cai, J.; Zhan, L.; Wu, X.; Liu, Q.; Wu, X. Multiple virus resistance using artificial trans-acting siRNAs. Journal of virological methods 2016, 228, 16–20. [Google Scholar] [CrossRef]
- López-Dolz, L.; Spada, M.; Daròs, J.A.; Carbonell, A. Fine-tune control of targeted RNAi efficacy by plant artificial small RNAs. Nucleic Acids Research 2020, 48, 6234–6250. [Google Scholar] [CrossRef] [PubMed]
- Voinnet, O. Origin, biogenesis, and activity of plant microRNAs. Cell 2009, 136, 669–687. [Google Scholar] [CrossRef] [PubMed]
- Axtell, M.J. Classification and comparison of small RNAs from plants. Annual review of plant biology 2013, 64, 137–159. [Google Scholar] [CrossRef]
- Bartel, D.P. MicroRNAs: target recognition and regulatory functions. Cell 2009, 136, 215–233. [Google Scholar] [CrossRef] [PubMed]
- Bazzini, A.A.; Hopp, H.E.; Beachy, R.N.; Asurmendi, S. Infection and coaccumulation of tobacco mosaic virus proteins alter microRNA levels, correlating with symptom and plant development. Proceedings of the National Academy of Sciences 2007, 104, 12157–12162. [Google Scholar] [CrossRef] [PubMed]
- Tagami, Y.; Inaba, N.; Kutsuna, N.; Kurihara, Y.; Watanabe, Y. Specific enrichment of miRNAs in Arabidopsis thaliana infected with Tobacco mosaic virus. DNA research 2007, 14, 227–233. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.; Yang, Z.; Wang, Y.; Zheng, L.; Ye, R.; Ji, Y.; Zhao, S.; Ji, S.; Liu, R.; Xu, L.; Zheng, H. Viral-inducible Argonaute18 confers broad-spectrum virus resistance in rice by sequestering a host microRNA. Elife 2015, 4, e05733. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Jiao, X.; Kong, X.; Hamera, S.; Wu, Y.; Chen, X.; Fang, R.; Yan, Y. A signaling cascade from miR444 to RDR1 in rice antiviral RNA silencing pathway. Plant Physiology 2016, 170, 2365–2377. [Google Scholar] [CrossRef] [PubMed]
- Yuan, Q.; Zhai, Y.; Zhou, L.; Ai, X.; Chen, J.; Yan, F. A Novel miRNA in Rice Associated with the Low Seed Setting Rate Symptom of Rice Stripe Virus. International Journal of Molecular Sciences 2023, 24, 3675. [Google Scholar] [CrossRef]
- Niu, Q.W.; Lin, S.S.; Reyes, J.L.; Chen, K.C.; Wu, H.W.; Yeh, S.D.; Chua, N.H. Expression of artificial microRNAs in transgenic Arabidopsis thaliana confers virus resistance. Nature biotechnology 2006, 24, 1420–1428. [Google Scholar] [CrossRef] [PubMed]
- Vaucheret, H.; Vazquez, F.; Crété, P.; Bartel, D.P. The action of ARGONAUTE1 in the miRNA pathway and its regulation by the miRNA pathway are crucial for plant development. Genes & development 2004, 18, 1187–1197. [Google Scholar]
- Lafforgue, G.; Martínez, F.; Sardanyés, J.; De La Iglesia, F.; Niu, Q.W.; Lin, S.S.; Solé, R.V.; Chua, N.H.; Daròs, J.A.; Elena, S.F. Tempo and mode of plant RNA virus escape from RNA interference-mediated resistance. Journal of virology 2011, 85, 9686–9695. [Google Scholar] [CrossRef] [PubMed]
- Cisneros, A.E.; Carbonell, A. Artificial small RNA-based silencing tools for antiviral resistance in plants. Plants 2020, 9, 669. [Google Scholar] [CrossRef]
- Schwab, R.; Ossowski, S.; Riester, M.; Warthmann, N.; Weigel, D. Highly specific gene silencing by artificial microRNAs in Arabidopsis. The Plant Cell 2006, 18, 1121–1133. [Google Scholar] [CrossRef]
- Ramesh, S.V.; Ratnaparkhe, M.B.; Kumawat, G.; Gupta, G.K.; Husain, S.M. Plant miRNAome and antiviral resistance: a retrospective view and prospective challenges. Virus Genes 2014, 48, 1–14. [Google Scholar] [CrossRef]
- Reynolds, A.; Leake, D.; Boese, Q.; Scaringe, S.; Marshall, W.S.; Khvorova, A. Rational siRNA design for RNA interference. Nature biotechnology 2004, 22, 326–330. [Google Scholar] [CrossRef]
- Duan, C.G.; Wang, C.H.; Fang, R.X.; Guo, H.S. Artificial microRNAs highly accessible to targets confer efficient virus resistance in plants. Journal of virology 2008, 82, 11084–11095. [Google Scholar] [CrossRef]
- Van Vu, T.; Choudhury, N.R.; Mukherjee, S.K. Transgenic tomato plants expressing artificial microRNAs for silencing the pre-coat and coat proteins of a begomovirus, Tomato leaf curl New Delhi virus, show tolerance to virus infection. Virus Research 2013, 172, 35–45. [Google Scholar]
- Bartel, D.P. MicroRNAs: genomics, biogenesis, mechanism, and function. Cell 2004, 116, 281–297. [Google Scholar] [CrossRef]
- Bartel, B. MicroRNAs directing siRNA biogenesis. Nature structural & molecular biology 2005, 12, 569–571. [Google Scholar]
- Wang, M.B.; Masuta, C.; Smith, N.A.; and Shimura, H. RNA Silencing and Plant Viral Diseases. 1275 MPMI 2012, 25, 1275–1285. [Google Scholar] [CrossRef] [PubMed]
- Pumplin, N.; Voinnet, O. RNA silencing suppression by plant pathogens: defence, counter-defence and counter-counter-defence. Nature Reviews Microbiology 2013, 11, 745–760. [Google Scholar] [CrossRef]
- Revers, F.; Nicaise, V. Plant resistance to infection by viruses. eLS 2014. [Google Scholar]
- Khalid, A.; Zhang, Q.; Yasir, M.; Li, F. Small RNA based genetic engineering for plant viral resistance: application in crop protection. Frontiers in microbiology 2017, 8, 43. [Google Scholar] [CrossRef] [PubMed]
- Lu, Y.D.; Gan, Q.H.; Chi, X.Y.; Qin, S. Roles of microRNA in plant defense and virus offense interaction. Plant cell reports 2008, 27, 1571–1579. [Google Scholar] [CrossRef]
- Waterhouse, P.M.; Graham, M.W.; Wang, M.B. Virus resistance and gene silencing in plants can be induced by simultaneous expression of sense and antisense RNA. Proceedings of the National Academy of Sciences 1998, 95, 13959–13964. [Google Scholar] [CrossRef]
- Rosa, C.; Kuo, Y.W.; Wuriyanghan, H.; Falk, B.W. RNA interference mechanisms and applications in plant pathology. Annual review of phytopathology 2018, 56, 581–610. [Google Scholar] [CrossRef] [PubMed]
- Gaffar, F.Y.; Koch, A. Catch me if you can! RNA silencing-based improvement of antiviral plant immunity. Viruses 2019, 11, 673. [Google Scholar] [CrossRef] [PubMed]
- Voloudakis, A.E.; Kaldis, A.; Patil, B.L. RNA-based vaccination of plants for control of viruses. Annual Review of Virology 2022, 9, 521–548. [Google Scholar] [CrossRef] [PubMed]
- Tenllado, F.; Dıaz-Ruız, J.R. Double-stranded RNA-mediated interference with plant virus infection. Journal of virology 2001, 75, 12288–12297. [Google Scholar] [CrossRef] [PubMed]
- Fuentes, A.; Carlos, N.; Ruiz, Y.; Callard, D.; Sánchez, Y.; Ochagavía, M.E.; Seguin, J.; Malpica-López, N.; Hohn, T.; Lecca, M.R.; Pérez, R. Field trial and molecular characterization of RNAi-transgenic tomato plants that exhibit resistance to tomato yellow leaf curl geminivirus. Molecular Plant-Microbe Interactions 2016, 29, 197–209. [Google Scholar] [CrossRef]
- Walsh, H.A.; Vanderschuren, H.; Taylor, S.; Rey, M.E.C. RNA silencing of South African cassava mosaic virus in transgenic cassava expressing AC1/AC4 hp-RNA induces tolerance. Biotechnology Reports 2019, 24, e00383. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Tanti, B.; Patil, B.L.; Mukherjee, S.K.; Sahoo, L. RNAi-derived transgenic resistance to Mungbean yellow mosaic India virus in cowpea. PLoS One 2017, 12, e0186786. [Google Scholar] [CrossRef] [PubMed]
- Wu, L.; Mao, L.; Qi, Y. Roles of dicer-like and argonaute proteins in TAS-derived small interfering RNA-triggered DNA methylation. Plant physiology 2012, 160, 990–999. [Google Scholar] [CrossRef]
- Allen, E.; Xie, Z.; Gustafson, A.M.; Carrington, J.C. microRNA-directed phasing during trans-acting siRNA biogenesis in plants. Cell 2005, 121, 207–221. [Google Scholar] [CrossRef] [PubMed]
- Yoshikawa, M.; Peragine, A.; Park, M.Y.; Poethig, R.S. A pathway for the biogenesis of trans-acting siRNAs in Arabidopsis. Genes & development 2005, 19, 2164–2175. [Google Scholar]
- Vazquez, F.; Gasciolli, V.; Crété, P.; Vaucheret, H. The nuclear dsRNA binding protein HYL1 is required for microRNA accumulation and plant development, but not posttranscriptional transgene silencing. Current Biology 2004, 14, 346–351. [Google Scholar] [CrossRef]
- Gasciolli, V.; Mallory, A.C.; Bartel, D.P.; Vaucheret, H. Partially redundant functions of Arabidopsis DICER-like enzymes and a role for DCL4 in producing trans-acting siRNAs. Current Biology 2005, 15, 1494–1500. [Google Scholar] [CrossRef]
- Carbonell, A.; Takeda, A.; Fahlgren, N.; Johnson, S.C.; Cuperus, J.T.; Carrington, J.C. New generation of artificial MicroRNA and synthetic trans-acting small interfering RNA vectors for efficient gene silencing in Arabidopsis. Plant physiology 2014, 165, 15–29. [Google Scholar] [CrossRef] [PubMed]
- Carbonell, A. Secondary small interfering RNA-based silencing tools in plants: an update. Frontiers in Plant Science 2019, 10, 687. [Google Scholar] [CrossRef] [PubMed]
- Wierzbicki, A.T.; Blevins, T.; Swiezewski, S. Long noncoding RNAs in plants. Annual Review of Plant Biology 2021, 72, 245–271. [Google Scholar] [CrossRef] [PubMed]
- Rai, M.I.; Alam, M.; Lightfoot, D.A.; Gurha, P.; Afzal, A.J. Classification and experimental identification of plant long non-coding RNAs. Genomics 2019, 111, 997–1005. [Google Scholar] [CrossRef] [PubMed]
- Budak, H.; Kaya, S.B.; Cagirici, H.B. Long non-coding RNA in plants in the era of reference sequences. Frontiers in Plant Science 2020, 11, 276. [Google Scholar] [CrossRef] [PubMed]
- Taliansky, M.; Samarskaya, V.; Zavriev, S.K.; Fesenko, I.; Kalinina, N.O.; Love, A.J. RNA-based technologies for engineering plant virus resistance. Plants 2021, 10, 82. [Google Scholar] [CrossRef]
- Gelaw, T.A.; Sanan-Mishra, N. Non-coding RNAs in response to drought stress. International journal of molecular sciences 2021, 22, 12519. [Google Scholar] [CrossRef] [PubMed]
- Mattick, J.S. The state of long non-coding RNA biology. Non-coding RNA 2018, 4, 17. [Google Scholar] [CrossRef]
- Prasad, A.; Prasad, M. Host-virus interactions mediated by long non-coding RNAs. Virus Research 2021, 298, 198402. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Yu, W.; Yang, Y.; Li, X.; Chen, T.; Liu, T.; Ma, N.; Yang, X.; Liu, R.; Zhang, B. Genome-wide analysis of tomato long non-coding RNAs and identification as endogenous target mimic for microRNA in response to TYLCV infection. Scientific reports 2015, 5, 16946. [Google Scholar] [CrossRef] [PubMed]
- Kang, S.H.; Sun, Y.D.; Atallah, O.O.; Huguet-Tapia, J.C.; Noble, J.D.; Folimonova, S.Y. A long non-coding RNA of Citrus tristeza virus: Role in the virus interplay with the host immunity. Viruses 2019, 11, 436. [Google Scholar] [CrossRef] [PubMed]
- Shrestha, N.; Bujarski, J.J. Long noncoding RNAs in plant viroids and viruses: a review. Pathogens 2020, 9, 765. [Google Scholar] [CrossRef]
- Navarro, B.; Gisel, A.; Rodio, M.E.; Delgado, S.; Flores, R.; Di Serio, F. Viroids: how to infect a host and cause disease without encoding proteins. Biochimie 2012, 94, 1474–1480. [Google Scholar] [CrossRef] [PubMed]
- Dadami, E.; Boutla, A.; Vrettos, N.; Tzortzakaki, S.; Karakasilioti, I.; Kalantidis, K. DICER-LIKE 4 but not DICER-LIKE 2 may have a positive effect on potato spindle tuber viroid accumulation in Nicotiana benthamiana. Molecular plant 2013, 6, 232–234. [Google Scholar] [CrossRef]
- Karlik, E.; Ari, S.; Gozukirmizi, N. LncRNAs: genetic and epigenetic effects in plants. Biotechnology & Biotechnological Equipment 2019, 33, 429–439. [Google Scholar]
- Yang, Y.; Liu, T.; Shen, D.; Wang, J.; Ling, X.; Hu, Z.; Chen, T.; Hu, J.; Huang, J.; Yu, W.; Dou, D. Tomato yellow leaf curl virus intergenic siRNAs target a host long noncoding RNA to modulate disease symptoms. PLoS pathogens 2019, 15, e1007534. [Google Scholar] [CrossRef] [PubMed]
- Bak, R.O.; Gomez-Ospina, N.; Porteus, M.H. Gene editing on center stage. Trends in Genetics 2018, 34, 600–611. [Google Scholar] [CrossRef]
- Saurabh, S. Genome editing: revolutionizing the crop improvement. Plant Molecular Biology Reporter 2021, 39, 752–772. [Google Scholar] [CrossRef]
- Mushtaq, M.; Mukhtar, S.; Sakina, A.; Dar, A.A.; Bhat, R.; Deshmukh, R.; Molla, K.; Kundoo, A.A.; Dar, M.S. Tweaking genome-editing approaches for virus interference in crop plants. Plant Physiology and Biochemistry 2020, 147, 242–250. [Google Scholar] [CrossRef] [PubMed]
- Jurica, M.S.; Stoddard, B.L. Homing endonucleases: structure, function and evolution. Cellular and Molecular Life Sciences CMLS 1999, 55, 1304–1326. [Google Scholar] [CrossRef] [PubMed]
- Mishra, R.; Joshi, R.K.; Zhao, K. Genome editing in rice: recent advances, challenges, and future implications. Frontiers in Plant Science 2018, 9, 1361. [Google Scholar] [CrossRef] [PubMed]
- Townsend, J.A.; Wright, D.A.; Winfrey, R.J.; Fu, F.; Maeder, M.L.; Joung, J.K.; Voytas, D.F. High-frequency modification of plant genes using engineered zinc-finger nucleases. Nature 2009, 459, 442–445. [Google Scholar] [CrossRef] [PubMed]
- Puchta, H. The repair of double-strand breaks in plants: mechanisms and consequences for genome evolution. Journal of experimental botany 2005, 56, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Osakabe, K.; Osakabe, Y.; Toki, S. Site-directed mutagenesis in Arabidopsis using custom-designed zinc finger nucleases. Proceedings of the National Academy of Sciences 2010, 107, 12034–12039. [Google Scholar] [CrossRef]
- Zhang, F.; Maeder, M.L.; Unger-Wallace, E.; Hoshaw, J.P.; Reyon, D.; Christian, M.; Li, X.; Pierick, C.J.; Dobbs, D.; Peterson, T.; Joung, J.K. High frequency targeted mutagenesis in Arabidopsis thaliana using zinc finger nucleases. Proceedings of the National Academy of Sciences 2010, 107, 12028–12033. [Google Scholar] [CrossRef] [PubMed]
- Carroll, D. Genome engineering with zinc-finger nucleases. Genetics 2011, 188, 773–782. [Google Scholar] [CrossRef] [PubMed]
- Curtin, S.J.; Zhang, F.; Sander, J.D.; Haun, W.J.; Starker, C.; Baltes, N.J.; Reyon, D.; Dahlborg, E.J.; Goodwin, M.J.; Coffman, A.P.; Dobbs, D. Targeted mutagenesis of duplicated genes in soybean with zinc-finger nucleases. Plant physiology 2011, 156, 466–473. [Google Scholar] [CrossRef] [PubMed]
- Gaj, T.; Gersbach, C.A.; Barbas, C.F. ZFN, TALEN, and CRISPR/Cas-based methods for genome engineering. Trends in biotechnology 2013, 31, 397–405. [Google Scholar] [CrossRef] [PubMed]
- Curtin, S.J.; Voytas, D.F.; Stupar, R.M. Genome engineering of crops with designer nucleases. The Plant Genome 2012, 5. [Google Scholar] [CrossRef]
- Qi, L.S.; Larson, M.H.; Gilbert, L.A.; Doudna, J.A.; Weissman, J.S.; Arkin, A.P.; Lim, W.A. Repurposing CRISPR as an RNA-guided platform for sequence-specific control of gene expression. Cell 2013, 152, 1173–1183. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.; Ren, S.; Yu, S.; Pan, H.; Li, T.; Ge, S.; Zhang, J.; Xia, N. Methods favoring homology-directed repair choice in response to CRISPR/Cas9 induced-double strand breaks. International Journal of Molecular Sciences 2020, 21, 6461. [Google Scholar] [CrossRef]
- Chen, W.; Qian, Y.; Wu, X.; Sun, Y.; Wu, X.; Cheng, X. Inhibiting replication of begomoviruses using artificial zinc finger nucleases that target viral-conserved nucleotide motif. Virus Genes 2014, 48, 494–501. [Google Scholar] [CrossRef]
- Sera, T. ; Inhibition of virus DNA replication by artificial zinc finger proteins. Journal of virology 2005, 79, 2614–2619. [Google Scholar] [CrossRef] [PubMed]
- Khan, Z.; Khan, S.H.; Ahmed, A.; Iqbal, M.U.; Mubarik, M.S.; Ghouri, M.Z.; Ahmad, F.; Yaseen, S.; Ali, Z.; Khan, A.A.; Azhar, M.T. Genome editing in cotton: challenges and opportunities. Journal of Cotton Research 2023, 6, 1–21. [Google Scholar] [CrossRef]
- Hanley-Bowdoin, L.; Bejarano, E.R.; Robertson, D.; Mansoor, S. Geminiviruses: masters at redirecting and reprogramming plant processes. Nature Reviews Microbiology 2013, 11, 777–788. [Google Scholar] [CrossRef]
- Lloyd, A.; Plaisier, C.L.; Carroll, D.; Drews, G.N. Targeted mutagenesis using zinc-finger nucleases in Arabidopsis. Proceedings of the National Academy of Sciences 2005, 102, 2232–2237. [Google Scholar] [CrossRef]
- Shukla, V.K.; Doyon, Y.; Miller, J.C.; DeKelver, R.C.; Moehle, E.A.; Worden, S.E.; Mitchell, J.C.; Arnold, N.L.; Gopalan, S.; Meng, X.; Choi, V.M. Precise genome modification in the crop species Zea mays using zinc-finger nucleases. Nature 2009, 459, 437–441. [Google Scholar] [CrossRef] [PubMed]
- Joung, J.K.; Sander, J.D. TALENs: a widely applicable technology for targeted genome editing. Nature reviews Molecular cell biology 2013, 14, 49–55. [Google Scholar] [CrossRef] [PubMed]
- Boch, J.; Bonas, U. Xanthomonas AvrBs3 family-type III effectors: discovery and function. Annual review of phytopathology 2010, 48, 419–436. [Google Scholar] [CrossRef]
- Kay, S.; Bonas, U. How Xanthomonas type III effectors manipulate the host plant. Current opinion in microbiology 2009, 12, 37–43. [Google Scholar] [CrossRef]
- Miller, J.C.; Tan, S.; Qiao, G.; Barlow, K.A.; Wang, J.; Xia, D.F.; Meng, X.; Paschon, D.E.; Leung, E.; Hinkley, S.J.; Dulay, G.P. A TALE nuclease architecture for efficient genome editing. Nature biotechnology 2011, 29, 143–148. [Google Scholar] [CrossRef] [PubMed]
- Yang, B.; Sugio, A.; White, F.F. Os8N3 is a host disease-susceptibility gene for bacterial blight of rice. Proceedings of the National Academy of Sciences 2006, 103, 10503–10508. [Google Scholar] [CrossRef] [PubMed]
- Cheng, X.; Li, F.; Cai, J.; Chen, W.; Zhao, N.; Sun, Y.; Guo, Y.; Yang, X.; Wu, X. Artificial TALE as a convenient protein platform for engineering broad-spectrum resistance to begomoviruses. Viruses 2015, 7, 4772–4782. [Google Scholar] [CrossRef] [PubMed]
- Ishino, Y.; Shinagawa, H.; Makino, K.; Amemura, M.; Nakata, A. Nucleotide sequence of the iap gene, responsible for alkaline phosphatase isozyme conversion in Escherichia coli, and identification of the gene product. Journal of bacteriology 1987, 169, 5429–5433. [Google Scholar] [CrossRef]
- Jansen, R.; Embden, J.D.V.; Gaastra, W.; Schouls, L.M. Identification of genes that are associated with DNA repeats in prokaryotes. Molecular microbiology 2002, 43, 1565–1575. [Google Scholar] [CrossRef]
- Wright, A.V.; Nuñez, J.K.; Doudna, J.A. Biology and applications of CRISPR systems: harnessing nature’s toolbox for genome engineering. Cell 2016, 164, 29–44. [Google Scholar] [CrossRef] [PubMed]
- Jinek, M.; Chylinski, K.; Fonfara, I.; Hauer, M.; Doudna, J.A.; Charpentier, E. A programmable dual-RNA–guided DNA endonuclease in adaptive bacterial immunity. Science 2012, 337, 816–821. [Google Scholar] [CrossRef] [PubMed]
- Lander, E.S. The heroes of CRISPR. Cell 2016, 164, 18–28. [Google Scholar] [CrossRef] [PubMed]
- Jiang, F.; Doudna, J.A. CRISPR–Cas9 structures and mechanisms. Annual review of biophysics 2017, 46, 505–529. [Google Scholar] [CrossRef]
- Konermann, S.; Shehata, S.; Dohmae, N.; Ishitani, R.; Zhang, F.; Nureki, O.; Nishimasu, H.; Ran, F.; Hsu, P. Crystal structure of Cas9 in complex with guide RNA and target DNA. Cell 2014, 156. [Google Scholar]
- Anders, C.; Niewoehner, O.; Duerst, A.; Jinek, M. Structural basis of PAM-dependent target DNA recognition by the Cas9 endonuclease. Nature 2014, 513, 569–573. [Google Scholar] [CrossRef]
- Mali, P.; Esvelt, K.M.; Church, G.M. Cas9 as a versatile tool for engineering biology. Nature methods 2013, 10, 957–963. [Google Scholar] [CrossRef] [PubMed]
- Sternberg, S.H.; Redding, S.; Jinek, M.; Greene, E.C.; Doudna, J.A. DNA interrogation by the CRISPR RNA-guided endonuclease Cas9. Biophysical Journal 2014, 106, 695a. [Google Scholar] [CrossRef]
- Abudayyeh, O.O.; Gootenberg, J.S.; Konermann, S.; Joung, J.; Slaymaker, I.M.; Cox, D.B.; Shmakov, S.; Makarova, K.S.; Semenova, E.; Minakhin, L.; Severinov, K. C2c2 is a single-component programmable RNA-guided RNA-targeting CRISPR effector. Science 2016, 353, aaf5573. [Google Scholar] [CrossRef] [PubMed]
- Aman, R.; Ali, Z.; Butt, H.; Mahas, A.; Aljedaani, F.; Khan, M.Z.; Ding, S.; Mahfouz, M. RNA virus interference via CRISPR/Cas13a system in plants. Genome biology 2018, 19, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Ali, Z.; Ali, S.; Tashkandi, M.; Zaidi, S.S.E.A.; Mahfouz, M.M. CRISPR/Cas9-mediated immunity to geminiviruses: differential interference and evasion. Scientific reports 2016, 6, 26912. [Google Scholar] [CrossRef]
- Ali, Z.; Abul-Faraj, A.; Li, L.; Ghosh, N.; Piatek, M.; Mahjoub, A.; Aouida, M.; Piatek, A.; Baltes, N.J.; Voytas, D.F.; Dinesh-Kumar, S. Efficient virus-mediated genome editing in plants using the CRISPR/Cas9 system. Molecular plant 2015, 8, 1288–1291. [Google Scholar] [CrossRef] [PubMed]
- Khan, Z.A.; Kumar, R.; Dasgupta, I. CRISPR/Cas-mediated resistance against viruses in plants. International Journal of Molecular Sciences 2022, 23, 2303. [Google Scholar] [CrossRef] [PubMed]
- Makarova, K.S.; Wolf, Y.I.; Alkhnbashi, O.S.; Costa, F.; Shah, S.A.; Saunders, S.J.; Barrangou, R.; Brouns, S.J.; Charpentier, E.; Haft, D.H.; Horvath, P. An updated evolutionary classification of CRISPR–Cas systems. Nature Reviews Microbiology 2015, 13, 722–736. [Google Scholar] [CrossRef]
- Zetsche, B.; Gootenberg, J.S.; Abudayyeh, O.O.; Slaymaker, I.M.; Makarova, K.S.; Essletzbichler, P.; Volz, S.E.; Joung, J.; Van Der Oost, J.; Regev, A.; Koonin, E.V. Cpf1 is a single RNA-guided endonuclease of a class 2 CRISPR-Cas system. Cell 2015, 163, 759–771. [Google Scholar] [CrossRef] [PubMed]
- Fonfara, I.; Richter, H.; Bratovič, M.; Le Rhun, A.; Charpentier, E. The CRISPR-associated DNA-cleaving enzyme Cpf1 also processes precursor CRISPR RNA. Nature 2016, 532, 517–521. [Google Scholar] [CrossRef] [PubMed]
- Selma, S.; Orzáez, D. Perspectives for epigenetic editing in crops. Transgenic Research 2021, 30, 381–400. [Google Scholar] [CrossRef] [PubMed]
- Uranga, M.; Daròs, J.A. Tools and targets: The dual role of plant viruses in CRISPR–Cas genome editing. The Plant Genome 2023, 16, e20220. [Google Scholar] [CrossRef] [PubMed]
- Dubois, A.; Roudier, F. Deciphering plant chromatin regulation via CRISPR/dCas9-based epigenome engineering. Epigenomes 2021, 5, 17. [Google Scholar] [CrossRef]
- Price, A.A.; Sampson, T.R.; Ratner, H.K.; Grakoui, A.; Weiss, D.S. Cas9-mediated targeting of viral RNA in eukaryotic cells. Proceedings of the National Academy of Sciences 2015, 112, 6164–6169. [Google Scholar] [CrossRef]
- Shmakov, S.; Abudayyeh, O.O.; Makarova, K.S.; Wolf, Y.I.; Gootenberg, J.S.; Semenova, E.; Minakhin, L.; Joung, J.; Konermann, S.; Severinov, K.; Zhang, F. Discovery and functional characterization of diverse class 2 CRISPR-Cas systems. Molecular cell 2015, 60, 385–397. [Google Scholar] [CrossRef]
- Zhang, T.; Zheng, Q.; Yi, X.; An, H.; Zhao, Y.; Ma, S.; Zhou, G. Establishing RNA virus resistance in plants by harnessing CRISPR immune system. Plant biotechnology journal 2018, 16, 1415–1423. [Google Scholar] [CrossRef]
- Marqués, M.C.; Sánchez-Vicente, J.; Ruiz, R.; Montagud-Martínez, R.; Márquez-Costa, R.; Gómez, G.; Carbonell, A.; Daròs, J.A.; Rodrigo, G. Diagnostics of infections produced by the plant viruses TMV, TEV, and PVX with CRISPR-Cas12 and CRISPR-Cas13. ACS Synthetic Biology 2022, 11, 2384–2393. [Google Scholar] [CrossRef]
- Shmakov, S.; Smargon, A.; Scott, D.; Cox, D.; Pyzocha, N.; Yan, W.; Abudayyeh, O.O.; Gootenberg, J.S.; Makarova, K.S.; Wolf, Y.I.; Severinov, K. Diversity and evolution of class 2 CRISPR–Cas systems. Nature reviews microbiology 2017, 15, 169–182. [Google Scholar] [CrossRef] [PubMed]
- Kavuri, N.R.; Ramasamy, M.; Qi, Y.; Mandadi, K. Applications of CRISPR/Cas13-based RNA editing in plants. Cells 2022, 11, 2665. [Google Scholar] [CrossRef] [PubMed]
- Zhang, T.; Zhao, Y.; Ye, J.; Cao, X.; Xu, C.; Chen, B.; An, H.; Jiao, Y.; Zhang, F.; Yang, X.; Zhou, G. Establishing CRISPR/Cas13a immune system conferring RNA virus resistance in both dicot and monocot plants. Plant biotechnology journal 2019, 17, 1185. [Google Scholar] [CrossRef] [PubMed]
- Gupta, R.; Ghosh, A.; Chakravarti, R.; Singh, R.; Ravichandiran, V.; Swarnakar, S.; Ghosh, D. Cas13d: a new molecular scissor for transcriptome engineering. Frontiers in Cell and Developmental Biology 2022, 10, 866800. [Google Scholar] [CrossRef]
- Komor, A.C.; Kim, Y.B.; Packer, M.S.; Zuris, J.A.; Liu, D.R. Programmable editing of a target base in genomic DNA without double-stranded DNA cleavage. Nature 2016, 533, 420–424. [Google Scholar] [CrossRef]
- Molla, K.A.; Yang, Y. CRISPR/Cas-mediated base editing: technical considerations and practical applications. Trends in biotechnology 2019, 37, 1121–1142. [Google Scholar] [CrossRef] [PubMed]
- Bastet, A.; Zafirov, D.; Giovinazzo, N.; Guyon-Debast, A.; Nogué, F.; Robaglia, C.; Gallois, J.L. Mimicking natural polymorphism in eIF 4E by CRISPR-Cas9 base editing is associated with resistance to potyviruses. Plant Biotechnology Journal 2019, 17, 1736–1750. [Google Scholar] [CrossRef] [PubMed]
- Gaudelli, N.M.; Komor, A.C.; Rees, H.A.; Packer, M.S.; Badran, A.H.; Bryson, D.I.; Liu, D.R. Programmable base editing of A• T to G• C in genomic DNA without DNA cleavage. Nature 2017, 551, 464–471. [Google Scholar] [CrossRef]
- Li, C.; Zhang, R.; Meng, X.; Chen, S.; Zong, Y.; Lu, C.; Qiu, J.L.; Chen, Y.H.; Li, J.; Gao, C. Targeted, random mutagenesis of plant genes with dual cytosine and adenine base editors. Nature biotechnology 2020, 38, 875–882. [Google Scholar] [CrossRef] [PubMed]
- Anzalone, A.V.; Koblan, L.W.; Liu, D.R. Genome editing with CRISPR–Cas nucleases, base editors, transposases and prime editors. Nature biotechnology 2020, 38, 824–844. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Z.; Hong, X.; Zhang, S.; Yao, R.; Xiao, Y.I. CRISPR base editing and prime editing: DSB and template-free editing systems for bacteria and plants. Synthetic and Systems Biotechnology 2020, 5, 277–292. [Google Scholar]
- Lin, Q.; Zong, Y.; Xue, C.; Wang, S.; Jin, S.; Zhu, Z.; Wang, Y.; Anzalone, A.V.; Raguram, A.; Doman, J.L.; Liu, D.R. Prime genome editing in rice and wheat. Nature biotechnology 2020, 38, 582–585. [Google Scholar] [CrossRef]
- Binyameen, B.; Khan, Z.; Khan, S.H.; Ahmad, A.; Munawar, N.; Mubarik, M.S.; Riaz, H.; Ali, Z.; Khan, A.A.; Qusmani, A.T.; Abd-Elsalam, K.A. Using multiplexed CRISPR/Cas9 for suppression of cotton leaf curl virus. International journal of molecular sciences 2021, 22, 12543. [Google Scholar] [CrossRef]
- Cao, J.; Xiao, Q.; Yan, Q. The multiplexed CRISPR targeting platforms. Drug Discovery Today: Technologies 2018, 28, 53–61. [Google Scholar] [CrossRef] [PubMed]
- Zhan, X.; Liu, W.; Nie, B.; Zhang, F.; Zhang, J. Cas13d-mediated multiplex RNA targeting confers a broad-spectrum resistance against RNA viruses in potato. Communications Biology 2023, 6, 855. [Google Scholar] [CrossRef] [PubMed]
- Eid, A.; Alshareef, S.; Mahfouz, M.M. CRISPR base editors: genome editing without double-stranded breaks. Biochemical Journal 2018, 475, 1955–1964. [Google Scholar] [CrossRef]
- Mubarik, M.S.; Wang, X.; Khan, S.H.; Ahmad, A.; Khan, Z.; Amjid, M.W.; Razzaq, M.K.; Ali, Z.; Azhar, M.T. Engineering broad-spectrum resistance to cotton leaf curl disease by CRISPR-Cas9 based multiplex editing in plants. GM Crops & Food 2021, 12, 647–658. [Google Scholar]
- Ji, X.; Si, X.; Zhang, Y.; Zhang, H.; Zhang, F.; Gao, C. Conferring DNA virus resistance with high specificity in plants using virus-inducible genome-editing system. Genome biology 2018, 19, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Baltes, N.J.; Gil-Humanes, J.; Cermak, T.; Atkins, P.A.; Voytas, D.F. DNA replicons for plant genome engineering. The Plant Cell 2014, 26, 151–163. [Google Scholar] [CrossRef] [PubMed]
- Takenaka, K.; Koshino-Kimura, Y.; Aoyama, Y.; Sera, T. Inhibition of tomato yellow leaf curl virus replication by artificial zinc-finger proteins. In Nucleic Acids Symposium Series, Oxford University Press 2007, Vol. 51, No. 1, pp. 429-430.
- Koshino-Kimura, Y.; Takenaka, K.; Domoto, F.; Ohashi, M.; Miyazaki, T.; Aoyama, Y.; Sera, T. Construction of plants resistant to TYLCV by using artificial zinc-finger proteins. In Nucleic Acids Symposium Series, Oxford University Press 2009, Vol. 53, No. 1, pp. 281-282. Oxford University Press.
- Mori, T.; Takenaka, K.; Domoto, F.; Aoyama, Y.; Sera, T. Inhibition of binding of tomato yellow leaf curl virus rep to its replication origin by artificial zinc-finger protein. Molecular biotechnology 2013, 54, 198–203. [Google Scholar] [CrossRef] [PubMed]
- Romay, G.; Bragard, C. Antiviral defenses in plants through genome editing. Frontiers in microbiology 2017, 8, 47. [Google Scholar] [CrossRef] [PubMed]
- Ordiz, M.I.; Magnenat, L.; Barbas, C.F.; Beachy, R.N. Negative regulation of the RTBV promoter by designed zinc finger proteins. Plant molecular biology 2010, 72, 621–630. [Google Scholar] [CrossRef] [PubMed]
- Ji, X.; Zhang, H.; Zhang, Y.; Wang, Y.; Gao, C. Establishing a CRISPR–Cas-like immune system conferring DNA virus resistance in plants. Nature Plants 2015, 1, 1–4. [Google Scholar] [CrossRef] [PubMed]
- Baltes, N.J.; Hummel, A.W.; Konecna, E.; Cegan, R.; Bruns, A.N.; Bisaro, D.M.; Voytas, D.F. Conferring resistance to geminiviruses with the CRISPR–Cas prokaryotic immune system. Nature Plants 2015, 1, 1–4. [Google Scholar] [CrossRef] [PubMed]
- Roy, A.; Zhai, Y.; Ortiz, J.; Neff, M.; Mandal, B.; Mukherjee, S.K.; Pappu, H.R. Multiplexed editing of a begomovirus genome restricts escape mutant formation and disease development. PloS one 2019, 14, e0223765. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Soyars, C.L.; Li, J.; Fei, Q.; He, G.; Peterson, B.A.; Meyers, B.C.; Nimchuk, Z.L.; Wang, X. CRISPR/Cas9-mediated resistance to cauliflower mosaic virus. Plant direct 2018, 2, e00047. [Google Scholar] [CrossRef]
- Kis, A.; Hamar, É.; Tholt, G.; Bán, R.; Havelda, Z. Creating highly efficient resistance against wheat dwarf virus in barley by employing CRISPR/Cas9 system. Plant biotechnology journal 2019, 17, 1004. [Google Scholar] [CrossRef]
- Tripathi, J.N.; Ntui, V.O.; Ron, M.; Muiruri, S.K.; Britt, A.; Tripathi, L. CRISPR/Cas9 editing of endogenous banana streak virus in the B genome of Musa spp. overcomes a major challenge in banana breeding. Communications Biology 2019, 2, 46. [Google Scholar] [CrossRef] [PubMed]
- Mehta, D.; Stürchler, A.; Anjanappa, R.B.; Zaidi, S.S.E.A.; Hirsch-Hoffmann, M.; Gruissem, W.; Vanderschuren, H. Linking CRISPR-Cas9 interference in cassava to the evolution of editing-resistant geminiviruses. Genome biology 2019, 20, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Honig, A.; Marton, I.; Rosenthal, M.; Smith, J.J.; Nicholson, M.G.; Jantz, D.; Zuker, A.; Vainstein, A. Transient expression of virally delivered meganuclease in planta generates inherited genomic deletions. Molecular plant 2015, 8, 1292–1294. [Google Scholar] [CrossRef] [PubMed]
- Gao, Q.; Xu, W.Y.; Yan, T.; Fang, X.D.; Cao, Q.; Zhang, Z.J.; Ding, Z.H.; Wang, Y.; Wang, X.B. Rescue of a plant cytorhabdovirus as versatile expression platforms for planthopper and cereal genomic studies. New Phytologist 2019, 223, 2120–2133. [Google Scholar] [CrossRef] [PubMed]
- Ma, X.; Zhang, X.; Liu, H.; Li, Z. Highly efficient DNA-free plant genome editing using virally delivered CRISPR–Cas9. Nature Plants 2020, 6, 773–779. [Google Scholar] [CrossRef] [PubMed]
- Noureen, A.; Zuhaib Khan, M.; Amin, I.; Zainab, T.; Ahmad, N.; Haider, S.; Mansoor, S. Broad-spectrum resistance against multiple PVY-strains by CRSIPR/Cas13 system in Solanum tuberosum crop. GM Crops & Food 2022, 13, 97–111. [Google Scholar]
- Ashraf, S.; Ahmad, A.; Khan, S.H.; Jamil, A.; Sadia, B.; Brown, J.K. LbCas12a mediated suppression of Cotton leaf curl Multan virus. Frontiers in Plant Science 2023, 14. [Google Scholar] [CrossRef] [PubMed]
- Uranga, M.; Vazquez-Vilar, M.; Orzáez, D.; Daròs, J.A. CRISPR-Cas12a genome editing at the whole-plant level using two compatible RNA virus vectors. The CRISPR Journal 2021, 4, 761–769. [Google Scholar] [CrossRef] [PubMed]
- Haider, S.; Faiq, A.; Khan, M.Z.; Mansoor, S.; Amin, I. Fully Transient CRISPR/Cas12f system in plants capable of broad-spectrum resistance against Begomovirus. bioRxiv 2022, 2022–06. [Google Scholar]
- Senthil-Kumar, M.; Mysore, K.S. New dimensions for VIGS in plant functional genomics. Trends in plant science 2011, 16, 656–665. [Google Scholar] [CrossRef] [PubMed]
- Dalakouras, A.; Ganopoulos, I. Induction of promoter DNA methylation upon high-pressure spraying of double-stranded RNA in plants. Agronomy 2021, 11, 789. [Google Scholar] [CrossRef]
- Mitter, N.; Worrall, E.A.; Robinson, K.E.; Li, P.; Jain, R.G.; Taochy, C.; Fletcher, S.J.; Carroll, B.J.; Lu, G.Q.; Xu, Z.P. Clay nanosheets for topical delivery of RNAi for sustained protection against plant viruses. Nature plants 2017, 3, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Liu, Q.; Li, Y.; Xu, K.; Li, D.; Hu, H.; Zhou, F.; Song, P.; Yu, Y.; Wei, Q.; Liu, Q.; Wang, W. Clay nanosheet-mediated delivery of recombinant plasmids expressing artificial miRNAs via leaf spray to prevent infection by plant DNA viruses. Horticulture Research 2020, 7. [Google Scholar] [CrossRef] [PubMed]
- Delgado-Martín, J.; Ruiz, L.; Janssen, D.; Velasco, L. Exogenous application of dsRNA for the control of viruses in cucurbits. Frontiers in Plant Science 2022, 13, 895953. [Google Scholar] [CrossRef] [PubMed]
- Rego-Machado, C.M.; Nakasu, E.Y.; Silva, J.M.; Lucinda, N.; Nagata, T.; Inoue-Nagata, A.K. siRNA biogenesis and advances in topically applied dsRNA for controlling virus infections in tomato plants. Scientific Reports 2020, 10, 22277. [Google Scholar] [CrossRef] [PubMed]
- Lau, S.E.; Mazumdar, P.; Hee, T.W.; Song, A.L.A.; Othman, R.Y.; Harikrishna, J.A. Crude extracts of bacterially-expressed dsRNA protect orchid plants against Cymbidium mosaic virus during transplantation from in vitro culture. The Journal of Horticultural Science and Biotechnology 2014, 89, 569–576. [Google Scholar] [CrossRef]
- Holeva, M.C.; Sklavounos, A.; Rajeswaran, R.; Pooggin, M.M.; Voloudakis, A.E. Topical application of double-stranded RNA targeting 2b and CP genes of Cucumber mosaic virus protects plants against local and systemic viral infection. Plants 2021, 10, 963. [Google Scholar] [CrossRef]
- Xu, X.; Yu, T.; Zhang, D.; Song, H.; Huang, K.; Wang, Y.; Shen, L.; Li, Y.; Wang, F.; Zhang, S.; Jiao, Y. Evaluation of the anti-viral efficacy of three different dsRNA nanoparticles against potato virus Y using various delivery methods. Ecotoxicology and Environmental Safety 2023, 255, 114775. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Schematic diagram to represent the RNAi pathway and the targets in genome editing strategies that ehance plant resistance to virus infection. (A) Infection of virus triggers Post-transcriptional Gene Silencing (PTGS) in plant cell. Long non-coding RNAs (incRNAs) like SILNR1 and viral encoded suppressor of RNA silencing (VSR) interfere with multiple steps in the silencing pathways. (B) Infection of DNA virus can also triggers Transcriptional Gene Silencing (TGS) in plant cells. Components of methylation machinery like Methyltransferase 1 (MET1), Domain rearranged methyltransferase 1/2 (DRM1/2), Chromomethylase 2/3 (CMT2/3) and Kryptonite 2 (KYP2) are involved in small RNA directed methylation. (C) The RNAi pathway can br artificially triggered by expression of hairpin RNA (hpRNA), artificial miRNA (amiRNA), synthetic trans acting siRNA (syn-tasiRNAs) and satellite or DI virus RNAs directed against the viral genome. (D) The GE strategies use nucleases such as FnCas9 and Cas13a,d to target the viral RNAs mainly the VSRs.
Figure 1.
Schematic diagram to represent the RNAi pathway and the targets in genome editing strategies that ehance plant resistance to virus infection. (A) Infection of virus triggers Post-transcriptional Gene Silencing (PTGS) in plant cell. Long non-coding RNAs (incRNAs) like SILNR1 and viral encoded suppressor of RNA silencing (VSR) interfere with multiple steps in the silencing pathways. (B) Infection of DNA virus can also triggers Transcriptional Gene Silencing (TGS) in plant cells. Components of methylation machinery like Methyltransferase 1 (MET1), Domain rearranged methyltransferase 1/2 (DRM1/2), Chromomethylase 2/3 (CMT2/3) and Kryptonite 2 (KYP2) are involved in small RNA directed methylation. (C) The RNAi pathway can br artificially triggered by expression of hairpin RNA (hpRNA), artificial miRNA (amiRNA), synthetic trans acting siRNA (syn-tasiRNAs) and satellite or DI virus RNAs directed against the viral genome. (D) The GE strategies use nucleases such as FnCas9 and Cas13a,d to target the viral RNAs mainly the VSRs.
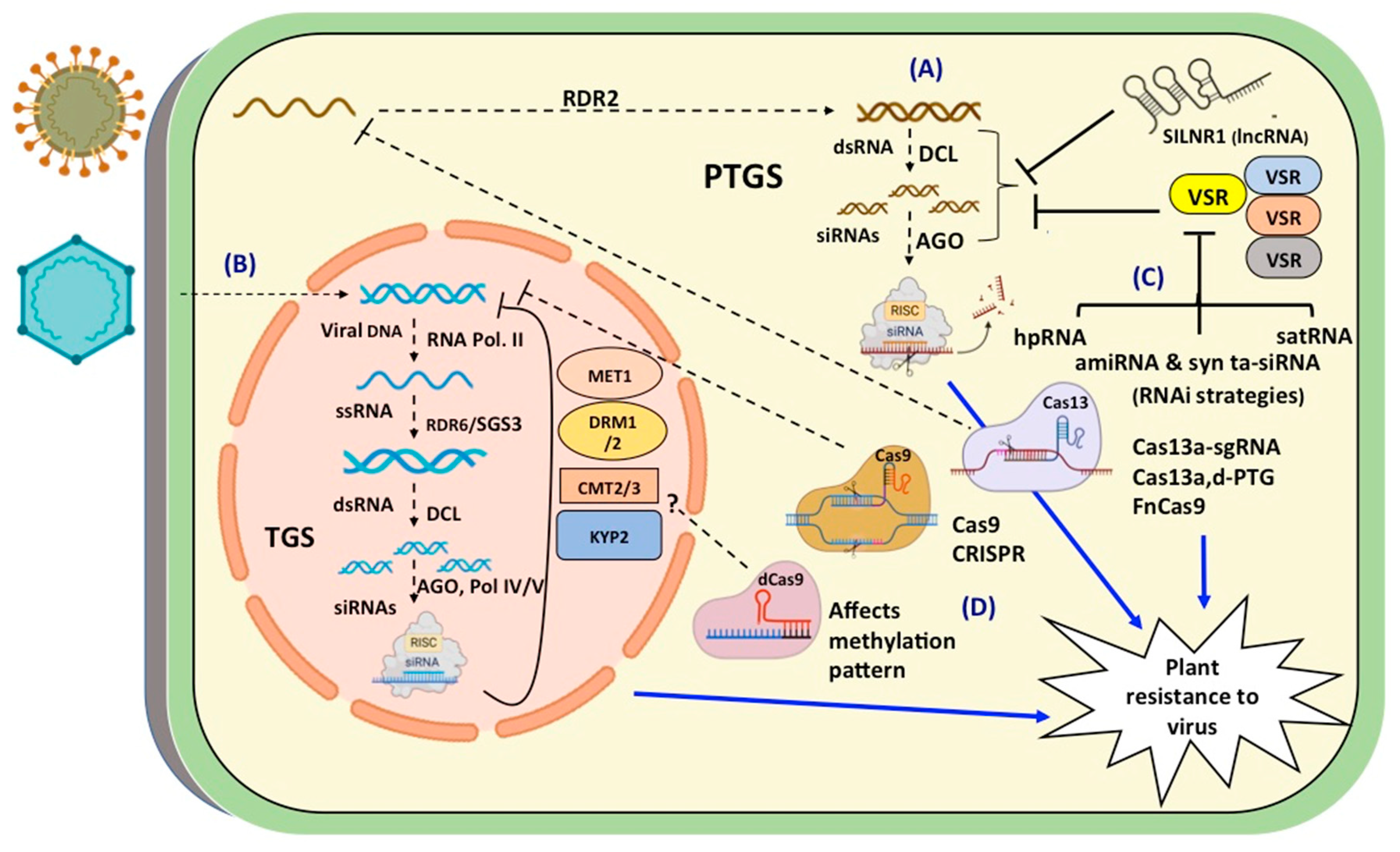
Table 1.
List of small RNA based GM approaches used against plant viruses.
| Strategy | Virus | Target | Plant | References |
|---|---|---|---|---|
|
hpRNA (IR) hpRNA |
TMV CMV PPV CGMMV PSTVd TYLCV PVX PVY PLRV PRSV SbDV SMV WCMV ACMV BBTV CTV |
Movement protein Replicase P1 and HC-Pro Coat protein Viroid sequence Replicase (C1) Coat protein ORF2 HC-Pro CP CP CP HC-Pro (VSR) Replicase Rep (AC1) Replicase P20, p23, p25 (VSRs) |
Tobacco Tomato Potato Melon Soyabean White clover Cassava Banana Mexican lime |
[91] [92] [93] [94] [95] [96] [97] [98] [99] [100] [101] [102] [103] [104] |
| DI Virus-derived RNA | PVY TBSV GLFV |
Coat protein (DI virus) Movement protein (DI virus) |
Tobacco | [105] |
| dsRNA | ACMV RGSV RTBV |
DNA-A and DNA-B pC5, pC6 ORF IV |
Tobacco Rice |
[106] [107] [53] |
|
amiRNA (precursor miR395) amiRNA (precursor miR319) amiRNA/single monocistronic AthMIR156 AthMIR159a AthMIR167b AthMIR169a amiRNA/single polycistronic OsaMIR395 amiRNA/single monocistronic in tandem repeats AthMIR159a amiRNA/multiple monocistronic in trans AthMIR159a AthMIR390a |
WSMV GFLV CGMMV WSMV PVX, PVY TuMV, TYMV RBSDV, RSV TuMV TBSV |
Virus genome (conserved region) CP CP 5’UTR+P1+HC-Pro+ P3 P25 (PVX) + HC-Pro (PVY) HC- Pro (TuMV) + P69 (TYMV) CP + HC-Pro 5’ terminal TBSV (+) RNA |
Wheat Grape Tobacco Wheat N. tabacum Arabidopsis Tobacco |
[89] [103] [108] [89] [109] [110] [111] |
| ta-siRNAs AthTAS3a | TuMV, CMV | Multiple genomic positions | Arabidopsis |
[112] |
|
syn-tasiRNA/ single polycistronic AthTAS1c |
TSWV |
RdRP |
Tobacco |
[113] |
Table 3.
List of exogenously delivered dsRNAs for induction of RNA silencing against plant viruses.
| dsRNA coating and inoculation | Plant | Virus | References |
|---|---|---|---|
| LDH nanosheets | Cowpea Tomato |
CMV TYLCV |
[250] [260] |
| Agroinfiltration and Direct spray | Cucumber | CGMMV ToLCNDV |
[261] |
| High pressure spraying of free siRNAs | Tobacco | CaMV 35S promoter | [258] |
| Mechanical inoculation of free dsRNAs in vivo and in vitro synthesized dsRNAs |
Tomato Orchid Tobacco Chenopodium Quinoa |
ToMV CymMV CMV |
[262] [263] [264] |
| CQAS-dsRNA nanoparticles and Carbon Quantum Dots (CQDs) through Root soaking, infiltration | Tobacco Tomato Pepper |
PVY | [265] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



